
miR-223 Deficiency Increases Eosinophil Progenitor Proliferation
8
of May 27, 2016. This information is current as Progenitor Proliferation miR-223 Deficiency Increases Eosinophil J. Aronow and Marc E. Rothenberg Itskovich, Andrew J. Plassard, Patricia C. Fulkerson, Bruce Thomas X. Lu, Eun-Jin Lim, John A. Besse, Svetlana http://www.jimmunol.org/content/190/4/1576 doi: 10.4049/jimmunol.1202897 January 2013; 2013; 190:1576-1582; Prepublished online 16 J Immunol Material Supplementary 7.DC1.html http://www.jimmunol.org/content/suppl/2013/01/16/jimmunol.120289 References http://www.jimmunol.org/content/190/4/1576.full#ref-list-1 , 14 of which you can access for free at: cites 27 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2013 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on May 27, 2016 http://www.jimmunol.org/ Downloaded from by guest on May 27, 2016 http://www.jimmunol.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of miR-223 Deficiency Increases Eosinophil Progenitor Proliferation

of May 27, 2016.This information is current as
Progenitor ProliferationmiR-223 Deficiency Increases Eosinophil
J. Aronow and Marc E. RothenbergItskovich, Andrew J. Plassard, Patricia C. Fulkerson, Bruce Thomas X. Lu, Eun-Jin Lim, John A. Besse, Svetlana
http://www.jimmunol.org/content/190/4/1576doi: 10.4049/jimmunol.1202897January 2013;
2013; 190:1576-1582; Prepublished online 16J Immunol
MaterialSupplementary
7.DC1.htmlhttp://www.jimmunol.org/content/suppl/2013/01/16/jimmunol.120289
Referenceshttp://www.jimmunol.org/content/190/4/1576.full#ref-list-1
, 14 of which you can access for free at: cites 27 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2013 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 27, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 27, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
miR-223 Deficiency Increases Eosinophil ProgenitorProliferation
Thomas X. Lu,*,† Eun-Jin Lim,* John A. Besse,* Svetlana Itskovich,* Andrew J. Plassard,‡
Patricia C. Fulkerson,* Bruce J. Aronow,‡ and Marc E. Rothenberg*
Recently, microRNAs have been shown to be involved in hematopoietic cell development, but their role in eosinophilopoiesis has
not yet been described. In this article, we show that miR-223 is upregulated during eosinophil differentiation in an ex vivo bone
marrow–derived eosinophil culture system. Targeted ablation of miR-223 leads to an increased proliferation of eosinophil pro-
genitors. We found upregulation of a miR-223 target gene, IGF1R, in the eosinophil progenitor cultures derived from miR-2232/2
mice compared with miR-223+/+ littermate controls. The increased proliferation of miR-2232/2 eosinophil progenitors was reversed
by treatment with an IGF1R inhibitor (picropodophyllin). Whole-genome microarray analysis of differentially regulated genes
between miR-223+/+ and miR-2232/2 eosinophil progenitor cultures identified a specific enrichment in genes that regulate hema-
tologic cell development. Indeed, miR-2232/2 eosinophil progenitors had a delay in differentiation. Our results demonstrate that
microRNAs regulate the development of eosinophils by influencing eosinophil progenitor growth and differentiation and identify
a contributory role for miR-223 in this process. The Journal of Immunology, 2013, 190: 1576–1582.
