
Leukocytic-Vascular Endothelial Growth Factor and Integrin ...
11
THE EGYPTIAN JOURNAL OF IMMUNOLOGY Vol. 15 (2), 2008 Page: 81-91 Leukocytic-Vascular Endothelial Growth Factor and Integrin α v ß 3 in Acute Myeloid Leukemia: Relation to Clinical Outcome 1 Lobna A. Abou-Shamaa and 2 Gihan N. Mahmoud Departments of 1 Immunology, 2 Hematology, Medical Research Institute, Alexandria University, Alexandria, Egypt. Different signaling routes seem to be simultaneously triggered in leukemia, with distinct and overlapping activities. Different reports emphasize the interaction between vascular endothelial growth factor (VEGF) and integrin αvß3 as a key control system of angiogenesis, oncogenesis and metatasis. The current study was undertaken to investigate leukocytic-VEGF and integrin αvß3 as correlated with clinical outcome in patients with acute myeloid leukemia (AML). The study groups included 10 newly diagnosed AML patients before the start of any chemotherapeutic medication and 10 normal healthy control subjects. The level of VEGF was estimated in culture supernatant of peripheral blood mononuclear cells (PBMN) of both groups using commercially available ELISA kit. The degree of integrin αvß3 expression on PBMN was estimated by indirect immunoflourescence. Obtained results showed that the level of VEGF and degree of expression of integrin αvß3 were significantly higher in AML patients than in normal healthy subjects. However, no significant correlation was observed between the levels of VEGF and the degree of expression of integrin αvß3.When clinical findings were concerned, there was a significant positive correlation between VEGF and the percentage of blasts, both in peripheral blood & bone marrow. On the other hand, such correlations were not observed in case of integrin αvß3. In addition no significant correlation was observed between either VEGF or integrin αvß3 and clinical staging, age, and sex. In conclusion, our results proved the importance of VEGF and integrin αvß3 in the pathogenesis of AML. However, the per se increased production or/and secretion of VEGF and integrin αvß3 by leukemic PBMN cells, respectively can not be used as independent predictor (s) for clinical outcome in AML patients. It is more comprehensive to study changes of intracellular signaling pathways when such critically interacting factors are concerned in the leukemic process. cute myelocytic leukemia (AML) is a malignant neoplasm of hematopoietic cells characterized by an abnormal proliferation of myeloid precursor cells, decreased rate of self-destruction and an arrest in cellular differentiation. The leukemic cells have an abnormal survival advantage. Thus, the bone marrow and peripheral blood are characterized by leukocytosis with a predominance of immature cells, primarily blasts. As the immature cells accumulate in the bone marrow, they replace the normal myelocytic cells, megakaryocytes, and erythrocytic cells. This leads to a loss of normal bone marrow function and associated complications of bleeding, anemia, and infection (Löwenberg et al., 1999). The endothelium forms a network of vasculature spanning the whole cavity of the bone marrow, thus providing a blood supply like in solid organs (Mazo et al., 1999). Accordingly, increased bone marrow angiogenesis, the process of new blood vessel formation from endothelial precursors, is crucial for the pathogenesis and progression of AML and other hematological malignancies similar to solid invasive tumors (Aguayo et al., 2000). A critical event in the regulation of angiogenesis is the signaling cascade involving vascular endothelium growth factor (VEGF-A, VEGF), which is a multifunctional, secreted cytokine that stimulates endothelial cells to proliferate, to migrate, and to increase their permeability to plasma proteins, (Karamysheva, 2008). The ability of this factor to enhance vascular permeability defines its important role in tumor cell penetration into vascular networks A
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Leukocytic-Vascular Endothelial Growth Factor and Integrin ...

THE EGYPTIAN JOURNAL OF IMMUNOLOGY Vol. 15 (2), 2008 Page: 81-91
Leukocytic-Vascular Endothelial Growth Factor and Integrin αvß3 in Acute Myeloid Leukemia: Relation to Clinical Outcome
1Lobna A. Abou-Shamaa and 2Gihan N. Mahmoud
Departments of 1Immunology, 2Hematology, Medical Research Institute, Alexandria University, Alexandria, Egypt.
Different signaling routes seem to be simultaneously triggered in leukemia, with distinct and overlapping activities. Different reports emphasize the interaction between vascular endothelial growth factor (VEGF) and integrin αvß3 as a key control system of angiogenesis, oncogenesis and metatasis. The current study was undertaken to investigate leukocytic-VEGF and integrin αvß3 as correlated with clinical outcome in patients with acute myeloid leukemia (AML). The study groups included 10 newly diagnosed AML patients before the start of any chemotherapeutic medication and 10 normal healthy control subjects. The level of VEGF was estimated in culture supernatant of peripheral blood mononuclear cells (PBMN) of both groups using commercially available ELISA kit. The degree of integrin αvß3 expression on PBMN was estimated by indirect immunoflourescence. Obtained results showed that the level of VEGF and degree of expression of integrin αvß3 were significantly higher in AML patients than in normal healthy subjects. However, no significant correlation was observed between the levels of VEGF and the degree of expression of integrin αvß3.When clinical findings were concerned, there was a significant positive correlation between VEGF and the percentage of blasts, both in peripheral blood & bone marrow. On the other hand, such correlations were not observed in case of integrin αvß3. In addition no significant correlation was observed between either VEGF or integrin αvß3 and clinical staging, age, and sex. In conclusion, our results proved the importance of VEGF and integrin αvß3 in the pathogenesis of AML. However, the per se increased production or/and secretion of VEGF and integrin αvß3 by leukemic PBMN cells, respectively can not be used as independent predictor (s) for clinical outcome in AML patients. It is more comprehensive to study changes of intracellular signaling pathways when such critically interacting factors are concerned in the leukemic process.
