
Age-associated stresses induce an anti-inflammatory senescent phenotype in endothelial cells
Upload
independentCategory
view
0download
0
Downloaded from www.microbiologyresearch.org by
IP: 54.90.14.242
On: Mon, 03 Oct 2016 07:50:14
Peripheral blood mononuclear cells increase thepermeability of dengue virus-infected endothelialcells in association with downregulation of vascularendothelial cadherin
Beti Ernawati Dewi,1,2,3 Tomohiko Takasaki1 and Ichiro Kurane1,3
Correspondence
Ichiro Kurane
1Laboratory of Vector-Borne Viruses, Department of Virology 1, National Institute of InfectiousDiseases, 1-23-1 Toyama, Shinjuku-ku, Tokyo 162-8640, Japan
2Department of Microbiology, Medical Faculty, University of Indonesia, Jalan Pegangsaan Timur no.16, Jakarta 10320, Indonesia
3Department of Infection Biology, Institute of Basic Medical Sciences, University of Tsukuba,Tsukuba 305-8575, Japan
Received 6 August 2007
Accepted 30 October 2007
Plasma leakage is one of the characteristic features of dengue haemorrhagic fever. The interaction
among peripheral blood mononuclear cells (PBMCs), dengue virus and endothelial cells was
analysed in vitro. Human umbilical vein endothelial cells (HUVECs) were infected with dengue-2
virus (DV-2) at an m.o.i. of 0.5 p.f.u. per cell. PBMCs were added to DV-2-infected HUVECs, and
transendothelial electrical resistance (TEER) and transalbumin permeability were assessed.
Dengue virus infection at an m.o.i. of 0.5 p.f.u. per cell alone did not decrease the TEER, but
addition of PBMCs decreased the TEER, increased the albumin permeability and induced
morphological changes of HUVECs. The extent of the decrease was more profound with adherent
PBMCs than with non-adherent PBMCs. The expression of vascular endothelial cadherin
(VE-cadherin) was examined using real-time RT-PCR and immunofluorescence. Addition of
PBMCs to DV-2-infected HUVECs decreased the levels of mRNA transcripts and cell-surface
expression of VE-cadherin. The results indicate that PBMCs increased the permeability of
DV-2-infected HUVECs and that the increased permeability was concomitant with morphological
change and the decrease in VE-cadherin expression. The results suggest that functional
impairment of the DV-2-infected HUVEC monolayer was caused by interaction with PBMCs.
INTRODUCTION
Dengue viruses are a serious cause of morbidity andmortality in tropical and subtropical areas of the world(Halstead, 1988). Dengue viruses usually cause a mild tosevere febrile illness, dengue fever (DF), but may causedengue haemorrhagic fever (DHF) and dengue shocksyndrome (DSS) in some cases (Gubler, 1998; Halstead,1979). It is estimated that up to 100 million individuals areinfected with dengue viruses, with 500 000 cases of DHFand 25 000 deaths annually (Gubler, 1998). It has beensuggested that antibody-dependent enhancement of den-gue virus infection contributes to the pathogenesis of DHF(Halstead, 1979, 1988). It has also been proposed that theseverity of the disease is related to the virulence of thedengue virus strain (Leitmeyer et al., 1999; Rico-Hesseet al., 1997).
Although the mechanism of pathogenesis of DHF has notbeen completely elucidated, it is apparent that profoundplasma leakage is one of the characteristic physiological
features. Due to the lack of a good animal model,endothelial cells have been used as a surrogate to elucidatethe pathogenesis of dengue virus interactions. Endothelialcells can be infected with dengue virus in vitro (Bunyaratvejet al., 1997; Huang et al., 2003; Killen & O’Sullivan, 1993).Dengue virus infection induces cytokine production(Avirutnan et al., 1998; Bosch et al., 2002) and alters theexpression of adhesion molecules on endothelial cells(Krishnamurti et al., 2002). Although endothelial cells arenot massively infected, there is evidence of dengue virusinfection of endothelial cells in vivo (Gubler & Zaki, 1998;Hall et al., 1991; Jessie et al., 2004). It is possible that lowlevels of dengue virus infection of endothelial cells mayinduce profound effects locally, along with other factorssuch as cytokines and peripheral blood mononuclear cells(PBMCs).
Endothelial cells form a continuous cell layer along the wallof blood vessels and control the movement of solutes andfluid from the vascular space to the tissue (Tretiach et al.,2003). Two main types of junctional structures, adherence
Journal of General Virology (2008), 89, 642–652 DOI 10.1099/vir.0.83356-0
642 0008-3356 G 2008 SGM Printed in Great Britain

Downloaded from www.microbiologyresearch.org by
IP: 54.90.14.242
On: Mon, 03 Oct 2016 07:50:14
and tight junctions, are involved in cell–cell contact. Theadherence junction is a multimolecular complex composedpredominantly of transmembrane proteins termed cadher-ins (Navarro et al., 1995; Steinberg & McNutt, 1999) and isa major contributor to the maintenance of monolayerintegrity (Allport et al., 2000; Dejana et al., 1999; DelMaschio et al., 1996). Vascular endothelial cells express aunique member of the cadherin family, vascular endothe-lial cadherin (VE-cadherin). VE-cadherin is a novelmember of the cadherin family and is selectively expressedat intercellular boundaries of all types of endothelial cells,but is not expressed by any other cell types (Dejana et al.,2001; Lampugnani et al., 1992). VE-cadherin plays animportant role in the maintenance of endothelial mono-layer integrity. Blocking of VE-cadherin functions withspecific antibodies results in increased vascular permeab-ility (Corada et al., 2001; Hordijk et al., 1999).
Activation of endothelial cells facilitates the adherence ofleukocytes to endothelial cells (McIntyre et al., 1985, 1997).Moreover, activated endothelial cells, in turn, activateadherent cells (McIntyre et al., 1985, 1997). Therefore, wehypothesized that adherence of leukocytes to endothelialcells infected with dengue virus might change the functionsof endothelial cells and increase permeability. In the presentstudy, we attempted to determine the effect of PBMCs onthe permeability of dengue virus-infected human umbilicalvein endothelial cells (HUVECs). A transendothelial elec-trical resistance (TEER) assay and albumin permeabilityassay were used to assess the permeability of HUVECs invitro (Dewi et al., 2004). In addition, we examined therelationship between the permeability of endothelial cellsand VE-cadherin expression.
METHODS
Human PBMCs. PBMCs were obtained from two healthy adult
donors. Donor A was dengue antibody-positive and had been
immunized with Japanese encephalitis vaccine 2 years previously.
Donor B was Japanese encephalitis antibody-positive but dengue
antibody-negative. PBMCs were separated by Ficoll-Paque
(Pharmacia Biotech AB) gradient centrifugation in LEUCOSEP tubes
(Greiner Labortechnik) according to the manufacturer’s instructions.
A ring of PBMCs in the tube were collected and washed twice in
30 ml RPMI 1640 medium. PBMCs were resuspended at a
concentration of 16106 cells ml21 in RPMI 1640 medium (Sigma)
supplemented with 10 % heat-inactivated fetal bovine serum (FBS)
(Sigma), L-glutamine and NaHCO3 (Gibco Invitrogen), 100 mg
streptomycin ml21 and 100 U penicillin ml21 (Gibco Invitrogen)
(Scott et al., 1980).
Separation of adherent and non-adherent PBMCs. Adherent
PBMCs were isolated as described by Malkovsky et al. (1987). One
million PBMCs in 1 ml RPMI supplemented with 10 % FBS were
added to 12-well tissue-culture plates (Nunc). After 2 h of incubation
at 37 uC in 5 % CO2, non-adherent cells were removed by pipetting and
plates were washed twice with RPMI medium without FBS. Adherent
PBMCs in the wells were removed by gentle scraping with a rubber cell
scraper. Adherent and non-adherent PBMCs were washed and
resuspended at a concentration of 16106 cells ml21 in RPMI
supplemented with 10 % FBS. Fifty microlitres non-adherent or
adherent PBMCs at a concentration of 16106 cells ml21 was addedto HUVECs on transwell membranes in the experiments. Thepercentage of monocytes in fractionated PBMCs was determined byimmunofluorescence with FITC-conjugated, anti-CD14 antibody(Biomedia). Anti-CD14 antibody reacts with the 55 kDa single-chainmembrane glycoprotein that is expressed primarily on monocytes andmacrophages.
