
Induction of Cardiogenesis in Embryonic Stem Cells via Downregulation of Notch1 Signaling
17
Induction of Cardiogenesis in Embryonic Stem Cells via Downregulation of Notch1 Signaling Mohamed Nemir, Adrien Croquelois, Thierry Pedrazzini, Freddy Radtke Abstract—Embryonic stem cells represent an attractive source of cardiomyocytes for cell-replacement therapies. However, before embryonic stem cells can be successfully used for the treatment of cardiac diseases, the precise molecular mechanisms that underlie their cardiogenic differentiation must be identified. A network of intrinsic and extrinsic factors regulates embryonic stem cell self-renewal and differentiation into a variety of different cell lineages. Here, we show that Notch signaling takes place in some but not all embryonic stem cells and that the Notch pathway is shut down during the course of differentiation concomitantly with downregulation of Notch receptor and ligand expression. Moreover, gain- and loss-of-function experiments for Notch signaling components show that this pathway is a crucial regulator of cardiomyocyte differentiation within ES cells. Differentiation of ES cells into cardiomyocytes is favored by inactivation of the Notch1 receptor, whereas endogenous Notch signaling promotes differentiation of ES cells into the neuronal lineage. We conclude that Notch signaling influences the cell fate decision between mesodermal and the neuroecto- dermal cell fates during embryonic stem cell differentiation. These findings should help to optimize the production of specific cell types via modulation of the Notch pathways and, in particular, to improve the production of embryonic stem cell-derived cardiomyocytes. (Circ Res. 2006;98:1471-1478.) Key Words: Notch embryonic stem cells cardiomyogenesis differentiation gene targeting H eart failure has become the leading cause of death in developed countries. Hundreds of thousands of new cases are diagnosed each year, and despite a large battery of pharmacological agents, heart transplant remains the ultimate therapy for patients with end-stage heart failure. However, the request for organs far exceeds the number of potential donors. As an alternative approach, the regeneration of the myocardium via controlled differentiation of cardiomyocyte progenitors is receiving much attention. Embryonic stem (ES) cells demonstrate several characteristics that suggest that they might serve as a source of cells for the therapeutic regener- ation of the heart. First, ES cells can be readily isolated from the inner cell mass of the blastocyst 1,2 and subsequently maintained indefinitely in vitro. 3–5 Second, ES cells are totipotent and can be induced to differentiate into a variety of cell types including cardiac myocytes. 6 Third, the recent generation of human ES cell lines has brought further support to the concept of regenerative medicine based on the con- trolled differentiation of ES cells to replace lost cells in damaged organs. 7 Nevertheless, because of their totipotency, ES cells could paradoxically represent a possible risk of producing teratomas following transfer in vivo. Therefore, knowledge of the precise control of the necessary differenti- ation processes will be required before ES cells can be used in therapy. The most commonly used method to induce differentiation of mouse ES cells is the formation of embryoid bodies (EBs), which contain precursors of all 3 embryonic germ layers. 8,9 Indeed, within EBs, ES cells differentiate into spontaneously beating cardiomyocytes that demonstrate structural and elec- trophysiological characteristics identical to those observed in cardiomyocytes of the developing heart. 10 –12 Induction of cardiogenesis appears to be, in part, under the control of morphogens of the transforming growth factor (TGF)- family, such as TGF- itself, nodal, activin, and bone morphogenic proteins (BMPs) 13 . BMP-dependent pathways, in turn, activate specific transcription factors including Nkx2.5, GATA-4, and mouse embryonic fibroblast (MEF)- 2c, which activate cardiac-specific genes such as the -myosin heavy chain (-MHC) and the myosin light chain (MLC)-2v. 14 In contrast, totipotency is maintained by cultur- ing ES cells on a feeder layer of MEFs or in the presence of leukemia inhibitory factor (LIF). 15 Two transcription factors, Oct3/4 and Nanog, have been classified as ES cell identity genes, as they are critical for the self-renewal of ES cells. 16 –18 The Notch pathway is involved in many differentiation processes and lineage decisions in fetal and postnatal devel- opment, as well as in adult self-renewing organs. 19 Notch receptor genes encode evolutionarily conserved transmem- brane proteins, which are activated by 2 types of cell surface Original received August 30, 2005; resubmission received December 13, 2005; revised resubmission received March 31, 2006; accepted May 2, 2006. From the Ludwig Institute for Cancer Research (M.N., A.C., F.R.), Lausanne Branch, University Lausanne, Epalinges; Department of Medicine (M.N., A.C., T.P.), University of Lausanne Medical School; and Swiss Institute for Experimental Cancer Research (ISREC) (F.R.), Epalinges, Switzerland. Correspondence to Freddy Radtke, Ludwig Institute for Cancer Research, Lausanne Branch, Chemin de Boveresses 155, Epalinges VA 1066, Switzerland. E-mail [email protected] © 2006 American Heart Association, Inc. Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000226497.52052.2a 1471 Molecular Medicine by guest on January 2, 2015 http://circres.ahajournals.org/ Downloaded from by guest on January 2, 2015 http://circres.ahajournals.org/ Downloaded from by guest on January 2, 2015 http://circres.ahajournals.org/ Downloaded from by guest on January 2, 2015 http://circres.ahajournals.org/ Downloaded from by guest on January 2, 2015 http://circres.ahajournals.org/ Downloaded from by guest on January 2, 2015 http://circres.ahajournals.org/ Downloaded from by guest on January 2, 2015 http://circres.ahajournals.org/ Downloaded from by guest on January 2, 2015 http://circres.ahajournals.org/ Downloaded from by guest on January 2, 2015 http://circres.ahajournals.org/ Downloaded from by guest on January 2, 2015 http://circres.ahajournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Induction of Cardiogenesis in Embryonic Stem Cells via Downregulation of Notch1 Signaling

Induction of Cardiogenesis in Embryonic Stem Cells viaDownregulation of Notch1 Signaling
Mohamed Nemir, Adrien Croquelois, Thierry Pedrazzini, Freddy Radtke
Abstract—Embryonic stem cells represent an attractive source of cardiomyocytes for cell-replacement therapies. However,before embryonic stem cells can be successfully used for the treatment of cardiac diseases, the precise molecularmechanisms that underlie their cardiogenic differentiation must be identified. A network of intrinsic and extrinsic factorsregulates embryonic stem cell self-renewal and differentiation into a variety of different cell lineages. Here, we showthat Notch signaling takes place in some but not all embryonic stem cells and that the Notch pathway is shut down duringthe course of differentiation concomitantly with downregulation of Notch receptor and ligand expression. Moreover,gain- and loss-of-function experiments for Notch signaling components show that this pathway is a crucial regulator ofcardiomyocyte differentiation within ES cells. Differentiation of ES cells into cardiomyocytes is favored by inactivationof the Notch1 receptor, whereas endogenous Notch signaling promotes differentiation of ES cells into the neuronallineage. We conclude that Notch signaling influences the cell fate decision between mesodermal and the neuroecto-dermal cell fates during embryonic stem cell differentiation. These findings should help to optimize the production ofspecific cell types via modulation of the Notch pathways and, in particular, to improve the production of embryonic stemcell-derived cardiomyocytes. (Circ Res. 2006;98:1471-1478.)
Key Words: Notch � embryonic stem cells � cardiomyogenesis � differentiation � gene targeting
Heart failure has become the leading cause of death indeveloped countries. Hundreds of thousands of new
cases are diagnosed each year, and despite a large battery ofpharmacological agents, heart transplant remains the ultimatetherapy for patients with end-stage heart failure. However,the request for organs far exceeds the number of potentialdonors. As an alternative approach, the regeneration of themyocardium via controlled differentiation of cardiomyocyteprogenitors is receiving much attention. Embryonic stem (ES)cells demonstrate several characteristics that suggest that theymight serve as a source of cells for the therapeutic regener-ation of the heart. First, ES cells can be readily isolated fromthe inner cell mass of the blastocyst1,2 and subsequentlymaintained indefinitely in vitro.3–5 Second, ES cells aretotipotent and can be induced to differentiate into a variety ofcell types including cardiac myocytes.6 Third, the recentgeneration of human ES cell lines has brought further supportto the concept of regenerative medicine based on the con-trolled differentiation of ES cells to replace lost cells indamaged organs.7 Nevertheless, because of their totipotency,ES cells could paradoxically represent a possible risk ofproducing teratomas following transfer in vivo. Therefore,knowledge of the precise control of the necessary differenti-ation processes will be required before ES cells can be usedin therapy.
