
Sox18 induces development of the lymphatic vasculature in mice
Upload
independentCategory
view
2download
0
2005;65:9261-9268. Cancer Res Meit A. Björndahl, Renhai Cao, Jeremy B. Burton, et al. Lymphangiogenesis and Lymphatic MetastasisVascular Endothelial Growth Factor-A Promotes Peritumoral
Updated version
http://cancerres.aacrjournals.org/content/65/20/9261
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2005/10/21/65.20.9261.DC1.html
Access the most recent supplemental material at:
Cited Articles
http://cancerres.aacrjournals.org/content/65/20/9261.full.html#ref-list-1
This article cites by 31 articles, 16 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/65/20/9261.full.html#related-urls
This article has been cited by 21 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on May 22, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from Research.
on May 22, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

Vascular Endothelial Growth Factor-A Promotes Peritumoral
Lymphangiogenesis and Lymphatic Metastasis
Meit A. Bjorndahl,1Renhai Cao,
1Jeremy B. Burton,
3Ebba Brakenhielm,
4Piotr Religa,
1
Dagmar Galter,2Lily Wu,
3,4and Yihai Cao
1
1Laboratory of Angiogenesis Research, Microbiology and Tumor Biology Center, and 2Department of Neuroscience, Karolinska Institutet,Stockholm, Sweden and Departments of 3Molecular and Medical Pharmacology and 4Urology, David Geffen School of Medicine atUniversity of California, Los Angeles, California
Abstract
Metastases are commonly found in the lymphatic system. Themolecular mechanism of lymphatic metastasis is, however,poorly understood. Here we report that vascular endothelialgrowth factor (VEGF)-A stimulated lymphangiogenesis in vivoand that overexpression of VEGF-A in murine T241 fibrosarco-mas induced the growth of peritumoral lymphatic vessels,which occasionally penetrated into the tumor tissue. As a resultof peritumoral lymphangiogenesis, metastases in lymphnodes of mice were detected. VEGF-A–overexpressing tumorscontained high numbers of infiltrating inflammatory cells suchas macrophages, which are known to express VEGF receptor(VEGFR)-1. It seemed that in the mouse cornea, VEGF-Astimulated lymphangiogenesis through a VEGF-C/-D/VEGFR-3–independent pathway as a VEGFR-3 antagonist selectivelyinhibited VEGF-C–induced, but not VEGF-A–induced, lym-phangiogenesis. Our data show that VEGF-A contributes tolymphatic mestastasis. Thus, blockage of VEGF-A–inducedlymphangiogenesis may provide a novel approach for preven-tion and treatment of lymphatic metastasis. (Cancer Res 2005;65(20): 9261-8)
Introduction
Metastasis is a hallmark of malignancy that distinguishesinvasive tumors from benign ones and constitutes a major causeof cancer mortality. In certain types of cancers, such as breastcancer, lymphatic metastasis is the major route for the spread ofcancer cells (1). Spontaneous lymphatic metastases are theconsequence of a complex metastatic process that includes (a)dissemination of malignant cells from a primary tumor to thelymphatics; (b) transport of tumor cells via the lymphatics to thelocal lymph nodes; (c) settlement of tumor cells in the lymph nodes;and (d) growth of metastatic tumors in the lymph nodes. Thecomplexity of this process implies the existence of multiple stepscontrolling cancer metastasis. Malignant tumors stimulate blood-and lymphatic vessel growth by producing angiogenic factors (2–4).Although it is known that induction of tumor angiogenesis involvesthe interplay of a dozen tumor-derived growth factors, it is poorlyunderstood how tumors induce lymphangiogenesis and what itsrole is in tumor spread.
Among the tumor-derived growth factors, vascular endothelialgrowth factor (VEGF)-A is a key angiogenic factor most frequentlyused by tumors and other tissues to switch on their angiogenicphenotypes. In fact, most, if not all, tumors express VEGF-A at highlevels (5). VEGF-A is the prototype of a growth factor family thatcontains at least four additional structurally related members,including placenta growth factor, VEGF-B, VEGF-C, and VEGF-D (6).The angiogenic signals triggered by members of the VEGF familyare mainly mediated by the activation of two homologous tyrosinekinase receptors, VEGF receptor (VEGFR)-1 and VEGFR-2, both ofwhich are expressed on blood vessel endothelial cells (7). VEGF-Abinds to VEGFR-1 and VEGFR-2, and induces vasculogenesis,angiogenesis, and vascular permeability. In contrast, placentagrowth factor and VEGF-B only bind to VEGFR-1, which mayparticipate in modulation of VEGF-A–induced angiogenesis (8, 9).In addition to VEGFR-1 and VEGFR-2, a lymphatic endothelial cellspecific tyrosine kinase receptor, VEGFR-3, has been identified.VEGF-C and VEGF-D interact with both VEGFR-2 and VEGFR-3 andinduce both blood angiogenesis and lymphangiogenesis (10, 11).High expression levels of VEGF-C and VEGF-D in tumors have
been correlated with enhanced lymphatic metastasis (11, 12).Interestingly, it has recently been found that VEGF-A also stimulateslymphangiogenesis (13–16). In this article, we show that in a murinexenograft tumor model, VEGF-A induces peritumoral lymphaticvessel growth and promotes lymphatic metastasis.