Eosinophils are multifunctional effector cells implicated inthe pathogenesis of a variety of diseases including asthma,eosinophil gastrointestinal disorders, and helminth in-
fection (1–3). They differentiate from a common myeloid pro-genitor in mice through an intermediate granulocyte/macrophageprogenitor and then via an eosinophil lineage-committed progen-itor (4). The cytokine IL-5 is particularly important in eosinophillineage development as it promotes the selective differentiation ofeosinophils and also stimulates the release of mature eosinophilsfrom the bone marrow (5). Indeed, eosinophil progenitors are asubset of granulocyte/macrophage progenitors that are IL-5Ra+,and IL-5 induces the growth and maturation of eosinophils (4, 6).MicroRNAs (miRNAs) are short, single-stranded RNA mole-
cules that silence target genes posttranscriptionally by eitherinhibiting protein translation or facilitating the degradation of
target mRNAs. Different hematopoietic lineages have significantdifferences in their miRNA expression (7). Various miRNAs havebeen reported to regulate the differentiation and lineage commit-ment of hematopoietic progenitor cells (8). However, there is alack of information on how miRNAs regulate the development ofhematopoietic cells after lineage commitment. To the best of ourknowledge, regulation of eosinophil progenitor cell proliferationby miRNAs has not been reported. Although multiple cytokines(e.g., IL-3, IL-5, and GM-CSF) and transcription factors (e.g.,GATA-1, PU.1) have been shown to regulate the growth of eo-sinophil progenitors (9), other regulatory mechanisms such asmiRNAs likely have a role in regulating or fine-tuning this pro-cess. Only one recent report has focused on this, showing thatmiR-21* could regulate the prosurvival effect of GM-CSF oneosinophils (10).The expression of miR-223 has been shown to be driven by the
myeloid transcription factors PU.1 and C/EBP, factors that areimportant in eosinophilopoiesis (11). In this article, we show thatmiR-223 was upregulated during eosinophil differentiation in anex vivo bone marrow–derived eosinophil culture model. Notably,miR-223-deficient eosinophil progenitor cells showed a hyper-proliferative capacity. Mechanistic analysis identified a contribu-tory role for the miR-223 target, IGF1R, in mediating eosinophilprogenitor cell proliferation. Gene expression analysis followedby systems biological analysis identified a role for miR-223 inhematopoietic development and cellular growth and function.Consistent with this prediction, miR-2232/2 eosinophils had a delayin eosinophil differentiation as assessed by CCR3 expression. Ourdata suggest that miRNAs can directly regulate the developmentof eosinophils by influencing the proliferation and differentiationof eosinophil progenitor cells.
Materials and MethodsMice
The miR-223 gene–targeted mice backcrossed for five generations into theC57BL/6 background was previously described; the mice were kindlyprovided to us by Dr. Eran Hornstein (Weizmann Institute of Science,Rehovot, Israel) (12). Littermate controls were used for all experiments.All animals used were housed under specific pathogen-free conditions in
*Division of Allergy and Immunology, Cincinnati Children’s Hospital MedicalCenter, University of Cincinnati College of Medicine, Cincinnati, OH 45229;†Physician Scientist Training Program, Cincinnati Children’s Hospital MedicalCenter, University of Cincinnati College of Medicine, Cincinnati, OH 45229; and‡Division of Biomedical Informatics, Cincinnati Children’s Hospital Medical Center,University of Cincinnati College of Medicine, Cincinnati, OH 45229
Received for publication October 18, 2012. Accepted for publication December 9,2012.
This work was supported by the Ruth L. Kirschstein National Research ServiceAward for individual predoctoral M.D./Ph.D. fellows from the National Heart Lungand Blood Institute (Award F30HL104892 to T.X.L.), the Ryan Fellowship from theAlbert J. Ryan Foundation (to T.X.L.), an Organogenesis Training Grant (NationalInstitutes of Health [NIH] Grant T32HD046387 to T.X.L.), the University of Cin-cinnati Medical Scientist Training Program (NIH Grant T32GM063483), NIH GrantR01AI083450 (to M.E.R.), the Campaign Urging Research for Eosinophilic Disease,the Buckeye Foundation, and the Food Allergy Research and Education (FARE;formerly Food Allergy Initiative and The Food Allergy & Anaphylaxis Network).
The sequences presented in this article have been submitted to Array Express data-base (http://www.ebi.ac.uk/arrayexpress/) under accession number E-MEXP-3350.
Address correspondence and reprint requests to Dr. Marc E. Rothenberg, Divisionof Allergy and Immunology, Cincinnati Children’s Hospital Medical Center, 3333Burnet Avenue, ML7028, Cincinnati, OH 45229-3039. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviation used in this article: miRNA, microRNA.
