
Drosha regulates neurogenesis by controlling Neurogenin 2 expression independent of microRNAs
Upload
independentCategory
view
1download
0
Statins Induce Angiogenesis, Neurogenesis,and Synaptogenesis after Stroke
Jieli Chen, MD,1 Zheng Gang Zhang, MD, PhD,1 Yi Li, MD,1 Ying Wang, MD,1 Lei Wang, MD,1
Hao Jiang, PhD,1 Chenling Zhang, BS,1 Mei Lu, PhD,2 Mark Katakowski, BS,1
Carolyn S. Feldkamp, PhD,3 and Michael Chopp, PhD1,4
We demonstrate that the 3-hydroxy-3-methyl-glutaryl-coenzyme A (HMG-CoA) reductase inhibitors atorvastatin andsimvastatin enhance functional outcome and induce brain plasticity when administered after stroke to rats. With ator-vastatin treatment initiated 1 day after stroke, animals exhibited significant increases in vascular endothelial growthfactor, cyclic guanosine monophosphate, angiogenesis, endogenous cell proliferation and neurogenesis, and an increase inthe synaptic protein, synaptophysin. Atorvastatin-induced angiogenesis in a tube formation assay was reduced by anantibody against the vascular endothelial growth factor receptor 2 (FIK-1) and by the nitric oxide synthase inhibitor,N-mono-methyl-L-arginine (L-NAME). Atorvastatin also induced phosphorylation of Akt and Erk in cultured primarycortical neurons. These data indicate that atorvastatin induced brain plasticity and has neurorestorative activity afterexperimental stroke.
Ann Neurol 2003;53:743–751
The 3-hydroxy-3-methyl-glutaryl-coenzyme A (HMG-CoA) reductase inhibitors (statins) reduce synthesis ofcholesterol and isoprenoids, molecules that modulatediverse cell function.1 Statin therapy has pleiotropiceffects, including protection against Alzheimer’s dis-ease and neuroprotection.2 Therapy with statins mod-ulates endothelial function and preserves blood flowto tissue exposed to an ischemic insult, by targetingthe vascular wall, inflammatory cells and platelets,and extravascular tissues (brain parenchyma) .2 Statinsalso induce angiogenesis in ischemic limbs of normo-cholesterolemic rabbits,3 upregulate endothelial nitricoxide synthase (NOS), and enhance cerebral bloodflow (CBF).2
Although statins have shown neuroprotective bene-fits in neurological injury and disease, there are nostudies to date that test the effects of statins on resto-ration of function after neural injury. In this study, wedemonstrate that widely used statins (atorvastatin andsimvastatin) when administered at 1 day after stroke ishighly effective in reducing neurological deficits in arat model of middle cerebral artery occlusion (MCAo).These benefits appear to be mediated by statin-induced
amplification of angiogenesis, neurogenesis, and synap-togenesis.
Materials and MethodsAnimal Middle Cerebral Artery Occlusion ModelAdult male Wistar rats weighing 270 to 300gm were used inall our experiments. We induced 2 hours of MCAo using amethod of intraluminal vascular occlusion modified in ourlaboratory.4
Experimental GroupsExperimental groups consist of rats subjected to 2 hours ofMCAo (n � 48). Twenty-four hours after MCAo, rats werefed atorvastatin (calcium, Pfizer, Groton, CT), dissolved insaline, 1mg/kg (n � 12), 3mg/kg (n � 12), or 8mg/kg (n �12) daily for 7 days. Rats subjected to MCAo and fed waterwithout atorvastatin served as a control group (n � 12). Allanimals received daily intraperitoneal injections of bromode-oxyuridine (BrdU; a thymidine analog, which labels newlysynthesized DNA, 100mg/kg; Sigma, St. Louis, MO) start-ing at 24 hours after MCAo and subsequently for 13 con-secutive days for identification of endothelial cell and endog-enous subventricular zone (SVZ) and dentate gyrus (DG)cell proliferation. All animals were killed at 14 days after
From the 1Department of Neurology, 2Biostatistics and ResearchEpidemiology, and 3Pathology Clinical Chemicology Division,Henry Ford Health Sciences Center, Detroit; and 4Department ofPhysics, Oakland University, Rochester, MI.
Received Oct 30, 2002, and in revised form Jan 16, 2003. Acceptedfor publication Jan 16, 2003.
Address correspondence to Dr Chopp, Neurology Research, E&RBuilding, Room 3056, Henry Ford Hospital, 2799 West GrandBoulevard, Detroit, MI 48202. E-mail: [email protected]
© 2003 Wiley-Liss, Inc. 743

MCAo. To test whether the effect of statin treatment ofstroke is specific to atorvastatin, we treated rats with simva-statin (1mg/kg, n � 8) using the identical treatment proto-col used for atorvastatin. Atrovastatin and simvastatin(Merck, Somerset, NJ) were purchased from Henry FordHospital pharmacy.
Functional TestsAdhesive-removal somatosensory test5,6 and a modified neu-rological severity score6,7 evaluation were performed beforeMCAo and at 1, 7, and 14 days after MCAo by an investi-gator who was blinded to the experimental groups.
