
Juvenile Administration of Methylphenidate Attenuates Adult Hippocampal Neurogenesis

www.e l sev i e r . com/ loca te /eu roneu ro
European Neuropsychopharmacology (2012) 22, 53–63
Methylphenidate administration determines enduringchanges in neuroglial network in ratsCarlo Cavalierea,1, Giovanni Cirillo a,1, Maria Rosaria Biancoa,Walter Adriani b, Antonietta De Simonea, Damiana Leoc,Carla Perrone-Capanod,e, Michele Papaa,⁎
a Department of Medicina Pubblica Clinica e Preventiva, Second University of Naples, Naples, Italyb Department of Cell Biology and Neurosciences, Istituto Superiore di Sanità, Roma, Italyc Department of Pharmacology, University of Montreal, QC, Canadad Department of Biological Sciences, University of Naples “Federico II”, Naples, Italye Institute of Genetics and Biophysics “A. Buzzati Traverso”, CNR, Napoli, Italy
Received 13 May 2010; received in revised form 31 March 2011; accepted 6 April 2011
Glutamate–Aspartate Transporter; nN⁎ Corresponding author at: Departme
80100 Naples, Italy. Tel.: +39 0812996E-mail address: michele.papa@unin
1 Contributed equally to this work.
0924-977X/$ - see front matter © 201doi:10.1016/j.euroneuro.2011.04.003
KEYWORDSADHD;Methylphenidate;Drug addiction;Glutamate transporters;Reactive astrocytosis;nNOS;Plasticity
Abstract
Repeated exposure to psychostimulant drugs induces complex molecular and structuralmodifications in discrete brain regions of the meso-cortico-limbic system. This structuralremodeling is thought to underlie neurobehavioral adaptive responses. Administration toadolescent rats of methylphenidate (MPH), commonly used in attention deficit and hyperactivitydisorder (ADHD), triggers alterations of reward-based behavior paralleled by persistent andplastic synaptic changes of neuronal and glial markers within key areas of the reward circuits. Byimmunohistochemistry, we observe a marked increase of glial fibrillary acidic protein (GFAP) and
neuronal nitric oxide synthase (nNOS) expression and a down-regulation of glial glutamatetransporter GLAST in dorso-lateral and ventro-medial striatum. Using electron microscopy, wefind in the prefrontal cortex a significant reduction of the synaptic active zone length, paralleledby an increase of dendritic spines. We demonstrate that in limbic areas the MPH-induced reactiveastrocytosis affects the glial glutamatergic uptake system that in turn could determineglutamate receptor sensitization. These processes could be sustained by NO production andsynaptic rearrangement and contribute to MPH neuroglial induced rewiring.© 2011 Published by Elsevier B.V.OS, neuronal Nitric Oxide Syntnt of Medicina Pubblica Clinica636; fax: +39 0815666011.a2.it (M. Papa).
1 Published by Elsevier B.V.
Abbreviations: ADHD, Attention Deficit and Hyperactivity Disorder; MPH, methylphenidate; GFAP, Glial Fibrillary Acidic Protein; GLAST,
hasee Preventiva, Institute of Human Anatomy, Second University of Naples,
54 C. Cavaliere et al.
1. Introduction
Repeated exposure to psychostimulant drugs causes complexmolecular, structural and plastic modifications in the brainreward circuitry (Nestler, 2005; Berman et al., 2008; Haydonet al., 2009), in both neuronal and glial compartments ofthe tripartite synapse (Araque et al., 1999; Newman, 2003;Robinson and Kolb, 2004; Machado-Vieira et al., 2009). Theneuroglial network (Alvarez-Maubecin et al., 2000; Giaumeet al., 2010) plays a key role in synaptic function and plas-ticity by participating to the reuptake and release of aminoacid transmitters (Ni and Parpura, 2009; Henneberger et al.,2010) and providing metabolic support to neurons (Rouachet al., 2008; Hamilton and Attwell, 2010).
Several studies support the role of both glutamatergic anddopaminergic transmissions in the development of sensiti-zation to psychostimulants, along with activation of bothNMDA and non-NMDA glutamate receptors (Pierce andKalivas, 1997; Wolf, 1998; Zweifel et al., 2008). Moreover,glutamate induced-neuronal stimulation and sensitizationinduce neuronal nitric oxide synthase (nNOS), triggeringneuronal as glial adaptation following repeated psychosti-mulants administration (Itzhak and Ali, 2006). These struc-tural remodeling events are similar to those associated withexperience-dependent plasticity, and they are thought tounderlie neurobehavioral adaptive responses and drugaddiction (Kelley, 2004; Hyman et al., 2006).
Recently, we have investigated in rats the consequences ofadolescent exposure to MPH, commonly used for attentiondeficit and hyperactivity disorder (ADHD), on neural architec-ture of the brain reward circuitry (Leo et al., 2009). Moreover,as we previously reported, modifications of the neuronalnetwork following glial activation strongly contribute toseveral mental disorders (Musholt et al., 2009), and neurode-generative as neuroinflammatory diseases (Giovannoni et al.,2007; Cavaliere et al., 2007; Cirillo et al., 2010) and representa crucial feature of many brain plasticity processes (Eng andGhirnikar, 1994; Lobsiger and Cleveland, 2007; Reissner andKalivas, 2010).
In different experimental conditions (Trotti et Al., 1998;Giovannoni et al., 2007; Cavaliere et al., 2007) glial activation,usually detected as glial fibrillary acidic protein (GFAP)upregulation, is accompanied by a reduction of glial glutamatetransporter (GLAST) expression, suggesting an impaired regu-lation of glutamate receptor activation during signal transduc-tion, thus promoting overstimulation of glutamate receptors, aprocess that could trigger long lasting plastic changes.
In this study we evaluated the modulation of behavioralimpulsivity by MPH administration and investigated GFAP,GLAST, and nNOS protein expressions as well as thecytoarchitecture in dorso-lateral and ventro-medial striatumof rats treated with repeated exposure to methylphenidateduring adolescence. Using immunohistochemistry, we found anincrease of GFAP and nNOS expressions and a down-regulationof glial glutamate transporter GLAST. Using electron micros-copy, we detected a reduction of the synaptic active zonelength and an increase of dendritic spines. Moreover, in thelimbic areas MPH-induced reactive astrocytosis was observedto affect the glial glutamatergic uptake system. We concludethat the observations could be sustained by NO production andsynaptic rearrangement and contribute to MPH neuroglialinduced rewiring.
