
Juvenile Administration of Methylphenidate Attenuates Adult Hippocampal Neurogenesis
10
Juvenile Administration of Methylphenidate Attenuates Adult Hippocampal Neurogenesis Diane C. Lagace, Jessica K. Yee, Carlos A. Bolaños, and Amelia J. Eisch Background: The neural consequences of early-life exposure to methylphenidate (MPH; Ritalin) are of great interest given the widespread, and sometimes inappropriate, use in children. Here we examine the impact of juvenile MPH exposure on adult hippocampal neurogenesis. Methods: Rats received MPH (2.0 mg/kg, intraperitoneal, twice daily) or saline (SAL) during preadolescence (postnatal days 20 –35). Hippocampal cell proliferation (Experiment 1), neurogenesis (Experiment 2), and stress-induced changes in cell proliferation (Experiment 3) were assessed at several developmental stages including adulthood. Results: Juvenile exposure to MPH did not alter proliferation at any developmental time point relative to control rats; however, exposure to MPH significantly decreased the long-term survival of newborn cells in adult rats, particularly in the temporal hippocampus. Although MPH-treated rats had higher levels of corticosterone after restraint stress, they did not show the expected greater decrease in hippocampal cell proliferation relative to control animals. Conclusions: Early-life exposure to MPH inhibits the survival of adult-generated neurons in the temporal hippocampus and may reduce progenitor sensitivity to corticosterone-induced decreases in proliferation. These findings suggest that decreased adult neurogenesis is an enduring consequence of early-life exposure to MPH and are discussed for their relevance to humans. Key Words: ADHD, BrdU, development, hippocampus, psycho- stimulant, Ritalin M ethylphenidate (MPH; Ritalin) is commonly prescribed for the treatment of attention-deficit/hyperactivity disor- der (ADHD). Treatment with MPH is generally well tolerated and effective in reducing symptoms associated with ADHD: inattentiveness, impulsivity, and hyperactivity (Abikoff et al 2004; Jensen et al 2001; Wilens and Dodson 2004). How- ever, given that the prescription rate of this psychostimulant for children, including preschoolers, is on the rise (Fogelman et al 2003; Frances et al 2002; Miller et al 2001; Olfson et al 2002; Schmidt-Troschke et al 2004; Zito et al 2000), researchers are actively investigating the long-term consequences of preadoles- cent MPH treatment on brain and behavior (Andersen 2005; Carlezon and Konradi 2004). The urgency of this issue is underscored by the prescription of MPH to children who do not meet full diagnostic criteria for ADHD (Angold et al 2000). Fueling speculation that exposure to MPH in early life may be detrimental to brain development, administration of MPH to young rodents produces notable changes in behavior, neuro- physiology, and biochemistry in adulthood. Behavioral changes seen after juvenile exposure to MPH (postnatal day [D] 20 –35) include decreased response to rewarding stimuli, as well as increases in depressive- and anxiety-like behaviors (Achat- Mendes et al 2003; Adriani et al 2005; Andersen et al 2002; Bolanos et al 2003; Carlezon et al 2003; Mague et al 2005). Physiologic and biochemical changes include decreased activity of dopaminergic neurons (Brandon et al 2001; Federici et al 2005), increased levels of cAMP (cyclic adenosine monophos- phate) response element-binding (CREB) protein and Homer 1A transcript (Adriani et al 2005; Andersen et al 2002), and enhanced corticosterone levels following restraint stress (Bolanos et al 2003). One interpretation of these studies is that juvenile admin- istration of MPH produces a depressive- or anxiety-like pheno- type in adulthood, marked by anhedonia and decreased physi- ological and behavioral correlates of reward (Andersen and Navalta 2004; Carlezon and Konradi 2004). Although this inter- pretation deserves careful testing for its validity and its applica- bility to humans (Hyman 2003; Kuczenski and Segal 2005; Volkow and Insel 2003), clearly these findings urge additional exploration of the long-term consequences of developmental administration of MPH in rodents, particularly in regard to neural substrates linked to affect and reward. Although the brain’s reward neurocircuitry has been exam- ined for adult biochemical and molecular adaptations seen after juvenile MPH exposure (Andersen 2005), the hippocampus has received relatively little attention. The hippocampus plays a prominent role in affective behaviors, drug taking, and relapse (Carlezon et al 2005; Duman et al 1997; Nestler et al 2002), making it a prime target in testing the hypothesis that juvenile exposure to MPH leads to a depressive-like phenotype in adulthood. Moreover, because acute MPH treatment in adoles- cent rodents dose-dependently increases norepinephrine levels in the hippocampus in the absence of altering dopamine levels in the nucleus accumbens (Kuczenski and Segal 2002), MPH ac- tions in the hippocampus may be central to MPH long-term effects. The hippocampus in turn can influence the brain’s reward neurocircuitry and psychostimulant-related behaviors (Charara and Grace 2003; Emerich and Walsh 1990; Mitchell et al 2000; Won et al 2003; Yang and Mogenson 1987). Thus, identi- fying MPH-induced changes in hippocampal physiology or func- tion will be critical in interpreting MPH-induced changes in reward circuitry and behavior. One aspect of hippocampal plasticity that has received much attention in regard to both depression and addiction is adult neurogenesis, or the ability to give rise to new neurons through- out life (Duman 2004; Eisch 2002). Chronic exposure to antide- pressant drugs increases the number of neural progenitors in the rodent subgranular zone (SGZ) of the dentate gyrus (DG) and can block the decrease in neurogenesis induced by stress (Czeh et al 2001; Malberg and Duman 2003; Malberg et al 2000; From the Department of Psychiatry (DCL, JKY, AJE), University of Texas South- western Medical Center, Dallas, Texas; Department of Psychology and Program in Neuroscience (CAB), Florida State University, Tallahassee, Florida. Address reprint requests to Dr. Amelia J. Eisch, Department of Psychiatry, University of Texas Southwestern Medical Center, Dallas, TX 75390-9070; E-mail: [email protected]. Received January 26, 2006; revised March 31, 2006; accepted April 4, 2006. BIOL PSYCHIATRY 2006;60:1121–1130 0006-3223/06/$32.00 doi:10.1016/j.biopsych.2006.04.009 © 2006 Society of Biological Psychiatry
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Juvenile Administration of Methylphenidate Attenuates Adult Hippocampal Neurogenesis

Juvenile Administration of MethylphenidateAttenuates Adult Hippocampal NeurogenesisDiane C. Lagace, Jessica K. Yee, Carlos A. Bolaños, and Amelia J. EischBackground: The neural consequences of early-life exposure to methylphenidate (MPH; Ritalin) are of great interest given thewidespread, and sometimes inappropriate, use in children. Here we examine the impact of juvenile MPH exposure on adulthippocampal neurogenesis.Methods: Rats received MPH (2.0 mg/kg, intraperitoneal, twice daily) or saline (SAL) during preadolescence (postnatal days 20–35).Hippocampal cell proliferation (Experiment 1), neurogenesis (Experiment 2), and stress-induced changes in cell proliferation(Experiment 3) were assessed at several developmental stages including adulthood.Results: Juvenile exposure to MPH did not alter proliferation at any developmental time point relative to control rats; however,exposure to MPH significantly decreased the long-term survival of newborn cells in adult rats, particularly in the temporalhippocampus. Although MPH-treated rats had higher levels of corticosterone after restraint stress, they did not show the expected greaterdecrease in hippocampal cell proliferation relative to control animals.Conclusions: Early-life exposure to MPH inhibits the survival of adult-generated neurons in the temporal hippocampus and mayreduce progenitor sensitivity to corticosterone-induced decreases in proliferation. These findings suggest that decreased adultneurogenesis is an enduring consequence of early-life exposure to MPH and are discussed for their relevance to humans.
Key Words: ADHD, BrdU, development, hippocampus, psycho-stimulant, Ritalin
Methylphenidate (MPH; Ritalin) is commonly prescribedfor the treatment of attention-deficit/hyperactivity disor-der (ADHD). Treatment with MPH is generally well
tolerated and effective in reducing symptoms associated withADHD: inattentiveness, impulsivity, and hyperactivity (Abikoffet al 2004; Jensen et al 2001; Wilens and Dodson 2004). How-ever, given that the prescription rate of this psychostimulant forchildren, including preschoolers, is on the rise (Fogelman et al2003; Frances et al 2002; Miller et al 2001; Olfson et al 2002;Schmidt-Troschke et al 2004; Zito et al 2000), researchers areactively investigating the long-term consequences of preadoles-cent MPH treatment on brain and behavior (Andersen 2005;Carlezon and Konradi 2004). The urgency of this issue isunderscored by the prescription of MPH to children who do notmeet full diagnostic criteria for ADHD (Angold et al 2000).
Fueling speculation that exposure to MPH in early life may bedetrimental to brain development, administration of MPH toyoung rodents produces notable changes in behavior, neuro-physiology, and biochemistry in adulthood. Behavioral changesseen after juvenile exposure to MPH (postnatal day [D] 20–35)include decreased response to rewarding stimuli, as well asincreases in depressive- and anxiety-like behaviors (Achat-Mendes et al 2003; Adriani et al 2005; Andersen et al 2002;Bolanos et al 2003; Carlezon et al 2003; Mague et al 2005).Physiologic and biochemical changes include decreased activityof dopaminergic neurons (Brandon et al 2001; Federici et al2005), increased levels of cAMP (cyclic adenosine monophos-phate) response element-binding (CREB) protein and Homer 1A
transcript (Adriani et al 2005; Andersen et al 2002), and enhancedcorticosterone levels following restraint stress (Bolanos et al2003). One interpretation of these studies is that juvenile admin-istration of MPH produces a depressive- or anxiety-like pheno-type in adulthood, marked by anhedonia and decreased physi-ological and behavioral correlates of reward (Andersen andNavalta 2004; Carlezon and Konradi 2004). Although this inter-pretation deserves careful testing for its validity and its applica-bility to humans (Hyman 2003; Kuczenski and Segal 2005;Volkow and Insel 2003), clearly these findings urge additionalexploration of the long-term consequences of developmentaladministration of MPH in rodents, particularly in regard to neuralsubstrates linked to affect and reward.