Materials and Methods
Reagents and animals. All animal studies done at the Karolinska
Institutet were reviewed and approved by the animal care and usecommittee of the North Stockholm Animal Board. All handling of animals
was done in accordance with University of California Los Angeles Animal
Research Committee guidelines (see Supplementary data for details).Vascular endothelial growth factor receptor-3/porcine aortic
endothelial cell shape changes and motility assay. Analysis of VEGFR-3/porcine aortic endothelial (PAE) cell shape changes and motility was
carried out according to our previously published methods, with theinclusion of the anti-mouse VEGFR-3 antibody (500 ng/mL; ref. 10; see
Supplementary data for details).
Mouse corneal neovascularization assay. The mouse corneal angio-
genesis assay was done according to procedures previously described (17).Mouse corneal tumor model. Implantation of tumor tissues in the
mouse cornea was carried out according to previously described procedures
(ref. 18; see Supplementary data for details).Retroviral vector design and tumor cell transduction. A murine
fibrosarcoma cell line, T241, was transfected with complementary cDNAs
coding for human VEGF (hVEGF)-A or hVEGF-C according to previously
published procedures (19).Tumor cell proliferation assay. Proliferation of nontransduced, empty
vector–, hVEGF-A– and hVEGF-C–transduced T241 fibrosarcoma cells was
done as previously published (19).
Note: M. Bjorndahl and R. Cao contributed equally to this work.Supplementary data for this article are available at Cancer Research Online (http://
cancerres.aacrjournals.org/).Requests for reprints: Yihai Cao, Microbiology and Tumor Biology Center,
Karolinska Institutet, 171 77 Stockholm, Sweden. Phone: 46-8-5248-7596; Fax: 46-8-31-94-70; E-mail: [email protected].
I2005 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-04-2345
www.aacrjournals.org 9261 Cancer Res 2005; 65: (20). October 15, 2005
Research Article
Research. on May 22, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

ELISA. Detection of endogenous mouse and human VEGF-A expressionlevels in conditioned media collected from wild-type (WT) and VEGF-A–
transduced T241 cells were determined using the Quantikine ELISA system
(R&D Systems, Inc., Minneapolis, MN) according to the instructions of the
manufacturer and our previously published methods (19). A 72-hourconditioned media was prepared from cell cultures (15 mL/flask) originally
seeded with 4 � 106 cells/flask.
Tumor growth assay. Approximately 1 � 106 tumor cells of WT, vector-,
hVEGF-A– or hVEGF-C–transduced tumor cells were implanted s.c. on theback of 6- to 7-week-old female C57BL/6 mouse, and tumor volumes were
measured as previously reported (20).
Lymphatic metastasis assay. The metastatic potential of s.c. implanted
T241-VEGF-A primary tumors was studied according to previouslydescribed procedures (17). In another experiment, 0.5 � 106 Renilla
luciferase–transduced VEGF-A T241 tumor cells (21) were implanted s.c. in
the right upper back of C57BL/6 mice (n = 10). Anesthetized mice wereimaged weekly using an optical imaging system immediately following i.v.
injection of Renilla luciferase substrate coelenterazine. About 2 to 4 weeksafter tumor implantation, the brachial lymph nodes from both sides were
dissected and imaged ex vivo for the presence of Renilla luciferase as well as
green fluorescent protein (GFP) signals according to previously describedtechniques (22). Optical signals were semiquantified using Living Image 2.5
(Xenogen) and IGOR (Wavemetrics, Lake Oswego, OR) image analysis
softwares. Bioluminescence ex vivo signal was considered positive if the
maximal signal in the region of interest exceeded 1 � 104 photons/s/cm2/steradian (see Supplementary data for details).
Histologic analysis of lymph nodes. Paraffin-embedded tissue sections
(5 Am) of the brachial lymph nodes were stained with Surgipath eosin and
Mayers’ hematoxylin (HistoLab products AB, Gothenburg, Sweden). Further,GFP-positive tumor cells were detected in the lymph nodes using confocal
microscopy (Zeiss Confocal LSM510 Microscope).
Whole-mount staining and confocal analysis. Growth factor– and
tumor-implanted mouse eyes and primary tumors were double stained forCD31 and LYVE-1 using a whole-mount staining protocol recently published
(ref. 17; see Supplementary data for details).
Detection of macrophages. Tumor sections of WT, VEGF-A, andVEGF-C were stained for macrophages according to standard immuno-
histochemical procedures (see Supplementary data for details).
In situ hybridization. Detection of mouse VEGF-C in tumor tissues by
in situ hybridization was done according to our recent published proceduresusing an oligo probe complementary to VEGF-C (nucleotides 536-585 of
murine VEGF-C mRNA; sequence reference no. NM-009506.1; ref. 17).
Statistical analyses. Statistical analyses of the in vitro and in vivo results
were made by a standard two-tailed Student’s t test using Microsoft EXCEL 5.P < 0.05 (*) and P < 0.001 (***) were deemed as significant and highly
significant, respectively.