2. Experimental procedures
2.1. Animal subjects and treatment design
Animal experimental protocols were approved by institutionalauthorities, on behalf of the Ministry of Health, in close agreementwith European Community Directives and Italian Law. All efforts weremade to minimize animal suffering, to reduce the number of animalsused and to use alternatives to in vivo testing. Ten pregnant Wistarfemale rats (Harlan, Italy) were housed in an air-conditioned room(temperature 211 °C, relative humidity 60±10%), with a 12-h light/dark cycle (lights on at 08:00 h). Water and food (Enriched StandardDiet; Mucedola, Settimo Milanese, Italy) were available ad libitum.The day of deliverywas considered as postnatal day (PND) zero. Even ifonly male subjects were used in the present experiments, cullingincluded the female gender to avoid carryover effects of rearinganimals in sexual segregation. Pups were then weaned on PND 21 andhoused in groups of two siblings, according to sex. Within litters, eachof the siblings was assigned to a different final group, according to asplit-litter design (Zorrilla, 1997). We used 20 male siblings for thepresent experiment. The consequences of adolescent MPH (Ritalin,CIBA-GEIGY SpA, Italy) exposure were investigated. We assigned fivepups toMPHgroup (n=5) to receiveMPH (2 mg/kg/day) solved in salinesolution intraperitoneally (i.p.) during adolescence (PND 30–44); fivesiblingswere assigned toCTRgroup (n=5) and received i.p. injection ofvehicle (saline solution). Animals were tested for motivational-choicebehavior in adulthood (PNDN60, about 4 weeks after the end oftreatment) in a drug-free state.
Given that learning and memory have significant effects onneuroplasticity, we decided animals for the ex vivo analyses to haveno experience at all between last treatment day and the killing. Thus,one CTR and one MPH sibling per litter (n=10 rats for each group) weresacrificed, at least 6 weeks after treatment (PNDN90), for ex vivoanalyses using light and electron microscopy. Fig. 1 clearly illustratesexperimental design.
2.2. Two-choice operant-behavior test
All animals (n=20) were tested in a probability-based reward-rarefaction protocol (Adriani et al., 2006). In brief, few days beforethe schedule started, animals were food-restricted for 2 days, tokeep them at 80–85% of their free-feeding weight, in order toincrease their motivation to work for food delivery. Each animal wasthen placed daily in a computer-controlled operant chamber(Coulbourn Instruments, Allentown, PA, USA), provided with twonose-poking holes, a chamber light, a feeder device, a magazinewhere pellets (45 mg, BioServ, Frenchtown, NJ, USA) were droppedand a magazine light. The nose poking in either hole was detected bya photocell and was recorded by a computer, which also controlledfood delivery. After the 20-min session, animals were returned totheir home cage, where they were given standard chow (approxi-mately 10 g/each).
For behavioral analysis, rats were tested in a drug-free state inadulthood (i.e. 4 weeks after the end of treatment). This was aimed toassess the long-term carryover consequences of adolescent MPHexposure on levels of adult self-control behavior.
During the training phase (1 week for both the groups), nose-poking in one of the two holes (called “small & certain” hole, H1)resulted in the delivery of one pellet of food, whereas nose-poking inthe other hole (“large & uncertain” hole, H5) resulted in the deliveryof five pellets of food. After nose-poking and before food delivery,the chamber light was turned on for 1 s. Following the food delivery,the magazine light was turned on for 25 s, during which nose-pokingwas recorded, but was without any scheduled consequence (time-out). During this 1-week training period, rats were kept for a fewdays at a percentage of actual food delivery (‘Pr’)=100%, to obtain astable baseline.

Figure 1 Experimental design. PND21 male siblings (n=20)were assigned to two different experimental groups: MPH group(n=10) (2 mg/kg/day i.p.) and CTR group (n=10) (saline i.p.administration) treated from PND30 to PND44. All rats under-went behavioral evaluation after treatment (PNDN60). Ratswere sacrificed at PNDN90 for ex vivo analyses, in particular fivesiblings for each group were used for light microscopy analysis(LM), and five for each group for electron microscopy (EM).
55Methylphenidate-induced changes of neuro-glial network
During the testing phase (1 week for both the groups), aprobabilistic dimension was associated to the delivery of the fivepellets following H5 nose-poking. The chamber and the magazinelights were turned on following the previous schedule. However,sometimes the delivery of food could be omitted according to a givenlevel of probability (‘Pr’) controlled by the computer. Theprobability level was kept fixed for each daily session, and wasdecreased progressively over subsequent days. During this 1-weektesting period, ‘Pr’ decreased regularly from 20 to 12.5%. This rangewas chosen as a stronger rarefaction challenge (Adriani et al., 2006).
All the treated animals were able to perform behavioral sessionand were sacrificed at PNDN90 for histological analyses.
2.3. Tissue preparation
For sacrifice, rats (n=20) were deeply anesthetized with chloralhydrate (300 mg/kg i.p.) and perfused transcardially with a solution(Tris–HCl 0.1 M, ethylenediaminetetraacetic acid 10 mM) followedby cold (4 °C) 4% paraformaldehyde plus 0.1% glutaraldehyde in0.01 M phosphate-buffered saline (PBS) at pH 7.4.
For light microscopy (n=5 for each group), the brain wasremoved and post-fixed 2 h in the same fixative, then soaked in30% sucrose PBS and frozen in chilled isopentane on dry ice. Serialsections were cut on a cooled sliding microtome at a thickness of25 μm and collected in cold PBS for immunohistochemistry.
For electron microscopy (n=5 for each group), the brain wasremoved and placed overnight in 4% paraformaldehyde added to 1%glutaraldehyde in 0.01 M PBS, pH 7.4 at 4 °C. Whole brain wasserially cut on vibratome at a thickness of 500 μm and collected incold PBS. Then, serial sections were reviewed, and tissue blocks foreach region were selected, according to Paxinos coordinates(Paxinos et al., 1980). Selected tissue blocks were rinsed inphosphate buffer and post-fixed in 2% osmium tetroxide inphosphate buffer for 20 min, dehydrated in ethanol and embeddedin araldite resin embedding kit (TAAB Laboratories, UK). Ultra-thinsections (1 μm thickness) including the 2nd to 4th cortical layerswere prepared within the interval +5.20 to +3.20 from bregma(Paxinos et al., 1980). Then, sections were stained with uranylacetate and lead citrate and examined with an EFTEM Lybra 120electron microscope (Zeiss, Oberchoken, Germany).
