
Mouse lung contains endothelial progenitors with high capacity to form blood and lymphatic vessels
Upload
independentCategory
view
1download
0
Thrombospondin-1 selectively inhibits early-stage carcinogenesis and
angiogenesis but not tumor lymphangiogenesis and lymphatic metastasis in
transgenic mice
Thomas Hawighorst1, Hajimu Oura1, Michael Streit1, Lauren Janes1, Lynh Nguyen1,Lawrence F Brown2, Guillermo Oliver3, David G Jackson4 and Michael Detmar*,1
1Cutaneous Biology Research Center and Department of Dermatology, Massachusetts General Hospital and Harvard MedicalSchool, Charlestown, Massachusetts, MA 02129, USA; 2Department of Pathology, Beth Israel Deaconess Medical Center andHarvard Medical School, Boston, Massachusetts, MA 02215, USA; 3Department of Genetics, St. Jude Children’s ResearchHospital, Memphis, Tennessee, TN 38105, USA; 4IMCR Human Immunology Unit, Institute of Molecular Medicine, JohnRadcliffe Hospital, Headington, Oxford OX3 9DS, UK
The roles played by the endogenous angiogenesisinhibitor thrombospondin-1 (TSP-1) in the early stagesof multi-step carcinogenesis and in the control ofhematogenous versus lymphatic metastasis are unknown.To investigate these issues we compared tumor develop-ment in normal mice and in transgenic mice withtargeted overexpression of TSP-1 in the epidermisfollowing a standard two-step chemical skin carcinogen-esis regimen. Overexpression of TSP-1 resulted indelayed and reduced development of premalignantepithelial hyperplasias, but did not inhibit the malignantconversion to squamous cell carcinomas. TSP-1 over-expression also suppressed tumor angiogenesis anddistant organ metastasis, but failed to inhibit tumor-associated lymphangiogenesis or lymphatic tumor spreadto regional lymph nodes. Concomitant with these results,we found that the endothelial TSP-1 receptor CD36 wasmostly absent from cutaneous lymphatic vessels. Ourfindings indicate the potential use of TSP-1 for theprevention of premalignant stages of tumorigenesis andare likely to have implications for the further develop-ment of anti-angiogenic cancer therapies.Oncogene (2002) 21, 7945 – 7956. doi:10.1038/sj.onc.1205956
Keywords: TSP-1; VEGF; Prox1; LYVE-1; chemopre-vention
Introduction
Thrombospondin-1 (TSP-1) is a 450 kD homotrimericmatricellular glycoprotein (for review, see Bornstein,2001; Lawler, 2000) that inhibits proliferation andmigration of vascular endothelial cells in vitro and
inhibits neovascularization in vivo, contributing to thenormal quiescence of the vasculature (Jimenez et al.,2000; Tolsma et al., 1993). Correlative expression datain human malignant melanomas (Zabrenetzky et al.,1994) and in cancers of the breast (Volpert et al.,1995), the cervix (Kodama et al., 2001) and the ovary(Alvarez et al., 2001), together with the results ofxenotransplant tumor studies in mice (Bleuel et al.,1999; Streit et al., 2000; Weinstat et al., 1994) indicatethat increased TSP-1 expression is associated withdecreased malignant tumor growth, invasion and organmetastasis. Conversely, antisense inhibition of TSP-1expression in squamous cell carcinomas (SCC) wasreported to suppress tumor growth in vivo (Castle etal., 1991). Thus, TSP-1 has been described as a tumorsuppressor (Volpert et al., 1995) and as a tumorpromoter (Tuszynski and Nicosia, 1996). Previousexperimental investigations into the importance ofTSP-1 for tumor growth and angiogenesis haveexclusively studied established malignant tumors(Bleuel et al., 1999; Castle et al., 1991; Streit et al.,2000; Weinstat et al., 1994). However, autochthonoushuman tumors usually develop through a series ofpreneoplastic and benign neoplastic stages beforeconverting into malignant cancer, and the impact ofTSP-1 on the early, premalignant, angiogenic stages ofmulti-step carcinogenesis is unknown. Based upon ourrecent findings that TSP-1 mRNA expression wasalready potently downregulated in the early premalig-nant stages of multi-step, chemically-induced skincarcinogenesis (Hawighorst et al., 2001), we hypothe-sized that TSP-1 might play an important role in thecontrol of early carcinogenic events.
Chemically-induced multistage epithelial carcinogen-esis is a well established model for skin cancer in mice(Ito et al., 1995) that allows a detailed evaluation ofdistinct premalignant and malignant stages of carcino-genesis (Marks and Furstenberger, 2000), and that hasalso provided a more in-depth understanding of humanepithelial cancer development (Balmain and Harris,2000; DiGiovanni, 1992; Yuspa, 1994). For manytumors, the initial dissemination occurs via lymphatic
Received 17 June 2002; revised 8 August 2002; accepted 8 August2002
*Correspondence: M Detmar, CBRC/Department of Dermatology,Massachusetts General Hospital, Building 149, 13th Street, Charles-town, MA 02129, USA;E-mail: [email protected]
Oncogene (2002) 21, 7945 – 7956ª 2002 Nature Publishing Group All rights reserved 0950 – 9232/02 $25.00
www.nature.com/onc

vessels to regional lymph nodes, following routes ofnatural tissue drainage (Sleeman, 2000). However, thebiological role of TSP-1 and of other endogenousinhibitors of angiogenesis in tumor lymphangiogenesisand lymphatic metastasis has remained unclear, in partdue to the lack of specific lymphatic markers thatdistinguish lymphatic from blood vascular endothe-lium.
In this present study, we have investigated the distinctbiological role of TSP-1 in multi-step epithelialcarcinogenesis and metastasis. We subjected mice withtargeted overexpression of TSP-1 in the epidermis (Streitet al., 2000) to a standard two-step chemical skincarcinogenesis regimen, using 7,12-dimethylbenz-(a)anthracene (DMBA) for tumor initiation andphorbol 12-myristate 13-acetate (PMA) to promotetumor growth (DiGiovanni, 1992). Here we report thatoverexpression of TSP-1 in epidermal keratinocytesdelayed and diminished the formation of pre-malignantepithelial hyperplasias but did not reduce the rate ofmalignant conversion to squamous cell carcinomas.TSP-1 overexpression was also associated withdecreased tumor angiogenesis and increased tumor cellapoptosis, but failed to inhibit tumor lymphangiogenesisand lymphatic tumor spread to regional lymph nodes.Concomitantly, the major vascular endothelial TSP-1receptor CD36 was only scarcely expressed or wasabsent from cutaneous lymphatic vessels. These resultsreveal a predominant inhibitory effect of TSP-1 on theearly stages of multi-step carcinogenesis, with specificinhibition of blood vascular angiogenesis but not oftumor lymphangiogenesis and lymphatic metastasis,suggesting that TSP-1 might most efficiently be used totarget early premalignant stages of tumorigenesis.