Histological and Immunohistochemical AssessmentAt 14 days after MCAo, animal (n � 5/group) brains wereembedded in paraffin. Lesion volume was measured usingthe Global Lab Image analysis system (Data Translation,Malboro, MA) and presented as a volume percentage of thelesion compared with the contralateral hemisphere.6
IMMUNOHISTOCHEMISTRY. BrdU, von Willebrand factor(dilution, 1:400; Dako, Carpinteria, CA) and synaptophysin(monoclonal antibody, clone SY 38, 1:40; BoehringerMannheim Biochemica, Indianapolis, IN) immunostainingwere performed, as previously described.6,7 To determinewhether BrdU-immunoreactive cells express neural pheno-type, we used double immunohistochemical staining to iden-tify BrdU (1:100; Boehringer Mannheim, Indianapolis, IN)with the progenitor neuronal marker, TUJ1 (1:400, Mono-clonal Anti–B-Tubulin Isotype III, Sigma). Using double im-munohistochemical staining, we counted a total of 500BrdU-positive cells in the SVZ per animal to obtain the per-centage of BrdU-reactive cells colocalized with TUJ1.
QUANTIFICATION. BrdU-immunostained sections weredigitized using a �40 objective (Olympus BX40) via theMCID computer imaging analysis system (Imaging Research,St. Catharines, Ontario, Canada). Cells within a total of 20enlarged and thin-walled vessels located in the boundary areaof the ischemic lesion were counted in each section. BrdU-reactive cell numbers were counted in the SVZ and DG ofthe ischemic hemisphere. Five sections and eight views ineach section were counted per rat with the number of BrdU-reactive cells averaged. Data are presented as the number ofthe BrdU-immunoreactive cells per section.
For measurement of vascular density and perimeters,8 eachvWF-immunostained coronal section was digitized using a�20 objective via the MCID computer imaging analysis sys-tem. The numbers of vessels and their perimeters werecounted throughout each field of view. The total number ofvessels was divided by the total tissue area to determine vas-cular density.
For semiquantification of synaptophysin immunoreactiv-ity, five immunostained coronal sections and eight fields ofview from the ischemic penumbra (cortex and striatum) andcontralateral hemisphere in each section were digitized undera �20 objective. The optical density was measured. All val-ues of density are presented as a percentage compared withthe contralateral homologous region on the same section.9
Vascular Endothelial Growth Factor DetectionVascular endothelial growth factor detection (VEGF) wasmeasured using a VEGF ELISA kit (mouse VEGF Immuno-assay, catalog no. MIMV00, Quantikine M; R & D Systems,Minneapolis, MN).7,10 Brain extract from control MCAoand atorvastatin- (1mg/kg) treated animals (n � 6/group)was obtained from the ischemic boundary zone at 14 daysafter MCAo.
Capillary-like Tube Formation AssayFormation of capillary-like tubes in mouse brain–derived en-dothelial cells was measured.11,12 Mouse brain–derived mi-crovessel endothelial cells (2 � 104 cells) in 1.5ml cultureregular medium or atorvastatin were added to each dish.These cells were incubated in (1) regular cell culture medium(Dulbecco’s modified Eagle medium) for control; atorvasta-tin or (2) 0.01�M, or (3) 0.1�M, or (4) 0.5�M, or (5)1.0�M, or (6) 10�M, or (7) 0.5�M with 10�g/ml anti–mouse neutralizing antibody to VEGFR-2 (DC101; Im-Clone System, New York, NY); (8) 0.5�M atorvastatin with5mM NOS inhibitor N-mono-methyl-L-arginine (L-NAME,Sigma); (9) 0.5�M atorvastatin with 10�g/ml anti–mouseneutralizing antibody to VEGFR-2 (DC101; Imclone Sys-tem) and 5mM L-NAME. All assays were performed in trip-licate and quantitated.
Corneal Assay for AngiogenesisTo test effect of atorvastatin on induction of angiogenesis inan in vivo system, we performed a corneal assay for angio-genesis (n � 6). Mixed atorvastatin (2�l, 10�g) with 12%hydrion (2�l) was inserted 2mm away from the limbus mar-gin of the cornea of the right eye.13 Twelve percent hydrion(4�l) alone was inserted into left eye for control. Corneaswere digitized under a �2.5 objective for measurement ofvessel length extension (in millimeters) and density using avideo camera interfaced with MCID image analysis system.
Cyclic Guanosine Monophosphate Measurement inBrain TissueLevels of cyclic guanosine monophosphate (cGMP) in theipsilateral hemisphere of ischemic brain with (1mg/kg for 7days) or without atorvastatin treatment after stroke in ratswere measured (n � 6/group). The animals were killed at 7days after stroke. cGMP was measured using a commerciallyavailable low pH Immunoassay kit (R & D Systems).14 Val-ues for cGMP were standardized by total protein (micromo-lars per milligrams protein).
Signaling Pathway of Atorvastatin-treated CulturedCortical NeuronsPhosphorylated Akt and Erk were measured using Westernblot analysis.15 Cultured primary cortical neurons were pre-pared from embryonic day 17 Wistar rats and pretreatedwith or without the phosphatidylinositol 3� kinase (PI3K)inhibitor, LY294002 (20�M; Calbiochem, San Diego, CA),followed by treatment with 0.5�M of atorvastatin. Lysateswere probed with anti-phospho-Akt (Ser473, Cell SignalingTechnology, Beverly, MA) and anti-phospho-Erk1 and Erk2(Santa Cruz Biotechnology, Santa Cruz, CA) antibodies anddetected using a SuperSignal protein detection kit (Pierce,
744 Annals of Neurology Vol 53 No 6 June 2003
New York, NY) with an horseradish peroxidase–conjugatedsecondary antibody. Three independent experiments wereperformed. The intensity of the bands was measured usingScion image analysis (Scion Cooperation, Frederick, MD).
Cholesterol, Blood Pressure, and Cerebral Blood FlowMeasurementsSerum total cholesterol was quantified (n � 3/group). Sys-temic blood pressure (BP) (Protocol Systems; INS, Beaver-ton, OR; n � 3/group) and regional CBF (rCBF) was mea-sured using laser-Doppler flowmetery16 30 minutes beforetreatment to 3 hours after treatment with atorvastatin (n �3/group). The data are presented as a percentage of beforetreatment baseline values.