2.4. Immunohistochemistry
Brain sections were pretreated with hydrogen peroxide prior toincubation of primary antibody to eliminate endogenous peroxidaseactivity. Then sections were blocked in 10% normal serum in 0.01 MPBS, 0.25% Triton X-100 for 1 h at room temperature (RT). Eachprimary antibody (monoclonal mouse antibodies directed to GFAP,1:400, Sigma-Aldrich; polyclonal rabbit antibodies directed to nNOS,1:500, Cayman; polyclonal guinea pig antibodies directed to GLAST,1:4000, Chemicon) was diluted in 0.01 M PBS containing 10% normalserum 0.25% Triton X-100. Free-floating slices were incubated for48 h at 4 °C on an oscillating plate. Sections were washed severaltimes in PBS and incubated with the appropriate biotinylatedsecondary antibody (Vector Labs Inc., Burlingame, CA, USA; 1:200)for 90 min at RT, washed in PBS and processed using the Vectastainavidin–biotin peroxidase kit (Vector Labs Inc.) for 90 min, also at RT.Sections were washed in 0.05 M Tris–HCl and reacted with 3,3-diaminobenzidine tetrahydrochloride (DAB; Sigma, Milan, Italy) in0.5 mg/ml Tris–HCl and 0.01% hydrogen peroxide. Sections weremounted on chrome-alum gelatine coated slides, dehydrated andcoverslipped. Appropriate controls lacking primary antibodies wereperformed for each of the antibodies. Adjacent sections were Nissl-stained. Slides were imaged with a Zeiss Axioskope 2 lightmicroscope (Zeiss) equipped with high-resolution digital camera(C4742-95, Hamamatsu Photonics, Italy).
A densitometry of the several markers and morphometricfeatures of cells in Caudate–Putamen (CPu), nucleus Accumbens(Acb) and Prefrontal Cortex (PFc) was accomplished using computer-assisted image analysis system (MCID 7.0; Imaging Res. Inc.,Canada). Sections of the CPu were analyzed within the interval+2.20 to −1.30 from bregma (Paxinos et al., 1980). The CPu wasdivided in two halves a dorsolateral (Dl-) and ventromedial (Vm-)one. The Acb was outlined for each section and analyzed accordingto Nissl-stained adjacent sections and the calbindin-treated sections(Zahm and Brog, 1992). The core was divided from the shellthroughout the posterior horn of the anterior commissure. Sectionsof the PFc were analyzed within the interval +5.20 to +3.20 frombregma (Paxinos et al., 1980).
For GFAP and nNOS expressions, allowing discrete imaging ofstructures, we counted the number of positive cell elements, andtherefore these two markers were expressed as proportion of thetotal target area relative to the scanned area (proportional area).For GLAST expression, where labeling appeared as diffuse staining, adensitometric approach was preferred. Briefly, a background image(a region of the slide not covered by the sample) was acquired andthen subtracted to each sample acquisition, according to the
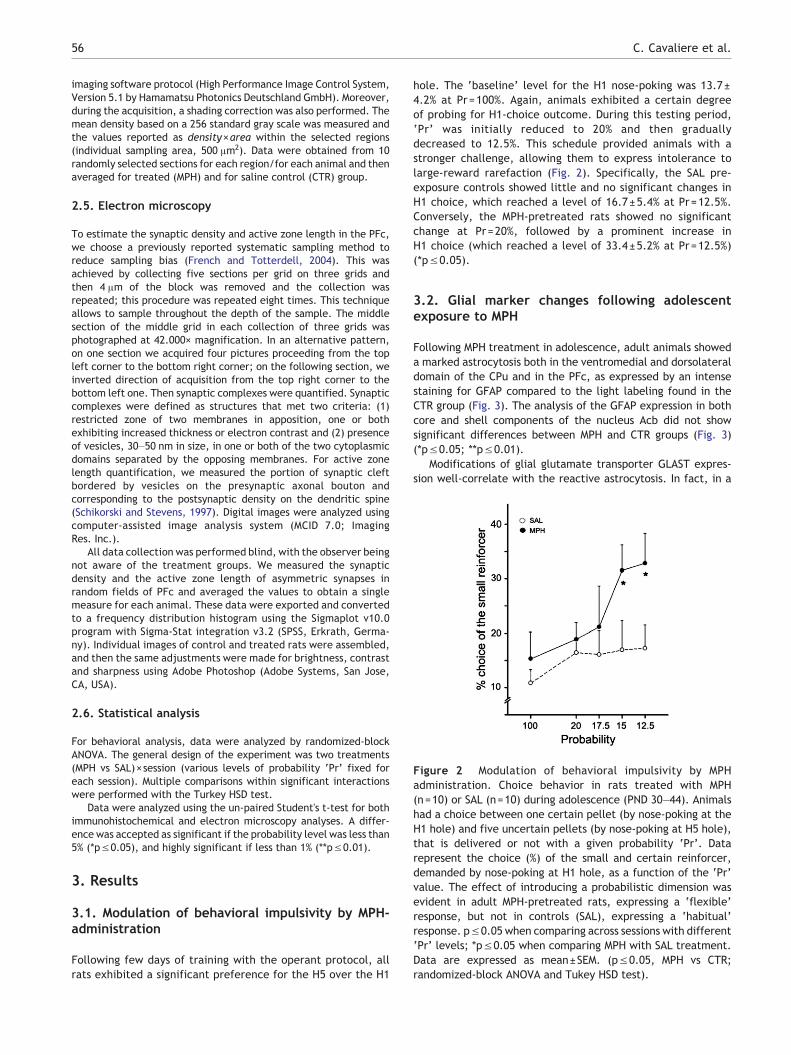
Figure 2 Modulation of behavioral impulsivity by MPHadministration. Choice behavior in rats treated with MPH(n=10) or SAL (n=10) during adolescence (PND 30–44). Animalshad a choice between one certain pellet (by nose-poking at theH1 hole) and five uncertain pellets (by nose-poking at H5 hole),that is delivered or not with a given probability ‘Pr’. Datarepresent the choice (%) of the small and certain reinforcer,demanded by nose-poking at H1 hole, as a function of the ‘Pr’value. The effect of introducing a probabilistic dimension wasevident in adult MPH-pretreated rats, expressing a ‘flexible’response, but not in controls (SAL), expressing a ‘habitual’response. p≤0.05 when comparing across sessions with different‘Pr’ levels; *p≤0.05 when comparing MPH with SAL treatment.Data are expressed as mean±SEM. (p≤0.05, MPH vs CTR;randomized-block ANOVA and Tukey HSD test).
