
Neuropeptide Y overexpression using recombinant adenoassociated viral vectors
Upload
independentCategory
view
0download
0
Neuropeptide Y Promotes Neurogenesis in Murine SubventricularZone
FABIENNE AGASSE,a LILIANA BERNARDINO,a HEIDI KRISTIANSEN,b SØREN H. CHRISTIANSEN,b
RAQUEL FERREIRA,a BRUNO SILVA,a SOFIA GRADE,a DAVID P.D. WOLDBYE,b JOAO O. MALVAa
aNeuroprotection and Neurogenesis in Brain Repair Group, Center for Neuroscience and Cell Biology, Faculty ofMedicine, University of Coimbra, Coimbra, Portugal; bLaboratory of Neuropsychiatry, Department of Neuroscienceand Pharmacology, University of Copenhagen and Rigshospitalet University Hospital, Copenhagen, Denmark
Key Words. Subventricular zone • Calcium imaging • Neuronal differentiation • Functional binding • Neurogenesis • Neuropeptide Y
ABSTRACT
Stem cells of the subventricular zone (SVZ) represent areliable source of neurons for cell replacement. Neuropep-tide Y (NPY) promotes neurogenesis in the hippocampalsubgranular layer and the olfactory epithelium and may beuseful for the stimulation of SVZ dynamic in brain repairpurposes. We describe that NPY promotes SVZ neurogen-esis. NPY (1 �M) treatments increased proliferation at 48hours and neuronal differentiation at 7 days in SVZ cellcultures. NPY proneurogenic properties are mediated viathe Y1 receptor. Accordingly, Y1 receptor is a major active
NPY receptor in the mouse SVZ, as shown by functionalautoradiography. Moreover, short exposure to NPY in-creased immunoreactivity for the phosphorylated form ofextracellular signal-regulated kinase 1/2 in the nucleus, com-patible with a trigger for proliferation, whereas 6 hours oftreatment amplified the phosphorylated form of c-Jun-NH2-terminal kinase signal in growing axons, consistent withaxonogenesis. NPY, as a promoter of SVZ neurogenesis, is acrucial factor for future development of cell-based braintherapy. STEM CELLS 2008;26:1636–1645
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Constitutive neurogenesis takes place in the rodent adult mam-malian brain, and particularly in the subventricular zone (SVZ).SVZ harbors a population of stem cells that proliferate and giverise to neurons and glial cells [1, 2]. SVZ-derived neuroblastsmigrate long distances along the rostral migratory stream towardthe olfactory bulb, where they functionally differentiate intoGABA and dopaminergic interneurons, involved in odor dis-crimination [3, 4]. Interestingly, following cerebral injuries suchas ischemia, epileptogenesis, or focal neuronal degeneration,SVZ neurogenesis increases, and SVZ-derived neurons are ableto repopulate damaged areas [5–8]. Therefore, SVZ cells rep-resent a source of repairing cells with the potential to be re-cruited through endogenous stimulation of neurogenesis orgrafted following in vitro manipulation. Therefore, it is criticalto identify proneurogenic factors able to promote neuronal pro-genitor expansion and neuronal differentiation.
Neuropeptide Y (NPY), a 36-amino acid peptide abundantlyand widely distributed in the brain, regulates a wide range ofphysiological processes, including food intake, sexual behavior,and mood [9–12]. At the cellular level, NPY displays neuro-
protective properties in the striatum and the hippocampus [13–15] and modulates neuronal activity [16, 17], inhibiting hip-pocampal hyperactivity in epilepsy; it has therefore has beenproposed as an endogenous anticonvulsant [18–21].
Recently, NPY has been shown to promote neurogenesis inthe dentate gyrus (DG) of the hippocampus and the olfactoryepithelium (OE) of the rodent brain [22–26]. Indeed, in the DG,NPY, secreted by interneurons, promotes the proliferation ofneuronal progenitor cells and, thereby, the production of newgranule neurons [26]. In the OE, olfactory receptor neurons areconstantly regenerated from a population of basal cells thatretain stem cell characteristics [22, 27]. In this system, NPYlocally produced by sustentacular cells, another cell type com-posing the OE, has been shown to stimulate proliferation of thebasal cells [24]. In both systems, the proneurogenic effects havebeen shown to be mediated through Y1 receptor activation [24,26]. Together, these recent data convincingly demonstrate thatNPY supports neurogenesis in the DG and the OE. However,very little is known about the action of NPY on SVZ neurogen-esis.
In the present work we investigated the biological effects ofNPY on neurogenesis in SVZ. Our data clearly show that NPYis an inducer of neurogenesis in SVZ cells, providing new
Author contributions: F.A.: conception and design, provision of study material, collection and/or assembly of data, data analysis andinterpretation, manuscript writing, final approval of manuscript; L.B.: provision of study material, collection and/or assembly of data, dataanalysis and interpretation; H.K. and S.H.C.: collection and/or assembly of data, data analysis and interpretation; R.F., B.S., and S.G.:collection and/or assembly of data; D.P.D.W.: conception and design, financial support, data analysis and interpretation, manuscript writing,final approval of manuscript; J.O.M.: conception and design, financial support, data analysis and interpretation, administrative support,manuscript writing, final approval of manuscript.
Correspondence: Joao O. Malva, Ph.D., Center for Neuroscience and Cell Biology, Institute of Biochemistry, Faculty of Medicine, Universityof Coimbra, 3004-504 Coimbra, Portugal. Telephone: 351-239-112254; Fax: 351-239-822776; e-mail: [email protected] ReceivedJanuary 17, 2008; accepted for publication March 24, 2008; first published online in STEM CELLS EXPRESS April 3, 2008. ©AlphaMed Press1066-5099/2008/$30.00/0 doi: 10.1634/stemcells.2008-0056
TISSUE-SPECIFIC STEM CELLS
STEM CELLS 2008;26:1636–1645 www.StemCells.com

perspectives and molecular targets for the development of newtreatments for neurodegenerative diseases and brain injuries.
MATERIALS AND METHODS
All experiments were performed in accordance with NIH and Eu-ropean (86/609/EEC) guidelines for the care and use of laboratoryanimals.
SVZ CulturesSVZ cells were cultured from 1–3-day-old C57Bl/6 donor mice.Fragments of SVZ were dissected out from 450-�m-thick coronalbrain sections, digested in 0.025% trypsin and 0.265 mM EDTA(Gibco, Rockville, MD, http://www.invitrogen.com) in Hanks’ bal-anced saline solution (Gibco), and dissociated by gentle triturationwith a P1000 pipette. The cell suspension was diluted in serum-freeculture medium (SFM) composed of Dulbecco’s modified Eagle’smedium/Ham’s F-12 medium with GlutaMAX-I (Gibco) supple-mented with 100 U/ml penicillin, 100 �g/ml streptomycin (Gibco),1% B27 (Gibco), 10 ng/ml epidermal growth factor (Gibco), and 10ng/ml basic fibroblast growth factor (Gibco). Single cells were thenplated on uncoated Petri dishes at a density of 3,000 cells per cm2.The neurospheres were allowed to develop in a 95% air-5% CO2humidified atmosphere at 37°C.
Six- to 8-day-old neurospheres were adhered for 48 hours ontopoly-D-lysine-coated glass coverslips in SFM devoid of growthfactors. Then, the neurospheres were allowed to develop for 48hours or 7 days at 37°C in the absence or in the presence of either1 �M NPY, 1 �M Y1 receptor agonist ([Leu31, Pro34]-NPY), 300nM Y2 receptor agonist (NPY13–36), 1 �M Y5 receptor agonist(NPY [19–23]-[Gly1, Ser3, Gln4, Thr6, Ala31, Aib32, Gln34]-PP), or1 �M NPY with 1 �M Y1 receptor antagonist (BIBP3226) (all fromBachem AG, Bubendorf, Switzerland, http://www.bachem.com).