cute myelocytic leukemia (AML) is a
malignant neoplasm of hematopoietic
cells characterized by an abnormal
proliferation of myeloid precursor cells,
decreased rate of self-destruction and an arrest
in cellular differentiation. The leukemic cells
have an abnormal survival advantage. Thus,
the bone marrow and peripheral blood are
characterized by leukocytosis with a
predominance of immature cells, primarily
blasts. As the immature cells accumulate in
the bone marrow, they replace the normal
myelocytic cells, megakaryocytes, and
erythrocytic cells. This leads to a loss of
normal bone marrow function and associated
complications of bleeding, anemia, and
infection (Löwenberg et al., 1999).
The endothelium forms a network of
vasculature spanning the whole cavity of the
bone marrow, thus providing a blood supply
like in solid organs (Mazo et al., 1999).
Accordingly, increased bone marrow
angiogenesis, the process of new blood vessel
formation from endothelial precursors, is
crucial for the pathogenesis and progression
of AML and other hematological
malignancies similar to solid invasive tumors
(Aguayo et al., 2000). A critical event in the
regulation of angiogenesis is the signaling
cascade involving vascular endothelium
growth factor (VEGF-A, VEGF), which is a
multifunctional, secreted cytokine that
stimulates endothelial cells to proliferate, to
migrate, and to increase their permeability to
plasma proteins, (Karamysheva, 2008). The
ability of this factor to enhance vascular
permeability defines its important role in
tumor cell penetration into vascular networks
A

Leukocytic-VEGF and Integrin αvß3 in AM 82
and metastasis (Sierra, 2005). VEGF exists as
five different isoforms of 121, 145, 165, 189
and 206 amino acids (Neufeld et al., 1996).
On adult endothelial cells it exhibits high-
affinity binding sites corresponding to two
distinct tyrosine kinase receptors, the VEGF
receptor (VEGFR)-1 encoded by Flt-1 and
VEGFR-2 encoded by KDR/ Flk-1 with much
greater binding affinity to VEGFR1.
(Robinson et al., 2001). It now appears that
VEGF also has autocrine functions acting as a
survival factor for tumour cells protecting
them from stresses such as hypoxia,
chemotherapy and radiotherapy. The
mechanisms of action of VEGF are still being
investigated with emerging insights into
overlapping pathways and cross-talk with
other receptors (Byrne et al., 2005).
Adhesion molecules, including integrins,
mediate critical cytosolic signaling events that
regulate both physiologic and pathologic
events, including complex processes such as
leucocyte migration, angiogenesis, tumor
growth, and metastasis (Moschos et al.,
2007). Several examples of crosstalk between
integrins and growth factor receptors indicate
that integrin ligation is required for growth
factor–induced biological processes.
Furthermore, integrins can directly associate
with growth factor receptors, thereby
regulating the capacity of integrin/growth
factor receptor complexes to propagate
downstream signaling (reviewed in Eliceiri,
2001). Integrin αvß3 (leukocyte response
integrin) is the cell surface receptor for
vitronectin, fibronectin, fibrinogen, and
denatured collagen and is widely distributed
on monocytes, vascular endothelial cells and
tumor cells (Capo et al., 1999; Kerr et al.,
2002; Kumar, 2003). It plays an important
role in regulation of cell growth,
differentiation, migration, and angiogenesis.
The adhesion of integrin αvß3 to the ECM
proteins induces focal adhesion contacts
which generate the cascade of
phosphorylation of many signal transduction
molecules, including tyrosine kinases (FAK,
pp60src), cytoskeletal proteins (vinculin,
paxillin), Grb-2, and MAP kinase (reviewed
in Hodivala-Dilke K. 2008). Previous reports
have provided evidences for the interaction
between VEGF and integrin αvß3 as regards
tumor progression (De et al., 2005; Soldi et
al., 1999). So, in the current study we
investigated leukocytic-VEGF and integrin
αvß3 as related to clinical outcome in patients
with AML.
Subjects and Methods
The present study was conducted on ten newly
diagnosed AML patients before the start of any
chemotherapeutic medication. Ten normal healthy
subjects were included as a normal control group.
Patients were obtained from Hematology Department,
Medical Research Institute, University of Alexandria.
Patients were subjected to detailed history taking and
thorough clinical examination with special emphasis on
lymph nodes, liver and spleen.