56 C. Cavaliere et al.
imaging software protocol (High Performance Image Control System,Version 5.1 by Hamamatsu Photonics Deutschland GmbH). Moreover,during the acquisition, a shading correction was also performed. Themean density based on a 256 standard gray scale was measured andthe values reported as density×area within the selected regions(individual sampling area, 500 μm2). Data were obtained from 10randomly selected sections for each region/for each animal and thenaveraged for treated (MPH) and for saline control (CTR) group.
2.5. Electron microscopy
To estimate the synaptic density and active zone length in the PFc,we choose a previously reported systematic sampling method toreduce sampling bias (French and Totterdell, 2004). This wasachieved by collecting five sections per grid on three grids andthen 4 μm of the block was removed and the collection wasrepeated; this procedure was repeated eight times. This techniqueallows to sample throughout the depth of the sample. The middlesection of the middle grid in each collection of three grids wasphotographed at 42.000× magnification. In an alternative pattern,on one section we acquired four pictures proceeding from the topleft corner to the bottom right corner; on the following section, weinverted direction of acquisition from the top right corner to thebottom left one. Then synaptic complexes were quantified. Synapticcomplexes were defined as structures that met two criteria: (1)restricted zone of two membranes in apposition, one or bothexhibiting increased thickness or electron contrast and (2) presenceof vesicles, 30–50 nm in size, in one or both of the two cytoplasmicdomains separated by the opposing membranes. For active zonelength quantification, we measured the portion of synaptic cleftbordered by vesicles on the presynaptic axonal bouton andcorresponding to the postsynaptic density on the dendritic spine(Schikorski and Stevens, 1997). Digital images were analyzed usingcomputer-assisted image analysis system (MCID 7.0; ImagingRes. Inc.).
All data collection was performed blind, with the observer beingnot aware of the treatment groups. We measured the synapticdensity and the active zone length of asymmetric synapses inrandom fields of PFc and averaged the values to obtain a singlemeasure for each animal. These data were exported and convertedto a frequency distribution histogram using the Sigmaplot v10.0program with Sigma-Stat integration v3.2 (SPSS, Erkrath, Germa-ny). Individual images of control and treated rats were assembled,and then the same adjustments were made for brightness, contrastand sharpness using Adobe Photoshop (Adobe Systems, San Jose,CA, USA).
2.6. Statistical analysis
For behavioral analysis, data were analyzed by randomized-blockANOVA. The general design of the experiment was two treatments(MPH vs SAL)×session (various levels of probability ‘Pr’ fixed foreach session). Multiple comparisons within significant interactionswere performed with the Turkey HSD test.
Data were analyzed using the un-paired Student's t-test for bothimmunohistochemical and electron microscopy analyses. A differ-ence was accepted as significant if the probability level was less than5% (*p≤0.05), and highly significant if less than 1% (**p≤0.01).
3. Results
3.1. Modulation of behavioral impulsivity by MPH-administration
Following few days of training with the operant protocol, allrats exhibited a significant preference for the H5 over the H1
hole. The ‘baseline’ level for the H1 nose-poking was 13.7±4.2% at Pr=100%. Again, animals exhibited a certain degreeof probing for H1-choice outcome. During this testing period,‘Pr’ was initially reduced to 20% and then graduallydecreased to 12.5%. This schedule provided animals with astronger challenge, allowing them to express intolerance tolarge-reward rarefaction (Fig. 2). Specifically, the SAL pre-exposure controls showed little and no significant changes inH1 choice, which reached a level of 16.7±5.4% at Pr=12.5%.Conversely, the MPH-pretreated rats showed no significantchange at Pr=20%, followed by a prominent increase inH1 choice (which reached a level of 33.4±5.2% at Pr=12.5%)(*p≤0.05).
3.2. Glial marker changes following adolescentexposure to MPH
Following MPH treatment in adolescence, adult animals showeda marked astrocytosis both in the ventromedial and dorsolateraldomain of the CPu and in the PFc, as expressed by an intensestaining for GFAP compared to the light labeling found in theCTR group (Fig. 3). The analysis of the GFAP expression in bothcore and shell components of the nucleus Acb did not showsignificant differences between MPH and CTR groups (Fig. 3)(*p≤0.05; **p≤0.01).
Modifications of glial glutamate transporter GLAST expres-sion well-correlate with the reactive astrocytosis. In fact, in a

Figure 3 GFAP immunostaining in brain reward regions of MPH-treated rats. Following MPH treatment in adolescence (PND 30–44),both the ventromedial and the dorsolateral domain of the CPu, and the PFc of adult animals showed a marked astrocytosis, asexpressed by an intense staining for GFAP compared to the light labeling found in the CTR group. The analysis of the GFAP expression inboth components of the nucleus Acb, the core and shell, did not show significative differences between the MPH group and the CTRone. Data are expressed as mean±SEM ; (* p≤0.05; **p≤0.01, MPH vs CTR; un-paired Student's t-test). Scale bar: 50 μm(20× objective).
57Methylphenidate-induced changes of neuro-glial network
close spatial correlation with the astrocytosis, we found asignificative decrease of GLAST in both the CPu and the PFc ofthe MPH group compared to the strong labeling found in the CTRgroup (Fig. 4A). In a same manner, the analysis of GLASTexpression in the nucleus Acb did not reveal significativedifferences between the two experimental groups (Fig. 4)(*p≤0.05; **p≤0.01). Higher magnification image (100×)revealed a specific and spotted pattern for GLAST staining(Fig. 4B).
3.3. Neuronal marker and cytoarchitecture in brainreward pathways are altered by MPH
Since NO represents a putative messenger for the neuroglialcross-talk, we investigated whether nNOS staining correlateswith the changes of glial markers observed in MPH-treated rats.We found an increase of nNOS expression both in theventromedial and dorsolateral domain of CPu as expressed byan intense staining for nNOS compared to the CTR group

Figure 4 GLAST expression in brain reward regions of MPH-exposed rats. (A) The glial glutamate transporter GLAST well-correlateswith increased GFAP levels. In fact, in a close spatial correlation with the astrocytosis, we found a significant decrease of GLAST inboth the CPu and the PFc of the MPH group compared to the strong labeling found in the CTR group. In a same manner, following thetrend of GFAP staining, the analysis of GLAST expression in the nucleus Acb did not reveal significative differences between the twoexperimental groups. (B) GLAST control antibody image of Dl-CPu region. Data are expressed as mean±SEM; (* p≤0.05; **p≤0.01,MPH vs CTR; un-paired Student's t-test). Scale bar: (A) 50 μm (20× objective); (B) 10 μm (100× objective).