Single-Cell Calcium ImagingTo functionally characterize neuronal differentiation in SVZ cells,variations of intracellular calcium concentration ([Ca2�]i) followingstimulation with 50 mM KCl and 100 �M histamine (Sigma-Aldrich, St. Louis, http://www.sigmaaldrich.com) were analyzed.KCl depolarization causes the increase of [Ca2�]i in neurons,whereas stimulation with histamine increases [Ca2�]i in stem/pro-genitor cells [28].
SVZ cultures were loaded for 40 minutes at 37°C with 5 �MFura-2 AM (Molecular Probes, Eugene, OR, http://probes.invitrogen.com), 0.1% fatty acid-free bovine serum albumin (BSA),and 0.02% pluronic acid F-127, in Krebs buffer (132 mM NaCl, 1mM KCl, 1 mM MgCl2, 2.5 mM CaCl2, 10 mM glucose, 10 mMHEPES, pH 7.4). After a 10-minute postloading period at roomtemperature, the coverslip was mounted on RC-20 chamber in aPH3 platform (Warner Instruments, Hamden, CT, http://www.warneronline.com) on the stage of an inverted Axiovert 200 fluo-rescence microscope (Carl Zeiss, Gottingen, Germany, http://www.zeiss.com). Cells (approximately 100 cells per field) werecontinuously perfused with Krebs and stimulated by applying 100�M histamine or high-potassium Krebs solution (containing 50 mMKCl, isosmotic substitution with NaCl) by the mean of a fast-pressurized (95% air, 5% CO2 atmosphere) system (AutoMateScientific Inc., Berkeley, CA, http://www.autom8.com). [Ca2�]iwas evaluated by quantifying the ratio of the fluorescence emitted at510 nm following alternate excitation (750 milliseconds) at 340 and380 nm, using a Lambda DG4 apparatus (Sutter Instrument, Novato,CA, http://www.sutter.com) and a 510-nm band-pass filter (CarlZeiss) before fluorescence acquisition with a �40 objective and aCoolSNAP digital camera (Roper Scientific, Tucson, AZ, http://www.roperscientific.com). Acquired values were processed usingthe MetaFluor software (Universal Imaging Corporation, Marlow,U.K., http://www.universal-imaging.co.uk). Histamine/KCl valuesfor Fura-2 ratio were calculated to determine the extent of neuronalmaturation in cultures. The percentage of cells displaying a neuro-nal-like profile was calculated on the basis of the histamine (Hist)/KCl ratio.
ImmunocytochemistryCells were fixed for 30 minutes in 4% paraformaldehyde in phos-phate-buffered saline (PBS); permeabilized in 0.25% Triton X-100(Sigma-Aldrich); incubated overnight with the following primaryantibodies: mouse monoclonal anti-microtubule-associated pro-tein-2 (anti-MAP-2) antibody (1:200; Sigma-Aldrich), mousemonoclonal anti-neuronal nuclear protein (anti-NeuN) antibody (1:100; Chemicon, Temecula, CA, http://www.chemicon.com), rabbitpolyclonal anti-phosphorylated form of stress-activated protein ki-nase (anti-P-SAPK)/c-Jun-NH2-terminal kinase (JNK) (1:100), rab-bit monoclonal anti-phosphorylated form of extracellular signal-regulated kinase 1/2 (anti-P-ERK1/2) (1:50), or mouse monoclonalanti-tau (1:500) (all from Cell Signaling Technology, Danvers, MA,http://www.cellsignal.com); and incubated for 1 hour at room tem-perature with the appropriate secondary antibodies: anti-rabbit IgGlabeled with Alexa Fluor 488 or Alexa Fluor 594 (1:200) or anti-mouse IgG labeled with Alexa Fluor 594 (1:200) (all from Molec-ular Probes). Nuclei were counterstained with Hoechst 33342 (2�g/ml in PBS containing 0.25% BSA; Molecular Probes). Prepara-tions were mounted in DakoCytomation fluorescent medium(DakoCytomation, Carpinteria, CA, http://www.dakocytomation.com).
Proliferation Assay5-Bromo-2�-deoxyuridine (BrdU; 10 �M; Sigma-Aldrich) wasadded in the last 4 hours of the culture session. BrdU was unmaskedand labeled following successive passages in 1% Triton X-100,ice-cold 0.1 M HCl, and 2 M HCl at 37°C and in borate buffer (0.1M Na2B4O7�10H2O, pH 8.5) and incubation with the primary ratmonoclonal anti-BrdU antibody (1:50; Oxford Biotechnology, Ra-leigh, NC, http://www.immunologicalsdirect.com/index) overnightat 4°C and with the secondary anti-rat IgG labeled with Alexa Fluor594 (1:200; Molecular Probes) for 1 hour at room temperature (RT).Nuclei counterstaining and mounting were performed as describedpreviously.
Apoptosis AssayThe terminal deoxynucleotidyl transferase dUTP nick-end labeling(TUNEL) method was used to detect apoptotic nuclei. Briefly,permeabilized cells were subsequently incubated in terminal deoxy-nucleotidyl transferase buffer (0.25 U/�l terminal transferase, 6 �Mbiotinylated dUTP, pH 7.5; Boehringer Mannheim, Mannheim,Germany, http://www.boehringer.com) for 1 hour 30 minutes at37°C, in 300 mM NaCl and 30 mM sodium citrate buffer for 15minutes, in avidin-biotin-peroxidase complex for 30 minutes (1:100; Vector Laboratories, Burlingame, CA, http://www.vectorlabs.com), and in 30 mM Tris-HCl (pH 7.6) buffer containing diamino-benzidine chromogen (0.025%; Sigma-Aldrich), 0.08% NiCl2, and0.003% H2O2. The preparations were mounted in Depex (BDH,Poole, U.K., http://pt.vwr.com/app/Home). Transmission imageswere recorded using a digital camera coupled to an Axioskopmicroscope (Carl Zeiss).
Quantitative Polymerase Chain Reaction AnalysisTotal cellular RNA from SVZ explants and SVZ neurospheres wasextracted using Trizol reagent (Invitrogen, Carlsbad, CA, http://www.invitrogen.com) according to the manufacturer’s instructions.Purified RNA was treated with DNA-Free (Ambion, Huntingdon,U.K., http://www.ambion.com). Total RNA (1 �g) was reversetranscribed using TaqMan RT master mix (Applied Biosystems,Naerum, Denmark, http://www.appliedbiosystems.com) and ran-dom hexamers in a 100-�l reaction on a PTC-200 DNA engineThermal Cycler (VWR International, Albertslund, Denmark). Rel-ative quantification of the cDNA was performed in 96-well plateson the iCycler (Bio-Rad, Grenaa, Denmark, http://www.bio-rad.com) programmed for 2 minutes at 50°C, 3 minutes at 95°C, and 40cycles of 15 seconds at 95°C and 1 minute at 60°C. Each wellcontained 2 �l of cDNA mixed with 10 �l of SYBR Green I mastermix (Bio-Rad), 4.8 �l of water, 0.2 �l of uracil-N-glycosylase (0.1U/�l; Epicentre Biotechnologies, Madison, WI, http://www.epibio.com), and 750 nM forward and reverse primers (MWG Biotech,Ebersberg, Germany, http://www.mwg-biotech.com). All samples
1637Agasse, Bernardino, Kristiansen et al.