Copyright� 2013 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/13/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1202897
by guest on May 27, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
accordance with institutional guidelines. The Institutional Animal Careand Use Committee of the Cincinnati Children’s Hospital Medical Centerapproved the use of animals in these experiments.
Bone marrow–derived eosinophil cultures
Bone marrow cells were collected from femur and tibia of the mice, and thestem/progenitor cell–enriched, low-density fraction was isolated by gra-dient centrifugation using Histopaque 1083 (Sigma) according to manu-facturer’s protocol. The low-density fraction of bone marrow cells wascultured in IMDM with 10% FBS, 100 U/ml penicillin, and 100 mg/mlstreptomycin supplemented with 100 ng/ml stem cell factor and 100 ng/mlFLT-3 ligand (PeproTech) from days 0–4. On day 4, the stem cell factorand FLT-3 ligand were replaced with 10 ng/ml IL-5 and cultured for anadditional 10–12 d (6). The culture media was changed every other day,and cells were counted and concentration was adjusted to 1 3 106/mlduring each media change. Eosinophil maturity was assessed by FACSstaining for CCR3 and Siglec-F and/or Diff-Quik staining of cytospin prep-arations. Eosinophil progenitor proliferation was assessed by counting thecells every 2 d using a hemacytometer.
Quantitative assessment of miRNA levels
Total RNA was isolated using miRNeasy mini Kit according to manu-facturer’s protocols (Qiagen). Levels of miRNA expression were mea-sured quantitatively by using the TaqMan MicroRNA Assay (AppliedBiosystems) following the manufacturer’s protocol and assayed on theApplied Biosystems 7900HT Real-Time PCR System. Normalization wasperformed using U6 small nuclear RNA. Relative expression was cal-culated using the comparative threshold cycle method as previously de-scribed (13).
Quantitative RT-PCR for mRNA
Total RNAwas reverse transcribed using the High Capacity cDNA ReverseTranscription kit (Applied Biosystems). All primer/probe sets were ob-tained from Applied Biosystems. Samples were analyzed by TaqManquantitative RT-PCR for Ccr3 (Assay ID: Mm01216172_m1) and nor-malized to Hprt1 (Assay ID: Mm00446968_m1). Relative expression wascalculated using the comparative threshold cycle method.
Flow cytometry analysis of eosinophil surface CCR3expression
One million cultured eosinophil progenitor cells were stained with CCR3-FITC (R&D Systems) and Siglec-F-PE (BD Bioscience), as CCR3 andSiglec-F are markers for mature eosinophils. Staining was performed onice for 30 min in staining buffer (0.5% BSA, 0.01% NaN3 in 13 HBSS)according to manufacturer’s protocol. Data were acquired on a BD FACSCanto I flow cytometer and analyzed using FlowJo.
Preparation of total cell lysates and Western blot
Cells were rapidly washed in PBS and lysed in M-PER mammalian proteinextraction reagent (Pierce) according to manufacturer’s protocol. Proteaseinhibitor cocktails (Pierce) and phosphatase inhibitor cocktails (Pierce)were added to the M-PER protein extraction reagent immediately beforelysis. Western blot analysis was performed as previously described (14).The anti-IGF1R Ab was obtained from Cell Signaling Technology. Theanti-GAPDH Ab was from Abcam.
Analysis of cell proliferation after picropodophyllin treatment
Bone marrow–derived eosinophils were resuspended at a concentration of1 3 106 cells/ml and treated with DMSO or 2 mM picropodophyllin at day8 (15). Cell proliferation was determined by cell counting using a hema-cytometer on days 10 and 12. Cell lysates were collected on day 10, andlevels of IGF1R expression were determined by Western blot. To deter-mine a dose–response curve of picropodophyllin, bone marrow–derivedeosinophils were resuspended at a concentration of 1 3 106 cells/ml andplated in a 96-well plate at 100 ml/well on day 9. The cells were treatedwith increasing concentrations of picropodophyllin, and the level of cellproliferation was determined using Cell-Titer Glo luminescent cell viabilityassay (Promega) according to manufacturer’s protocol.