Although the brain’s reward neurocircuitry has been exam-ined for adult biochemical and molecular adaptations seen afterjuvenile MPH exposure (Andersen 2005), the hippocampus hasreceived relatively little attention. The hippocampus plays aprominent role in affective behaviors, drug taking, and relapse(Carlezon et al 2005; Duman et al 1997; Nestler et al 2002),making it a prime target in testing the hypothesis that juvenileexposure to MPH leads to a depressive-like phenotype inadulthood. Moreover, because acute MPH treatment in adoles-cent rodents dose-dependently increases norepinephrine levelsin the hippocampus in the absence of altering dopamine levels inthe nucleus accumbens (Kuczenski and Segal 2002), MPH ac-tions in the hippocampus may be central to MPH long-termeffects. The hippocampus in turn can influence the brain’sreward neurocircuitry and psychostimulant-related behaviors(Charara and Grace 2003; Emerich and Walsh 1990; Mitchell et al2000; Won et al 2003; Yang and Mogenson 1987). Thus, identi-fying MPH-induced changes in hippocampal physiology or func-tion will be critical in interpreting MPH-induced changes inreward circuitry and behavior.
One aspect of hippocampal plasticity that has received muchattention in regard to both depression and addiction is adultneurogenesis, or the ability to give rise to new neurons through-out life (Duman 2004; Eisch 2002). Chronic exposure to antide-pressant drugs increases the number of neural progenitors in therodent subgranular zone (SGZ) of the dentate gyrus (DG) andcan block the decrease in neurogenesis induced by stress (Czehet al 2001; Malberg and Duman 2003; Malberg et al 2000;
From the Department of Psychiatry (DCL, JKY, AJE), University of Texas South-western Medical Center, Dallas, Texas; Department of Psychology andProgram in Neuroscience (CAB), Florida State University, Tallahassee,Florida.
Address reprint requests to Dr. Amelia J. Eisch, Department of Psychiatry,University of Texas Southwestern Medical Center, Dallas, TX 75390-9070;E-mail: [email protected].
Received January 26, 2006; revised March 31, 2006; accepted April 4, 2006.
BIOL PSYCHIATRY 2006;60:1121–11300006-3223/06/$32.00doi:10.1016/j.biopsych.2006.04.009 © 2006 Society of Biological Psychiatry

Santarelli et al 2003). In contrast, numerous drugs of abuse (Eischet al 2000; Eisch and Harburg 2006; Opanashuk et al 2001;Yamaguchi et al 2004), including psychostimulants such asmethamphetamine (Teuchert-Noodt et al 2000), decrease neuro-genesis. Negative early-life experiences can also have a negativeimpact on adult hippocampal neurogenesis and function (Kartenet al 2005; Lemaire et al 2000; Mirescu et al 2004). It is unknownhow early-life exposure to MPH affects adult hippocampalneurogenesis. However, given that juvenile MPH exposure pro-duces a depressive-like phenotype in adult rats (Bolanos et al2003; Carlezon et al 2003) and MPH is a psychostimulant, it isreasonable to hypothesize that juvenile MPH exposure willdiminish adult hippocampal neurogenesis.
We performed three experiments to assess adult hippocampalneurogenesis after juvenile exposure to MPH. Because adultneurogenesis is a process, not a time point (Kempermann et al2004), the first and second experiments explore the impact ofjuvenile MPH exposure on the proliferation and the survival ofnewborn cells, respectively. Because early exposure to MPH isalso associated with enhanced corticosterone response to stress(Bolanos et al 2003) and corticosterone inhibits cell proliferation(Gould and Tanapat 1999), the third experiment investigateswhether proliferation is altered in MPH rats after stress. Althoughthese studies explore the specific hypothesis that early-lifeexposure to MPH is detrimental to hippocampal neurogenesis inadulthood, in general they begin to provide much-needed insightinto hippocampal neuroadaptations induced after juvenile expo-sure to MPH.
Methods and Materials
Rats and Drug TreatmentLactating female Sprague–Dawley rats with their male pups
were purchased from Harlan (Indianapolis, Indiana). Pups wereat D14 on arrival and were given 4 days of acclimation beforeweaning at D18. Drug treatment lasted for 16 days (D20–35),during which the rats received intraperitoneal (IP) injectionstwice a day (12 AM and 6 PM) of either MPH hydrochloride (MPHHCl, 2.0 mg/kg, dissolved in .9% saline as 1 mL/kg, SigmaLaboratories, St. Louis, Missouri) or .9% saline vehicle injections(SAL, 1.0 mg/kg). Rats were weighed daily during drug treat-ment, and, as previously reported (Bolanos et al 2003), weightsbetween SAL and MPH rats did not differ significantly during theperiod of SAL or MPH administration or on the day of sacrifice(data not shown). The animal colony was in an Association forthe Assessment and Accreditation of Laboratory Animal Care–approved facility and followed a 12–12 light–dark cycle. All ratswere provided with food and water ad libitum. Experimentswere conducted in compliance with the National Institute ofHealth and University of Texas Southwestern’s institutional re-view committee.
Experimental DesignAs depicted in Table 1, each treatment group (SAL or MPH)
was divided into six subgroups that were named according to thepostnatal day of sacrifice: D46 (adolescence), D77 (early adult-hood), or D90A, D90B, D90C, and D112 (adulthood).
Locomotor Activity TestingTo examine whether juvenile MPH affects motor activity
during different developmental stages, separate groups of ratswere assessed for locomotor behavior during adolescence (D46),early adulthood (D77), or adulthood (D90C and D112). This testwas performed to confirm and augment previous findings that
juvenile administration of MPH reduces reactivity to a novelenvironment in adulthood (Bolanos et al 2003; Carlezon et al2003). Locomotor response was assessed in a dedicated group(D90C) run in parallel with rats given bromodeoxyuridine (BrdU)(D90B), or several days before BrdU injection and sacrifice (D46,D77, D112). Each rat was placed inside a circular activitychamber, a setting novel to the rat (75 cm ! 15 cm; diameter !width, Med Associates, St. Albans, Vermont). Activity inside thenovel environment was detected by four photocell beams, whichwere computer-monitored (Med Associates) to record move-ment. Each rat was assessed for 90 min at 5-min intervals. Dataare reported as movement counts.
BrdU Injections and Tissue PreparationTo label SGZ progenitors at various developmental stages,
rats received an IP injection of BrdU (150 mg/kg; BoehringerMannheim, Mannheim, Germany) in adolescence (D46), earlyadulthood (D77), and adulthood (D90A, D90B, D112). To assesslevels of SGZ progenitor proliferation or survival, rats weresacrificed either 2 hours (D46, D77, D90A, D90B) or 4 weeks(D112) following BrdU injection. All rats were anesthetized withchloral hydrate and perfused transcardially with cold .1 mmol/mLphosphate buffered saline (PBS) followed by 4% paraformaldehydein .1 mmol/mL PBS (pH 7.4) at a rate of 7 mL/min and processed aspreviously described (Eisch et al 2000; Mandyam et al 2004). Brainswere sectioned coronally on a freezing microtome (Leica, Wet-zlar, Germany) at 30 "m through the hippocampus. Nine serialsets of sections were stored in .1% NaN3 in 1X PBS at 4oC untilprocessing.
ImmunohistochemistryTo assess levels of cell proliferation, cell survival, and cell fate,
BrdU was detected via diaminobenzidine (DAB) or fluorescentimmunohistochemistry (IHC; Eisch et al 2000). Serial sets of sectionsthrough the hippocampus were ordered from rostral to caudal,mounted on glass slides (Fisher Superfrost/Plus, Hampton, New
Table 1. Experiment Design and Time Course
Group
MPH or SALDay 20–35
(n # 69)Locomotion
TestingRestraint
StressBrdU
Injection Perfusion
D46 MPH (n # 6) Day 44 — Day 46 Day 46Saline (n # 6)
D77 MPH (n # 6) Day 74 — Day 77 Day 77Saline (n # 6)
D90A MPH (n # 4) — — Day 90 Day 90Saline (n # 6)
D90B MPH (n # 5) — Day 90 Day 90 Day 90Saline (n # 6)
D90C MPH (n # 6) Day 90 — — —Saline (n # 6)
D112 MPH (n # 6) Day 109 — Day 85 Day 112Saline (n # 6)
Juvenile rats (n # 69) received methylphenidate (MPH; 2.0 mg/kg) orsaline (SAL, 1.0 mg/kg) treatment twice daily from postnatal (D) 20 to D35.They were divided into six groups and named occurring to the postnatal dayof sacrifice. Locomotor activity in a novel environment was assessed in ratsduring adolescence (D46), young adulthood (D77), and adulthood (D90C,D112). Experiment 1 assessed dentate gyrus (DG) cell proliferation duringadolescence (D46), young adulthood (D77), and adulthood (D90A) 2 hoursafter BrdU injection. Experiment 2 assessed DG cell survival in adulthood(D112) 4 weeks after BrdU injection at D85. Experiment 3 assessed DG cellproliferation in adult rats that had undergone restraint stress (D90B). — #not performed or assessed. BrdU, bromodeoxyuridine.