www.StemCells.com

were run in duplicate. Collected data were analyzed using theiCycler software, and a cycle threshold (Ct) for each sample wasdetermined. Relative quantification was achieved by subtractingeach Ct sample with the in-plate Ct of the reference control geneglyceraldehyde-3-phosphate dehydrogenase (GAPDH). Using the�-� method, we also determined fold mRNA expression of Y2, Y5,and NPY compared with Y1 mRNA. Regulations were verifiedusing RNA polymerase II as a reference gene. A group of referencegenes was tested, and GAPDH and RNA polymerase II were foundto be the least regulated genes. Water controls and genomic DNAcontrols were run simultaneously with the samples on all plates anddid not differ significantly from background. The following primersequences were used: Y1 (forward, GCT TAT GGG GCG GTGATT ATT CT; reverse, ACC GCA ACG AGC AAG TCT GAG),Y2 (forward, AAG TGG CCT GGG GAA GAG AAG AGT;reverse, GAG GCA AAA CGT ACA GGA TGA GCA), Y5 (for-ward, CAA CCT GGC CTT CTC CGA CAT; reverse, GCT TTGCCG AAC ATC CAC TGA), NPY (forward, CTC TGC GAC ACTACA TCA ATC TCA TCA; reverse, GGG CGT TTT CTG TGCTTT CCT T), GAPDH (forward, TGC ACC ACC AAC TGC TTAG; reverse, GGA TGC AGG GAT GAT GTT C), and RNA poly-merase II (forward, TGC GCA CCA CGT CCA ATG ATA; re-verse, GGA GCG CCA AAT GCC GAT AA). The efficiency of allprimers was between 90% and 110%.
Y1 Agonist-Stimulated [35S]-GTP�S FunctionalBindingThe functional binding method was described previously [19, 29].Brains of six naıve adult C57Bl/6 mice (25–35 g; Taconic, LilleSkensved, Denmark, http://www.taconic.com) were quickly re-moved following decapitation, frozen in cold isopentane, and storedat �80°C until use. The brains were coronally sliced, using acryostat (Thermo Shandon Inc., Pittsburgh, http://www.thermo.com), at 15 �m at the level of the SVZ: �0.98 to �0.02 mm relativeto bregma [30]. Sections were incubated in assay buffer A (50 mMTris-HCl, 3 mM MgCl2, 0.2 mM EGTA, 100 mM NaCl, pH 7.4) for10 minutes at RT and preincubated in assay buffer B (assay bufferA � 0.2 mM dithiothreitol, 1 �M 1,3-dipropyl-8-cyclopentyl-xan-thine [no. C-101, PerkinElmer Life and Analytical Sciences,Waltham, MA, http://www.perkinelmer.com], 0.5% wt/vol BSA,and 2 mM GDP [no. G7127; Sigma-Aldrich]) for 20 minutes at RTto shift all G-proteins into the inactivated state. Subsequently,incubation was performed in assay buffer B � 40 pM [35S]-GTP�S(1,250 Ci/mmol; NEG030H250UC; PerkinElmer) for 2 hours at25°C with 10�5 M human [Leu31, Pro34]-NPY (Bachem), a Y1receptor agonist. Basal and nonspecific binding were determined byincubation in assay buffer B � 40 pM [35S]-GTP�S and in buffer B� 40 pM [35S]-GTP�S � 10 �M nonlabeled GTP�S (no. 89378;Sigma-Aldrich), respectively. Specificity of the Y1 binding wasconfirmed by adding 10�6 M BIBP3226 (no. E3620; Bachem)during the preincubation and incubation. Incubation was terminatedby washing twice for 5 minutes in ice-cold 50 mM Tris-HCl buffer(pH 7.4). After a brief rinse in deionized H2O, sections were driedunder a cooled steam of air and exposed to Kodak Biomax MRfilms (Kodak, Rochester, NY, http://www.kodak.com) together with14C-microscales (GE Healthcare, Hillerød, Denmark, http://www.gehealthcare.com) for 5 days at �20°C. The films were developedin Kodak GBX developer. Finally, the films were digitalized usinga COHU, high-performance charge-coupled device camera, andquantifications were performed measuring the optical density usingcomputer-assisted image analysis (Scion Image, NIH) calibrated tothe 14C-microscale (nCi/g equivalent tissue). Means of absolutevalues in nCi/g minus basal binding values were determined.
Data AnalysisFluorescence images were recorded using an LSM 510 Meta con-focal microscope or an Axioskop 2 Plus fluorescence microscope(both from Carl Zeiss). In all the in vitro experiments, measure-ments were performed at the border of the neurospheres, wheremigrating cells emerged, forming a dense cell monolayer. In all invitro experiments, each experimental condition was assayed in threedifferent wells. Except where otherwise specified, the experimentswere replicated three times. Percentages of BrdU-, TUNEL-,
P-ERK1/2- (at 1 hour), and NeuN-immunoreactive cells were de-rived from cells counted in five independent microscopy fields ineach coverslip with a �40 objective (approximately 200 cells perfield). Quantifications of P-ERK1/2- and P-SAPK/JNK-positivenuclei at both 5 and 15 minutes were done in two independentcultures in at least 20 nonoverlapping fields (magnification, �40).Measurements of total length (micrometers) of the ramifications andquantification of the number of ramifications per neurosphere weredone in approximately 20 nonoverlapping fields in each coverslipusing digital images (magnification, �20) (n � 3 coverslips fromthree different cultures). In single-cell calcium imaging experi-ments, the percentage of neuron-like cells was calculated in amicroscopy field containing approximately 100 cells in each cov-erslip and using a �40 objective. Statistical significance was deter-mined using two-tailed Student’s t test or one-way analysis ofvariance followed by Bonferroni-corrected post hoc t test for mul-tiple comparisons. All data are presented as means � SEM. Statis-tical significance level was set for p values �.05.
RESULTS
NPY Promotes SVZ Cell Proliferation and ExertsNo Effect on Cell DeathTo investigate the effect of NPY on proliferation, SVZ neuro-spheres derived from newborn mice were treated for 48 hoursand 7 days in the absence (control condition) or presence of 1�M NPY. The thymidine analogue BrdU that incorporates inDNA in S-phase of the cell cycle was added for the last 4 hoursof both culture sessions. Nuclei were then immunostained forBrdU, as shown in Figure 1A. NPY induced a significantincrease in the percentage of BrdU-positive cells compared withcontrol after 48 hours (control 48 hours, 6.0% � 0.7%; NPY 48hours, 9.1% � 0.7%; p � .01; Fig. 1B). The effect was notmaintained after 7 days (control 7 days, 4.8% � 0.6%; NPY 7days, 3.9% � 0.3%), probably because of a shift from prolifer-ation to neuronal differentiation (described below).
The effect of NPY on cell death was evaluated after 48hours and 7 days of treatment with 1 �M NPY. Apoptotic nucleiwere darkly stained by the TUNEL method (Fig. 1C). Nosignificant differences in the numbers of TUNEL-positive nu-clei were found at either 48 hours or 7 days, indicating that NPYis not toxic to the cells (control 48 hours, 10.6% � 1.1%; NPY48 hours, 9.2% � 1.0%, Fig. 1D; control 7 days, 5.9% � 0.9%;NPY 7 days, 6.7% � 0.9%).
Y1, Y2, and Y5 Receptors and NPY mRNAs AreDetected in the SVZTo determine the expression of Y1, Y2, and Y5 receptors in theSVZ, quantitative polymerase chain reactions (q-PCRs) wereperformed in cDNAs from both neonatal SVZ neurospheres andadult SVZ explants. The Ct (i.e., the number of cycles necessaryfor a gene to be linearly expanded) was determined for eachgene and normalized by subtracting each Ct sample from in-plate Ct of the housekeeping reference gene GAPDH. Thesmaller the Ct value the more the gene is expressed comparedwith in-plate tested genes.
In SVZ neonatal cultures and adult SVZ explants, the Y1gene was found to be expressed at higher levels than Y5 and Y2,respectively (in neonatal cultures, normalized CtY1, 13.7 � 0.2;normalized CtY5, 17.5 � 0.6; normalized CtY2, 17.8 � 0.4; inadult SVZ explants, normalized CtY1, 7.2 � 0.7; normalizedCtY5, 8.5 � 0.6; normalized CtY2, 9.3 � 0.5; Fig. 2A, 2B). NPYmRNA was also detected in both SVZ cultures and explants(normalized CtNPY, 15.0 � 0.5 and 3.1 � 0.6, respectively; Fig.2A, 2B).