Diagnosis of AML based on standard morphology
and cytochemistry of peripheral blood and bone
marrow films according to the French-American-
British (FAB) criteria (Bennett et al., 1985). Diagnosis
was confirmed by immunophenotyping using a
comprehensive panel of monoclonal antibodies against
myeloid associated antigens as proposed by the EGIL
group (Bene et al., 1995).
Routine Hematological Investigations
Complete blood picture including hemoglobin
concentration, platelet count as well as total and
differential leukocytic counts was done using
automated blood cell counter (Hema Star II). Films
from fresh blood samples and bone marrow aspirate
were stained by Leishmann stain and cytochemical
staining to confirm the differential blood leukocytic
count and detect abnormal leukocyte morphology and
characteristic abnormalities in the bone marrow (Bain
et al., 2001).
Cell Culture
Peripheral blood mononuclear cells (PBMN) and
leukemic blast cells were isolated from normal control
subjects and AML patients, respectively. Simply,
heparinized venous blood samples were centrifuged
over ficoll-hypaque density gradient (Sigma) at 1800
rpm for 45 min., washed three times with saline (NaCl
0.9%), and resuspended in RPMI-1640 medium
(Gibco, UK) supplemented with 10% fetal calf serum,

THE EGYPTIAN JOURNAL OF IMMUNOLOGY 83
100 IU/mL penicillin, 100 mg/mL streptomycin and 2
mM glutamine. Viability of the isolated cells was tested
using dye exclusion technique which is based on the
impermeability of viable cells to trypan blue.
Resuspended cells were cultured in 96 wells flat bottom
polystyrene culture plates (2x105
cells/well) and
stimulated with 20µg/ml phytohaemagglutinin (PHA;
Sigma). After incubation for 72 hrs in a humidified 5%
CO2 incubator at 37°C, culture supernatant was
collected and stored at -70°C till time of assay for
VEGF (Fiedler et al., 1997).
Measurement of Leukocytic-VEGF-A (VEGF)
The level of VEGF was measured in culture
supernatant using commercially available enzyme
linked immunosorbent assay [ELISA] kit [Human
VEGF-A ELISA Kit, Bender MedSystems, Austria]
and following manufacturer recommendations. The
sensitivity of the applied ELISA kit is 20 pg/ml.
Concentrations were deduced from a manually
constructed standard curve using supplied standard
concentrations and corresponding optical densities.
Expression of Leukocytic-Integrin αvß3
The degree of expression of integrin αvß3 on PBMN
was assessed by indirect immunoflourescent technique
as previously described (Ylänne et al., 1990) with mild
modification. Briefly, PBMN cells were attached to
glass slides by cytospin centrifugation and fixed in
100% ethanol. PBMN monolayers were washed in PBS
(pH 7.4) containing 0.2% bovine serum albumin and
then incubated with the first monoclonal antibody
[1/100, mouse anti-human integrin alphavbeta3,
Chemicon International, Austria] for 45 min at room
temperature. After washing with PBS, pH 7.4, samples
were incubated with goat anti-mouse IgG coupled to
fluorescein isothiocyanate [1/50, Goat-anti-mouse IgG
(Fc specific), FITC conjugate, Sigma] for 45 min at
room temperature in the dark. Slides were then washed
in PBS (pH 7.4) and mounted in glycerol-PBS (9–1).
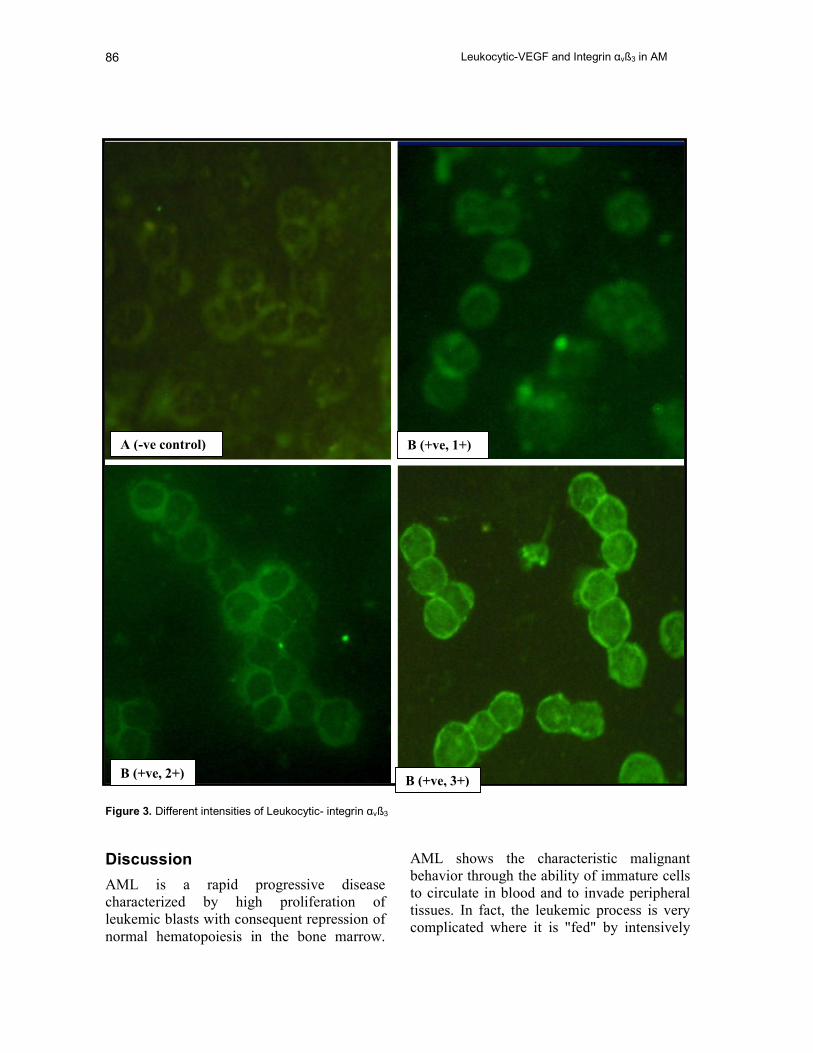
Expression was scored blindly [(+), (++), and (+++)]
by fluorescent microscopic examination considering
intensity and density of fluorescence on the periphery
of the cells.