Mouse genome-wide mRNA microarray
The Affymetrix Mouse Gene 1.0ST array was used to compare gene ex-pression profiles between miR-223+/+ and miR-2232/2 eosinophil pro-genitor cultures at days 4, 8, and 12. Microarray data were analyzed usingthe GeneSpring software (Agilent Technologies). Global scaling was per-formed to compare genes from chip to chip, and a base set of probes
was generated by requiring a minimum raw expression level of 20th per-centile out of all probes on the microarray. The resulting probe sets werethen baseline transformed and filtered on at least 1.5-fold difference be-tween miR-223+/+ and miR-2232/2 eosinophil progenitor cultures. Sta-tistical significance was determined at p , 0.05 with Benjamini–Hochbergfalse discovery rate correction. The resulting list of genes was clusteredusing hierarchical clustering, and a heat map was generated. Biologicalfunctional enrichment analysis was carried out using Ingenuity PathwayAnalysis (Ingenuity Systems) and ToppGene/ToppCluster (16, 17). Themicroarray data have been deposited into the Array Express database(http://www.ebi.ac.uk/arrayexpress) with accession number E-MEXP-3350 in compliance with minimum information about microarray experimentstandards.
Statistical analysis
Statistical analyses were performed with Student t test or one-way ANOVAwith Tukey post hoc test where appropriate. Statistical significance andthe p values were indicated on the figures where appropriate. The p values,0.05 were considered statistically significant.
ResultsExpression of miR-223 in an ex vivo culture of bone marrow–derived eosinophils
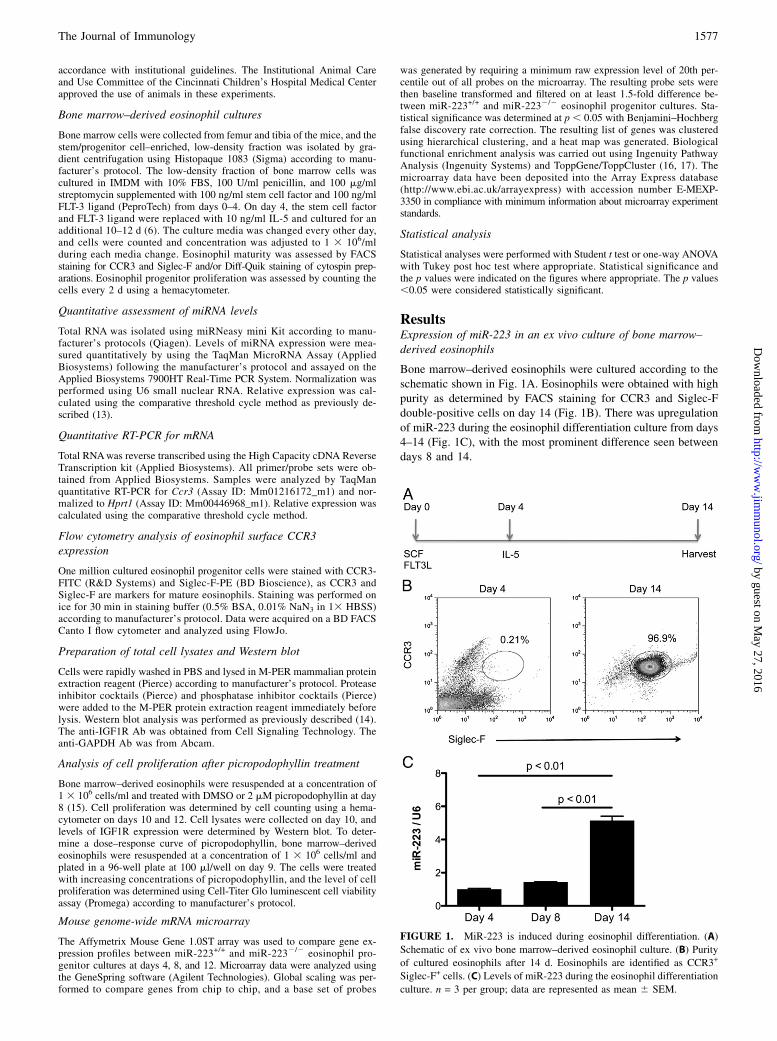
Bone marrow–derived eosinophils were cultured according to theschematic shown in Fig. 1A. Eosinophils were obtained with highpurity as determined by FACS staining for CCR3 and Siglec-Fdouble-positive cells on day 14 (Fig. 1B). There was upregulationof miR-223 during the eosinophil differentiation culture from days4–14 (Fig. 1C), with the most prominent difference seen betweendays 8 and 14.