1638 NPY Promotes SVZ Neurogenesis

NPY Induces Neuronal Differentiation Through Y1ActivationThe effect of NPY on neuronal cell differentiation was assessedon SVZ cells maintained for 7 days in the presence or absenceof 1 �M NPY and processed for MAP-2 and NeuN immuno-cytochemistry. A denser network of ramified MAP-2-positiveneurons was observed in SVZ cultures treated with NPY com-pared with control conditions (Fig. 3A, 3B). In the presence ofNPY, a significant increase in the number of NeuN-positiveneurons was observed compared with control (control, 9.1% �1.6%; NPY, 24.5% � 1.9%; p � .0001), indicating that NPYinduces neuronal differentiation (Fig. 3C–3E).
To assess functional neuronal differentiation, we measured[Ca2�]i variations in single cells, following KCl and histaminestimulation. Briefly, this method is based on the profiles of[Ca2�]i variations, according to cell type, following KCl depo-larization and stimulation with histamine [28]. KCl depolariza-tion leads to the massive entry of Ca2� through voltage-depen-dent calcium channels and is used as a measure of neuronaldifferentiation, since differentiated neurons robustly expressthese channels [31, 32]. Histamine perfusion increases [Ca2�]i
in SVZ immature cells but not in neurons or in glial cells [28,33]. We previously showed that Hist/KCl ratios in mature neu-
rons are low (below 0.8), being characteristic of SVZ-derivedneuronal-like cells [28].
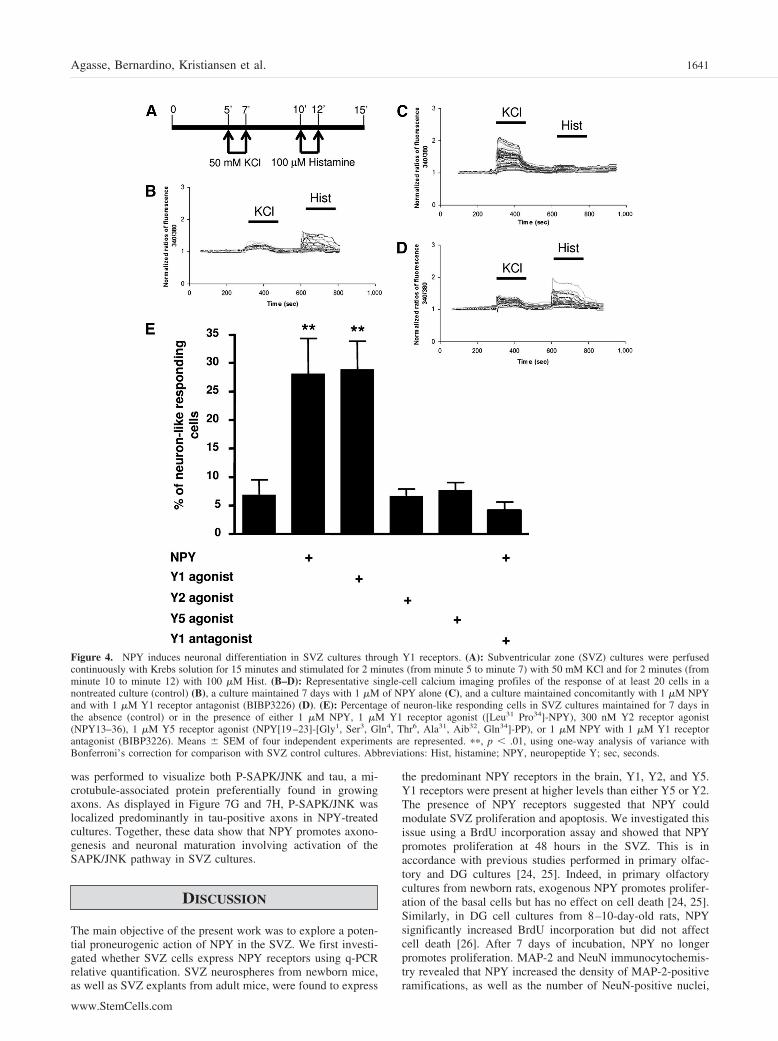
SVZ cell cultures were loaded with the Fura-2AM calciumprobe, perfused continuously for 15 minutes with Krebs solu-tion, and subsequently stimulated for 2 minutes with 50 mMKCl and with 100 �M histamine, as shown in Figure 4A. Figure4B–4D represents characteristic profiles of fluorescence recordsdisplayed by at least 20 cells of control, NPY-treated, andNPY/BIBP3226-treated cultures (Fig. 4B–4D, respectively). Incontrol cultures, most of the cells responded to histamine but notto KCl, whereas in NPY-treated cultures, the scenario wascompletely different (Fig. 4B, 4C, respectively). Indeed, follow-ing 7 days of treatment, the percentage of neuronal-like cellsincreased significantly compared with control cultures, suggest-ing a proneurogenic effect of NPY (control, 6.8% � 2.8%;NPY, 27.8% � 6.4%; p � .01; Fig. 4E).
To determine which receptor mediates the proneurogeniceffect of NPY, SVZ cells were treated for 7 days with either 1�M Y1 receptor agonist ([Leu31, Pro34]-NPY), 300 nM Y2receptor agonist (NPY13–36), 1 �M Y5 receptor agonist (NPY[19–23]-[Gly1, Ser3, Gln4, Thr6, Ala31, Aib32, Gln34]-PP), or 1�M NPY in combination with 1 �M Y1 receptor antagonist(BIBP3226). The same single-cell calcium experiments were
Figure 1. NPY promotes proliferation in subventricular zone (SVZ)culture but exerts no effect on cell death. (A): Representative confocalphotos of SVZ cell nuclei in SVZ cell cultures maintained for 48 hoursin the absence (control) or in the presence of 1 �M NPY and immuno-labeled for BrdU (red nuclei). (B): Percentage of BrdU-immunostainednuclei. (C): Representative transmission photography of SVZ cell nucleiin a control SVZ culture stained using the Tunel method (dark nuclei).(D): Percentages of Tunel-stained nuclei in cultures maintained for 48hours in the absence (control) or the presence of 1 �M NPY. (A, C):Mean percentages � SEM of two independent experiments are repre-sented. ��, p � .01, using analysis of variance with Bonferroni’scorrection for comparison with SVZ control cultures exposed to normalmedium for 48 hours. Abbreviations: BrdU, 5-bromo-2�-deoxyuridine;NPY, neuropeptide Y; Tunel, terminal deoxynucleotidyl transferasedUTP nick-end labeling.
Figure 2. Y1R, Y2R, and Y5R cDNA relative quantification in sub-ventricular zone (SVZ). (A): Y1R, Y2R, and Y5R cDNA relativequantification in SVZ neonatal cultures. Ct values are means � SEM ofquantitative polymerase chain reactions (q-PCRs) in four different8–10-day-old SVZ cultures. Fold expression compared with Y1 isshown as the inset. (B): Y1R, Y2R, and Y5R cDNA relative quantifi-cation in SVZ explants from adult mice. Ct values are means � SEM ofq-PCRs in at least eight mice for each receptor. All samples were run induplicate. Fold expression compared with Y1 is shown as the inset.Reference gene cDNA: glyceraldehyde-3-phosphate dehydrogenase(GAPDH). Normalization was achieved by subtracting Ct of GAPDHfrom Ct of target gene. Abbreviations: Ct, cycle threshold; NPY, neu-ropeptide Y; R, receptor.