Statistical AnalysisGiven a lack of normality of the data, the generalized esti-mation equation approach was used for analysis of functionaltest scores and lesion volume. For each outcome, one-wayanalysis of variance or covariance was used to test the doseeffect adjusting for its pretreatment score and was followedby the pairwise dose comparisons at the 0.05 level, if a doseeffect was detected at the 0.05 level. Two-sample t test wasused to test for VEGF, neurogenesis, and angiogenesis be-tween groups with and without treatment of atorvastatin atthe significance level of 0.05. Data are presented as mean �standard error.
ResultsNeurological Outcome and Lesion VolumeAll animals survived elective surgery. Compared withthe control treated group, rats treated with 1 and3mg/kg atorvastatin and 1mg/kg simvastatin had sig-nificantly (p � 0.05) improved functional recovery,with no functional benefit detected with 8mg/kg ator-vastatin (Table). No significant differences of ischemiclesion volumes in atorvastatin and simvastatin-treatedgroups (29 � 7.0% for 1mg/kg atorvastatin; 28.1 �4.9% for 3mg/kg atorvastatin; 42.3 � 6.9% for8mg/kg atorvastatin; 29.4�4.0% for 1mg/kg simvasta-
tin) were detected compared with control treated rats(34.7 � 5.6 %), respectively.
Endothelial Cell Proliferation and AngiogenesisEnlarged and thin-walled vessels, containing BrdU-reactive endothelial cells indicative of angiogenesis,17
significantly increased in the ipsilateral hemisphere ofthe low-dose atorvastatin (1 and 3mg/kg) treatmentgroup (Fig 1c, d), but not in the high-dose atorvastatin(8mg/kg, see Fig 1d) compared with the ipsilateralhemisphere of control animals (see Fig 1b, d) or thecontralateral hemisphere of treated animals (see Fig1a). Treatment with low-dose (1 and 3mg/kg) but notin high-dose (8mg/kg) atorvastatin (see Fig 1e, f) sig-nificantly (p � 0.05) increased vascular perimeter anddensity in the ipsilateral hemisphere compared with thecontrol rats (see Fig 1e, f). These data indicate thatlow-dose atorvastatin enhances angiogenesis in isch-emic brain.
Atorvastatin Promotes Vascular Endothelial GrowthFactor Detection Secretion in the Ischemic BoundaryAreaWe tested whether atorvastatin evokes production ofVEGF, a principal angiogenic factor,18 in ischemicbrain. Using ELISA, we found that VEGF productionwithin the ischemic boundary area at 14 days afterstroke significantly increased in the low-dose atorvasta-tin (1mg/kg) treatment group (25.7 � 2.7ng/ml) com-pared with the MCAo control group (10.5 � 1.7ng/ml) and normal brain (13.4 � 1.5ng/ml, p � 0.05).
Atorvastatin Promotes Corneal NeovascularizationAs a complementary measurement of atorvastatin me-diated angiogenesis, we used a corneal angiogenesis as-say. Atorvastatin significantly augmented corneal neo-vascularization (see Fig 1h) compared with controlanimals (see Fig 1g). The total vascular lengths signif-
Table. Functional Tests: Treatment with or without Atorvastatin or Simvastatin after MCAo (n � 8/group, mean � SE)
MCAo
Adhesive-Removal Test (sec) NSS Test
Base 1 day 7 day 14 day Base 1 day 7 day 14 day
Control 7.1 � 0.6 98.8 � 8.9 73.0 � 12.9 63.6 � 12.0 0 � 0 9.9 � 0.4 8.6 � 1.5 7.8 � 1.61 mg Atorva-
statin7.3 � 0.5 108.7 � 7.9 41.3 � 8.8a 24.0 � 4.3a 0 � 0 10.3 � 0.6 6.8 � 0.4a 5.4 � 0.6a
3mg Atorva-statin
7.3 � 0.5 105.3 � 10 45.5 � 9.4a 22.5 � 5.8a 0 � 0 10.3 � 1.0 5.0 � 0.7a 4.0 � 0.5a
8 mg Atorva-statin
7.8 � 0.5 113.8 � 4.9 106.2 � 10 90.2 � 16.3 0 � 0 10.2 � 0.9 7.6 � 0.7 6.2 � 0.9
1mg Simva-statin
7.5 � 0.7 97.63 � 9.7 31.8 � 4.9a 19.9 � 3.7a 0 � 0 9.63 � 0.6 5.9 � 0.6a 4.0 � 0.4a
ap � 0.05 compared with control MCAo group.
MCAo � middle cerebral artery occlusion; SE � standard error; NSS � neurological severity score.
Chen et al: Statins Induce Brain Plasticity 745

icantly increased in the atorvastatin treatment group(29.9 � 7.9mm/cornea) compared with control ani-mals (1.5 � 1.0mm/cornea).
