
Pharmacological Strategies to Overcome HER2 Cross-Talk and Trastuzumab Resistance
Upload
independentCategory
view
4download
0
1
Down-regulation of HER2 by RIG1 Involves the PI3K/Akt Pathway in Ovarian Cancer Cells Chien-Chih Ou1, Shih-Chung Hsu2, Yin-Hui Hsieh3, Wan-Ling Tzou3, Tzu-Chao Chuang4,5, Jah-Yao
Liu6*, and Ming-Ching Kao3, 7*
1Graduate Institute of Medical Sciences and 3Graduate Institute of Biochemistry, National Defense
Medical Center, Taipei, Taiwan 2The Graduate Institute of Medical Sciences, Taipei Medical University, Taipei, Taiwan
4Department of Chemistry and 5Graduate Institute of Life Sciences, Tamkang University, Tamsui, Taipei, Taiwan
6Department of Obstetrics & Gynecology, Tri-Service General Hospital, Taipei, Taiwan 7Graduate Institute of Clinical Medical Science, China Medical University College of Medicine,
Taichung, Taiwan
Running Title: Down-regulation of HER2 by RIG1 *Corresponding author: Dr. Ming-Ching Kao, Graduate Institute of Clinical Medical Science, China Medical University College of Medicine, No. 91 Hsueh-Shih Road, Taichung, Taiwan 40402, ROC, E-Mail: [email protected]; or Dr. Jah-Yao Liu, Department of Obstetrics & Gynecology, Tri-Service General Hospital, No. 325, Sec. 2, Cherng-Kung Road, Neihu, Taipei, Taiwan 114, ROC.
1
© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
Carcinogenesis Advance Access published January 3, 2008 by guest on M
arch 18, 2014http://carcin.oxfordjournals.org/
Dow
nloaded from

2
Abstract
Interferon-γ (IFN-γ) is known to down-regulate HER2 oncoprotein (p185HER2 or briefly p185) in
prostate cancer cells. We demonstrate that the IFN-γ-induced retinoid-inducible gene 1 (RIG1) acts as
a transrepressor of p185. Furthermore, we exhibit that RIG1 down-regulates the activated
(phosphorylated) form of p185 and PI3K/Akt and the mammalian target of rapamycin (mTOR),
downstream substrates of HER2. We also elucidate that heregulin (HRG) specifically restores the
activation of p185 and Akt after their activities are reduced by RIG1. Additionally, expression of
vascular endothelial growth factor (VEGF) increases through the HER2 and Akt/mTOR signaling
pathways, indicating that VEGF is down-regulated by RIG1 within the cell. These findings suggest
that RIG1 plays a role in IFN-γ-mediated therapy by down-regulating p185 and its downstream
PI3K/Akt/mTOR/VEGF signaling pathway. These results may provide a new therapeutic mechanism
for the clinical use of IFN-γ and RIG1.
2
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

3
Introduction
Interferon-γ (IFN-γ), one of three IFN cytokines, inhibited the proliferation and regulation of
oncogenes and tumor suppressor genes (1, 2). Receptor-bound IFN-γ induces JAK1 and JAK2
phosphorylation and activates STAT1, resulting in translocation to the nucleus and the turning on of
downstream genes. Although many actions of IFN-γ have been studied, the underlying mechanisms
are still not well known.
Retinoid-inducible gene 1 (RIG1), also known as tazarotene-induced gene 3 (TIG3) or retinoic acid
receptor responder 3 (RARRES3), belongs to the class II tumor suppressor gene HREV107 family and
is one of the IFN-γ downstream genes (3-5). RIG1 is expressed in high levels in well-differentiated
skin and colon mucosal tissues but at low levels in cancer tissues derived from the skin, liver,
colorectum, and biliary tract (6, 7). Overexpression of RIG1 suppresses colony formation in T47D
breast cancer cells and HaCaT keratinocytes. Ectopic RIG1 expression leads to cellular apoptosis and
suppression of growth in cancer cells and human keratinocytes (3, 8, 9, 10). RIG1 also facilitates the
terminal stages in keratinocyte differentiation by activating type I transglutaminase (11). Previous
studies have demonstrated that RIG1 decreases protein levels and activities of Ras, one of the HER2
downstream proteins, while interacting with Ras protein in the cytoplasmic compartment in HtTA
cervical cancer cells (9, 12). However, the molecular mechanisms responsible for the effects of RIG1
remain unclear.
3
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

4
The HER2 gene, also known as neu or erbB2, encodes a 185 kDa transmembrane receptor tyrosine
kinase belonging to the epidermal growth factor receptor (EGFR) family (13-18). Overexpression of
p185 is found in about 30% of human breast cancers and in many other cancer types (19, 20). The
p185 phosphorylates downstream substrates and activates a variety of signaling cascades, including
the phosphatidylinositol-3 kinase (PI3K)/Akt and Ras/MAPK pathways. These regulatory signal
cascades promote cell survival and tumor growth and metastasis (20-22).
We demonstrated previously that SV40 LT425 (LT425) acts as a transforming and metastasis
suppressor of HER2 oncogene (23, 24). Microarray and proteomic analysis revealed that both of
IFN-γ and RARRES3 genes are up-regulated in LT425 stable transfectants (Kao et al., unpublished
data). Through a literature search and database comparison, we found that IFN-γ down-regulates p185
expression in prostate cancer cells and induces RIG1 expression in ovarian cancer cells (2, 5). Thus,
we hypothesize that IFN-γ-induced RIG1 may contribute to repression of HER2. We also investigate
the effects of RIG1 on the HER2 signaling cascade.
Materials and Methods
Cell Culture and Plasmids — SKOV-3, OVCAR-3, and TOV-21G human ovarian cancer cell lines
were grown in McCoy’s 5A, RPMI 1640 (Gibco BRL), and MCDB 105/Medium 199
(Sigma-Aldrich) media, respectively, supplemented with 10-20% fetal bovine serum (FBS, HyClone).
BT-474, MCF-7/HER (MCF-7 of a HER2-transfected stable line), and MCF-7 human breast cancer
4
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