Infection of HUVECs with dengue virus. HUVECs were purchasedfrom Clonetics (catalogue no. CC-2517) and propagated insupplemented endothelial cell basal medium-2 (Dewi et al., 2004).HUVECs were infected with dengue-2 virus (DV-2) New Guinea Cstrain on transwell membranes at an m.o.i. of 0.5 p.f.u. per cell.Briefly, medium in the upper transwell polycarbonate membraneculture dish was removed and the cells were infected by adding 50 mlDV-2 per well. Cells were incubated for 2 h at 37 uC in 5 % CO2, andwashed with PBS without CaCl2 and MgCl2 [PBS (2)]. Culturesupernatant fluid from uninfected C6/36 cells and heat-inactivatedDV-2 (Putnak et al., 1991) were used as controls. HUVECs were alsoinfected with DV-2 on slide chambers for determining the percentageof DV-2 antigen-positive cells and for immunofluorescence stainingof VE-cadherin.
Effect of PBMCs and culture supernatant fluids from DV-2-
infected HUVECs and PBMCs on the permeability of HUVECs.HUVECs were infected with DV-2 for 2 h, as described above, and50 ml of 106 to 102 PBMCs ml21 was added to the infected HUVECs.In other experiments, HUVECs on transwell membranes were washedand 100 ml culture supernatant fluid from DV-2-infected HUVECsand PBMCs were added. After various incubation times, the TEERwas assessed. Culture supernatant fluids from uninfected HUVECstreated with PBMCs were used as controls. Human TNF-a (1 mgml21) with a titre of 26107 U mg21 was used as a positive control(Dewi et al., 2004).
Transendothelial electrical resistance (TEER). The TEER ofHUVECs was measured using the electrical cell sensor systemEndohm chamber (Millicell-ERS; Millipore) (World PrecisionInstruments), as reported previously (Dewi et al., 2004). The TEERwas first measured at 24 h after seeding, and this time point wasdefined as 0 h. The TEER was measured at various incubation timepoints after addition of PBMCs or culture supernatant fluids fromDV-2-infected HUVECs and PBMCs. The TEER included theresistance of the interelectrode solution and blank membrane;therefore, a transwell polycarbonate membrane culture dish withoutcells was always included as the blank. Every test was done induplicate and repeated at least three times. The mean and SD werecalculated from two independent experiments with two transwellculture dishes per group.
Transendothelial albumin permeability. Transendothelial albuminpermeability on transwell polycarbonate membrane culture disheswas also examined using trypan blue-bound BSA as described byBonner & O’Sullivan (1998) with a minor modification (Dewi et al.,2004). Two transwell culture dishes per group were washed with PBSand 100 ml trypan blue-bound BSA was added to the upper chamberof each transwell polycarbonate membrane culture dish containingHUVEC monolayers (Dewi et al., 2004). The amount of trypan blue-bound BSA detected in the lower chamber was considered torepresent the level of HUVEC permeability.
Counting the PBMCs attached to HUVECs. In order to determinewhether dengue virus infection increases the number of PBMCsadherent to HUVECs, HUVEC-attached PBMCs were counted.HUVECs were infected with DV-2 at an m.o.i. of 0.5 p.f.u. per cellfor 2 h. PBMCs were added to HUVECs and incubated for 24 h. Cellswere washed twice with PBS (2), fixed with methanol at 37 uC for
Permeability of dengue virus-infected endothelial cells
http://vir.sgmjournals.org 643

Downloaded from www.microbiologyresearch.org by
IP: 54.90.14.242
On: Mon, 03 Oct 2016 07:50:14
30 min and washed twice with PBS (2). Cells were then stained with
Giemsa for 30 min at room temperature and washed once with water
and once with 30 % methanol in PBS (2). After air drying, the
PBMCs on HUVECs were counted in five fields for each treatment
with a magnification of 6200 under a light microscope.
Morphology of HUVECs. HUVECs were seeded into 96-well flat-
bottomed microplates (Nunc) and infected with DV-2 at an m.o.i. of
0.5 p.f.u. per cell. PBMCs were added and incubated for 24 h.
HUVECs were then fixed and stained as reported previously (Dewi
et al., 2004). After air drying, the morphology of HUVECs was
observed under a light microscope with magnification of 6200.
Indirect immunofluorescence staining. PBMCs and HUVECs
were stained for DV-2 antigen by indirect immunofluorescence
staining as described previously (Dewi et al., 2004). Cells were
incubated with 1 : 250-diluted hyperimmune mouse ascites fluid to
DV-2 (Brandt et al., 1967) at 37 uC for 60 min. After washing, the
cells were reacted with 1 : 500-diluted FITC-conjugated sheep anti-
mouse IgG (Cappel) in PBS containing 5 % Block Ace (AbD Serotec)
at 37 uC for 60 min. The slide was washed and covered with Syva
microtech mounting fluid which contained PBS with glycerol
(Boehring Diagnostic Inc.). Cells were observed under a fluorescence
microscope.
Immunofluorescence staining of VE-cadherin. HUVECs on slide
chambers were washed with PBS and air-dried for 20 min. Cells were
fixed in acetone at –20 uC for 20 min and rinsed in PBS. Cells
were treated with Block Ace at 37 uC for 60 min. After washing
three times for 3 min with PBS, cells were incubated with 1 : 500-
diluted, FITC-labelled antibody to VE-cadherin (Bender
MedSystems) in PBS containing 5 % Block Ace at 37 uC for
60 min. Slides were washed and covered with Syva microtech
mounting fluid containing phosphate buffer with glycerol (Boehring
Diagnostic Inc.). Cells were observed under a fluorescence micro-
scope at 6400 magnification.
RNA isolation from HUVECs. HUVECs on 12-well plates (Nunc)
were lysed with 1 ml Isogen (Wako Nippon Gene) and 200 ml
chloroform was then added. RNA isolation was done according to the
manufacturer’s instructions (Wako Nippon Gene). The RNA was
dissolved in 20 ml ultrapure DNase-free, RNase-free distilled water
(Invitrogen Life Technologies) and then treated with DNase I
(Invitrogen Life Technologies) at room temperature for 15 min to
remove contaminating DNA. The RNA concentration was measured
using a spectrophotometer and adjusted to 100 mg ml21 and used for
real-time RT-PCR. Glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) was used as a housekeeping gene to confirm the use of
equal amounts of HUVEC mRNA.
Real-time RT-PCR. TaqMan primers and probes for detection of the
VE-cadherin gene were prepared according to the sequence of
GenBank accession no. NM_001795 and those for the GAPDH gene
used GenBank accession no. BC004109. Primers used for amplifica-
tion of VE-cadherin were forward primer 59-TCACCTGGTCGCCA-
ATCC-39 and reverse primers 59-AGGCCACATCTTGGGTTCCT-39.
The probe specific for VE-cadherin was 59-TCTCTTTCTTTTCTC-
TGTCTACTCCTTATCCCTTGGTT-39. Primers used for amplifica-
tion of GAPDH were forward primer 59-GAAATCCCATCACC-
ATCTTCCA-39 and reverse primer 59-CCAGCATCGCCCCACTT-39.
The probe specific for GAPDH was 59-AGCGAGATCCCTCC-39.
The ABI PRISM 7000 sequence detection system (Applied
Biosystems) was used for one-step RT-PCR and hybridization.
Aliquots (5–10 ng) of purified RNA were used as the template in
25 ml total reaction volumes. The reagents were added to the TaqMan
one-step RT-PCR master mix reagents (PE Applied Biosystems). The
real-time RT-PCR program was: initial incubation for 30 min at48 uC for reverse transcription and denaturation at 95 uC for 10 minfollowed by 45 cycles of 95 uC for 15 s and 57 uC for 1 min. All testswere performed in duplicate.
Statistical analysis. The significance of differences was determinedby Student’s t-tests compared with control HUVECs withouttreatment. The data were expressed as means±SD, and differencesare considered to be significant when the P value was less than 0.05.The TEER test consisted of at least three independent experimentswith two transwell polycarbonate membrane culture dishes.