The most commonly used method to induce differentiationof mouse ES cells is the formation of embryoid bodies (EBs),which contain precursors of all 3 embryonic germ layers.8,9
Indeed, within EBs, ES cells differentiate into spontaneouslybeating cardiomyocytes that demonstrate structural and elec-trophysiological characteristics identical to those observed incardiomyocytes of the developing heart.10–12 Induction ofcardiogenesis appears to be, in part, under the control ofmorphogens of the transforming growth factor (TGF)-�family, such as TGF-� itself, nodal, activin, and bonemorphogenic proteins (BMPs)13. BMP-dependent pathways,in turn, activate specific transcription factors includingNkx2.5, GATA-4, and mouse embryonic fibroblast (MEF)-2c, which activate cardiac-specific genes such as the�-myosin heavy chain (�-MHC) and the myosin light chain(MLC)-2v.14 In contrast, totipotency is maintained by cultur-ing ES cells on a feeder layer of MEFs or in the presence ofleukemia inhibitory factor (LIF).15 Two transcription factors,Oct3/4 and Nanog, have been classified as ES cell identitygenes, as they are critical for the self-renewal of ES cells.16–18
The Notch pathway is involved in many differentiationprocesses and lineage decisions in fetal and postnatal devel-opment, as well as in adult self-renewing organs.19 Notchreceptor genes encode evolutionarily conserved transmem-brane proteins, which are activated by 2 types of cell surface
Original received August 30, 2005; resubmission received December 13, 2005; revised resubmission received March 31, 2006; accepted May 2, 2006.From the Ludwig Institute for Cancer Research (M.N., A.C., F.R.), Lausanne Branch, University Lausanne, Epalinges; Department of Medicine (M.N.,
A.C., T.P.), University of Lausanne Medical School; and Swiss Institute for Experimental Cancer Research (ISREC) (F.R.), Epalinges, Switzerland.Correspondence to Freddy Radtke, Ludwig Institute for Cancer Research, Lausanne Branch, Chemin de Boveresses 155, Epalinges VA 1066,
Switzerland. E-mail [email protected]© 2006 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000226497.52052.2a
1471
Molecular Medicine
by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from

ligands, named Jagged and Delta. Mouse and humans have 4Notch receptors (Notch1 to -4) and 5 ligands: Jagged1,Jagged2, Delta-like 1 (Dll-1), Dll-3, and Dll-4.19,20 Signalingis initiated via ligand–receptor interactions on neighboringcells. This interaction leads to the proteolytic cleavage of thereceptor, a process that releases the Notch intracellulardomain (NIC), which then translocates to the nucleus andheterodimerizes with the transcription factor RBP-Jk, con-verting it from a transcriptional repressor to an activator.19
Notch signaling plays an important role in the cellulariza-tion21 of the heart during cardiac development, and mice inwhich the canonical Notch pathway is inhibited die duringembryonic life, in part, from heart defects.22 Interestingly, theblockade of the Notch pathways in ES cells was recentlyshown to favor commitment to a mesodermal cell fate.23
Therefore, in an attempt to define the molecular eventscontrolling the production of ES cell–derived cardiomyo-cytes, we investigated the ability of the Notch pathway tocontrol ES cell cardiogenic differentiation. We report thatNotch signaling mediated by the Notch1 receptor is respon-sible for suppressing cardiogenesis in ES cells. Notch doesnot appear to be a general inhibitor of ES cell differentiationbut functions as a regulator of cell fate decisions in multipo-tent ES cells that must choose between mesodermal andneuroectodermal fates.
Materials and MethodsCell CultureIn this study, Notch1�/� ES cell clones and the parental Notch1lox/lox
cell line, the E14/T, and the 293T cell lines were used. The cultureconditions, transfections, and differentiation protocols are describedin the expanded Materials and Methods section in the online datasupplement, available at http://circres.ahajournals.org. Details ofcloning and expression of DNA constructs, immunofluorescence,fluorescence-activated cell sorting (FACS), Western blot, and RT-PCR analyses are also described therein.
ResultsNotch Signaling Is Downregulated During ESCell DifferentiationTo explore the functions of Notch signaling in ES cells,expression of the different Notch receptors and ligands wasdetermined in undifferentiated ES cells as well as in EBs thatexhibited spontaneous contraction, an indication of cardio-myocyte differentiation. Undifferentiated ES cells expressedrelatively high levels of Notch1 and Notch4, whereas Notch2was expressed to a lesser extent (Figure 1). Notch3 did notappear to be expressed (data not shown). Among the Notchligands, Jagged2 was expressed at high levels, whereasJagged1 and Dll-1 were only detected at moderate levels(Figure 1). During ES cell differentiation Notch1, Notch4,and Jagged2 expression appeared to be downregulated similarto that of Oct3/4 and Nanog (Figure 1). In contrast, expres-sion of the other Notch family members such as Notch2,Jagged1, or Dll-1 was not significantly altered. Similarly, nodifferences in the expression of putative target genes of Notchpathways, such as Hes1, Hey-1, and Hey-2, were observed inundifferentiated or differentiated ES cells, indicating thatregulation of these target genes is either independent of Notch
signaling or mediated by Notch2, the only receptor that is notdown regulated during differentiation (Figure 1).
To visualize Notch signaling in ES cells, a reporter geneexpressing the red fluorescent protein DsRed under thecontrol of 12 Notch responsive RBP-J DNA binding elementswas constructed (pCAGIP-12RBP-DsRed). The specificity ofthis reporter was first assessed by cotransfection experimentsin 293T cells using constructs encoding a constitutivelyactive form of the Notch1 receptor (NIC, pCAGIPNIC) and adominant negative mutant of the RBP-J protein lacking DNAbinding activity (RBP-JDN, pCAGIPRBP-JDN; Figure 2A).Figure 2B shows that 293T cells display low levels of Notchsignaling under basal conditions. In contrast, DsRed fluores-cence markedly increased with the constitutive activation ofthe Notch pathway. Finally, reporter expression was totallyshut down after cotransfecting the dominant negative RBP-JDN gene. Taken together, these results demonstrate that theDsRed reporter system is Notch responsive and specific.
Next, we transfected this reporter plasmid into ES cells.Representative pictures of undifferentiated ES cell coloniesgrown in the presence of LIF and of early EBs and terminallydifferentiated EBs are shown in Figure 3A. The punctuateexpression pattern of DsRed in undifferentiated ES cellcolonies and in early EBs indicates that Notch signaling
Figure 1. Notch signaling in mouse embryonic stem cells. TotalRNA was isolated from undifferentiated ES cells (Undiff. ES),from 6-day EBs (6-d EBs) derived from ES cells grown in hang-ing drops for 3 days and in suspension for another 3 days, andfrom beating EBs (Diff. EBs) and was subjected to RT-PCR anal-ysis using primers specific for the indicated genes.
1472 Circulation Research June 23, 2006
by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from

occurred only in small, defined areas. Immunostaining forNotch1-IC also demonstrated a dotted staining pattern inwild-type ES cells, consistent with the results obtained withthe Notch-DsRed reporter system (Figure 3C). Furthermore,in agreement with the expression data depicted in Figure 1,terminally differentiated EBs showed little or no fluores-cence, indicating that Notch signaling is shut down duringdifferentiation. To ascertain that the DsRed� cells wereindeed receiving a Notch signal, fluorescent (DsRed�) andnonfluorescent (DsRed�) cells were FACS sorted and ana-lyzed for the expression of Notch1, which is itself a directtarget gene of Notch signaling.24 Figure 3B shows that theDsRed� cells receiving a Notch signal expressed higher levelsof Notch1 mRNA compared with DsRed� ES cells, confirm-ing that activation of the reporter is a valid readout of Notchsignaling. Interestingly, the expression of Hes1, and Hey-2,but not Hey-1 was moderately increased in DsRed� relative toDsRed� ES cells (Figure 3B), suggesting that these genes areonly partially regulated by Notch in ES cells.