58 C. Cavaliere et al.
(Fig. 5). The analysis of the nNOS expression in the PFc and inboth components of the nucleus Acb did not show significantdifferences between MPH and CTR group (Fig. 5).
To evaluate if adolescent MPH exposure was able to inducelong lasting changes in synaptic number and shape, we analyzedby electron microscopy the synaptic density and the active zonelength in the PFc of adult animals. This brain structure waschosen because of its central role in the reward circuitry. Wemeasured the synaptic density and the active zone length ofasymmetric synapses in random fields of PFc and averaged the
values to obtain a single measure for each animal (Fig. 6). MPHtreated animals had a higher synaptic density compared with salinecontrols (4.7±0.33 synapses/9 μm2 of neuropil vs 3.71±0.18synapses/9 μm2; t=−2.63, degrees of freedom=8, *p=0.03;Fig. 6a,b,c). We then measured the active zone length, composedof the synaptic cleft bordered by vesicles on the presynaptic axonalbouton and corresponding to the postsynaptic density on thedendritic spine (Schikorski and Stevens, 1997). The length of theactive zone measured in asymmetric spine synapses found in arandomsampleof neuropil fieldswithinPFcwas significantly shorter

Figure 5 nNOS immunostaining. nNOS was increased both in the ventromedial and in dorsolateral domain of CPu as expressed by anintense staining for nNOS compared to that found in the CTR group. The analysis of the nNOS expression in the PFc and in bothcomponents of the nucleus Acb, the core and shell, did not show significant differences between the MPH group and the CTR one. Dataare expressed as mean±SEM; (*p≤0.05; **p≤0.01, MPH vs CTR; un-paired Student's t-test). Scale bar: 50 μm (20× objective).
59Methylphenidate-induced changes of neuro-glial network
in MPH-treated rats, compared to CTR ones (67.96±1.69 vs 83.521.91 μm; t=6.09, degrees of freedom=8, **p≤0.01; Fig. 6a,b,d).
4. Discussion
Growing evidence suggests the role of tripartite synapse tothe pathophysiology of several cognitive and behavioraldisorders (Gray et al., 2007; Machado-Vieira et al., 2009;Musholt et al., 2009) establishing that several drugs of abuseinterfere with synaptic plasticity mechanisms in rewardregions, thus influencing their function (Wolf et al., 2004;Kauer and Malenka, 2007; Beique, 2009).
In the present study, we focused on behavioral adapta-tions and neuroglial modifications within the brain rewardcircuitry of rats chronically treated during adolescence withthe psychostimulant methylphenidate (MPH), that wasshown to produce alterations of reward-based behavior andpersistent region-specific changes in brain reward circuitry(Adriani et al., 2006; Leo et al., 2009). When testingadolescent animals, control subjects showed increased H5demands and a clear tendency towards disappearance of H1responses. In other words, control animals seemed to havelearned that, despite a great number of lost rewards, a largefood reinforcer will eventually come and preferred to waitfor this ‘lucky’ but ‘rare’ event, and kept demanding at the

Figure 6 Synaptic organization in the PFc of MPH-treated rats. Changes in the number and structure of synaptic contacts in adult malerats exposed to saline (CTR) orMPH (2 mg/kg/day) during adolescence (PND 30–44). The synaptic complexes are shown (arrow) for CTR (a)and MPH (b) treated groups (individual sampling area, 9 μm2; total sampling area, 270 μm2). (c) The number of PFc synapses shows asignificant increase (*p=0.03) in MPH-exposed rats (filled column) (4.7±0.33 synapses/9 μm2) compared to controls (CTR, open column)(3.71±0.18 synapses/9 μm2). (d) The active zone (AZ) length (post-synaptic density) of the prefrontal synapses shows a significantdecrease (**p≤0.01) in MPH-exposed rats (filled column) (67.96±1.69 μm) compared to controls (CTR, open column) (83.52±1.91 μm).Data are expressed as mean±SEM; (*p≤0.05; **p≤0.01, MPH vs CTR; un-paired Student's t-test). Scale bar: 1 μm.
60 C. Cavaliere et al.
H5 hole. Consistently, it has been shown that the proportionof ‘risky’ choices does increase as the scheduled availabilityof a feeding resource decreases (Hastjarjo et al., 1990;Kaminski and Ator, 2001). Interestingly, MPH-treated sub-jects did not apparently react to the stochastic-rarefactionchallenge and were less dependent upon being actually food-rewarded, possibly due to direct MPH-induced hedonicactivation (Marsteller et al., 2002). MPH treated ratsexhibited sensitivity to the enhancement of large-reinforcerrarefaction and a marked shift towards H1 nose-poking,delivering the smaller but certain reward. In line withprevious evidence (Adriani et al., 2004), MPH administrationallowed adolescent animals to increase their ability to copewith adverse reinforcement contingencies, represented hereby the unpredictable omission of large-reward delivery. Anincrease in H1 responding, shown by MPH-pretreated sub-jects, is consistent with enhanced impulsivity (Mobini et al.,2000) but also it should be interpreted in terms of (a)increased ‘economical’ efficiency, and/or (b) enhancedbehavioral flexibility. While the latter is possibly due to animprovement of striatal plasticity (Leo et al., 2009), theformer feature may result from a lower impact of ‘subcor-tical’ drives (the binge-reward preference) and/or a morecareful ‘cortical’ evaluation of the long-term payoff
(Ridderinkhof et al., 2004). MPH-exposed rats may perceivethe ‘global’ effects of accumulating omitted rewards (the‘losses’) across time, rather than being merely driven by the‘unitary’ size of the eventual outcome (the binge but rare‘wins’) (McClure et al., 2004; Ridderinkhof et al., 2004).Alternatively, MPH-pretreated rats may be less dependent onestablished behavioral habits compared to the SAL-pretreatedsiblings. Interestingly, enhanced flexibility may representanother ‘adolescent-like’ feature in adult rats pre-exposed toMPH.