Results
Vascular endothelial growth factor-A stimulated lymphaticvessel growth in vivo . We developed a mouse corneal lymph-angiogenic model to evaluate the lymphangiogenic activity ofVEGF-A. The newly formed lymphatic vessels were detected usinga specific antibody against LYVE-1, a receptor almost exclusivelyexpressed on lymphatic endothelial cells (23), with the exception ofliver sinusoids (24). Implantation of a slow release polymer lackinggrowth factors did not result in the growth of new lymphaticvessels (Supplementary Fig. S1A , LYVE-1). In contrast, VEGF-Apotently stimulated the growth of new lymphatic vessels sproutingfrom the existing limbal lymphatics (Fig. 1C). These newly formedlymphatic vessels were larger in diameter than newly formed bloodvessels. The lymphatic vessels expressed only low, or undetectable,levels of platelet endothelial cell adhesion molecule 1 (PECAM-1;
Figure 1. Stimulation of lymphangiogenesis by VEGF-A. Confocal analysisof mouse corneal blood- and lymphatic vessels, induced by VEGF-A(A, C, E , and G ) or FGF-2 (B, D, F , and H ) at day 25, using immunofluorescentdouble labeling for CD31 (red, yellow arrows ) and LYVE-1 (blue, white arrows ).Bar, 100 Am. P, pellet.
Figure 2. Stimulation of lymphangiogenesis without blood vessels. VEGF-A wasimplanted on one (front) side (A ) of the cornea and lymphatic vessel growth in theabsence of blood angiogenesis was detected on the opposite side (B ) of thecornea. Corneal whole-mount sections were double stained by immunofluorescentlabeling for CD31 (red) and LYVE-1 (blue ). Bar, 100 Am. P, pellet.
Cancer Research
Cancer Res 2005; 65: (20). October 15, 2005 9262 www.aacrjournals.org
Research. on May 22, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
CD31), and their distribution did not generally overlap with bloodvessels (Fig. 1E and G ; CD31). Similar to VEGF-A, fibroblast growthfactor (FGF)-2, a known lymphangiogenic factor, also induced arobust lymphangiogenic response in this model (Fig. 1D, F, and H).These findings show that VEGF-A is a potent lymphangiogenicfactor.Remodeling of newly formed lymphatic vessels. Similar to
blood vessels, new lymphatic structures were detectable at day 5after VEGF-A implantation (Supplementary Fig. S1B). At this timepoint, the early lymphatic sprouts lacked a vascular tree-likestructure and the blood vessels appeared as disorganized vascularplexuses. At day 14, robust angiogenic and lymphangiogenicresponses were detected, as the early primitive blood- and lymphaticvessels present at day 5 had undergone remarkable remodeling toform vascular tree-like structures mainly consisting of distinctvessels (Supplementary Fig. S1C). Unlike VEGF-A, FGF-2–inducedlymphatic- and blood vessels appeared as well-organized vascula-tures already by day 5, and they were further remodeled into maturevascular networks by day 14 after implantation (SupplementaryFig. S1D and E). Quantification analysis showed that VEGF-A and
FGF-2 induced approximately equal amounts of blood- andlymphatic vessels at day 14 although FGF-2 was more potent atday 5 (Supplementary Fig. S1F and G).Lymphangiogenesis in the absence of blood angiogenesis.