Inhibition of Notch Signaling in ES Cells FavorsCardiomyocyte DifferentiationTo investigate whether Notch signaling can control thedifferentiation of ES cells into particular cell lineages, wemodulated the levels of Notch signaling in differentiating EScells by transfecting either the RBP-JDN inhibitory protein orthe constitutively active NIC domain into ES cells. Quanti-tative analysis showed that 40% of the EBs expressing theRBP-JDN protein showed spontaneous beating activity as earlyas 24 hours after plating on gelatin (day 7) compared withonly 10% in controls (Figure 4 A). After 4 days, the beatingactivity was observed in nearly 100% of the RBP-JDN express-
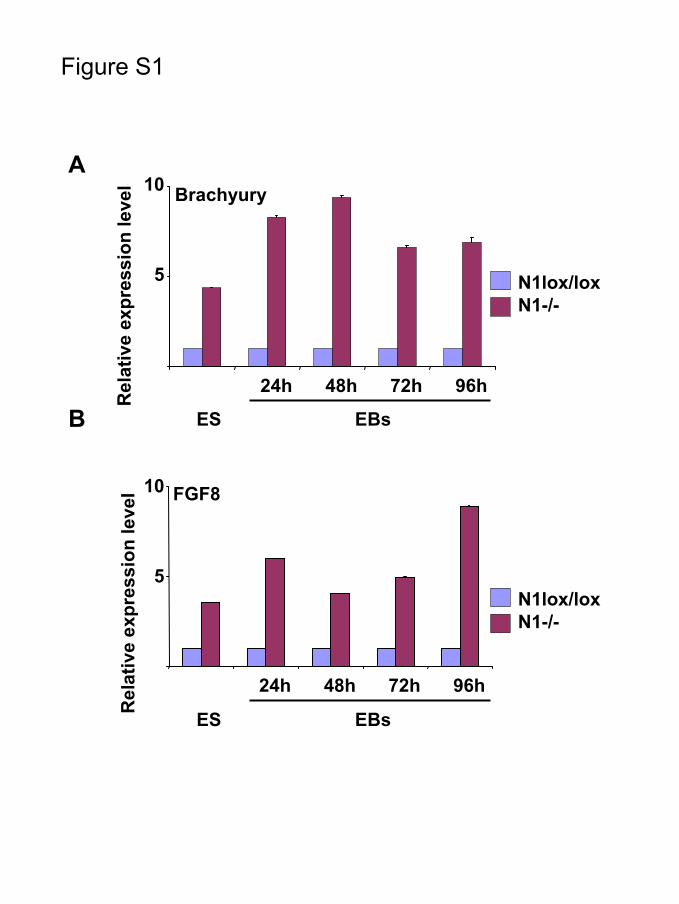
ing EBs and only in 50% to 60% of controls (Figure 4A). Inadditional experiments, we tested whether forced Notchsignaling would inhibit cardiogenesis. As shown in Figure4B, forced expression of the NIC construct significantlyinhibits differentiation of ES cells into cardiomyocytes. Todetermine whether Notch-mediated inhibition preferentiallyaffected the cardiac lineage or blocked commitment toward amesodermal fate, a time-course experiment was performed toquantify the expression of 2 mesodermal markers, Brachyuryand fibroblast growth factor (FGF)-8, in wild-type EBs aswell as in NIC-expressing, RBP-JDN-expressing (Figure 4Cand 4D), and Notch1-deficient EBs (supplemental Figure I).Inhibition of Notch signaling or loss of Notch1 functionresulted in upregulation of Brachyury and FGF-8 expression.In contrast, forced Notch-IC expression produced inhibitionof Brachyury and FGF-8 expression (Figure 4C and 4D).Taken together, these data indicated that Notch signalingoperated in mesodermal progenitors.
Figure 2. Characterization of Notch reporter system. A, Westernblot analysis showing NIC-myc and RBPJDN protein expressionin HEK 293T cells transfected with the indicated combinationsof CAGIP-NIC-myc and CAGIP-RBP-JDN plasmids, in addition tothe Notch reporter p12RBPDsRed and the pEGFPN3 plasmid,used as a control for transfection efficiency. B, Representativemicrographs of HEK 293T cells transfected as in A. The bottompanels with red fluorescence show cells with Notch reporteractivity, whereas the top panels with green fluorescence showequal transfection efficiency. Scale bar�50 �m.
Figure 3. A, The Notch reporter plasmid pCAG-12RBPDsRedwas transfected into E14/T ES cells that were selected withpuromycin and photographed 48 to 96 hours posttransfection.E14/T cells were grown on gelatin or on MEFs, as indicated.Top row, right, shows transfected cells that were photographedat high magnification to indicate alternating DsRed� andDsRed� cells representing Notch signaling and responding cells,respectively. B, RT-PCR analysis of Notch target genes in sig-naling and responding cells. Transfected ES cells were fluores-cence-activated cell sorted based on DsRed fluorescence, andRT-PCR analysis was preformed on DsRed� and DsRed� cellsas indicated. Scale bars�100 �m; scale bar in inset in A,25 �m. C, Immunostaining for activated NIC on wild-type andNotch1�/� ES cells grown on either gelatin or MEFs.
Nemir et al Notch1 Signaling and Cardiogenic Differentiation 1473
by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from
Notch Signaling and CardiomyocyteDifferentiation Are Mutually ExclusiveThe experiments described above suggest that the activationof Notch signaling pathways is incompatible with commit-ment to a cardiac fate. To further address this question, wecotransfected ES cells with the Notch reporter as well as witha cardiac-specific reporter plasmid expressing EGFP geneunder the control of the cardiac-specific �-MHC promoter.Figure 4D shows that in early EBs, Notch signaling (redfluorescence) and cardiac differentiation (green fluorescence)occurred in distinct areas and never colocalized, indicatingthat these 2 events cannot occur in the same cell simulta-
neously. To exclude the possibility that the ES cells had lostthe episomal reporter constructs during differentiation,EGFP� and DsRed� cells were sorted from primary EBs andassessed for their ability to express the reporters in secondaryEBs. Both EGFP� and DsRed� cells were able to formsecondary EBs showing green and red fluorescence, respec-tively, indicating that they had not lost the reporter plasmids(Figure 4F).
Notch1 Mediates Inhibition of CardiogenicDifferentiation in ES CellsWe next investigated which of the Notch receptors mightmediate inhibition of cardiomyocyte differentiation. BecauseNotch1 was downregulated in differentiating EBs, this recep-tor represented an attractive candidate. Therefore, Notch1-deficient ES cells were generated from blastocysts of floxedNotch1 mice that had been transfected with a Cre-combinaseexpression plasmid (Figure 5A and 5B). Loss of Notch1protein and mRNA (Figure 5C and 5F) demonstrated thesuccessful generation of Notch1-deficient ES cell clones. Thegrowth rates (Figure 5D), morphology (Figure 5E), andundifferentiated status of Notch1-deficient ES cells, as indi-cated by the expression levels of Oct3/4 and Nanog (Figure5F), were similar to those of parental control ES cells. The
Figure 4. Notch signaling inhibits cardiac differentiation. A,Time-course analysis of beating activity in empty vector–trans-fected (circles, and red line) or CAGIP-RBP-JDN–transfected (dia-monds, and dark blue line) E14/T cells. ES cell differentiationwas monitored daily, starting 24 hours postplating on gelati-nized plates (day 7). B, Quantitative analysis of cardiomyocytedifferentiation of wild-type (WT) (empty vector–transfected),CAGIP-RBP-JDN–, and CAGIP-NIC–expressing ES cells. Thegraph shows percentage of EBs with beating activity at 6 dayspostplating on gelatin-coated plates. Error bars indicate SD.P�0.005 for WT vs RBPJDN-expressing ES cells; P�0.02 for WTvs NIC expressing ES cells. C and D, Quantitative analysis ofBrachyury (C) and FGF8 (D) expression in WT, NIC-, and RBP-JDN–expressing ES cells at indicated time points during differen-tiation. The expression levels, normalized to GAPDH, are relativeto those of undifferentiated ES cells. E, E14/T ES cells werecotransfected with the Notch (red fluorescence) and cardiacreporter (green fluorescence) plasmids, selected in puromycinand differentiated into EBs. Top row shows merge of fluorescentand phase-contrast micrographs of representative 6-day EBsshowing no overlap between Notch signaling and cardiac differ-entiation. Bottom row shows high-magnification fluorescentmicrographs of EBs after plating on gelatin. F, ES cells weretransfected either with pCAG-12RBP-J-DsRed or with pCAG-MHC-EGFP and were differentiated into 6-day EBs, which weresubsequently trypsinized into single cells. DsRed� and EGFP�
cells were isolated by FACS and used to make secondary EBs,which were photographed at 6 days. Scale bars�100 �m.
Figure 5. Generation of Notch1-deficient ES cells. A, upper line,Schematic representation of the Notch1 allele with exon 1 (E1)flanked by 2 LoxP sites (P). Lower line represents the Cre-recombined allele, without E1. B, PCR analysis of isolatedNotch1lox/lox and Notch�/� ES cell clones. PCR primers are sche-matically shown in A (arrows). C, Western blot analysis ofNotch1 protein expression in Notch1lox/lox and Notch1�/� EScells. The myoblast cell line C2C12 is used as a positive control.D, Analysis of ES cell growth. Two independent ES cell cloneswith either floxed (N1l/L #2 and N1l/L #10) and inactivated (N1�/�
#1 and N1�/� #3) Notch1 alleles were plated in triplicates ongelatin-coated 24-well plates and counted every 24 hours. E,Representative ES cell colonies of Notch1lox/lox and Notch1�/�
ES cells are shown. F, RT-PCR analysis for Notch1, Nanog, andOct3/4 gene expression derived from Notch1lox/lox and Notch1�/�
ES cells. Scale bar�100 �m.