RESULTS
Effect of PBMCs on the permeability of HUVECs
The effect of PBMCs on the TEER of HUVECs wasexamined. HUVECs on a transwell membrane wereinfected with DV-2 at an m.o.i. of 0.5 to 0.0005 p.f.u. percell or treated with C6/36 supernatant or inactivated DV-2for 2 h. After washing, 56105 to 56102 PBMCs wereadded to DV-2-infected HUVECs and the TEER wasassessed. Addition of 56105 or 56104 PBMCs to HUVECsinfected with DV-2 at an m.o.i. of 0.5 p.f.u. per celldecreased the TEER, but addition of 56103 or fewerPBMCs did not (Tables 1 and 2). The results were similarusing PBMCs from donor B.
HUVECs infected with DV-2 at an m.o.i. of 0.5 p.f.u. percell did not demonstrate a decrease in the TEER or anincrease in albumin permeability compared with untreatedHUVECs unless PBMCs were added (Table 1). Addition ofPBMCs to untreated HUVECs or to HUVECs treated withC6/36 supernatant or inactivated DV-2 did not decreasethe TEER or increase the albumin permeability. The TEERof PBMCs alone was at the similar level to wells containingno cells (data not shown).
Changes in permeability were also assessed by the albuminpermeability assay. Addition of 56104 PBMCs to HUVECsinfected with DV-2 at an m.o.i. of 0.5 p.f.u. per cellincreased the albumin permeability across HUVECs(Fig. 1), but addition of PBMCs to HUVECs treated withC6/36 supernatant or inactivated DV-2 did not. Thus, theincrease in the albumin permeability across HUVECs wasconsistent with the decrease in the TEER. The increase inthe albumin permeability across HUVECs become moreprofound on days 2–5.
Effects of adherent and non-adherent PBMCs onthe permeability of HUVECs
PBMCs were separated into adherent and non-adherentfractions and used in the assays. The adherent and non-adherent fractions contained CD14-positive monocytes at94±4 % and 5±2 %, respectively. Both adherent and non-adherent fractions decreased the TEER of DV-2-infectedHUVECs. The extent of the decrease was more profoundwith the adherent fraction than with non-adherent fraction(Table 1).
B. E. Dewi, T. Takasaki and I. Kurane
644 Journal of General Virology 89
Downloaded from www.microbiologyresearch.org by
IP: 54.90.14.242
On: Mon, 03 Oct 2016 07:50:14
Effect of the supernatant fluid from DV-2-infectedHUVECs and PBMCs on the permeability ofHUVECs
HUVECs were infected with DV-2 at an m.o.i. of 0.5 p.f.u.per cell for 2 h, 56104 PBMCs were added and super-natants were collected at various time points. The collectedculture supernatant fluids were then added to HUVECs ontranswell membranes and the permeability was assessed for3 days. Culture supernatant fluids collected on days 1 and2 did not decrease the TEER, but those collected on days 3and 4 did (Table 2). The decrease in the TEER started 4 hafter addition of the supernatant fluids. Those collected ondays 5, 6 and 7 also decreased the TEER (data not shown).Supernatant fluids collected on days 3–7 from the culturecontaining HUVECs treated with inactivated DV-2 or C6/36 and PBMCs did not decrease the TEER (data notshown).
Effect of the level of DV-2 infection on thepermeability of HUVECs
To determine the relationship between the level of DV-2infection of HUVECs and the PBMC-induced decrease inthe TEER, HUVECs were infected with DV-2 at variousm.o.i. and then 56104 PBMCs were added. Infection ofHUVECs with DV-2 at an m.o.i. of 0.5 p.f.u. per celldecreased the TEER when PBMCs were added, butinfection at an m.o.i. of 0.05 p.f.u. per cell or less did not(Table 2). HUVECs treated with C6/36 supernatant or
treated with inactivated DV-2 did not decrease the TEERwhen 56104 PBMCs were added.
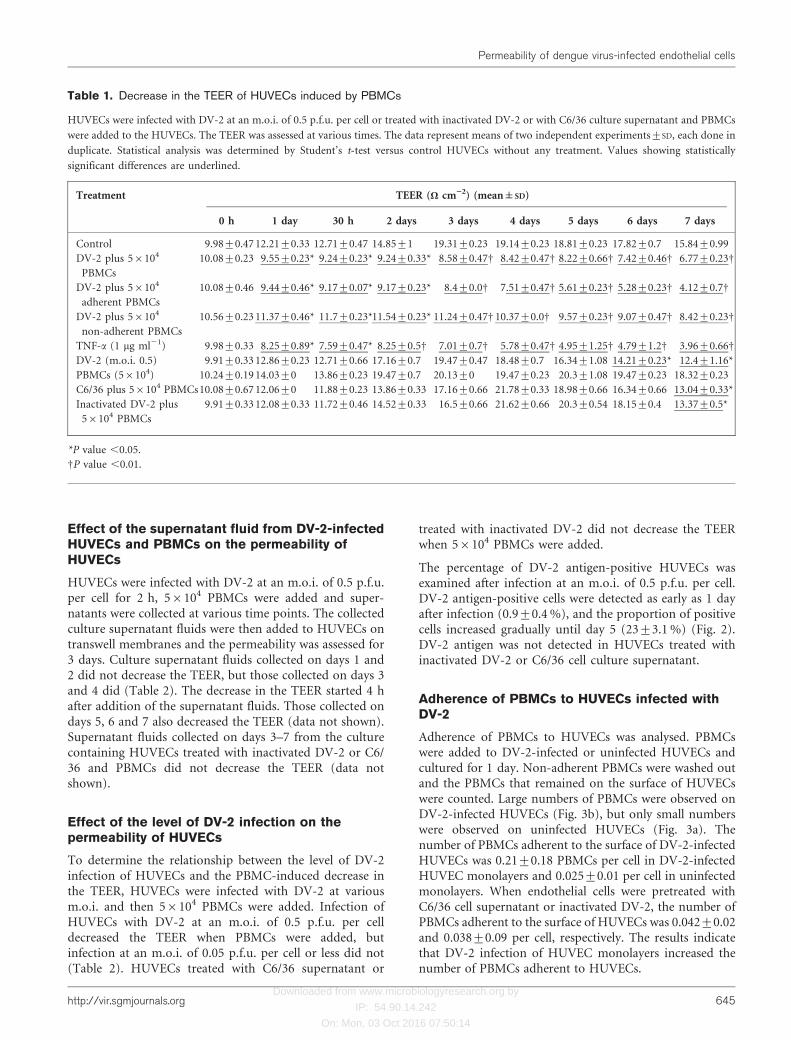
The percentage of DV-2 antigen-positive HUVECs wasexamined after infection at an m.o.i. of 0.5 p.f.u. per cell.DV-2 antigen-positive cells were detected as early as 1 dayafter infection (0.9±0.4 %), and the proportion of positivecells increased gradually until day 5 (23±3.1 %) (Fig. 2).DV-2 antigen was not detected in HUVECs treated withinactivated DV-2 or C6/36 cell culture supernatant.