Vascular Endothelial Growth Factor MediatesAtorvastatin-induced Angiogenesis In VitroAtorvastatin induced mouse brain–derived endothelialcells to form a network of capillary-like structures withnumerous intercellular contacts (in 0.5�M atorvasta-tin, Fig 2c). Low concentrations of atorvastatin (0.01–1�M), but not a high concentration (10�M) signifi-cantly induced tube formation compared with control
medium (see Fig 2a). Total capillary tube length sig-nificantly increased in medium containing atorvastatin(0.5�M, see Fig 2a, c) compared with the control me-dium (see Fig 2a, b). Both anti–VEGF receptor 2(DC101; see Fig 2d, g) and NOS inhibitor L-NAME(see Fig 2e, g) significantly decreased atorvastatin-induced capillary tube formation compared with ator-vastatin alone (p � 0.05, see Fig 2c, g). The combina-tion of anti–VEGF receptor 2 (DC101) with L-NAMEsignificantly inhibited atorvastatin-induced capillary tubeformation (see Fig 2f, g) compared with atorvastatinalone (p � 0.05), and tube formation was significantly
Fig 1. Atorvastatin induces angiogenesis, as indicated by increased bromodeoxyuridine (BrdU)–immunoreactive endothelial cells, vascu-lar density, and perimeter. Panels a to c show BrdU immunoreactivity in the contralateral hemisphere (a), and ipsilateral hemispherein control MCAo representative rats (b) or in rats with MCAo and treated with 1mg/kg atorvastatin (c). BrdU-immunoreactive endo-thelial cells were detected in enlarged vessels (arrows in b and c). Panel d shows the percentage of BrdU-immunoreactive cells in vessels.Panels e and f show vascular perimeters (e) and vessel density in the ischemic boundary area treated with or without atorvastatin.Treatment with low-dose atorvastatin (1 and 3mg) significantly (p � 0.05) increased the number of BrdU-immunoreactive cells invessels (d), vascular perimeter (e), and density (f) in the ipsilateral hemisphere compared with control MCAo alone (n � 8/group).p value less than 0.05 compared with MCAo alone. Scale bar in a � 25�m. Panels g and h show angiogenesis in rat cornea treatedwith (h) or without (g) atorvastatin in vivo. Atorvastatin induces neovascularization in cornea (h).
746 Annals of Neurology Vol 53 No 6 June 2003

reduced compared with atorvastatin with L-NAME(p � 0.05). These data suggest that both VEGF andNO facilitate atorvastatin-induced angiogenesis.
Cell Proliferation in the Subventricular Zone andDentate GyrusBrdU-positive cells significantly increased in the ipsilat-eral DG (Fig 3b, c) and SVZ (see Fig 3e, f) in low-dose atorvastatin (1 and 3mg/kg) but not high-dose(8mg/kg, see Fig 3c, f) treated groups compared withthe control treatment group (see Fig 3a, c for DG, seeFig 3d–f for SVZ). Using confocal imaging, we foundthat double staining immunohistochemistry shows thatsome BrdU-positive cells (see Fig 3g and i) colocalizedwith the developmental neuronal marker TUJ1 (see Fig3h and i) in the low dose (1 and 3mg/kg) atorvastatintreatment group. The percentage of BrdU-reactive cellsthat express TUJ1 in the ipsilateral SVZ increased inlow-dose atorvastatin-treated animals (1 and 3mg/kg;25.4 � 4.5%, 21.4 � 2.1%, p � 0.05), but not inhigh-dose–treated animals (8mg/kg, 15 � 5.8%) com-pared with control group (14.4 � 6.2%).
Effects of Atorvastatin on Ischemic Brain CyclicGuanosine MonophosphatecGMP promotes neurogenesis.19 Statins act as NO do-nors20 and thereby may enhance NO-mediated in-
crease in cGMP. To test this hypothesis, we measuredcGMP in ischemic brain in both control andatorvastatin-treated rats at 8 days after stroke. Admin-istration of low-dose (1mg/kg) atorvastatin at 24 hoursafter MCAo for 7 days significantly increased cGMP(3.9 � 0.5�mol/mg protein, p � 0.05) compared withthe control group (2.3 � 0.1�mol/mg protein) in theipsilateral hemisphere.
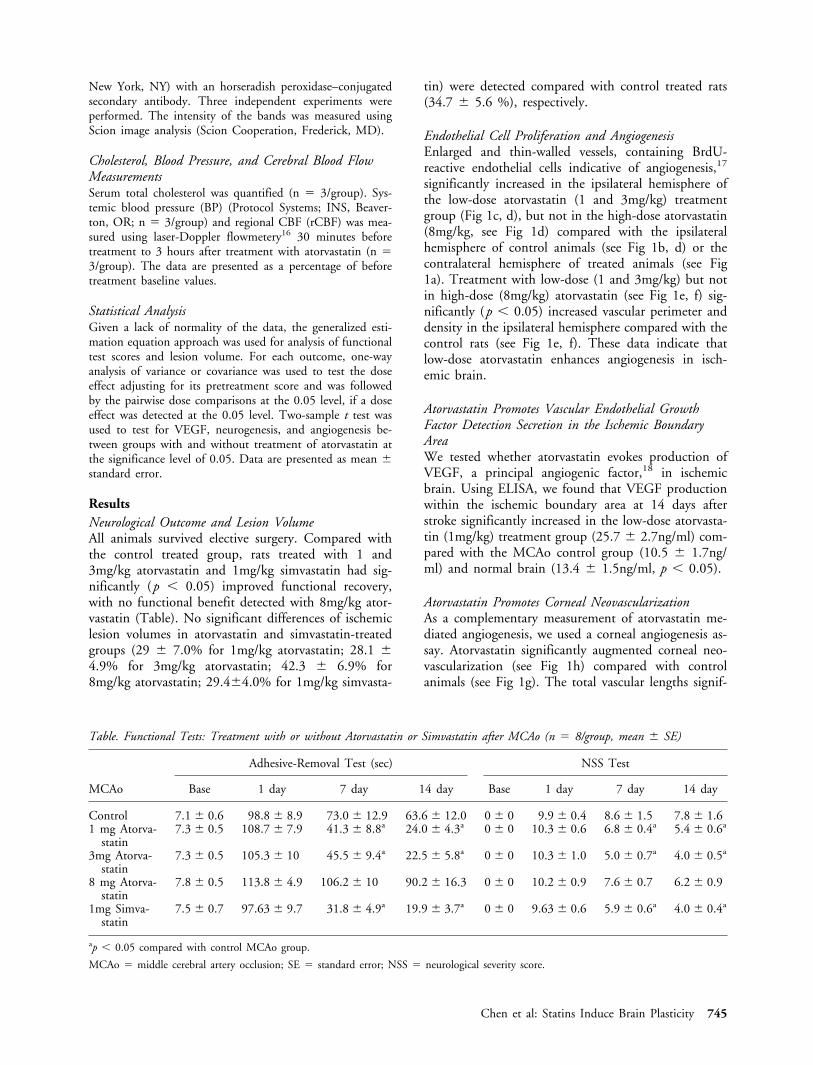
SynaptophysinFunctional benefit derived from treatment of strokewith atorvastatin suggests an effect of atorvastatin onsynaptic plasticity. Synaptophysin is a marker for pre-synaptic plasticity and synaptogenesis.21 Figure 4shows synaptophysin expression significantly increasedin the low-dose (see Fig 4b, c) but not in high-doseatorvastatin treatment rats compared with control rats(see Fig 4a, c).