Statistical Analysis
Statistical analysis was done using Statistical Package
for Social Sciences (SPSS/version 15) software. Chi-
square test and student t-test were used for comparing
qualitative and quantitative data of each two groups,
respectively. Person correlation coefficient was used to
detect the correlation between different variables. The
level of significant was 0.05. Numerical values are
presented as mean ± SD.
Results
Subjects under study included ten newly
diagnosed AML patients (2 males and 8
females, mean age 43.5±13.52 years) before
the start of any chemotherapeutic medication
and ten normal healthy control subjects (4
males and 6 females, mean age 41.6±10.33
years). No significant difference was observed
between the two groups as regards age and
sex.
Clinical data
All control subjects were completely healthy
with normal complete blood picture. Total
leukocytic count (TLC) and percentage of
leukemic blasts in peripheral blood (PB) and
bone marrow (BM) in AML patients at
presentation are presented in Table (1).
According to FAB classification, AML
patients were either M1, M2, or M4 (number=
4, 2, and 4, respectively).
Table 1. TLC and leukemic blasts in AML patients.
Min. Max. Mean S.D.
TLC(x109 / L) 8600.0 75000.0 34551.0 22502.96
Blasts in PB (%) 12.0 80.0 51.30 22.80
Blasts in BM (%) 22.0 95.0 60.8 31.70

Leukocytic-VEGF and Integrin αvß3 in AM 84
Leucocytic-VEGF
The level of VEGF in culture supernatant of
PBMN from AML patients was significantly
higher than corresponding level in normal
control subjects, (253.8±121.49 vs
143.5±20.94 pg/ml, respectively, P= 0.0093).
(Table 2, Figure 1). In AML patients,
estimated level of VEGF had a significant
positive correlation with the percentage of
blasts, both in PB & BM. (P= .033 & .011,
respectively). However, no significant
correlation was observed when clinical
staging, age, and sex were concerned in
relation to VEGF.
Leucocytic- integrin αvß3
The intensity of expression of integrin αvß3 on
PBMN from AML patients was significantly
higher than that in normal control subjects, P=
0.0006. (Table 2, Figure 2, 3). However, there
was no significant correlation between
leukocytic- VEGF and integrin αvß3.
Similarly, no significant correlation was
observed when percentage of blasts, clinical
staging, age, and sex were concerned in
relation to integrin αvß3 in AML patients.
1010N =
Normal controlAML patients
VE
GF
600
500
400
300
200
100
0
2
5
Figure 1. Leukocytic-VEGF in AML patients and normal control subjects.
Le
uko
cytic-V
EG
F p
g/m
l

THE EGYPTIAN JOURNAL OF IMMUNOLOGY 85
Table 2. Leukocytic-VEGF and integrin αvß3 in AML patients and normal control subjects.
AML patients
n=10
Normal control
n=10
VEGF(pg/ml)
Range
Mean ± S.D.
68 – 480
253.8±121.49
120 – 184
143.5±20.94
P 0.0093*
Integrin αvß3
+
++
+++
0 (0.0%)
2 (20.0%)
8 (80.0%)
6 (60.0%)
4 (40.0%)
0 (0.0%)
P 0.0006*
P<0.05 is significant.
+60.0%
++40.0%
++20.0%
+++80.0%
Normal controlAML patients
Figure 2. Leukocytic- integrin αvß3 in AML patients and normal control subjects.
Leukocytic-VEGF and Integrin αvß3 in AM 86
Figure 3. Different intensities of Leukocytic- integrin αvß3
Discussion
AML is a rapid progressive disease
characterized by high proliferation of
leukemic blasts with consequent repression of
normal hematopoiesis in the bone marrow.