1639Agasse, Bernardino, Kristiansen et al.
www.StemCells.com

then conducted. Incubation with the Y1 agonist but neither withY2 nor with Y5 agonists resulted in an increase in the percent-age of neuronal-like cells of similar magnitude as culturestreated with NPY (control, 6.8% � 2.8%; Y1 agonist, 28.7% �5.1%; p � .01; Y2 agonist, 6.6% � 1.3%; Y5 agonist, 7.6% �1.4%; Fig. 4E). As further evidence for a Y1 receptor-mediatedmechanism, interestingly, the general profiles determined incultures treated with both NPY and BIBP3226 were similar tothose of the control condition (Fig. 4D). Accordingly, coincu-bation of NPY with the Y1 antagonist (BIBP3226) abolished thecapacity of NPY in promoting neuronal differentiation (NPY �Y1 antagonist, 4.1% � 1.5%; Fig. 4C, 4D). As a control,profiles displayed by cells incubated with BIBP3226 alone weresimilar to the control condition (data not shown). Moreover, theY2 receptor antagonist BIIE0246 did not prevent the proneuro-genic effect of NPY (data not shown). Taken together, thesedata indicate a proneurogenic role for NPY mediated via Y1receptors in SVZ cells.
Evidence for Functional In Vivo Y1 Activity in theSVZDetection of Y1 receptor mRNA in adult SVZ explants (Fig. 2)together with the results obtained in vitro indicated that, in vivo,this receptor is present and functionally active in the SVZ.Evaluation of Y1 receptor function in the SVZ was performed invivo, in adult mice brain slices, using [Leu31, Pro34]-NPY-stimulated [35S]-GTP�S binding, a method that allows the de-tection of G-protein-coupled NPY receptor activation [19, 29].
Figure 5 shows pseudocolored autoradiograms of unstimu-lated (basal binding; Fig. 5A) and [Leu31, Pro34]-NPY-stimu-lated (Fig. 5B) mouse brain slices, nonspecific binding (Fig.
5C), and after specific blocking of [Leu31, Pro34]-NPY-stimu-lated [35S]-GTP�S binding by the Y1 antagonist BIBP3226(Fig. 5D). A significant increase in [Leu31, Pro34]-NPY-stimu-lated [35S]-GTP�S binding compared with basal binding wasfound in the SVZ ([Leu31, Pro34]-NPY-stimulated, 222.6 � 5.0nCi/g; basal, 194.7 � 2.8 nCi/g; p � .0005). For comparison,high levels of [Leu31, Pro34]-NPY-stimulated [35S]-GTP�Sbinding were observed in the same slices in the neocortex(motor cortex, M1, layers I–III), consistent with a previousstudy [34] ([Leu31, Pro34]-NPY-stimulated, 317.6 � 18.9 nCi/g;basal, 188.7 � 7.3 nCi/g; p � .0027). To test the specificity ofthe [Leu31, Pro34]-NPY-stimulated [35S]-GTP�S binding, sliceswere also coincubated with the Y1 antagonist BIBP3226. Meansof absolute values of radioactivity minus basal binding werecalculated. BIBP3226 incubation completely abolished [Leu31,Pro34]-NPY-stimulated [35S]-GTP�S binding ([Leu31, Pro34]-NPY-stimulated, 27.9 � 3.5 nCi/g; [Leu31, Pro34]-NPY-stimu-lated � BIBP3226, 2.0 � 2.6 nCi/g; p � .0018; Figure 5).Likewise, in the neocortex, an area known to contain high levelsof functional Y1 receptors, BIBP3226 almost completelyblocked [Leu31, Pro34]-NPY-stimulated [35S]-GTP�S binding inthe same slices ([Leu31, Pro34]-NPY-stimulated, 128.9 � 23.3nCi/g; [Leu31, Pro34]-NPY-stimulated � BIBP3226, 26.3 �13.4 nCi/g; p � .0029). Taken together, these results show thatthe SVZ neurogenic niche contains functional Y1 receptors.
NPY Activates Extracellular Signal-RegulatedKinase and Stress-Activated Protein Kinase/JNKPathways in SVZ CulturesExtracellular signal-regulated kinase 1/2 (ERK1/2) and stress-activated protein kinase (SAPK)/JNK are MAPKs involved inproliferation and neuronal differentiation. To investigatewhether NPY activates the ERK1/2 and SAPK/JNK signalingpathways, SVZ cell cultures were exposed to 1 �M NPY for 5minutes, 15 minutes, 1 hour, or 6 hours. Cells were processedfor immunocytochemistry against the phosphorylated (acti-vated) forms of ERK1/2 and SAPK/JNK kinases (i.e.,P-ERK1/2 and P-SAPK/JNK). Following 5 minutes of incuba-tion with 1 �M of NPY, the number of P-ERK1/2-positivenuclei per neurosphere was significantly increased comparedwith control (control, 7.0 � 1.4; NPY, 33.4 � 1.1; p � .0043)(Fig. 6A, 6B). Following 1 hour of NPY treatment, P-ERK1/2immunoreactivity was no longer found in nuclei but in thecytoplasm (Fig. 6A). Percentage of P-ERK1/2-positive cellsincreased significantly in NPY-treated cultures compared withcontrol (control, 9.3% � 1.8%; NPY, 17.2% � 3.0%; p � .047;Fig. 6C).
Similarly, 15 minutes of incubation with 1 �M NPY signif-icantly increased the number of P-SAPK/JNK-positive nucleiper neurosphere compared with control (control, 1.5 � 0.3;NPY, 4.7 � 0.3; p � .017). Exposure of SVZ cells to NPY for6 hours induced a robust P-SAPK/JNK immunoreactivity inneuritic and growth cone-like structures emerging from theneurospheres and no longer in the nuclei (Fig. 7B–7D), com-pared with control cultures showing a diffuse P-SAPK/JNKstaining throughout the cytoplasm, with only few positivegrowth cone-like structures immunoreactive for P-SAPK/JNK(Fig. 7A). Quantification of the total length of ramifications perneurosphere (Fig. 7E), as well as the number of ramificationsper neurosphere (Fig. 7F), showed that 1 �M NPY increasessignificantly both parameters compared with control cultures(total length of ramification per neurosphere: control, 50.3 � 7.0�m; NPY, 296.6 � 98.9 �m; p � .032; number of ramificationsper neurosphere: control, 0.6 � 0.1; NPY, 1.9 � 0.4; p � .0077;Fig. 7E, 7F). To ascertain that P-SAPK/JNK localization wasassociated with axons, double labeling immunocytochemistry
Figure 3. Neuronal differentiation in control subventricular zone(SVZ) cultures and in cultures exposed to 1 �M NPY. (A–D): Immu-nodetection of microtubule-associated protein-2-positive (A, B) andneuronal nuclear protein (NeuN)-positive (C, D) neurons. (E): Percent-age of NeuN-immunostained neurons. Shown are means � SEM of twoindependent experiments. ���, p � .0001 using two-tailed Student’s ttest for comparison with SVZ control cultures. Abbreviation: NPY,neuropeptide Y.
1640 NPY Promotes SVZ Neurogenesis
was performed to visualize both P-SAPK/JNK and tau, a mi-crotubule-associated protein preferentially found in growingaxons. As displayed in Figure 7G and 7H, P-SAPK/JNK waslocalized predominantly in tau-positive axons in NPY-treatedcultures. Together, these data show that NPY promotes axono-genesis and neuronal maturation involving activation of theSAPK/JNK pathway in SVZ cultures.