5
cell lines were grown in DMEM/F12 medium supplemented with 10% FBS. Cells were grown at 37
°C in a humidified incubator with 5% CO2. The plasmids pRIG1-EGFP and pRIG1-myc were kindly
provided by Dr. SY Jiang (Department of Education and Research, Buddhist Tzu Chi General
Hospital, Taipei, Taiwan, ROC).
Cell Transfection — A liposome-mediated transfection method was used to transfect cells. In
brief, siRNAs (Dharmacon and Ambion) or plasmids were diluted in Opti-MEM (Gibco BRL)
medium and then mixed with Lipofectamine 2000 (Gibco BRL) at room temperature for 20 min. The
lipoplex complex was added to the cells for 6 h, and the cells refreshed with complete medium.
Western blot analysis — Cell lysates were prepared and sedimented as described previously (23).
The supernatant was used for SDS-PAGE analysis. Sixty micrograms of each cell sample was loaded
onto the NuPAGE 4-12% Bis-Tris Gel (NP0322, Invitrogen). After electrophoresis, the gels were
blotted onto nitrocellulose (NC) membranes. The NC membranes were subjected to blocking with 5%
skim milk in TBST buffer (10 mM Tris, pH 8.0, 150 mM NaCl, 0.1% Tween-20) for 2 h at room
temperature. The membranes were then incubated with the primary anti-neu antibody (Ab-3; 1:1000
dilution; Oncogene Science), anti-phospho-neu (Tyr1248) antibody (Ab-18; 1:500 dilution;
NeoMarkers), anti-Akt antibody (1:1000 dilution; Cell Signaling), anti-phospho-Akt (Ser473)
antibody (1:500 dilution; Cell Signaling), anti-GFP antibody (Santa Cruz Biotechnology), anti-PI3K
antibody (Cell Signaling), anti-phospho-(Tyr) p85 PI3K antibody (Cell Signaling), anti-mTOR
5
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

6
antibody (Cell Signaling), anti-phospho-mTOR (Ser2448) antibody (Cell Signaling), anti-β-actin
(Chemicon, 1:5000 dilution) antibody, or other antibodies (Cell Signaling) in fresh 5% skim
milk-TBST buffer for 1 h at room temperature or 4 °C overnight. The membranes were washed and
incubated with the secondary antibody of HRP-conjugated goat anti-mouse or goat anti-rabbit IgG
(1:1000 dilution; Santa Cruz Biotechnology). Visualization was performed in an Amersham ECL
chemiluminescence system. Bands were quantified by Adobe Photoshop software. IFN-γ, heregulin
(HRG), and all chemicals were purchased from Sigma-Aldrich.
RNA isolation and reverse transcription polymerase chain reaction (RT-PCR) — Total RNAs were
extracted using TRIzol solution (Invitrogen). cDNA was prepared by incubation at 37 °C for 1 h in 20
μl of a mixture containing 3 μg total RNA, 1 μl Moloney murine leukemia virus reverse transcriptase
(Promega), 1 μl dNTP, 1 μl oligo-dT, and 1 U RNaseout recombinant RNase inhibitor. PCR was
performed in 25 μl of a reaction mixture containing 100 ng cDNA, and 200 nM each of HER2 (sense:
5′-CCAGAGCAAGAGAGGTATCC-3′; antisense: 5′-CTGTGGTGGTGAAGCTGTAG-3′), RIG1
(sense: 5′-ACCAGACCTCCTCTTGGC-3′; antisense: 5′-GAAGGGGCAGATGGCTGT-3′), or
β-actin (sense: 5′-CCAGAGCAAGAGAGGTATCC-3′; antisense:
5′-CTGTGGTGGTGAAGCTGTAG-3′) primers. The PCR products were quantified by ethidium
bromide staining on a 0.8% agarose gel.
6
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

7
Quantitative PCR — Quantitative PCR was performed on a LightCycler (Roche Diagnostics)
using SYBR Green I as a double-strand DNA-specific binding dye and continuous fluorescence
monitoring. Amplification was carried out in a total volume of 20 μl containing 0.5 μM of each
primer, 4 μM MgCl2, 2 μl LightCycler DNA Master SYBR Green I (containing 1.25 U Taq
polymerase, 10× Taq buffer [500 μM KC1 and 100 μM Tris-HC1, pH 8.3], 2 μM of each dNTP, and
10× SYBR Green; Roche Diagnostics), and 1 μl of cDNA prepared as described above. The PCR
primers were identical to those used for semiquantitative RT-PCR. Melting curve analysis of
amplification products was performed at the end of each PCR reaction by cooling the samples to
40 °C and then increasing the temperature to 95 °C at 0.2 °C/s.
Fluorescence-activated Cell Sorter (FACS) analysis — The FACS analysis of p185 was performed
as described (25). In brief, PE-conjugated anti-HER2 monoclonal antibody (Becton Dickinson) was
diluted with washing buffer (0.1% NaN3 and 1.0% FBS in PBS) and the cell pellets were resuspended
to a concentration of 2 × 107 cells/ml. The cell–antibody complex were incubated at 4 °C for 30 min
in the dark and then washed twice with washing buffer. The supernatant was removed after each
centrifugation. Five hundred microliters of washing buffer was added to transfer the cell pellets to
Falcon round bottom tubes (Becton Dickinson), and the cells were analyzed using either a FACS
Caliber or FACS Vantage SE (Becton Dickinson).
7
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

8
ELISA Assay— Sorted cells (1 × 106) were plated in a 60-mm tissue culture plate and allowed to
attach overnight. The cells were then washed and incubated in serum-free medium for 24 h. The
serum-free conditioned medium was collected and centrifuged at 1000 x g for 15 min to remove
debris. ELISA assay was performed using the VEGF Quantikine ELISA kit (R&D Systems) (26).
HER2 Promoter Activity Assay — Cells were cotransfected with pNulit (kindly provided by Dr.
MC Hung, Department of Molecular and Cellular Oncology, University of Texas MD Anderson
Cancer Center, Houston, Texas) and pCMV-β-gal plasmids and various amounts of pRIG1-myc. Cell
lysates were prepared before the luciferase activity assay by following the manufacturer’s (Promega)
instructions. The measured luciferase activity levels were normalized against the β-galactosidase
(β-gal) standard. The β-gal activity was measured by mixing equal amounts of cell extract and the 2×
assay buffer (120 mM Na2HPO4, 80 mM NaH2PO4, 2 mM MgCl2, 100 mM β-mercaptoethanol, and
1.33 mg/ml orthonitrophenyl galactoside). After incubation at 37°C, the enzyme activity of β-gal was
determined at A420 nm (27).
Statistical Analysis — Data were analyzed by using the paired t-test for comparison of
independent means.
8
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