Although molecular evidence suggests that this inducedphenotype could be related to stable changes in geneexpression and persistent alterations within the mesocorti-colimbic DA system, the main pathway of the brain-rewardcircuitry (Nestler, 2005; Robinson and Kolb, 2004), the wholemorphological and molecular changes of chronic exposure toMPH during critical periods of development are poorlyunderstood (Carlezon and Konradi, 2004). As also elsewherereported (Guilarte et al., 2003; Lyles and Cadet, 2003), weshow here an astrocytic activation and a significant GFAPincrease in rat striatum and PFc after MPH administration.The lack of a significant astrogliosis in the nucleusAccumbens demonstrated by other authors (Narita et al.,2008, 2006) is probably due to the different experimental

61Methylphenidate-induced changes of neuro-glial network
follow-up and/or treatment protocol. GFAP increase wasparalleled by a reduction of glial glutamate transporterGLAST, probably as consequence of morpho-functionalrearrangement of astrocytic network, as already demon-strated (Rao et al., 2003; Cavaliere et al., 2007; Cirillo et al.,2010). These data are also strengthened by recent studies(Karisson et al., 2008) that demonstrated that a loss ofglutamate transporter GLAST causes locomotor hyperactivityand exaggerated responses to psychotomimetics, and pro-pose glutamate homeostasis as a target for treating addictivedisorders (Bunch et al., 2009; Reissner and Kalivas, 2010).Both GLAST (or EAAT1) and EAAT2 (or GLT1), almostexclusively expressed on astroglial cells, account for themajority of the glial glutamate transport capacity in thedeveloping brain (Maragakis and Rothstein, 2004; Schousboe etal., 2004; Kugler and Schleyer, 2004). Glial glutamate trans-porters provide an efficient and rapid removal of glutamatefrom the synaptic cleft, prevent the spread of excitation butalso limit the potential detrimental effects (Jiang and Amara,2011), preventing over-activation of glutamate receptors,mainly the NMDA subtype (Schousboe et al., 2004). Recentevidence suggests that nitridergic system, through nNOSinduction, and glutamatergic system, via NMDA activation,triggers astrocytic activation (Brahmachari et al., 2006). Forthis purpose, we evaluated nNOS expression in the rewardcircuitry andwe found that nNOS levels well correlated to glialrearrangement in fronto-striatal regions. The increase of nNOSexpression and subsequently NO could activate astrocytes(Brahmachari et al., 2006) and impair neuronal transmissionthrough changes of GLAST expression. On the other side, theincrease of glutamate availability to the post-synapticinterface could sensitize NMDA-receptor functionally linkedto nNOSenzyme. In both cases the neuronal overstimulation byexcess of glutamate could lead to sensitization of theglutamatergic system.
Among the persistent consequences of adolescent MPHexposure, we previously described region-specific changes ofselected neuronal markers within CPu and PFc, causingadaptations in neurons of the direct striatal output pathway(Adriani et al., 2006; Leo et al., 2009). This prompted us toevaluate by electronmicroscopy themorphological rearrange-ment of the striatal neuropil. The administration of MPHduringadolescence determineda significant increase of spine numberand a reduction of the active zone spine interface in PFc inPNDN90 rats. The increase of synaptic contacts in MPH treatedrats could be interpreted as the plastic changes of brain rewardcircuitry following the reduction of active zone length. Theselong-lasting effects indicate an enduring remodeling ofsynaptic morphology in this brain area. While it is known thatmorphological remodeling of dendritic spines following co-caine, amphetamine and nicotine administration (Robinsonand Kolb, 2004; Hernandez and Kelley, 2005), to ourknowledge our works first demonstrated MPH-induced persis-tent changes of striatal synaptic structure. In line with currentunderstanding (Russell et al., 2006; Narita et al., 2008; Cirilloet al., 2010), the present study provides evidence that long-term MPH administration to adolescent rats leads to long-lasting changes in the astrocytic activation, the expression ofglial glutamate transporter, the induction of neuronal NOS inbrain regions implicated in reward circuitry.
These findings suggest an important role for the glialcompartment in the tripartite synapse plasticity. Further
studies are needed to clarify the paradoxical, sedativeeffect of MPH-treatment in ADHD affected patients, and therole of astrocytic morpho-functional rearrangement in thisadaptation.
Role of the funding source
This work was supported by grants from Regione Campania, by PRIN/MIUR no 2007_STRNHK_002, Italian Ministry of Health “Under 40”2007. Funding sources had no further role in study design, in thecollection, analysis and interpretation of data, in the writing of thereport, and in the decision to submit the paper for publication.
Contributors
C.C., G.C., and M.R.B. realized the experiments on neuronal andglial markers and morphology, performed the statistical analysesand wrote the first draft of the manuscript. D.L. and W.A.contributed to experimental design, treated the animals and carriedout the behavioral experiments. A.DeS. contributed to electronmicroscopy image acquisition. C.P.-C. contributed to experimentaldesign, and coordinated the experimental work. M.P. contributed toexperimental design, devised the experiments on neuronal and glialmarkers and morphology and coordinated the experimental work. Allauthors contributed to and have approved the final manuscript.
Conflict of interest
None.
Acknowledgments
This work was supported by a postdoctoral fellowship from theAssociazione Levi-Montalcini (M.R.B. and G.C.), by grants fromRegione Campania (L.R. N.5 Bando 2003 to M.P.), Regione Campania(Prog. Spec art 12 E.F. 2000 to M.P.), by PRIN/MIUR no2007_STRNHK_002 (to CPC), and the Italian Ministry of Health“Under 40” 2007 (to WA).
References
Adriani, W., Rea, M., Baviera, M., Invernizzi, W., Carli, M., Ghirardi,O., Caprioli, A., Laviola, G., 2004. Acetyl-L-carnitine reducesimpulsive behaviour in adolescent rats. Psychopharmacology176, 296–304.
Adriani, W., Leo, D., Greco, D., Rea, M., di Porzio, U., Laviola, G.,Perrone-Capano, C., 2006. Methylphenidate administration toadolescent rats determines plastic changes on reward-relatedbehavior and striatal gene expression. Neuropsychopharmacol-ogy 31, 1946–1956.
Alvarez-Maubecin, V., Garcia-Hernandez, F., Williams, J.T., VanBockstaele, E.J., 2000. Functional coupling between neurons andglia. J. Neurosci. 20, 4091–4098.
