
Insulin resistance in penile arteries from a rat model of metabolic syndrome
Upload
independentCategory
view
5download
0
746 Exp Physiol 95.6 pp 746–755
Experimental Physiology – Research Paper
Exercise training attenuates ageing-induced BKCa channeldownregulation in rat coronary arteries
Sulayma Albarwani, Sultan Al-Siyabi, Hajar Baomar and Mohammed O. Hassan
Department of Physiology, College of Medicine and Health Sciences, Sultan Qaboos University, PO Box 35, PC 123, Muscat, Sultanate of Oman
Physical inactivity and ageing are widely recognized as risk factors for development of coronaryartery disease. One of the characteristic changes that occurs in aged coronary artery isdownregulation of their large-conductance voltage- and calcium-activated K+ (BKCa) channels.In this study, we investigated the effects of moderate exercise training (ET) on the activity ofBKCa channels in coronary arteries of aged rats. Old Fischer 344 rats (23–26 months old) wererandomly assigned to sedentary (O-SED, n = 24) or exercise-trained groups (O-ET, n = 28). TheO-ET rats underwent a progressive treadmill exercise-training programme for 60 min day−1,5 days week−1 for 12 weeks. Young animals were used for comparison. Coronary arteries weremounted on a wire myograph, and contractions in response to 1, 10, 30, 50 and 100 nmol l−1
iberiotoxin were compared. Iberiotoxin (100 nmol l−1) contracted coronary arteries of young,O-SED and O-ET rats by 115 ± 14, 36 ± 5.6 and 61 ± 5% of 5-hydroxytryptamine-inducedcontractions, respectively. Patch-clamp studies revealed a larger magnitude of BKCa currentin young (104 ± 15.6 pA pF−1) compared with O-ET (44 ± 9 pA pF−1) and least in O-SEDcoronary smooth muscle cells (8.6 ± 2 pA pF−1). Western immunoblotting was performed tostudy expression levels of BKCa channel proteins. The α and β1 subunits of the BKCa channelwere reduced by 40 ± 3.5 and 30 ± 2.6%, respectively, in coronary arteries of old comparedwith young rats, and ET attenuated this reduction in expression level to 28 ± 2 and 12 ± 4%,respectively. Our results showed that ageing was associated with a reduction in BKCa channels,and ET partly reversed this reduction. We conclude that low-intensity ET may be beneficial inrestoring age-related decline in coronary vasodilatory properties mediated by BKCa channels.
(Received 2 November 2009; accepted after revision 5 February 2010; first published online 5 February 2010)Corresponding author S. Albarwani: Department of Physiology, College of Medicine and Health Sciences, SultanQaboos University, PO Box 35, PC 123, Muscat, Sultanate of Oman. Email: [email protected]
Physical inactivity is well recognized as an independentrisk factor for the development of coronary artery disease(Fletcher et al. 1996). Substantial evidence exists tosupport the beneficial effect of regular exercise in reducingthe incidence and severity of coronary vascular diseasein young and old humans and animals (Chandrashekhar& Anand, 1991; Stewart, 2005). The effect of exerciseon coronary vascular tone of humans and animalsdemonstrated enhancement of endothelium-dependentand -independent vasorelaxation (DeSouza et al. 2000;Laughlin et al. 2001), altered myogenic tone (Laughlin &Muller, 1998), augmented voltage-gated calcium channel(VGCC) currents (Bowles et al. 1998a) and enhancedcontribution of K+ channels to vascular tone (Bowles et al.1998b). However, since most of these studies were carriedout in young age groups, more information is needed on
the effects of exercise training on coronary vascular controlin old age.
As age advances, coronary arteries become stiffer andexhibit diminished vasodilatory responses (Ishida et al.2003) and exaggerated vasoconstrictor responses (Korzicket al. 2005). Most of the cardiovascular complicationsassociated with ageing are due to alterations in coronaryvascular function (Yildiz, 2007). One of the key elementsthat regulates vascular smooth muscle cell excitability isthe activity of its K+ channels. The large-conductancevoltage- and calcium-activated K+ (BKCa) channels are themost abundant of the vascular K+ channels (Nelson et al.1990). In humans and rats, the expression level of BKCa
channels has been shown to be significantly lower in agedcoronary arteries (Marijic et al. 2001; Nishamura et al.2004).
DOI: 10.1113/expphysiol.2009.051250 C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

Exp Physiol 95.6 pp 746–755 Ageing, exercise training and BKCa channels 747
The BKCa channels are composed of pore-formingα and cytoplasmic regulatory β subunits (Tanaka et al.1997). The β1 subunits are expressed predominantly invascular smooth muscle cells (Brenner et al. 2000) andhave been shown to affect gating kinetics (Wallner et al.1999), voltage and calcium sensitivity (Cox & Aldrich,2000), pharmacological properties (Dick et al. 2001)and membrane trafficking of α subunits (Kim et al.2007). In vascular smooth muscle cells, because of theirabundance and high conductance, small changes in theiropen probabilities cause significant hyperpolarization,which results in decreased Ca2+ influx throughL-type VGCC, thereby causing vasorelaxation (Brayden &Nelson, 1992; Nelson & Quayle, 1995). Inhibition of BKCa
channels by the specific blocker iberiotoxin (IbTx; Garciaet al. 1999) causes constriction of isolated animal andhuman coronary arteries (Nishimaru et al. 2004). Theseobservations indicate that in the coronary circulation theBKCa channels are tonically active at resting membranepotential and therefore contribute to the regulation ofvascular tone.
