
Uncaria tomentosa stimulates the proliferation of myeloid progenitor cells

�-Catenin Signaling Promotes Proliferation of Progenitor Cells inthe Adult Mouse Subventricular Zone
KAZUHIDE ADACHI,a,b,c ZAMAN MIRZADEH,d MASANORI SAKAGUCHI,a TORU YAMASHITA,a,b,e
TANIA NIKOLCHEVA,f YUKIKO GOTOH,g GARY PELTZ,f LEYI GONG,h TAKESHI KAWASE,c
ARTURO ALVAREZ-BUYLLA,d HIDEYUKI OKANO,a KAZUNOBU SAWAMOTOa,b,i
aDepartment of Physiology, bBridgestone Laboratory of Developmental and Regenerative Neurobiology, andcDepartment of Neurosurgery, Keio University School of Medicine, Tokyo, Japan; dDepartment of NeurologicalSurgery and Program in Developmental and Stem Cell Biology, University of California San Francisco, SanFrancisco, California, USA; eDepartment of Neurology, Okayama University Graduate School of Medicine,Dentistry and Pharmaceutical Science, Okayama, Japan; fDepartments of Genetics and Genomics and gInstitute ofMolecular and Cellular Biosciences, University of Tokyo, Tokyo, Japan; hDepartment of Chemistry, Roche PaloAlto, Palo Alto, California, USA; iDepartment of Developmental and Regenerative Biology, Nagoya City UniversityGraduate School of Medical Sciences, Nagoya, Japan
Key Words. �-Catenin • Neurogenesis • Cell proliferation • Subventricular zone • Olfactory bulb • Mash1
ABSTRACT
The subventricular zone (SVZ) is the largest germinal zonein the mature rodent brain, and it continuously producesyoung neurons that migrate to the olfactory bulb. Neuralstem cells in this region generate migratory neuroblasts viahighly proliferative transit-amplifying cells. The Wnt/�-catenin signaling pathway partially regulates the prolifera-tion and neuronal differentiation of neural progenitor cellsin the embryonic brain. Here, we studied the role of �-cate-nin signaling in the adult mouse SVZ. �-Catenin-dependentexpression of a destabilized form of green fluorescent pro-tein was detected in progenitor cells in the adult SVZ of
Axin2-d2EGFP reporter mice. Retrovirus-mediated expres-sion of a stabilized �-catenin promoted the proliferation ofMash1� cells and inhibited their differentiation into neuro-blasts. Conversely, the expression of Dkk1, an inhibitor ofWnt signaling, reduced the proliferation of Mash1� cells. Inaddition, an inhibitor of GSK3� promoted the proliferationof Mash1� cells and increased the number of new neuronsin the olfactory bulb 14 days later. These results suggest that�-catenin signaling plays a role in the proliferation of pro-genitor cells in the SVZ of the adult mouse brain. STEMCELLS 2007;25:2827–2836
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
The largest germinal zone of the adult mammalian forebrain isthe subventricular zone (SVZ) of the lateral ventricles, wherenew neurons are continuously generated [1, 2]. This region iscomposed of four cell types [3]: slowly dividing SVZ astro-cytes (type B cells), rapidly dividing transit-amplifying cells(type C cells), migrating neuroblasts (type A cells), andependymal cells (type E cells). Each of these cell typesregulates its cell cycle by a different mechanism [4 –7].However, little is known about the signaling mechanisms thatcontrol cell proliferation in the adult SVZ.
The canonical Wnt pathway is an important regulator ofmammalian neural development [8], and �-catenin is a criticaldownstream component of this pathway. Depending on its phos-phorylation state, �-catenin can be found in the membrane,cytoplasm, or nucleus. In the absence of a Wnt signal, �-cateninis phosphorylated by casein kinase I and GSK3� and degraded
by the ubiquitin-proteasome system. In the presence of a Wntsignal, GSK3� activity is inhibited, and unphosphorylated�-catenin accumulates in the cytoplasm and translocates into thenucleus where it promotes the transactivation of a variety ofdevelopmentally important genes [9].
Both in vitro and in vivo studies demonstrate that the Wnt/�-catenin pathway regulates the proliferation and differentiationof neural progenitor cells [10]. Several studies have demon-strated that the context-dependent role of this pathway in neu-ronal differentiation. For example, neuronal differentiation isinduced in the pluripotent P19 cell line by overexpression of�-catenin [11] or pharmacological inhibition of GSK3� [12].Neuronal differentiation is also induced in vitro by Wnt3a [13]or Wnt7a or by expression of a stabilized form of �-catenin [14].
In contrast, proliferation of neural progenitor cells is in-duced by activation of the �-catenin signaling pathway, and aGSK3� inhibitor can inhibit differentiation and maintain theself-renewal capacity of human and mouse embryonic stem cells[15]. Likewise, Wnt7a and Wnt7b stimulate the proliferation of
Correspondence: Kazunobu Sawamoto, Ph.D., Department of Developmental and Regenerative Biology, Nagoya City University GraduateSchool of Medical Sciences, 1-Kawasumi, Mizuho-cho, Mizuho-ku, Nagoya 467-8601, Japan. Telephone: 81-52-853-8532; Fax: 81-52-851-1898; e-mail: [email protected]; Hideyuki Okano, M.D., Ph.D., Department of Physiology, Keio University School ofMedicine, 35 Shinanomachi, Shinjuku-ku, Tokyo 160-8582, Japan. Telephone: 81-3-5363-3747; Fax: 81-3-3357-5445; e-mail: [email protected] Received March 19, 2007; accepted for publication July 23, 2007; first published online in STEM CELLS EXPRESS August 2, 2007.©AlphaMed Press 1066-5099/2007/$30.00/0 doi: 10.1634/stemcells.2007-0177
TISSUE-SPECIFIC STEM CELLS
STEM CELLS 2007;25:2827–2836 www.StemCells.com

neurogenic progenitors and primary neurospheres [16], and thegrowth of the hippocampus is inhibited in mice lacking Wnt3a[17] or LEF1 [18]. Stabilized �-catenin also causes an over-growth of neural progenitor cells, resulting in a grossly enlargedbrain [19]. A recent study has identified Wnt signaling as aregulatory pathway in adult hippocampal neurogenesis involvedin the control of neuronal differentiation and the proliferation ofprogenitor cells [20]. Wnt3a and Wnt5a both increase the pro-liferation of cultured progenitor cells isolated from postnatal andadult mouse SVZ and promote their neuronal differentiation[21]. Thus, the Wnt/�-catenin signal can regulate the prolifer-ation and/or differentiation of neural progenitor cells duringdevelopment in a stage- and tissue-dependent manner. Here, weused pharmacological inhibitors and retroviral overexpression tostudy the in vivo effect of �-catenin signaling on neurogenesisin the adult SVZ. We found that �-catenin signaling regulatescell proliferation in the SVZ of mice in vivo.
MATERIALS AND METHODS
AnimalsAdult male C57/BL6 mice (9 weeks old) and Axin2-d2EGFP trans-genic mice [22] were maintained on a 12-hour light/dark cycle withunlimited access to food and water. All animal-related procedureswere approved by the Laboratory Animal Care and Use Committeeof Keio University and the University of California San Franciscoand were conducted in accordance with the guidelines of the NIH.