1122 BIOL PSYCHIATRY 2006;60:1121–1130 D.C. Lagace et al
www.sobp.org/journal

Hampshire), and dried overnight. Slides were coded to ensureobjectivity, and the code was not broken until after data collec-tion was complete. Pretreatment for both types of IHC was asfollows: antigen retrieval (.01 mol citric acid, pH 6.0, 95°C, 10min), membrane permeabilization (.1% trypsin in .1 mol Tris and.1% CaCl2, 10 min), and DNA denaturation (2NHCl in 1! PBS, 30min). Following pretreatment, sections for DAB IHC were blockedwith 3% normal goat serum (NGS, Vector Laboratories, Burlingame,California) and incubated with mouse-anti-BrdU primary antibody(1:100, Becton-Dickinson Laboratories, San Jose, California) over-night. The following day, sections were incubated with 1.5% NGSand goat-anti-mouse secondary antibody (1:200, Sigma Laborato-ries, St. Louis, Missouri) for 60 min, .3% hydrogen peroxide(Sigma Laboratories) for 30 min, and avidin biotin complex(Vector Laboratories) for 90 min. Staining of BrdU-labeled cellswas visualized using DAB (Pierce, Rockford, Illinois) for 30 min.Nuclear Fast Red (Vector Laboratories) was used as a counter-stain. For triple-labeling immunoflourescence, sections wereblocked with 3% normal donkey and goat sera (Vector Labora-tories) for 60 min before overnight incubation with rat-anti-BrdU(1:100, Accurate Chemical, Westbury, New York), mouse-anti-neuronal nuclei (NeuN, 1:50, Chemicon International, Temecula,CA) and rabbit-anti-glial fibrillary acidic protein (GFAP, 1:500,DAKO, Glostrup, Denmark). Sections were then incubated influorescent donkey-anti-rat, donkey-anti-rabbit, and goat-anti-mouse secondary antibodies (1:200, Jackson ImmunoResearch,West Grove, Pennsylvania) for 3 hours, followed by counter-staining with DAPI (1:5000, Roche, Basel, Switzerland). All slideswere dehydrated and coverslipped.
Experiment 1: Juvenile MPH and Adult HippocampalCell Proliferation
To assess whether juvenile MPH exposure alters the numberof DG progenitors in the adult hippocampus, BrdU-labeled cellswere counted in the DG of adolescent, early adult, and adult rats(D46, D77, D90A, D90B). Four regions of the DG (bregma –1.5 to–6.3 mm) were examined as previously described (Mandyam etal 2004): molecular layer (Mol), SGZ, outer granule cell layer(oGCL), and hilus (H). The habenula was also examined as anonneurogenic region. Counting of BrdU-labeled cells was donein bright field by an examiner blind to treatment at 400! with anOlympus BX-51 microscope (Olympus, Tokyo). The BrdU-la-beled cells counted within the four regions of the dentate gyrus(DG) and the habenula were irregular in shape, dark brown, andcommonly found in clusters (Cameron and McKay 2001; Eisch etal 2000). Because counting of cells was conducted on every ninthsection of the hippocampus bilaterally, the number of countedcells in each region was multiplied by nine to obtain an estimateof the total number of cells per region (Eisch et al 2000). Inaddition to estimating the total number of BrdU-labeled cells, thenumber of cells was also quantified across the longitudinal axisof the hippocampus. For this analysis by bregma, data arepresented as total number of cells in the SGZ per section at eachseptotemporal level analyzed (Paxinos and Watson 1997).
Experiment 2: Juvenile MPH Exposure and AdultHippocampal Neurogenesis
To determine whether juvenile MPH exposure alters hip-pocampal neurogenesis, tissue from rats sacrificed 4 weeks afterBrdU injection (D112) was examined for BrdU-labeled cellcounts within the four DG regions and the habenula, as well asacross the longitudinal axis of the hippocampus (see Experiment 1).The BrdU-labeled cells from D112 rats displayed characteristics of
mature granule cell neurons: large, round, individual (rather than inclusters), with dark or punctate BrdU staining reflecting divisionsince initial BrdU incorporation (Cameron and McKay 2001;Eisch et al 2000). In addition, BrdU-labeled cells were classifiedas neuronal or glial based on the colocalization of BrdU with themature neuronal protein NeuN or the asytrocyte protein GFAP.Colocalization of immunofluorescence was determined with aconfocal microscope (Zeiss Axiovert 200 and LSM510-META, CarlZeiss, Oberkochen, Germany; excitation wavelengths 488, 543,and 633 nm) at 630! using multitrack scanning and an opticalsection thickness of approximately .5 "m in the Z plane. Approxi-mately 32 $ 3 cells per rat (387 cells SGZ and oGCL) were selectedby a blind examiner to assess colocalization of BrdU with eitherNeuN or GFAP. To guard against false positives (Raff 2003),colocalization was verified by importing stacks of Z images into athree-dimensional reconstruction program, Volocity (Improvi-sion, Lexington, Massachusetts). Three-dimensional renderingswere rotated and colocalization was examined from all perspec-tives. For presentation, images were imported into Photoshop(Adobe Systems 7.0, Carlsbad, California), and adjustments weremade only via changes in gamma in the level function.
Experiment 3: Juvenile MPH Exposure and RestraintStress-Induced Alterations in Cell Proliferation
Rats from D90B received restraint stress on D90 to determinewhether a previously described MPH-induced increase in stressresponse (Bolanos et al 2003) corresponded to altered hip-pocampal cell proliferation. Restraint stress for 20 min wasperformed as previously described (Bolanos et al 2003). Briefly,tail blood samples were collected at initiation of restraint (t # 0)and near completion of restraint (t # 15 min). The rats werereturned to home cage after restraint. Blood samples werecollected in heparin-coated tubes and immediately centrifuged(1000 g for 15 min). Plasma was divided into aliquots and frozenat –20°C until assayed for corticosterone levels using the OCTEIAcorticosterone competitive enzyme immunoassay (EIA, ALPCODiagnostics, Windham, New Hampshire). Four hours after re-straint stress, rats were injected with BrdU and perfused transcar-dially 2 hours later for assessment of cell proliferation.
Statistical AnalysesThe data are reported as mean $ SEM for MPH versus SAL
rats. Statistical analyses were performed using a multiple variableanalysis of variance (ANOVA) followed by a Bonferroni post hoctest. For repeated measure variables (90 min of locomotionassessment, BrdU-labeled cell counts across bregma levels), arepeated-measures ANOVA was used to assess the effect 1) of therepeated measure (time or bregma), 2) group (SAL vs. MPH), and3) any interaction effects. Post hoc analyses were performedusing the Bonferroni comparison. To assess differences betweenthe number of BrdU-labeled cells colabeled with either NeuN orGFAP, a chi-square analysis was performed. All statistical analy-ses were performed using either SPSS (version 11.0.2) or Prismsoftware. Statistical significance was defined as p % .05.
Results
Juvenile MPH Decreases Locomotor Responseto Novelty in Adulthood
Locomotor activity in a novel environment was significantlydifferent between SAL and MPH rats [Figure 1A; F (1,40) # 4.3,p % .05], with the D112 MPH rats displaying less locomotioncompared with D112 SAL rats (p % .01). During the 90-min test,there was also a significant difference between MPH and SAL rats
D.C. Lagace et al BIOL PSYCHIATRY 2006;60:1121–1130 1123
www.sobp.org/journal

[F (1,9) # 8.1, p % .05], with MPH rats displaying less locomotoractivity at the beginning of the test (Figure 1B; 15, 20, 25, and35 min, p % .05). These results confirm previous work(Bolanos et al 2003) that juvenile MPH exposure significantlyreduces locomotor activity in a novel environment duringadulthood. Moreover, this finding demonstrates that juvenileMPH administration does not produce any significant alter-ation in locomotion in response to novel environment prior toD112.
Experiment 1: Juvenile MPH Does Not Affect NeuralProgenitor Cell Proliferation
Between MPH and SAL rats, there was no difference in thenumber of BrdU-labeled cells in the SGZ at any developmentalstage (Figure 2A). There was also no significant differencebetween SAL and MPH rats in the number of newly born cells inthe other examined regions (data not shown) or across thelongitudinal axis of the hippocampus (Figure 2B). Theseresults show that repeated juvenile MPH treatment does notalter hippocampal progenitor proliferation, as assessed byBrdU-labeled cell counts, in adolescence, early adulthood, oradulthood.
Independent of early-life exposure to MPH or SAL, there wasan age-dependent decrease in BrdU-labeled cells [Figure 2A;F (2,28) # 31.3, p % .001]. Similar to previous results (Bizon andGallagher 2003; Heine et al 2004a), we find approximately halfthe number of BrdU-labeled cells in adult rats compared withadolescent rats (D46 vs. D77 and D46 vs. D90A; p % .05).Moreover, across the longitudinal axis of the hippocampus there
was a significant difference in BrdU-labeled cells within the SGZin all three groups [Figure 2B; data from D46, F (16,144) # 19.4,p % .001; similar results for D77 or D90A, data not shown]. Therewere a greater number of cells located in the temporal sections,with the greatest number of SGZ BrdU-labeled cells at bregma–5.3 and –5.8. Although these results demonstrate a gradient inproliferation along the axis of the hippocampus with moreproliferation occurring in the temporal hippocampus, juvenileexposure to MPH did not result in altered proliferation in later lifeanywhere along the longitudinal axis of the hippocampus.