9
Results
Down-regulation of p185 by IFN-γ induced RIG1
IFN-γ has been shown to have inhibitory effects on p185 in prostate cancer cells (2). Thus, it is
reasonable to ask whether IFN-γ down-regulates p185 in ovarian and breast cancer cell lines. As
expected, IFN-γ down-regulated p185 expression in SKOV-3, OVCAR-3, and TOV-21G ovarian
cancer cell lines, as well as in BT-474, MCF-7/HER, and MCF-7 breast cancer cell lines (Figure 1A).
IFN-γ also down-regulated p185 expression in a dose- and time-dependent manner in
HER2-overexpressing SKOV-3 cells, as shown in the western blot analysis (Figures 1B and 1C). We
then examined whether IFN-γ could up-regulate RIG1 gene expression in SKOV-3 cells. Since there
is no effective anti-RIG1 antibody available, we used a quantitative RT-PCR approach to measure the
RIG1 mRNA levels. As expected, increasing RIG1 mRNA levels after treatment with increasing
doses of IFN-γ were observed. The level of RIG1 mRNA increased more than two folds after
treatment with 1000 U/ml of IFN-γ (Figure 2A). The suppressive effects of IFN-γ on p185 expression
by induction of RIG1 were examined by treating several ovarian cancer cell lines with different doses
of IFN-γ (100 and 1000 U/ml). After IFN-γ-treatment for 24 h, cells were transfected with RIG1
siRNA or negative control siRNA. In cells treated with 1000 U/ml of IFN-γ, RIG1 siRNA restored the
IFN-γ-mediated repression of p185 level to 290% in TOV-21G and 51% in OVCAR-3 cell lines. In
addition, the p185 level in SKOV-3 cells could be restored by 40% by RIG1 siRNA with 100 U/ml of
IFN-γ treatment (Figure 2B). These results were also revealed by different set of RIG1 siRNA
(Supplement 1). Taken together, these data suggest that IFN-γ down-regulates p185 by induction of
RIG1 (Figures 1 and 2).
To further explore whether RIG1 is an important suppressor of p185 in SKOV-3 cells, the protein
levels of p185 detected with the PE-conjugated anti-HER2 monoclonal antibody were analyzed by
FACS. We transfected the pEGFP vector only, or pRIG1–EGFP into SKOV-3 cells, and then
9
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

10
measured the red fluorescence (p185) and green fluorescence (pEGFP or pRIG1–EGFP) in the cells.
RIG1 could significantly down-regulate p185 expression by about 52% after 24 h of transfection of
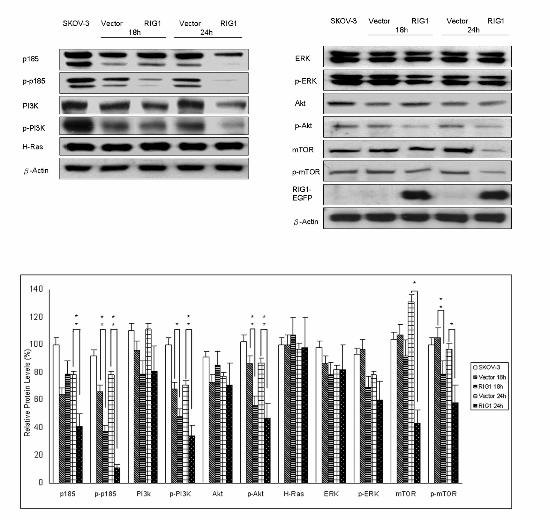
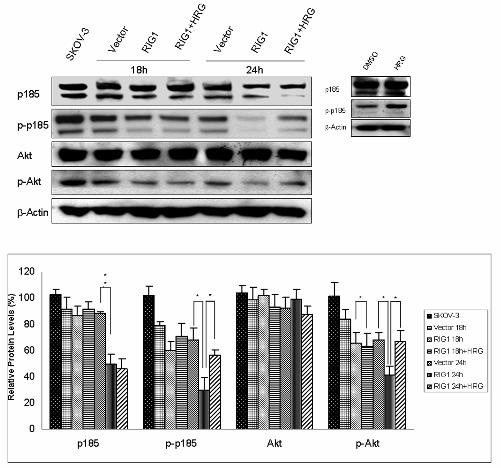
RIG1 (Figure 3A). At the same time, we isolated the successfully transfected RIG1-overexpressing
cells 18 h and 24 h after plasmid transfection (Figure 4A). Western blot analysis demonstrated that
these FACS-sorted RIG1-overexpressing cells were able to effectively reduce the p185 level by 24 h
(Figure 4A). In addition, phospho-p185 (the activated form of p185) was remarkably down-regulated
by RIG1 at both 18 and 24 h (Figure 4A). These results suggest that RIG1 down-regulates p185
protein levels and suppresses p185 activation.
RIG1 represses the HER2 gene activity
To unravel the mechanisms of down-regulation of p185 by RIG1, we assayed the mRNA levels of
different sorted transcripts. As shown in Figure 3B, consistent with the p185 protein levels, the HER2
mRNA levels were down-regulated by RIG1 at 24 h but not at 18 h. RIG1 obviously reduced HER2
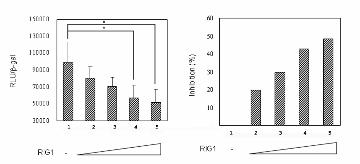
RNA by more than 50% at 24 h (Figure 3B, right). The transcriptional activity of the HER2 promoter
was also analyzed. In transient expression assays, using NIH3T3 (Figure 3C) and SKOV-3 (Figure
3D) as recipient cells, cotransfection of pNulit and pCMV-β-gal and different amounts of pRIG1-myc
for 24 h led to a significant decrease in luciferase activity (Figures 3C and 3D, left), which was
normalized by β-gal. The HER2 promoter inhibitory rates increased in relation to an increasing
amount of RIG1, and an inhibition rate of more than two times was observed at a ratio of
pNulit:pRIG1-myc as low as 1:1 (Figures 3C and 3D, right). Taken together, these data suggest that
down-regulation of p185 protein by RIG1 occurs because of repression of HER2 promoter activity.
Down-regulation of phospho-PI3K/Akt and phospho-mTOR by IFN-γ and RIG1
Among HER2 stimulated intracellular signaling pathways, two main pathways, the Ras/MAPK
and the PI3K/Akt cascades, were shown to be activated by HER2 (22). RIG1 was reported to inhibit
the Ras/MAPK pathway by suppressing the activity of Ras and Ras protein in HtTA cervical cancer
10
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