Results
Delayed and diminished skin carcinogenesis in TSP-1transgenic mice
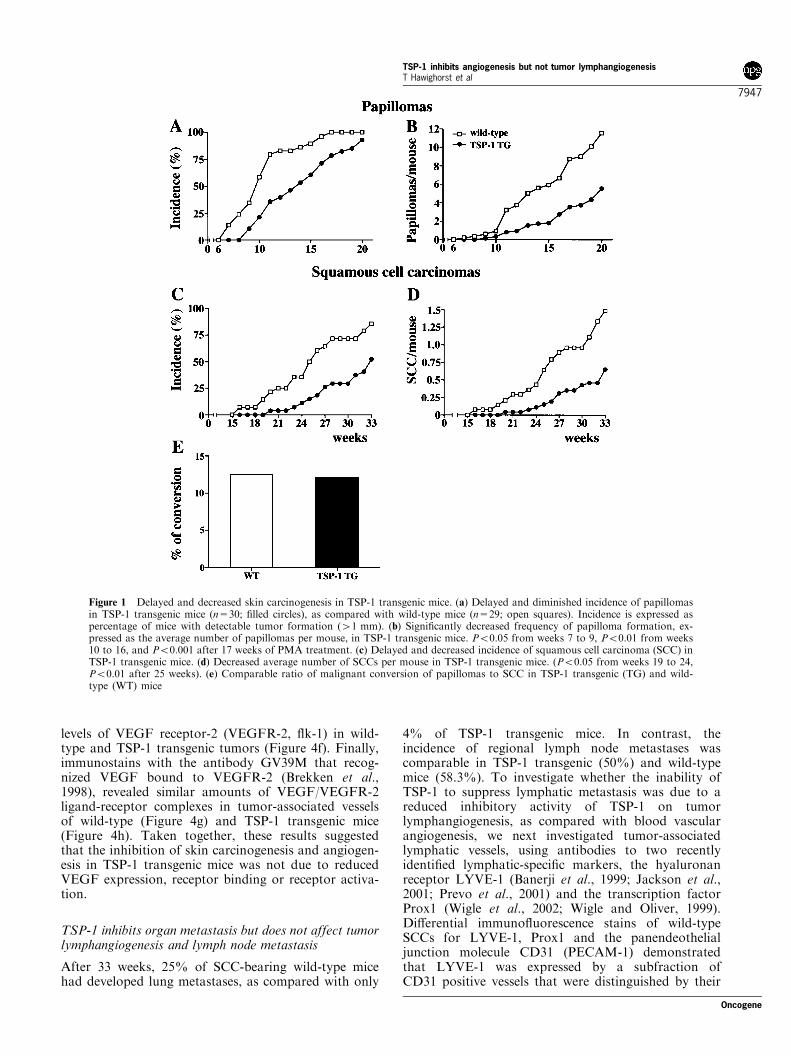
We subjected wild-type and TSP-1 transgenic mice to astandard two-step skin carcinogenesis protocol byusing topical application of a single subcarcinogenicdose of DMBA for initiation, followed by 20 weeklytopical applications of PMA for tumor promotion.TSP-1 transgenic mice showed delayed formation ofpre-malignant skin papillomas, with an average latencyperiod of 14 weeks after the first PMA application, ascompared with 10 weeks in wild-type mice (Figure 1a).At 17 weeks, all of the wild-type mice, but only 79% ofTSP-1 transgenic mice, had developed papillomas.After 20 weeks of PMA promotion, TSP-1 transgenicmice had developed less than six papillomas per mouse,as compared with more than 11 papillomas in wild-type mice (Figure 1b). Malignant conversion ofpapillomas to SCC was first detected at 16 weeks afterTPA promotion in wild-type mice and after 20 weeksin TSP-1 transgenic mice (Figure 1c). After 33 weeks,85% of wild-type mice but only 50% of TSP-1transgenic mice had developed SCC. The averagenumber of SCC per mouse was significantly decreased
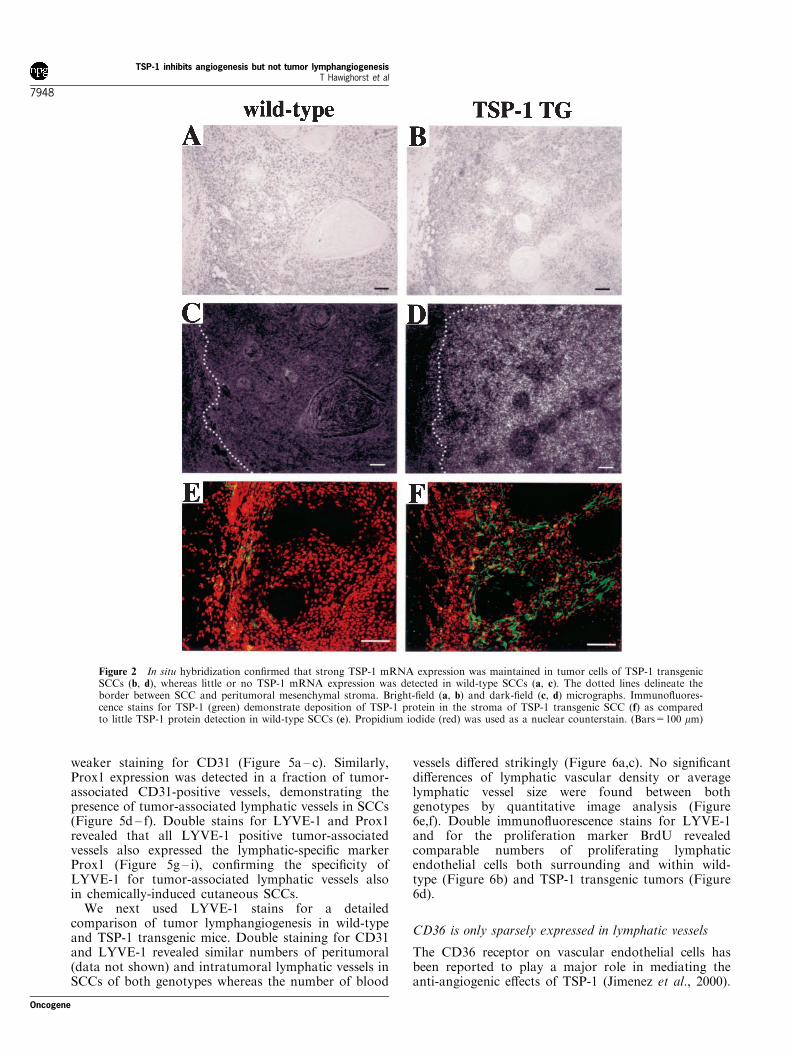
in TSP-1 transgenic mice (0.7+0.15 at week 33;P50.01), as compared with wild-type mice(1.5+0.19; Figure 1d). However, the ratio of malignantconversion of papillomas to SCC was comparable inwild-type (12.5%) and TSP-1 transgenic mice (12.1%;Figure 1e). No tumors were observed in wild-type orTSP-1 transgenic mice treated with DMBA or withPMA alone (data not shown). In situ hybridizationstudies and immunohistochemistry confirmed thattransgenic expression of TSP-1 mRNA and proteinwas maintained throughout the successive steps ofepithelial carcinogenesis (Figure 2). Immunofluores-cence stains for the early and late epidermal terminaldifferentiation markers keratin 10 and loricrin showedcomparable differentiation of normal skin and ofepithelial tumors in both genotypes (data not shown).