Adherence of PBMCs to HUVECs infected withDV-2
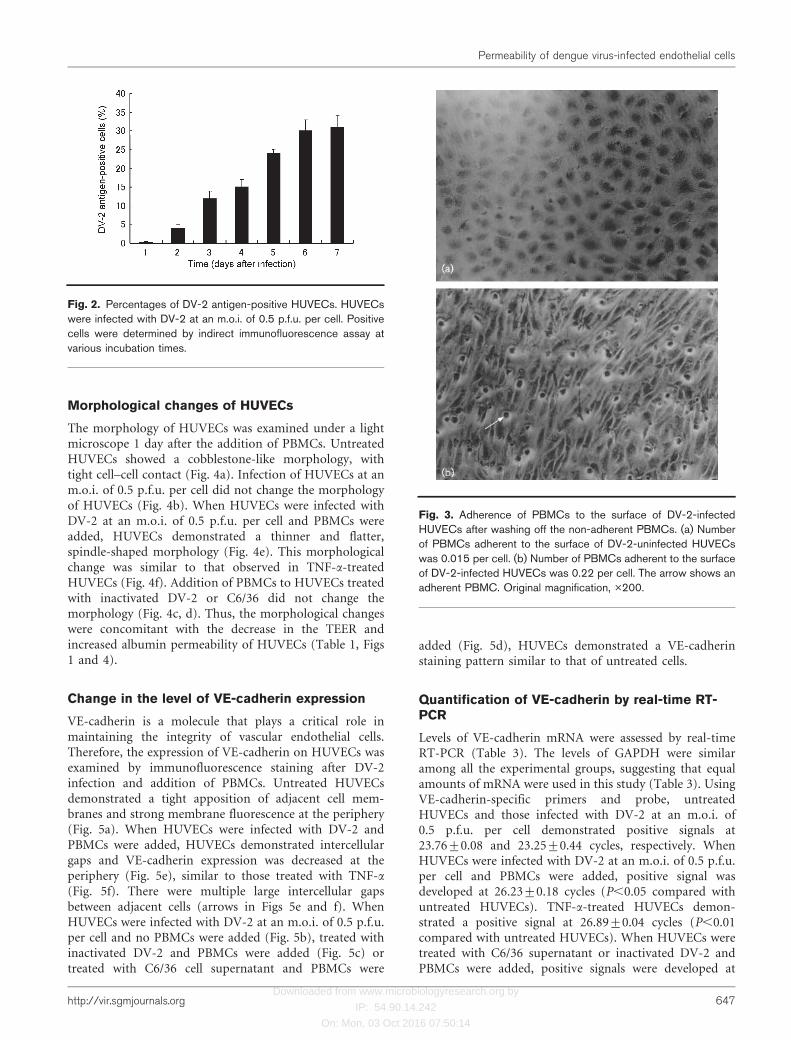
Adherence of PBMCs to HUVECs was analysed. PBMCswere added to DV-2-infected or uninfected HUVECs andcultured for 1 day. Non-adherent PBMCs were washed outand the PBMCs that remained on the surface of HUVECswere counted. Large numbers of PBMCs were observed onDV-2-infected HUVECs (Fig. 3b), but only small numberswere observed on uninfected HUVECs (Fig. 3a). Thenumber of PBMCs adherent to the surface of DV-2-infectedHUVECs was 0.21±0.18 PBMCs per cell in DV-2-infectedHUVEC monolayers and 0.025±0.01 per cell in uninfectedmonolayers. When endothelial cells were pretreated withC6/36 cell supernatant or inactivated DV-2, the number ofPBMCs adherent to the surface of HUVECs was 0.042±0.02and 0.038±0.09 per cell, respectively. The results indicatethat DV-2 infection of HUVEC monolayers increased thenumber of PBMCs adherent to HUVECs.
Table 1. Decrease in the TEER of HUVECs induced by PBMCs
HUVECs were infected with DV-2 at an m.o.i. of 0.5 p.f.u. per cell or treated with inactivated DV-2 or with C6/36 culture supernatant and PBMCs
were added to the HUVECs. The TEER was assessed at various times. The data represent means of two independent experiments±SD, each done in
duplicate. Statistical analysis was determined by Student’s t-test versus control HUVECs without any treatment. Values showing statistically
significant differences are underlined.
Treatment TEER (V cm”2) (mean±SD)
0 h 1 day 30 h 2 days 3 days 4 days 5 days 6 days 7 days
Control 9.98±0.47 12.21±0.33 12.71±0.47 14.85±1 19.31±0.23 19.14±0.23 18.81±0.23 17.82±0.7 15.84±0.99
DV-2 plus 56104
PBMCs
10.08±0.23 9.55±0.23* 9.24±0.23* 9.24±0.33* 8.58±0.47D 8.42±0.47D 8.22±0.66D 7.42±0.46D 6.77±0.23D
DV-2 plus 56104
adherent PBMCs
10.08±0.46 9.44±0.46* 9.17±0.07* 9.17±0.23* 8.4±0.0D 7.51±0.47D 5.61±0.23D 5.28±0.23D 4.12±0.7D
DV-2 plus 56104
non-adherent PBMCs
10.56±0.23 11.37±0.46* 11.7±0.23*11.54±0.23* 11.24±0.47D 10.37±0.0D 9.57±0.23D 9.07±0.47D 8.42±0.23D
TNF-a (1 mg ml21) 9.98±0.33 8.25±0.89* 7.59±0.47* 8.25±0.5D 7.01±0.7D 5.78±0.47D 4.95±1.25D 4.79±1.2D 3.96±0.66D
DV-2 (m.o.i. 0.5) 9.91±0.33 12.86±0.23 12.71±0.66 17.16±0.7 19.47±0.47 18.48±0.7 16.34±1.08 14.21±0.23* 12.4±1.16*
PBMCs (56104) 10.24±0.19 14.03±0 13.86±0.23 19.47±0.7 20.13±0 19.47±0.23 20.3±1.08 19.47±0.23 18.32±0.23
C6/36 plus 56104 PBMCs10.08±0.67 12.06±0 11.88±0.23 13.86±0.33 17.16±0.66 21.78±0.33 18.98±0.66 16.34±0.66 13.04±0.33*
Inactivated DV-2 plus
56104 PBMCs
9.91±0.33 12.08±0.33 11.72±0.46 14.52±0.33 16.5±0.66 21.62±0.66 20.3±0.54 18.15±0.4 13.37±0.5*
*P value ,0.05.
DP value ,0.01.
Permeability of dengue virus-infected endothelial cells
http://vir.sgmjournals.org 645

Downloaded from www.microbiologyresearch.org by
IP: 54.90.14.242
On: Mon, 03 Oct 2016 07:50:14
Table 2. Comparison of the decrease in the TEER observed with various numbers of PBMCs, various m.o.i. of DV-2 and supernatantof DV-2-infected HUVECs and PBMCs
The TEER was assessed at various incubation times. Data are means of two independent experiments±SD, each done in duplicate. Statistical
analysis was determined by Student’s t-test versus control HUVECs without any treatment. Values showing statistically significant differences are
underlined.
Treatment TEER (V cm”2) (mean±SD)
0 h 1 day 30 h 2 days 3 days 4 days 5 days 6 days 7 days
Control 9.98±0.47 12.21±0.33 12.71±0.47 14.85±1 19.31±0.23 19.14±0.23 18.81±0.23 17.82±0.7 15.84±0.99
TNF-a (1 mg ml21) 9.98±0.33 8.25±0.89* 7.59±0.47* 8.25±0.5D 7.01±0.7D 5.78±0.47D 4.95±1.25D 4.79±1.2D 3.96±0.66D
DV-2 (m.o.i. 0.5)
+PBMCs (56105)
10.24±0.33 9.04±0.05* 8.91±0* 8.54±0.33D 8.05±0.33D 8.09±0D 6.59±0.33D 6.76±0.66D 5.12±0.33D
DV-2 (m.o.i. 0.5)
+PBMCs (56104)
10.08±0.23 9.55±0.23* 9.24±0.23* 9.24±0.33* 8.58±0.47D 8.42±0.47D 8.22±0.66D 7.42±0.46D 6.77±0.23D
DV-2 (m.o.i. 0.5)
+PBMCs (56103)
9.91±0.33 11.22±0.33 10.56±0.99 13.45±2.64 17.68±0.66 18.57±1.65 15.87±0.33 13.04±0* 10.4±0.33*
DV-2 (m.o.i. 0.5)
+PBMCs (56102)
10.24±0.33 12.54±0.33 12.54±0.33 18.83±0.99 19.31±0.33 19.82±0.33 17.86±0.23 15.53±0.33 14.39±0.33
DV-2 (m.o.i. 0.05)
+PBMCs (56104)
9.91±0.99 12.54±0.99 12.05±0.66 14.52±0.33 17.66±0.99 24.92±0.66 20.3±2.31 18.32±0.23 14.19±0.33
DV-2 (m.o.i. 0.005)
+PBMCs (56104)
9.91±0.33 13.2±0.99 13.7±0.33 16.83±0.99 20.63±1.32 26.89±0.33 23.1±0 16.83±2.31 14.36±2.64
DV-2 (m.o.i. 0.0005)
+PBMCs (56104)
10.24±0.66 13.2±0.33 13.53±0.66 17.16±1.98 20.46±0.33 27.39±0.33 24.26±2.64 19.47±0.66 17.16±0.33
DV-2 (m.o.i. 0.00005)
+PBMCs (56104)
9.99±0.12 12.87±0.47 13.2±0.7 17.41±0.82 20.46±0.23 26.32±0.82 23.02±0.11 18.77±1.75 15.26±1.28
Supernatant DV-2
+PBMCs (day 1)
9.08±0.33 12.21±0.99 12.72±0.46 14.85±0.33 19.3±0.66 19.1±0.66 18.8±0.54 17.85±0.4 15.8±0.5
Supernatant DV-2
+PBMCs (day 2)
9.31±0.47 11.89±0.33 12.34±0.46 13.56±0.33 16.98±0.66 18.62±0.66 17.9.3±0.54 17.15±0.4 14.37±0.66
Supernatant DV-2
+PBMCs (day 3)
9.08±0.33 8.35±0.33* 9.72±0.46* 8.15±0.66D 8.2±0.66D 8.54±0.83D 8.12±0.66D 8.15±0.33D 7.37±0.5D
Supernatant DV-2
+PBMCs (day 4)
9.05±0.66 8.3±0.5* 8.23±0.66* 8.09±0.5D 8.89±1.32D 7.62±0.66D 7.3±0.54D 8.15±0.4D 7.37±0.33D
*P value ,0.05.