11
cells (12). Here, we examined whether RIG1 could interfere with the Ras/MAPK pathway in ovarian
cancer cells. As demonstrated in Figure 4A, both Ras and ERK proteins were not altered in
RIG1-overexpressing cells. The activation of ERK was not significantly reduced after ectopic
expression of RIG1 (Figure 4A). We then investigated if RIG1 affected the PI3K/Akt signaling
cascade. After 18 and 24 h of transfection with RIG1, phospho-PI3k, phospho-Akt and
phospho-mTOR were all dramatically down-regulated (p < 0.01). More than 40% of inhibition rate
was observed at 24 h (Figure 4A). In addition, a new finding that RIG1 has an inhibitory effect on
mTOR protein (Figure 4A) suggests that mTOR is another target for RIG1 inhibition. These data
clearly demonstrate that RIG1 exerts inhibitory effects on
phospho-PI3K/phospho-Akt/phospho-mTOR signaling pathway. This is the first report showing that
RIG1 could affect PI3K/Akt pathway.
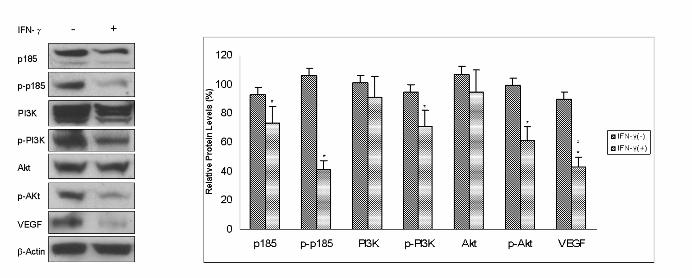
To further evaluate the effects of IFN-γ on the PI3K/Akt pathway, SKOV-3 cells were treated with
1000 U/ml of IFN-γ. After 24 h of IFN-γ administration, the expression of p185, phospho-p185,
phospho-PI3K, phospho-Akt, and VEGF all remarkably decreased, but the protein levels of PI3K and
Akt did not change (Figure 4B). These results were identical to those obtained for RIG1 ectopic
expression, as shown in Figure 4A. In summary, these data demonstrate that both IFN-γ and RIG1 can
abrogate the HER2 and PI3K/Akt pathways.
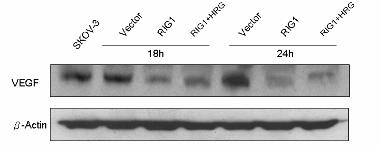
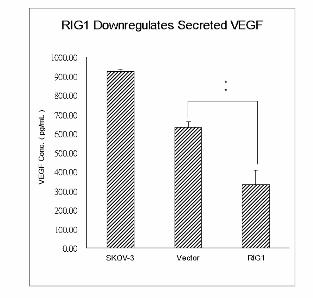
Down-regulation of VEGF by RIG1
HER2-overexpressing tumors tend to be more angiogenic than other tumors (28). VEGF is one of
the most potent inducers of angiogenesis (28). The HER2 and PI3K/Akt signaling pathway has been
implicated in the regulation of VEGF through activation of mTOR/p70S6K (26). Thus, we proposed
that RIG1 down-regulated VEGF by inactivating mTOR. As expected, western blot analysis of
intracellular VEGF demonstrated that RIG1 down-regulated VEGF at 18 and 24 h after transfection
(Figure 5A). To further explore the participation of VEGF in the medium, we used ELISA to quantify
11
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

12
and compare the VEGF secreted into serum-free conditioned medium after 24 h of incubation with
different transfectants. As shown in Figure 5B, RIG1 significantly reduced the secreted VEGF level
by about 50%, indicating that RIG1 could decrease VEGF expression by down-regulating the HER2
and PI3K/Akt/mTOR signaling pathway.
Heregulin restores the RIG1-induced decrease in phospho-p185 and phospho-Akt
HRG is a member of the neuregulin family of ligands that interact with and activate EGFR family
members. We found that RIG1 suppresses the activation of p185 and wondered whether HRG could
reactivate p185. As shown in Figure 6A, phospho-p185 was down-regulated by RIG1 at 24 h and
HRG restored phospho-p185 at this time point. Since RIG1 down-regulated both p185 levels and
phospho-p185 expression, HRG might not effectively restore the phosphorylation levels to the vector
control level. In addition, HRG restored the down-regulated phospho-Akt to the control level,
implying that RIG1 acts as a suppressor of the HER2 and PI3K/Akt pathway. HRG was added to the
cells before they were applied to FACS so VEGF expression would not have been increased by HRG
(Figure 5A). We also found p185 protein expression slightly decreased 24 h after addition of HRG
(Figure 6A, lower panel). This finding is consistent with a previous report that ligand can induce
endocytosis and degradation of HER2 (32).
Discussion
Type II IFNs possess antiproliferative effects in various tumor cell types. The binding of IFN-γ to
its cognate receptor activates the JAK/STAT signal transduction pathway, in which STAT1 molecules
are phosphorylated and translocated to the nucleus where they bind to IFN-γ-activated sites and
activate transcription of target genes. Here, we demonstrated that IFN-γ suppresses p185 expression in
both ovarian and breast cancer cell lines. The phosphoprotein levels of p185, PI3K, and Akt, and the
protein levels of p185 and VEGF were all down-regulated by IFN-γ treatment (Figure 4B). The
repressive effect of IFN-γ-induced RIG1 on p185 expression was verified by the RIG1 siRNA.
12
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