FIGURE 1. MiR-223 is induced during eosinophil differentiation. (A)
Schematic of ex vivo bone marrow–derived eosinophil culture. (B) Purity
of cultured eosinophils after 14 d. Eosinophils are identified as CCR3+
Siglec-F+ cells. (C) Levels of miR-223 during the eosinophil differentiation
culture. n = 3 per group; data are represented as mean 6 SEM.
The Journal of Immunology 1577
by guest on May 27, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
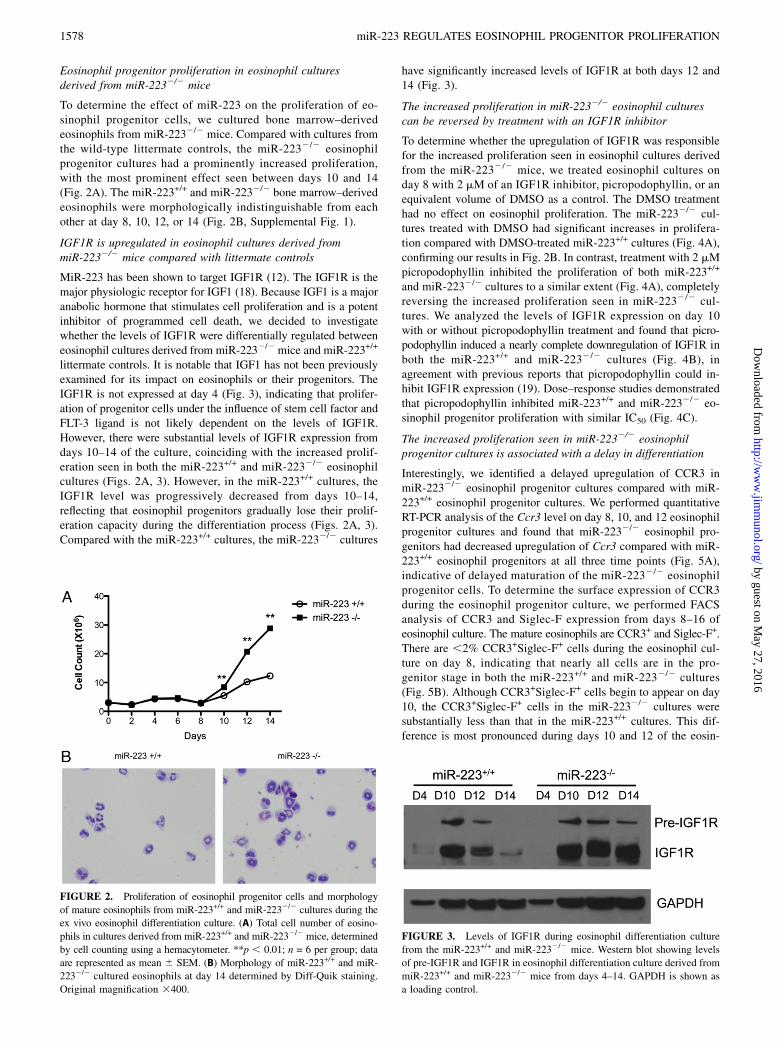
Eosinophil progenitor proliferation in eosinophil culturesderived from miR-2232/2 mice
To determine the effect of miR-223 on the proliferation of eo-sinophil progenitor cells, we cultured bone marrow–derivedeosinophils from miR-2232/2 mice. Compared with cultures fromthe wild-type littermate controls, the miR-2232/2 eosinophilprogenitor cultures had a prominently increased proliferation,with the most prominent effect seen between days 10 and 14(Fig. 2A). The miR-223+/+ and miR-2232/2 bone marrow–derivedeosinophils were morphologically indistinguishable from eachother at day 8, 10, 12, or 14 (Fig. 2B, Supplemental Fig. 1).