Experiment 2: Juvenile MPH Reduces the Survival of NewlyBorn Cells in Adulthood
Having established that neural progenitor proliferation is notaltered after juvenile exposure to MPH, we allowed a parallelgroup of rats injected with BrdU in adulthood (D85) to survivefor an additional 4 weeks. Although no gross morphologicdifferences were evident between BrdU-labeled cells in SAL andMPH rats (Figure 3A), there was a significant difference betweenthe number of BrdU-labeled cells between the regions examined[Figure 3B; F (4,50) # 66.72, p % .005], as well as a significantdifference between SAL and MPH rats [Figure 3B; F (1,50) #10.08, p % .01]. Post hoc analysis demonstrated MPH rats hadsignificantly fewer BrdU-labeled cells in the SGZ compared withSAL rats (p % .05), emphasizing the region-specificity of theeffects of MPH on survival of cells within the hippocampus. In
Figure 1. Juvenile methylphenidate (MPH) exposure reduced locomotoractivity in a novel environment when assessed during adulthood. (A) Therewas a significant difference between saline (SAL) and MPH rats, with MPHrats having significantly less locomotion compared with the SAL rats at day(D)112. Data presented represent the mean $ SEM photocell counts duringthe 90 min test. (B) Time course of locomotor behavior from D112 group.Movement counts over the course of the 90-min test revealed that MPH ratsexplored less than SAL rats, with a significant reduction in movement in 15,20, 25, and 35 min following placement into the chamber. * p % .05, ** p %.01.
Figure 2. Juvenile methylphenidate (MPH) administration did not affecthippocampal progenitor proliferation in the subgranular zone (SGZ) at anystage of development or alter the distribution of proliferation along theseptotemporal axis. (A) Proliferation, as assessed by bromodeoxyuridine(BrdU)-labeled cell counts in groups D46, 77, and 90A, did not differ be-tween MPH and saline (SAL) rats in the SGZ. Other regions of the DG and thehabenula were similarly unaffected by MPH treatment (data not shown).Subgranular zone (SGZ): Septotemporal analysis of SGZ proliferation re-vealed that sections in the temporal hippocampus had more BrdU-labeledcells than the septal hippocampus with no significant difference betweenMPH and SAL rats. Data shown represented mean BrdU-labeled cell countsper section for D46 SAL and MPH rats. Similar patterns of distribution bybregma were observed in D77 and D90A rats (data not shown).
1124 BIOL PSYCHIATRY 2006;60:1121–1130 D.C. Lagace et al
www.sobp.org/journal
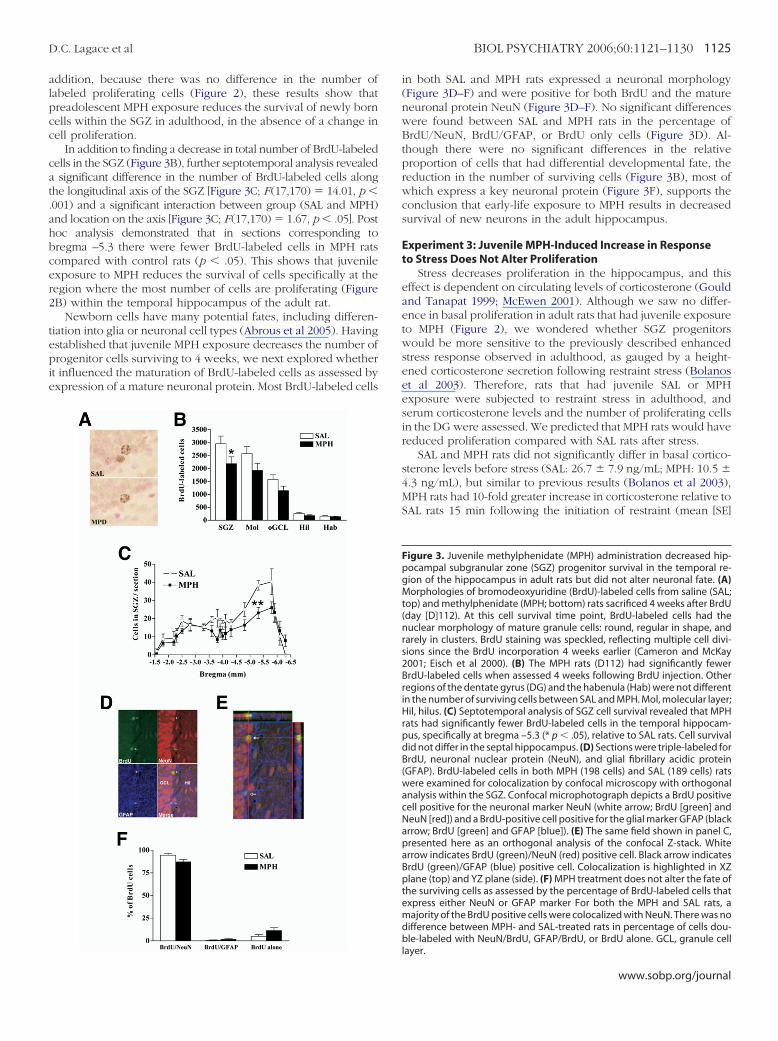
addition, because there was no difference in the number oflabeled proliferating cells (Figure 2), these results show thatpreadolescent MPH exposure reduces the survival of newly borncells within the SGZ in adulthood, in the absence of a change incell proliferation.
In addition to finding a decrease in total number of BrdU-labeledcells in the SGZ (Figure 3B), further septotemporal analysis revealeda significant difference in the number of BrdU-labeled cells alongthe longitudinal axis of the SGZ [Figure 3C; F(17,170) # 14.01, p %.001) and a significant interaction between group (SAL and MPH)and location on the axis [Figure 3C; F(17,170) # 1.67, p % .05]. Posthoc analysis demonstrated that in sections corresponding tobregma –5.3 there were fewer BrdU-labeled cells in MPH ratscompared with control rats (p % .05). This shows that juvenileexposure to MPH reduces the survival of cells specifically at theregion where the most number of cells are proliferating (Figure2B) within the temporal hippocampus of the adult rat.
Newborn cells have many potential fates, including differen-tiation into glia or neuronal cell types (Abrous et al 2005). Havingestablished that juvenile MPH exposure decreases the number ofprogenitor cells surviving to 4 weeks, we next explored whetherit influenced the maturation of BrdU-labeled cells as assessed byexpression of a mature neuronal protein. Most BrdU-labeled cells
in both SAL and MPH rats expressed a neuronal morphology(Figure 3D–F) and were positive for both BrdU and the matureneuronal protein NeuN (Figure 3D–F). No significant differenceswere found between SAL and MPH rats in the percentage ofBrdU/NeuN, BrdU/GFAP, or BrdU only cells (Figure 3D). Al-though there were no significant differences in the relativeproportion of cells that had differential developmental fate, thereduction in the number of surviving cells (Figure 3B), most ofwhich express a key neuronal protein (Figure 3F), supports theconclusion that early-life exposure to MPH results in decreasedsurvival of new neurons in the adult hippocampus.
Experiment 3: Juvenile MPH-Induced Increase in Responseto Stress Does Not Alter Proliferation
Stress decreases proliferation in the hippocampus, and thiseffect is dependent on circulating levels of corticosterone (Gouldand Tanapat 1999; McEwen 2001). Although we saw no differ-ence in basal proliferation in adult rats that had juvenile exposureto MPH (Figure 2), we wondered whether SGZ progenitorswould be more sensitive to the previously described enhancedstress response observed in adulthood, as gauged by a height-ened corticosterone secretion following restraint stress (Bolanoset al 2003). Therefore, rats that had juvenile SAL or MPHexposure were subjected to restraint stress in adulthood, andserum corticosterone levels and the number of proliferating cellsin the DG were assessed. We predicted that MPH rats would havereduced proliferation compared with SAL rats after stress.