Effect of Atorvastatin on Activation of Akt and Erkin Cultured Primary Cortical NeuronsStatins activate PI3K/Akt and Ras/Erk pathways.22
Phosphorylation of Akt and Erk transduces cell survivalsignals and promotes synaptic plasticity.23–25 We there-fore measured phosphorylation of Akt and Erk in pri-mary cortical neurons. Incubation of atorvastatin(0.5�M) in cultured primary cortical neurons mark-
Fig 2. Capillary-like tube formation. When mouse brain endothelial cells were incubated with a regular medium, capillary-like tubeformation was not detected (b; n � 3). However, incubation with low concentrations (0.01–1.0�M) but not high concentrationatorvastatin (10�M) significantly induced capillary-like tube formation (a, c, 0.5�M atorvastatin; n � 3). A rat anti–mouse neu-tralizing antibody to VEGFR2 (DC101) significantly inhibited atorvastatin-induced capillary-like tube formation (d, g; n � 3).L-NAME partially inhibited atorvastatin-induced capillary-like tube formation (e, g; n � 3). Capillary tube formation was mostinhibited by the combination of DC101 and L-NAME (f, g). Panels a and g show quantitative analysis of capillary-like tube for-mation. Scale bar in b � 100�m.
Chen et al: Statins Induce Brain Plasticity 747

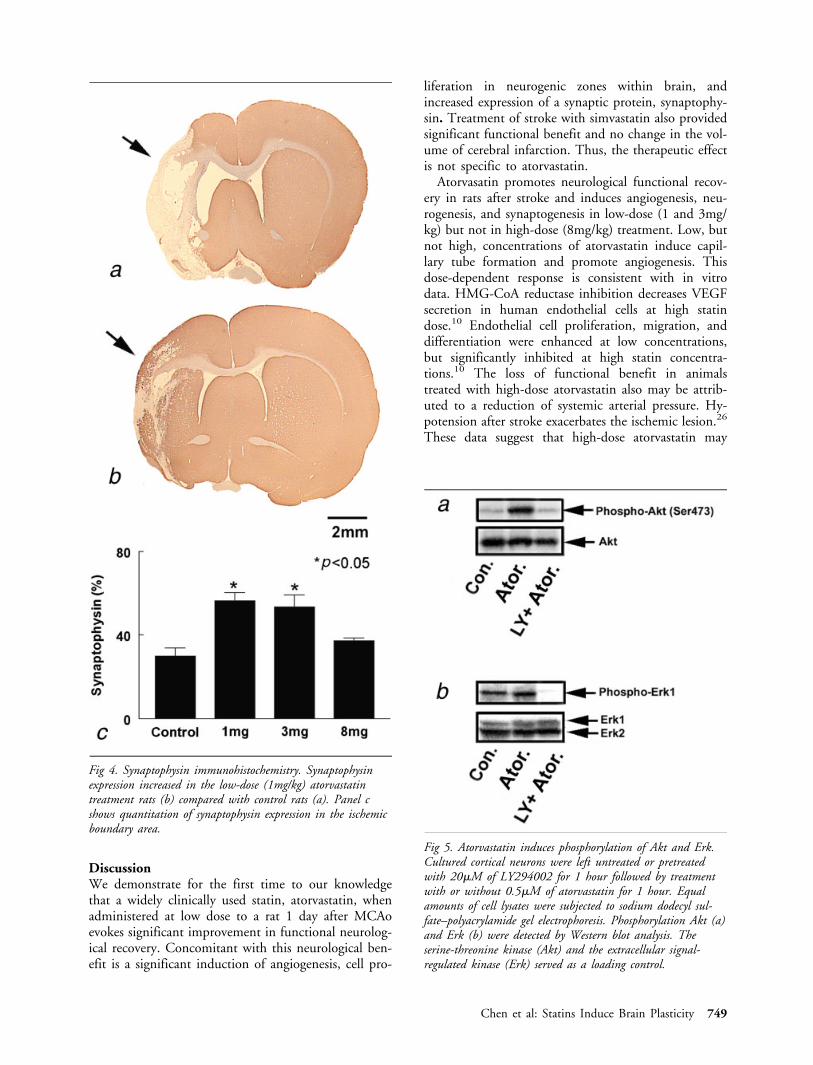
edly increased phosphorylation of Akt (265%, Fig 5a)and Erk (140%, see Fig 5b) compared with control(see Fig 5a, b). Pretreatment with LY294002, a stablePI3K inhibitor, significantly reduced atorvastatin-induced phosphorylation of Akt and Erk (75% and2.7%, respectively; see Fig 5a, b). These data indicatethat atorvastatin activates the PI3K/Akt and Erk path-ways in cortical neurons.