13
Treatment with 1000 U/ml of IFN-γ together with RIG1 siRNA restored the IFN-γ-mediated
repression of the p185 level to 290% in TOV-21G cells and to 51% in OVCAR-3 cells (Figure 2B) ,
whereas p185 was only partially restored by RIG1 siRNA in SKOV-3 cells after treatment with 1000
U/ml of IFN-γ (data not shown). This phenomenon was also observed in the HER2-overexpressing
breast cancer BT-474 cell line when treated with 1000 U/ml of IFN-γ (data not shown). However, the
p185 level in both SKOV-3 and BT-474 cells could be restored by RIG1 siRNA under a low dose of
IFN-γ (100 U/ml). Therefore, inhibition of RIG1 appeared insufficient to restore the repression of
p185 expression after a high dose of IFN-γ treatment. It has been reported that down-regulation of
p185 by a high dose (5000 U/ml) of IFN-γ occurs through the interaction of phosphorylated STAT1
with p300 (2). Thus, RIG1 may play an important role after IFN-γ treatment, but other mechanisms
may be involved in the regulation of p185, especially with a high dose of IFN-γ.
Using a transient transfection method and cell sorter, we isolated RIG1-overexpressing SKOV-3
cells. Inside these cells, both p185 protein and its phosphorylated form were down-regulated. We also
showed that repression of p185 protein was caused by the inhibition of HER2 promoter activities
(Figures 3C and 3D). Therefore, we propose that RIG1 may possess phosphatase activities, and the
interaction and colocalization of RIG1 with p185 in the cytoplasmic compartment may contribute to
the decreased p185 activation (data not shown). In addition to down-regulation of phospho-p185, the
amount of activated PI3K, Akt and mTOR decreased 18 h after transfection of RIG1 (Figure 4A).
Thus, IFN-γ induced RIG1 may play an important role in the HER2 and PI3K/Akt pathways (Figure
6B).
RIG1 belongs to the HREV107 family of class II tumor suppressor genes. A notable feature of
class II tumor suppressor is loss of the growth-constraining function in tumor cells because of blocked
expression, but not because of DNA mutation or deletion, such as p53 (29, 30). Previous studies have
shown the induction of RIG1 protein by retinoids (3, 4), but the mechanisms of induction of RIG1 by
IFN-γ are not well understood. Evidence suggests that the RIG1 promoter contains a retinoic acid
13
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

14
response element (RARE), which is also referred to as a putative p53 response element (p53RE) (31).
The presence of p53RE suggests that the class II tumor suppressor, RIG1, might be a direct
transcriptional target of the class I tumor suppressor p53. However, SKOV-3 is a p53-null cell line,
meaning that the regulation of RIG1 by IFN-γ is p53-independent. Treatment of A27/80 and
OVCAR-3 human ovarian carcinoma cells with IFN-γ can stimulate HREV107-1 expression, which is
directly related to activation of IFN regulatory factor-1 (IRF-1), indicating that HREV107-1 is a target
of IRF-1 and is involved in IFN-γ-induced cell death (5). In this study, we demonstrated that RIG1
mRNA levels increased by more than two folds after administration of IFN-γ for 2 h (Figure 2A). This
result confirmed an earlier report that IRF-1 is detectable within 3 h after IFN-γ administration (5).
Thus, the mechanisms responsible for the IFN-γ induction of RIG1 probably involve IRF-1 and
should be investigated further.
It has been hypothesized that RIG1 is involved in a negative feedback loop that normally controls
the level of MAPK activation (33). Overexpression of RIG1 down-regulates MAPK activities in
HtTA cervical and T47D breast cancer cells, and enhances MAPK-mediated suppression of RIG1
(33). In our study, the activity of ERK is not significantly reduced by RIG1. Therefore, we focused on
PI3K/Akt pathway. As shown in Figure 4A, PI3k/Akt activities were dramatically reduced (p < 0.01)
after ectopic expression of RIG1. In SKOV-3 cells, Akt signaling is dominantly activated as a result
of increased HER2 activation and in the absence of p53 and PTEN (34, 35, 43, 44). p53 and PTEN are
known to repress PI3K/Akt activities (34, 35, 43, 44). Although MAPK is also activated in ovarian
carcinoma cells, it is not the major pathway for RIG1-mediated suppression of HER2 signaling in
SKOV-3 cells in our study.
HER2 overexpression increases the metastatic potential of human cancer cells (20, 21). Patients
with HER2-overexpressing tumors have increased incidences of metastasis and a poor survival rate
compared with HER2 normal patients (19). Angiogenesis is a key component of cancer metastasis
(36) and is tightly controlled under normal physiological conditions. However, in pathological
14
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

15
diseases such as cancer, the fine balance between proangiogenic and antiangiogenic factors is
disrupted. VEGF is one of the most potent inducers of angiogenesis and induces endothelial cell
proliferation and migration (28). VEGF expression in human breast cancers correlates with increased
microvessel density and reduced survival (37). HER2 has been implicated in the regulation of VEGF,
which is activated by the mTOR/p70S6K pathway. mTOR is a 289 kDa phosphoinositide
kinase-related serine/threonine kinase (38). Through the formation of multimolecular complexes,
rictor or raptor, the evolutionarily conserved TOR pathway controls an array of fundamental cell
functions, such as translation initiation, protein stability, transcription of ribosome and stress response
genes, ribosomal biogenesis, and tRNA synthesis, thereby playing a central role in the regulation of
cell growth, proliferation, and survival (39, 40). Several pharmaceutical and biotechnology companies
are actively pursuing the clinical development of inhibitors of the mTOR pathway for cancer
therapies, such as rapamycin. Our data show that RIG1 effectively down-regulates both
phospho-mTOR and mTOR protein (Figure 4A), suggesting mTOR as a target for RIG1 inhibitory
effects. mTOR localizes in the endoplasmic reticulum and Golgi apparatus (GA) membranes (41, 42).
Coincidentally, RIG1 also resides in the GA membrane (12). Further investigations may be needed to
understand whether RIG1 associates with mTOR in the GA membrane and whether this regulates its
protein level. In addition, activation of mTOR/p70S6K increases angiogenesis and spontaneous
metastasis by increasing VEGF synthesis. We found that RIG1 significantly reduced VEGF
expression in serum-free conditioned medium and intracellular protein levels (Figure 5), implicating a
role for RIG1 in decreasing angiogenesis in tumor growth. This is the first report that RIG1 exerts its
function to down-regulate mTOR and VEGF (Figure 6B). However, the underlying mechanisms
remain to be elucidated.
Acknowledgements
15
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