DISCUSSION
The main objective of the present work was to explore a poten-tial proneurogenic action of NPY in the SVZ. We first investi-gated whether SVZ cells express NPY receptors using q-PCRrelative quantification. SVZ neurospheres from newborn mice,as well as SVZ explants from adult mice, were found to express
the predominant NPY receptors in the brain, Y1, Y2, and Y5.Y1 receptors were present at higher levels than either Y5 or Y2.The presence of NPY receptors suggested that NPY couldmodulate SVZ proliferation and apoptosis. We investigated thisissue using a BrdU incorporation assay and showed that NPYpromotes proliferation at 48 hours in the SVZ. This is inaccordance with previous studies performed in primary olfac-tory and DG cultures [24, 25]. Indeed, in primary olfactorycultures from newborn rats, exogenous NPY promotes prolifer-ation of the basal cells but has no effect on cell death [24, 25].Similarly, in DG cell cultures from 8–10-day-old rats, NPYsignificantly increased BrdU incorporation but did not affectcell death [26]. After 7 days of incubation, NPY no longerpromotes proliferation. MAP-2 and NeuN immunocytochemis-try revealed that NPY increased the density of MAP-2-positiveramifications, as well as the number of NeuN-positive nuclei,
Figure 4. NPY induces neuronal differentiation in SVZ cultures through Y1 receptors. (A): Subventricular zone (SVZ) cultures were perfusedcontinuously with Krebs solution for 15 minutes and stimulated for 2 minutes (from minute 5 to minute 7) with 50 mM KCl and for 2 minutes (fromminute 10 to minute 12) with 100 �M Hist. (B–D): Representative single-cell calcium imaging profiles of the response of at least 20 cells in anontreated culture (control) (B), a culture maintained 7 days with 1 �M of NPY alone (C), and a culture maintained concomitantly with 1 �M NPYand with 1 �M Y1 receptor antagonist (BIBP3226) (D). (E): Percentage of neuron-like responding cells in SVZ cultures maintained for 7 days inthe absence (control) or in the presence of either 1 �M NPY, 1 �M Y1 receptor agonist ([Leu31 Pro34]-NPY), 300 nM Y2 receptor agonist(NPY13–36), 1 �M Y5 receptor agonist (NPY[19–23]-[Gly1, Ser3, Gln4, Thr6, Ala31, Aib32, Gln34]-PP), or 1 �M NPY with 1 �M Y1 receptorantagonist (BIBP3226). Means � SEM of four independent experiments are represented. ��, p � .01, using one-way analysis of variance withBonferroni’s correction for comparison with SVZ control cultures. Abbreviations: Hist, histamine; NPY, neuropeptide Y; sec, seconds.
1641Agasse, Bernardino, Kristiansen et al.
www.StemCells.com

indicating that there is a shift from proliferation toward differ-entiation. Consistently, studies in DG and primary olfactory cell
cultures exposed to NPY have shown that the number of � IIItubulin cells, a neuron-specific protein, increases [24–26].
We also quantified functional neuronal differentiation usinga method recently developed by our group, based on the mea-surements of [Ca2�]i amplitude responses following stimulationwith KCl or histamine [28]. Briefly, high KCl solution inneurons leads to an increase in [Ca2�]I, whereas in immatureSVZ cells or glial cells the [Ca2�]i remained unchanged. On theother hand, perfusion with 100 �M histamine increases [Ca2�]i
in SVZ immature cells but neither in neurons nor in glial cells[28, 31–33]. The percentage of cells displaying a neuronal-likeprofile (i.e., with a Hist/KCl ratio below 0.8) was calculated[28]. Using this method, we showed that NPY increases thenumber of functional SVZ neurons. Moreover, this effect ismediated by Y1 receptors, since the increase in neuronal differ-entiation was reproduced using the Y1 receptor agonist [Leu31,Pro34]-NPY but abolished in the presence of NPY and the Y1antagonist BIBP3226.
Y1-mediated neurogenesis has been shown to occur in theOE and DG. Indeed, in DG and primary olfactory cell cultures,increase in neuronal proliferation is mimicked by incubationwith Y1 selective agonists and inhibited in the presence ofBIBP3226 [24, 26]. In addition, proliferation is decreased in theDG of Y1 receptor KO mice [26]. Moreover, when functionalbinding assays were performed in adult naıve mice, functionalY1 receptors were shown to be present in the SVZ, consistentwith the concept that endogenous NPY may promote neurogen-esis in vivo via Y1 receptors.
NPY triggered the phosphorylation of both the MAPKs,ERK1/2 and SAPK/JNK. Activation of the ERK pathway byNPY has already been shown in other neurogenic systems andhas been correlated with NPY-induced neuroproliferation. InNPY-treated DG and primary olfactory cell cultures, inhibitionof ERK1/2 phosphorylation or upstream kinase activation abol-ished the neuroproliferative effect of NPY [24, 26]. In ourexperimental model, increase in SVZ proliferation by NPY may,in part, be mediated through ERK1/2 phosphorylation, asERK1/2 activation is required for SVZ cell proliferation [35].Moreover, nuclear localization of P-ERK1/2 is correlated toproliferation, as P-ERK1/2 substrates are mainly transcription
Figure 6. NPY binding activates the extracellular signal-regulated kinase 1/2 (ERK1/2) signaling pathway. (A): Representative fluorescenceconfocal photos of the P-ERK1/2 (red and green) immunocytochemistry and Hoechst staining (blue nuclei), in control cultures and in culturesmaintained for 5 minutes and 1 hour in the presence of 1 �M NPY. Arrows depict P-ERK1/2-positive nuclei (red) and cell cytoplasm (green). (B):Number of P-ERK1/2-positive nuclei per neurosphere. Measurements were done in at least five neurospheres in each coverslip. Means � SEM oftwo independent cultures per condition are represented. ��, p � .005 using two-tailed Student’s t test for comparison with subventricular zone (SVZ)control cultures exposed to normal medium for 5 minutes. (C): Number of P-ERK1/2-positive cells, expressed as percentage of total number of cells,in control cultures and in cultures maintained for 1 hour in the presence of 1 �M NPY. Means � SEM of three independent experiments arerepresented. �, p � .05 using two-tailed Student’s t test for comparison with SVZ control cultures exposed to normal medium for 1 hour.Abbreviations: NPY, neuropeptide Y; P-ERK, phosphorylated form of extracellular signal-regulated kinase 1/2.
Figure 5. Y1 receptor functional binding in the subventricular zone(SVZ). Pseudocolored autoradiograms show Y1 receptor functionalbinding in the SVZ of mice using [Leu31, Pro34]-NPY-stimulated [35S]-GTP�S binding. (A): Basal binding. (B): Y1 agonist: [Leu31, Pro34]-NPY (10�5 M). (C): Nonspecific binding. (D): [Leu31, Pro34]-NPY(10�5 M) � BIBP3226 (10�6 M). (E): Magnification of dashed sectionin (A), corresponding to the SVZ. (F): Magnification of dashed sectionin (B). (G): Magnification of dashed section in (D). Scale bar � 1 mm.(H): Levels of [Leu31, Pro34]-NPY-stimulated [35S]-GTP�S binding andspecificity of Y1 binding in the SVZ of mice. Means � SEM of absolutevalues minus basal binding values obtained in a total of six naıve mice.���, p � .0005, using paired t test for comparison with [Leu31, Pro34]-NPY-stimulated [35S]-GTP�S binding.
1642 NPY Promotes SVZ Neurogenesis

factors involved in the activation of gene expression related tocell-cycle progression [36–38]. After 1 hour of treatment withNPY, P-ERK1/2 is found in the cytoplasm, where it can phos-phorylate neurofilament heavy chain proteins and is associatedwith microtubules regulating cytoskeleton organization and neu-ronal function [38–40].