DP value ,0.01.
Fig. 1. Clearance of labelled BSA acrosscontrol and treated HUVECs at variousincubation times. Addition of PBMCs toinfected HUVECs showed increased clear-ance of labelled BSA (P,0.05). A similarresult was also shown for HUVECs treatedwith 1 mg TNF-a ml”1.
B. E. Dewi, T. Takasaki and I. Kurane
646 Journal of General Virology 89
Downloaded from www.microbiologyresearch.org by
IP: 54.90.14.242
On: Mon, 03 Oct 2016 07:50:14
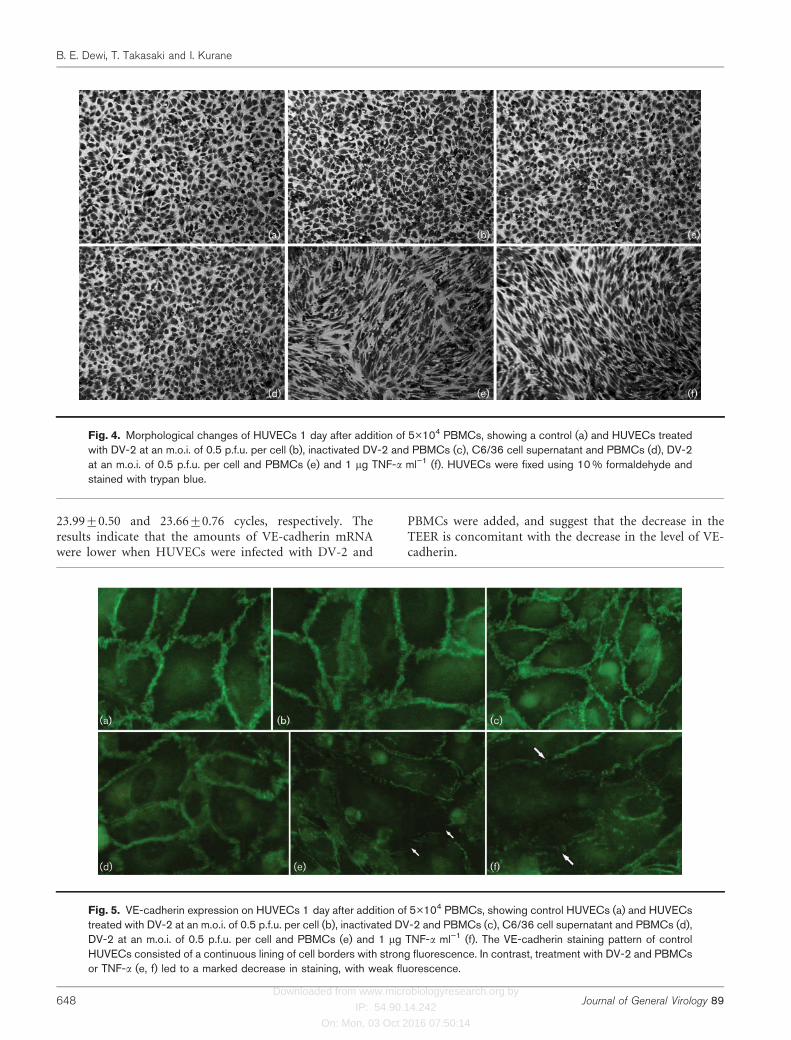
Morphological changes of HUVECs
The morphology of HUVECs was examined under a lightmicroscope 1 day after the addition of PBMCs. UntreatedHUVECs showed a cobblestone-like morphology, withtight cell–cell contact (Fig. 4a). Infection of HUVECs at anm.o.i. of 0.5 p.f.u. per cell did not change the morphologyof HUVECs (Fig. 4b). When HUVECs were infected withDV-2 at an m.o.i. of 0.5 p.f.u. per cell and PBMCs wereadded, HUVECs demonstrated a thinner and flatter,spindle-shaped morphology (Fig. 4e). This morphologicalchange was similar to that observed in TNF-a-treatedHUVECs (Fig. 4f). Addition of PBMCs to HUVECs treatedwith inactivated DV-2 or C6/36 did not change themorphology (Fig. 4c, d). Thus, the morphological changeswere concomitant with the decrease in the TEER andincreased albumin permeability of HUVECs (Table 1, Figs1 and 4).
Change in the level of VE-cadherin expression
VE-cadherin is a molecule that plays a critical role inmaintaining the integrity of vascular endothelial cells.Therefore, the expression of VE-cadherin on HUVECs wasexamined by immunofluorescence staining after DV-2infection and addition of PBMCs. Untreated HUVECsdemonstrated a tight apposition of adjacent cell mem-branes and strong membrane fluorescence at the periphery(Fig. 5a). When HUVECs were infected with DV-2 andPBMCs were added, HUVECs demonstrated intercellulargaps and VE-cadherin expression was decreased at theperiphery (Fig. 5e), similar to those treated with TNF-a
(Fig. 5f). There were multiple large intercellular gapsbetween adjacent cells (arrows in Figs 5e and f). WhenHUVECs were infected with DV-2 at an m.o.i. of 0.5 p.f.u.per cell and no PBMCs were added (Fig. 5b), treated withinactivated DV-2 and PBMCs were added (Fig. 5c) ortreated with C6/36 cell supernatant and PBMCs were
added (Fig. 5d), HUVECs demonstrated a VE-cadherinstaining pattern similar to that of untreated cells.
Quantification of VE-cadherin by real-time RT-PCR
Levels of VE-cadherin mRNA were assessed by real-timeRT-PCR (Table 3). The levels of GAPDH were similaramong all the experimental groups, suggesting that equalamounts of mRNA were used in this study (Table 3). UsingVE-cadherin-specific primers and probe, untreatedHUVECs and those infected with DV-2 at an m.o.i. of0.5 p.f.u. per cell demonstrated positive signals at23.76±0.08 and 23.25±0.44 cycles, respectively. WhenHUVECs were infected with DV-2 at an m.o.i. of 0.5 p.f.u.per cell and PBMCs were added, positive signal wasdeveloped at 26.23±0.18 cycles (P,0.05 compared withuntreated HUVECs). TNF-a-treated HUVECs demon-strated a positive signal at 26.89±0.04 cycles (P,0.01compared with untreated HUVECs). When HUVECs weretreated with C6/36 supernatant or inactivated DV-2 andPBMCs were added, positive signals were developed at
Fig. 3. Adherence of PBMCs to the surface of DV-2-infectedHUVECs after washing off the non-adherent PBMCs. (a) Numberof PBMCs adherent to the surface of DV-2-uninfected HUVECswas 0.015 per cell. (b) Number of PBMCs adherent to the surfaceof DV-2-infected HUVECs was 0.22 per cell. The arrow shows anadherent PBMC. Original magnification, �200.
Fig. 2. Percentages of DV-2 antigen-positive HUVECs. HUVECswere infected with DV-2 at an m.o.i. of 0.5 p.f.u. per cell. Positivecells were determined by indirect immunofluorescence assay atvarious incubation times.
Permeability of dengue virus-infected endothelial cells
http://vir.sgmjournals.org 647
Downloaded from www.microbiologyresearch.org by
IP: 54.90.14.242
On: Mon, 03 Oct 2016 07:50:14
23.99±0.50 and 23.66±0.76 cycles, respectively. Theresults indicate that the amounts of VE-cadherin mRNAwere lower when HUVECs were infected with DV-2 and
PBMCs were added, and suggest that the decrease in theTEER is concomitant with the decrease in the level of VE-cadherin.