AML shows the characteristic malignant
behavior through the ability of immature cells
to circulate in blood and to invade peripheral
tissues. In fact, the leukemic process is very
complicated where it is "fed" by intensively
B (+ve, 1+) A (-ve control)
B (+ve, 2+) B (+ve, 3+)

THE EGYPTIAN JOURNAL OF IMMUNOLOGY 87
interacting leukemic cells, endothelial cells
and extracellular matrix. Among the
important candidates of this interplay are
agiogenic growth factors and adhesion
molecules (McKenzie, 2005; Wood, 2007). In
the current study we are concerned with two
important candidates; VEGF and integrin
αvβ3.
Our results revealed that leukemic PBMN
produce significantly higher levels of VEGF
than normal PBMN. Such data support
pervious evidences for the importance of
leukocyte-associated VEGF in the
pathogenesis of AML. It was previously
observed that leukemic blasts from AML
patients express and produce significantly
increased levels of VEGF (Aguayo et al.,
2000; Dias et al., 2000; Fiedler et al., 1997;
Hussong et al, 2000; Loges et al., 2006).
It is now quite evidenced that acute
leukemia cells use angiogenic growth factor
signaling pathways, namely those activated by
VEGF in autocrine and paracrine fashions.
The presence of VEGFR-1 and -2 on many of
leukemic blasts implies that expansion of the
leukemic population may be facilitated by an
autocrine loop (Bellamy et al., 2001). Such
autocrine action of VEGF was supported by
other more recent studies (Fragoso et al.,
2007, Vales et al., 2007). Moreover, the
leukemia-derived VEGF can also stimulate
the production of growth factors, including
interleukin-6 [IL-6] and granulocyte-
macrophage colony stimulating factor [GM-
CSF], by human endothelial cells, which in
turn further promotes the growth of leukemia
cells (the paracrine loop). In a system where
leukemia cells are co-cultured with
endothelial cells, IMC-2C6, a fully human
anti-VEGFR2 antibody, inhibits both the
production of IL-6 and GM-CSF by
endothelial cells and the growth of leukemia
cells. In addition, IMC-2C6 effectively blocks
VEGF-induced migration of KDR+ human
leukemia cells, and when administered in
vivo, significantly prolonged survival of mice
inoculated with KDR+ human leukemia cells
(Zhang et al., 2004). Autocrine and paracrine
actions of VEGF results in leukemic cell
proliferation, increased survival and
migration. Paracrine growth stimulation may
not only be restricted to the bone marrow
microenvironment, but may also take place at
extramedullary sites. Circulating AML blasts
may profit from paracrine provision of growth
factors in various capillary beds. This may
result in their expansion in peripheral blood
(Fragoso et al., 2007).
On the contrary, Litwin et al., (2002);
showed a discrepancy between bone marrow
vascularity and VEGF expression in vivo and
VEGF expression and angiogenesis from 2-
day conditioned medium ex vivo. They
suggested that angiogenesis in AML likely
represents a response to microenvironmental
factors in vivo, rather than being an intrinsic
property of leukemic cells. Actually, the
control of angiogenesis involves not only
changes in the profiles of environmental
angiogenic cues, such as the increase in
VEGFR2 in response to VEGF, but also
changes in the adhesive capacity of the
endothelial cells. In particular, the changes in
one family of adhesion molecules, integrins,
are thought to regulate several of these
angiogenic steps. In this respect the
interaction of co-expressed endothelial
VEGFR2 and integrin αvβ3 is suggested to
play an important role (Hodivala-Dilke,
2008). Recent studies have identified a
significant increase in activated αvβ3 on
angiogenic vessels, and these cells also appear
to express high levels of VEGFR2
(Mahabeleshwar et al., 2008). It was
previously shown that VEGF increases the
expression of αvβ3 by human microvascular
endothelial cells and also enhances cell
adhesion and migration mediated by αvβ3
(Byzova et al., 2000; Senger et al., 1996).
Current results also showed that the
expression of leucocytic- integrin αvß3 was
significantly higher in AML patients than in

Leukocytic-VEGF and Integrin αvß3 in AM 88
normal control subjects. Up to our knowledge,
current study is the first report looking for the
expression of integrin αvß3 by peripheral
blood blast cells in AML patients. It was
previously observed that integrin αvß3 is
expressed by leukemic cell lines (Aswald et
al., 2004, Atkins et al., 1998, Benedetto et al.,
2006) and other metastatic malignant cells
(Felding-Habermann et al., 2001, 2002).
In cancer growth, the modification of
integrin structure is often associated with both
quantitative and qualitative alterations in
integrin cell surface patterns (Mizejewski,
1999). A characteristic feature of integrins,
including αvß3, is the capacity to transmit
signals bidirectionally, both inside-out and
outside-in. (Byzova et al., 2000). Integrin
activation, or inside-out signaling, is a tightly
governed process involving conformational
changes within the highly conserved
cytoplasmic tail of integrin receptor ß
subunits (Hughes et al., 1996) and provides a
mechanism of integrin regulation. This
process occurs when an agonist binds to a
traditional receptor that ultimately changes the
activation state of an integrin (Banno et al.,
2008). Previous studies emphasize the role of
integrin αvß3 as a ‘‘gatekeeper’’ of VEGF-
mediated processes (Borges et al., 2000,
Reynolds et al., 2004, Soldi et al., 1999).