IGF1R is upregulated in eosinophil cultures derived frommiR-2232/2 mice compared with littermate controls
MiR-223 has been shown to target IGF1R (12). The IGF1R is themajor physiologic receptor for IGF1 (18). Because IGF1 is a majoranabolic hormone that stimulates cell proliferation and is a potentinhibitor of programmed cell death, we decided to investigatewhether the levels of IGF1R were differentially regulated betweeneosinophil cultures derived from miR-2232/2 mice and miR-223+/+
littermate controls. It is notable that IGF1 has not been previouslyexamined for its impact on eosinophils or their progenitors. TheIGF1R is not expressed at day 4 (Fig. 3), indicating that prolifer-ation of progenitor cells under the influence of stem cell factor andFLT-3 ligand is not likely dependent on the levels of IGF1R.However, there were substantial levels of IGF1R expression fromdays 10–14 of the culture, coinciding with the increased prolif-eration seen in both the miR-223+/+ and miR-2232/2 eosinophilcultures (Figs. 2A, 3). However, in the miR-223+/+ cultures, theIGF1R level was progressively decreased from days 10–14,reflecting that eosinophil progenitors gradually lose their prolif-eration capacity during the differentiation process (Figs. 2A, 3).Compared with the miR-223+/+ cultures, the miR-2232/2 cultures
have significantly increased levels of IGF1R at both days 12 and14 (Fig. 3).
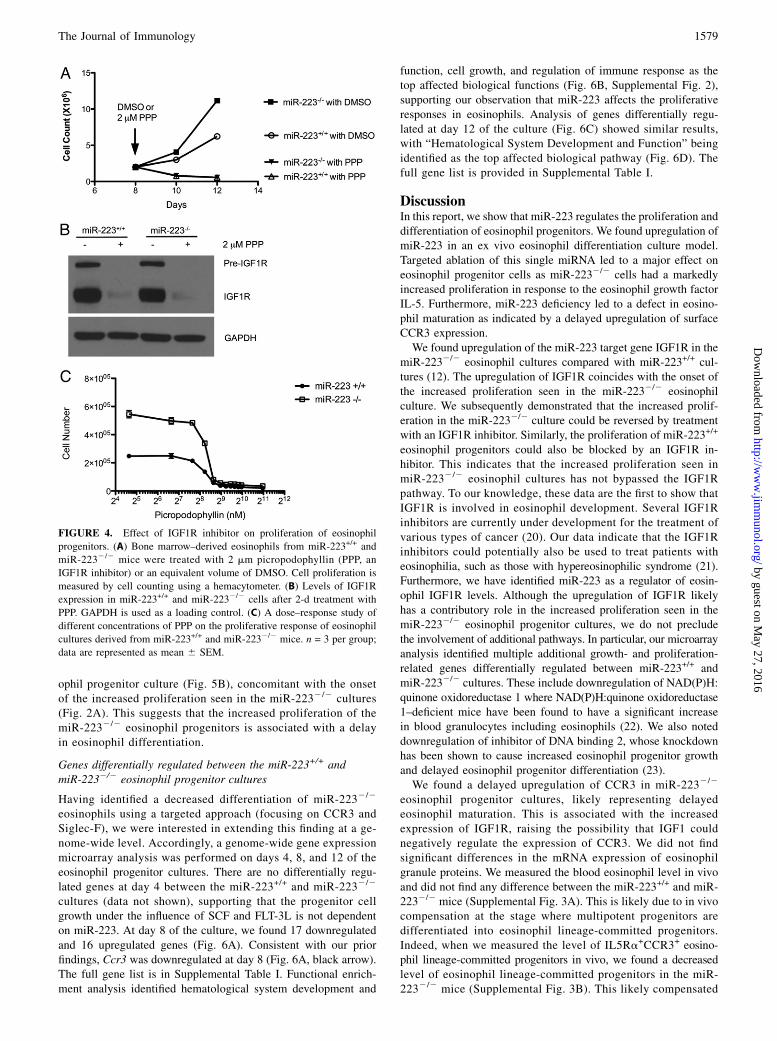
The increased proliferation in miR-2232/2 eosinophil culturescan be reversed by treatment with an IGF1R inhibitor
To determine whether the upregulation of IGF1R was responsiblefor the increased proliferation seen in eosinophil cultures derivedfrom the miR-2232/2 mice, we treated eosinophil cultures onday 8 with 2 mM of an IGF1R inhibitor, picropodophyllin, or anequivalent volume of DMSO as a control. The DMSO treatmenthad no effect on eosinophil proliferation. The miR-2232/2 cul-tures treated with DMSO had significant increases in prolifera-tion compared with DMSO-treated miR-223+/+ cultures (Fig. 4A),confirming our results in Fig. 2B. In contrast, treatment with 2 mMpicropodophyllin inhibited the proliferation of both miR-223+/+