Fig. 4. Morphological changes of HUVECs 1 day after addition of 5�104 PBMCs, showing a control (a) and HUVECs treatedwith DV-2 at an m.o.i. of 0.5 p.f.u. per cell (b), inactivated DV-2 and PBMCs (c), C6/36 cell supernatant and PBMCs (d), DV-2at an m.o.i. of 0.5 p.f.u. per cell and PBMCs (e) and 1 mg TNF-a ml”1 (f). HUVECs were fixed using 10 % formaldehyde andstained with trypan blue.
Fig. 5. VE-cadherin expression on HUVECs 1 day after addition of 5�104 PBMCs, showing control HUVECs (a) and HUVECstreated with DV-2 at an m.o.i. of 0.5 p.f.u. per cell (b), inactivated DV-2 and PBMCs (c), C6/36 cell supernatant and PBMCs (d),DV-2 at an m.o.i. of 0.5 p.f.u. per cell and PBMCs (e) and 1 mg TNF-a ml”1 (f). The VE-cadherin staining pattern of controlHUVECs consisted of a continuous lining of cell borders with strong fluorescence. In contrast, treatment with DV-2 and PBMCsor TNF-a (e, f) led to a marked decrease in staining, with weak fluorescence.
B. E. Dewi, T. Takasaki and I. Kurane
648 Journal of General Virology 89

Downloaded from www.microbiologyresearch.org by
IP: 54.90.14.242
On: Mon, 03 Oct 2016 07:50:14
DISCUSSION
The pathological mechanisms of DHF/DSS are notcompletely understood. In order to elucidate thesemechanisms, interactions among endothelial cells,PBMCs and DV-2 were analysed in vitro using HUVECs.Addition of PBMCs to DV-2-infected HUVECs decreasedthe TEER. The decrease in the TEER was consistent withthe increased albumin permeability across HUVECs andconcomitant with morphological changes observed inHUVECs and the decreases in VE-cadherin expressionand VE-cadherin mRNA.
Infection of HUVECs with DV-2 at an m.o.i. of 0.5 p.f.u.per cell did not decrease the TEER unless PBMCs wereadded. It is possible that dengue virus mediates endothelialcell barrier function via an indirect route rather than directinfection of endothelial cells. In general, however, the tissuedamage observed in pathological studies is not very severecompared with the severity of illness (Bhamarapravati,1989). Furthermore, the lack of structural damage, theshort-lived nature of the plasma leakage syndrome and theremarkably rapid recovery of children with DSS all suggestthat the alteration of permeability is effected by a solublemediator (Innis, 1995).
Given the increased permeability of endothelial cells in thepresence of PBMCs, it is likely that increased permeabilityof HUVECs was in part due to the cytokines which wereproduced by DV-2-infected PBMCs. Dengue viruses canreplicate in monocyte-derived macrophages and mono-cyte-like cell lines (Kurane et al., 1990; O’Sullivan & Killen,1994) and induce cytokines (Bosch et al., 2002; Espina et al.,2003) and cytotoxic factors (Mukerjee et al., 1995; Shaioet al., 1995). Culture fluids from dengue virus-infectedperipheral blood monocytes activated endothelial cells, as
measured by upregulation of cell adhesion molecules(Anderson et al., 1997), and increased the permeability ofendothelial cells in vitro (Carr et al., 2003). It has also beenreported that cytokines play an important role in thepathogenesis of DHF (Rothman & Ennis, 1999), andplasma leakage in DHF patients is associated with theelevation of plasma levels of various cytokines (Green et al.,1999; Kurane et al., 1991).
There was a report that only the supernatant collected fromdengue virus-infected monocyte-macrophages at 72 hincreased permeability, and not that collected before 72 h(Carr et al., 2003), and similar results were observed in thepresent study. In this experiment, we found that the TEERwas decreased (P,0.05) on day 1 after the addition ofPBMCs. Furthermore, larger numbers of PBMCs attachedto the surface of DV-2-infected HUVEC monolayers afterwashing than to uninfected HUVECs. It is possible that thedecrease in the TEER before 48 h after addition of PBMCsis due to activation of PBMCs by adherence to the DV-2-infected endothelial cell monolayer. After infection withdengue virus, endothelial cells express adhesion molecules(Krishnamurti et al., 2002). PBMCs which attached toinfected endothelial cells are activated, and activatedPBMCs may produce cytokines that act locally and increaseplasma leakage. Activation of endothelial cells, in turn,might activate the adherent leukocytes (McIntyre et al.,1997; McIntyre, 2000) and produce cytokines, chemokinesand tissue factors in a contact-dependent manner (Monacoet al., 2002).
Addition of PBMCs to HUVECs infected with DV-2 at anm.o.i. of less than 0.5 p.f.u. per cell did not decrease theTEER. Although the underlying mechanism remains poorlyunderstood, it seems that the number of viruses isassociated with the severity of disease. Epidemiologicalstudies of DHF patients showed that DV-3 viraemia wasgreater in more severe clinical disease (Libraty et al., 2002).Previous studies examining DV-1 and DV-2 infectionshave reported higher circulation levels of replicating virus(Vaughn et al., 2000) or viral RNA copies (Murgue et al.,2000) in DHF than in DF patients. Addition of PBMCs tocontrol HUVECs or HUVECs pretreated with C6/36 cellssupernatant or pretreated with inactivated DV-2 did notdecrease the TEER. These results indicate that infection ofHUVECs is necessary for the decrease of the TEER in thisexperimental system.
We separated adherent and non-adherent cell fractionsfrom PBMCs and tested their effects on the decrease of theTEER. The majority of adherent PBMCs on plastic wellplates were determined to be monocytes by immunofluor-escence staining with anti-CD14 antibody. AdherentPBMCs decreased the TEER to a greater extent than non-adherent PBMCs. It was reported that adherent cells are themajor cell population that supports dengue virus replica-tion in vivo (Scott et al., 1980). Allport et al. (2000) alsoshowed that the adherence of monocytes on endothelialcells induces focal disruption of the VE-cadherin.
Table 3. Real-time quantitative RT-PCR analysis to determinethe level of VE-cadherin mRNA in control and treated HUVECs
Data are means±SD. Statistical analysis was done in comparison with
control HUVECs; values showing statistically significant differences
are underlined.
Treatment CT value (cycle)
GAPDH
detector
VE-cadherin
detector
Control HUVECs 28.41±0.10 23.76±0.08
DV-2+PBMCs (56104) 28.39±0.11 26.23±0.18*
Inactivated DV-2+PBMCs
(56104)
28.43±0.02 23.66±0.76
C6/36+PBMCs (56104) 28.52±0.04 23.99±0.50
DV-2 alone 29.03±0.24 23.25±0.44
PBMCs (56104) alone 28.47±0.05 23.12±0.57
TNF-a (1 mg ml21) 28.48±0.11 26.89±0.04D
*P value ,0.05.
DP value ,0.01.
Permeability of dengue virus-infected endothelial cells
http://vir.sgmjournals.org 649

Downloaded from www.microbiologyresearch.org by
IP: 54.90.14.242
On: Mon, 03 Oct 2016 07:50:14
Consistent with these reports, the extent of the decreasewas more profound with adherent PBMCs than with non-adherent PBMCs in our study. We suggest that attachmentof PBMCs leads to production of cytokines that act locallyand increase the permeability of DV-2-infected HUVECs.
The morphology of HUVECs changed when HUVECs wereinfected with DV-2 at an m.o.i. of 0.5 p.f.u. per cell andPBMCs were added. A similar result was found onHUVECs treated with TNF-a. It was reported that thelow TEER was associated with irregular cell shape, whenstudied morphologically (Tretiach et al., 2003), andalterations in the permeability of the vascular endothelium(Bucana et al., 1988). McKenzie & Ridley (2007) found thatTNF-a subsequently caused a progressive increase inpermeability and in stress fibre reorganization, cellelongation and intercellular gap formation over 8–24 h.Consistent with the increased permeability, occludin andJAM-A were removed from tight junctions and ZO-1 waspartially redistributed (McKenzie & Ridley, 2007). Similarresults were obtained in the present study. Morphologicalchanges were observed concomitantly with the decrease inthe TEER and increased permeability.