Various studies have demonstrated that BKCa channelsremodel in different physiological and pathologicalconditions. For instance, Nishimaru et al. (2004) reporteda decrease in the β1 subunit of the BKCa channel withcoronary artery ageing in humans and rats. Severalpathological conditions, such as hypertension (Amberg &Santana, 2003), diabetes mellitus (Burnham et al. 2006),metabolic syndrome (Borbouse et al. 2009) and hypoxia(Navarro-Antolın et al. 2005), have also been shown toalter expression levels of BKCa channels in different arterialregions.
Based on the important contribution of BKCa channelsto the regulation of vascular tone in health and disease,and their downregulation with ageing, this study wasdesigned to examine the hypothesis that moderate exercisetraining may attenuate age-related downregulation ofBKCa channels.
Methods
The study and surgical procedures were approved by theMedical Research and Ethics Committee of Sultan QaboosUniversity, Sultanate of Oman.
Experimental animals and exercise training protocol
Twenty-six young (12–16 weeksold) and 52 old (23–26 months old) Fischer 344 male rats (Harlan Inc.,Indianapolis, IN, USA) were housed in a temperature-controlled room (22 ± 2◦C) with a 12 h–12 h light–darkcycle, and received food and water ad libitum. For the oldrats, the mortality rate at this age is reported to be about45–50% (Tanaka et al. 2000).
Old rats were randomly assigned to a sedentary (O-SED, n = 24) or an exercise-training group (O-ET, n = 28).The exercise-training (ET) sessions consisted of walkingon a treadmill at speed of 15 m min−1, five times aweek for 12 weeks. Exercise time was increased graduallyto 10 min day−1, 20 min day−1, 20 min day−1 with 5%gradient and 40 min day−1 by the end of week 1, 2, 3and 4, respectively. The final level of 60 min day−1 with5% gradient was maintained for the remainder of theprogramme (weeks 5–12; Ziada et al. 2005). To reducestress, rats were allowed a 5 min rest every 20 min (Blanco-Centurion & Shiromani, 2006). Heart rate (HR) andsystolic blood pressure were measured at the beginningand at the end of the ET period using a tail cuff (ColumbusInstruments, Columbus, OH, USA). The effectiveness ofET was assessed by reductions of HR of more than 5% ofthe basal rate for each rat (Wei et al. 1987). Rats were killedwithin 24–48 h after the last exercise session.
Isolation of coronary arteries and isometric tensionrecording
Rats were killed with an overdose of a mixture of theanaesthetics ketamine (140 mg kg−1 I.M.) and xylazine(40 mg kg−1 I.M.). Hearts were then excised and placedin cold physiological saline solution (PSS; 0–4◦C). Third-order branches of left coronary arteries were isolated,cleaned free of fat and connective tissue, cut into shortsegments (100–180 μm in diameter, 2.0–2.3 cm in length),and mounted on an isometric wire myograph (DanishMyo Technology, Aarhus, Denmark). Arteries were placedinto the myograph bath, where two parallel tungstenwires (40 μm diameter) were gently passed longitudinallythrough each artery. One wire was attached to the forcetransducer and the other to the micrometer for precisemeasurements of changes in vessel circumference. Arterieswere then superfused with warm (37◦C) PSS of thefollowing composition (mmol l−1): 119 NaCl, 4.7 KCl,1.18 KH2PO4, 1.17 MgSO4, 25 NaHCO3, 5.5 glucose and1.6 CaCl2, pH 7.4 adjusted with NaOH. Arteries werestretched progressively to an internal circumferenceequivalent to 90% of the circumference that the vesselwould have reached if exposed to 100 mmHg transmuralpressure (Halpern et al. 1978). After the normalizationprocedure, arteries were left to equilibrate for 1 h at 37◦Cbefore subsequent evaluation.
Arteries were then contracted with 5 μmol l−1 5-hydroxytryptamine (5-HT), washed and subsequentlyexposed to IbTx in a dose–response manner (1, 10, 30,50 and 100 nmol l−1). Differences in tensions between 5-HT-induced contractions and basal tones were consideredas maximal tension (100%) and were used to normalizeIbTx-induced tensions. At the end of each experiment,arteries were relaxed with Ca2+-free PSS to assess calcium-dependent tone. The Ca2+-free PSS had CaCl2 omitted and
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

748 S. Albarwani and others Exp Physiol 95.6 pp 746–755
1 mmol l−1 EGTA added and the pH corrected accordinglywith NaOH. Throughout the experiments, arteries werecontinuously bubbled with 95% O2 and 5% CO2.
Cell isolation and electrophysiological recording
Arteries were placed into 2 ml of dissociation medium(DM) composed of (in mmol l−1): 145 NaCl, 1 MgCl2,10 Hepes, 10 glucose and 0.1 CaCl2. The cell dissociationprocedure was modified from that reported previously(Albarwani et al. 2003). Briefly, coronary arteries wereplaced in 2 ml DM and the following enzymes (in mgml−1) were added; 1.5 papain, 1 bovine serum albumin(BSA) and 1 dithioerythritol, and coronary arteries wereincubated for 1 h at 4◦C and then for 20 min at 37◦C.They were then transferred to a fresh 2 ml DM containinga second mixture of enzymes (in mg ml−1); 2 collagenase(type XI), 1 trypsin inhibitor and 1 BSA for 10 min at 37◦C.Digested tissue was triturated in an enzyme-free solution,kept in ice and used within 6 h.
Outward currents were elicited by progressive 10 mVvoltage steps (500 ms duration, 5 s intervals) from aholding potential of −70 mV to +90 mV using 3–5 M�
patch electrodes. The pipette solution contained (inmmol l−1): 145 glutamate, 1 MgCl2, 10 Hepes, 1 EGTA,1 Na2ATP and 0.78 CaCl2 (free [Ca2+] ≈ 0.5 μmol l−1).The pH was buffered to 7.1 with KOH (final K+
concentration of ∼145 mmol l−1). The bath solutioncomprised (in mmol l−1): 140 NaCl, 4 KCl, 0.1 CaCl2,1 MgCl2, 10 Hepes and 10 glucose. The solution wasbuffered using NaOH to a final pH of 7.4.