Synthesis and Characterization of Ro3303544The indole maleimide Ro3303544 (supplemental online Fig. 1) isone of a series of highly selective GSK3� inhibitors (inhibitoryconcentration50 of 0.6 nM) that were prepared according to methodsdescribed in a published patent [23]. In brief, Ro3303544 wassynthesized by the conversion of the corresponding furan-2,5-dionewith ammonia in reflux in dimethylformamide. The furan-2,5-diones were synthesized from N-methyl-indol-3-yl-oxo-acetyl chlo-ride and substituted with phenylacetic acid in dichloromethane inthe presence of triethylamine.
For the assay of GSK3� enzyme activity, rabbit GSK3� cDNAwas expressed as a GST fusion in BL21DE3 cells from thepGEX-3X vector [24]. To produce constitutively active GSK3�enzyme, 10 NH2-terminal amino acids were deleted [25]. Trans-formed BL21 DE3 cells were grown at 37°C to mid log-phase, andprotein production was induced with isopropyl-beta-(D)-thiogalac-topyranoside (0.4 mM) at 30°C for 2 hours. After the cells werehomogenized, the extract was loaded on a glutathione-Sepharose 4Bcolumn, and GSK3� was eluted with glutathione buffer (50 mMTris, pH 8, and 10 mM reduced glutathione). The eluate wascollected in 3-minute fractions and assayed for GSK3� content ona 10% SDS-polyacrylamide gel electrophoresis. Fractions above20% peak height were pooled and used in the assay. The GSK3�assay was performed in 50-�l reactions in a 96-well polypropyleneplate; each reaction contained 20 mM magnesium chloride, 40 �MATP, 2 mM dithiothreitol, 88.5 �M biotinylated and phosphory-lated CREB-peptide substrate (biotin-KRREILSRRPS (p) YR),[�-33P]ATP (1 �Ci), and 2 �l of inhibitors in dimethyl sulfoxide(DMSO). Fifteen microliters of the pooled GSK3� elute was added,and the reaction mixture was incubated at 30°C for 1 hour. Thereaction was stopped by transferring 25 �l of the reaction mixtureto a phosphocellulose plate containing 130 �l of 1.85% phosphoricacid. The free radionucleotides in the membrane were washed offunder vacuum with 1.85% phosphoric acid (five times). After thelast wash, the plate was transferred to an adapter plate, 50 �l ofscintillation cocktail (Microscint-20; catalog no. 20-133;PerkinElmer Life and Analytical Sciences, Waltham, MA, http://las.perkinelmer.com) was added to each well, and the amount ofradioactivity was counted in a Top Count scintillation counter.Protein kinase C assays were performed using recombinant proteinkinase C-� (catalog no. 14-232; Upstate, Charlottesville, VA, http://
www.upstate.com ) and substrate (catalog no. 17-139) according tothe manufacturer’s instructions. To assay the inhibitor’s activity onother kinases, Ro3303544 was sent to Upstate Cell Signaling So-lutions (Charlottesville, VA, http://www.upstate.com) for kinasepanel testing.
�-Catenin Immunoblotting7F2 mouse osteoblasts (ATCC CRL-12557) were cultured ingrowth medium in a six-well plate. Upon reaching confluence, thegrowth medium was replaced with differentiation medium (10 mM�-glycerophosphate and 50 �g/ml ascorbic acid) with 100, 300, or500 nM Ro3303544 or vehicle control (DMSO). Twenty-four hourslater, the cells were harvested, and nuclear and cytoplasmic extractswere prepared using a NE-PER kit (Pierce, Rockford, IL, http://www.piercenet.com), according to the manufacturer’s instructions.Lysates (20 �g each) were subjected to immunoblotting with amonoclonal antibody against �-catenin (Transduction Laboratories,Lexington, KY, http://www.bdbiosciences.com/pharmingen). As aloading control, the blots were reprobed with either anti-�-actin(Sigma-Aldrich, St. Louis, http://www.sigmaaldrich.com) or anti-Lamin A/C (Santa Cruz Biotechnology Inc., Santa Cruz, CA, http://www.scbt.com) monoclonal antibodies.
�-Catenin Enzyme ImmunoassayAfter 24 hours of incubation in six-well plates containing GSK3�inhibitors, Jurkat T cells (5 � 105 cells per milliliter) were washedin phosphate-buffered saline (PBS), harvested, and lysed in 0.3 ml ofradioimmunoprecipitation assay buffer (catalog no. 1 920 693; Boehr-inger Mannheim, Mannheim, Germany, http://www.boehringer.com)according to the manufacturer’s instructions. �-Catenin protein in thelysate was analyzed by enzyme immunoassay (EIA). The EIA wasperformed in 96-microwell plates; capture antibody (mouse mono-clonal anti-�-catenin; Zymed, Carlsbad, CA, http://www.invitrogen.com)-coated wells were washed three times (PBS, 0.05% Tween20), blocked with assay diluent (blocked with assay diluent; BDPharmingen, San Diego, http://www.bdbiosciences.com/index_us.shtml), and then incubated at room temperature for at least 72 hours.The wells were washed, 100 �l of cell extract or various concen-trations of purified �-catenin protein were added, and the plateswere incubated for 2 hours at room temperature. The wells werewashed again, 100 �l of anti-�-catenin antibody (�-catenin H-102,sc-7199, rabbit IgG; Santa Cruz Biotechnology) in assay diluent(1:1,250) was added, and the plates were incubated at room tem-perature for 2 hours. Antibody binding was measured using 100 �lof a 1:2,000 dilution of biotinylated mouse monoclonal anti-rabbitIgG antibody (B5283; Sigma-Aldrich) and 3,3�,5,5�-tetramethyl-benzidine (catalog no. 2642KK; BD Pharmingen) for color devel-opment, according to the manufacturer’s instructions.
GSK3� Inhibitor and BromodeoxyuridineAdministrationsMice received two intraperitoneal injections per day of vehicle(saline) or Ro3303544 GSK3� inhibitor (500 �M) for 5 consecutivedays. Bromodeoxyuridine (BrdU) (Boehringer Mannheim) (50 mg/kg) was given intraperitoneally to mice at 1, 3, or 14 days beforesacrifice (n � 5 each). For intracerebroventricular administration,the GSK3� inhibitor (100 �M) or vehicle alone was infused into theright lateral ventricle of the brain for 7 days with a mini-osmoticpump (model 1007D; flow rate, 0.5 �l/hour; Alzet Osmotic Pumps,Cupertino, CA, http://www.alzet.com). The cannula was implantedstereotaxically at the following coordinates: anterior, 0 mm; lateral,1.1 mm; depth, 2.3 mm (relative to the bregma and the surface of thebrain). After 6 days of infusion, BrdU was injected into the mice (50mg/kg i.p.). Twenty-four hours later, the pump was removed, andthe mice were sacrificed for analyses.
Retroviral Injections into the SVZPMX-IRES-GFP, a replication-incompetent retroviral vector con-taining an internal ribosome entry site (IRES) sequence followed bythe coding sequence for enhanced green fluorescent protein (GFP)[26], was used for the production of recombinant retroviruses.