In the corneal lymphangiogenesis model, a VEGF-A pellet wasimplanted into a micropocket created on one side of the circum-ferential corneal globe, as previously described (17). Immuno-histologic analysis on this side of the cornea showed a delayedlymphangiogenic response as compared with blood angiogenesis,suggesting that blood vessels might be a prerequisite for lymphan-giogenesis (Fig. 2A). Surprisingly, on the opposite side of thecircumferential eye globe, VEGF-A stimulated lymphatic vesselgrowth with almost no blood neovascularization (Fig. 2B). Thisfinding shows that VEGF-A–induced lymphangiogenesis can occurin the absence of blood vessels.Anti–vascular endothelial growth factor receptor-3 neutral-
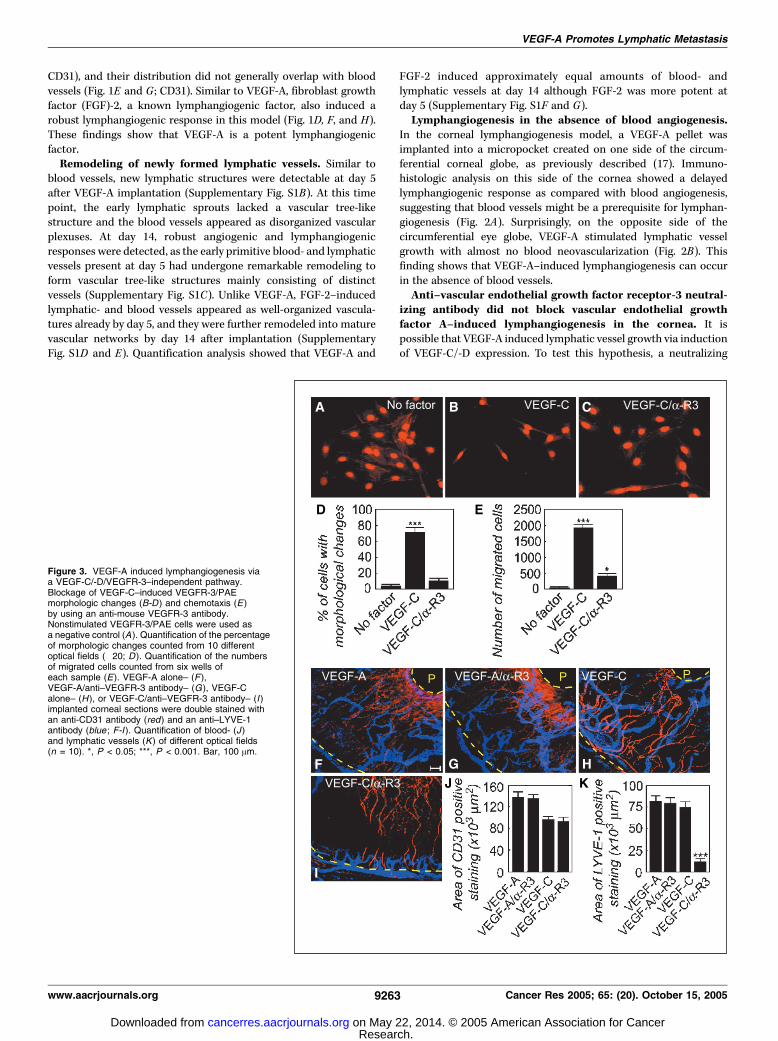
izing antibody did not block vascular endothelial growthfactor A–induced lymphangiogenesis in the cornea. It ispossible that VEGF-A induced lymphatic vessel growth via inductionof VEGF-C/-D expression. To test this hypothesis, a neutralizing
Figure 3. VEGF-A induced lymphangiogenesis viaa VEGF-C/-D/VEGFR-3–independent pathway.Blockage of VEGF-C–induced VEGFR-3/PAEmorphologic changes (B-D ) and chemotaxis (E)by using an anti-mouse VEGFR-3 antibody.Nonstimulated VEGFR-3/PAE cells were used asa negative control (A ). Quantification of the percentageof morphologic changes counted from 10 differentoptical fields (�20; D ). Quantification of the numbersof migrated cells counted from six wells ofeach sample (E). VEGF-A alone– (F ),VEGF-A/anti–VEGFR-3 antibody– (G), VEGF-Calone– (H ), or VEGF-C/anti–VEGFR-3 antibody– (I )implanted corneal sections were double stained withan anti-CD31 antibody (red) and an anti–LYVE-1antibody (blue ; F-I ). Quantification of blood- (J )and lymphatic vessels (K) of different optical fields(n = 10). *, P < 0.05; ***, P < 0.001. Bar, 100 Am.
VEGF-A Promotes Lymphatic Metastasis
www.aacrjournals.org 9263 Cancer Res 2005; 65: (20). October 15, 2005
Research. on May 22, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

anti–VEGFR-3 antibody was examined for its potential effect onVEGF-A–induced lymphangiogenesis in the cornea. To first confirmthe neutralizing activity, the anti–VEGFR-3 antibody was used toblock VEGF-C–induced VEGFR-3/PAE cell activity in vitro . Theanti–VEGFR-3 antibody significantly inhibited VEGF-C–induced
morphologic changes (Fig. 3A-D) and cell migration (Fig. 3E),clearly demonstrating that the anti–VEGFR-3 antibody sufficientlyblocked VEGF-C–induced endothelial cell activity in vitro .To test the blocking capacities of the anti–VEGFR-3 antibody
in vivo , VEGF-A or VEGF-C was coimplanted with the antibody in
Figure 4. Stimulation of tumor growth,angiogenesis, and lymphangiogenesis. A, growthrates of vector-, VEGF-A–, or VEGF-C–transducedtumor cells, or WT tumor cells in vitro. B, tumorvolumes were measured at indicated time pointsand the data are presented as mean determinants(F SE). C, tumors were weighed at day 11 afterimplantation and the data are presented as meandeterminants (F SE). ***, P < 0.001. D to H,peritumoral (D-F ) and intratumoral (G and H)blood- and lymphatic vessels were double labeledfor CD31 (red) and LYVE-1 (blue ). Green signalsare GFP-positive tumor cells. White arrows,tumor borders. Quantification of peritumoral (I )and intratumoral (J ) lymphatic vessels of differentoptical fields (n = 7-9). **, P < 0.01; ***,P < 0.001. Bar, 100 Am.