1474 Circulation Research June 23, 2006
by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from

expression of neuronal neurofilament 200 (NF200) andendodermal �-fetoprotein (AFP) markers during ES celldifferentiation was indistinguishable between control andNotch1-deficient ES cells (Figure 6E), indicating thatNotch1�/� ES cells are still pluripotent.
Notch1-deficient and control ES cells were then assessedfor their differentiation potential into cardiomyocytes (Figure
6). On average, 70% of EBs formed by Notch1-deficient EScells showed differentiation into cardiomyocytes with beatingactivity, compared with only 30% of their control counter-parts (Figure 6A). In addition, we also noted control EBsshowed an average of 2 to 3 beating areas per EB, whereasNotch1-deficient EBs had 6 to 12 beating areas per EB(Figure 6B). Intracellular staining and subsequent FACSanalysis of single cell suspensions derived from Notch1-deficient beating EBs showed an average of 66% �-actininpositive cells compared with 38% of control EBs (Figure 6C).Furthermore, to visualize cardiomyocytes within the beatingareas, differentiated EBs were fixed and stained for �-actinin.This staining revealed that beating areas of a Notch1-deficient EB was much larger and contained on average 3times more cardiac myocytes than that of control EBs (Figure6D, right column). Notch1-deficient early EBs also showed amore mature morphology characterized by elongated andspread-out cardiac myocytes. These also had well-organizedmyofibrils with aligned sarcomeres (Figure D6, inset). Incontrast, control EBs contained relatively more round cellswith weak �-actinin staining, indicative of immature cardio-myocytes (Figure 6D, left column). These results are furthersupported by gene expression analysis for cardiac-specificmarkers such as MLC-2v, MLC-2a, and MEF-2c in wild-typeand Notch1-deficient ES cells, 6-day EBs, and 12-day EBs.Absence of Notch1 results in increased and/or earlier onset ofexpression of these cardiac specific markers (Figure 6E). Inaddition, quantitative RT-PCR analysis of early mesodermalexpression markers, such as GATA4, Nkx-2.5, or �- and�-MHC, but not �-skeletal actin (ActaA), shows increasedexpression during ES cell differentiation in absence ofNotch1 (Figure 6F). Moreover, the ratios between �- and�-MHC expression shows increased and earlier onset of�-MHC expression confirming the more mature sarcomericpattern of cardiomyocytes derived from Notch1�/� ES cells.Taken together, these results clearly show that Notch1 sig-naling inhibits cardiomyocyte differentiation.
Notch Signaling in ES Cells FacilitatesDifferentiation Toward theNeuroectodermal LineageBecause Notch signaling has been shown to influence binarycell fate decisions in many developmental systems, it couldconceivably play a similar role during ES cell differentiation.To assess whether Notch-mediated prevention of cardiomyo-cyte differentiation occurs at the expense of another celllineage, ES cells were transfected with either the Notchsignaling or the cardiac differentiation reporter plasmids anddifferentiated into primary EBs. Cells receiving a Notchsignal (red) or cells starting to differentiate toward cardio-myocytes (green) were fluorescence-activated cell sorted andused to form secondary EBs. These secondary EBs were thenassessed for their ability to differentiate into either mesoder-mal (cardiomyocyte) or neuroectodermal (neuronal) precur-sors. Secondary EBs formed by DsRed� cells were negativefor �-actinin (Figure 7C), indicating that they were no longerable to differentiate into cardiomyocytes. In contrast, a largeproportion of these secondary EBs stained positive for nestin(Figure 7E), suggesting that they have adopted a neuronal cell
Figure 6. Increased cardiac differentiation of Notch1�/� ES cells.ES cells were differentiated, and spontaneous beating activitywas quantified and expressed as percentage of beating EBs (A)or as number of beating areas per EB (B). Error bars indicateSD. P�0.005 for A and B. C, Quantitative FACS analysis of car-diomyocyte differentiation using �-actinin, a cardiomyocyte-specific marker. Notch1lox/lox (clone #5) and Notch1�/� (clone #1)ES cells were differentiated, dissociated into single cells, andpermeabilized, and intracellular staining was performed using anAlexa488-conjugated monoclonal anti-sarcomeric �-actinin anti-body. D, Immunofluorescence staining of differentiated EBsderived from Notch1lox/lox (clone #2) or Notch1�/� (clone #1) EScells. Cells were differentiated as in A and stained with anti–�-actinin antibody and with Cy3-conjugated secondary antibody.Staining was performed on early EBs (24 hours after plating ongelatin; top panels) and on late EBs (12 days; bottom panels).Inset at top-right is a high magnification showing the striatedpattern of staining corresponding to the Z-disks of the sarco-meres. E, Semiquantitative RT-PCR analysis of cardiac markers(MLC-2v, MLC-2a, MEF-2c), a neuronal marker (NF200), and anendodermal marker (AFP) and tubulin in ES cells, 6-day EBs and12-day EBs formed by Notch1lox/lox and Notch1�/� ES cells. F,Quantitative RT-PCR analysis of cardiac and mesodermalmarker genes. Expression levels in 6-day and 12-day EBs arerelative to levels detected in ES cells. Values represent an aver-age of duplicate determinations in 2 independentexperiments�SD. **Significant at P�0.05. Scale bars in D,100 �m; scale bar in inset, 5 �m.
Nemir et al Notch1 Signaling and Cardiogenic Differentiation 1475
by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from
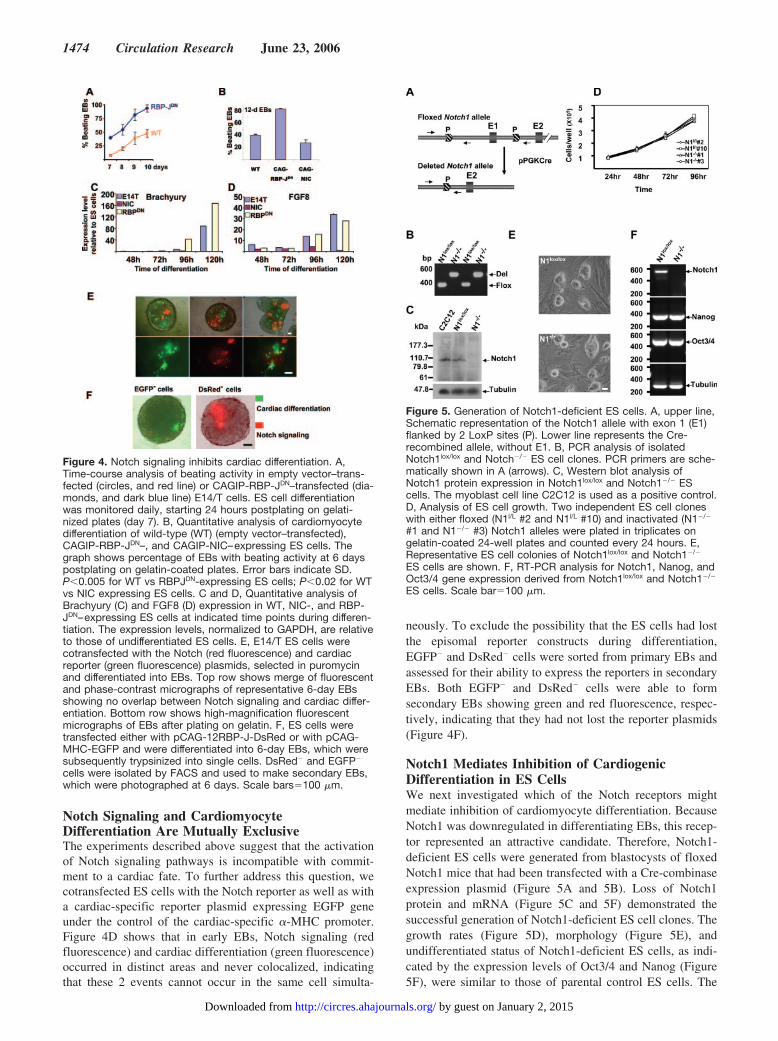
fate. In parallel experiments, sorted EGFP� cells were stillable to form secondary EBs, which stained positively for�-actinin (Figure 7D) and negatively for nestin (Figure 7F).