2828 �-Catenin Promotes Adult Neurogenesis

pMX-�N90�-catenin-IRES-GFP and pMX-Dkk1-IRES-GFP weredescribed previously [14]. Ecotropic virus-packaging (PLAT-E)cells were transfected with each plasmid using FuGENE 6 (RocheDiagnostics, Basel, Switzerland, http://www.roche-applied-science.com ) and then cultured for 3 days at 37°C. Retroviral particles werecollected from the culture supernatant by centrifugation at 6,000gfor 16 hours at 4°C and were resuspended in serum-free Dulbecco’smodified Eagle’s medium-Ham’s F-12 medium (1:1) (Invitrogen,Carlsbad, CA, http://www.invitrogen.com). The titers of the retro-viral stocks used in this study were 7.0–7.3 � 105 colony-formingunits per milliliter. Two hundred nanoliters of a viral suspensionwas injected into the right SVZ at the following coordinates: ante-rior, 1 mm; lateral, 1 mm; and depth, 3.4–1.8 mm.
ImmunocytochemistryBrains were perfusion-fixed with 4% paraformaldehyde and post-fixed in the same fixative overnight, and 50 �m sections were cuton a Vibratome sectioning system (VT1000S; Leica, Heidelberg,Germany, http://www.leica.com). After three rinses in PBS, somesections were incubated either in acetone for 20 seconds on ice (forstaining �-catenin) or in 2 N HCl for 30 minutes (for staining BrdU,proliferating cell nuclear antigen [PCNA], and Mash1). The sec-tions were then incubated for 1 hour in TNB blocking solution(Vector Laboratories, Burlingame, CA, http://www.vectorlabs.com), overnight with primary antibodies, followed by a 60-minuteincubation at room temperature with biotinylated secondary anti-bodies (1:200) or Alexa Fluor-conjugated secondary antibodies(1:200; Molecular Probes Inc., Eugene, OR, http://probes.invitrogen.com), unless otherwise noted. Biotinylated antibodieswere visualized using the ABC Elite kit (Vector Laboratories) andTSA (PerkinElmer Life and Analytical Sciences). The primaryantibodies (final dilution and source) used in this study were asfollows: mouse monoclonal anti-glial fibrillary acidic protein(GFAP) (1:100) (catalog number G3893; Sigma-Aldrich), rabbitpolyclonal anti-GFAP (1:1,000) (catalog number Z0334; Dako,Glostrup, Denmark, http://www.dako.com), mouse monoclonal an-ti-PSA-NCAM IgM (1:250, a gift from Tatsunori Seki) [27], mousemonoclonal anti-Mash1 (1:100) (catalog number 556604; BDPharmingen), rat monoclonal anti-BrdU (1:100) (catalog numberab6326; Abcam, Cambridge, U.K., http://www.abcam.com), rabbitanti-GFP (1:200) (catalog number 598; MBL International Corp.,Woburn, MA, http://www.mblintl.com), goat anti-doublecortin (anti-DCX) (1:200) (catalog number sc-8066; Santa Cruz Biotechnol-ogy), anti-phosphohistone H3 (1:200) (catalog number 06-570; Up-state), anti-�-catenin (1:200) (catalog number 610154; Becton,Dickinson and Company, Franklin Lakes, NJ, http://www.bd.com),anti-PCNA (1:200) (catalog number NAO3T; Siemens MedicalSolutions Diagnostics, Oncogene Science Biomarker Group, Cam-bridge, MA, http://www.oncogene.com), anti-NeuN (1:200) (cata-log number MAB377; Chemicon, Temecula, CA, http://www.chemicon.com), and mouse monoclonal anti-cleaved Caspase-3 (1:200) (catalog number 9661; Cell Signaling Technology, Beverly,MA, http://www.cellsignal.com).
These antibodies have been shown to be specific in the follow-ing ways. The anti-GFP and anti-BrdU antibodies used in this studydid not show any staining in brain sections of normal, untreatedmice. The antibodies against anti-GFAP [28], PSA-NCAM [28],Mash1 [29], DCX [30], phosphohistone H3 [31], PCNA [32], NeuN[33], and cleaved Caspase-3 [34] have been widely used as specificmarkers for the immunohistochemistry of brain sections. The label-ing patterns we obtained with these antibodies were consistent withprevious reports. The anti-�-catenin antibody used in this study waspurchased from Becton Dickinson and stains a single band of 92kDa on Western blots (according to the manufacturer’s technicalinformation). This antibody also produced a staining pattern similarto that previously reported [35]. The number of SVZ cells contain-ing nuclear �-catenin, which were detected using this antibody, wasincreased by GSK3� inhibitor injection, as expected for the specificlabeling of �-catenin (Fig. 3). Finally, we further confirmed thespecificity of this antibody by staining brain sections of mice that hadreceived injections of pMX-IRES-GFP or pMX-�N90�-catenin-IRES-GFP retrovirus into the SVZ. The �-catenin signal was clearly colo-calized with GFP in the SVZ treated with pMX-�N90�-catenin-IRES-
GFP but not that treated with pMX-IRES-GFP (supplemental onlineFig. 1), indicating that the signal produced by this antibody representsspecific immunoreactivity with the �-catenin protein.
Quantitative Analysis of ImmunohistochemistryUsing a Vibratome sectioning system, we first prepared a completeset of serial sections of the forebrain. We then selected coronalsections from two brains at the same level of the SVZ or olfactorybulb sections from five brains, all taken at the same level, and usedthem to count the number of cells positive for each marker. Quan-titative analyses of marker expression were performed using a ZeissLSM 510 Laser Scanning Confocal Microscope. For SVZ analyses,we chose two coronal sections from each brain from two anteriorSVZ regions: one contained the anterior half of the anterior horn(between 1.18 mm anterior and 0.58 mm anterior to the bregma) andthe other contained the posterior half (between 0.58 mm anteriorand 0.02 mm posterior to the bregma). We counted all the detectablecells labeled with each marker on both sides of the SVZ, which wasidentified by its higher cell density. For counting nuclear �-catenin-containing cells, we used three-dimensional image stacks.
Statistical AnalysisAll the quantified data are expressed as the mean � SD. Retroviralresults were evaluated for statistical significance by analysis ofvariance with Dunnett’s test. Differences between two means wereanalyzed by the unpaired Student’s t-test and were consideredstatistically significant when the p value was less than 0.05.
RESULTS
Axin2-d2EGFP Reporter Is Expressed in Type Band C Cells in SVZWe first analyzed activation of �-catenin-induced gene expres-sion in each cell type in the SVZ using Ax2-d2EGFP transgenicmice. In these mice, the expression of a green fluorescentprotein with reduced stability (d2EGFP) is driven by the pro-moter and first intron of the Axin2 gene, a direct target of theWnt pathway whose activation is mediated by Tcf/LEF factors[22]. The 5.6-kilobase Axin2 genomic sequence, including thepromoter and first intron, which contains eight Tcf/LEF con-sensus-binding sites, is sufficient to direct the tissue-specificexpression of d2EGFP in transgenic embryos. Therefore, theAx2-d2EGFP reporter is a sensitive and reliable reporter formonitoring the activation of �-catenin signaling in vivo. Coro-nal sections of the Ax2-d2EGFP reporter mouse brains showedd2EGFP expression in the SVZ within the lateral wall of thelateral ventricles (Fig. 1A). To identify each cell type in theSVZ, we used the following markers: GFAP, which is expressedin type B cells [3]; Mash1, which is expressed in type C cells[29, 36] and in a subpopulation of type B cells (Z.M. andA.A.-B., unpublished data); and DCX, which is expressed intype A cells [3] (Fig. 1B–1M). GFP was detected in 42.5% �5.4% of Mash1� cells. GFP was also detectable in a subpopu-lation of GFAP� type B cells, although the fibrous and frag-mentary pattern of GFAP staining prevented quantitative anal-ysis. GFP was not detected in DCX� type A cells. These resultssuggest that the �-catenin signaling is activated in type B and Ccells in the adult SVZ.