Cancer Research
Cancer Res 2005; 65: (20). October 15, 2005 9264 www.aacrjournals.org
Research. on May 22, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

the mouse cornea. As expected, the anti–VEGFR-3 antibody almostcompletely blocked VEGF-C–induced lymphangiogenesis, but notblood angiogenesis (Fig. 3H-K). In contrast, neither VEGF-A–induced angiogenesis nor lymphangiogenesis was affected by thisantibody treatment (Fig. 3F, G, J, and K), suggesting that theVEGFR-3 signaling pathway is not critical for the lymphangiogenicactivity of VEGF-A.Vascular endothelial growth factor-A stimulated peritu-
moral lymphangiogenesis in a xenograft tumor model. To testif VEGF-A was able to induce tumoral lymphangiogenesis, wetransfected a murine T241 fibrosarcoma cell line with VEGF-AcDNA. As determined by a sensitive ELISA assay, the VEGF-A–transfected cell line showed an f285-fold increase in theexpression of hVEGF-A (567 ng/mL) as compared with theendogenous levels of mVEGF-A (2.0 ng/mL) in these cells (19).The high expression levels of VEGF-A seemed to have no effect ontumor cell growth rate in vitro (Fig. 4A). However, implantation ofVEGF-A T241 tumor cells in syngeneic mice resulted in acceleratedtumor growth as measured by tumor volume and weight (Fig. 4Band C). Interestingly, VEGF-A T241 tumors contained a largenumber of peritumoral lymphatic vessels (Fig. 4E , LYVE-1). Theperitumoral lymphatic vessels were of larger diameters than thoseof the blood vasculature. Similarly, overexpression of VEGF-C in theT241 fibrosarcoma cells also resulted in a remarkable growth oflymphatic vessels in the peritumoral area (Fig. 4F, LYVE-1). Inaddition to peritumoral lymphatic vessels, the VEGF-C T241tumors also contained a high density of intratumoral lymphaticvessels (Fig. 4H , LYVE-1), whereas in the VEGF-A T241 tumorsintratumoral lymphatic vessels were rarely detectable (Fig. 4G ,LYVE-1). In the vector-transfected (Fig. 4D , LYVE-1) or non-transfected (data not shown) tumors, only a few peritumorallymphatic vessels were detected around the border of the tumors.VEGF-C was more potent than VEGF-A in inducing lymphangio-genesis in the peritumoral as well as the intratumoral area of thetumors (Fig. 4I and J).Vascular endothelial growth factor-A stimulated tumoral
lymphangiogenesis in a corneal tumor model. To further studyVEGF-A–induced tumoral lymphangiogenesis, we established amouse corneal tumor model. Because of the corneal avascularity,tumor growth in the cornea excludes the involvement of any
preexisting blood or lymphatic vessels. Implantation of WT orVEGF-A–transduced tumor tissues into the corneal micropocketsresulted in the growth of tumors, expanding from the micropocketsto the limbus. Corneal tumor neovascularization became visible bydirect gross examination 2 weeks after implantation (Fig. 5Aand F). The implanted VEGF-A–transfected tumor cells induced arobust angiogenic response, a pseudo-hemorrhagic phenotype, witha trimmed leading edge growing towards the implanted tumor(Fig. 5F). In contrast, WT tumors induced relatively well-definedtumor vasculatures in this model (Fig. 5A). Immunohistologicanalysis revealed that VEGF-A tumors contained a honeycomb-likevascular network extending throughout the entire tumor tissue(Fig. 5G, I-K, M , and N). LYVE-1 staining revealed the existence ofgigantic lymphatic vessels growing throughout the entire VEGF-Atumor (Fig. 5H-J and L-N). In the WT tumor, lymphatic vessels onlygrew in the surrounding area of the tumor, and they rarelypenetrated into the tumor tissue (Fig. 5C-E). This corneal tumormodel further shows that VEGF-A stimulates tumoral lymphaticvessel growth. As corneal tumors are relatively flat structures,as compared with those implanted s.c., the peritumoral andintratumoral lymphatic vessels might be difficult to distinguishcompletely in this model system.Vascular endothelial growth factor-A promoted lymphatic
metastasis. Stimulation of peritumoral lymphangiogenesis byVEGF-A raised the possibility that this factor might promotelymphatic metastasis. Indeed, resection of s.c. primary VEGF-AT241 tumors implanted in the middle dorsum of mice resulted inmetastatic lesions of brachial lymph nodes in the majority ofanimals. Of 21 mice, 14 mice had metastases in both brachiallymph nodes, 6 mice had metastases in one brachial lymph node,and 1 mouse had no metastasis. These lymphatic tumor lesionswere significantly greater in size than the lymph nodes of healthyanimals or mice with WT or vector-transfected tumors asmeasured by volume and weight (Fig. 6D and E). Similarly,removal of VEGF-C T241 primary tumors also resulted in brachiallymphatic metastasis on both sides of all mice (n = 16). Incontrast, resection of WT or vector-transfected tumors did notresult in visible metastases in regional lymph nodes at the timepoint when these mice were sacrificed. Histologic examination ofthe lymph nodes confirmed the presence of GFP-positive VEGF-A
Figure 5. VEGF-A–induced corneal tumorangiogenesis and lymphangiogenesis. A piece oftumor tissue (f0.4 mm3) was implanted into eachcorneal micropocket. At day 14 after implantation, WT(A-E) and VEGF-A–overexpressing (F-N ) tumors werephotographed (A and F, yellow arrows) and tumorblood- (B, G , and K ) and lymphatic vessels (C, H , andL) were stained for CD31 (red) and LYVE-1 (blue ).Green, GFP-positive tumor cells. Dashed line, WTtumor border (B-E ). T, tumors. White arrows,lymphatic vessels. Bar, 100 Am.