16
We wish to thank Drs. SY Jiang (Department of Education and Research, Buddhist Tzu Chi
General Hospital, Taipei, Taiwan, ROC) and MC Hung (Department of Molecular and Cellular
Oncology, University of Texas MD Anderson Cancer Center, Houston, Texas) for providing plasmids
as well as Dr. JJ Yen (Institute of Biomedical Sciences, Academia Sinica, Taipei, Taiwan, ROC) for
assisting FACS sorting experiments. This work was supported in part by the “C.Y. Foundation” and
the National Science Council, Taiwan, ROC, granted to MC Kao (NSC 95-2320-B-039-047).
Reference 1. Hobeika, A.C., Etienne, W., Cruz, P.E., Subramaniam, P.S., and Johnson, H.M. (1998) IFN-γ
induction of p21WAF1 in prostate cancer cells: role in cell cycle, alteration of phenotype and invasive potential. Int J Cancer 77, 138-145
2. Kominsky, S., Hobeika, A.C., Lake, F.A., Torres, B.A., and Johnson, H.M. (2000) Down-regulation of neu/HER-2 by interferon-γ in prostate cancer cells. Cancer Res 60, 3904-3908
3. DiSepio, D., Ghosn, C., Eckert, R.L., Deucher, A., Robinson, N., Duvic, M., Chandraratna, R.A., and Nagpal, S. (1998) Identification and characterization of a retinoid-induced class II tumor suppressor/growth regulatory gene. Proc Natl Acad Sci U S A. 95, 14811-14815
4. Huang, S.L., Shyu, R.Y., Yeh, M.Y., and Jiang, S.Y. (2000) Cloning and characterization of a novel retinoid-inducible gene 1(RIG1) deriving from human gastric cancer cells. Mol Cell Endocrinol 159, 15-24
5. Sers, C., Husmann, K., Nazarenko, I., Reich, S., Wiechen, K., Zhumabayeva, B, Adhikari, P., Schrode, K., Gontarewicz, A., and Schafer, R. (2002) The class II tumour suppressor gene H-REV107-1 is a target of interferon-regulatory factor-1 and is involved in IFNgamma-induced cell death in human ovarian carcinoma cells. Oncogene 21, 2829-2839
6. Duvic, M., Helekar, B., Schulz, C., Cho, M., DiSepio, D., Hager, C. DiMao, D., Hazarika, P., Jackson, B., Breuer-McHam, J., Young, J., Clayman, G., Lippman, SM., Chandraratna, R.A., Robinson, N.A., Deucher, A., Eckert, R.L., and Nagpal, S. (2000) Expression of a retinoid-inducible tumor suppressor, Tazarotene-inducible gene-3, is decreased in psoriasis and skin cancer. Clin Cancer Res 6, 3249-3259
7. Shyu, R.Y., Jiang, S.Y., Chou, J.M., Shih, Y.L., Lee, M.S., Yu, J.C. Chao, P.C., Hsu, Y.J., and Jao, S.W. (2003) RARRES3 expression positively correlated to tumour differentiation in tissues of colorectal adenocarcinoma. Br J Cancer 89, 146-151
8. Deucher, A., Nagpal, S., Chandraratna, R.A., DiSepio, D., Robinson, N.A., Dashti, S.R., and Eckert, R.L. (2000) The carboxy-terminal hydrophobic domain of TIG3, a class II tumor suppressor protein, is required for appropriate cellular localization and optimal biological activity. Int J Oncol 17, 1195-1203
9. Huang, S.L., Shyu, R.Y., Yeh, M.Y., and Jiang, S.Y. (2002) The retinoid-inducible gene I: effect on apoptosis and mitogen-activated kinase signal pathways. Anticancer Res 22, 799-804
10. Higuchi, E., Chandraratna, R.A., Hong, W.K., and Lotan, R. (2003) Induction of TIG3, a putative
16
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