Decreased tumor angiogenesis and increased apoptosis inTSP-1 transgenic mice
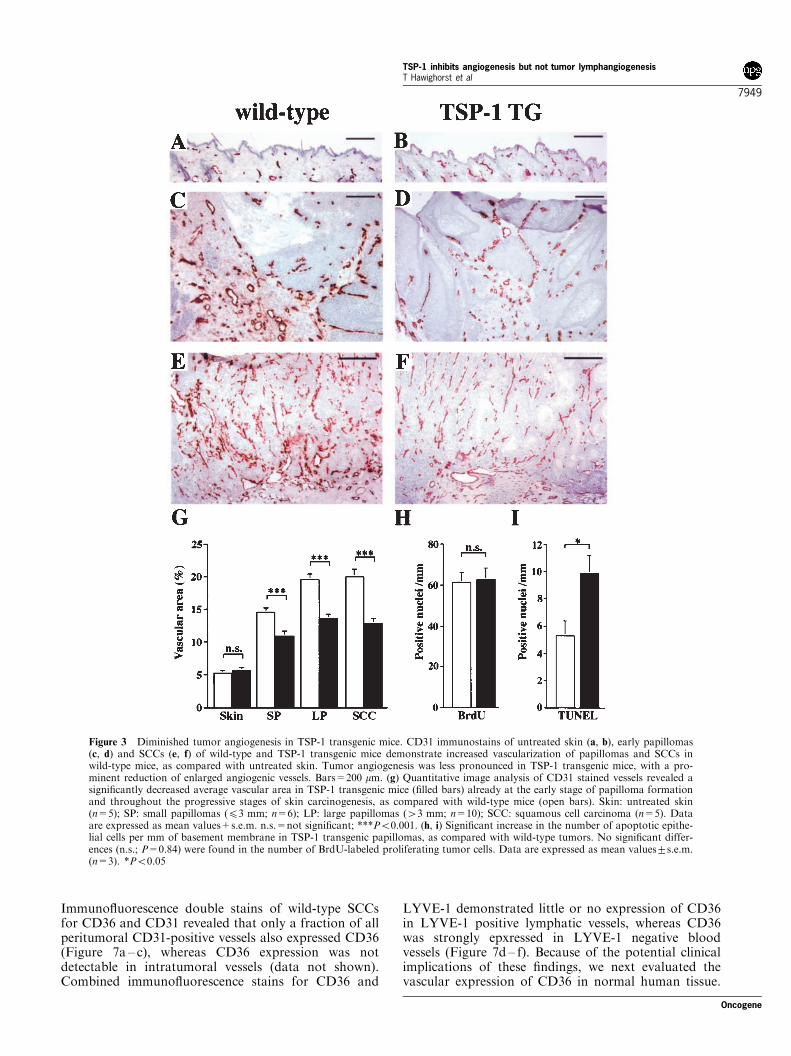
The ‘angiogenic switch’ from vascular quiescence toupregulation of angiogenesis was already detected atthe early stages of epithelial tumorigenesis in wild-typemice, as shown by comparative analysis of CD31-positive vessels in normal skin (Figure 3a), early-stagepapillomas (Figure 3c) and SCC (Figure 3e). Incontrast, the extent of tumor angiogenesis was lesspronounced in TSP-1 transgenic mice (Figure 3b,d,f)which were characterized by reduced numbers of largeangiogenic vessels (3d,f). Computer-assisted morpho-metric image analysis revealed that the relative areaoccupied by CD31-positive vessels was significantlylower (P50.001) in TSP-1 transgenic mice throughoutthe successive stages of skin carcinogenesis (Figure 3g).In agreement with our previous results (Streit et al.,2000), the vascularization of normal skin was compar-able in TSP-1 transgenic mice and wild-type mice(Figure 3a,b,g), suggesting that the reduced angiogenicresponse in early TSP-1 transgenic tumors was not dueto already decreased vascularity in untreated skin. Thenumber of apoptotic tumor cells was more than 1.8-fold (P50.05) increased in TSP-1 transgenic papillo-mas, as compared with wild-type tumors (Figure 3i),whereas no significant differences of tumor cellproliferation were detected (P=0.84; Figure 3h).
TSP-1 does not modulate VEGF expression, receptorbinding or receptor activation during skin carcinogenesis
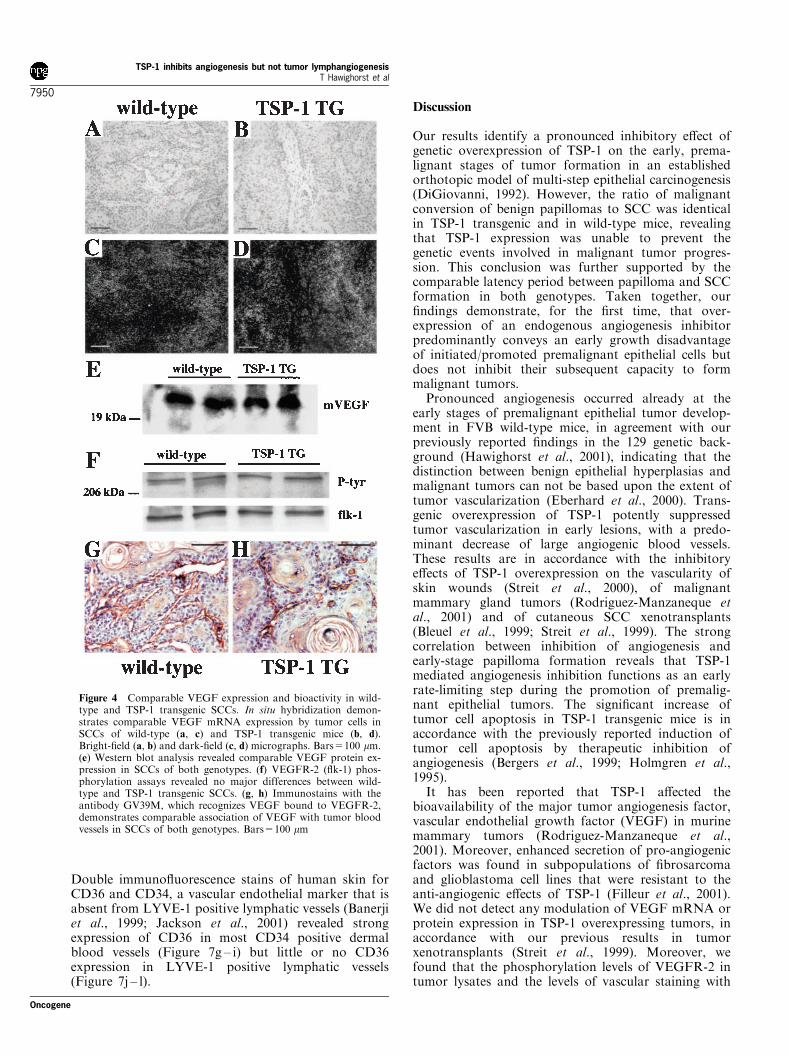
We next investigated whether the TSP-1 mediatedinhibition of vascularization was due to direct orindirect effects of TSP-1 on VEGF expression orbioavailability. In situ hybridization (Figure 4a – d)revealed only minor differences in VEGF mRNAexpression between wild-type (Figure 4a,c) and TSP-1transgenic SCCs (Figure 4b,d), with little or noexpression in the tumor stroma of both genotypes.Moreover, Western blot analyses of SCC lysatesshowed comparable VEGF protein levels in bothgenotypes (Figure 4e), and immunoprecipitationstudies demonstrated comparable phosphorylation
TSP-1 inhibits angiogenesis but not tumor lymphangiogenesisT Hawighorst et al
7946
Oncogene
levels of VEGF receptor-2 (VEGFR-2, flk-1) in wild-type and TSP-1 transgenic tumors (Figure 4f). Finally,immunostains with the antibody GV39M that recog-nized VEGF bound to VEGFR-2 (Brekken et al.,1998), revealed similar amounts of VEGF/VEGFR-2ligand-receptor complexes in tumor-associated vesselsof wild-type (Figure 4g) and TSP-1 transgenic mice(Figure 4h). Taken together, these results suggestedthat the inhibition of skin carcinogenesis and angiogen-esis in TSP-1 transgenic mice was not due to reducedVEGF expression, receptor binding or receptor activa-tion.