More recently, De S. et al., (2005), have
defined a mechanism where by integrin αvß3,
through activation, clustering, and signaling,
regulates the production of VEGF in tumor
cells expressing this integrin. Tumors with
‘‘activatable’’ but not ‘‘inactive’’ ß3 integrin
secrete high levels of VEGF, which in turn
promotes extensive neovascularization and
augments tumor growth in vivo. This
stimulation of VEGF expression depends
upon the ability of αvß3 integrin to cluster and
promote phosphorylation of p66 Shc (Src
homology 2 domain containing).
We could not find a significant correlation
between released VEGF and expressed
integrin αvß3.When clinical and hematological
findings were concerned, our data revealed a
significant positive correlation between
VEGF and the percentage of blasts, both in
PB & BM. On the other hand, such
correlations were not observed in case of
integrin αvß3. In addition, no significant
correlation was observed between either
VEGF or integrin αvß3 and clinical staging,
age, and sex. Several reports pointed to the
correlation between increased VEGF and
worse prognosis (Aguayo et al, 1999, 2002;
de Bont et al., 2002). In addition, increased
expression of integrin αvß3 was associated
with poor prognosis and increased metastasis
in solid tumors (Gasparini et al., 1998;
Kageshita et al., 2000). However, our results
suggested that it is not just the quantitative
analysis which reflects the importance of
VEGF and integrin αvß3 in the leukemic
process. Looking for qualitative changes as
well as intracellular signaling pathways is
more indicative for the ultimate interaction
between these two importantly cooperating
factors.
In conclusion, obtained data confirm the
importance of VEGF and integrin αvß3 VEGF
in the pathogenesis of AML. The above
mentioned discussion reflects the complex
nature of interaction between VEGF and
intgrin αvß3 in controlling the leukemic
process. So, it is better to rely on related
qualitative rather than just quantitative
changes. The intimate link between intgrin
αvß3 and VEGF in tumor growth and
angiogenesis may influence anti-integrin as
well as anti-VEGF therapeutic strategies.
References
1. Aguayo A, Estey E, Kantarjian K, Taghi
Mansouri T, Gidel C, Keating M, Giles F, Estrov
Z, Barlogie B, Albitar M. (1999). Cellular Vascular
Endothelial Growth Factor Is a Predictor of
Outcome in Patients With Acute Myeloid
Leukemia. Blood. 94: 3717-21
2. Aguayo A, Kantarjian H, Manshouri T, Gidel C,
Estey E, Thomas D, Koller C, Estrov Z, O'Brien S,
Keating M, Freireich E, Albitar M. (2000).

THE EGYPTIAN JOURNAL OF IMMUNOLOGY 89
Angiogenesis in acute and chronic leukemias and
myelodysplastic syndromes. Blood. 96: 2240-45.
3. Aguayo, A., Kantarjian, H.M., Estey, E.H., Giles,
F.J., Verstovsek, S., Manshouri, T., Gidel, C.,
O’Brien, S., Keating, M.J., Albitar, M. (2002)
Plasma vascular endothelial growth factor levels
have prognostic significance in patients with acute
myeloid leukemia but no in patients with
myelodysplastic syndromes. Cancer. 95: 1923–30.
4. Aswald S, Lutynski A, Moaddeli N, Wells RA,
Schuh AC. (2004). Reduced expression of the
coxsackievirus and adenovirus receptor and of
alpha(v) integrins differentiates myelodysplasia-
related and primary acute myeloid leukaemia.
Leukemia.18:1316-9.
5. Atkins K, Berry JE, Zhang WZ, Harris JF,
Chambers AF, Simpson RU, Somerman MJ.
(1998). Coordinate expression of OPN and
associated receptors during monocyte/macrophage
differentiation of HL-60 cells. J Cell Physiol. 175:
229-37.
6. Bain B, Bates I. (2001) Basic hematological
techniques. In: Lewis SM, Bain B, Bates I. Dacie
and Lewis Practical Hematology 9th ed. Churchill
living stone, UK; pp. 19.
7. Banno A, Ginsberg MH. (2008). Integrin
activation. Biochem Soc Trans. 36: 229-34.
8. Bellamy WT, Richter L, Sirjani D, Roxas C,
Glinsmann-Gibson B, Frutiger Y, Grogan TM, List
AF. (2001). Vascular endothelial cell growth factor
is an autocrine promoter of abnormal localized
immature myeloid precursors and leukemia
progenitor formation in myelodysplastic
syndromes. Blood. 97:1427-34.
9. Bene MC, Castoldi G, Knapp W, Ludwig WD,
Matutes E, Orfao A, van't Veer MB. (1995).
Proposals for the immunological classification of
acute leukemias. European Group for the
Immunological Characterization of Leukemias
(EGIL). Leukemia. 9: 1783-86.