To assess the BKCa current amplitudes, K+ currents werefirst elicited in a drug-free bath solution and then in asolution containing 100 nmol l−1 IbTx. In each cell, peakcurrent amplitudes in each experimental condition wereaveraged from three trials. Peak current was expressed ascurrent density (in pA pF−1) to normalize for differencesbetween cell membrane areas. Membrane capacitance(in pF) was obtained by integrating the average chargeelicited by hyperpolarizing pulses and dividing this valueby the input voltage. Data acquisition and analysis wasperformed using pClamp 9 (Axon instruments, FosterCity, CA, USA).
Western immunoblotting
Western immunoblotting was performed as described byAlbarwani et al. (2003). Membranes were blocked in 10%skimmed milk for 1 h at room temperature and thenincubated overnight at 8◦C with monoclonal anti-BKCa-α (dilution 1:200; BD Bioscience, USA) or polyclonalβ1 (dilution 1:200; Affinity Bioreagents, USA). Afterwashing, membranes were incubated for 2 h at 25◦Cwith the horseradish peroxidase-conjugated secondaryantibodies (dilution 1:5000; Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA). Immunoreactive bandscorresponding to the molecular mass were detectedby chemiluminescence (Advance ECL, Amersham, UK).Membranes were stripped (RestoreTM Western BlotStripping Buffer; Pierce, Rockford, IL, USA) and probedwith β-actin antibody (dilution 1:5000). Channel proteinswere quantified using densitometry analysis, normalizedfor loading differences to β-actin signal and expressedrelative to young coronary density.
Chemicals
Chemicals used in this study were obtained from SigmaChemicals (Steinheim, Germany) unless otherwise stated.Iberiotoxin was prepared in 100 μM stock solutions in PSS,and stored in aliquots at −20◦C.
Statistical analysis
Values are expressed as means ± S.E.M. Statisticalcomparisons between groups were made with ANOVAwith subsequent Student–Newman–Keuls post hocanalysis test. Significance was accepted at P < 0.05.
Results
Efficacy of exercise training
At end of week 12 of ET, the HR of O-SED rats remainedunchanged at 399 ± 8 beats min−1, while that of the O-ETgroup showed a small but significant reduction of 6.6%(from 395 ± 9 to 369 ± 1.3 beats min−1). The O-ET ratsalso showed a reduction in body weight from 432 ± 9.0to 362 ± 2.3 g, while the O-SED group showed a slightinsignificant gain by the end of the ET period.
Exercise training enhanced BKCa channel contributionto basal tone
The basal tones of arteries from the three groups of ratswere compared after 1 h of equilibration at 37◦C. Figure 1Ashows that arteries of young rats exhibited significantlyhigher resting tone (3 ± 0.2 mN mm−1, n = 17) thanarteries of O-SED (2 ± 0.4 mN mm−1, n = 17) or O-ETrats (2.6 ± 0.2 mN mm−1, n = 13). Exposing the arteriesto calcium-free PSS resulted in significant reduction inresting tone in arteries of young (53 ± 6%) compared witharteries of O-SED (37 ± 4.9%) and O-ET rats (41 ± 3.9%;Fig. 1B). Exercise training did not significantly alter restingtension or calcium-dependent tone.
The contribution of BKCa channels to regulation ofcoronary arteriolar tone was examined by calculatingchanges in tension generated by blocking BKCa channelswith the specific blocker, IbTx. Increases in tensionselicited by 5-HT (5 μmol l−1) were not significantlydifferent amongst the three groups of arteries (young
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

Exp Physiol 95.6 pp 746–755 Ageing, exercise training and BKCa channels 749
Figure 1. Basal tone,calcium-dependent tone and effect of5-HT on coronary arteries of young, oldsedentary (O-SED) and oldexercise-trained (O-ET) ratsAverage basal tone (A) andcalcium-dependent tone (B) of O-SEDcoronary arteries were significantly lowerthan those of young rats. Exercise trainingof old rats did not significantly alter basaltone or calcium-dependent tone. C, 5-HT(5 μmol l−1) produced comparable maximalcontractions in coronary arteries of young,SED and O-ET groups. Each value representsthe mean + S.E.M. ∗ Significantly differencefrom young (P < 0.05).
3.02 ± 0.48 mN mm−1, O-SED 2.8 ± 0.47 mN mm−1 andO-ET 3 ± 0.3 mN mm−1; Fig. 1C).
Iberiotoxin caused dose-dependent contractions inarteries form all three groups. However, coronary arteriesof young rats contracted more in response to IbTxcompared with those of O-SED and O-ET rats (Fig. 2A).Mean data of dose-dependent IbTx-induced contractionsin the three groups of rats are shown in Fig. 2B.Iberiotoxin caused significantly greater contraction incoronary arteries of young rats than in those of O-SEDrats. Exercise training of old rats significantly enhancedthese contractions. Iberiotoxin (100 nmol l−1) contractedarteries of young, O-SED and O-ET rats to a maximalvalue of 115 ± 14, 36 ± 5.6 and 61 ± 5% of 5-HT-inducedcontractions, respectively. The EC50 of the young group(8.44 ± 0.18 nmol l−1) was significantly different from that
Figure 2. Effects of iberiotoxin (IbTx) on tensions of coronary arteriesA, examples of isometric tension recordings showing the ability of IbTx to cause cumulative dose–responsecontractions of coronary arteries in young, O-SED and O-ET groups. Iberiotoxin caused greater contraction inthe artery from the young than O-ET rat and least in the artery from the O-SED rat. Dotted lines indicate base-line tension. B, mean dose–response curves of IbTx-induced contractions normalized to contractions induced by5 μmol l−1 5-HT in young, O-ET and O-SED groups. Each data point represents the mean ± S.E.M.
of O-SED (12.5 ± 0.2 nmol l−1) and O-ET (14 ± 0.3 nmoll−1). The EC50 values of the two old groups were notsignificantly different. These results indicate that thecontribution of BKCa channel activity to the regulation ofbasal vascular tone in rat coronary arteries is reduced withageing, as indicated by the small increases in tension inresponse to blockade of BKCa channels with IbTx; exercisetraining reversed this reduction.