TSP-1 inhibits organ metastasis but does not affect tumorlymphangiogenesis and lymph node metastasis
After 33 weeks, 25% of SCC-bearing wild-type micehad developed lung metastases, as compared with only
4% of TSP-1 transgenic mice. In contrast, theincidence of regional lymph node metastases wascomparable in TSP-1 transgenic (50%) and wild-typemice (58.3%). To investigate whether the inability ofTSP-1 to suppress lymphatic metastasis was due to areduced inhibitory activity of TSP-1 on tumorlymphangiogenesis, as compared with blood vascularangiogenesis, we next investigated tumor-associatedlymphatic vessels, using antibodies to two recentlyidentified lymphatic-specific markers, the hyaluronanreceptor LYVE-1 (Banerji et al., 1999; Jackson et al.,2001; Prevo et al., 2001) and the transcription factorProx1 (Wigle et al., 2002; Wigle and Oliver, 1999).Differential immunofluorescence stains of wild-typeSCCs for LYVE-1, Prox1 and the panendeothelialjunction molecule CD31 (PECAM-1) demonstratedthat LYVE-1 was expressed by a subfraction ofCD31 positive vessels that were distinguished by their
Figure 1 Delayed and decreased skin carcinogenesis in TSP-1 transgenic mice. (a) Delayed and diminished incidence of papillomasin TSP-1 transgenic mice (n=30; filled circles), as compared with wild-type mice (n=29; open squares). Incidence is expressed aspercentage of mice with detectable tumor formation (41 mm). (b) Significantly decreased frequency of papilloma formation, ex-pressed as the average number of papillomas per mouse, in TSP-1 transgenic mice. P50.05 from weeks 7 to 9, P50.01 from weeks10 to 16, and P50.001 after 17 weeks of PMA treatment. (c) Delayed and decreased incidence of squamous cell carcinoma (SCC) inTSP-1 transgenic mice. (d) Decreased average number of SCCs per mouse in TSP-1 transgenic mice. (P50.05 from weeks 19 to 24,P50.01 after 25 weeks). (e) Comparable ratio of malignant conversion of papillomas to SCC in TSP-1 transgenic (TG) and wild-type (WT) mice
TSP-1 inhibits angiogenesis but not tumor lymphangiogenesisT Hawighorst et al
7947
Oncogene
weaker staining for CD31 (Figure 5a – c). Similarly,Prox1 expression was detected in a fraction of tumor-associated CD31-positive vessels, demonstrating thepresence of tumor-associated lymphatic vessels in SCCs(Figure 5d – f). Double stains for LYVE-1 and Prox1revealed that all LYVE-1 positive tumor-associatedvessels also expressed the lymphatic-specific markerProx1 (Figure 5g – i), confirming the specificity ofLYVE-1 for tumor-associated lymphatic vessels alsoin chemically-induced cutaneous SCCs.
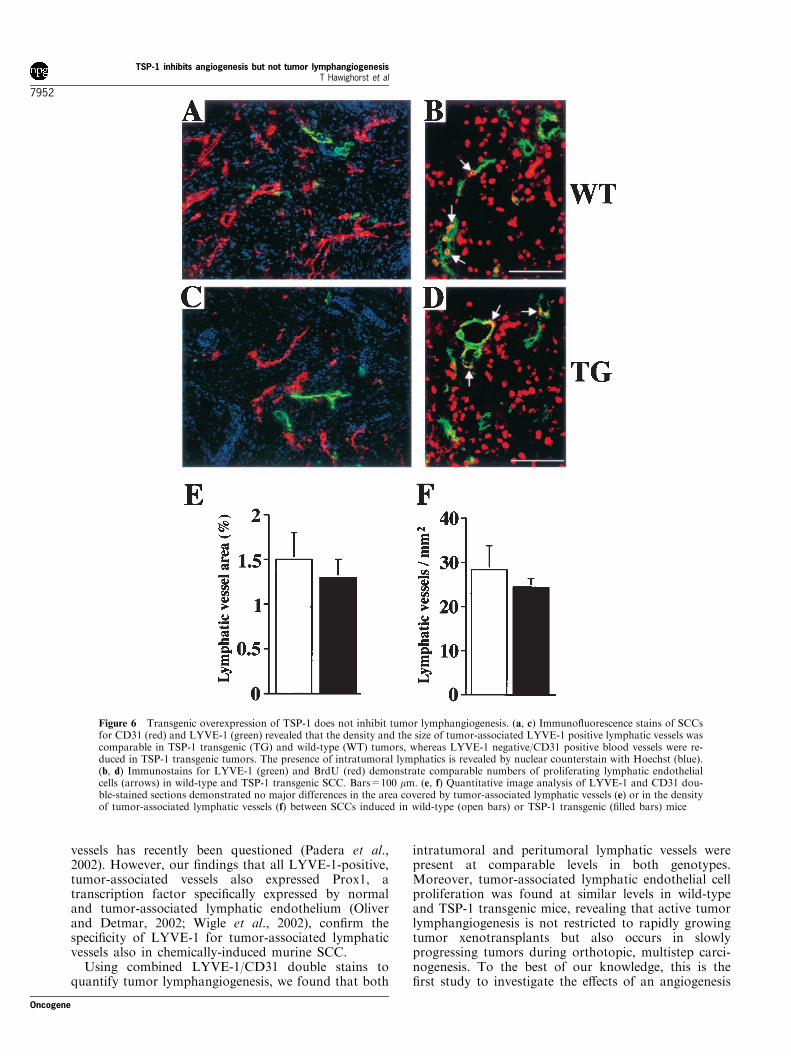
We next used LYVE-1 stains for a detailedcomparison of tumor lymphangiogenesis in wild-typeand TSP-1 transgenic mice. Double staining for CD31and LYVE-1 revealed similar numbers of peritumoral(data not shown) and intratumoral lymphatic vessels inSCCs of both genotypes whereas the number of blood
vessels differed strikingly (Figure 6a,c). No significantdifferences of lymphatic vascular density or averagelymphatic vessel size were found between bothgenotypes by quantitative image analysis (Figure6e,f). Double immunofluorescence stains for LYVE-1and for the proliferation marker BrdU revealedcomparable numbers of proliferating lymphaticendothelial cells both surrounding and within wild-type (Figure 6b) and TSP-1 transgenic tumors (Figure6d).
CD36 is only sparsely expressed in lymphatic vessels
The CD36 receptor on vascular endothelial cells hasbeen reported to play a major role in mediating theanti-angiogenic effects of TSP-1 (Jimenez et al., 2000).