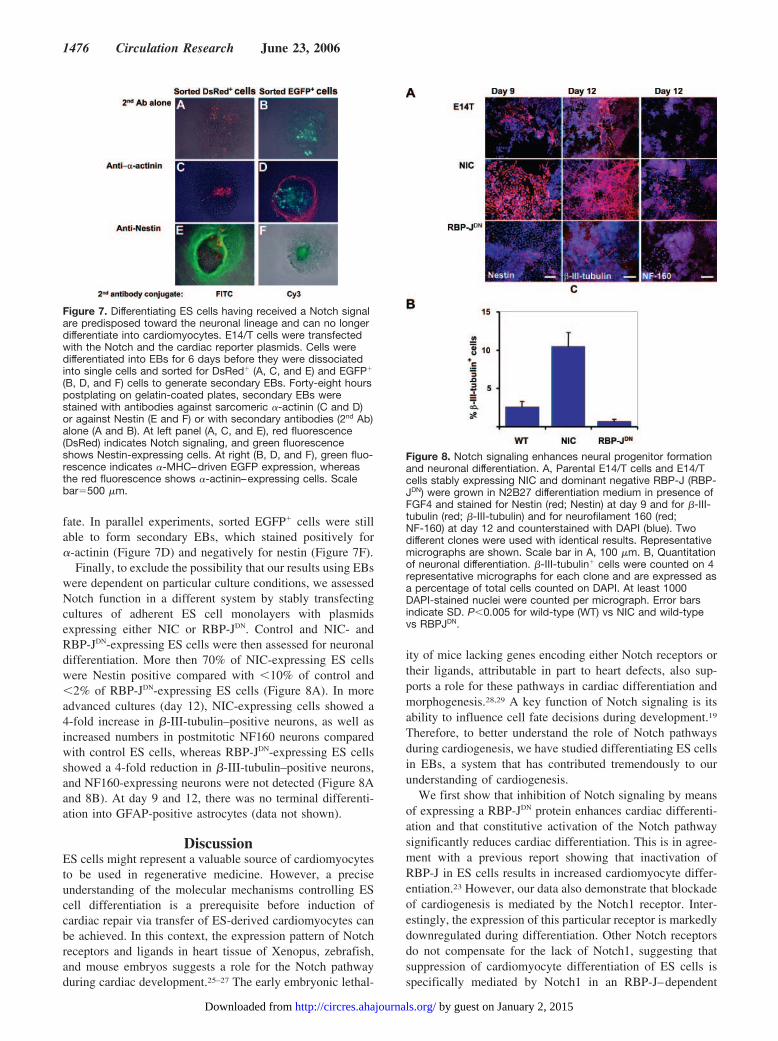
Finally, to exclude the possibility that our results using EBswere dependent on particular culture conditions, we assessedNotch function in a different system by stably transfectingcultures of adherent ES cell monolayers with plasmidsexpressing either NIC or RBP-JDN. Control and NIC- andRBP-JDN-expressing ES cells were then assessed for neuronaldifferentiation. More then 70% of NIC-expressing ES cellswere Nestin positive compared with �10% of control and�2% of RBP-JDN-expressing ES cells (Figure 8A). In moreadvanced cultures (day 12), NIC-expressing cells showed a4-fold increase in �-III-tubulin–positive neurons, as well asincreased numbers in postmitotic NF160 neurons comparedwith control ES cells, whereas RBP-JDN-expressing ES cellsshowed a 4-fold reduction in �-III-tubulin–positive neurons,and NF160-expressing neurons were not detected (Figure 8Aand 8B). At day 9 and 12, there was no terminal differenti-ation into GFAP-positive astrocytes (data not shown).
DiscussionES cells might represent a valuable source of cardiomyocytesto be used in regenerative medicine. However, a preciseunderstanding of the molecular mechanisms controlling EScell differentiation is a prerequisite before induction ofcardiac repair via transfer of ES-derived cardiomyocytes canbe achieved. In this context, the expression pattern of Notchreceptors and ligands in heart tissue of Xenopus, zebrafish,and mouse embryos suggests a role for the Notch pathwayduring cardiac development.25–27 The early embryonic lethal-
ity of mice lacking genes encoding either Notch receptors ortheir ligands, attributable in part to heart defects, also sup-ports a role for these pathways in cardiac differentiation andmorphogenesis.28,29 A key function of Notch signaling is itsability to influence cell fate decisions during development.19
Therefore, to better understand the role of Notch pathwaysduring cardiogenesis, we have studied differentiating ES cellsin EBs, a system that has contributed tremendously to ourunderstanding of cardiogenesis.
We first show that inhibition of Notch signaling by meansof expressing a RBP-JDN protein enhances cardiac differenti-ation and that constitutive activation of the Notch pathwaysignificantly reduces cardiac differentiation. This is in agree-ment with a previous report showing that inactivation ofRBP-J in ES cells results in increased cardiomyocyte differ-entiation.23 However, our data also demonstrate that blockadeof cardiogenesis is mediated by the Notch1 receptor. Inter-estingly, the expression of this particular receptor is markedlydownregulated during differentiation. Other Notch receptorsdo not compensate for the lack of Notch1, suggesting thatsuppression of cardiomyocyte differentiation of ES cells isspecifically mediated by Notch1 in an RBP-J–dependent
Figure 7. Differentiating ES cells having received a Notch signalare predisposed toward the neuronal lineage and can no longerdifferentiate into cardiomyocytes. E14/T cells were transfectedwith the Notch and the cardiac reporter plasmids. Cells weredifferentiated into EBs for 6 days before they were dissociatedinto single cells and sorted for DsRed� (A, C, and E) and EGFP�
(B, D, and F) cells to generate secondary EBs. Forty-eight hourspostplating on gelatin-coated plates, secondary EBs werestained with antibodies against sarcomeric �-actinin (C and D)or against Nestin (E and F) or with secondary antibodies (2nd Ab)alone (A and B). At left panel (A, C, and E), red fluorescence(DsRed) indicates Notch signaling, and green fluorescenceshows Nestin-expressing cells. At right (B, D, and F), green fluo-rescence indicates �-MHC–driven EGFP expression, whereasthe red fluorescence shows �-actinin–expressing cells. Scalebar�500 �m.
Figure 8. Notch signaling enhances neural progenitor formationand neuronal differentiation. A, Parental E14/T cells and E14/Tcells stably expressing NIC and dominant negative RBP-J (RBP-JDN) were grown in N2B27 differentiation medium in presence ofFGF4 and stained for Nestin (red; Nestin) at day 9 and for �-III-tubulin (red; �-III-tubulin) and for neurofilament 160 (red;NF-160) at day 12 and counterstained with DAPI (blue). Twodifferent clones were used with identical results. Representativemicrographs are shown. Scale bar in A, 100 �m. B, Quantitationof neuronal differentiation. �-III-tubulin� cells were counted on 4representative micrographs for each clone and are expressed asa percentage of total cells counted on DAPI. At least 1000DAPI-stained nuclei were counted per micrograph. Error barsindicate SD. P�0.005 for wild-type (WT) vs NIC and wild-typevs RBPJDN.
1476 Circulation Research June 23, 2006
by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from

manner. Transfection of a Notch reporter construct into EScells also shows that Notch signaling is reduced duringdifferentiation, concomitantly with the reduction of the ex-pression of Notch1 itself. In addition, cotransfection of Notchand cardiac differentiation reporter constructs shows thatcells that have received a Notch signal are prevented fromcommitting to a cardiac fate. These data indicate that Notchsignaling inhibits cardiac differentiation of ES cells throughthe action of Notch1. However, we also show that theexpression of Brachyury and FGF-8, 2 markers for mesoder-mal cell lineage, are also repressed in Notch-responsive cellsand, together with other mesodermal markers such asGATA4, Nkx-2.5, or �- and �-MHC, markedly increased incells lacking Notch signaling, suggesting that Notch influ-ences commitment of mesodermal progenitors.
The expression of Notch1, Notch4, and Jagged2 is down-regulated during ES cell differentiation, similar to that of the2 ES cell identity genes Oct3/416 and Nanog17 (Figure 1).However, it is unlikely that Notch pathways play a role in EScell maintenance or self-renewal. This is supported by thepunctuate pattern of Notch signaling that is restricted to someareas of ES cell colonies, making it unlikely that an entirecolony could be maintained in an undifferentiated statethrough only a few Notch signal–receiving cells (Figure 3).Second, expression of Oct3/4 and Nanog are not affected bythe presence or absence of Notch1. Third, the morphologyand growth rate of Notch1-deficient ES cells, as well as theirpluripotency, is identical to wild-type cells (Figures 5 and 6).Similarly, RBP-J–deficient or RBP-JDN ES cell lines havecharacteristics identical to wild-type cells. Finally, embryoslacking components of the Notch cascade develop beyondgastrulation to a late stage (embryonic day 9.5) (reviewedpreviously28,29).