NPY was found to increase P-SAPK/JNK immunoreactivityfirst in the nucleus and then in the cytoplasm. This is consistentwith a previous study where NPY induced phosphorylation ofSAPK/JNK in synaptosomes prepared from dentate gyrus [41].SAPK/JNKs appear to play an important role in neural differ-entiation and maturation. Indeed, in the nucleus, P-SAPK/JNKphosphorylates transcription factors, such as c-jun, involved inexpression of genes related to neuronal differentiation [38, 42].The JNK pathway is involved in C17.2 neural progenitor celldifferentiation induced by interferon-� [43], and JNK1 is re-quired for neurogenesis in mouse embryonic bodies [44]. In thepresent study, after 6 hours of NPY treatment, P-SAPK/JNKwas found in the cytoplasm, in neurite-like structures, colocal-izing with tau, a microtubule-associated protein mainly presentin axons. Moreover, NPY treatment increased the total lengthand number of P-SAPK/JNK-positive ramifications. Indeed, theSAPK/JNK pathway is involved in neurite outgrowth and axono-genesis [42, 45]. JNK3 expression, for instance, significantlyincreases the number and length of neurites in NGF-inducedneuronal differentiation of PC12 cells [46]. JNK1 is respon-sible for MAP-2 phosphorylation and regulates microtubuleassembly [47]. Consistent with our results, P-SAPK/JNKimmunoreactivity is associated with tau-positive axons andgrowth cones in rat embryonic day 18 hippocampal neurons[45]. Together, these data suggest that NPY promotes neu-ronal differentiation and axonal sprouting through activationof the SAPK/JNK pathway.
Previous studies on the OE and the DG, together with theresults shown in the present work in the SVZ, highlight theproneurogenic properties of NPY [24–26]. NPY is secretedlocally in the DG by interneurons and in the OE by sustentacular
cells. In this study, NPY mRNA was detected in both SVZneurospheres and brain explants. In vivo, NPY may be secretedby SVZ cells acting locally in an autocrine/paracrine manner tostimulate both proliferation and neuronal differentiation. Indeed,the SVZ microenvironment, where stem and progenitor cellsreside, is endowed with factors regulating neurogenesis, referredto as the neurogenic niche [48]. For instance, explants of adultSVZ secrete soluble factors that enhance proliferation, cellsurvival, and neuronal differentiation in SVZ cell cultures [49],and NPY may be part of the SVZ soluble factor repertoire. Tofurther support this hypothesis, it has recently been shown thatproliferation is reduced in the caudal part of the SVZ of Y1knockout mice, suggesting that endogenous secretion of NPYpromotes proliferation in normal mice [50]. Although neighbor-ing striatum contains NPYergic neurons [51], SVZ endogenousproduction of NPY may contribute to basal neurogenesis. More-over, in single-cell calcium imaging studies, the number ofneurons in cultures incubated with both NPY and BIBP3226was found to be slightly lower than in control conditions,probably because of the blocking of the endogenous and exog-enous effect of NPY by BIBP3226.
CONCLUSION
The present work highlights for the first time the proneurogenicproperties of NPY in the SVZ, the major source of repairingcells in the mammalian brain. The use of NPY, as a neurogen-esis activator, opens new perspectives for the development ofSVZ cell-based therapies for brain repair.
ACKNOWLEDGMENTS
We thank Dr. Luisa Cortes for help and expertise in confocalmicroscopy. This work was supported by FCT Portugal and
Figure 7. NPY binding activates the stress-activated protein kinase (SAPK)/c-Jun-NH2-terminal kinase (JNK) signaling pathway.(A–D): Representative fluorescence confo-cal photos of the phosphorylated form ofstress-activated protein kinase (P-SAPK)/JNK (green) and Hoechst staining (blue nu-clei) in cultures maintained for 6 hours incontrol medium (A) or in the presence of 1�M NPY (B–D). (D): High-magnificationfluorescence confocal photos of a growthcone-like structure of a P-SAPK/JNK-posi-tive cell (C) in a subventricular zone (SVZ)culture treated with 1 �M NPY for 6 hours.(E, F): Total length (�m) of the ramifica-tions per neurosphere (E) and number oframifications per neurosphere (F). Means �SEM are shown. Measurements were donein approximately 20 nonoverlapping fieldsin each coverslip using digital images (mag-nification, �20) (from three different cul-tures; total number of neurospheres countedwas at least 20 neurospheres in each culture).�, p � .05; ��, p � .01, using two-tailedStudent’s t test for comparison with SVZcontrol cultures. (G, H): Representative flu-orescence confocal photographs of P-SAPK/JNK (green), tau (red), and Hoechst staining(blue nuclei) in control cultures (G) and incultures maintained for 6 hours in the pres-ence of 1 �M NPY (H). Abbreviation: NPY,neuropeptide Y.
1643Agasse, Bernardino, Kristiansen et al.
www.StemCells.com

by FEDER, POCI/NSE/58492/2004, PTDC/SAU-NEU/68465/2006, FRH/PBD/26462/2006, SFRH/BD/12731/2003, DanishNational Research Council Grant 64750, the Lundbeck Foun-dation, Friis Foundation, and Elsass Foundation.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Garcia AD, Doan NB, Imura T et al. GFAP-expressing progenitors arethe principal source of constitutive neurogenesis in adult mouse fore-brain. Nat Neurosci 2004;7:1233–1241.
2 Taupin P, Gage FH. Adult neurogenesis and neural stem cells of thecentral nervous system in mammals. J Neurosci Res 2002;69:745–749.
3 Betarbet R, Zigova T, Bakay RA et al. Dopaminergic and GABAergicinterneurons of the olfactory bulb are derived from the neonatal subven-tricular zone. Int J Dev Neurosci 1996;14:921–930.
4 Gheusi G, Cremer H, McLean H et al. Importance of newly generatedneurons in the adult olfactory bulb for odor discrimination. Proc NatlAcad Sci U S A 2000;97:1823–1828.
5 Chen J, Magavi SS, Macklis JD. Neurogenesis of corticospinal motorneurons extending spinal projections in adult mice. Proc Natl Acad SciU S A 2004;101:16357–16362.
6 Parent JM, Valentin VV, Lowenstein DH. Prolonged seizures increaseproliferating neuroblasts in the adult rat subventricular zone-olfactorybulb pathway. J Neurosci 2002;22:3174–3188.
7 Romanko MJ, Rola R, Fike JR et al. Roles of the mammalian subven-tricular zone in cell replacement after brain injury. Prog Neurobiol2004;74:77–99.
8 Zhang R, Zhang Z, Wang L et al. Activated neural stem cellscontribute to stroke-induced neurogenesis and neuroblast migrationtoward the infarct boundary in adult rats. J Cereb Blood Flow Metab2004;24:441– 448.
9 Kalra SP, Kalra PS. NPY—An endearing journey in search of a neuro-chemical on/off switch for appetite, sex and reproduction. Peptides2004;25:465–471.
10 Sparta DR, Fee JR, Knapp DJ et al. Elevated anxiety-like behaviorfollowing ethanol exposure in mutant mice lacking neuropeptide Y(NPY). Drug Alcohol Depend 2007;90:297–300.
11 Redrobe JP, Dumont Y, Fournier A et al. The neuropeptide Y (NPY) Y1receptor subtype mediates NPY-induced antidepressant-like activity inthe mouse forced swimming test. Neuropsychopharmacology 2002;26:615–624.
12 Karl T, Herzog H. Behavioral profiling of NPY in aggression andneuropsychiatric diseases. Peptides 2007;28:326–333.
13 Silva AP, Pinheiro PS, Carvalho AP et al. Activation of neuropeptide Yreceptors is neuroprotective against excitotoxicity in organotypic hip-pocampal slice cultures. FASEB J 2003;17:1118–1120.
14 Thiriet N, Deng X, Solinas M et al. Neuropeptide Y protects againstmethamphetamine-induced neuronal apoptosis in the mouse striatum.J Neurosci 2005;25:5273–5279.
15 Xapelli S, Silva AP, Ferreira R et al. Neuropeptide Y can rescue neuronsfrom cell death following the application of an excitotoxic insult withkainate in rat organotypic hippocampal slice cultures. Peptides 2007;28:288–294.