Figure 2 In situ hybridization confirmed that strong TSP-1 mRNA expression was maintained in tumor cells of TSP-1 transgenicSCCs (b, d), whereas little or no TSP-1 mRNA expression was detected in wild-type SCCs (a, c). The dotted lines delineate theborder between SCC and peritumoral mesenchymal stroma. Bright-field (a, b) and dark-field (c, d) micrographs. Immunofluores-cence stains for TSP-1 (green) demonstrate deposition of TSP-1 protein in the stroma of TSP-1 transgenic SCC (f) as comparedto little TSP-1 protein detection in wild-type SCCs (e). Propidium iodide (red) was used as a nuclear counterstain. (Bars=100 mm)
TSP-1 inhibits angiogenesis but not tumor lymphangiogenesisT Hawighorst et al
7948
Oncogene
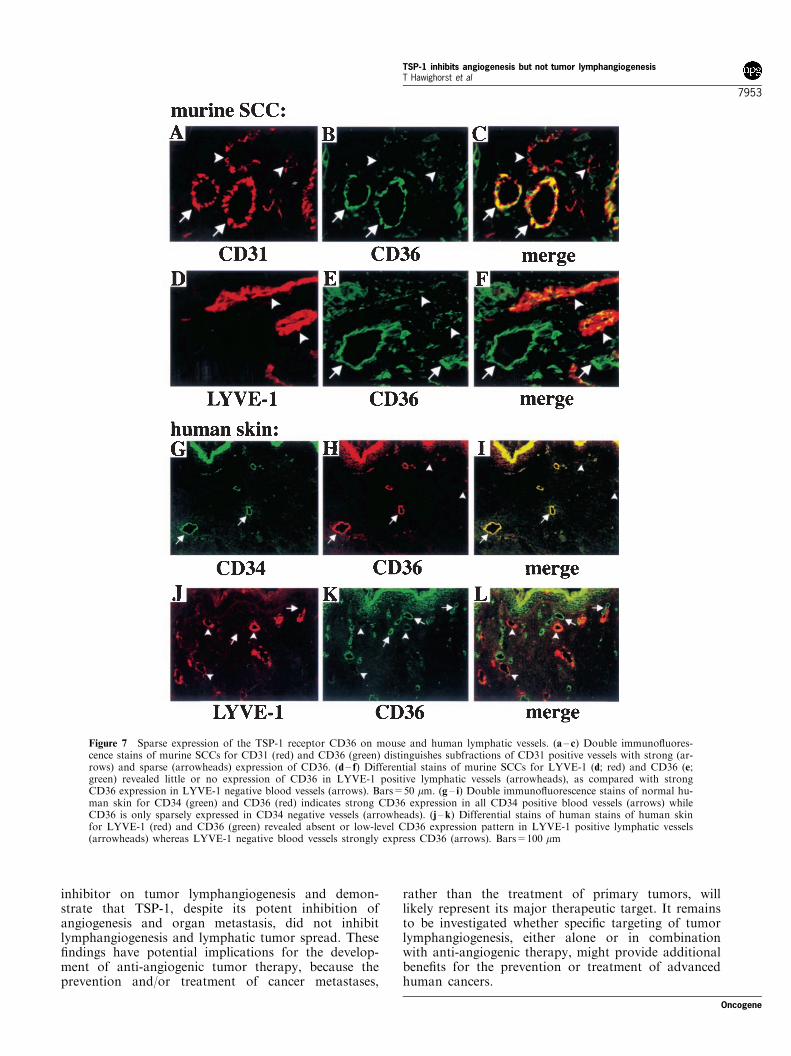
Immunofluorescence double stains of wild-type SCCsfor CD36 and CD31 revealed that only a fraction of allperitumoral CD31-positive vessels also expressed CD36(Figure 7a – c), whereas CD36 expression was notdetectable in intratumoral vessels (data not shown).Combined immunofluorescence stains for CD36 and
LYVE-1 demonstrated little or no expression of CD36in LYVE-1 positive lymphatic vessels, whereas CD36was strongly epxressed in LYVE-1 negative bloodvessels (Figure 7d – f). Because of the potential clinicalimplications of these findings, we next evaluated thevascular expression of CD36 in normal human tissue.
Figure 3 Diminished tumor angiogenesis in TSP-1 transgenic mice. CD31 immunostains of untreated skin (a, b), early papillomas(c, d) and SCCs (e, f) of wild-type and TSP-1 transgenic mice demonstrate increased vascularization of papillomas and SCCs inwild-type mice, as compared with untreated skin. Tumor angiogenesis was less pronounced in TSP-1 transgenic mice, with a pro-minent reduction of enlarged angiogenic vessels. Bars=200 mm. (g) Quantitative image analysis of CD31 stained vessels revealed asignificantly decreased average vascular area in TSP-1 transgenic mice (filled bars) already at the early stage of papilloma formationand throughout the progressive stages of skin carcinogenesis, as compared with wild-type mice (open bars). Skin: untreated skin(n=5); SP: small papillomas (43 mm; n=6); LP: large papillomas (43 mm; n=10); SCC: squamous cell carcinoma (n=5). Dataare expressed as mean values+s.e.m. n.s.=not significant; ***P50.001. (h, i) Significant increase in the number of apoptotic epithe-lial cells per mm of basement membrane in TSP-1 transgenic papillomas, as compared with wild-type tumors. No significant differ-ences (n.s.; P=0.84) were found in the number of BrdU-labeled proliferating tumor cells. Data are expressed as mean values+s.e.m.(n=3). *P50.05
TSP-1 inhibits angiogenesis but not tumor lymphangiogenesisT Hawighorst et al
7949
Oncogene
Double immunofluorescence stains of human skin forCD36 and CD34, a vascular endothelial marker that isabsent from LYVE-1 positive lymphatic vessels (Banerjiet al., 1999; Jackson et al., 2001) revealed strongexpression of CD36 in most CD34 positive dermalblood vessels (Figure 7g – i) but little or no CD36expression in LYVE-1 positive lymphatic vessels(Figure 7j – l).
Discussion
Our results identify a pronounced inhibitory effect ofgenetic overexpression of TSP-1 on the early, prema-lignant stages of tumor formation in an establishedorthotopic model of multi-step epithelial carcinogenesis(DiGiovanni, 1992). However, the ratio of malignantconversion of benign papillomas to SCC was identicalin TSP-1 transgenic and in wild-type mice, revealingthat TSP-1 expression was unable to prevent thegenetic events involved in malignant tumor progres-sion. This conclusion was further supported by thecomparable latency period between papilloma and SCCformation in both genotypes. Taken together, ourfindings demonstrate, for the first time, that over-expression of an endogenous angiogenesis inhibitorpredominantly conveys an early growth disadvantageof initiated/promoted premalignant epithelial cells butdoes not inhibit their subsequent capacity to formmalignant tumors.
Pronounced angiogenesis occurred already at theearly stages of premalignant epithelial tumor develop-ment in FVB wild-type mice, in agreement with ourpreviously reported findings in the 129 genetic back-ground (Hawighorst et al., 2001), indicating that thedistinction between benign epithelial hyperplasias andmalignant tumors can not be based upon the extent oftumor vascularization (Eberhard et al., 2000). Trans-genic overexpression of TSP-1 potently suppressedtumor vascularization in early lesions, with a predo-minant decrease of large angiogenic blood vessels.These results are in accordance with the inhibitoryeffects of TSP-1 overexpression on the vascularity ofskin wounds (Streit et al., 2000), of malignantmammary gland tumors (Rodriguez-Manzaneque etal., 2001) and of cutaneous SCC xenotransplants(Bleuel et al., 1999; Streit et al., 1999). The strongcorrelation between inhibition of angiogenesis andearly-stage papilloma formation reveals that TSP-1mediated angiogenesis inhibition functions as an earlyrate-limiting step during the promotion of premalig-nant epithelial tumors. The significant increase oftumor cell apoptosis in TSP-1 transgenic mice is inaccordance with the previously reported induction oftumor cell apoptosis by therapeutic inhibition ofangiogenesis (Bergers et al., 1999; Holmgren et al.,1995).
It has been reported that TSP-1 affected thebioavailability of the major tumor angiogenesis factor,vascular endothelial growth factor (VEGF) in murinemammary tumors (Rodriguez-Manzaneque et al.,2001). Moreover, enhanced secretion of pro-angiogenicfactors was found in subpopulations of fibrosarcomaand glioblastoma cell lines that were resistant to theanti-angiogenic effects of TSP-1 (Filleur et al., 2001).We did not detect any modulation of VEGF mRNA orprotein expression in TSP-1 overexpressing tumors, inaccordance with our previous results in tumorxenotransplants (Streit et al., 1999). Moreover, wefound that the phosphorylation levels of VEGFR-2 intumor lysates and the levels of vascular staining with
Figure 4 Comparable VEGF expression and bioactivity in wild-type and TSP-1 transgenic SCCs. In situ hybridization demon-strates comparable VEGF mRNA expression by tumor cells inSCCs of wild-type (a, c) and TSP-1 transgenic mice (b, d).Bright-field (a, b) and dark-field (c, d) micrographs. Bars=100 mm.(e) Western blot analysis revealed comparable VEGF protein ex-pression in SCCs of both genotypes. (f) VEGFR-2 (flk-1) phos-phorylation assays revealed no major differences between wild-type and TSP-1 transgenic SCCs. (g, h) Immunostains with theantibody GV39M, which recognizes VEGF bound to VEGFR-2,demonstrates comparable association of VEGF with tumor bloodvessels in SCCs of both genotypes. Bars=100 mm
TSP-1 inhibits angiogenesis but not tumor lymphangiogenesisT Hawighorst et al
7950
Oncogene

the antibody GV39M, that recognizes VEGF bound toVEGFR-2 (Brekken et al., 1998), were comparable inSCCs induced in wild-type and TSP-1 transgenic mice.These results suggest that the inhibition of tumorvascularization during multi-step epithelial carcinogen-esis by transgenic TSP-1 was not mediated byinhibitory effects on VEGF expression, bioactivity, orbioavailability.