SAL and MPH rats did not significantly differ in basal cortico-sterone levels before stress (SAL: 26.7 $ 7.9 ng/mL; MPH: 10.5 $4.3 ng/mL), but similar to previous results (Bolanos et al 2003),MPH rats had 10-fold greater increase in corticosterone relative toSAL rats 15 min following the initiation of restraint (mean [SE]
Figure 3. Juvenile methylphenidate (MPH) administration decreased hip-pocampal subgranular zone (SGZ) progenitor survival in the temporal re-gion of the hippocampus in adult rats but did not alter neuronal fate. (A)Morphologies of bromodeoxyuridine (BrdU)-labeled cells from saline (SAL;top) and methylphenidate (MPH; bottom) rats sacrificed 4 weeks after BrdU(day [D]112). At this cell survival time point, BrdU-labeled cells had thenuclear morphology of mature granule cells: round, regular in shape, andrarely in clusters. BrdU staining was speckled, reflecting multiple cell divi-sions since the BrdU incorporation 4 weeks earlier (Cameron and McKay2001; Eisch et al 2000). (B) The MPH rats (D112) had significantly fewerBrdU-labeled cells when assessed 4 weeks following BrdU injection. Otherregions of the dentate gyrus (DG) and the habenula (Hab) were not differentin the number of surviving cells between SAL and MPH. Mol, molecular layer;Hil, hilus. (C) Septotemporal analysis of SGZ cell survival revealed that MPHrats had significantly fewer BrdU-labeled cells in the temporal hippocam-pus, specifically at bregma –5.3 (* p % .05), relative to SAL rats. Cell survivaldid not differ in the septal hippocampus. (D) Sections were triple-labeled forBrdU, neuronal nuclear protein (NeuN), and glial fibrillary acidic protein(GFAP). BrdU-labeled cells in both MPH (198 cells) and SAL (189 cells) ratswere examined for colocalization by confocal microscopy with orthogonalanalysis within the SGZ. Confocal microphotograph depicts a BrdU positivecell positive for the neuronal marker NeuN (white arrow; BrdU [green] andNeuN [red]) and a BrdU-positive cell positive for the glial marker GFAP (blackarrow; BrdU [green] and GFAP [blue]). (E) The same field shown in panel C,presented here as an orthogonal analysis of the confocal Z-stack. Whitearrow indicates BrdU (green)/NeuN (red) positive cell. Black arrow indicatesBrdU (green)/GFAP (blue) positive cell. Colocalization is highlighted in XZplane (top) and YZ plane (side). (F) MPH treatment does not alter the fate ofthe surviving cells as assessed by the percentage of BrdU-labeled cells thatexpress either NeuN or GFAP marker For both the MPH and SAL rats, amajority of the BrdU positive cells were colocalized with NeuN. There was nodifference between MPH- and SAL-treated rats in percentage of cells dou-ble-labeled with NeuN/BrdU, GFAP/BrdU, or BrdU alone. GCL, granule celllayer.
D.C. Lagace et al BIOL PSYCHIATRY 2006;60:1121–1130 1125
www.sobp.org/journal

fold increase; SAL # 5.7(1.4) vs. MPH # 14.71(2.7); t9 # –3.03,p % .05). Consistent with previous research, stress reduced thenumber of BrdU-labeled cells (Figure 4A; D90A vs. D90B,F (1,105) # 30.9, p % .001) in a region-specific manner (Figure4A; F (4,105) # 9.2, p % .001). Regardless of juvenile exposure toSAL or MPH, stressed rats had fewer BrdU-labeled cells in theSGZ, Mol, and Hil in adulthood (ps % .001) relative to non-stressed rats (D90A). These findings show that restraint stress inrats is associated with regional decreases in proliferation withinthe hippocampus. Opposite to our hypothesis. However, therewas no significant difference in the number of proliferating cellsbetween SAL and MPH restrained rats in any of the regionscounted (Figure 4A). When compared with rats that were notsubjected to restraint stress (D90A), there was also no significantinteraction between stress (D90A vs. D90B) and group (SAL vs.MPH; Figure 4A). These data show that early-life exposure toMPH is associated with heightened corticosterone response inresponse to restraint stress in the absence of a greater decrease incell proliferation within the hippocampus.
Because MPH exposure decreased the survival of BrdU-labeled cells to a greater degree in the temporal hippocampus(Figure 3B), we also analyzed proliferation along the longitudinalaxis of the hippocampus between stressed MPH and SAL rats.Similar to our previous analysis there was a significant septotem-poral difference [Figure 4B; F (15,135) # 7.4, p % .001], with thelargest number of cells at bregma –5.3. However, there was no
significant difference in proliferation between MPH and SAL ratsat any bregma level.
Discussion
Repeated exposure to MPH in early life affects a variety ofbehavioral and physiological responses in adulthood (Achat-Mendes et al 2003; Adriani et al 2005; Andersen et al 2002; Bolanoset al 2003; Carlezon et al 2003; Mague et al 2005). Given the impactof psychostimulants on brain reward circuitry (Nestler 2005; Nestlerand Malenka 2004; Robinson 2004) and the decreased response torewarding stimuli in adulthood seen after early MPH exposure(Carlezon et al 2003; Mague et al 2005), most studies focus on thestriatum, nucleus accumbens, and other brain regions related to thereward pathway in exploring the consequences of adolescent MPHexposure (Adriani et al 2005; Andersen et al 2002; Brandon et al2003; Carlezon et al 2003; Chase et al 2005a, 2005b; Shen andChoong 2005). Decreased adult hippocampal neurogenesis nowjoins the growing list of long-term effects of early MPH exposure,and the findings presented here emphasize that MPH-inducedneuroadaptations are not limited to the primary sites of reward inthe adult brain.
Of the thousands of new cells generated in the rat DG eachday (Cameron and McKay 2001), half of SGZ progenitors may diebetween 6 and 22 days after labeling with BrdU (Dayer et al2003). The newborn granule neurons that survive this period ofcell death integrate into hippocampal circuitry (Gould et al 1999).Here we find that juvenile MPH exposure leads to decreasedsurvival of new neurons in the adult hippocampus, withoutinfluencing the number of proliferating cells. We speculate thatthis distinction points to MPH decreasing survival by increasingapoptosis of maturing, not proliferating, neurons. In support ofthis theory, decreases in survival in the absence of changes inproliferation have been associated with increases in apoptosis(Ambrogini et al 2005; Kuhn et al 2005; Sun et al 2004). Work isongoing in our laboratory to address the hypothesis that early-lifeexposure to juvenile MPH increases apoptosis of nascent neu-rons leading to reduced survival in MPH rats. Alternative possi-bilities exist to explain the reduced neuronal survival in MPHrats, such as the effect of decreased reactivity to a novelenvironment or the stress of the locomotor testing. The firstpossibility is unlikely because reduced reactivity does not altersurvival and indeed is correlated with increased, not decreased,proliferation (Lemaire et al 1999). The second possibility isparticularly important to consider because juvenile MPH expo-sure enhances serum corticosterone levels in response to stressin adulthood (Bolanos et al 2003; present data), and stressgenerally decreases neurogenesis (Mirescu and Gould 2006).Unlike proliferation, however, decreases in survival appear torequire chronic exposure to a changed environment (Ambroginiet al 2005; Lee et al 2006; Lichtenwalner et al 2006); this is incontrast to the single exposure to a novel environment usedhere. In addition, exposure to chronic unpredictable stress,which decreases proliferation, does not alter the number ofsurviving hippocampal cells (Heine et al 2004b). Taken togetherwith the fact that restraint stress did not influence proliferationdifferentially between the MPH and control animals (Figure 4),these data suggest that the relatively mild stressor of locomotiontesting for 90 min would not alter survival in itself. This evidencetherefore suggests the reduced survival in MPH rats is not due toa differential response to the possibly stressful novel environ-ment associated with locomotor testing.
Relatively little is known about factors within the SGZ microen-
Figure 4. Restraint stress reduces proliferation, yet juvenile methylpheni-date (MPH) administration does not alter proliferation in stressed adult rats.(A) Compared with unstressed rats (90A), stressed rats (90B) had a signifi-cant reduction in the number of proliferating cells in the subgranular layer(SGZ), molecular layer (Mol), and hilus (Hil; ps % .001) with no difference inthe outer granule cell layer (oGCL) or habenula (Hab). Although stressedMPH rats (90B MPH) had 10-fold greater stress-induced increase in serumcortisoterone (see Results, Experiment 3), stressed MPH rats did not showthe expected greater decrease in proliferation compared to SAL rats. (B)Septotemporal analysis of SGZ proliferation revealed that sections in thetemporal hippocampus had more bromodeoxyuridine-labeled cells in theseptal hippocampus with no significant difference between MPH and SALrats. Data shown are mean number of BrdU-labeled cell counts per sectionfor D90B SAL and MPH rats.
1126 BIOL PSYCHIATRY 2006;60:1121–1130 D.C. Lagace et al
www.sobp.org/journal

vironment that regulate cell survival independent of proliferation(Abrous et al 2005; Ming and Song 2005). Several recent factors havebeen identified, however, that preferentially influence survivalrelative to proliferation (Ambrogini et al 2005; Kuhn et al 2005; Leeet al 2006; Lichtenwalner et al 2006; Sun et al 2004). The datapresented here add juvenile exposure to MPH to the growing list offactors that alter survival independent of proliferation. BecauseMPH exposure selectively diminished the number of surviving BrdUcells in the temporal hippocampus, additional studies could buildon these findings to examine how the septal versus temporal SGZmicroenvironment is altered after juvenile MPH exposure to helpelucidate possible mechanisms for the effect of MPH on survival.This work also has tremendous potential to shed light on howexperiences, such as maternal care (Bredy et al 2003) and hip-pocampal-dependent learning (Gould et al 1999), similarly influ-ence the survival of adult generated neurons independent ofproliferation.