16 Silva AP, Malva JO, Ambrosio AF et al. Role of kainate receptoractivation and desensitization on the [Ca2�]i changes in cultured rathippocampal neurons. J Neurosci Res 2001;65:378–386.
17 Silva AP, Carvalho AP, Carvalho CM et al. Functional interactionbetween neuropeptide Y receptors and modulation of calcium channels inthe rat hippocampus. Neuropharmacology 2003;44:282–292.
18 Silva AP, Xapelli S, Pinheiro PS et al. Up-regulation of neuropeptide Ylevels and modulation of glutamate release through neuropeptide Yreceptors in the hippocampus of kainate-induced epileptic rats. J Neuro-chem 2005;93:163–170.
19 Silva AP, Lourenco J, Xapelli S et al. Protein kinase C activity blocksneuropeptide Y-mediated inhibition of glutamate release and contributesto excitability of the hippocampus in status epilepticus. FASEB J 2007;21:671–681.
20 Woldbye DP, Larsen PJ, Mikkelsen JD et al. Powerful inhibition ofkainic acid seizures by neuropeptide Y via Y5-like receptors. Nat Med1997;3:761–764.
21 Woldbye DP, Nanobashvili A, Sørensen AT et al. Differential suppres-sion of seizures via Y2 and Y5 neuropeptide Y receptors. Neurobiol Dis2005;20:760–772.
22 Calof AL, Mumm JS, Rim PC et al. The neuronal stem cell of theolfactory epithelium. J Neurobiol 1998;36:190–205.
23 Gage FH, Kempermann G, Palmer TD et al. Multipotent progenitor cellsin the adult dentate gyrus. J Neurobiol 1998;36:249–266.
24 Hansel DE, Eipper BA, Ronnett GV. Neuropeptide Y functions as aneuroproliferative factor. Nature 2001;410:940–944.
25 Hansel DE, Eipper BA, Ronnett GV. Regulation of olfactory neurogen-esis by amidated neuropeptides. J Neurosci Res 2001;66:1–7.
26 Howell OW, Doyle K, Goodman JH et al. Neuropeptide Y stimulatesneuronal precursor proliferation in the post-natal and adult dentate gyrus.J Neurochem 2005;93:560–570.
27 Beites CL, Kawauchi S, Crocker CE et al. Identification and molecularregulation of neural stem cells in the olfactory epithelium. Exp Cell Res2005;306:309–316.
28 Agasse F, Bernardino L, Silva BA et al. Response to histamine allows thefunctional identification of neuronal progenitors, neurons, astrocytes andimmature cells in subventricular zone cell cultures. Rejuvenation Res2008;11:187–200.
29 Christensen DZ, Olesen MV, Kristiansen H et al. Unaltered neuropeptideY (NPY)-stimulated [35S]GTPgammaS binding suggests a net increase inNPY signalling after repeated electroconvulsive seizures in mice. J Neu-rosci Res 2006;84:1282–1291.
30 Franklin KBJ, Paxinos G. The Mouse Brain in Stereotaxic Coordinates.New York: Academic Press, 1997.
31 Maric D, Maric I, Barker JL. Developmental changes in cell calciumhomeostasis during neurogenesis of the embryonic rat cerebral cortex.Cereb Cortex 2000;10:561–573.
32 Fiorio Pla A, Maric D, Brazer SC et al. Canonical transient receptorpotential 1 plays a role in basic fibroblast growth factor (bFGF)/FGFreceptor-1-induced Ca2� entry and embryonic rat neural stem cellproliferation. J Neurosci 2005;25:2687–2701.
33 Tran PB, Ren D, Veldhouse TJ et al. Chemokine receptors are expressedwidely by embryonic and adult neural progenitor cells. J Neurosci Res2004;76:20–34.
34 Shaw JL, Gackenheimer SL, Gehlert DR. Functional autoradiographyof neuropeptide Y Y1 and Y2 receptor subtypes in rat brain usingagonist stimulated [35S]GTPgammaS binding. J Chem Neuroanat2003;26:179 –193.
35 Learish RD, Bruss MD, Haak-Frendscho M. Inhibition of mitogen-activated protein kinase kinase blocks proliferation of neural progenitorcells. Brain Res Dev Brain Res 2000;122:97–109.
36 Kalluri HS, Eickstaedt J, Dempsey RJ. Oxygen glucose deprivationinhibits the growth and ERK phosphorylation of neural progenitor cellsin vitro. Neurosci Lett 2007;426:145–148.
37 Kim-Kaneyama JR, Nose K, Shibanuma M. Significance of nuclearrelocalization of ERK1/2 in reactivation of c-fos transcription and DNAsynthesis in senescent fibroblasts. J Biol Chem 2000;275:20685–20692.
38 Pearson G, Robinson F, Beers Gibson T et al. Mitogen-activated protein(MAP) kinase pathways: Regulation and physiological functions. EndocrRev 2001;22:153–183.
39 De Girolamo LA, Billett EE. Role of extracellular-regulated kinase andc-Jun NH2-terminal kinase in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyri-dine-induced neurofilament phosphorylation. J Neurosci Res 2006;83:680–693.
40 Reszka AA, Bulinski JC, Krebs EG et al. Mitogen-activated proteinkinase/extracellular signal-regulated kinase 2 regulates cytoskeletal or-ganization and chemotaxis via catalytic and microtubule-specific inter-actions. Mol Biol Cell 1997;8:1219–1232.
41 Whittaker E, Vereker E, Lynch MA. Neuropeptide Y inhibits glutamaterelease and long-term potentiation in rat dentate gyrus. Brain Res 1999;827:229–233.
42 Waetzig V, Zhao Y, Herdegen T. The bright side of JNKs-Multitalentedmediators in neuronal sprouting, brain development and nerve fiberregeneration. Prog Neurobiol 2006;80:84–97.
43 Kim SJ, Son TG, Kim K et al. Interferon-gamma promotes differentia-tion of neural progenitor cells via the JNK pathway. Neurochem Res2007;32:1399–1406.
44 Amura CR, Marek L, Winn RA et al. Inhibited neurogenesis in JNK1-deficient embryonic stem cells. Mol Cell Biol 2005;25:10791–10802.
45 Oliva AA, Atkins CM, Copenagle L et al. Activated c-Jun N-termi-nal kinase is required for axon formation. J Neurosci 2006;26:9462–9470.
46 Waetzig V, Herdegen T. A single c-Jun N-terminal kinase isoform(JNK3–p54) is an effector in both neuronal differentiation and cell death.J Biol Chem 2003;278:567–572.
47 Chang L, Jones Y, Ellisman MH et al. JNK1 is required for maintenanceof neuronal microtubules and controls phosphorylation of microtubule-associated proteins. Dev Cell 2003;4:521–533.
1644 NPY Promotes SVZ Neurogenesis

48 Alvarez-Buylla A, Lim DA. For the long run: Maintaining germinalniches in the adult brain. Neuron 2004;41:683–686.
49 Agasse F, Roger M, Coronas V. Neurogenic and intact or apoptoticnon-neurogenic areas of adult brain release diffusible molecules thatdifferentially modulate the development of subventricular zone cell cul-tures. Eur J Neurosci 2004;19:1459–1468.
50 Howell OW, Silva S, Scharfman HE et al. Neuropeptide Y is importantfor basal and seizure-induced precursor cell proliferation in the hip-pocampus. Neurobiol Dis 2007;26:174–188.
51 Aoki C, Pickel VM. Neuropeptide Y in the cerebral cortex and the caudate-putamen nuclei: Ultrastructural basis for interactions with GABAergic andnon-GABAergic neurons. J Neurosci 1989;9:4333–4354.
1645Agasse, Bernardino, Kristiansen et al.
www.StemCells.com
Copyright © 2022 FDOKUMEN




