The metastatic spread of tumor cells is responsiblefor the majority of cancer-related deaths. Continuoustransgenic overexpression of TSP-1 during multi-stepepithelial carcinogenesis resulted in a reduced incidenceof SCC metastasis to the lungs, associated with reducedvascularity of the primary tumors. These results are inagreement with the previously reported inhibition ofspontaneous pulmonary metastases by stable over-expression of TSP-1 in MDA-MB-435 breast cancerxenotransplants (Weinstat et al., 1994), and theysuggest that the rate of hematogenous metastasis wasreduced because of the pronounced inhibitory effect ofTSP-1 on blood vascular tumor angiogenesis.
It has remained unclear whether angiogenesisinhibitors currently developed for clinical therapy
might also affect lymphangiogenesis, the growth oflymphatic vessels. Lymphatic tumor spread to regionallymph nodes represents the most common pathway ofinitial tumor dissemination, and the detection oflymphatic metastasis serves as an important prognosticindicator of tumor aggressiveness and frequentlydetermines the choice of anti-tumoral therapy. Incontrast to the reduced incidence of lung metastases,we did not detect any difference in the incidence oflymphatic SCC metastasis to regional lymph nodesbetween TSP-1 transgenic and wild-type mice. Becauserecent experimental studies have revealed a correlationbetween tumor lymphangiogenesis and lymphaticmetastasis (Karpanen et al., 2001; Mandriota et al.,2001; Skobe et al., 2001; Stacker et al., 2001), we nextinvestigated the density and size of tumor-associatedlymphatic vessels by using an antibody against themurine hyaluronan receptor LYVE-1 (Jackson et al.,2001) that specifically stains lymphatic vessels, but notblood vessels, in experimental tumor xenotransplants inmice (Skobe et al., 2001; Wigle et al., 2002) and inhuman tumors (Beasley et al., 2002). The specificity ofthe LYVE-1 antibody for tumor-associated lymphatic
Figure 5 LYVE-1 is a specific marker for tumor-associated lymphatic vessels in multistage skin carcinogenesis. (a – c) Double im-munofluorescence stains of chemically-induced SCCs for the endothelial junction molecule CD31 (red) and LYVE-1 (green) demon-strated that only a fraction of all CD31-positive vessels, characterized by low-level expression of CD31, expressed LYVE-1 (arrows).(d – f) Differential stains of SCCs for the lymphatic-specific transcription factor Prox1 (green) and CD31 (red) revealed Prox1 po-sitive tumor-associated lymphatic vessels (arrows), characterized by low-level CD31 expression. (g – i) Double immunofluorescencestains of SCCs for LYVE-1 (red) and Prox1 (green) revealed that all LYVE-1 positive lymphatic vessels also expressed Prox1(arrows)
TSP-1 inhibits angiogenesis but not tumor lymphangiogenesisT Hawighorst et al
7951
Oncogene
vessels has recently been questioned (Padera et al.,2002). However, our findings that all LYVE-1-positive,tumor-associated vessels also expressed Prox1, atranscription factor specifically expressed by normaland tumor-associated lymphatic endothelium (Oliverand Detmar, 2002; Wigle et al., 2002), confirm thespecificity of LYVE-1 for tumor-associated lymphaticvessels also in chemically-induced murine SCC.
Using combined LYVE-1/CD31 double stains toquantify tumor lymphangiogenesis, we found that both
intratumoral and peritumoral lymphatic vessels werepresent at comparable levels in both genotypes.Moreover, tumor-associated lymphatic endothelial cellproliferation was found at similar levels in wild-typeand TSP-1 transgenic mice, revealing that active tumorlymphangiogenesis is not restricted to rapidly growingtumor xenotransplants but also occurs in slowlyprogressing tumors during orthotopic, multistep carci-nogenesis. To the best of our knowledge, this is thefirst study to investigate the effects of an angiogenesis
Figure 6 Transgenic overexpression of TSP-1 does not inhibit tumor lymphangiogenesis. (a, c) Immunofluorescence stains of SCCsfor CD31 (red) and LYVE-1 (green) revealed that the density and the size of tumor-associated LYVE-1 positive lymphatic vessels wascomparable in TSP-1 transgenic (TG) and wild-type (WT) tumors, whereas LYVE-1 negative/CD31 positive blood vessels were re-duced in TSP-1 transgenic tumors. The presence of intratumoral lymphatics is revealed by nuclear counterstain with Hoechst (blue).(b, d) Immunostains for LYVE-1 (green) and BrdU (red) demonstrate comparable numbers of proliferating lymphatic endothelialcells (arrows) in wild-type and TSP-1 transgenic SCC. Bars=100 mm. (e, f) Quantitative image analysis of LYVE-1 and CD31 dou-ble-stained sections demonstrated no major differences in the area covered by tumor-associated lymphatic vessels (e) or in the densityof tumor-associated lymphatic vessels (f) between SCCs induced in wild-type (open bars) or TSP-1 transgenic (filled bars) mice
TSP-1 inhibits angiogenesis but not tumor lymphangiogenesisT Hawighorst et al
7952
Oncogene
inhibitor on tumor lymphangiogenesis and demon-strate that TSP-1, despite its potent inhibition ofangiogenesis and organ metastasis, did not inhibitlymphangiogenesis and lymphatic tumor spread. Thesefindings have potential implications for the develop-ment of anti-angiogenic tumor therapy, because theprevention and/or treatment of cancer metastases,
rather than the treatment of primary tumors, willlikely represent its major therapeutic target. It remainsto be investigated whether specific targeting of tumorlymphangiogenesis, either alone or in combinationwith anti-angiogenic therapy, might provide additionalbenefits for the prevention or treatment of advancedhuman cancers.
Figure 7 Sparse expression of the TSP-1 receptor CD36 on mouse and human lymphatic vessels. (a – c) Double immunofluores-cence stains of murine SCCs for CD31 (red) and CD36 (green) distinguishes subfractions of CD31 positive vessels with strong (ar-rows) and sparse (arrowheads) expression of CD36. (d – f) Differential stains of murine SCCs for LYVE-1 (d; red) and CD36 (e;green) revealed little or no expression of CD36 in LYVE-1 positive lymphatic vessels (arrowheads), as compared with strongCD36 expression in LYVE-1 negative blood vessels (arrows). Bars=50 mm. (g – i) Double immunofluorescence stains of normal hu-man skin for CD34 (green) and CD36 (red) indicates strong CD36 expression in all CD34 positive blood vessels (arrows) whileCD36 is only sparsely expressed in CD34 negative vessels (arrowheads). (j –k) Differential stains of human stains of human skinfor LYVE-1 (red) and CD36 (green) revealed absent or low-level CD36 expression pattern in LYVE-1 positive lymphatic vessels(arrowheads) whereas LYVE-1 negative blood vessels strongly express CD36 (arrows). Bars=100 mm
TSP-1 inhibits angiogenesis but not tumor lymphangiogenesisT Hawighorst et al
7953
Oncogene

TSP-1 is thought to inhibit angiogenesis by anumber of different mechanisms whose relative impor-tance for different types of angiogenesis in benign ormalignant conditions remain unclear (reviewed inBornstein, 2001; Lawler, 2000). However, a recentstudy in CD36-deficient mice has provided strongevidence for an important role of the CD36 receptoron vascular endothelial cells for mediating the in vivoanti-angiogenic effects of TSP-1 (Jimenez et al., 2000).Using differential immunostains of murine SCC and ofhuman skin for CD36, the lymphatic LYVE-1 and theblood vascular marker CD34 (Oliver and Detmar,2002; Prevo et al., 2001), our study revealed little or noexpression of CD36 in lymphatic vessels as comparedwith its strong expression in blood vessels, suggestingthat different CD36 expression levels contribute to thedifferential effects of TSP-1 on angiogenesis versuslymphangiogenesis.