17
class II tumor suppressor gene, by retinoic acid in head and neck and lung carcinoma cells and its association with suppression of the transformed phenotype. Oncogene 22, 4627-4635
11. Sturniolo, M.T., Dashti, S.R., Deucher, A., Rorke, E.A., Broome, A.M., Chandraratna, R.A. Keepers, T., and Eckert, R.L. (2003) A novel tumor suppressor protein promotes keratinocyte terminal differentiation via activation of type I transglutaminase. J Biol Chem 28, 48066-48073
12. Tsai, F.M., Shyu, R.Y., and Jiang, S.Y. (2006) RIG1 inhibits the Ras/mitogen-activated protein kinase pathway by suppressing the activation of Ras. Cell Signal 18, 349-58
13. Shih, C., Padhy, L.C., Murray, M., and Weinberg, R.A. (1981) Transforming genes of carcinomas and neuroblastomas introduced into mouse fibroblasts. Nature 290, 261-264
14. Coussens, L., Yang-Feng, T.L., Liao, Y.C., Chen, E., Gray, A., McGrath, J., Seeburg, P.H., Libermann, T.A., Schlessinger, J., and Francke, U. (1985) Tyrosine kinase receptor with extensive homology to EGF receptor shares chromosomal location with neu oncogene. Science 230, 1132-1139
15. King, C.R., Kraus, M.H., and Aaronson, S.A. (1985) Amplification of a novel v-erbB-related gene in a human mammary carcinoma. Science 229, 974-976
16. King, C.R., Swain, S.M., Porter, L., Steinberg, S.M., Lippman, M.E., and Gelmann, E.P. (1989) Heterogeneous expression of erbB-2 messenger RNA in human breast cancer. Cancer Res 49, 4185-4191
17. Yamamoto, T., Ikawa, S., Akiyama, T., Semba, K., Nomura, N., and Miyajima, N. Saito, T., and Toyoshima, K. (1986) Similarity of protein encoded by the human c-erb-B-2 gene to epidermal growth factor receptor. Nature 319, 230-234
18. Hung, M.C., Zhang, X., Yan, D.H., Zhang, H.Z., He, G.P., Zhang, T.Q., and Shi, D.R. (1992) Aberrant expression of the c-erbB-2/neu protooncogene in ovarian cancer. Cancer Lett 61, 95-103
19. Slamon, D.J., Clark, G.M., Wong, S.G., Levin, W.J., Ullrich, A., and McGuire, W.L. (1987) Human breast cancer: correlation of relapse and survival with amplification of the HER-2/neu oncogene. Science 235, 177-182
20. Yarden, Y., and Sliwkowski, M.X. (2001) Untangling the ErbB signalling network. Nat Rev Mol Cell Biol 2, 127-137
21. Yu, D., and Hung, M.C. (2000) Role of erbB2 in breast cancer chemosensitivity. BioEssay 22, 673-680
22. Hynes, N.E., and Lane, H.A. (2005) ERBB receptors and cancer: the complexity of targeted inhibitors. Nat Rev Cancer 5, 341-354
23. Kao, M.C., Liu, G.Y., Chuang, T.C., Lin, Y.S., Wuu, J.A., and Law, S.L. (1998) The N-terminal 178-amino-acid domain only of the SV40 large T antigen acts as a transforming suppressor of the HER-2/neu oncogene. Oncogene 16, 547-554
24. Chuang, T.C., Yu, Y.H., Lin, Y.S., Wang, S.S., and Kao, M.C. (2002) The N-terminal domain of SV40 large T antigen represses the HER2-mediated transformation and metastatic potential in breast cancers. FEBS Lett 511, 46-50
25. Menendez, J.A., Vellon, L., Mehmi, I., Oza, B.P., Ropero, S., Colomer, R., and Lupu, R. (2004) Inhibition of fatty acid synthase (FAS) suppresses HER2 (erbB-2) oncogene overexpression in cancer cells. Proc Natl Acad Sci USA 101, 10715-10720
26. Klos, K.S., Wyszomierski, S.L., Sun, M., Tan, M., Zhou, X., and Li, P. Yang, W., Yin, G., Hittelman, W.N., and Yu, D. (2006) ErbB2 increases vascular endothelial growth factor protein synthesis via activation of mammalian target of rapamycin/p70S6K leading to increased angiogenesis and spontaneous metastasis of human breast cancer cells. Cancer Res 15, 2028-2037
27. Xing, X., Wang, S.C., Xia, W., Zou, Y., Shao, R., Kwong, K.Y., Yu, Z., Zhang, S., Miller, S., Huang, L., and Hung, M.C. (2000) The ets protein PEA3 suppresses HER-2/neu overexpression and inhibits tumorigenesis. Nat Med 6, 189-195
28. Ferrara, N., and Davis-Smyth, T. (1997) The biology of vascular endothelial growth factor. Endocr Rev 18, 4–25
29. Krzyzosiak, W.J., Shindo-Okada, N., Teshima, H., Nakajima, K., and Nishimura, S. (1992)
17
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

18
Isolation of genes specifically expressed in flat revertant cells derived from activated ras-transformed NIH 3T3 cells by treatment with azatyrosine. Proc Natl Acad Sci USA 89, 4879–4883
30. Roder, K., Latasa, M.J., and Sul, H.S. (2002) Silencing of the mouse H-rev107 gene encoding a class II tumor suppressor by CpG methylation. J Biol Chem 277, 30543-30550
31. Jiang, S.Y., Wu, M.S., Chen, L.M., Hung, M.W., Lin, H.E., Chang, G.G., and Chang, T.C. (2005) Identification and characterization of the retinoic acid response elements in the human RIG1 gene promoter. Biochem Biophys Res Commun 331, 630-639
32. Citri, A., and Yarden, Y. (2006) EGF-ERBB signalling: towards the systems level. Nat Rev Mol Cell Biol 7, 505-516
33. Lotz, K., Kellner, T., Heitmann, M., Nazarenko, I., Noske, A., and Malek, A. Gontarewicz, A., Schäfer, R., and Sers, C. (2005) Suppression of the TIG3 tumor suppressor gene in human ovarian carcinomas is mediated via mitogen-activated kinase-dependent and -independent mechanisms. Int J Cancer 116, 894-902
34. Harris, S.L., and Levine, A.J. (2005) The p53 pathway: positive and negative feedback loops. Oncogene 24, 2899-2908
35. Gottlieb, T.M., Leal, J.F., Seger, R., Taya, Y., and Oren, M. (2002) Cross-talk between Akt, p53 and Mdm2: possible implications for the regulation of apoptosis. Oncogene 21, 1299-303
36. Folkman, J. (2002) Role of angiogenesis in tumor growth and metastasis. Semin Oncol 29, 15–18 37. Gasparini, G., Toi, M., Gion, M., Verderio, P., Dittadi, R., and Hanatani, M. Matsubara, I.,
Vinante, O., Bonoldi, E., Boracchi, P., Gatti, C., Suzuki, H., and Tominaga, T. (1997) Prognostic significance of vascular endothelial growth factor protein in node-negative breast carcinoma. J Natl Cancer Inst 89, 139–147
38. Shaw, R.J., and Cantley, L.C. (2006) Ras, PI(3)K and mTOR signalling controls tumour cell growth. Nature 441, 421-430
39. Sawyers, C.L. (2003) Will mTOR inhibitors make it as cancer drugs? Cancer Cell 4, 343-8 40. Hay, N., and Sonenberg, N. (2004) Upstream and downstream of mTOR. Genes Dev 18, 1926-45 41. Drenan, R.M., Liu, X., Bertram, P.G., and Zheng, X.F. (2004) FKBP12-rapamycin-associated
protein or mammalian target of rapamycin (FRAP/mTOR) localization in the endoplasmic reticulum and the Golgi apparatus. J Biol Chem 279, 772-778
42. Liu, X., and Zheng, X,F. (2007) Endoplasmic reticulum and Golgi localization sequences for mammalian target of rapamycin. Mol Biol Cell 18, 1073-1082
43. Wang, H.Q., Altomare, D.A., Skele, K.L., Poulikakos, P.I., Kuhajda, F.P., Di Cristofano, A., and Testa, J.R. (2005) Positive feedback regulation between AKT activation and fatty acid synthase expression in ovarian carcinoma cells. Oncogene 24, 3574-3582
44. Longva, K.E., Pedersen, N.M., Haslekas, C., Stang, E., and Madshus, I.H. (2005) Herceptin-induced inhibition of ErbB2 signaling involves reduced phosphorylation of Akt but not endocytic down-regulation of ErbB2. Int J Cancer 116, 359-367
18
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