and miR-2232/2 cultures to a similar extent (Fig. 4A), completelyreversing the increased proliferation seen in miR-2232/2 cul-tures. We analyzed the levels of IGF1R expression on day 10with or without picropodophyllin treatment and found that picro-podophyllin induced a nearly complete downregulation of IGF1R inboth the miR-223+/+ and miR-2232/2 cultures (Fig. 4B), inagreement with previous reports that picropodophyllin could in-hibit IGF1R expression (19). Dose–response studies demonstratedthat picropodophyllin inhibited miR-223+/+ and miR-2232/2 eo-sinophil progenitor proliferation with similar IC50 (Fig. 4C).
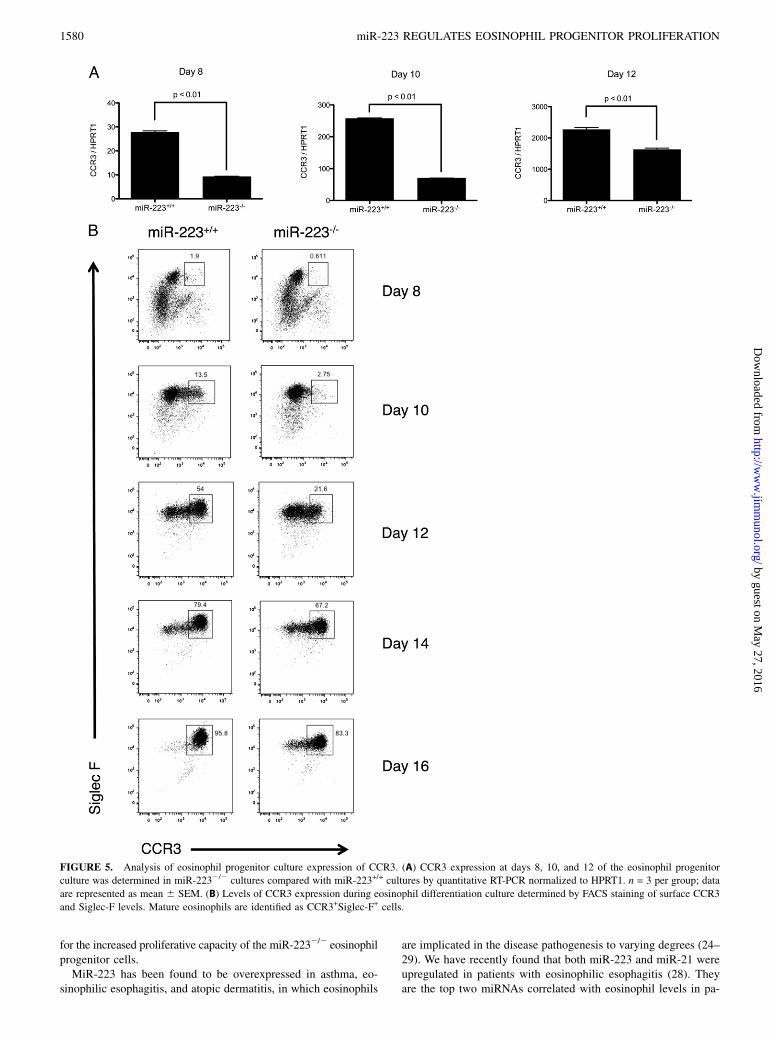
The increased proliferation seen in miR-2232/2 eosinophilprogenitor cultures is associated with a delay in differentiation
Interestingly, we identified a delayed upregulation of CCR3 inmiR-2232/2 eosinophil progenitor cultures compared with miR-223+/+ eosinophil progenitor cultures. We performed quantitativeRT-PCR analysis of the Ccr3 level on day 8, 10, and 12 eosinophilprogenitor cultures and found that miR-2232/2 eosinophil pro-genitors had decreased upregulation of Ccr3 compared with miR-223+/+ eosinophil progenitors at all three time points (Fig. 5A),indicative of delayed maturation of the miR-2232/2 eosinophilprogenitor cells. To determine the surface expression of CCR3during the eosinophil progenitor culture, we performed FACSanalysis of CCR3 and Siglec-F expression from days 8–16 ofeosinophil culture. The mature eosinophils are CCR3+ and Siglec-F+.There are ,2% CCR3+Siglec-F+ cells during the eosinophil cul-ture on day 8, indicating that nearly all cells are in the pro-genitor stage in both the miR-223+/+ and miR-2232/2 cultures(Fig. 5B). Although CCR3+Siglec-F+ cells begin to appear on day10, the CCR3+Siglec-F+ cells in the miR-2232/2 cultures weresubstantially less than that in the miR-223+/+ cultures. This dif-ference is most pronounced during days 10 and 12 of the eosin-