Serum Total Cholesterol, Blood Pressure, andRegional Cerebral Blood FlowSerum total cholesterol was not significantly differentin any group (1mg/kg, 66.3 � 4.9mg/dl; 3mg/kg,71 � 11.5mg/dl; 8mg/kg, 64 � 10.1mg/dl; and con-trol group, 76.4 � 9.4mg/dl). This suggests thatatorvastatin-induced brain plasticity is not dependenton the lowering of cholesterol. High-dose (8mg/kg)
treatment with atorvastatin caused a significant reduc-tion (p � 0.05) of systemic arterial BP levels at 1 hourafter treatment (51 � 10.6mm Hg) compared withlevels obtained before treatment (85.0 � 3.3mm Hg).BP was not decreased at 1 hour after low-dose (1 and3mg/kg) atorvastatin administration (78.3 � 4.7 and81.6 � 12.1mm Hg, respectively) compared with BPmeasured before treatment (84.3 � 4.0 and 76.3 �6.3mm Hg, respectively). rCBF significantly increasedin the ischemic hemisphere in low-dose atorvastatin(1mg/kg) treatment from 40 to 120 minutes after ator-vastatin administration (range, 165 � 22% to 130 �3.9%, p � 0.05) but not in high-dose (8mg/kg; range,86.2 � 9.4% to 115.3 � 15.1%) treatment comparedwith pretreatment levels. No significant differences inrCBF were detected 2 hours after atorvastatin treat-ment among all groups.
Fig 3. Atorvastatin induces neurogenesis. Panels a to e show bromodeoxyuridine (BrdU) immunoreactivity. Cell proliferation in thesubventricular zone (SVZ) and dentate gyrus of the ischemic hemisphere is identified by BrdU immunoreactivity. A few BrdU-immunoreactive cells in the dentate gyrus (a, arrow) and SVZ (d, arrow) were present in the middle cerebral artery occlusion(MCAo) control animal. Panels b and e show increased BrdU-reactive cells in the dentate gyrus (b, arrow) and SVZ (e, arrow) inatorvastatin- (1mg/kg) treated rat. Panels c and f show quantitative analysis of the number of BrdU-positive cells in the dentategyrus and SVZ is significantly increased in low-dose atorvastatin treatment (1 and 3mg/kg, p � 0.05), but not in the high-dose(8mg/kg) group compared with MCAo control. Panels g to i show confocal images of double immunostaining BrdU and TUJ1 inthe SVZ of atorvastatin- (1mg/kg) treated animals. BrdU-reactive cells (g, arrow) were TUJ1 immunoreactive (h, arrow), andpanel i shows colocalization of BrdU with TUJ1 immunoreactivity. Scale bars in a and g � 200�m and 10�m, respectively.
748 Annals of Neurology Vol 53 No 6 June 2003
DiscussionWe demonstrate for the first time to our knowledgethat a widely clinically used statin, atorvastatin, whenadministered at low dose to a rat 1 day after MCAoevokes significant improvement in functional neurolog-ical recovery. Concomitant with this neurological ben-efit is a significant induction of angiogenesis, cell pro-
liferation in neurogenic zones within brain, andincreased expression of a synaptic protein, synaptophy-sin. Treatment of stroke with simvastatin also providedsignificant functional benefit and no change in the vol-ume of cerebral infarction. Thus, the therapeutic effectis not specific to atorvastatin.
Atorvasatin promotes neurological functional recov-ery in rats after stroke and induces angiogenesis, neu-rogenesis, and synaptogenesis in low-dose (1 and 3mg/kg) but not in high-dose (8mg/kg) treatment. Low, butnot high, concentrations of atorvastatin induce capil-lary tube formation and promote angiogenesis. Thisdose-dependent response is consistent with in vitrodata. HMG-CoA reductase inhibition decreases VEGFsecretion in human endothelial cells at high statindose.10 Endothelial cell proliferation, migration, anddifferentiation were enhanced at low concentrations,but significantly inhibited at high statin concentra-tions.10 The loss of functional benefit in animalstreated with high-dose atorvastatin also may be attrib-uted to a reduction of systemic arterial pressure. Hy-potension after stroke exacerbates the ischemic lesion.26
These data suggest that high-dose atorvastatin may
Fig 5. Atorvastatin induces phosphorylation of Akt and Erk.Cultured cortical neurons were left untreated or pretreatedwith 20�M of LY294002 for 1 hour followed by treatmentwith or without 0.5�M of atorvastatin for 1 hour. Equalamounts of cell lysates were subjected to sodium dodecyl sul-fate–polyacrylamide gel electrophoresis. Phosphorylation Akt (a)and Erk (b) were detected by Western blot analysis. Theserine-threonine kinase (Akt) and the extracellular signal-regulated kinase (Erk) served as a loading control.
Fig 4. Synaptophysin immunohistochemistry. Synaptophysinexpression increased in the low-dose (1mg/kg) atorvastatintreatment rats (b) compared with control rats (a). Panel cshows quantitation of synaptophysin expression in the ischemicboundary area.
Chen et al: Statins Induce Brain Plasticity 749

have an adverse effect on the ischemic lesion and onfunctional recovery.