Given the diverse anatomic connections and functions of theseptal versus temporal hippocampus (Amaral 1993; Bannermanet al 2004), it is interesting that less than a dozen studies haveevaluated adult neurogenesis along the longitudinal axis of thehippocampus in the rat (Ambrogini et al 2000; Banasr et al, inpress; Cameron et al 1995; Kim et al 2005; Rao and Shetty 2004),mouse (Ferland et al 2002; Hayes and Nowakowski 2002), orgerbil (Dawirs et al 1998; Hildebrandt et al 1999; Teuchert-Noodtet al 2000). The data presented here are the first to showproliferation and survival across the entire longitudinal axis aswell as regulation across this axis, and it is notable that thedecreased survival shown here occurs specifically in the tempo-ral hippocampus. This is in keeping with recent papers showingthat the temporal hippocampus is the preferential site of regula-tion of proliferation by restraint stress (Kim et al 2005) andagomelatine, a newer antidepressant drug (Banasr et al 2006).Taken together with our data demonstrating regulation of sur-vival, these data highlight an underappreciated heterogeneity ofthe hippocampus in regard to regulation of neurogenesis. Thedifferential response of the septal versus temporal hippocampusmay be related to the distinct anatomic connections and func-tions of these regions. The septal, or rostral, hippocampus isconnected to sensory areas and is implicated in spatial memory,whereas the temporal, or caudal, hippocampus is connected tolimbic-related areas and is implicated in anxiety- and depression-related behaviors (Bannerman et al 2004). Future studies mighttherefore examine whether the reduced number of survivingcells in the temporal hippocampus seen after juvenile MPHexposure correlates with anxiety- and depressive-like behaviorsin the adult. Moreover, as studies identify the molecular hetero-geneity along the axis of the rat dentate gyrus, as has recentlybeen done for the mouse hippocampus (Lein et al 2004, 2005;Leonardo et al 2006), proposed mechanisms for the region-specific effects observed in this study could be tested. Althoughcertainly out of the scope of this study, such studies promise tobe fruitful in exploring how regional differences in neurogenesiscontribute to distinct hippocampal processes.
We found no influence of juvenile MPH on the number ofproliferating cells at any developmental time point; however, oneaspect of our work suggests that the proliferating cells are perhapsnot completely unaffected by juvenile MPH exposure. The reactivityof the HPA axis to restraint stress was greatly increased in MPH rats,replicating previous work (Bolanos et al 2003), and stress decreasedthe number of proliferating cells (Figure 4A), in agreement withprevious studies (Bain et al 2004). Despite MPH rats having in-creased corticosterone response to stress, MPH rats did not have the
expected greater decrease in number of proliferating cells. Thiswas somewhat surprising because high levels of glucocorticoids,whether resulting from administered hormones (Cameron andGould 1994; Gould et al 1992) or experimental stressors (Czeh etal 2002; Malberg and Duman 2003; Tanapat et al 2001), decreasethe number of proliferating cells within the hippocampus. Al-though the relationship between neurogenesis and corticoste-rone is complex (Brunson et al 2005; Pham et al 2003), oneinterpretation of our data is that proliferating cells in MPH ratsmay have a blunted response to corticosterone. Initial studies inthe rat found no corticoid receptors on proliferating cells (Cam-eron et al 1993), but recent technical advances in receptordetection and progenitor subtype analysis show that a subpopu-lation of progenitors in the mouse have corticoid receptors(Garcia et al 2004). If future studies do find corticoid receptors onrat progenitor cells, additional work is warranted to examinewhether corticoid receptor distribution or function is altered inthe SGZ progenitors of rats exposed to juvenile MPH. Importantfor our results, corticosterone is also a recognized player in cellsurvival in the adult mouse SGZ. Neurons that express a gluo-cocorticoid receptor during key phases of maturation appear tobe targeted for cell death (Garcia et al 2004). Therefore, studiesare needed to examine whether altered gluococorticoid receptorexpression on maturing progenitor cells may contribute to thediminished survival reported here in MPH rats.
Adult-generated SGZ cells become DG neurons and functionallyintegrate into hippocampal circuitry (Hastings and Gould 2003;Markakis et al 2004; van Praag et al 2002), raising speculations thatthese neurons play an important role in hippocampal-dependentbehaviors. Correlative studies support this because the addition ofnew neurons to the adult DG has positive effects on learning andmemory, whereas inhibition of new neuron birth contributes tomemory loss or lack of clearance of old memories (Abrous et al2005). Others have shown that early-life experiences such asprenatal stress (Lemaire et al 2000) and perinatal maternal depriva-tion (Bredy et al 2003) attenuate proliferation and neurogenesis andlead to impairments in hippocampal-related learning and memorytasks in adulthood (Lemaire et al 2000; Liu et al 2000). Because wedemonstrate a reduction in the survival of new neurons in MPH rats,it would be of interest to identify whether a decrease in survivaloccurs throughout the developmental period and whether thiscoincides with impaired learning of hippocampal-dependent tasks(Shors et al 2002). If adult rats treated with MPH earlier in life havedeficits in learning and memory in adulthood, this finding wouldsupport that a reduction in survival of newly born cells may havefunctional effects.
Direct extrapolations of our findings to the human condition areunwarranted given the lack of animal models to assess appropri-ately psychostimulant exposure and the difficultly in achievingsimilar pharmacokinetic profiles for psychotropic drugs in rodents(Andersen 2005; Kuczenski and Segal 2005); however, discussion ofthese results would be incomplete without mention of how the dataintegrate with three established avenues of research into the long-term consequences of MPH treatment. First, our findings of juvenileexposure to MPH leading to decreased adult hippocampal neuro-genesis are in parallel with ongoing studies examining the impact oflong-term MPH exposure on human hippocampal volume. Whileyouth treated with MPH appear to not have diminished hippocam-pal volume, longitudinal follow-up studies are addressing thehypothesis that altered hippocampal volume may not appear untiladulthood (Judith Rapoport, personal communication, 2005). Sec-ond, because neurogenesis is presumed to contribute to hippocam-pal function, our findings encourage the assessment of juvenile
D.C. Lagace et al BIOL PSYCHIATRY 2006;60:1121–1130 1127
www.sobp.org/journal

MPH exposure in adult rodents and humans on outcomes related tohippocampal function, such as learning and memory. Third, studiesimplicate adult hippocampal neurogenesis in depression (Duman2004; Eisch 2002; Henn and Vollmayr 2004), and juvenile MPHproduces a depressive-like phenotype in adult rodents (Bolanos etal 2003; Carlezon et al 2003) Therefore, our finding of decreasedhippocampal neurogenesis with no obvious impact on proliferationraises the hypothesis that depression is marked by diminishedsurvival of adult-generated neurons independent of an effect onproliferation. Indeed, this hypothesis may explain why depressive-like behavior in animal models does not always correlate withdecreased proliferation of hippocampal progenitors (Malberg andDuman 2003; Vollmayr et al 2003). Studies that assess neurogenesisin animal models of depression are encouraged to include measuresof survival in addition to proliferation to address this hypothesisfurther.
In sum, when considered in light of previous work, this studyprovides additional evidence that early-life exposure to MPH canhave complex effects that endure later into life. These findingsurge further exploration of hippocampal neuroadaptations afterjuvenile MPH exposure and emphasize the need for continuedappropriate use of MPH in youth.
This work was supported by a NARSAD Young InvestigatorAward (AJE), an RO1 (AJE), and a National Research ServiceAward (CAB) from the National Institute on Drug Abuse; a post-doctoral research fellowship from the Canadian Institute of HealthResearch (DCL); and a summer fellowship from the Stanley Foun-dation (JKY). The authors thank Rebekah Norris for outstandingtechnical assistance and Michele Noonan and Stephanie Fischer forcritical reading of the manuscript.
Abikoff H, Hechtman L, Klein RG, Weiss G, Fleiss K, Etcovitch J, et al (2004):Symptomatic improvement in children with ADHD treated with long-term methylphenidate and multimodal psychosocial treatment. J AmAcad Child Adolesc Psychiatry 43:802– 811.
Abrous DN, Koehl M, Le Moal M (2005): Adult neurogenesis: From precursorsto network and physiology. Physiol Rev 85:523–569.
Achat-Mendes C, Anderson KL, Itzhak Y (2003): Methylphenidate and MDMAadolescent exposure in mice: Long-lasting consequences on cocaine-induced reward and psychomotor stimulation in adulthood. Neurophar-macology 45:106 –115.
Adriani W, Leo D, Greco D, Rea M, di Porzio U, Laviola G, Perrone-Capano C(2005): Methylphenidate administration to adolescent rats determinesplastic changes on reward-related behavior and striatal gene expres-sion. Neuropsychopharmacology 1–11 [Epub ahead of print; doi:10.1038/sj.npp.1300962].
Amaral DG (1993): Emerging principles of intrinsic hippocampal organiza-tion. Curr Opin Neurobiol 3:225–229.
Ambrogini P, Cuppini R, Cuppini C, Ciaroni S, Cecchini T, Ferri P, et al (2000):Spatial learning affects immature granule cell survival in adult rat den-tate gyrus. Neurosci Lett 286:21–24.
Ambrogini P, Cuppini R, Ferri P, Mancini C, Ciaroni S, Voci A, et al (2005):Thyroid hormones affect neurogenesis in the dentate gyrus of adult rat.Neuroendocrinology 81:244 –253.
Andersen SL (2005): Stimulants and the developing brain. Trends PharmacolSci 26:237–243.
Andersen SL, Arvanitogiannis A, Pliakas AM, LeBlanc C, Carlezon WA Jr(2002): Altered responsiveness to cocaine in rats exposed to methyl-phenidate during development. Nat Neurosci 5:13–14.
Andersen SL, Navalta CP (2004): Altering the course of neurodevelopment: Aframework for understanding the enduring effects of psychotropicdrugs. Int J Dev Neurosci 22:423– 440.
Angold A, Erkanli A, Egger HL, Costello EJ (2000): Stimulant treatment forchildren: A community perspective. J Am Acad Child Adolesc Psychiatry39:975–984; discussion 984 –994.
Bain MJ, Dwyer SM, Rusak B (2004): Restraint stress affects hippocampal cellproliferation differently in rats and mice. Neurosci Lett 368:7–10.
Banasr M, Soumier A, Hery M, Mocaer E, Daszuta A (in press): Agomelatine, anew antidepressant, induces regional changes in hippocampal neuro-genesis. Biol Psychiatry.