�-Catenin Signaling Increases the Number ofMash1� CellsWe next tested the effects of �-catenin signaling on SVZ cells.To activate or inactivate �-catenin signaling specifically inmitotic SVZ cells, we injected the following retroviral vectors[14]: pMX-IRES-GFP (as a control retrovirus), pMX-�N90�-catenin-IRES-GFP (to express a constitutively active form of�-catenin) [37], and pMX-mDKK-IRES-GFP (to express
2829Adachi, Mirzadeh, Sakaguchi et al.
www.StemCells.com

DKK-1, which antagonizes frizzled receptors and inhibits�-catenin signaling) [38–40]. Two days after the injections, wequantified the percentage of infected cells expressing GFP thatwere also positive for Mash1 (for type C cells and a subset of Bcells) or DCX (type A cells) [41]. The percentage of Mash1�
cells was significantly increased by �N90�-catenin and de-creased by mDKK-1 (Fig. 2A–2K; �, p � .05; ��, p � .01; n �5). On the other hand, the percentage of DCX� type A cells wasdecreased after �N90�-catenin expression and was increasedafter mDKK-1 expression (Fig. 2L–2U; ��, p � .01; n � 5).Taken together, these results indicate that �-catenin signalingincreases the percentage of Mash1-positive cells in the SVZ.
�-Catenin Signaling Promotes Cell Proliferation inthe SVZWe next examined the effect of a novel highly potent and selectiveinhibitor of GSK3� enzyme activity (Ro3303544; supplementalonline Fig. 2) on cellular proliferation in the adult SVZ. Sincepharmacological inhibition of GSK3� enzyme activity results inactivation of �-catenin signaling, this enabled us to examine thefunctional importance of this pathway in cells within the SVZ.Ro3303544 inhibits the GSK3� kinase activity at nanomolar con-centrations but does not inhibit other kinases tested (supplementalonline Table 1). In vitro treatment of two different cell types withthis inhibitor produced a dose-dependent increase in cytoplasmicand nuclear �-catenin (supplemental online Fig. 3A, 3B), indicat-ing activation of the canonical Wnt pathway.
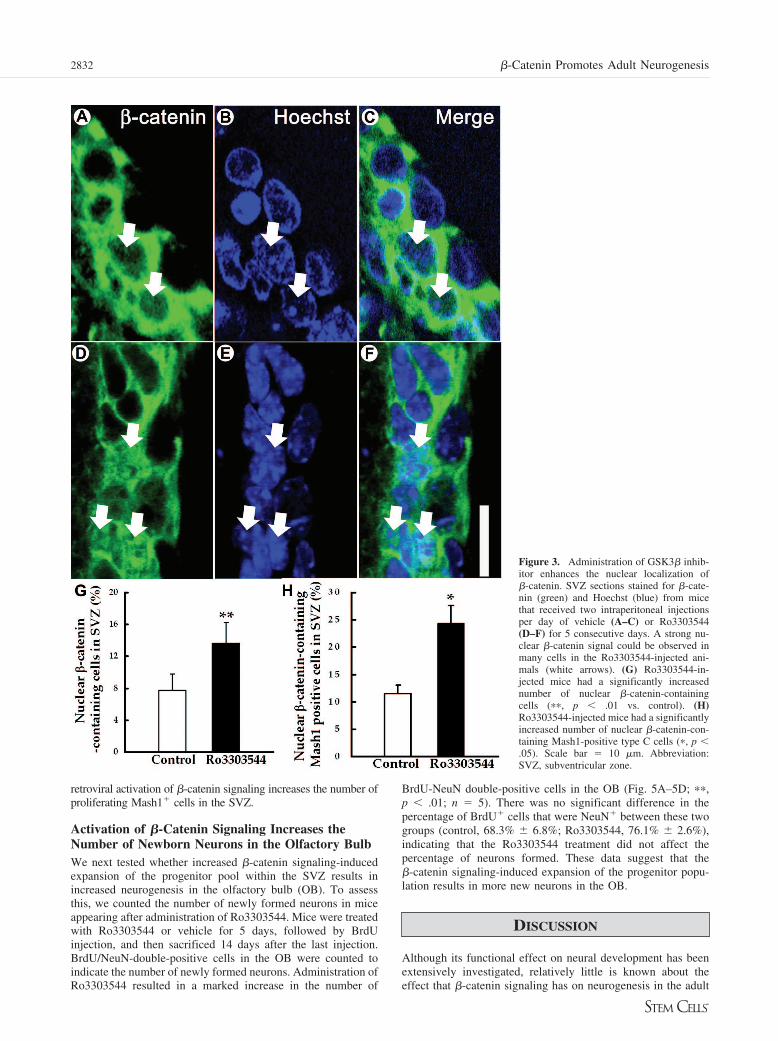
Administration of Ro3303544 over a 5-day period resultedin a marked increase in the number of cells with nuclear �-cate-
nin relative to those that received vehicle control (Fig. 3A–3G;��, p � .01; n � 5). The number of Mash1� cells withdetectable nuclear �-catenin was increased after Ro3303544administration (Fig. 3H; �, p � .05; n � 5). These resultsindicate that administration of Ro3303544 activated �-cateninsignaling in Mash1� cells in the adult SVZ.
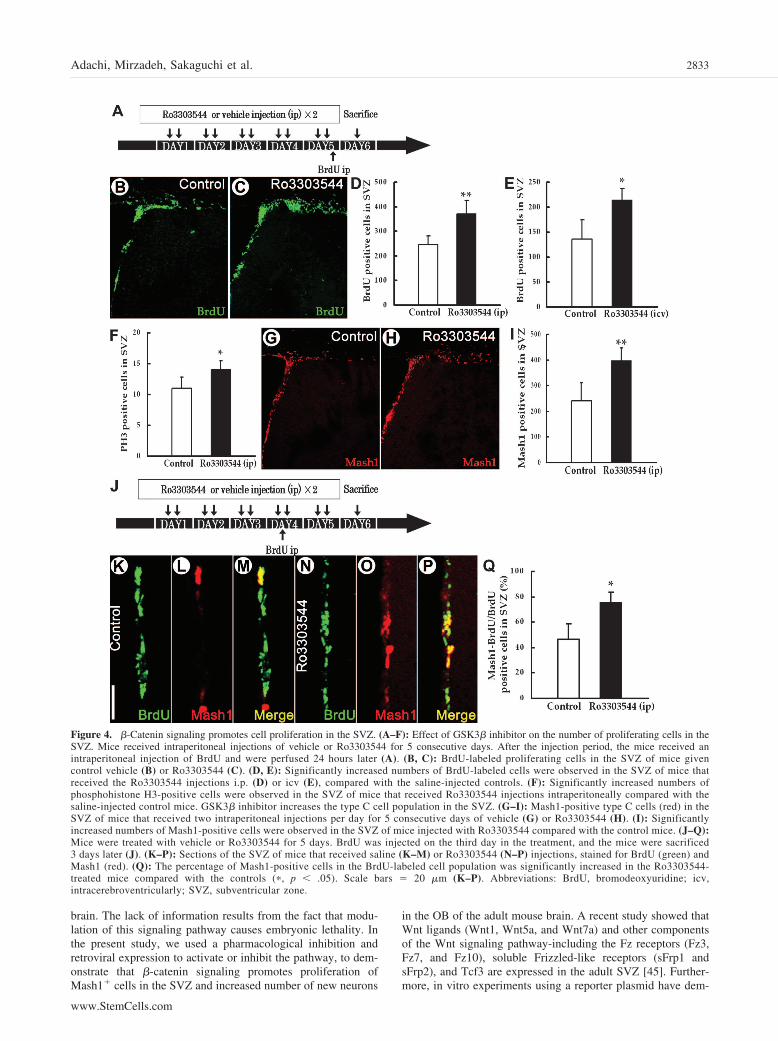
To study the effect of this drug on cell proliferation, mice weretreated with Ro3303544 by intraperitoneal infusion for 5 days (Fig.4A–4D, 4F) or by intracerebroventricular administration for 7 days(Fig. 4E), followed by BrdU injections to label proliferating cellsand sacrificed 1 day later. The number of SVZ cells that werelabeled with BrdU or phosphohistone H3, which is a M phase cellcycle marker [42], was assessed in drug- and vehicle-treated mice.There was a significant increase in the number of labeled cells inthe SVZ of Ro3303544-treated mice relative to the vehicle-infusedcontrol group (Fig. 4A–4F; �, p � .05; ��, p � .01; n � 5 miceeach). There was no significant difference in the number ofcleaved-Caspase3-positive cells in the SVZ between the animalstreated with Ro3303544 and vehicle alone (data not shown; p �.17; n � 5). This suggests that the drug-induced increase in thenumber of BrdU� cells was caused by increased proliferationrather than increased cell survival [43, 44].