VEGF-A Promotes Lymphatic Metastasis
www.aacrjournals.org 9265 Cancer Res 2005; 65: (20). October 15, 2005
Research. on May 22, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

or VEGF-C tumor cells in brachial lymph nodes (Fig. 6B and C ,GFP), whereas lymph nodes of control mice lacked detectableGFP-positive tumor cells (Fig. 6A , GFP). H&E staining revealedthe presence of invasive tumor cells in lymph nodes of VEGF-Aand VEGF-C tumor–bearing mice but not in the healthy lymphnodes of control mice (Fig. 6A-C, H , and E). To further validatethese findings, VEGF-A T241 tumor cells were labeled in vitro bytransducing with a lentivirus expressing Renilla luciferase reportergene (21). Two weeks after implantation of the tumor cells in theright upper back of mice, bioluminescence signals were detectablein ex vivo dissected brachial lymph nodes and revealed thepresence of lymphatic metastases (Fig. 6F and G). The opticalsignals were intensified more than 10-fold at 4 weeks afterimplantation (4 � 104 versus 5 � 105 photons/s/cm2/steradian),indicating the in situ growth of the metastatic lesions in thelymph nodes (Fig. 6H). The metastases were mainly confined tothe ipsilateral side, although contralateral involvement was also
observed after 4 weeks. The finding that VEGF-A promotes lymphnode metastasis is further confirmed by our unpublished datafrom a prostate cancer xenograft model where f40% of the micedeveloped brachial lymph node lesions.Vascular endothelial growth factor-A tumors contain high
numbers of inflammatory macrophages. It has recently beenreported that VEGF-A activates inflammatory cells that contributeto VEGF-A–induced lymphangiogenesis (13). To study if VEGF-Acould also induce inflammatory cells in tumors, tissue sections ofvarious tumors were stained with an antimacrophage specificantibody. Interestingly, VEGF-A induced higher numbers ofintratumoral macrophages as compared with WT or VEGF-Ctumors (Supplementary Fig. S2A, C, E , and G). However, VEGF-Cwas also able to significantly induce macrophage infiltration(Supplementary Fig. S2C and G). In situ hybridization analysisshowed that VEGF-A did not up-regulate VEGF-C mRNAexpression in VEGF-A–overexpressing tumors as compared with
Figure 6. Lymphatic metastasis. A to C,implanted s.c. tumors were removed andbrachial lymph node metastases were visible afterf4 weeks in VEGF-C and VEGF-A tumor–bearingmice, but not in vector-bearing mice. Histologicexamination of lymph nodes using H&E staining.LN, lymph node tissue; T, tumor tissue. Dashedlines, borders between tumor tissues and lymphnode tissues. After removal of primary tumors,resected lymph nodes and lymphatic metastaseswere weighed (D ) and the volumes were measured(E). ***, P < 0.001. Bar, 25 Am. VEGF-A T241tumor cells were labeled with Renilla luciferaseand implanted dorsally. Optical signals wereapparent in the primary tumor 1 week afterimplantation (F ). The color scale indicatesmaximal bioluminescence intensity(photons/s/cm2/steradian). Following 2 or 4weeks of tumor growth, the brachial lymphnodes were dissected and the optical signalwas determined ex vivo (G ). Relative opticalintensity was calculated as ipsilateral/contralateralluciferase signal from brachial lymph nodes inthe same animal (H ).
Cancer Research
Cancer Res 2005; 65: (20). October 15, 2005 9266 www.aacrjournals.org
Research. on May 22, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

control tumors (Supplementary Fig. S2B, F, and H). As a positivecontrol, we used a VEGF-C T241 cell line expressing high levels ofVEGF-C mRNA (Supplementary Fig. S2D and H). This findingindicates that VEGF-A does not up-regulate VEGF-C expressionand thus supports our conclusion that VEGF-A induces lymphan-giogenesis via a VEGF-C–independent pathway.
Discussion
Most solid malignant tumors metastasize to regional lymphnodes. However, at present, it remains controversial on the issuewhether tumor lymphatics are functional or not (11, 12, 25). Tumorcells may either access into the lymphatic system by inducingintratumoral lymphangiogenesis or by co-opting preexistinglymphatics in the surrounding tissue (26). As the key function ofthe lymphatic system is immune surveillance, intratumorallymphangiogenesis could be a part of our body’s defense systemagainst cancer. However, cancer cells have attained the ability toescape our immune surveillance system and have adopted growthadvantages in different environments. As a consequence, the lymphnode can become a site for tumor growth instead of a site fortumor elimination.VEGF-A was previously thought to act as a specific blood
angiogenic factor through activation of VEGFR-1 and VEGFR-2.However, a couple of recent studies have shown that VEGF-A isable to induce lymphangiogenesis in animal models (13, 15, 16).In a recent article, Cursiefen et al. (13) report that VEGF-A–activated inflammatory macrophages release VEGF-C/-D thatcontributes to lymphangiogenesis. In support of this finding, weobserved significantly increased number of inflammatory macro-phages in the VEGF-A tumors. However, we were not able toblock VEGF-A–induced lymphangiogenesis with a neutralizinganti–VEGFR-3 antibody in the cornea assay, suggesting thatVEGF-A stimulates lymphatic vessel growth via a VEGF-C/-D/VEGFR-3–independent pathway. In support of our finding, it hasbeen reported that the VEGFR-2 receptor is expressed occasion-ally on lymphatic endothelial cells, suggesting a direct stimula-tory effect of VEGF-A on lymphatic vessels (27). We found thaton the opposite side of the implanted corneal globe, lymphaticvessels can grow without blood vessels, suggesting thatlymphangiogenesis is not entirely dependent on the blood vesselsystem.We observed that tumors overexpressing VEGF-A were highly