Exercise training enhanced ageing-reducedBKCa currents
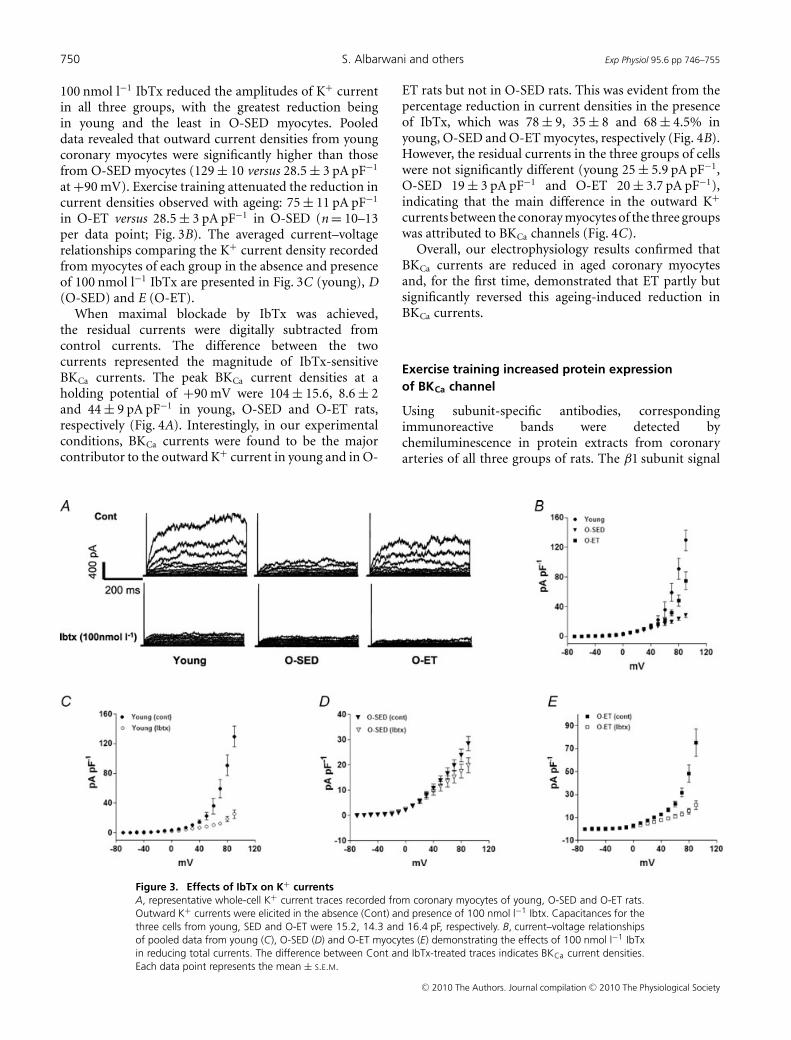
Figure 3A shows representative current traces of young,O-SED and O-ET myocytes in the absence (top) andpresence of 100 nmol l−1 IbTx (bottom). Addition of
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society
750 S. Albarwani and others Exp Physiol 95.6 pp 746–755
100 nmol l−1 IbTx reduced the amplitudes of K+ currentin all three groups, with the greatest reduction beingin young and the least in O-SED myocytes. Pooleddata revealed that outward current densities from youngcoronary myocytes were significantly higher than thosefrom O-SED myocytes (129 ± 10 versus 28.5 ± 3 pA pF−1
at +90 mV). Exercise training attenuated the reduction incurrent densities observed with ageing: 75 ± 11 pA pF−1
in O-ET versus 28.5 ± 3 pA pF−1 in O-SED (n = 10–13per data point; Fig. 3B). The averaged current–voltagerelationships comparing the K+ current density recordedfrom myocytes of each group in the absence and presenceof 100 nmol l−1 IbTx are presented in Fig. 3C (young), D(O-SED) and E (O-ET).
When maximal blockade by IbTx was achieved,the residual currents were digitally subtracted fromcontrol currents. The difference between the twocurrents represented the magnitude of IbTx-sensitiveBKCa currents. The peak BKCa current densities at aholding potential of +90 mV were 104 ± 15.6, 8.6 ± 2and 44 ± 9 pA pF−1 in young, O-SED and O-ET rats,respectively (Fig. 4A). Interestingly, in our experimentalconditions, BKCa currents were found to be the majorcontributor to the outward K+ current in young and in O-
Figure 3. Effects of IbTx on K+ currentsA, representative whole-cell K+ current traces recorded from coronary myocytes of young, O-SED and O-ET rats.Outward K+ currents were elicited in the absence (Cont) and presence of 100 nmol l−1 Ibtx. Capacitances for thethree cells from young, SED and O-ET were 15.2, 14.3 and 16.4 pF, respectively. B, current–voltage relationshipsof pooled data from young (C), O-SED (D) and O-ET myocytes (E) demonstrating the effects of 100 nmol l−1 IbTxin reducing total currents. The difference between Cont and IbTx-treated traces indicates BKCa current densities.Each data point represents the mean ± S.E.M.