We next studied the effects of pharmacologically activating�-catenin signaling on SVZ progenitor cells. Intraperitoneal admin-istration of Ro3303544 for 5 days (two injections per day) resultedin a marked increase in the number of Mash1� cells in the SVZ(Fig. 4G–4I; �, p � .05; ��, p � .01; n � 5). Of note, thepercentage of Mash1� cells that were labeled with BrdU on day 4was significantly increased after Ro3303544 administration (Fig.
Figure 1. �-Catenin-responsive cells in the adult SVZ. (A): Coronal section of Ax2dEGFP reporter mouse shows GFP expression in cells of theSVZ, on the lateral wall of the lateral ventricle. (B–M): At higher magnification, GFP-expressing cells were found coexpressing GFAP (B–E) orMash1 (F–I), but not Dcx (J–M). Abbreviations: Ax2dEGFP, Axin2–d2EGFP; DAPI, 4,6-diamidino-2-phenylindole; Dcx, doublecortin; GFAP, glialfibrillary acidic protein; GFP, green fluorescent protein.
2830 �-Catenin Promotes Adult Neurogenesis

4J–4Q; �, p � .05; n � 5). Similarly, retroviral injection studiesindicated that �-catenin signaling is required and sufficient for
promoting cell proliferation in the SVZ (supplemental online Fig.4). Taken together, these results suggest that pharmacological or
Figure 2. The �-catenin signal increasesthe number of type C cells in the subven-tricular zone. (A–K): Effects of a retrovi-rally introduced stabilized form of �-cateninor Dkk1 on the maintenance of type C cells.(A): Timeline of the experiment. (B–J): Sec-tions of subventricular zone harvested 2 daysafter the stereotaxic injection of pMX-IRES-GFP (B–D), pMX-�N90�-catenin-IRES-GFP (E–G), or pMX-mDKK-IRES-GFP(H–J), stained for Mash1 (red). Infectedcells and their progenies expressed GFP(green). (K): The percentage of Mash1-pos-itive cells in the GFP-positive cell popula-tion was significantly decreased bymDKK-1 (�, p � .05) and increased by�N90�-catenin (��, p � .01). (L–U): Ef-fects of a retrovirally introduced stabilizedform of �-catenin or Dkk1 on the differen-tiation of type C cells into type A cells.(L–T): Sections of subventricular zone har-vested 2 days after the stereotaxic injectionof pMX-IRES-GFP (L–N), pMX-�N90�-catenin-IRES-GFP (O–Q), or pMX-mDKK-IRES-GFP (R–T), stained for Dcx (red). In-fected cells and their progenies expressedGFP (green). (U): The percentage of Dcx-positive cells in the GFP-positive cell popu-lation was significantly increased by DKK1(��, p � .05) and decreased by �N90�-catenin (��, p � .05). Scale bar � 50 �m.Abbreviations: Dcx, doublecortin; GFP,green fluorescent protein; SVZ, subventricu-lar zone.
2831Adachi, Mirzadeh, Sakaguchi et al.
www.StemCells.com
retroviral activation of �-catenin signaling increases the number ofproliferating Mash1� cells in the SVZ.
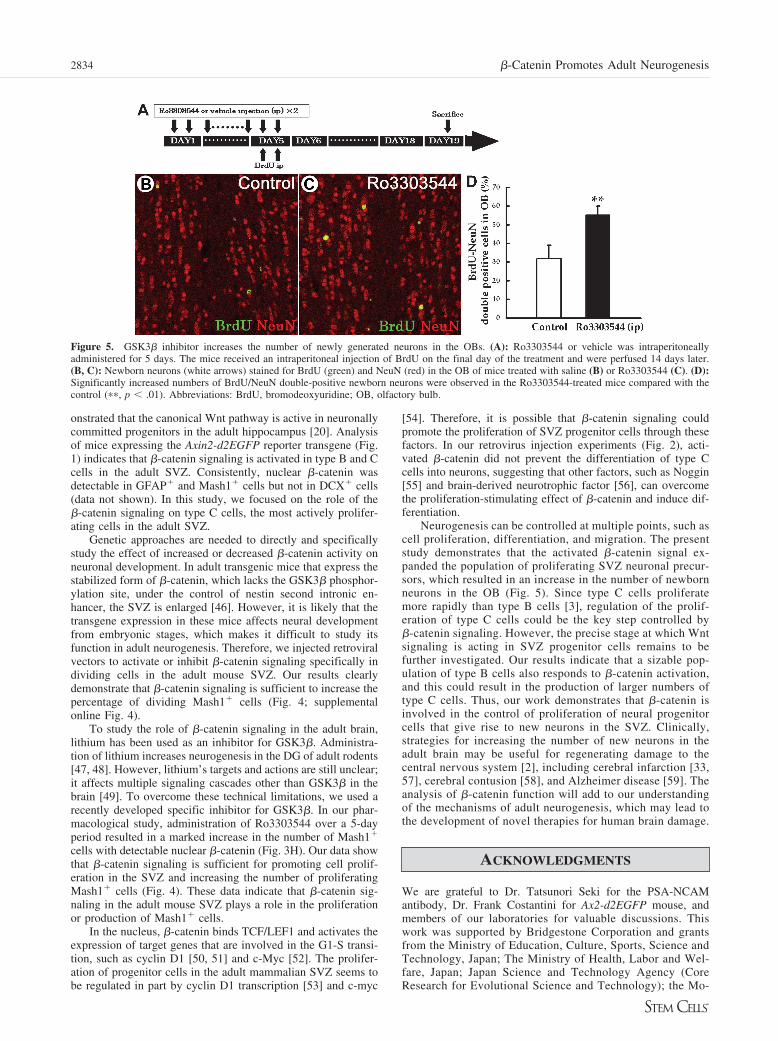
Activation of �-Catenin Signaling Increases theNumber of Newborn Neurons in the Olfactory BulbWe next tested whether increased �-catenin signaling-inducedexpansion of the progenitor pool within the SVZ results inincreased neurogenesis in the olfactory bulb (OB). To assessthis, we counted the number of newly formed neurons in miceappearing after administration of Ro3303544. Mice were treatedwith Ro3303544 or vehicle for 5 days, followed by BrdUinjection, and then sacrificed 14 days after the last injection.BrdU/NeuN-double-positive cells in the OB were counted toindicate the number of newly formed neurons. Administration ofRo3303544 resulted in a marked increase in the number of
BrdU-NeuN double-positive cells in the OB (Fig. 5A–5D; ��,p � .01; n � 5). There was no significant difference in thepercentage of BrdU� cells that were NeuN� between these twogroups (control, 68.3% � 6.8%; Ro3303544, 76.1% � 2.6%),indicating that the Ro3303544 treatment did not affect thepercentage of neurons formed. These data suggest that the�-catenin signaling-induced expansion of the progenitor popu-lation results in more new neurons in the OB.