infiltrated by macrophages. This finding is consistent with thefact that macrophages are responsive to VEGF-A–induced chemo-taxis via the VEGFR-1 signaling pathway (28). Inflammatory cells,such as macrophages, are recruited to tumors by a wide rangeof tumor cell–derived cytokines and growth factors, including
VEGF-A (29). Recent publications have provided experimentalevidence that macrophages promote progression of solid tumorsand metastasis (30, 31). If monocytes/macrophages play a criticalrole in mediating VEGF-A–induced lymphangiogenesis, what arethe active lymphatic cytokines produced by these cells? Amongknown lymphangiogenic factors, VEGF-C and VEGF-D are thebest-characterized growth factors that act directly on lymphaticendothelial cells via activation of VEGFR-3. Our data show thatan anti–VEGFR-3 neutralizing antibody was unable to block VEGF-A–induced lymphangiogenesis in the cornea tissue, suggestingthat VEGF-C/-D are not major players in mediating VEGF-A–stimulated lymphangiogenesis. The fact that tumors overexpress-ing VEGF-A do not display increased levels of VEGF-C expressionfurther supports this finding.Our results provide convincing evidence that VEGF-A induces a
lymphangiogenic response in tumors expressing this factor at ahigh level. As a result of the lymphangiogenic effect, VEGF-A–expressing tumors metastasize to the local lymph nodes. Consis-tent with our findings, a recent report using a genetic skin tumormodel showed that VEGF-A promotes metastasis to regional lymphnodes (29). Thus, our work, together with this report, providesnovel insights for the understanding of the role of VEGF-A inpromoting primary tumor growth and cancer metastasis. VEGF-Ais probably the most frequently up-regulated angiogenic factorduring tumor growth. Thus, it might also be the most frequentlyoccurring lymphangiogenic factor contributing to lymphaticmetastasis as nearly all solid tumors express VEGF-A at highlevels. Recently, clinical evaluation of a VEGF-A antagonist, avastin,used in the treatment of human cancers, has provided encouragingresults in the field of antiangiogenic therapy. Taken together withour work, further development of VEGF antagonists in thetreatment of human cancers may improve the therapeutic outcomefor cancer patients.
Acknowledgments
Received 7/5/2004; revised 6/30/2005; accepted 8/5/2005.Grant support: The Swedish Research Council, the Swedish Heart and Lung
Foundation, the Swedish Cancer Foundation, the Soderberg Foundation, and theKarolinska Institute Foundation; National Cancer Institute Specialized Program ofResearch Excellence program P50 CA092131 (L. Wu); University of California at LosAngeles Research Training in Pharmacological Sciences Training program T32-GM008652 (J. Burton); and the Swedish Wenner-Gren Foundations (E. Brakenhielm).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Dr. David Jackson (John Radcliffe Hospital, Oxford, United Kingdom)for providing us valuable anti–LYVE-1 antibodies for our study. We thank IrvinChen and John Colicelli (Department of Biological Chemistry, University ofCalifornia, Los Angeles, CA) for providing materials for the generation of lentiviralconstructs.
References1. Fidler IJ. The pathogenesis of cancer metastasis: the‘‘seed and soil’’ hypothesis revisited. Nat Rev Cancer2003;3:453–8.
2. Carmeliet P, Jain RK. Angiogenesis in cancer and otherdiseases. Nature 2000;407:249–57.
3. Folkman J. Angiogenesis in cancer, vascular, rheuma-toid and other disease. Nat Med 1995;1:27–31.
4. Hanahan D, Folkman J. Patterns and emergingmechanisms of the angiogenic switch during tumori-genesis. Cell 1996;86:353–64.
5. Senger DR, Galli SJ, Dvorak AM, Perruzzi CA, Harvey
VS, Dvorak HF. Tumor cells secrete a vascularpermeability factor that promotes accumulation ofascites fluid. Science 1983;219:983–5.
6. Eriksson U, Alitalo K. Structure, expression andreceptor-binding properties of novel vascular endothe-lial growth factors. Curr Top Microbiol Immunol1999;237:41–57.
7. Mustonen T, Alitalo K. Endothelial receptor tyrosinekinases involved in angiogenesis. J Cell Biol 1995;129:895–8.
8. Makinen T, Olofsson B, Karpanen T, et al. Differentialbinding of vascular endothelial growth factor B spliceand proteolytic isoforms to neuropilin-1. J Biol Chem1999;274:21217–22.
9. Hattori K, Heissig B, Wu Y, et al. Placental growthfactor reconstitutes hematopoiesis by recruitingVEGFR1(+) stem cells from bone-marrow microenvi-ronment. Nat Med 2002;8:841–9.
10. Cao Y, Linden P, Farnebo J, et al. Vascular endothelialgrowth factor C induces angiogenesis in vivo . Proc NatlAcad Sci U S A 1998;95:14389–94.