ET rats but not in O-SED rats. This was evident from thepercentage reduction in current densities in the presenceof IbTx, which was 78 ± 9, 35 ± 8 and 68 ± 4.5% inyoung, O-SED and O-ET myocytes, respectively (Fig. 4B).However, the residual currents in the three groups of cellswere not significantly different (young 25 ± 5.9 pA pF−1,O-SED 19 ± 3 pA pF−1 and O-ET 20 ± 3.7 pA pF−1),indicating that the main difference in the outward K+
currents between the conoray myocytes of the three groupswas attributed to BKCa channels (Fig. 4C).
Overall, our electrophysiology results confirmed thatBKCa currents are reduced in aged coronary myocytesand, for the first time, demonstrated that ET partly butsignificantly reversed this ageing-induced reduction inBKCa currents.
Exercise training increased protein expressionof BKCa channel
Using subunit-specific antibodies, correspondingimmunoreactive bands were detected bychemiluminescence in protein extracts from coronaryarteries of all three groups of rats. The β1 subunit signal
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

Exp Physiol 95.6 pp 746–755 Ageing, exercise training and BKCa channels 751
Figure 4. Histograms showingdifferential blockade of coronarymyocyte current densities by100 nmol l−1 IbTx at a holding potentialof +90 mVA shows BKCa current densities obtained bydigital subtraction of residual currents in thepresence of 100 nmol l−1 IbTx from controlcurrents. The BKCa current densities weresignificantly different in the three groups ofrats. B depicts the BKCa current as apercentage of the total current in thecoronary myocytes of the three animalgroups. A large proportion of the current inyoung and in O-ET myocytes but not in O-SEDmyocytes was due to BKCa channels. C, theresidual currents after full blockade of BKCa
currents were not significantly different in thethree groups of cells. Bars represent themean + S.E.M. ∗ Significantly different fromyoung (P < 0.05) and † significantly differentfrom O-ET (P < 0.05).
was detected as doublets at molecular mass 45–47 and37 kDa. Pre-adsorption of primary anti-β1 subunit ofBKCa channel with its neutralizing peptide (NP) abolishedthe lower immunoreactive band, indicating the specifityof antibody to the protein corresponding to the lowerband (Fig. 5A).
Figure 5. Western immunoblots comparing the expression of BKCa channel protein in the three ratgroupsA, β1 subunit signal was detected as doublets at molecular mass 45–47 and 37 kDa. Pre-adsorption of primaryanti-β1 subunit with its neutralizing peptide (NP) abolished the lower immunoreactive band, indicating the specifityof antibody to protein corresponding to the lower band (37 kDa). B, reduced band densities were detected forboth α and β1 subunits of BKCa channels in O-SED arteries. Exercise training increased the expression of bothsubunits. C and D, average densitometric values normalized to β-actin densities from 4 Western blots, indicatingthat α subunit (C) and β1 subunits of BKCa channels (D) are significantly reduced in aged coronary arteries and thatexercise training increased the expression of both subunits. Bars represent means + S.E.M. ∗ Significantly differentfrom young (P < 0.05) and † significantly different from O-ET (P < 0.05).
Representative immunoblots of α (n = 4) andβ1 subunit proteins (n = 4) show that protein expressionof both subunits decreased with ageing, and ET attenuatedthis reduction (Fig. 5B). The relative amount of the α
and β1 subunits of the BKCa channel normalized to theβ-actin signal in four different membranes are shown in
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

752 S. Albarwani and others Exp Physiol 95.6 pp 746–755
Fig. 5C and D. On average, in O-SED coronary arteries, theprotein expression level of α and β1 subunits was reducedby 40 ± 3.5 and 30 ± 2.6%, respectively, compared withthe coronary arteries of young rats. Exercise trainingattenuated this decline in expression level to 28 ± 2 and12 ± 4%, respectively. This indicated that ET improvedthe expression level of α and β1 subunits by 20 and 28%,respectively, from the sedentary level.
Discussion
The aim of this study was to test the hypothesis thatexercise training of moderate intensity would attenuateage-induced downregulation of BKCa channels in coronaryarteries of old rats. Our results demonstrated threemain observations that support our hypothesis. First,the contraction of isolated coronary arteries in responseto IbTx was reduced in old rats compared with younganimals. Second, the magnitude of BKCa current densitieswas reduced in old compared with young myocytes. Third,the expression of α and β1 protein of BKCa channelsdecreased in old rats. Exercise training partly reversed allthese effects of ageing. Thus, from these observations itcould be deduced that BKCa channel activity declines withageing, and the present study provides, for the first time,evidence that exercise training attenuates this reduction.
The ability of IbTx to induce contractions of isolatedarteries indicated that BKCa channels are open in theresting state, and their closure depolarized the membrane,leading to influx of Ca2+ through VGCC channels andthereby causing contraction (Nelson & Quayle 1995). Inour experiments, arteries from the three animal groupscontracted in response to IbTx. However, arteries of youngrats contracted significantly more than those of O-ET ratsand to a greater degree compared with arteries of O-SEDrats.
Based on our results, one would expect that coronaryarteries isolated from O-SED rats would exhibit higherresting tone compared with those from young rats, andET would reduce this tone. On the contrary, our resultsshowed that the basal tone of coronary arteries isolatedfrom O-SED rats was lower than that of young rats. Thecause of such paradoxical results is not clear at this stage.The increased BKCa channel activity induced by ET isexpected to be associated with hyperpolarization, reducedcalcium influx and therefore reduced resting tone (Nelson& Quayle, 1995; Gollasch & Nelson, 1997). In this study,we found no significant difference in resting tone and,consistent with this, in Ca2+-dependent tone between O-SED and O-ET groups. This observation is in agreementwith the findings of Heaps et al. (2000), who demonstratedthat ET enhanced VGCC currents without a change in thebasal tone or the net cytoplasmic Ca2+ in sedentary andexercise-trained pigs. The authors, to explain their finding,suggested the presence of other cellular mechanism(s) that
keep intracellular calcium low. It is possible that increasedBKCa channel density is, at least in part, responsiblefor maintaining intracellular calcium low in the face ofincreased VGCC channel activity and greater spontaneoussarcoplasmic reticulum calcium release, which was shownto occur with ET (Stehno-Bittel et al. 1991).