In summary, our studies provide evidence that TSP-1mediated angiogenesis inhibition during multistepcarcinogenesis predominantly conveys an early growthdisadvantage of initiated/promoted premalignantepithelial cells but does not inhibit the subsequentdevelopment of malignant tumors, indicating thatTSP-1 may prove efficacious in the chemoprevention ofthe early premalignant stages of epithelial tumorigenesis.Despite its potent suppression of blood vascularangiogenesis and distant organ metastasis, however,TSP-1 was unable to inhibit tumor lymphangiogenesisand lymphatic metastasis, with potential implications forthe development of clinical anti-angiogenic therapies.
Materials and methods
Chemical skin carcinogenesis protocol
The construction of the keratin 14 (K14) promoter/TSP-1transgene vector and the generation of transgenic FVB micewith targeted overexpression of TSP-1 in the skin have beenpreviously described (Streit et al., 2000). Chemically-inducedskin carcinogenesis was performed in heterozygous TSP-1transgenic mice (n=30) and their wild-type littermates(n=29) as previously described (Hawighorst et al., 2001).The ratio of malignant conversion was calculated as the totalnumber of carcinomas divided by the total number ofpapillomas, expressed as a percentage. At autopsy, allaxillary and inguinal lymph nodes as well as the lungs ofSCC-bearing mice were examined for the presence ofmetastases. The two-sided unpaired Student’s t-test was usedto analyse differences in the number of tumors per mousebetween the two genotypes. In vivo metastasis data wereevaluated using the Mann-Whiney test and the AlternateWelch test. All animal studies were approved by theMassachusetts General Hospital Subcommittee on ResearchAnimal Care.
Histology, immunohistochemistry and in situ hybridization
Hematoxylin and eosin-stained sections were prepared fromlymph nodes and from lungs at three random depths. Theslides were evaluated using a Nikon E-600 microscope(Nikon, Melville, NY, USA), and mice were designatedpositive for the presence of metastasis when at least one
colony of tumor cells was detected in the draining lymphnodes or lungs as previously described (Leone et al., 1993).Immunohistochemical staining was performed on 6 mmparaffin sections or cryostat sections as previously described(Hawighorst et al., 2001), using a rat monoclonal antibodyagainst CD31 (Pharmingen, San Diego, CA, USA) and theantibody GV39M (kindly provided by P Thorpe, Dallas, TX,USA and R Brekken, Seattle, WA, USA) that recognizesVEGF bound to VEGFR-2. For immunofluorescence singleand double stains, 6 mm frozen sections were stained withantibodies to human TSP-1 (kindly provided by R Swerlick,Atlanta, GA, USA), murine and human LYVE-1 (Prevo etal., 2001), Prox1 (Wigle et al., 2002), mouse keratin 10 andloricrin (Babco, Richmond, CA, USA), mouse CD31(Pharmingen), human CD34 (Pharmingen), human CD36(Immunotech, Marseille, France), and mouse CD36 (kindlyprovided by R Silverstein, New York NY, USA), and withcorresponding secondary antibodies labeled with AlexaFluor488 or 594 (Molecular Probes, Eugene, Oregon, OR,USA). In situ hybridization for human TSP-1 and murineVEGF was performed as described (Detmar et al., 1998;Streit et al., 2000). The VEGF riboprobe used recognizes allisoforms of mouse VEGF mRNA.
Computer-assisted morphometric vessel analysis
Representative CD31-stained frozen sections, obtained frombiopsies of normal untreated skin (n=5), small papillomas(1 – 3 mm in diameter; n=6), large papillomas (43 mm indiameter; n=10), and squamous cell carcinomas (SCC; n=5)were analysed as described (Hawighorst et al., 2001). Forevaluation of lymphatic vessels, frozen sections of fivedifferent representative SCCs per group were double-stainedwith antibodies to LYVE-1 and CD31. Lymphatic vesselswere quantified in tumor areas with the highest density oflymphatic vessels, using the IP lab software (Scanalytics).Statistical analysis was performed using the two-sidedunpaired student’s t-test.
Western blot analyses
Six different wild-type and TSP-1 transgenic SCCs wereanalysed, and three tumors each were pooled and homo-genized in lysis buffer (2% SDS, 50 mM Tris pH 7.4, 20 mM
phenylmethylsulfonyl fluoride, 50 mg/ml leupeptin and 50 mg/ml aproptinin). Samples were analysed by denaturing SDS –PAGE and were immunoblotted with an antibody againstmouse VEGF (R&D Systems Inc., Minneapolis, MN, USA).Immunoreactive proteins were visualized using a chemilumi-nescense detection system (ECL; Amersham ArlingtonHeights, IL, USA). Levels of VEGF protein expression werequantified by densitometry.
Receptor phosphorylation assays
In vivo receptor phosphorylation assays for flk-1 wereperformed as recently described (Hawighorst et al., 2002).Briefly, six different wild-type and TSP-1 transgenic SCCswere homogenized in RIPA lysis buffer containing proteaseinhibitors, followed by immunoprecipitation with polyclonalantibody against mouse VEGFR-2 (flk-1; Santa Cruz).Immunocomplexes were recovered on protein-G-sepharoseseparated by 7.5% SDS–PAGE, and blotted onto anitrocellulose membrane (Bio-Rad). Flk-1 phosphorylationwas analysed by probing the membranes with a monoclonalanti-phosphotyrosine antibody (PY-20; ICN Biomedicals,Inc., Aurora, OH, USA). The same membranes were then
TSP-1 inhibits angiogenesis but not tumor lymphangiogenesisT Hawighorst et al
7954
Oncogene

stripped using TBST (10 mM Tris-HCl, 150 mM NaCl, 0.1%Tween 20, pH 2.4) and were probed for VEGFR-2 (flk-1;Santa Cruz). Immunoreactive proteins were visualized usingthe ECL Chemiluminescence detection system. Levels ofVEGFR-2 phosphorylation were quantified by densitometryand were normalized to the expression levels of VEGFR-2.
Cell proliferation and apoptosis assays
Large papillomas of similar size were obtained from TSP-1transgenic (n=3) and wild-type (n=3) mice. Proliferatingtumor cells were visualized by double immunofluorescencestains with an FITC-conjugated anti-BrdU antibody (Phar-mingen) and a rabbit antibody to mouse keratin K14, andwere counted in an area overlying a total length of 21.3 mmof basement membrane in TSP-1 transgenic mice and of25.4 mm in wild-type mice as described (Hawighorst et al.,2001). Apoptotic cells were identified by TUNEL stainingusing the Fluorescein-FragEL DNA fragmentation kit(Oncogene, Cambridge, MA, USA) according to themanufacturer’s instructions. Nuclei were stained with propi-dium iodide in PBS (2 mg/ml). Labeled tumor cells werecounted in an area overlying a total length of 43.2 mm ofbasement membrane in TSP-1 transgenic mice and of
39.4 mm in wild-type mice. Results are expressed as themean+s.e.m. of BrdU- or TUNEL-positive nuclei per mmbasement membrane. The two-sided unpaired Student’s t-testwas used to analyse differences of proliferation and apoptosisrates.