Bannerman DM, Rawlins JN, McHugh SB, Deacon RM, Yee BK, Bast T, et al(2004): Regional dissociations within the hippocampus—memory andanxiety. Neurosci Biobehav Rev 28:273–283.
Bizon JL, Gallagher M (2003): Production of new cells in the rat dentategyrus over the lifespan: Relation to cognitive decline. Eur J Neurosci18:215–219.
Bolanos CA, Barrot M, Berton O, Wallace-Black D, Nestler EJ (2003): Methyl-phenidate treatment during pre- and periadolescence alters behavioralresponses to emotional stimuli at adulthood. Biol Psychiatry 54:1317–1329.
Brandon CL, Marinelli M, Baker LK, White FJ (2001): Enhanced reactivity andvulnerability to cocaine following methylphenidate treatment in adoles-cent rats. Neuropsychopharmacology 25:651– 661.
Brandon CL, Marinelli M, White FJ (2003): Adolescent exposure to methyl-phenidate alters the activity of rat midbrain dopamine neurons. BiolPsychiatry 54:1338 –1344.
Bredy TW, Grant RJ, Champagne DL, Meaney MJ (2003): Maternal care influ-ences neuronal survival in the hippocampus of the rat. Eur J Neurosci18:2903–2909.
Brunson KL, Baram TZ, Bender RA (2005): Hippocampal neurogenesis is notenhanced by lifelong reduction of glucocorticoid levels. Hippocampus15:491–501.
Cameron HA, Gould E (1994): Adult neurogenesis is regulated by adrenalsteroids in the dentate gyrus. Neuroscience 61:203–209.
Cameron HA, McEwen BS, Gould E (1995): Regulation of adult neurogenesisby excitatory input and NMDA receptor activation in the dentate gyrus.J Neurosci 15:4687– 4692.
Cameron HA, McKay RD (2001): Adult neurogenesis produces a large pool ofnew granule cells in the dentate gyrus. J Comp Neurol 435:406 – 417.
Cameron HA, Woolley CS, Gould E (1993): Adrenal steroid receptor immu-noreactivity in cells born in the adult rat dentate gyrus. Brain Res 611:342–346.
Carlezon WA Jr, Duman RS, Nestler EJ (2005): The many faces of CREB. TrendsNeurosci 28:436 – 445.
Carlezon WA Jr, Konradi C (2004): Understanding the neurobiological con-sequences of early exposure to psychotropic drugs: Linking behaviorwith molecules. Neuropharmacology 47(suppl 1):47– 60.
Carlezon WA Jr, Mague SD, Andersen SL (2003): Enduring behavioral effectsof early exposure to methylphenidate in rats. Biol Psychiatry 54:1330 –1337.
Charara A, Grace AA (2003): Dopamine receptor subtypes selectivelymodulate excitatory afferents from the hippocampus and amygdalato rat nucleus accumbens neurons. Neuropsychopharmacology 28:1412–1421.
Chase TD, Carrey N, Brown RE, Wilkinson M (2005a): Methylphenidate differ-entially regulates c-fos and fosB expression in the developing rat stria-tum. Brain Res Dev Brain Res 157:181–191.
Chase TD, Carrey N, Brown RE, Wilkinson M (2005b): Methylphenidate regu-lates c-fos and fosB expression in multiple regions of the immature ratbrain. Brain Res Dev Brain Res 156:1–12.
Czeh B, Michaelis T, Watanabe T, Frahm J, de Biurrun G, van Kampen M, et al(2001): Stress-induced changes in cerebral metabolites, hippocampalvolume, and cell proliferation are prevented by antidepressant treat-ment with tianeptine. Proc Natl Acad Sci U S A 98:12796 –12801.
Czeh B, Welt T, Fischer AK, Erhardt A, Schmitt W, Muller MB, et al (2002):Chronic psychosocial stress and concomitant repetitive transcranialmagnetic stimulation: Effects on stress hormone levels and adult hip-pocampal neurogenesis. Biol Psychiatry 52:1057–1065.
Dawirs RR, Hildebrandt K, Teuchert-Noodt G (1998): Adult treatment withhaloperidol increases dentate granule cell proliferation in the gerbilhippocampus. J Neural Transm 105:317–327.
Dayer AG, Ford AA, Cleaver KM, Yassaee M, Cameron HA (2003): Short-termand long-term survival of new neurons in the rat dentate gyrus. J CompNeurol 460:563–572.
Duman RS (2004): Depression: A case of neuronal life and death? Biol Psychi-atry 56:140 –145.
Duman RS, Heninger GR, Nestler EJ (1997): A molecular and cellular theory ofdepression. Arch Gen Psychiatry 54:597– 606.
Eisch AJ (2002): Adult neurogenesis: Implications for psychiatry. Prog BrainRes 138:315–342.
1128 BIOL PSYCHIATRY 2006;60:1121–1130 D.C. Lagace et al
www.sobp.org/journal

Eisch AJ, Barrot M, Schad CA, Self DW, Nestler EJ (2000): Opiates inhibitneurogenesis in the adult rat hippocampus. Proc Natl Acad Sci U S A97:7579 –7584.
Eisch AJ, Harburg GC (2006): Opiates, psychostimulants, and adult hip-pocampal neurogenesis: Insights for addiction and stem cell biology.Hippocampus 16:271–286.
Emerich DF, Walsh TJ (1990): Hyperactivity following intradentate injectionof colchicine: A role for dopamine systems in the nucleus accumbens.Pharmacol Biochem Behav 37:149 –154.
Federici M, Geracitano R, Bernardi G, Mercuri NB (2005): Actions of methyl-phenidate on dopaminergic neurons of the ventral midbrain. Biol Psychi-atry 57:361–365.
Ferland RJ, Gross RA, Applegate CD (2002): Differences in hippocampalmitotic activity within the dorsal and ventral hippocampus followingflurothyl seizures in mice. Neurosci Lett 332:131–135.
Fogelman Y, Vinker S, Guy N, Kahan E (2003): Prevalence of and change inthe prescription of methylphenidate in Israel over a 2-year period. CNSDrugs 17:915–919.
Frances C, Hoizey G, Millart H, Trenque T (2002): Utilization of methylpheni-date (Ritalin) in France. Therapie 57:189 –193.
Garcia A, Steiner B, Kronenberg G, Bick-Sander A, Kempermann G (2004):Age-dependent expression of glucocorticoid- and mineralocorticoid re-ceptors on neural precursor cell populations in the adult murine hip-pocampus. Aging Cell 3:363–371.
Gould E, Beylin A, Tanapat P, Reeves A, Shors TJ (1999): Learning enhancesadult neurogenesis in the hippocampal formation. Nat Neurosci 2:260 –265.
Gould E, Cameron HA, Daniels DC, Woolley CS, McEwen BS (1992): Adrenalhormones suppress cell division in the adult rat dentate gyrus. J Neurosci12:3642–3650.
Gould E, Tanapat P (1999): Stress and hippocampal neurogenesis. Biol Psy-chiatry 46:1472–1479.
Hastings NB, Gould E (2003): Neurons inhibit neurogenesis. Nat Med 9:264 –266.
Hayes NL, Nowakowski RS (2002): Dynamics of cell proliferation in the adultdentate gyrus of two inbred strains of mice. Brain Res Dev Brain Res134:77– 85.
Heine VM, Maslam S, Joels M, Lucassen PJ (2004a): Prominent decline ofnewborn cell proliferation, differentiation, and apoptosis in the agingdentate gyrus, in absence of an age-related hypothalamus-pituitary-adrenal axis activation. Neurobiol Aging 25:361–375.
Heine VM, Maslam S, Zareno J, Joels M, Lucassen PJ (2004b): Suppressedproliferation and apoptotic changes in the rat dentate gyrus after acuteand chronic stress are reversible. Eur J Neurosci 19:131–144.
Henn FA, Vollmayr B (2004): Neurogenesis and depression: etiology or epi-phenomenon? Biol Psychiatry 56:146 –150.
Hildebrandt K, Teuchert-Noodt G, Dawirs RR (1999): A single neonatal doseof methamphetamine suppresses dentate granule cell proliferation inadult gerbils which is restored to control values by acute doses of halo-peridol. J Neural Transm 106:549 –558.
Hyman SE (2003): Methylphenidate-induced plasticity: What should we belooking for? Biol Psychiatry 54:1310 –1311.
Jensen PS, Hinshaw SP, Swanson JM, Greenhill LL, Conners CK, Arnold LE, etal (2001): Findings from the NIMH Multimodal Treatment Study of ADHD(MTA): Implications and applications for primary care providers. J DevBehav Pediatr 22:60 –73.
Karten YJ, Olariu A, Cameron HA (2005): Stress in early life inhibits neurogen-esis in adulthood. Trends Neurosci 28:171–172.
Kempermann G, Jessberger S, Steiner B, Kronenberg G (2004): Milestones ofneuronal development in the adult hippocampus. Trends Neurosci 27:447– 452.
Kim SJ, Lee KJ, Shin YC, Choi SH, Do E, Kim S, et al (2005): Stress-induceddecrease of granule cell proliferation in adult rat hippocampus: Assess-ment of granule cell proliferation using high doses of bromodeoxyuri-dine before and after restraint stress. Mol Cells 19:74 – 80.
Kuczenski R, Segal DS (2002): Exposure of adolescent rats to oral methyl-phenidate: Preferential effects on extracellular norepinephrine and ab-sence of sensitization and cross-sensitization to methamphetamine.J Neurosci 22:7264 –7271.