Based on the differential increase in resting tone inresponse to IbTx, we hypothesized that BKCa currentsare reduced in coronary myocytes of O-SED rats. In thisstudy, we report significant reduction in IbTx-sensitiveBKCa currents in myocytes of O-SED rats comparedwith young rats. Ageing-induced reduction BKCa channelactivity has previously been reported in coronary myocytes(Marijic et al. 2001) and in cardiac mitochondrialmembranes (Heinen et al. 2008). We also demonstratedthat ET increased BKCa currents in coronary myocytes.Interestingly, the net contribution of BKCa currents tototal K+ currents was also significantly increased in O-ET compared with O-SED rats.
Bowles et al. (1998b) showed that conduit coronaryarteries from young ET pigs contracted more in responseto IbTx compared with arteries from sedentary animals.However, contrary to our electrophysiological findings,they found no significant difference in K+ currentsbetween the two groups. This discrepancy is not yet fullyunderstood; however, it may arise in the following manner.First, our study compared the effects of ET on old rats. Itis possible that while ageing reduces the expression ofBKCa channels, old animals still retain the ability to adaptpositively to ET by increasing the activity of BKCa channelsby a mechanism that appears to relate to the expressionlevel of BKCa channels. Young animals, on the other hand,increase activity of the BKCa channel via a mechanismthat does not involve changes in its expression, such aschanges in channel gating or in channel modulation byintracellular messengers (Schubert & Nelson, 2001; Houet al. 2009). Second, it is well accepted that ET-associatedadaptations are not uniform in different segments of thecoronary arterial tree. For example, ET has been shown toenhance endothelium-dependent vasorelaxation in smallarterioles, but not in conduit coronary arteries (Parkeret al. 1994; Duncker & Bache, 2008). Therefore, it ispossible that ET induces BKCa channel upregulation insmall arteries, where they are thought to play a major rolein regulating vascular tone and resistance, whereas theyplay a minor role in larger conduit arteries. Third, it is alsolikely that one of the potential causes of this discrepancybetween studies is the difference in animal species usedand in ET protocols.
To find out whether the differences in BKCa currentswere associated with changes in the pore-formingchannel protein (α subunit) or the regulatory subunit(β1 subunit), we compared the expression levels of thesetwo proteins in the three groups of rats. Our results showedthat ageing downregulates both α and β1 subunits of the
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

Exp Physiol 95.6 pp 746–755 Ageing, exercise training and BKCa channels 753
BKCa channel, as reported earlier (Marijic et al. 2001;Nishamura et al. 2004). Furthermore, and for the firsttime, we demonstrated that ET increased the expressionlevel of both α and β1 subunit protein. However, thedegree of change in expression levels of both subunitswas not comparable to changes in basal tones in responseto blockade of BKCa channels in O-SED and O-ET groups.In O-SED, α and β1 subunit proteins were reduced by40 and 30%, respectively, while the contraction inducedby 100 nmol l−1 IbTx was reduced by 76%. Similarly, ETincreased α and β1 subunit proteins by 20 and 28%,respectively, compared with O-SED, while contractioninduced by 100 nmol l−1 IbTx was increased by 70%.One possible reason for this descepancy is that coronarytissue used for Western immunoblotting was pooledfrom the main right and left coronary arteries and theirbranches. Thus, these results may not precisely representthe adaptation that took place in arteries studied byelectrophysiology and tension-recording experiments.
It is important to address a few methodologicallimitations of our study that may have influenced ourfindings. First, our study did not include an exercise-trained group of young rats. Therefore, it is not possibleto determine whether upregulation of BKCa channels isa specific adaptation due to interaction between ageingand exercise or whether it is a general adaptation thatoccurs in all ages in response to exercise. Second, ourisolated arteries experiment was performed in the presenceof intact endothelium. Different endothelium-derivedfactors have been reported to alter the activity of vascularBKCa channels (Tanaka et al. 2004). In our study, thesefactors may have influenced the reduced or enhancedcontribution of BKCa channels to basal tone in O-SEDand O-ET, respectively. For instance, endothelin-1 hasbeen shown to have an inhibitory effect on BKCa channels(Betts & Kozlowski, 2000). Expression of endothelin-1in vascular endothelial cells increases in aged sedentarymen (Donato et al. 2009). This increase is abolished byaerobic exercise training (Van Guilder et al. 2007; Maedaet al. 2009). Therefore, in our experiments endothelin-1 may have contributed to the reduced BKCa channelactivity observed in O-SED rats. Similarly, many studieshave demonstrated the role of BKCa channels in NO- andprostacylin-induced blood vessel relaxation (Tanaka et al.2004). Although coronary NO and endothelial nitric oxidesynthase were shown equivocally to be reduced in ageing(Korzick et al. 2005; Shipley & Muller-Delp 2005) andincreased in exercise training (Sessa et al. 1994; Laughlinet al. 2001) this remains as a possible contributory factorto our observations.
The benefits of exercise in reducing the risk ofcoronary artery disease in young and old humans are welldocumented. However, the interaction between ageingand ET remains incompletely understood. In this study,we have provided evidence that in old rats, coronary
myocytes responded to graded exercise by upregulation ofBKCa channels. This enhancement of channel activity mayprovide a more efficient negative feedback system to meetthe metabolic demands of aged coronary myocytes duringexercise. Since the activation of vascular BKCa channelsacts as an important feedback mechanism that opposesexcitation and limits contraction, increased activity mayhave a large impact on limiting excessive excitability, thuspreventing vasospasm of aged coronary vasculature. Morestudies on the molecular mechanisms involved in delayingthe ageing changes in coronary vasculature are required todevelop strategies to improve the quality of life of theever-increasing ageing population.