Notch signaling could inhibit cardiogenesis either byblocking mesodermal differentiation per se or by inhibiting amesodermal progenitor from adopting its final cardiomyocytefate. Analysis of Brachyury and FGF-8 expression, 2 markersof mesodermal differentiation, suggests that Notch pathwaysacts rather at an early step in the differentiation process toinhibit commitment toward the mesodermal lineage. Theincrease in MLC-2a, as well as in �- and �-MHC, expressionin Notch1-deficient EBs, together with upregulation of earlycardiac markers such as GATA4 and Nkx2.5, likely reflects ageneral relief of a block in mesodermal and then cardiacdifferentiation, generating in turn cardiac cells not yet spec-ified in ventricular or atrial myocytes.
Moreover, the observation that cardiac differentiation andNotch signaling are mutually exclusive suggests that Notch-expressing cells are not direct precursors of cardiomyocytes.In contrast, the presence of cells expressing nestin but not�-actinin in secondary EBs formed by Notch responsive cells(DsRed�) indicates that cells receiving Notch signals haveadopted a neuronal fate and are no longer able to differentiateinto cardiomyocytes. This interpretation is further supportedby the ability of a dominant active form of Notch to induceand enhance neuronal differentiation, whereas inhibition ofNotch signaling results in suppression of neurogenesis. Theseobservations are compatible with the ability of Notch toinfluence binary cell fate decisions, among the best-
documented roles of Notch signaling in many developmentalsystems (reviewed previously30). Although our data clearlydemonstrate that Notch signaling influences neuronal versuscardiac differentiation, it remains to be shown whether thiscontrol is exerted on 1 common or 2 distinct neural/mesoder-mal progenitors. Although such a common neural/cardiacprogenitor has not yet been described in ES cells, bipotentprogenitors have been shown to exit in the neural crest-derived cardiac side population.31
Several endoderm-derived factors, such as TGF-�,32
BMPs,33 and FGFs,34 that regulate cardiogenesis in theadjacent mesoderm have been used to enhance cardiacdifferentiation of ES cells in vitro.35 Indeed, a recent reportshows that timely inhibition of BMP signaling in ES cells byNoggin greatly enhances cardiogenesis via induction ofBrachyury and Nkx2.5.33 Our study demonstrates that inhi-bition of Notch signaling promotes cardiogenesis and simul-taneously enhances Brachyury expression. Interestingly, inseveral model systems, such as in differentiating myoblasts,Notch and BMP pathways cooperate to block downstreamdifferentiation events.36 Whether or not Notch signaling candirectly modulate Noggin expression and/or release to furtherenhance cardiogenesis in ES cells requires further investiga-tion. Nevertheless, the present data support the hypothesisthat inhibition of Notch1-mediated signaling is essential formesodermal induction of ES cells and is, therefore, a man-datory step for their cardiogenic differentiation.
AcknowledgmentsWe are grateful to Dr Anne Wilson (LICR, Epalinges, Switzerland)for help with FACS analysis and for critical reading ofthe manuscript.
Sources of FundingThis work was supported by the Leenaards Foundation (to F.R. andT.P.) and the Swiss National Science Foundation (grant no. 32-00B0-102154/1 to T.P.).
DisclosuresNone.
References1. Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells
from mouse embryos. Nature. 1981;292:154–156.2. Martin GR. Isolation of a pluripotent cell line from early mouse embryos
cultured in medium conditioned by teratocarcinoma stem cells. Proc NatlAcad Sci U S A. 1981;78:7634–7638.
3. Burdon T, Smith A, Savatier P. Signalling, cell cycle and pluripotency inembryonic stem cells. Trends Cell Biol. 2002;12:432–438.
4. Bradley A, Evans M, Kaufman MH, Robertson E. Formation of germ-linechimaeras from embryo-derived teratocarcinoma cell lines. Nature. 1984;309:255–256.
5. Nagy A, Gocza E, Diaz EM, Prideaux VR, Ivanyi E, Markkula M,Rossant J. Embryonic stem cells alone are able to support fetal devel-opment in the mouse. Development. 1990;110:815–821.
6. Rippon HJ, Bishop AE. Embryonic stem cells. Cell Prolif. 2004;37:23–34.
7. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ,Marshall VS, Jones JM. Embryonic stem cell lines derived from humanblastocysts. Science. 1998;282:1145–1147.
8. Abe K, Niwa H, Iwase K, Takiguchi M, Mori M, Abe SI, Yamamura KI.Endoderm-specific gene expression in embryonic stem cells differen-tiated to embryoid bodies. Exp Cell Res. 1996;229:27–34.
Nemir et al Notch1 Signaling and Cardiogenic Differentiation 1477
by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from

9. Leahy A, Xiong JW, Kuhnert F, Stuhlmann H. Use of developmentalmarker genes to define temporal and spatial patterns of differentiationduring embryoid body formation. J Exp Zool. 1999;284:67–81.
10. Hescheler J, Halbach M, Egert U, Lu ZJ, Bohlen H, Fleischmann BK,Reppel M. Determination of electrical properties of ES cell-derived car-diomyocytes using MEAs. J Electrocardiol. 2004;37(suppl):110–116.
11. Kehat I, Kenyagin-Karsenti D, Snir M, Segev H, Amit M, Gepstein A,Livne E, Binah O, Itskovitz-Eldor J, Gepstein L. Human embryonic stemcells can differentiate into myocytes with structural and functional prop-erties of cardiomyocytes. J Clin Invest. 2001;108:407–414.
12. Wobus AM, Kaomei G, Shan J, Wellner MC, Rohwedel J, Ji G,Fleischmann B, Katus HA, Hescheler J, Franz WM. Retinoic acid accel-erates embryonic stem cell-derived cardiac differentiation and enhancesdevelopment of ventricular cardiomyocytes. J Mol Cell Cardiol. 1997;29:1525–1539.
13. Foley A, Mercola M. Heart induction: embryology to cardiomyocyteregeneration. Trends Cardiovasc Med. 2004;14:121–125.
14. Lev S, Kehat I, Gepstein L. Differentiation pathways in human embryonicstem cell-derived cardiomyocytes. Ann N Y Acad Sci. 2005;1047:50–65.
15. Williams RL, Hilton DJ, Pease S, Willson TA, Stewart CL, Gearing DP,Wagner EF, Metcalf D, Nicola NA, Gough NM. Myeloid leukaemiainhibitory factor maintains the developmental potential of embryonicstem cells. Nature. 1988;336:684–687.
16. Niwa H, Miyazaki J, Smith AG. Quantitative expression of Oct-3/4defines differentiation, dedifferentiation or self-renewal of ES cells. NatGenet. 2000;24:372–376.
17. Chambers I, Colby D, Robertson M, Nichols J, Lee S, Tweedie S, SmithA. Functional expression cloning of Nanog, a pluripotency sustainingfactor in embryonic stem cells. Cell. 2003;113:643–655.
18. Mitsui K, Tokuzawa Y, Itoh H, Segawa K, Murakami M, Takahashi K,Maruyama M, Maeda M, Yamanaka S. The homeoprotein Nanog isrequired for maintenance of pluripotency in mouse epiblast and ES cells.Cell. 2003;113:631–642.
19. Artavanis-Tsakonas S, Rand MD, Lake RJ. Notch signaling: cell fatecontrol and signal integration in development. Science. 1999;284:770–776.
20. Radtke F, Raj K. The role of Notch in tumorigenesis: oncogene or tumoursuppressor? Nat Rev Cancer. 2003;3:756–767.
21. Timmerman LA, Grego-Bessa J, Raya A, Bertran E, Perez-Pomares JM,Diez J, Aranda S, Palomo S, McCormick F, Izpisua-Belmonte JC, de laPompa JL. Notch promotes epithelial-mesenchymal transition duringcardiac development and oncogenic transformation. Genes Dev. 2004;18:99–115.
22. Oka C, Nakano T, Wakeham A, de la Pompa JL, Mori C, Sakai T,Okazaki S, Kawaichi M, Shiota K, Mak TW, Honjo T. Disruption of the
mouse RBP-J kappa gene results in early embryonic death. Development.1995;121:3291–3301.
23. Schroeder T, Fraser ST, Ogawa M, Nishikawa S, Oka C, Bornkamm GW,Honjo T, Just U. Recombination signal sequence-binding protein Jkappaalters mesodermal cell fate decisions by suppressing cardiomyogenesis.Proc Natl Acad Sci U S A. 2003;100:4018–4023.