19
Figure Legends
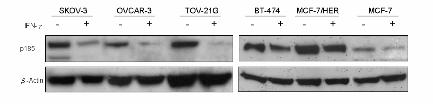
Figure 1 Down-regulation of p185 expression by IFN-γ in ovarian and breast cancer cell lines. (A)
SKOV-3, OVCAR-3, and TOV-21G human ovarian cancer cell lines, and BT-474, MCF-7/HER, and
MCF-7 human breast cancer cell lines were treated with 1000 U/ml of IFN-γ or media alone for 72 h.
Cell lysates were immunoblotted with antibodies specific for neu or β-actin. (B) SKOV-3 cells were
treated with 1000 U/ml of IFN-γ or media for the indicated time. Densitometric analysis of the
percentage decrease was determined using Adobe Photoshop software. (C) SKOV-3 cells were treated
with the indicated doses of IFN-γ for 72 h. Western blot and densitometric analysis were performed as
described above. p185 expression was down-regulated by IFN-γ in SKOV-3 cell line in a dose- and
time-dependent manner. The experiment was repeated three times. The data are presented as the mean
± SE. *, p < 0.05. **, p < 0.01.
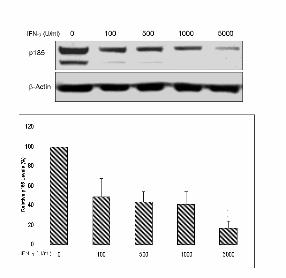
Figure 2 (A) Up-regulation of RIG1 expression by IFN-γ treatment in SKOV-3 ovarian cancer cells.
Cells were harvested after 2 h incubation with the indicated doses of IFN-γ. The PCR products were
stained with ethidium bromide on a 0.8% agarose gel (upper panel). The q-PCR quantification was
performed by adding the CT values of RIG1 cDNA and normalizing them to GAPDH cDNA (Roche)
(lower panel). (B) RIG1 siRNA restored the reduction of p185 caused by IFN-γ. Cells were treated
with 1000 or 100 U/ml of IFN-γ for 24 h and then transfected with negative control siRNA (NCi) or
19
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

20
RIG1 siRNA (RIG1i). Cell lysates were immunoblotted with antibodies specific for neu or β-actin
(left). RT-PCR for RIG1 shows the inhibitory effect of RIG1 siRNA (right).
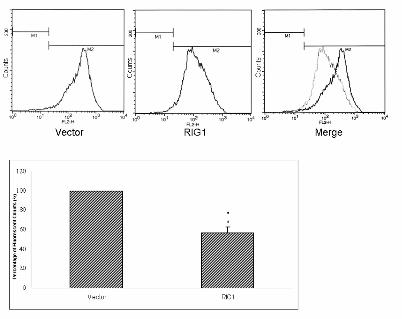
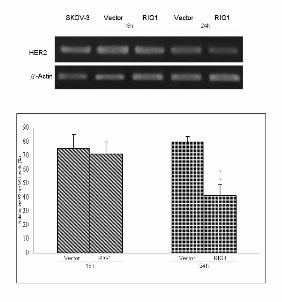
Figure 3 Down-regulation of the HER2 gene and protein by RIG1. (A) The amount of p185 in
RIG1-containing or vector control cells was quantified by flow cytometry using PE-conjugated
anti-HER2 monoclonal antibody. The mean fluorescence intensity was 237.27 for the vector and
116.98 for RIG1. The 52% (exponential scale) reduction of fluorescence by RIG1 was quantified
using the GeoMean fluorescence parameter provided with CELLQUEST software. (B) SKOV-3 cells
were transfected and sorted at the indicated time. mRNA quantification was performed as described
previously. The data are presented as the mean ± SE. NIH3T3 (C) and SKOV-3 (D) cell lines were
cotransfected with 2 μg of pNulit with different amounts of RIG1 plasmid DNA (0 μg, 0.5 μg, 1 μg,
1.5 μg, or 2 μg on the abscissa). The experiments were repeated four times. The relative light units
(RLU) activity of luciferase was normalized relative to β-galactosidase activity (left), and the
inhibition of promoter activities was also determined (right).
Figure 4 Down-regulation of p185 and phospho-PI3K/Akt/mTOR by IFN-γ and RIG1. (A) RIG1 or
vector transfectants or SKOV-3 parental cells were collected 18 and 24 h after transfection.
Successfully transfected cells were sorted by FACS Vantage SE. Western blot analysis of cell lysates
was performed with antibodies recognizing p185, phospho-p185-Y1248 (p-p185), p85 PI3K,
phospho-Y p85 PI3K (p-PI3K), Akt, phospho-Akt-S473 (p-Akt), Ras, ERK,
20
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

21
phospho-ERK-T202/Y204 (p-ERK), mTOR, phospho-mTOR-S2448 (p-mTOR), GFP (RIG1), and
β-actin proteins in the transfected cells. The densitometric analysis was done as previously. (B)
SKOV-3 cells were treated with 1000 U/ml of IFN-γ or media alone. Cells were harvested after 24 h,
and western blot analysis of cellular proteins was performed as described previously. The experiments
were repeated three times. The data are presented as the mean ± SE. Both IFN-γ and RIG1 could
down-regulate p185 and its signaling pathway.
Figure 5 Down-regulation of VEGF by RIG1. (A) Protein lysates were prepared using similar
procedures from the vector control and RIG1 transfectants. Antibodies specific to VEGF (A-20) and
β-actin were applied. (B) Cells were seeded overnight, and then incubated for 24 h in serum-free
medium. VEGF concentration was measured by Quantikine ELISA kits. RIG1 produced the lowest
VEGF levels.
Figure 6 (A) Heregulin restored the RIG1-induced reduction of phospho-p185 and phospho-Akt. Cells
were transfected for the indicated time, 5 nM of HRG was added to the cells for 30 min before
harvest, and the cells were applied to the FACS Vantage SE. HRG specifically restored the activated
levels of p185 and Akt after 24 h of RIG1 transfection (upper left). Densitometry was performed as
described above (lower panel). Treatment with 5 nM HRG for 30 min was enough to increase
phospho-p185 but not p185 (upper right). (B) A schematic model of the effects of IFN-γ and RIG1 on
the regulation of HER2.
21
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from

© The Author 2008 . Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected]
by guest on March 18, 2014
http://carcin.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