References
Albarwani S, Nemetz LT, Madden JA, Tobin AA, England SK,Pratt PF & Rusch NJ (2003). Voltage-gated K+ channels inrat small cerebral arteries: molecular identity of thefunctional channels. J Physiol 551, 751–763.
Amberg GC & Santana LF (2003). Down regulation of the BKchannel β1 subunit in genetic hypertension. Circ Res 93,965–971.
Betts LC & Kozlowski RZ (2000). Electrophysiological effects ofendothelin-1 and their relationship to contraction in ratrenal arterial smooth muscle. Br J Pharmacol 130, 787–796.
Blanco-Centurion CA & Shiromani PJ (2006). Beneficial effectsof regular exercise on sleep in old F344 rats. Neurobiol Aging27, 1859–1869.
Borbouse L, Dick GM, Asano S, Bender SB, Dincer UD, PayneGA, Neeb ZP, Bratz IN, Sturek M & Tune JD (2009).Impaired function of coronary BKCa channels in metabolicsyndrome. Am J Physiol Heart Circ Physiol 297,H1629–H1637.
Bowles DK, Hu Q, Laughlin MH & Sturek M (1998a). Exercisetraining increases L-type calcium current density incoronary smooth muscle. Am J Physiol Heart Circ Physiol275, H2159–H2169.
Bowles DK, Laughlin MH & Sturek M (1998b). Exercisetraining increases K+-channel contribution to regulation ofcoronary arterial tone. J Appl Physiol 84, 1225–1233.
Brayden JE & Nelson MT (1992). Regulation of arterial tone byactivation of calcium-dependent potassium channels. Science256, 532–535.
Brenner R, Perez GJ, Bonev AD, Eckman DM, Kosek JC, WilerSW, Patterson AJ, Nelson MT & Aldrich RW (2000).Vasoregulation by the β1 subunit of the calcium-activatedpotassium channel. Nature 407, 870–876.
Burnham MP, Johnson IT & Weston AH (2006). ReducedCa2+-dependent activation of large-conductanceCa2+-activated K+ channels from arteries of Type 2 diabeticZucker diabetic fatty rats. Am J Physiol Heart Circ Physiol290, H1520–H1527.
Chandrashekhar Y & Anand IS (1991). Exercise as a coronaryprotective factor. Am Heart J 122, 1723–1739.
Cox DH & Aldrich RW (2000). Role of the β1 subunit in largeconductance Ca2+-activated K+ channel gating energetics.Mechanisms of enhanced Ca2+ sensitivity. J Gen Physiol 116,411–432.
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

754 S. Albarwani and others Exp Physiol 95.6 pp 746–755
DeSouza CA, Shapiro LF, Clevenger CM, Dinenno FA,Monahan KD, Tanaka H & Seals DR (2000). Regular aerobicexercise prevents and restores age-related declines inendothelium-dependent vasodilation in healthy men.Circulation 102, 1351–1357.
Dick GM, Rossow CF, Smirnov S, Horowitz B & Sanders KM(2001). Tamoxifen activates smooth muscle BK channelsthrough the regulatory β1 subunit. J Biol Chem 276,34594–34599.
Donato AJ, Gano LB, Eskurza I, Silver AE, Gates PE, JablonskiK & Seals DR (2009). Vascular endothelial dysfunction withaging: endothelin-1 and endothelial nitric oxide synthase.Am J Physiol Heart Circ Physiol 297, H425–H432.
Duncker DJ & Bache RJ (2008). Regulation of coronary bloodflow during exercise. Physiol Rev 88, 1009–1086.
Fletcher GF, Balady G, Blair SN, Blumenthal J, Caspersen C,Chaitman B, Epstein S, Sivarajan Froelicher ES, FroelicherVF, Pina IL & Pollock ML (1996). Statement on exercise:benefits and recommendations for physical activityprograms for all Americans. A statement for healthprofessionals by the Committee on Exercise and CardiacRehabilitation of the Council on Clinical Cardiology,American Heart Association. Circulation 94, 857–862.
Garcia ML, Hanner M, Knaus HG, Slaughter R & KaczorowskiGJ (1999). Scorpion toxins as tools for studying potassiumchannels. Methods Enzymol 294, 624–639.
Gollasch M & Nelson MT (1997). Voltage-dependent Ca2+channels in arterial smooth muscle cells. Kidney Blood PressRes 20, 355–371.
Halpern W, Mulvany MJ & Warshaw DM (1978). Mechanicalproperties of smooth muscle cells in the walls of arterialresistance vessels. J Physiol 275, 85–101.
Heaps CL, Bowles DK, Sturek M, Laughlin MH & Parker JL(2000). Enhanced L-type Ca2+ channel current density incoronary smooth muscle of exercise-trained pigs iscompensated to limit myoplasmic free Ca2+ accumulation.J Physiol 528, 435–445.
Heinen A, Winning A, Schlack W, Hollmann MW, Preckel B,Fraßdorf J & Weber NC (2008). The regulation ofmitochondrial respiration by opening of mKCa channels isage-dependent. Eur J Pharmacol 578, 108–113.
Hou S, Heinemann SH & Hoshi T (2009). Modulation of BKCa
channel gating by endogenous signaling molecules.Physiology (Bethesda) 24, 26–35.