Increased angiogenesis promotes neurological func-tional recovery after stroke.17 Our data demonstratethat low-dose atorvastatin promotes endothelial cellproliferation and increased vessel perimeter and den-sity, induces angiogenesis, increases CBF, and improvesfunctional recovery. Statins may enhance recovery fromcerebral ischemia through beneficial modulation of thebrain vascular system and the promotion of angiogen-esis.
Administration of atorvastatin significantly increasedthe number of BrdU-immunoreactive cells in the SVZand DG, and some cells colocalized with TUJ1, sug-gesting that atorvastatin promotes neurogenesis in theadult brain after stroke. This enhanced neural progen-itor production may play a role in neurological func-tional recovery.27 However, we have not demonstratedthat these newly produced cells integrate into the cere-bral architecture and are electrically active. Therefore,the data associating neurogenesis with functional recov-ery should be the interpreted with caution.
Reduction of neurological deficits after stroke hasbeen attributed to extensive synaptic and functional re-organization.28 Treatment with low-dose atorvastatininitiated 24 hours after MCAo significantly promotessynaptophysin expression in the ischemic boundaryand thereby may improve functional outcome.
Activation of PI3K/Akt and Erk signaling in neuronsplays a central role in controlling synaptic plasticityand memory23 and may be implicated in atorvastatininduction of angiogenesis, neurogenesis, and synapto-genesis. Activated Akt differentiates PC12 cells, accel-erates motor axon regeneration in vivo,25 and increasesaxon caliber and branching in sensory neurons.24
p-Erk-2 fusion protein enters the nucleus and pro-motes axon elongation.24 We demonstrate that atorva-statin promotes p-Akt and p-Erk in primary corticalneurons. Phosphorylation of PI3K/Akt leads to post-transcriptional activation of endothelial NOS, whichpromotes NO production.29 NO activates soluble gua-nylyl cyclase leading to the formation of cGMP, as wehave found. In the central nervous system, NO/cGMPsignaling promotes synaptic plasticity, axonal out-growth and neurogenesis, and angiogenesis during de-velopment and also in the mature animal.30–33 NO/cGMP regulates expression of transcriptional genes andsome of these genes, such as the cAMP response el-ement–binding protein, are involved in cellular sur-vival, neurogenesis,31,32 differentiation, synaptogenesis,neurotransmitter release, and synaptic plasticity.33,34
Atorvastatin also promotes VEGF secretion in the isch-emic brain. VEGF is an angiogenic factor and stimu-lates axon outgrowth.35 The significant inhibition ofcapillary tube formation by anti–VEGFR2 antibodyand by L-NAME and the added inhibition of angio-
genesis by L-NAME in combination with anti-VEGFR2 indicate that angiogenesis is promoted byboth VEGF and NO with contributions from thePI3K/Akt pathway.36,37 Thus, the atorvastatin influ-enced PI3K/Akt/NO/cGMP and Ras/Erk pathwaysmay be related to angiogenesis, neurogenesis, brainplasticity, and synaptogenesis.
In summary, we demonstrate that treatment of ex-perimental stroke with low-dose atorvastatin at 24hours after stroke significantly improves functionaloutcome. This functional benefit may be related to en-hancement of angiogenesis, neurogenesis, and synapto-genesis after stroke.
This work was supported by NINDS grants (PO1 NS23393, M.C.;RO1 NS35504, M.C.; RO1HL64766, Z.G.Z.).
We thank Dr Z. Zhu for kindly providing anti–VEGFR2 antibody(DC101; Imclone Systems Incorporated, New York, NY), and C.Powers and C. Roberts for technical assistance.
References1. Istvan ES, Deisenhofer J. Structural mechanism for statin inhi-
bition of HMG-CoA reductase. Science 2001;292:1160–1164.2. Amin-Hanjani S, Stagliano NE, Yamada M, et al. Mevastatin,
an HMG-CoA reductase inhibitor, reduces stroke damage andupregulates endothelial nitric oxide synthase in mice. Stroke2001;32:980–986.
3. Kureishi Y, Luo Z, Shiojima I, et al. The HMG-CoA reductaseinhibitor simvastatin activates the protein kinase Akt and pro-motes angiogenesis in normocholesterolemic animals. Nat Med2000;6:1004–1010.
4. Chen H, Chopp M, Zhang ZG, Garcia JH. The effect of hy-pothermia on transient middle cerebral artery occlusion in therat. J Cereb Blood Flow Metab 1992;12:621–628.
5. Schallert T, Hernandez TD, Barth TM. Recovery of functionafter brain damage: severe and chronic disruption by diazepam.Brain Res 1986;379:104–111.
6. Chen J, Li Y, Wang L, et al. Therapeutic benefit of intravenousadministration of bone marrow stromal cells after cerebral isch-emia in rats. Stroke 2001;32:1005–1011.
7. Li Y, Chen J, Chen XG, et al. Human marrow stromal celltherapy for stroke in rat: neurotrophins and functional recovery.Neurology 2002;59:514–523.
8. Zhang ZG, Zhang L, Jiang Q, Chopp M. Bone marrow-derived endothelial progenitor cells participate in cerebral neo-vascularization after focal cerebral ischemia in the adult mouse.Circ Res 2002;90:284–288.
9. King DL, Arendash GW. Maintained synaptophysin immuno-reactivity in Tg2576 transgenic mice during aging: correlationswith cognitive impairment. Brain Res 2002;926:58–68.
10. Weis M, Heeschen C, Glassford AJ, Cooke JP. Statins havebiphasic effects on angiogenesis. Circulation 2002;105:739–745.
11. Haralabopoulos GC, Grant DS, Kleinman HK, Lelkes PI, et al.Inhibitors of basement membrane collagen synthesis preventendothelial cell alignment in matrigel in vitro and angiogenesisin vivo. Lab Invest 1994;71:575–582.