24. Deftos ML, He YW, Ojala EW, Bevan MJ. Correlating notch signalingwith thymocyte maturation. Immunity. 1998;9:777–786.
25. Rones MS, McLaughlin KA, Raffin M, Mercola M. Serrate and Notchspecify cell fates in the heart field by suppressing cardiomyogenesis.Development. 2000;127:3865–3876.
26. Loomes KM, Taichman DB, Glover CL, Williams PT, Markowitz JE,Piccoli DA, Baldwin HS, Oakey RJ. Characterization of Notch receptorexpression in the developing mammalian heart and liver. Am J MedGenet. 2002;112:181–189.
27. Loomes KM, Underkoffler LA, Morabito J, Gottlieb S, Piccoli DA,Spinner NB, Baldwin HS, Oakey RJ. The expression of Jagged1 in thedeveloping mammalian heart correlates with cardiovascular disease inAlagille syndrome. Hum Mol Genet. 1999;8:2443–2449.
28. Gridley T. Notch signaling in vertebrate development and disease. MolCell Neurosci. 1997;9:103–108.
29. Gridley T. Notch signaling and inherited disease syndromes. Hum MolGenet. 2003;12(spec no. 1):R9–R13.
30. Greenwald I. LIN-12/Notch signaling: lessons from worms and flies.Genes Dev. 1998;12:1751–1762.
31. Tomita Y, Matsumura K, Wakamatsu Y, Matsuzaki Y, Shibuya I,Kawaguchi H, Ieda M, Kanakubo S, Shimazaki T, Ogawa S, Osumi N,Okano H, Fukuda K. Cardiac neural crest cells contribute to the dormantmultipotent stem cell in the mammalian heart. J Cell Biol. 2005;170:1135–1146.
32. Behfar A, Zingman LV, Hodgson DM, Rauzier JM, Kane GC, Terzic A,Puceat M. Stem cell differentiation requires a paracrine pathway in theheart. FASEB J. 2002;16:1558–1566.
33. Yuasa S, Itabashi Y, Koshimizu U, Tanaka T, Sugimura K, Kinoshita M,Hattori F, Fukami S, Shimazaki T, Okano H, Ogawa S, Fukuda K.Transient inhibition of BMP signaling by Noggin induces cardiomyocytedifferentiation of mouse embryonic stem cells. Nat Biotechnol. 2005;23:607–611.
34. Dell’Era P, Ronca R, Coco L, Nicoli S, Metra M, Presta M. Fibroblastgrowth factor receptor-1 is essential for in vitro cardiomyocyte devel-opment. Circ Res. 2003;93:414–420.
35. Heng BC, Haider H, Sim EK, Cao T, Ng SC. Strategies for directing thedifferentiation of stem cells into the cardiomyogenic lineage in vitro.Cardiovasc Res. 2004;62:34–42.
36. Kluppel M, Wrana JL. Turning it up a Notch: cross-talk between TGFbeta and Notch signaling. Bioessays. 2005;27:115–118.
1478 Circulation Research June 23, 2006
by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from

1
Online Data Supplements
Expanded Materials and Methods
Floxed Notch1 (N1lox/lox) ES cells were derived from blastocysts carrying floxed
Notch1 gene 1 and Notch1 -/- (N1-/-) ES cells were obtained by transfecting N1lox/lox
cells with pPGK-Cre plasmid 2. Notch1 deletion was detected by PCR analysis as
described 3. E14/T ES cell line stably expressing the polyoma large T antigen allows
maintenance of episomal vectors carrying a polyoma origin of replication 4 was a
generous gift from Dr. Austin Smith (Stem cell Laboratory, Edinburgh, Scotland).
Cells were cultured on mouse embryonic fibroblast feeders (N1lox/lox and N1-/- cells)
or on gelatinized plates (E14/T cells) in standard ES cell medium supplemented with
1000U/ml of LIF. Cardiac differentiation of ES cells was induced by aggregating
aliquots containing ~500 cells in hanging drops to form embryoid bodies 5 .
Quantitation of cardiac differentiation was done by counting the number of beating
EBs and counting the number of beating areas per EB. Directed differentiation of ES
cells into neurones was carried out in monolayer cultures as described 6 with the
exception of the addition of 1 ng/ml of FGF4 (Sigma). After 3, 6, 9 or 12 days of
growth in differentiation medium, cells were processed for immunofluorescence
analysis.
Plasmids and transfections
Notch and cardiac reporter plasmids: To avoid silencing observed with stably
transfected DNA in ES cells, we used episomal DNA constructs 7 in E147T ES cells
described above. For Notch reporter plasmids, DsRed cDNA was isolated from
pDsRedN1 (Clontech) using XhoI and NotI and cloned downstream of a DNA

2
fragment containing 12 RBP DNA-binding sites. The fragment containing the Notch
responsive promoter plus DsRed was cut with PvuI (blunted) and SfiI and cloned into
pPyCAGIP (a kind gift from Dr. Austin Smith) via SfiI and PstI (blunted) resulting in
pCAGIP-12RBPDsRed. For the Cardiac reporter plasmid, Alpha myosin heavy chain
(α−MHC) promoter 8 was isolated from the Cl22-MHC plasmid (a kind gift from Dr.
Jeffery Robbins; University of Cincinnati, College of Medicine, Cincinnati, Ohio,
USA) using BamHI and XhoI and the 2XHS4 insulator fragment of chicken β-globin
gene 9 was cut from pJC13-1 plasmid (a kind gift from Dr. Adam West; NIDDK,
NIH, Bethesda, Maryland, USA) using NdeI and BamHI. These two fragments have
been subsequently cloned into pEGFPN1 digested with AseI- SalI using a 2-insert
ligation. The fragment containing the HS4 insulator, the α-MHC promoter and
EGFPN1 cDNA were digested with SfiI and PvuI (blunted) and cloned into
pPyCAGIP vector digested with PstI (blunted) and SfiI to yield pCAGIP-Tri.
NotchIC and dominant negative RBPJ expression plasmids: NotchIC-myc Tag
10 was cut from pMI-Notch IC (a kind gift from Professor Mike Bevan; University of
Washington, Seattle, USA) using SmaI and a partial digest with ApaI and
subsequently cloned into pBSII (Stratagene). pBSII-NIC was then cut with SmaI and
KpnI in order to clone NIC into pPGKEGFP. NIC was isolated via SmaI and HpaI
and ligated into pPyCAGIP via blunted XhoI, giving rise to the plasmid pCAG-NIC.
The cDNA encoding a dominant negative RBP-J protein (RBPJDN) 11 was isolated
from pCMX-RBPJ/R218H (a kind gift from Professor Tasuko Honjo, University of
Kyoto, Japan) using EcoRI and BamHI, blunted with Klenow DNA polymerase and
ligated into the blunted XhoI site of the pPyCAGIP giving rise to pCAG-RBPDN.
E14/T cells were transfected with plasmid DNA using Lipofectamine2000
reagent (Invitrogen) and were selected for 2 days using 2 µg/ml of puromycin

3
(Sigma). Cells were continuously maintained in medium containing 0.5 µg/ml of
puromycin. For stable expression of NotchIC and RBPJDN proteins, plasmids were
linearized with PvuI and transfected into ES cells by electroporation using BioRad
Gene Pulser at 960 µF and 250mV settings. Transfected cells were selected using 2
µg/ml of puromycin and resistant clones were picked and screened for protein
expression. Stable transfectants were also maintained under 0.5 µg/ml puromycin
selection throughout the experiments.
Immunofluorescence analysis
Differentiated EBs and monolayer cultures were fixed for 10 minutes in 4%
paraformaldehyde in PBS and permeabilised with 0.2% Triton X100 in PBS. After
blocking in blocking buffer (PBS containing 0.001% Triton X100, 1% BSA and 1%
FCS) they were incubated overnight at 4 °C with the following antibodies: anti α-
actinin (1:500, Sigma A7811), anti β-III-tubulin (1:200, Sigma, T8660); anti
Neurofilament 160 (NF160; 1:100, Sigma N5264) anti-nestin (1:200, Chemicon MAB
363) and anti-activated Notch1 IC (Abcam ab8925). For quantification of neuronal
differentiation, β-III-tubulin-positive cells were counted on 4 representative
micrographs containing each 1000-3000 cells using the NIH Image software.
FACS analysis
Differentiated EBs were trypsinized into single cells, fixed in 2% PFA, permabilized
with 0.5% Saponin and intracellularly stained with Alexa488-conjugated anti
sarcomeric α-actinin antibody (made in house and diluted 1:30 in PBS/0.5%
Saponin/3%FCS) for 45 min. Cells were analyzed using a BD FACS Calibur.