Araque, A., Parpura, V., Sanzgiri, R.P., Haydon, P.G., 1999.Tripartite synapses: glia, the unacknowledged partner. TrendsNeurosci. 22, 208–215.
Beique, J.C., 2009. Homeostatic plasticity in a reward processingregion: accumbens neurons scale too! (Commentary on Sun &Wolf). Eur. J. Neurosci. 30, 537–538.
Berman, S., O'Neill, J., Fears, S., Bartzokis, G., London, E.D., 2008.Abuse of amphetamines and structural abnormalities in thebrain. Ann. N. Y. Acad. Sci. 1141, 195–220.

62 C. Cavaliere et al.
Brahmachari, S., Fung, Y.K., Pahan, K., 2006. Induction of glialfibrillary acidic protein expression in astrocytes by nitric oxide.J. Neurosci. 26, 4930–4939.
Bunch, L., Erichsen, M.N., Jensen, A.A., 2009. Excitatory amino acidtransporters as potential drug targets. Expert Opin. Ther.Targets 13, 719–731.
Carlezon Jr., W.A., Konradi, C., 2004. Understanding the neurobi-ological consequences of early exposure to psychotropic drugs:linking behavior with molecules. Neuropharmacology 47, 47–60.
Cavaliere, C., Cirillo, G., Bianco, M.R., Rossi, F., De Novellis, V.,Maione, S., Papa, M., 2007. Gliosis alters expression and uptakeof spinal glial amino acid transporters in a mouse neuropathicpain model. Neuron Glia Biol. 3, 141–153.
Cirillo, G., Maggio, N., Bianco, M.R., Vollono, C., Sellitti, S., Papa,M., 2010. Discriminative behavioral assessment unveils remark-able reactive astrocytosis and early molecular correlates in basalganglia of 3-nitropropionic acid subchronic treated rats. Neu-rochem. Int. 56, 152–160.
Eng, L.F., Ghirnikar, R.S., 1994. GFAP and astrogliosis. Brain Pathol.4, 229–237.
French, S.J., Totterdell, S., 2004. Quantification of morphologicaldifferences in boutons from different afferent populations to thenucleus accumbens. Brain Res. 1007, 167–177.
Giaume, C., Koulakoff, A., Roux, L., Holcman, D., Rouach, N., 2010.Astroglial networks: a step further in neuroglial and gliovascularinteractions. Nat. Rev. Neurosci. 11, 87–99.
Giovannoni, R., Maggio, N., Bianco, M.R., Cavaliere, C., Cirillo, G.,Lavitrano, M., Papa, M., 2007. Reactive astrocytosis and glial gluta-mate transporter clustering are early changes in a spinocerebellarataxia type 1 transgenic mousemodel. Neuron Glia Biol. 3, 335–351.
Gray, J.D., Punsoni, M., Tabori, N.E., Melton, J.T., Fanslow, V.,Ward, M.J., Zupan, B., Menzer, D., Rice, J., Drake, C.T., Romeo,R.D., Brake, W.G., Torres-Reveron, A., Milner, T.A., 2007.Methylphenidate administration to juvenile rats alters brainareas involved in cognition, motivated behaviors, appetite, andstress. J. Neurosci. 27, 7196–7207.
Guilarte, T.R., Nihei, M.K., McGlothan, J.L., Howard, A.S., 2003.Methamphetamine-induced deficits of brain monoaminergicneuronal markers: distal axotomy or neuronal plasticity. Neuro-science 122, 499–513.
Hamilton, N.B., Attwell, D., 2010. Do astrocytes really exocytoseneurotransmitters? Nat. Rev. Neurosci. 11, 227–238.
Hastjarjo, T., Silberberg, A., Hursh, S.R., 1990. Risky choice as afunction of amount and variance in food supply. J. Exp. Anal.Behav. 53, 155–161.
Haydon, P.G., Blendy, J., Moss, S.J., Rob Jackson, F., 2009. Astrocyticcontrol of synaptic transmission and plasticity: a target for drugs ofabuse? Neuropharmacology 56 (Suppl. 1), 83–90.
Henneberger, C., Papouin, T., Oliet, S.H., Rusakov, D.A., 2010.Long-term potentiation depends on release of D-serine fromastrocytes. Nature 463, 232–236.
Hernandez, P.J., Kelley, A.E., 2005. Cracking addiction the secondtime around: reconsolidation of drug-related memories. Neuron47, 772–775.
Hyman, S.E., Malenka, R.C., Nestler, E.J., 2006. Neural mechanismsof addiction: the role of reward-related learning and memory.Annu. Rev. Neurosci. 29, 565–598.
Itzhak, Y., Ali, S.F., 2006. Role of nitrergic system in behavioral andneurotoxic effects of amphetamine analogs. Pharmacol. Ther.109, 246–262.
Jiang, J., Amara, S.G., 2011. New views of glutamate transporterstructure and function: advances and challenges. Neuropharma-cology 60, 172–181.
Kaminski, B.J., Ator, N.A., 2001. Behavioral and pharmacologicalvariables affecting risky choice in rats. J. Exp. Anal. Behav. 75,275–297.
Karisson, R.-M., Heilig, M., Holmes, A., 2008. Loss of glutamatetransporter GLAST (EAAT1) causes locomotor hyperactivity and
exaggerated responses to psychotomimetics: rescue by haloper-idol and mGlu2/3 agonist. Biol. Psychiatry 64, 810–814.
Kauer, J.A., Malenka, R.C., 2007. Synaptic plasticity and addiction.Nat. Rev. Neurosci. 8, 844–858.
Kelley, A.E., 2004. Memory and addiction: shared neural circuitryand molecular mechanisms. Neuron 44, 161–179.
Kugler, P., Schleyer, V., 2004. Developmental expression ofglutamate transporters and glutamate dehydrogenase in astro-cytes of the postnatal rat hippocampus. Hippocampus 14,975–985.
Leo, D., Adriani, W., Cavaliere, C., Cirillo, G., Marco, E.M., Romano,E., di Porzio, U., Papa, M., Perrone-Capano, C., Laviola, G.,2009. Methylphenidate to adolescent rats drives enduringchanges of accumbal Htr7 expression: implications for impulsivebehavior and neuronal morphology. Genes Brain Behav. 8,356–368.
Lobsiger, C.S., Cleveland, D.W., 2007. Glial cells as intrinsiccomponents of non-cell-autonomous neurodegenerative disease.Nat. Neurosci. 10, 1355–1360.