10. Benedetto S, Pulito R, Crich SG, Tarone G, Aime
S, Silengo L, Hamm J. (2006). Quantification of
the expression level of integrin receptor
alpha(v)beta3 in cell lines and MR imaging with
antibody-coated iron oxide particles. Magn Reson
Med. 56: 711-6.
11. Bennett J, Catovsky D, Daniel M, Flandrin G,
Galton D, Gralnick H, Sultan C. (1985). Proposed
revised criteria for the classification of acute
myeloid leukemia. A report of the French-
American-British Cooperative Group. Ann Intern
Med. 103: 620-25.
12. Borges E, Jan Y, Ruoslahti E. (2000) Platelet-
derived growth factor receptor-ß and vascular
endothelial growth factor receptor-2 bind to the ß3
integrin through its extracellular domain. J Biol
Chem. 275: 39867–73.
13. Byrne AM, Bouchier-Hayes DJ, Harmey JH.
(2005). Angiogenic and cell survival functions of
vascular endothelial growth factor (VEGF). J Cell
Mol Med. 9: 777-94.
14. Byzova TV, Goldman CK, Pampori N, Thomas
KA, Bett A, Shattil SJ, Plow EF. (2000). A
mechanism for modulation of cellular responses to
VEGF: activation of the integrins. Mol Cell. 6:
851–60
15. Capo C, Lindberg FP, Meconi S, Zaffran Y, Tardei
G, Brown EJ, Raoult D, Mege JL. (1999).
Subversion of monocyte functions by coxiella
burnetii: impairment of the cross-talk between
alphavbeta3 integrin and CR3. J Immunol. 163:
6078-85.
16. de Bont, E.S., Fidler, V., Meeuwsen, T., Scherpen,
F., Hahlen, K. & Kamps, W.A. (2002). Vascular
endothelial growth factor secretion is an
independent prognostic factor for relapse-free
survival in pediatric acute myeloid leukemia
patients. Clinical Cancer Research. 8: 2856–61
17. De S, Razorenova O, McCabe NP, O'Toole T, Qin
J, Byzova TV. (2005). VEGF-integrin interplay
controls tumor growth and vascularization. Proc
Natl Acad Sci U S A. 102: 7589-94.
18. Dias S, Hattori K, Zhu Z, Heissig B, Choy M,
Lane W, Wu Y, Chadburn A, Hyjek E, Gill M,
Hicklin DJ, Witte L, Moore MA, Rafii S. (2000).
Autocrine stimulation of VEGFR-2 activates
human leukemic cell growth and migration. J Clin
Invest. 106: 511-21.
19. Eliceiri BP. (2001). Integrin and growth factor
receptor crosstalk. Circ Res. 89: 1104-10.
20. Felding-Habermann, B; Fransvea, E; O’Toole, TE;
Manzuk, L; Faha, B; Hensler, M. (2002).
Involvement of tumor cell integrin alpha v beta 3 in
hematogenous metastasis of human melanoma
cells. Clin Exp Metastasis. 19: 427–36.
21. Felding-Habermann, B; O’Toole, TE; Smith, JW;
Fransvea, E; Ruggeri, ZM; Ginsberg, MH; Hughes,
PE; Pampori, N; Shattil, SJ; Saven, A; Mueller,
BM. (2001) Integrin activation controls metastasis

Leukocytic-VEGF and Integrin αvß3 in AM 90
in human breast cancer. Proc Natl Acad Sci USA.
98: 1853–58.
22. Fiedler W, Graeven U, Ergün S, Verago S, Kilic
N, Stockschläder M, Hossfeld DK. (1997).
Vascular endothelial growth factor, a possible
paracrine growth factor in human acute myeloid
leukemia. Blood. 89: 1870-75.
23. Fragoso R, Elias AP, Dias S. (2007). Autocrine
VEGF loops, signaling pathways, and acute
leukemia regulation. Leuk Lymphoma. 48: 481-88.
24. Gasparini G, Brooks PC, Biganzoli E, Vermeulen
PB, Bonoldi E, Dirix LY, Ranieri G, Miceli R,
Cheresh DA. (1998). Vascular integrin
alpha(v)beta3: a new prognostic indicator in breast
cancer. Clin Cancer Res.4: 2625-34.
25. Hodivala-Dilke K. (2008). αvß3 integrin and
angiogenesis: a moody integrin in a changing
environment. Cur Opin Cell Biol. 20:514–19
26. Hughes PE, Diaz-Gonzalez F, Leong L, Wu C,
McDonald JA, Shattil SJ, Ginsberg MH. (1996).
Breaking the integrin hinge. A defined structural
constraint regulates integrin signaling. J Biol
Chem. 271: 6571–74
27. Hussong, J.W., Rodgers, G.M., Shami, P.J. (2000)
Evidence of increased angiogenesis in patients with
acute myeloid leukemia.Blood.95: 309–13.
28. Kageshita T, Hamby CV, Hirai S, Kimura T, Ono
T, Ferrone S. (2000). Alpha(v)beta3 expression on
blood vessels and melanoma cells in primary
lesions: differential association with tumor
progression and clinical prognosis. Cancer
Immunol Immunother. 49: 314-18.