DISCUSSION
Although its functional effect on neural development has beenextensively investigated, relatively little is known about theeffect that �-catenin signaling has on neurogenesis in the adult
Figure 3. Administration of GSK3� inhib-itor enhances the nuclear localization of�-catenin. SVZ sections stained for �-cate-nin (green) and Hoechst (blue) from micethat received two intraperitoneal injectionsper day of vehicle (A–C) or Ro3303544(D–F) for 5 consecutive days. A strong nu-clear �-catenin signal could be observed inmany cells in the Ro3303544-injected ani-mals (white arrows). (G) Ro3303544-in-jected mice had a significantly increasednumber of nuclear �-catenin-containingcells (��, p � .01 vs. control). (H)Ro3303544-injected mice had a significantlyincreased number of nuclear �-catenin-con-taining Mash1-positive type C cells (�, p �.05). Scale bar � 10 �m. Abbreviation:SVZ, subventricular zone.
2832 �-Catenin Promotes Adult Neurogenesis
brain. The lack of information results from the fact that modu-lation of this signaling pathway causes embryonic lethality. Inthe present study, we used a pharmacological inhibition andretroviral expression to activate or inhibit the pathway, to dem-onstrate that �-catenin signaling promotes proliferation ofMash1� cells in the SVZ and increased number of new neurons
in the OB of the adult mouse brain. A recent study showed thatWnt ligands (Wnt1, Wnt5a, and Wnt7a) and other componentsof the Wnt signaling pathway-including the Fz receptors (Fz3,Fz7, and Fz10), soluble Frizzled-like receptors (sFrp1 andsFrp2), and Tcf3 are expressed in the adult SVZ [45]. Further-more, in vitro experiments using a reporter plasmid have dem-
Figure 4. �-Catenin signaling promotes cell proliferation in the SVZ. (A–F): Effect of GSK3� inhibitor on the number of proliferating cells in theSVZ. Mice received intraperitoneal injections of vehicle or Ro3303544 for 5 consecutive days. After the injection period, the mice received anintraperitoneal injection of BrdU and were perfused 24 hours later (A). (B, C): BrdU-labeled proliferating cells in the SVZ of mice givencontrol vehicle (B) or Ro3303544 (C). (D, E): Significantly increased numbers of BrdU-labeled cells were observed in the SVZ of mice thatreceived the Ro3303544 injections i.p. (D) or icv (E), compared with the saline-injected controls. (F): Significantly increased numbers ofphosphohistone H3-positive cells were observed in the SVZ of mice that received Ro3303544 injections intraperitoneally compared with thesaline-injected control mice. GSK3� inhibitor increases the type C cell population in the SVZ. (G–I): Mash1-positive type C cells (red) in theSVZ of mice that received two intraperitoneal injections per day for 5 consecutive days of vehicle (G) or Ro3303544 (H). (I): Significantlyincreased numbers of Mash1-positive cells were observed in the SVZ of mice injected with Ro3303544 compared with the control mice. (J–Q):Mice were treated with vehicle or Ro3303544 for 5 days. BrdU was injected on the third day in the treatment, and the mice were sacrificed3 days later (J). (K–P): Sections of the SVZ of mice that received saline (K–M) or Ro3303544 (N–P) injections, stained for BrdU (green) andMash1 (red). (Q): The percentage of Mash1-positive cells in the BrdU-labeled cell population was significantly increased in the Ro3303544-treated mice compared with the controls (�, p � .05). Scale bars � 20 �m (K–P). Abbreviations: BrdU, bromodeoxyuridine; icv,intracerebroventricularly; SVZ, subventricular zone.
2833Adachi, Mirzadeh, Sakaguchi et al.
www.StemCells.com
onstrated that the canonical Wnt pathway is active in neuronallycommitted progenitors in the adult hippocampus [20]. Analysisof mice expressing the Axin2-d2EGFP reporter transgene (Fig.1) indicates that �-catenin signaling is activated in type B and Ccells in the adult SVZ. Consistently, nuclear �-catenin wasdetectable in GFAP� and Mash1� cells but not in DCX� cells(data not shown). In this study, we focused on the role of the�-catenin signaling on type C cells, the most actively prolifer-ating cells in the adult SVZ.
Genetic approaches are needed to directly and specificallystudy the effect of increased or decreased �-catenin activity onneuronal development. In adult transgenic mice that express thestabilized form of �-catenin, which lacks the GSK3� phosphor-ylation site, under the control of nestin second intronic en-hancer, the SVZ is enlarged [46]. However, it is likely that thetransgene expression in these mice affects neural developmentfrom embryonic stages, which makes it difficult to study itsfunction in adult neurogenesis. Therefore, we injected retroviralvectors to activate or inhibit �-catenin signaling specifically individing cells in the adult mouse SVZ. Our results clearlydemonstrate that �-catenin signaling is sufficient to increase thepercentage of dividing Mash1� cells (Fig. 4; supplementalonline Fig. 4).
To study the role of �-catenin signaling in the adult brain,lithium has been used as an inhibitor for GSK3�. Administra-tion of lithium increases neurogenesis in the DG of adult rodents[47, 48]. However, lithium’s targets and actions are still unclear;it affects multiple signaling cascades other than GSK3� in thebrain [49]. To overcome these technical limitations, we used arecently developed specific inhibitor for GSK3�. In our phar-macological study, administration of Ro3303544 over a 5-dayperiod resulted in a marked increase in the number of Mash1�
cells with detectable nuclear �-catenin (Fig. 3H). Our data showthat �-catenin signaling is sufficient for promoting cell prolif-eration in the SVZ and increasing the number of proliferatingMash1� cells (Fig. 4). These data indicate that �-catenin sig-naling in the adult mouse SVZ plays a role in the proliferationor production of Mash1� cells.