12. Witte L, Hicklin DJ, Zhu Z, et al. Monoclonal antibodies tar-geting the VEGF receptor-2 (Flk1/KDR) as an anti-angiogenictherapeutic strategy. Cancer Metastasis Rev 1998;17:155–161.
750 Annals of Neurology Vol 53 No 6 June 2003

13. Fournier GA, Lutty GA, Watt S, et al. A corneal micropocketassay for angiogenesis in the rat eye. Invest Ophthalmol Vis Sci1981;21:351–354.
14. Zhang R, Zhang L, Zhang Z, et al. A nitric oxide donor in-duces neurogenesis and reduces functional deficits after strokein rats. Ann Neurol 2001;50:602–611.
15. Monshipouri M, Jiang H, Lazarovici P. NGF stimulation of erkphosphorylation is impaired by a point mutation in the trans-membrane domain of trkA receptor. J Mol Neurosci 2000;14:69–76.
16. Zhang RL, Chopp M, Zhang ZG, et al. A rat model of focalembolic cerebral ischemia. Brain Res 1997;766:83–92.
17. Zhang ZG, Zhang L, Tsang W, et al. Correlation of VEGF andangiopoietin expression with disruption of blood-brain barrierand angiogenesis after focal cerebral ischemia. J Cereb BloodFlow Metab 2002;22:379–392
18. He H, Venema VJ, Gu X, et al.. Vascular endothelial growthfactor signals endothelial cell production of nitric oxide andprostacyclin through flk-1/KDR activation of c-Src. J BiolChem 1999;274:25130–25135.
19. Firestein BL, Bredt DS. Regulation of sensory neuron precursorproliferation by cyclic GMP-dependent protein kinase. J Neu-rochem 1998;71:1846–1853.
20. Laufs U, Endres M, Liao JK. Regulation of endothelial NOproduction by Rho GTPase. Med Klin 1999;94:211–218.
21. Ujike H, Takaki M, Kodama M, Kuroda S. Gene expressionrelated to synaptogenesis, neuritogenesis, and MAP kinase inbehavioral sensitization to psychostimulants. Ann N Y Acad Sci2002;965:55–67.
22. Urbich C, Dernbach E, Zeiher AM, Dimmeler S. Double-edged role of statins in angiogenesis signaling. Circ Res 2002;90:737–744.
23. Daw MI, Bortolotto ZA, Saulle E, et al. Phosphatidylinositol 3kinase regulates synapse specificity of hippocampal long-termdepression. Nat Neurosci 2002;5:835–836.
24. Markus A, Zhong J, Snider WD. Raf and akt mediate distinctaspects of sensory axon growth. Neuron 2002;35:65–76.
25. Namikawa K, Honma M, Abe K, et al.. Akt/protein kinase Bprevents injury-induced motoneuron death and accelerates ax-onal regeneration. J Neurosci 2000;15:2875–2886.
26. Zhu CZ, Auer RN. Graded hypotension and MCA occlusionduration: effect in transient focal ischemia. J Cereb Blood FlowMetab 1995;15:980–988.
27. van Praag H, Christie BR, Sejnowski TJ, Gage FH. Runningenhances neurogenesis, learning, and long-term potentiation inmice. Proc Natl Acad Sci USA 1999;96:13427–13431.
28. Gage FH, Coates PW, Palmer TD, et al. Survival and differ-entiation of adult neuronal progenitor cells transplanted to theadult brain. Proc Natl Acad Sci USA 1995;92:11879–11883.
29. Ciani E, Virgili M, Contestabile A. Akt pathway mediates acGMP-dependent survival role of nitric oxide in cerebellargranule neurones. J Neurochem 2002;81:218–228.
30. Truman JW, De Vente J, Ball EE. Nitric oxide-sensitive guan-ylate cyclase activity is associated with the maturational phase ofneuronal development in insects. Development 1996;122:3949–3958.
31. Nandagopal K, Dawson TM, Dawson VL. Critical role for nitricoxide signaling in cardiac and neuronal ischemic preconditioningand tolerance. J Pharmacol Exp Ther 2001;297:474–478.
32. Koyano-Nakagawa N, Wettstein D, Kintner C. Activation ofXenopus genes required for lateral inhibition and neuronal dif-ferentiation during primary neurogenesis. Mol Cell Neurosci1999;14:327–339.
33. Bredt DS, Snyder SH. Nitric oxide: a physiologic messengermolecule. Annu Rev Biochem 1994;63:175–195.
34. Contestabile A. Roles of NMDA receptor activity and nitricoxide production in brain development. Brain Res Brain ResRev 2000;32:476–509.
35. Sondell M, Sundler F, Kanje M. Vascular endothelial growth fac-tor is a neurotrophic factor which stimulates axonal outgrowththrough the flk-1 receptor. Eur J Neurosci 2000;12:4243–4254.
36. Gerber HP, McMurtrey A, Kowalski J, et al. Vascular endothe-lial growth factor regulates endothelial cell survival through thephosphatidylinositol 3�-kinase/Akt signal transduction pathway.Requirement for Flk-1/KDR activation. J Biol Chem 1998;273:30336–30343.
37. Suzuma K, Naruse K, Suzuma I, et al. Vascular endothelialgrowth factor induces expression of connective tissue growthfactor via KDR, Flt1, and phosphatidylinositol 3-kinase-akt-dependent pathways in retinal vascular cells. J Biol Chem 2000;275:40725–40731.
Chen et al: Statins Induce Brain Plasticity 751
Copyright © 2022 FDOKUMEN




