Ishida S, Hamasaki S, Kamekou M, Yoshitama T, Nakano F,Yoshikawa A, Kataoka T, Saihara K, Minagoe S & Tei C(2003). Advancing age is associated with diminished vascularremodeling and impaired vasodilation in resistance coronaryarteries. Coron Artery Dis 14, 443–449.
Kim EY, Zou S, Ridgway LD & Dryer SE (2007). β1-Subunitsincrease surface expression of a large-conductanceCa2+-activated K+ channel isoform. J Neurophysiol 97,3508–3516.
Korzick DH, Muller-Delp JM, Dougherty P, Heaps CL, BowlesDK & Krick KK (2005). Exaggerated coronary vasoreactivityto endothelin-1 in aged rats: role of protein kinase C.Cardiovasc Res 66, 384–392.
Laughlin MH & Muller JM (1998). Vasoconstrictor responsesof coronary resistance arteries in exercise-trained pigs. J ApplPhysiol 84, 884–889.
Laughlin MH, Pollock JS, Amann JF, Hollis ML, Woodman CR& Price EM (2001). Training induces nonuniform increasesin eNOS content along the coronary arterial tree. J ApplPhysiol 90, 501–510.
Maeda S, Sugawara J, Yoshizawa M, Otsuki T, Shimojo N,Jesmin S, Ajisaka R, Miyauchi T & Tanaka H (2009).Involvement of endothelin-1 in habitual exercise-inducedincrease in arterial compliance. Acta Physiol (Oxf) 196,223–229.
Marijic J, Li Q, Song M, Nishimaru K, Stefani E & Toro L(2001). Decreased expression of voltage- and Ca2+-activatedK+ channels in coronary smooth muscle during aging. CircRes 88, 210–216.
Navarro-Antolın J, Levitsky KL, Calderon E, Ordonez A &Lopez-Barneo J (2005). Decreased expression of maxi-K+channel β1-subunit and altered vasoregulation in hypoxia.Circulation 112, 309–315.
Nelson MT, Patlak JB, Worley JF & Standen NB (1990).Calcium channels, potassium channels, and voltagedependence of arterial smooth muscle tone. Am J Physiol CellPhysiol 259, C3–C18.
Nelson MT & Quayle JM (1995). Physiological roles andproperties of potassium channels in arterial smooth muscle.Am J Physiol Cell Physiol 268, C799–C822.
Nishimaru K, Eghbali M, Lu R, Marijic J, Stefani E & Toro L(2004). Functional and molecular evidence of MaxiKchannel β1 subunit decrease with coronary artery ageing inthe rat. J Physiol 559, 849–862.
Parker JL, Oltman CL, Muller JM, Myers PR, Adams HR &Laughlin MH (1994). Effects of exercise training onregulation of tone in coronary arteries and arterioles. MedSci Sports Exerc 26, 1252–1261.
Schubert R & Nelson MT (2001). Protein kinases: tuners of theBKCa channel in smooth muscle. Trends Pharmacol Sci 22,505–512.
Sessa WC, Pritchard K, Seyedi N, Wang J & Hintze TH (1994).Chronic exercise in dogs increases coronary vascular nitricoxide production and endothelial cell nitric oxide synthasegene expression. Circ Res 74, 349–353.
Shipley RD & Muller-Delp JM (2005). Aging decreasesvasoconstrictor responses of coronary resistance arteriolesthrough endothelium-dependent mechanisms. CardiovascRes 66, 374–383.
Stehno-Bittel L, Laughlin MH & Sturek M (1991). Exercisetraining depletes sarcoplasmic reticulum calcium incoronary smooth muscle. J Appl Physiol 71,1764–1773.
Stewart KJ (2005). Physical activity and aging. Ann N Y AcadSci 1055, 193–206.
Tanaka S, Tamaya N, Matsuzawa K & Miyaishi O (2000).Differences in survivability among F344 rats. Exp Anim 49,141–145.
Tanaka Y, Koike K & Toro L (2004). MaxiK channel roles inblood vessel relaxations induced by endothelium-derivedrelaxing factors and their molecular mechanisms. J SmoothMuscle Res 40, 125–153.
Tanaka Y, Meera P, Song M, Knaus HG & Toro L (1997).Molecular constituents of maxi KCa channels in humancoronary smooth muscle: predominant α + β subunitcomplexes. J Physiol 502, 545–557.
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society

Exp Physiol 95.6 pp 746–755 Ageing, exercise training and BKCa channels 755
Van Guilder GP, Westby CM, Greiner JJ, Stauffer BL &DeSouza CA (2007). Endothelin-1 vasoconstrictor toneincreases with age in healthy men but can be reduced byregular aerobic exercise. Hypertension 50, 403–409.
Wallner M, Meera P & Toro L (1999). Molecular basis of fastinactivation in voltage and Ca2+-activated K+ channels: atransmembrane β-subunit homolog. Proc Natl Acad Sci USA96, 4137–4142.
Wei JY, Li Y & Ragland J (1987). Effect of exercise training onresting blood pressure and heart rate in adult and aged rats.J Gerontol 42, 11–16.
Yildiz O (2007). Vascular smooth muscle and endothelialfunctions in aging. Ann N Y Acad Sci 1100, 353–360.
Ziada AM, Hassan MO, Tahlilkar KI & Inuwa IM (2005).Long-term exercise training and angiotensin-convertingenzyme inhibition differentially enhance myocardialcapillarization in the spontaneously hypertensive rat.J Hypertens 23, 1233–1240.
Acknowledgements
We would like to thank the staff of the Small Animal House ofSultan Qaboos University for their great help in exercising therats. This work was funded by Sultan Qaboos University, grantnumber IG/MED/PHY/05/02, to S. Albarwani.
C© 2010 The Authors. Journal compilation C© 2010 The Physiological Society
Copyright © 2022 FDOKUMEN




