In the nucleus, �-catenin binds TCF/LEF1 and activates theexpression of target genes that are involved in the G1-S transi-tion, such as cyclin D1 [50, 51] and c-Myc [52]. The prolifer-ation of progenitor cells in the adult mammalian SVZ seems tobe regulated in part by cyclin D1 transcription [53] and c-myc
[54]. Therefore, it is possible that �-catenin signaling couldpromote the proliferation of SVZ progenitor cells through thesefactors. In our retrovirus injection experiments (Fig. 2), acti-vated �-catenin did not prevent the differentiation of type Ccells into neurons, suggesting that other factors, such as Noggin[55] and brain-derived neurotrophic factor [56], can overcomethe proliferation-stimulating effect of �-catenin and induce dif-ferentiation.
Neurogenesis can be controlled at multiple points, such ascell proliferation, differentiation, and migration. The presentstudy demonstrates that the activated �-catenin signal ex-panded the population of proliferating SVZ neuronal precur-sors, which resulted in an increase in the number of newbornneurons in the OB (Fig. 5). Since type C cells proliferatemore rapidly than type B cells [3], regulation of the prolif-eration of type C cells could be the key step controlled by�-catenin signaling. However, the precise stage at which Wntsignaling is acting in SVZ progenitor cells remains to befurther investigated. Our results indicate that a sizable pop-ulation of type B cells also responds to �-catenin activation,and this could result in the production of larger numbers oftype C cells. Thus, our work demonstrates that �-catenin isinvolved in the control of proliferation of neural progenitorcells that give rise to new neurons in the SVZ. Clinically,strategies for increasing the number of new neurons in theadult brain may be useful for regenerating damage to thecentral nervous system [2], including cerebral infarction [33,57], cerebral contusion [58], and Alzheimer disease [59]. Theanalysis of �-catenin function will add to our understandingof the mechanisms of adult neurogenesis, which may lead tothe development of novel therapies for human brain damage.
ACKNOWLEDGMENTS
We are grateful to Dr. Tatsunori Seki for the PSA-NCAMantibody, Dr. Frank Costantini for Ax2-d2EGFP mouse, andmembers of our laboratories for valuable discussions. Thiswork was supported by Bridgestone Corporation and grantsfrom the Ministry of Education, Culture, Sports, Science andTechnology, Japan; The Ministry of Health, Labor and Wel-fare, Japan; Japan Science and Technology Agency (CoreResearch for Evolutional Science and Technology); the Mo-
Figure 5. GSK3� inhibitor increases the number of newly generated neurons in the OBs. (A): Ro3303544 or vehicle was intraperitoneallyadministered for 5 days. The mice received an intraperitoneal injection of BrdU on the final day of the treatment and were perfused 14 days later.(B, C): Newborn neurons (white arrows) stained for BrdU (green) and NeuN (red) in the OB of mice treated with saline (B) or Ro3303544 (C). (D):Significantly increased numbers of BrdU/NeuN double-positive newborn neurons were observed in the Ro3303544-treated mice compared with thecontrol (��, p � .01). Abbreviations: BrdU, bromodeoxyuridine; OB, olfactory bulb.
2834 �-Catenin Promotes Adult Neurogenesis

chida Memorial Foundation for Medical and PharmaceuticalResearch; the Inamori Foundation; and the Takeda Founda-tion. Work in the Alvarez-Buylla laboratory was supportedby NIH Grant HD32116 and a gift from Frances and JohnBowes, and by NIH Grant P30 DK063720 to the DiabetesCenter Microscopy Core.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Alvarez-Buylla A, Garcia-Verdugo JM. Neurogenesis in adult subven-tricular zone. J Neurosci 2002;22:629–634.
2 Okano H. Stem cell biology of the central nervous system. J NeurosciRes 2002;69:698–707.
3 Doetsch F, Garcia-Verdugo JM, Alvarez-Buylla A. Cellular compositionand three-dimensional organization of the subventricular germinal zonein the adult mammalian brain. J Neurosci 1997;17:5046–5061.
4 Doetsch F, Verdugo JM, Caille I et al. Lack of the cell-cycle inhibitorp27Kip1 results in selective increase of transit-amplifying cells for adultneurogenesis. J Neurosci 2002;22:2255–2264.
5 Smith CM, Luskin MB. Cell cycle length of olfactory bulb neuronalprogenitors in the rostral migratory stream. Dev Dyn 1998;213:220–227.
6 Morshead CM, van der Kooy D. Postmitotic death is the fate of consti-tutively proliferating cells in the subependymal layer of the adult mousebrain. J Neurosci 1992;12:249–256.
7 Schultze B, Korr H. Cell kinetic studies of different cell types in thedeveloping and adult brain of the rat and the mouse: A review. CellTissue Kinet 1981;14:309–325.
8 Patapoutian A, Reichardt LF. Roles of Wnt proteins in neural develop-ment and maintenance. Curr Opin Neurobiol 2000;10:392–399.
9 Wodarz A, Nusse R. Mechanisms of Wnt signaling in development.Annu Rev Cell Dev Biol 1998;14:59–88.
10 Hirabayashi Y, Gotoh Y. Stage-dependent fate determination of neuralprecursor cells in mouse forebrain. Neurosci Res 2005;51:331–336.
11 Israsena N, Hu M, Fu W et al. The presence of FGF2 signaling deter-mines whether beta-catenin exerts effects on proliferation or neuronaldifferentiation of neural stem cells. Dev Biol 2004;268:220–231.
12 Ding S, Wu TY, Brinker A et al. Synthetic small molecules that controlstem cell fate. Proc Natl Acad Sci U S A 2003;100:7632–7637.
13 Otero JJ, Fu W, Kan L et al. Beta-catenin signaling is required forneural differentiation of embryonic stem cells. Development 2004;131:3545–3557.
14 Hirabayashi Y, Itoh Y, Tabata H et al. The Wnt/beta-catenin pathwaydirects neuronal differentiation of cortical neural precursor cells. Devel-opment 2004;131:2791–2801.
15 Sato N, Meijer L, Skaltsounis L et al. Maintenance of pluripotency inhuman and mouse embryonic stem cells through activation of Wntsignaling by a pharmacological GSK-3-specific inhibitor. Nat Med 2004;10:55–63.
16 Viti J, Gulacsi A, Lillien L. Wnt regulation of progenitor maturation inthe cortex depends on Shh or fibroblast growth factor 2. J Neurosci2003;23:5919–5927.
17 Lee SM, Tole S, Grove E et al. A local Wnt-3a signal is required fordevelopment of the mammalian hippocampus. Development 2000;127:457–467.
18 Galceran J, Miyashita-Lin EM, Devaney E et al. Hippocampus develop-ment and generation of dentate gyrus granule cells is regulated by LEF1.Development 2000;127:469–482.
19 Chenn A, Walsh CA. Regulation of cerebral cortical size by control ofcell cycle exit in neural precursors. Science 2002;297:365–369.
20 Lie DC, Colamarino SA, Song HJ et al. Wnt signalling regulates adulthippocampal neurogenesis. Nature 2005;437:1370–1375.
21 Yu JM, Kim JH, Song GS et al. Increase in proliferation and differen-tiation of neural progenitor cells isolated from postnatal and adult micebrain by Wnt-3a and Wnt-5a. Mol Cell Biochem 2006;288:17–28.
22 Jho EH, Zhang T, Domon C et al. Wnt/beta-catenin/Tcf signaling in-duces the transcription of Axin2, a negative regulator of the signalingpathway. Mol Cell Biol 2002;22:1172–1183.