Additionally, we examined the levels of VE-cadherinmRNA using real-time RT-PCR. Only when PBMCs wereadded, HUVECs infected with DV-2 demonstrated lowerlevels of VE-cadherin mRNA. In immunofluorescenceassays, infection of HUVECs with DV-2 at an m.o.i. of0.5 p.f.u. per cell and addition of PBMCs resulted in amarked decrease in immunofluorescence for membrane-associated VE-cadherin. This treatment also led to thedevelopment of multiple intercellular gaps between adja-cent cells. Since VE-cadherin plays a crucial role in themaintenance of endothelial cell–cell contact and monolayerintegrity (Breviario et al., 1995; Del Maschio et al., 1996),the appearance of multiple intercellular gaps betweenadjacent cells probably indicates an increase in permeab-ility. These results are consistent with previous reportsusing TNF-a (Friedl et al., 2002; Hofmann et al., 2002) andthrombin (Rabiet et al., 1996).
The disassembly of the adherence-junction component, asindicated by the decrease in VE-cadherin expression,probably contributes to the increase in endothelialpermeability in this study. In addition to adherencejunctions, endothelial cells also possess tight junctions.Studies are also needed to understand the role of the tightjunction of endothelial cells in the pathogenesis of DHF.Furthermore, the relevance of the results in the presentstudy to the pathogenesis of DHF in vivo needs to beelucidated in future studies.
ACKNOWLEDGEMENTS
We thank Mr Takagi, Dr M. Ito, Dr S. Tajima, Mr A. Kotaki and MsR. Nerome, Laboratory of Vector-Borne Viruses, Department ofVirology 1, National Institute of Infectious Diseases, for theirtechnical help. This study was supported by a grant-in-aid (no.12877045) from the Ministry of Education, Sciences, Sports and
Culture, Japan, and a grant for Research on Emerging andReEmerging Infectious Diseases from the Ministry of Health, Laborand Welfare, Japan (H18-Shinkou-ippan-009).
REFERENCES
Allport, J. R., Muller, W. A. & Luscinskas, F. W. (2000). Monocytesinduce reversible focal changes in vascular endothelial cadherincomplex during transendothelial migration under flow. J Cell Biol148, 203–216.
Anderson, R., Wang, S., Osiowy, C. & Issekutz, A. C. (1997).Activation of endothelial cells via antibody-enhanced dengue virusinfection of peripheral blood monocytes. J Virol 71, 4226–4232.
Avirutnan, P., Malasit, P., Seliger, B., Bhakdi, S. & Husmann, M.(1998). Dengue virus infection of human endothelial cells leads tochemokine production, complement activation, and apoptosis.J Immunol 161, 6338–6346.
Bhamarapravati, N. (1989). Hemostatic defects in dengue hemor-rhagic fever. Rev Infect Dis 11, S826–S829.
Bonner, S. M. & O’Sullivan, M. A. (1998). Endothelial cell monolayersas a model system to investigate dengue shock syndrome. J VirolMethods 71, 159–167.
Bosch, I., Xhaja, K., Estevez, L., Raines, G., Melichar, H., Warke, R. V.,Fournier, M. V., Ennis, F. A. & Rothman, A. L. (2002). Increasedproduction of interleukin-8 in primary human monocytes and inhuman epithelial and endothelial cell lines after dengue viruschallenge. J Virol 76, 5588–5597.
Brandt, W. E., Buescher, E. L. & Hetrick, F. M. (1967). Production andcharacterization of arbovirus antibody in mouse ascitic fluid. Am JTrop Med Hyg 16, 339–347.
Breviario, F., Caveda, L., Corada, M., Martin-Padura, I., Navarro, P.,Golay, J., Introna, M., Gulino, D., Lampugnani, M. G. & Dejana, E.(1995). Functional properties of human vascular endothelial cadherin(7B4/cadherin-5), an endothelium-specific cadherin. ArteriosclerThromb Vasc Biol 15, 1229–1239.
Bucana, C. D., Trial, J., Papp, A. C. & Wu, K. K. (1988). Bovine aortaendothelial cell incubation with interleukin 2: morphological changescorrelate with enhanced vascular permeability. Scanning Microsc 2,1559–1566.
Bunyaratvej, A., Butthep, P., Yoksan, S. & Bhamarapravati, N.(1997). Dengue viruses induce cell proliferation and morphologicalchanges of endothelial cells. Southeast Asian J Trop Med Public Health28, 32–37.
Carr, J. M., Hocking, H., Bunting, K., Wright, P. J., Davidson, A.,Gamble, J., Burrell, C. J. & Li, P. (2003). Supernatants from denguevirus type-2 infected macrophages induce permeability changes inendothelial cell monolayers. J Med Virol 69, 521–528.
Corada, M., Liao, F., Lindgren, M., Lampugnani, M. G., Breviario, F.,Frank, R., Muller, W. A., Hicklin, D. J., Bohlen, P. & Dejana, E. (2001).Monoclonal antibodies directed to different regions of vascularendothelial cadherin extracellular domain affect adhesion andclustering of the protein and modulate endothelial permeability.Blood 97, 1679–1684.
Dejana, E., Bazzoni, G. & Lampugnani, M. G. (1999). The role ofendothelial cell-to-cell junctions in vascular morphogenesis. ThrombHaemost 82, 755–761.
Dejana, E., Spagnuolo, R. & Bazzoni, G. (2001). Interendothelialjunctions and their role in the control of angiogenesis, vascular perme-ability and leukocyte transmigration. Thromb Haemost 86, 308–315.
Del Maschio, A., Zanetti, A., Corada, M., Rival, Y., Ruco, L.,Lampugnani, M. G. & Dejana, E. (1996). Polymorphonuclear
B. E. Dewi, T. Takasaki and I. Kurane
650 Journal of General Virology 89

Downloaded from www.microbiologyresearch.org by
IP: 54.90.14.242
On: Mon, 03 Oct 2016 07:50:14
leukocyte adhesion triggers the disorganization of endothelial cell-to-cell adherens junctions. J Cell Biol 135, 497–510.
Dewi, B. E., Takasaki, T. & Kurane, I. (2004). In vitro assessment ofhuman endothelial cell permeability: effects of inflammatorycytokines and dengue virus infection. J Virol Methods 121, 171–180.
Espina, L. M., Valero, N. J., Hernandez, J. M. & Mosquera, J. A. (2003).Increased apoptosis and expression of tumor necrosis factor-alphacaused by infection of cultured human monocytes with dengue virus.Am J Trop Med Hyg 68, 48–53.
Friedl, J., Puhlmann, M., Bartlett, D. L., Libutti, S. K., Turner, E. N.,Gnant, M. F. X. & Alexander, H. R. (2002). Induction of permeabilityacross endothelial cell monolayers by tumor necrosis factor (TNF)occurs via a tissue factor-dependent mechanism: relationship betweenthe procoagulant and permeability effects of TNF. Blood 100,1334–1349.
Green, S., Vaughn, D. W., Kalayanarooj, S., Nimmannitya, S.,Suntayakorn, S., Nisalak, A., Rothman, A. L. & Ennis, F. A. (1999).Elevated plasma interleukin-10 levels in acute dengue correlate withdisease severity. J Med Virol 59, 329–334.
Gubler, D. J. (1998). Dengue and dengue hemorrhagic fever. ClinMicrobiol Rev 11, 480–496.
Gubler, D. J. & Zaki, S. R. (1998). Dengue and other viral hemorrhagicfevers. In Pathology of Emerging Infections, vol. 2, pp. 43–71. Edited byA. M. Nelson & C. R. Horsburgh, Jr. Washington, DC: AmericanSociety for Microbiology.
Hall, W. C., Crowell, T. P., Watts, D. M., Barros, V. L. R., Kruger, H.,Pinheiro, F. & Peters, C. J. (1991). Demonstration of yellow fever anddengue antigens in formalin-fixed paraffin-embedded human liver byimmunohistochemical analysis. Am J Trop Med Hyg 45, 408–417.
Halstead, S. B. (1979). In vivo enhancement of dengue virus infectionin rhesus monkeys by passively transferred antibody. J Infect Dis 140,527–533.
Halstead, S. B. (1988). Pathogenesis of dengue: challenges tomolecular biology. Science 239, 476–481.