29. Karamysheva AF. (2008). Mechanisms of
angiogenesis. Biochemistry (Mosc). 8: 247-57.
30. Kerr JS, Slee AM, Mousa SA. (2002). The alpha v
integrin antagonists as novel anticancer agents: an
update. Expert Opin Investig Drugs. 11: 1765-74.
31. Kumar CC. (2003). Integrin alpha v beta 3 as a
therapeutic target for blocking tumor-induced
angiogenesis. Curr Drug Targets. 2: 123-31.
32. Litwin C, Leong KG, Zapf R, Sutherland H,
Naiman SC, Karsan A. (2002). Role of the
microenvironment in promoting angiogenesis in
acute myeloid leukemia. Am J Hematol. 70: 22-30.
33. Loges S, Tinnefeld H, Metzner A, Jücker M,
Butzal M, Bruweleit M, Fischer U, Draab E,
Schuch G, O'-Farrel AM, Hossfeld DK,
Bokemeyer C, Fiedler W. (2006). Downregulation
of VEGF-A, STAT5 and AKT in acute myeloid
leukemia blasts of patients treated with SU5416.
Leuk Lymphoma. 47: 2601-09.
34. Löwenberg B, Downing JR, Burnett A. (1999).
Acute myeloid leukemia. N Engl J Med. 341:
1051-106
35. Mahabeleshwar GH, Chen J, Feng W, Somanath
PR, Razorenova OV, Byzova TV. (2008). Integrin
affinity modulation in angiogenesis. Cell Cycle. 7:
335-47.
36. Mazo IB, von Andrian UH. (1999). Adhesion and
homing of blood-borne cells in bone marrow
microvessels. J Leukoc Biol. 66: 25-32.
37. McKenzie SB. (2005). Advances in understanding
the biology and genetics of acute myelocytic
leukemia. Clin Lab Sci. 18: 28-37.
38. Mizejewski GJ.(1999). Role of integrins in cancer:
survey of expression patterns. Proc Soc Exp Biol
Med.222: 124-38.
39. Moschos SJ, Drogowski LM, Reppert SL,
Kirkwood JM. (2007). Integrins and cancer.
Oncology. 21: 13-20.
40. Neufeld G, Cohen T, Gitay-Goren H, Poltorak Z,
Tessler S, Sharon R, Gengrinovitch S, Levi BZ.
(1996). Similarities and differences between the
vascular endothelial growth factor (VEGF) splice
variants. Cancer Metastasis Rev. 15: 153-158.
41. Reynolds AR, Reynolds LE, Nagel TE, Lively JC,
Robinson SD, Hicklin D J, Bodary, S. C. &
Hodivala-Dilke, K. M. (2004). Elevated Flk1
(vascular endothelial growth factor receptor 2)
signaling mediates enhanced angiogenesis in beta3-
integrin-deficient mice. Cancer Res. 64: 8643–50.
42. Robinson CJ, Stringer SE. (2001). The splice
variants of vascular endothelial growth factor
(VEGF) and their receptors. J Cell Sci. 114: 853-
65.
43. Senger DR, Ledbetter SR, Claffey KP,
Papadopoulos-Sergiou A, Peruzzi CA, Detmar M.
(1996). Stimulation of endothelial cell migration
by vascular permeability factor/vascular
endothelial growth factor through cooperative
mechanisms involving the αvß3 integrin,
osteopontin, and thrombin. Am J Pathol. 149: 293–
305.
44. Sierra A. (2005). Metastases and their
microenvironments: linking pathogenesis and
therapy. Drug Resist Updat. 73: 751-62.
45. Soldi R, Mitola S, Strasly M, Defilippi P, Tarone
G, Bussolino F. (1999). Role of alphavbeta3

THE EGYPTIAN JOURNAL OF IMMUNOLOGY 91
integrin in the activation of vascular endothelial
growth factor receptor-2. EMBO J. 15: 882-92.
46. Vales A, Kondo R, Aichberger KJ, Mayerhofer M,
Kainz B, Sperr WR, Sillaber C, Jäger U, Valent P.
(2007). Myeloid leukemias express a broad
spectrum of VEGF receptors including neuropilin-
1 (NRP-1) and NRP-2. Leuk Lymphoma. 48:
1997-2007.
47. Wood BL. (2007). Myeloid malignancies:
myelodysplastic syndromes, myeloproliferative
disorders, and acute myeloid leukemia. Clin Lab
Med. 27: 551-75.
48. Ylänne J, Cheresh DA, Virtanen I. (1990).
Localization of beta 1, beta 3, alpha 5, alpha v, and
alpha IIb subunits of the integrin family in
spreading human erythroleukemia cells. Blood. 76:
570-7.
49. Zhang H, Li Y, Li H, Bassi R, Jimenez X, Witte L,
Bohlen P, Hicklin D, Zhu Z. (2004). Inhibition of
both the autocrine and the paracrine growth of
human leukemia with a fully human antibody
directed against vascular endothelial growth factor
receptor 2. Leuk Lymphoma. 45: 1887-97.




