Analysis was carried out on an FSC/SSC gate to select for cells resembling newborn
cardiomyocytes.

4
Western blot analysis
Total cellular proteins (80 µg) were detected by Western blotting using following
antibodies: anti-Notch1, 1:200 (C20; Santa Cruz), anti-myc Tag 1:200 (9E10; Santa
Cruz) and anti-RBPJ-k protein 1:250 (T6709; Institute of Immunology Co. Tokyo,
Japan). Antibody binding was detected using HRP-conjugated antibody and the ECL
reagent (Amersham).
RT-PCR and quantitative RT-PCR analysis
RNA was isolated using Trizol reagent (Invitrogen). Aliquots containing 250-500 ng
of total cellular RNA were analyzed using the GeneAmp kit (Applied Biosystems).
The sequences of specific primers used for Notch1, Notch2, Notch3, Notch4,
Jagged1, Jagged2, Dll4, Nanog, Oct3/4, Brachyury, MLC2v, MLC2a, MEF2c,
NF200, AFP, and tubulin are listed in the supplementary Table 1 below. Each RT-
PCR was performed with optimized conditions allowing amplification within the
exponential phase of the PCR reaction. For quantitative Real-Time RT-PCR, total
cellular RNA was reverse transcribed using Omnisctipt RT Kit (Qiagen) and oligo dT
primers (Promega). Quantitative PCR amplification of the resulting cDNAs was
performed using primers, probes and the Universal Master Mix of TaqMan Gene
Expression Assays on an ABI PRISM 7700 Sequence Detection System following the
manufacturer’s instructions (PE, Applied Biosystems). The data represent an average
of at least two independent experiments done in duplicates. The values of the relative
expression levels of individual genes are normalized to those of GAPDH.
Statistical analysis

5
For statistical significance of beating EB counts, comparison between different
experimental groups was performed by Student’s T-test using two-tailed distribution
and unequal variances. For Real time PCR data, Statistical comparisons among
values of all groups was performed using Two-way ANOVA with p < 0.05.
Supplementary Table S1
List of RT-PCR primers
Notch1 LP: 5’-TGT GAC AGC CAG TGC AAC TC-3’
RP: 5’-GCA GTG CTT CCA GAG TGC CA-3’
Notch2 LP: 5’-ACA TCA TCA CAG ACT TGG TC-3’
RP: 5’-GGC AGC TGC TGT CAA TAA TG-3’
Notch3 LP: 5’-GAT GTC AAT GCA GTG GAT GAG-3’
RP: 5’-CTT TGA GGC CAG GAA GGA AG-3’
Notch4 LP: 5’-CAC CTA GCT GCC AGA TTC TCT C-3’
RP: 5’-ACT GTC CTG GGC ATC TTT ATC C-3’
Jagged1 LP: 5’-ATT CGA TCT ACA TAG CCT GTG AG-3’
RP: 5’-ACC GGA TGG AAT ACA TCG TAT A-3’
Jagged2 LP: 5’-TGT CAG CCA CGG AGC AGT CAT T-3’
RP: 5’-AAG CGC AGG AAA GAA CGT GAG A-3’
Dll4 LP: 5’-GGG TTA ATA CCT GCA CCT GTC TC-3’
RP: 5’-CTG ATG TGC AGT TCA CAG TGG-3’
Nanog LP: 5’-AGG GTC TGC TAC TGA GAT GCT CTG-3’
RP: 5’-CAA CCA CTG GTT TTT CTG CCA CCG-3’
Oct3/4 LP: 5’-CTG AGG GCC AGG CAG GAG CAC GAG-3’
RP: 5’-CTG TAG GGA GGG CTT CGG GCA CTT-3’

6
Brachyury LP: 5’-AAG GAA CAA CCG GTC ATC-3’
RP:5’-GTG TGC GTC AGT GGT GTG TAA TG-3’
MLC2v LP: 5′-GCC AAG AAG CGG ATA GAA GG-3′
RP: 5′-CTT GTG GAA ACG TTT CTC TTG CGG-3′
MLC2a LP: 5′-CGT GGC TCT TCT AAT GTC TTC TC-3′
RP: 5′-AAA CAG TTG CTC TAC CTC AGC AG-3′
MEF2c LP: 5’-GGT TTC CGT AGC AAC TCC TAC TT-3′
RP:5′-CTT GAC TGA AGG ACT TTC CCT TT-3′
NF-200 LP: 5’- GAGTGGTTCCGAGTGAGGTTGGAC-3’
RP: 5’- GAAGCTCCATATCCCTGGGTGGAAAG-3’
AFP LP: 5’- CCTTGGCTGCTCAGTACGACAAGG-3’
RP : 5’- CCTGCAGACACTCCAGCGAGTTTC-3’
Tubulin LP: 5’-TCA CTG TGC CTG AAC TTA CC-3’
RP: 5’-GGA ACA TAG CCG TAA ACT GC-3’
Supplementary legend to Figure S1
The expression of the mesodermal marker genes Brachyury and FGF8 is elevated in
Notch1-/- ES cells compared to the parental Notch1lox/lox ES cells during
differentiation. Time course analysis of Brachyury (A) and FGF8 (B) gene expression
was performed using real-time quantitative PCR at the indicated time points. The data
represents expression values in Notch1-/- relative to Notch1lox/lox ES cells (set at 1),
after normalizing to GAPDH.

7
Supplementary References
1. Radtke F, Wilson A, Stark G, Bauer M, van Meerwijk J, MacDonald HR, Aguet M. Deficient T cell fate specification in mice with an induced inactivation of Notch1. Immunity. 1999;10:547-58.
2. Gu H, Marth JD, Orban PC, Mossmann H, Rajewsky K. Deletion of a DNA polymerase beta gene segment in T cells using cell type-specific gene targeting. Science. 1994;265:103-6.
3. Wolfer A, Wilson A, Nemir M, MacDonald HR, Radtke F. Inactivation of Notch1 impairs VDJbeta rearrangement and allows pre-TCR-independent survival of early alpha beta Lineage Thymocytes. Immunity. 2002;16:869-79.
4. Chambers I, Colby D, Robertson M, Nichols J, Lee S, Tweedie S, Smith A. Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell. 2003;113:643-55.
5. Wobus AM, Kaomei G, Shan J, Wellner MC, Rohwedel J, Ji G, Fleischmann B, Katus HA, Hescheler J, Franz WM. Retinoic acid accelerates embryonic stem cell-derived cardiac differentiation and enhances development of ventricular cardiomyocytes. J Mol Cell Cardiol. 1997;29:1525-39.
6. Ying QL, Smith AG. Defined conditions for neural commitment and differentiation. Methods Enzymol. 2003;365:327-41.
7. Gassmann M, Donoho G, Berg P. Maintenance of an extrachromosomal plasmid vector in mouse embryonic stem cells. Proc Natl Acad Sci U S A. 1995;92:1292-6.
8. Gulick J, Subramaniam A, Neumann J, Robbins J. Isolation and characterization of the mouse cardiac myosin heavy chain genes. J Biol Chem. 1991;266:9180-5.
9. Chung JH, Whiteley M, Felsenfeld G. A 5' element of the chicken beta-globin domain serves as an insulator in human erythroid cells and protects against position effect in Drosophila. Cell. 1993;74:505-14.
10. Deftos ML, He YW, Ojala EW, Bevan MJ. Correlating notch signaling with thymocyte maturation. Immunity. 1998;9:777-86.
11. Chung CN, Hamaguchi Y, Honjo T, Kawaichi M. Site-directed mutagenesis study on DNA binding regions of the mouse homologue of Suppressor of Hairless, RBP-J kappa. Nucleic Acids Res. 1994;22:2938-44.
FGF8
5
10
24h 48h 72h 96h
Brachyury
5
10
24h 48h 72h 96h
N1lox/loxN1-/-
N1lox/loxN1-/-
A
B
Figure S1
Rel
ativ
e ex
pres
sion
leve
lR
elat
ive
expr
essi
on le
vel
ES EBs
ES EBs

Mohamed Nemir, Adrien Croquelois, Thierry Pedrazzini and Freddy RadtkeSignaling
Induction of Cardiogenesis in Embryonic Stem Cells via Downregulation of Notch1
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2006 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/01.RES.0000226497.52052.2a2006;98:1471-1478; originally published online May 11, 2006;Circ Res.
http://circres.ahajournals.org/content/98/12/1471World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2006/05/11/01.RES.0000226497.52052.2a.DC1.htmlData Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on January 2, 2015http://circres.ahajournals.org/Downloaded from




