Hofmann, S., Grasberger, H., Jung, P., Bidlingmaier, M., Vlotides, J.,Janssen, O. E. & Landgraf, R. (2002). The tumour necrosis factor-alpha induced vascular permeability is associated with a reduction ofVE-cadherin expression. Eur J Med Res 7, 171–176.
Hordijk, P. L., Anthony, E., Mul, F. P., Rientsma, R., Oomen, L. C. &Roos, D. (1999). Vascular-endothelial-cadherin modulates endothe-lial monolayer permeability. J Cell Sci 112, 1915–1923.
Huang, Y. H., Lei, H. Y., Liu, H. S., Lin, Y. S., Chen, S. H., Liu, C. C. &Yeh, T. M. (2003). Tissue plasminogen activator induced by denguevirus infection of human endothelial cells. J Med Virol 70, 610–616.
Innis, B. L. (1995). Dengue and dengue hemorrhagic fever. In ExoticViral Infections, p. 103. Edited by J. S. Porterfield. London: Chapman& Hall.
Jessie, K., Fong, M. Y., Devi, S., Lam, S. K. & Wong, K. T. (2004).Localization of dengue virus in naturally infected human tissues, byimmunohistochemistry and in situ hybridization. J Infect Dis 189,1411–1418.
Killen, H. & O’Sullivan, M. A. (1993). Detection of dengue virus by insitu hybridization. J Virol Methods 41, 135–146.
Krishnamurti, C., Peat, R. A., Cutting, M. A. & Rothwell, S. W. (2002).Platelet adhesion to dengue-2 virus-infected endothelial cells. Am JTrop Med Hyg 66, 435–441.
Kurane, I., Kontny, U., Janus, J. & Ennis, F. A. (1990). Dengue-2 virusinfection of human mononuclear cell lines and establishment ofpersistent infections. Arch Virol 110, 91–101.
Kurane, I., Innis, B. L., Nimmannitya, S., Nisalak, A., Meager, A.,Janus, J. & Ennis, F. A. (1991). Activation of T lymphocytes in dengue
virus infections. High levels of soluble interleukin 2 receptor, soluble
CD4, soluble CD8, interleukin 2, and interferon-gamma in sera of
children with dengue. J Clin Invest 88, 1473–1480.
Lampugnani, M. G., Resnati, M., Raiteri, M., Pigott, R., Pisacane, A.,Houen, G., Ruco, L. P. & Dejana, E. (1992). A novel endothelial-
specific membrane protein is a marker of cell-cell contacts. J Cell Biol
118, 1511–1522.
Leitmeyer, K. C., Vaughn, D. W., Watts, D. M., Salas, R., Villalobos deChacon, I., Ramos, C. & Rico-Hesse, R. (1999). Dengue virus
structural differences that correlate with pathogenesis. J Virol 73,
4738–4747.
Libraty, D. H., Young, P. R., Pickering, D., Endy, T. P., Kalayanarooj, S.,Green, S., Vaughn, D. W., Nisalak, A., Ennis, F. A. & Rothman, A. L.(2002). High circulating levels of the dengue virus nonstructural
protein NS1 early in dengue illness correlate with the development of
dengue hemorrhagic fever. J Infect Dis 186, 1165–1168.
Malkovsky, M., Loveland, B., North, M., Asherson, G. L., Gao, L.,Ward, P. & Fiers, W. (1987). Recombinant interleukin-2 directly
augments the cytotoxicity of human monocytes. Nature 325, 262–265.
McIntyre, T. M. (2000). Molecular mechanisms of the inflammatory
response. In New Frontiers in Vascular Biology. Thrombosis and
Hemostatis, Edited by K. Suzuki, Y. Ikeda & I. Maruyama. Osaka:
Eibun Press.
McIntyre, T. M., Zimmerman, G. A., Satoh, K. & Prescott, S. M.(1985). Cultured endothelial cells synthesize both platelet-activating
factor and prostacyclin in response to histamine, bradykinin, and
adenosine triphosphate. J Clin Invest 76, 271–280.
McIntyre, T. M., Modur, V., Prescott, S. M. & Zimmerman, G. A.(1997). Molecular mechanisms of early inflammation. Thromb
Haemost 78, 302–305.
McKenzie, J. A. & Ridley, A. J. (2007). Roles of Rho/ROCK and MLCK
in TNF-alpha-induced changes in endothelial morphology and
permeability. J Cell Physiol 213, 221–228.
Monaco, C., Andreakos, E., Young, S., Feldmann, M. & Paleolog, E.(2002). T cell-mediated signaling to vascular endothelium: induction
of cytokines, chemokines, and tissue factor. J Leukoc Biol 71, 659–668.
Mukerjee, R., Chaturvedi, U. C. & Dhawan, R. (1995). Dengue virus-
induced human cytotoxic factor: production by peripheral blood
leucocytes in vitro. Clin Exp Immunol 102, 262–267.
Murgue, B., Roche, C., Chungue, E. & Deparis, X. (2000). Prospective
study of the duration and magnitude of viraemia in children
hospitalised during the 1996–1997 dengue-2 outbreak in French
Polynesia. J Med Virol 60, 432–438.
Navarro, P., Caveda, L., Breviario, F., Mandoteanu, I., Lampugnani,M. G. & Dejana, E. (1995). Catenin-dependent and -independent
functions of vascular endothelial cadherin. J Biol Chem 270,
30965–30972.
O’Sullivan, M. A. & Killen, H. M. (1994). The differentiation state of
monocytic cells affects their susceptibility to infection and the effects
of infection by dengue virus. J Gen Virol 75, 2387–2392.
Putnak, R., Feighny, R., Burrous, J., Cochran, M., Hackett, C., Smith, G.& Hoke, C. (1991). Dengue-1 virus envelope glycoprotein gene
expressed in recombinant baculovirus elicits virus-neutralizing anti-
body in mice and protects them from virus challenge. Am J Trop Med
Hyg 45, 159–167.
Rabiet, M. J., Plantier, J. L., Rival, Y., Genoux, Y., Lampugnani, M. G. &Dejana, E. (1996). Thrombin-induced increase in endothelial
permeability is associated with changes in cell-to-cell junction
organization. Arterioscler Thromb Vasc Biol 16, 488–496.
Rico-Hesse, R., Harrison, L. M., Salas, R. A., Tovar, D., Nisalak, A.,Ramos, C., Boshell, J., de Mesa, M. T., Nogueira, R. M. & Rosa, A. T.
Permeability of dengue virus-infected endothelial cells
http://vir.sgmjournals.org 651

Downloaded from www.microbiologyresearch.org by
IP: 54.90.14.242
On: Mon, 03 Oct 2016 07:50:14
(1997). Origins of dengue type 2 viruses associated with increasedpathogenicity in the Americas. Virology 230, 244–251.
Rothman, A. L. & Ennis, F. A. (1999). Immunopathogenesis of denguehemorrhagic fever. Virology 257, 1–6.
Scott, R. M., Nisalak, A., Cheamudon, U., Seridhoranakul, S. &Nimmannitya, S. (1980). Isolation of dengue viruses from peripheralblood leukocytes of patients with hemorrhagic fever. J Infect Dis 141,1–6.
Shaio, M. F., Cheng, S. N., Yuh, Y. S. & Yang, K. D. (1995). Cytotoxicfactors released by dengue virus-infected human blood monocytes.J Med Virol 46, 216–223.
Steinberg, M. S. & McNutt, P. M. (1999). Cadherins and their
connections: adhesion junctions have broader functions. Curr Opin
Cell Biol 11, 554–560.
Tretiach, M., van Driel, D. & Gillies, M. C. (2003). Transendothelial
electrical resistance of bovine retinal capillary endothelial cells is
influenced by cell growth patterns: an ultrastructural study. Clin
Experiment Ophthalmol 31, 348–353.
Vaughn, D. W., Green, S., Kalayanarooj, S., Innis, B. L., Nimmannitya, S.,Suntayakorn, S., Endy, T. P., Raengsakulrach, B., Rothman, A. L. &
other authors (2000). Dengue viremia titer, antibody response pattern,
and virus serotype correlate with disease severity. J Infect Dis 181, 2–9.
B. E. Dewi, T. Takasaki and I. Kurane
652 Journal of General Virology 89
Copyright © 2022 FDOKUMEN




