Acknowledgments
The authors thank R Silverstein, P Thorpe, R Brekken andR Swerlick for the generous gift of antibodies, M Mihmand S Dadras for advice on mouse skin pathology, J Nagyfor statistical advice and M Constant for technicalassistance. This work was supported by NIH/NCI grantsCA69184, CA86410 and CA91861 (M Detmar), by NIHgrants GM58462 and EY12162 (G Oliver), by AmericanCancer Society Program Project Grant 99-23901 (MDetmar), by the Deutsche Forschungsgemeinschaft (THawighorst), by Deutscher Akademischer Austauschdienst(M Streit), by the Dermatology Foundation (M Streit), bythe American Lebanese Syrian Associated Charities (GOliver) and by the Cutaneous Biology Research Centerthrough the Massachusetts General Hospital/Shiseido Co.Ltd., Agreement (M Detmar).
References
Alvarez AA, Axelrod JR, Whitaker RS, Isner PD, BentleyRC, Dodge RK and Rodriguez GC. (2001). Gynecol.,Oncol., 82, 273 – 278.
Balmain A and Harris CC. (2000). Carcinogenesis, 21, 371 –377.
Banerji S, Ni J, Wang SX, Clasper S, Su J, Tammi R, JonesM and Jackson DG. (1999). J. Cell Biol., 144, 789 – 801.
Beasley NJ, Prevo R, Banerji S, Leek RD, Moore J, vanTrappen P, Cox G, Harris AL and Jackson DG. (2002).Cancer Res., 62, 1315 – 1320.
Bergers G, Javaherian K, Lo KM, Folkman J and HanahanD. (1999). Science, 284, 808 – 812.
Bleuel K, Popp S, Fusenig NE, Stanbridge EJ and BoukampP. (1999). Proc. Natl. Acad. Sci. USA, 96, 2065 – 2070.
Bornstein P. (2001). J. Clin. Invest., 107, 929 – 934.Brekken RA, Huang X, King SW and Thorpe PE. (1998).Cancer Res., 58, 1952 – 1959.
Castle V, Varani J, Fligiel S, Prochownik EV and Dixit V.(1991). J. Clin. Invest., 87, 1883 – 1888.
Detmar M, Brown LF, Schon MP, Elicker BM, Velasco P,Richard L, Fukumura D, Monsky W, Claffey KP and JainRK. (1998). J. Invest. Dermatol., 111, 1 – 6.
DiGiovanni J. (1992). Pharmacol. Ther., 54, 63 – 128.Eberhard A, Kahlert S, Goede V, Hemmerlein B, Plate KHand Augustin HG. (2000). Cancer Res., 60, 1388 – 1393.
Filleur S, Volpert OV, Degeorges A, Voland C, Reiher F,Clezardin P, Bouck N and Cabon F. (2001). Genes Dev.,15, 1373 – 1382.
Hawighorst T, Skobe M, Streit M, Hong YK, Velasco P,Brown LF, Riccardi L, Lange-Asschenfeldt D and DetmarM. (2002). Am. J. Pathol., 160, 1381 – 1392.
Hawighorst T, Velasco P, Streit M, Hong YK, KyriakidesTR, Brown LF, Bornstein P and Detmar M. (2001).EMBO J., 20, 2631 – 2640.
Holmgren L, O’Reilly MS and Folkman J. (1995).Nat. Med.,1, 149 – 153.
Ito N, Hasegawa R, Imaida K, Hirose M, Asamoto M andShirai T. (1995). Crit. Rev. Oncol. Hematol., 21, 105 – 133.
Jackson DG, Prevo R, Clasper S and Banerji S. (2001).Trends Immunol., 22, 317 – 321.
Jimenez B, Volpert OV, Crawford SE, Febbraio M,Silverstein RL and Bouck N. (2000). Nature Med., 6,
41 – 48.Karpanen T, Egeblad M, Karkkainen MJ, Kubo H, Yla-Herttuala S, Jaattela M and Alitalo K. (2001). Cancer Res.,61, 1786 – 1790.
Kodama J, Hashimoto I, Seki N, Hongo A, Yoshinouchi M,Okuda H and Kudo T. (2001). Clin. Cancer Res., 7, 2826 –2831.
Lawler J. (2000). Curr. Opin. Cell Biol., 12, 634 – 640.Leone A, Flatow U, VanHoutte K and Steeg PS. (1993).Oncogene, 8, 2325 – 2333.
Mandriota SJ, Jussila L, Jeltsch M, Compagni A, Baetens D,Prevo R, Banerji S, Huarte J, Montesano R, Jackson DG,Orci L, Alitalo K, Christofori G and Pepper MS. (2001).EMBO J., 20, 672 – 862.
Marks F and Furstenberger G. (2000). Eur. J. Cancer., 36,314 – 329.
Oliver G and Detmar M. (2002). Genes Dev., 16, 773 – 783.Padera TP, Kadambi A, di Tomaso E, Carreira CM, BrownEB, Bouscher Y, Choi NC, Mathisen D, Wain J, Mark EJ,Munn LL and Jain RK. (2002). Science, 296, 1883 – 1886.
Prevo R, Banerji S, Ferguson DJ, Clasper S and Jackson DG.(2001). J. Biol. Chem., 276, 19420 – 19430.
Rodriguez-Manzaneque JC, Lane TF, Ortega MA, HynesRO, Lawler J and Iruela-Arispe ML. (2001). Proc. Natl.Acad. Sci. USA, 98, 12485 – 12490.
Skobe M, Hawighorst T, Jackson DG, Prevo R, Janes L,Velasco P, Riccardi L, Alitalo K, Claffey K and DetmarM. (2001). Nat. Med., 7, 192 – 198.
Sleeman JP. (2000). Recent Results Cancer Res., 157, 55 – 81.Stacker SA, Caesar C, Baldwin ME, Thornton GE, WilliamsRA, Prevo R, Jackson DG, Nishikawa S, Kubo H andAchen MG. (2001). Nat. Med., 7, 186 – 191.
TSP-1 inhibits angiogenesis but not tumor lymphangiogenesisT Hawighorst et al
7955
Oncogene

Streit M, Velasco P, Brown LF, Skobe M, Richard L,Riccardi L, Lawler J and Detmar M. (1999). Am. J. Pathol,155, 441 – 452.
Streit M, Velasco P, Riccardi L, Spencer L, Brown LF, JanesL, Lange-Asschenfeldt B, Yano K, Hawighorst T, Iruela-Arispe L and Detmar M. (2000). EMBO J., 19, 3272 –3282.
Tolsma SS, Volpert OV, Good DJ, Frazier WA, Polverini PJand Bouck N. (1993). J. Cell Biol., 122, 497 – 511.
Tuszynski GP and Nicosia RF. (1996). Bioessays, 18, 71 – 76.Volpert OV, Stellmach V and Bouck N. (1995). BreastCancer Res. Treat., 36, 119 – 126.
Weinstat SD, Zabrenetzky VS, VanHoutte K, Frazier WA,Roberts DD and Steeg PS. (1994). Cancer Res., 54, 6504 –6511.
Wigle JT, Harvey N, Detmar M, Lagutina I, Grosveld G,Gunn MD, Jackson DG and Oliver G. (2002). EMBO J.,21, 1505 – 1513.
Wigle JT and Oliver G. (1999). Cell, 98, 769 – 778.Yuspa SH. (1994). Cancer Res., 54, 1178 – 1189.Zabrenetzky V, Harris CC, Steeg PS and Roberts DD.(1994). Int. J. Cancer, 59, 191 – 195.
TSP-1 inhibits angiogenesis but not tumor lymphangiogenesisT Hawighorst et al
7956
Oncogene
Copyright © 2022 FDOKUMEN




