11. Stacker SA, Caesar C, Baldwin ME, et al. VEGF-Dpromotes the metastatic spread of tumor cells via thelymphatics. Nat Med 2001;7:186–91.
12. Skobe M, Hawighorst T, Jackson DG, et al. Inductionof tumor lymphangiogenesis by VEGF-C promotesbreast cancer metastasis. Nat Med 2001;7:192–8.
VEGF-A Promotes Lymphatic Metastasis
www.aacrjournals.org 9267 Cancer Res 2005; 65: (20). October 15, 2005
Research. on May 22, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from

13. Cursiefen C, Chen L, Borges LP, et al. VEGF-Astimulates lymphangiogenesis and hemangiogenesis ininflammatory neovascularization via macrophage re-cruitment. J Clin Invest 2004;113:1040–50.
14. Hirakawa S, Kodama S, Kunstfeld R, Kajiya K, BrownLF, Detmar M. VEGF-A induces tumor and sentinellymph node lymphangiogenesis and promotes lymphat-ic metastasis. J Exp Med 2005;201:1089–99.
15. Hong YK, Lange-Asschenfeldt B, Velasco P, et al.VEGF-A promotes tissue repair-associated lymphaticvessel formation via VEGFR-2 and the a1h1 and a2h1integrins. FASEB J 2004;18:1111–3.
16. Nagy JA, Vasile E, Feng D, et al. Vascular permeabilityfactor/vascular endothelial growth factor induces lym-phangiogenesis as well as angiogenesis. J Exp Med2002;196:1497–506.
17. Cao R, Bjorndahl MA, Religa P, et al. PDGF-BBinduces intratumoral lymphangiogenesis and promoteslymphatic metastasis. Cancer Cell 2004;6:333–45.
18. Muthukkaruppan V, Auerbach R. Angiogenesis in themouse cornea. Science 1979;205:1416–8.
19. Eriksson A, Cao R, Pawliuk R, et al. Placentagrowth factor-1 antagonizes VEGF-induced angiogen-
esis and tumor growth by the formation of function-ally inactive PlGF-1/VEGF heterodimers. Cancer Cell2002;1:99–108.
20. Cao R, Wu HL, Veitonmaki N, et al. Suppression ofangiogenesis and tumor growth by the inhibitor K1-5generated by plasmin-mediated proteolysis. Proc NatlAcad Sci U S A 1999;96:5728–33.
21. Morizono K, Xie Y, Ringpis GE, et al. Lentiviral vectorretargeting to P-glycoprotein on metastatic melanomathrough intravenous injection. Nat Med 2005;11:346–52.
22. Bhaumik S, Gambhir SS. Optical imaging of Renillaluciferase reporter gene expression in living mice. ProcNatl Acad Sci U S A 2002;99:377–82.
23. Prevo R, Banerji S, Ferguson DJ, Clasper S, JacksonDG. Mouse LYVE-1 is an endocytic receptor forhyaluronan in lymphatic endothelium. J Biol Chem2001;276:19420–30.
24. Mouta Carreira C, Nasser SM, di Tomaso E, et al.LYVE-1 is not restricted to the lymph vessels: expressionin normal liver blood sinusoids and down-regulation inhuman liver cancer and cirrhosis. Cancer Res 2001;61:8079–84.
25. Padera TP, Kadambi A, di Tomaso E, et al. Lymphatic
metastasis in the absence of functional intratumorlymphatics. Science 2002;296:1883–6.
26. Beasley NJ, Prevo R, Banerji S, et al. Intratumorallymphangiogenesis and lymph node metastasis in headand neck cancer. Cancer Res 2002;62:1315–20.
27. Partanen TA, Makinen T, Arola J, Suda T, Weich HA,Alitalo K. Endothelial growth factor receptors in humanfetal heart. Circulation 1999;100:583–6.
28. Barleon B, Sozzani S, Zhou D, Weich HA, Mantovani A,Marme D. Migration of human monocytes in response tovascular endothelial growth factor (VEGF) is mediated viathe VEGF receptor flt-1. Blood 1996;87:3336–43.
29. Bingle L, Brown NJ, Lewis CE. The role of tumour-associated macrophages in tumour progression: impli-cations for new anticancer therapies. J Pathol 2002;196:254–65.
30. Pollard JW. Tumour-educated macrophages promotetumour progression and metastasis. Nat Rev Cancer2004;4:71–8.
31. Schoppmann SF, Birner P, Stockl J, et al. Tumor-associated macrophages express lymphatic endothelialgrowth factors and are related to peritumoral lymphan-giogenesis. Am J Pathol 2002;161:947–56.
Cancer Research
Cancer Res 2005; 65: (20). October 15, 2005 9268 www.aacrjournals.org
Research. on May 22, 2014. © 2005 American Association for Cancercancerres.aacrjournals.org Downloaded from
Copyright © 2022 FDOKUMEN











![Detection of Pancreatic Carcinomas by Imaging Lactose-Binding Protein Expression in Peritumoral Pancreas Using [18F]Fluoroethyl-Deoxylactose PET/CT](https://static.fdokumen.com/doc/165x107/631be6bc93f371de19012dfd/detection-of-pancreatic-carcinomas-by-imaging-lactose-binding-protein-expression.jpg)