Kuczenski R, Segal DS (2005): Stimulant actions in rodents: implications forattention-deficit/hyperactivity disorder treatment and potential sub-stance abuse. Biol Psychiatry 57:1391–1396.
Kuhn HG, Biebl M, Wilhelm D, Li M, Friedlander RM, Winkler J (2005): In-creased generation of granule cells in adult Bcl-2-overexpressing mice: Arole for cell death during continued hippocampal neurogenesis. EurJ Neurosci 22:1907–1915.
Lee KJ, Kim SJ, Kim SW, Choi SH, Shin YC, Park SH, et al (2006): Chronic mildstress decreases survival, but not proliferation, of new-born cells in adultrat hippocampus. Exp Mol Med 38:44 –54.
Lein ES, Callaway EM, Albright TD, Gage FH (2005): Redefining the bound-aries of the hippocampal CA2 subfield in the mouse using gene expres-sion and 3-dimensional reconstruction. J Comp Neurol 485:1–10.
Lein ES, Zhao X, Gage FH (2004): Defining a molecular atlas of the hippocam-pus using DNA microarrays and high-throughput in situ hybridization.J Neurosci 24:3879 –3889.
Lemaire V, Aurousseau C, Le Moal M, Abrous DN (1999): Behavioural trait ofreactivity to novelty is related to hippocampal neurogenesis. Eur J Neu-rosci 11:4006 – 4014.
Lemaire V, Koehl M, Le Moal M, Abrous DN (2000): Prenatal stress produceslearning deficits associated with an inhibition of neurogenesis in thehippocampus. Proc Natl Acad Sci U S A 97:11032–11037.
Leonardo ED, Richardson-Jones JW, Sibille E, Kottman A, Hen R (2006):Molecular heterogeneity along the dorsal-ventral axis of the murinehippocampal CA1 field: A microarray analysis of gene expression. Neu-roscience 137:177–186.
Lichtenwalner RJ, Forbes ME, Sonntag WE, Riddle DR (2006): Adult-onsetdeficiency in growth hormone and insulin-like growth factor-I decreasessurvival of dentate granule neurons: Insights into the regulation of adulthippocampal neurogenesis. J Neurosci Res 83:199 –210.
Liu D, Diorio J, Day JC, Francis DD, Meaney MJ (2000): Maternal care, hip-pocampal synaptogenesis and cognitive development in rats. Nat Neu-rosci 3:799 – 806.
Mague SD, Andersen SL, Carlezon WA, Jr. (2005): Early developmental expo-sure to methylphenidate reduces cocaine-induced potentiation of brainstimulation reward in rats. Biol Psychiatry 57:120 –125.
Malberg JE, Duman RS (2003): Cell proliferation in adult hippocampus isdecreased by inescapable stress: Reversal by fluoxetine treatment. Neu-ropsychopharmacology 28:1562–1571.
Malberg JE, Eisch AJ, Nestler EJ, Duman RS (2000): Chronic antidepressanttreatment increases neurogenesis in adult rat hippocampus. J Neurosci20:9104 –9110.
Mandyam CD, Norris RD, Eisch AJ (2004): Chronic morphine induces prema-ture mitosis of proliferating cells in the adult mouse subgranular zone.J Neurosci Res 76:783–794.
Markakis EA, Palmer TD, Randolph-Moore L, Rakic P, Gage FH (2004): Novelneuronal phenotypes from neural progenitor cells. J Neurosci 24:2886–2897.
McEwen BS (2001): Plasticity of the hippocampus: Adaptation to chronicstress and allostatic load. Ann N Y Acad Sci 933:265–277.
Miller AR, Lalonde CE, McGrail KM, Armstrong RW (2001): Prescription ofmethylphenidate to children and youth, 1990 –1996. Cmaj 165:1489 –1494.
Ming GL, Song H (2005): Adult neurogenesis in the mammalian centralnervous system. Annu Rev Neurosci 28:223–250.
Mirescu C, Gould E (2006): Stress and adult neurogenesis. Hippocampus16:233–238.
Mirescu C, Peters JD, Gould E (2004): Early life experience alters response ofadult neurogenesis to stress. Nat Neurosci 7:841– 846.
Mitchell SN, Yee BK, Feldon J, Gray JA, Rawlins JN (2000): Activation of theretrohippocampal region in the rat causes dopamine release in thenucleus accumbens: Disruption by fornix section. Eur J Pharmacol 407:131–138.
Nestler EJ (2005): Is there a common molecular pathway for addiction? NatNeurosci 8:1445–1449.
Nestler EJ, Barrot M, DiLeone RJ, Eisch AJ, Gold SJ, Monteggia LM (2002):Neurobiology of depression. Neuron 34:13–25.
Nestler EJ, Malenka RC (2004): The addicted brain. Sci Am 290:78 – 85.Olfson M, Marcus SC, Weissman MM, Jensen PS (2002): National trends in the
use of psychotropic medications by children. J Am Acad Child AdolescPsychiatry 41:514 –521.
Opanashuk LA, Pauly JR, Hauser KF (2001): Effect of nicotine on cerebellargranule neuron development. Eur J Neurosci 13:48 –56.
Paxinos G, Watson C (1997): The Rat Brain in Stereotaxic Coordinates. 3rdedition. San Diego, CA: Academic Press.
D.C. Lagace et al BIOL PSYCHIATRY 2006;60:1121–1130 1129
www.sobp.org/journal

Pham K, Nacher J, Hof PR, McEwen BS (2003): Repeated restraint stresssuppresses neurogenesis and induces biphasic PSA-NCAM expression inthe adult rat dentate gyrus. Eur J Neurosci 17:879 – 886.
Raff M (2003): Adult stem cell plasticity: fact or artifact? Annu Rev Cell Dev Biol19:1–22.
Rao MS, Shetty AK (2004): Efficacy of doublecortin as a marker to analyse theabsolute number and dendritic growth of newly generated neurons inthe adult dentate gyrus. Eur J Neurosci 19:234 –246.
Robinson TE (2004): Neuroscience. Addicted rats. Science 305:951–953.Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S, et al (2003):
Requirement of hippocampal neurogenesis for the behavioral effects ofantidepressants. Science 301:805– 809.
Schmidt-Troschke SO, Ostermann T, Melcher D, Schuster R, Erben CM,Matthiessen PF (2004): [The use of methylphenidate in children: anal-ysis of prescription usage based in routine data of the statutoryhealth insurance bodies concerning drug prescriptions]. Gesund-heitswesen 66:387–392.
Shen RY, Choong KC (2005): Different adaptations in ventral tegmental areadopamine neurons in control and ethanol exposed rats after methyl-phenidate treatment. Biol Psychiatry 59: 635– 642.
Shors TJ, Townsend DA, Zhao M, Kozorovitskiy Y, Gould E (2002): Neurogen-esis may relate to some but not all types of hippocampal-dependentlearning. Hippocampus 12:578 –584.
Sun W, Winseck A, Vinsant S, Park OH, Kim H, Oppenheim RW (2004): Pro-grammed cell death of adult-generated hippocampal neurons is medi-ated by the proapoptotic gene Bax. J Neurosci 24:11205–11213.
Tanapat P, Hastings NB, Rydel TA, Galea LA, Gould E (2001): Exposure tofox odor inhibits cell proliferation in the hippocampus of adult rats
via an adrenal hormone-dependent mechanism. J Comp Neurol437:496 –504.
Teuchert-Noodt G, Dawirs RR, Hildebrandt K (2000): Adult treatment withmethamphetamine transiently decreases dentate granule cell prolifera-tion in the gerbil hippocampus. J Neural Transm 107:133–143.
van Praag H, Schinder AF, Christie BR, Toni N, Palmer TD, Gage FH (2002):Functional neurogenesis in the adult hippocampus.Nature 415:1030–1034.
Volkow ND, Insel TR (2003): What are the long-term effects of methylpheni-date treatment? Biol Psychiatry 54:1307–1309.
Vollmayr B, Simonis C, Weber S, Gass P, Henn F (2003): Reduced cell prolifer-ation in the dentate gyrus is not correlated with the development oflearned helplessness. Biol Psychiatry 54:1035–1040.
Wilens TE, Dodson W (2004): A clinical perspective of attention-deficit/hyperactivity disorder into adulthood. J Clin Psychiatry 65:1301–1313.
Won M, Minabe Y, Tani K, Suzuki K, Kawai M, Sekine Y, et al (2003): The effectsof dentate granule cell destruction on behavioral activity and Fos pro-tein expression induced by systemic MDMA in rats. Neurosci Res 46:153–160.
Yamaguchi M, Suzuki T, Seki T, Namba T, Juan R, Arai H, et al (2004): Repeti-tive cocaine administration decreases neurogenesis in adult rat hip-pocampus. Ann N Y Acad Sci 1025:351–362.
Yang CR, Mogenson GJ (1987): Hippocampal signal transmission to thepedunculopontine nucleus and its regulation by dopamine D2 recep-tors in the nucleus accumbens: An electrophysiological and behaviouralstudy. Neuroscience 23:1041–1055.
Zito JM, Safer DJ, dosReis S, Gardner JF, Boles M, Lynch F (2000): Trends in theprescribing of psychotropic medications to preschoolers. JAMA 283:1025–1030.
1130 BIOL PSYCHIATRY 2006;60:1121–1130 D.C. Lagace et al
www.sobp.org/journal




