Lyles, J., Cadet, J.L., 2003. Methylenedioxymethamphetamine(MDMA, Ecstasy) neurotoxicity: cellular and molecular mecha-nisms. Brain Res. Brain Res. Rev. 42, 155–168.
Machado-Vieira, R., Manji, H.K., Zarate, C.A., 2009. The role of thetripartite glutamatergic synapse in the pathophysiology andtherapeutics of mood disorders. Neuroscientist 15, 525–539.
Maragakis, N.J., Rothstein, J.D., 2004. Glutamate transporters:animal models to neurologic disease. Neurobiol. Dis. 15,461–473.
Marsteller, D.A., Gerasimov, M.R., Schiffer, W.K., Geiger, J.M.,Barnett, C.R., Borg, J.S., Scott, S., Ceccarelli, J., Volkow, N.D.,Molina, P.E., Alexoff, D.L., Dewey, S.L., 2002. Acute handlingstress modulates methylphenidate-induced catecholamine over-flow in the medial prefrontal cortex. Neuropsychopharmacology27, 163–170.
McClure, S.M., Laibson, D.I., Loewenstein, H., Cohen, J.D., 2004.Separate neural systems value immediate and delayed monetaryrewards. Science 306, 503–507.
Mobini, S., Chiang, T.J., Al-Ruwaitea, A.S., Ho, M.Y., Bradshaw,C.M., Szabadi, E., 2000. Effect of central 5-hydroxytryptaminedepletion on inter-temporal choice: a quantitative analysis.Psychopharmacology 149, 313–318.
Musholt, K., Cirillo, G., Cavaliere, C., Rosaria, B.M., Bock, J.,Helmeke, C., Braun, K., Papa, M., 2009. Neonatal separationstress reduces glial fibrillary acidic protein- and S100betaimmunoreactive astrocytes in the rat medial precentral cortex.Dev. Neurobiol. 69, 203–211.
Narita, M., Miyatake, M., Narita, M., Shibasaki, M., Shindo, K.,Nakamura, A., Kuzumaki, N., Nagumo, Y., Suzuki, T., 2006.Direct evidence of astrocytic modulation in the development ofrewarding effects induced by drugs of abuse. Neuropsychophar-macology 31, 2476–2488.
Narita, M., Suzuki, M., Kuzumaki, N., Miyatake, M., Suzuki, T., 2008.Implication of activated astrocytes in the development of drugdependence differences between methamphetamine and mor-phine. Ann. N. Y. Acad. Sci. 1141, 96–104.
Nestler, E.J., 2005. Is there a common molecular pathway foraddiction? Nat. Neurosci. 8, 1445–1449.
Newman, E.A., 2003. New roles for astrocytes: regulation of synaptictransmission. Trends Neurosci. 26, 536–542.
Ni, Y., Parpura, V., 2009. Dual regulation of Ca2+−dependentglutamate release from astrocytes: vesicular glutamate trans-porters and cytosolic glutamate levels. Glia 57, 1296–1305.
Paxinos, G., Watson, C.R., Emson, P.C., 1980. AChE-stainedhorizontal sections of the rat brain in stereotaxic coordinates.J. Neurosci. Methods 3, 129–149.
Pierce, R.C., Kalivas, P.W., 1997. A circuitry model of the expressionof behavioral sensitization to amphetamine-like psychostimu-lants. Brain Res. Brain Res. Rev. 25, 192–216.

63Methylphenidate-induced changes of neuro-glial network
Rao, S.D., Yin, H.Z., Weiss, J.H., 2003. Disruption of glial glutamatetransport by reactive oxygen species produced in motor neurons.J. Neurosci. 23, 2627–2633.
Reissner, K.J., Kalivas, P.W., 2010. Using glutamate homeostasis asa target for treating addictive disorders. Behav. Pharmacol. 21,514–522.
Ridderinkhof, K.R., van der Wildenberg, W.P.M., Segalowitz, S.J.,Carter, C.S., 2004. Neurocognitive mechanisms of cognitivecontrol: the role of prefrontal cortex in action selection,response inhibition, performance monitoring and reward-basedlearning. Brain Cogn. 56, 129–140.
Robinson, T.E., Kolb, B., 2004. Structural plasticity associated withexposure to drugs of abuse. Neuropharmacology 47, 33–46.
Rouach, N., Koulakoff, A., Abudara, V., Willecke, K., Giaume, C.,2008. Astroglial metabolic networks sustain hippocampal synap-tic transmission. Science 322, 1551–1555.
Russell, V.A., Oades, R.D., Tannock, R., Killeen, P.R., Auerbach,J.G., Johansen, E.B., Sagvolden, T., 2006. Response variability inAttention-Deficit/Hyperactivity Disorder: a neuronal and glialenergetics hypothesis. Behav. Brain Funct. 2, 30.
Schikorski, T., Stevens, C.F., 1997. Quantitative ultrastructuralanalysis of hippocampal excitatory synapses. J. Neurosci. 17,5858–5867.
Schousboe, A., Sarup, A., Bak, L.K., Waagepetersen, H.S., Larsson,O.M., 2004. Role of astrocytic transport processes in glutama-tergic and GABAergic neurotransmission. Neurochem. Int. 45,521–527.
Trotti, D., Danbolt, N.C., Volterra, A., 1998. Glutamate transportersare oxidant-vulnerable: a molecular link between oxidative andexcitotoxic neurodegeneration? Trends Pharmacol. Sci. 19,328–334.
Wolf, M.E., 1998. The role of excitatory amino acids in behavioralsensitization to psychomotor stimulants. Prog. Neurobiol. 54,679–720.
Wolf, M.E., Sun, X., Mangiavacchi, S., Chao, S.Z., 2004. Psychomotorstimulants and neuronal plasticity. Neuropharmacology 47,61–79.
Zahm, D.S., Brog, J.S., 1992. On the significance of subterritories inthe "accumbens" part of the rat ventral striatum. Neuroscience50, 751–767.
Zorrilla, E.P., 1997. Multiparous species present problems (andpossibilities) to developmentalists. Dev. Psychobiol. 30,141–150.
Zweifel, L.S., Argilli, E., Bonci, Zweifel, Palmiter, R.D., 2008. Roleof NMDA receptors in dopamine neurons for plasticity andaddictive behaviors. Neuron 59, 486–496.
Copyright © 2022 FDOKUMEN




