23 Gong L, Grupe A, Peltz G, inventors; Syntex, LLC, assignee. 3-Indolyl-4-phenyl-1H-pyrrole-2,5-dione derivatives as inhibitors of glycogen syn-thase kinase-3�. US patent 6479490. November 12, 2002.
24 Wang QM, Fiol CJ, DePaoli-Roach AA et al. Glycogen synthasekinase-3 beta is a dual specificity kinase differentially regulated bytyrosine and serine/threonine phosphorylation. J Biol Chem 1994;269:14566 –14574.
25 Murai H, Okazaki M, Kikuchi A. Tyrosine dephosphorylation of glyco-gen synthase kinase-3 is involved in its extracellular signal-dependentinactivation. FEBS Lett 1996;392:153–160.
26 Morita S, Kojima T, Kitamura T. Plat-E: An efficient and stable systemfor transient packaging of retroviruses. Gene Ther 2000;7:1063–1066.
27 Seki T, Arai Y. The persistent expression of a highly polysialylatedNCAM in the dentate gyrus of the adult rat. Neurosci Res 1991;12:503–513.
28 Doetsch F, Caille I, Lim DA et al. Subventricular zone astrocytes areneural stem cells in the adult mammalian brain. Cell 1999;97:703–716.
29 Sakaguchi M, Shingo T, Shimazaki T et al. A carbohydrate-bindingprotein, Galectin-1, promotes proliferation of adult neural stem cells.Proc Natl Acad Sci U S A 2006;103:7112–7117.
30 Lee SH, Kim YJ, Lee KM et al. Ischemic preconditioning enhancesneurogenesis in the subventricular zone. Neuroscience 2007;146:1020 –1031.
31 Liu X, Chen N, Wang X et al. Apoptosis and proliferation markers indiffusely infiltrating astrocytomas: Profiling of 17 molecules. J Neuro-pathol Exp Neurol 2006;65:905–913.
32 Nagai R, Tsunoda S, Asada H et al. Proliferating cell nuclear antigenpositive cells in the hippocampal subgranular zone decline after irradi-ation in a rodent model. Neurol Res 2002;24:517–520.
33 Yamashita T, Ninomiya M, Hernandez Acosta P et al. Subventricularzone-derived neuroblasts migrate and differentiate into mature neurons inthe post-stroke adult striatum. J Neurosci 2006;26:6627–6636.
34 He XJ, Nakayama H, Dong M et al. Evidence of apoptosis in thesubventricular zone and rostral migratory stream in the MPTP mousemodel of Parkinson disease. J Neuropathol Exp Neurol 2006;65:873– 882.
35 Cleary MA, Uboha N, Picciotto MR et al. Expression of ezrin in glialtubes in the adult subventricular zone and rostral migratory stream.Neuroscience 2006;143:851–861.
36 Parras CM, Galli R, Britz O et al. Mash1 specifies neurons and oligo-dendrocytes in the postnatal brain. EMBO J 2004;23:4495–4505.
37 Morin PJ, Sparks AB, Korinek V et al. Activation of beta-catenin-Tcfsignaling in colon cancer by mutations in beta-catenin or APC. Science1997;275:1787–1790.
38 Bafico A, Liu G, Yaniv A et al. Novel mechanism of Wnt signallinginhibition mediated by Dickkopf-1 interaction with LRP6/Arrow. NatCell Biol 2001;3:683–686.
39 Mao B, Wu W, Li Y et al. LDL-receptor-related protein 6 is a receptorfor Dickkopf proteins. Nature 2001;411:321–325.
40 Semenov MV, Tamai K, Brott BK et al. Head inducer Dickkopf-1 is aligand for Wnt coreceptor LRP6. Curr Biol 2001;11:951–961.
41 Nacher J, Crespo C, McEwen BS. Doublecortin expression in the adultrat telencephalon. Eur J Neurosci 2001;14:629–644.
42 Mahadevan LC, Willis AC, Barratt MJ. Rapid histone H3 phosphoryla-tion in response to growth factors, phorbol esters, okadaic acid, andprotein synthesis inhibitors. Cell 1991;65:775–783.
43 Kuan CY, Schloemer AJ, Lu A et al. Hypoxia-ischemia induces DNAsynthesis without cell proliferation in dying neurons in adult rodentbrain. J Neurosci 2004;24:10763–10772.
44 Namura S, Zhu J, Fink K et al. Activation and cleavage of caspase-3 inapoptosis induced by experimental cerebral ischemia. J Neurosci 1998;18:3659–3668.
45 Shimogori T, VanSant J, Paik E et al. Members of the Wnt, Fz, and Frpgene families expressed in postnatal mouse cerebral cortex. J CompNeurol 2004;473:496–510.
46 Chenn A, Walsh CA. Increased neuronal production, enlarged forebrainsand cytoarchitectural distortions in beta-catenin overexpressing trans-genic mice. Cereb Cortex 2003;13:599–606.
47 Chen G, Rajkowska G, Du F et al. Enhancement of hippocampal neu-rogenesis by lithium. J Neurochem 2000;75:1729–1734.
48 Son H, Yu IT, Hwang SJ et al. Lithium enhances long-term potentiationindependently of hippocampal neurogenesis in the rat dentate gyrus.J Neurochem 2003;85:872–881.
49 Coyle JT, Duman RS. Finding the intracellular signaling pathways af-fected by mood disorder treatments. Neuron 2003;38:157–160.
50 Tetsu O, McCormick F. Beta-catenin regulates expression of cyclin D1in colon carcinoma cells. Nature 1999;398:422–426.
51 Shtutman M, Zhurinsky J, Simcha I et al. The cyclin D1 gene is a targetof the beta-catenin/LEF-1 pathway. Proc Natl Acad Sci U S A 1999;96:5522–5527.
52 He TC, Sparks AB, Rago C et al. Identification of c-MYC as a target ofthe APC pathway. Science 1998;281:1509–1512.
2835Adachi, Mirzadeh, Sakaguchi et al.
www.StemCells.com

53 Coronas V, Bantubungi K, Fombonne J et al. Dopamine D3 receptorstimulation promotes the proliferation of cells derived from the post-natalsubventricular zone. J Neurochem 2004;91:1292–1301.
54 Lemkine GF, Raj A, Alfama G et al. Adult neural stem cell cycling invivo requires thyroid hormone and its alpha receptor. FASEB J 2005;19:863–865.
55 Lim DA, Tramontin AD, Trevejo JM et al. Noggin antagonizes BMPsignaling to create a niche for adult neurogenesis. Neuron 2000;28:713–726.
56 Louissaint A Jr, Rao S, Leventhal C et al. Coordinated interaction of
neurogenesis and angiogenesis in the adult songbird brain. Neuron 2002;34:945–960.
57 Ninomiya M, Yamashita T, Araki N et al. Enhanced neurogenesis in theischemic striatum following EGF-induced expansion of transit-amplify-ing cells in the subventricular zone. Neurosci Lett 2006;403:63–67.
58 Shear DA, Tate MC, Archer DR et al. Neural progenitor cell transplantspromote long-term functional recovery after traumatic brain injury. BrainRes 2004;1026:11–22.
59 Sugaya K. Possible use of autologous stem cell therapies for Alzheimer’sdisease. Curr Alzheimer Res 2005;2:367–376.
See www.StemCells.com for supplemental material available online.
2836 �-Catenin Promotes Adult Neurogenesis
Copyright © 2022 FDOKUMEN




