FIGURE 2. Proliferation of eosinophil progenitor cells and morphology
of mature eosinophils from miR-223+/+ and miR-2232/2 cultures during the
ex vivo eosinophil differentiation culture. (A) Total cell number of eosino-
phils in cultures derived from miR-223+/+ and miR-2232/2 mice, determined
by cell counting using a hemacytometer. **p , 0.01; n = 6 per group; data
are represented as mean 6 SEM. (B) Morphology of miR-223+/+ and miR-
2232/2 cultured eosinophils at day 14 determined by Diff-Quik staining.
Original magnification 3400.
FIGURE 3. Levels of IGF1R during eosinophil differentiation culture
from the miR-223+/+ and miR-2232/2 mice. Western blot showing levels
of pre-IGF1R and IGF1R in eosinophil differentiation culture derived from
miR-223+/+ and miR-2232/2 mice from days 4–14. GAPDH is shown as
a loading control.
1578 miR-223 REGULATES EOSINOPHIL PROGENITOR PROLIFERATION
by guest on May 27, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
ophil progenitor culture (Fig. 5B), concomitant with the onsetof the increased proliferation seen in the miR-2232/2 cultures(Fig. 2A). This suggests that the increased proliferation of themiR-2232/2 eosinophil progenitors is associated with a delayin eosinophil differentiation.
Genes differentially regulated between the miR-223+/+ andmiR-2232/2 eosinophil progenitor cultures
Having identified a decreased differentiation of miR-2232/2
eosinophils using a targeted approach (focusing on CCR3 andSiglec-F), we were interested in extending this finding at a ge-nome-wide level. Accordingly, a genome-wide gene expressionmicroarray analysis was performed on days 4, 8, and 12 of theeosinophil progenitor cultures. There are no differentially regu-lated genes at day 4 between the miR-223+/+ and miR-2232/2
cultures (data not shown), supporting that the progenitor cellgrowth under the influence of SCF and FLT-3L is not dependenton miR-223. At day 8 of the culture, we found 17 downregulatedand 16 upregulated genes (Fig. 6A). Consistent with our priorfindings, Ccr3 was downregulated at day 8 (Fig. 6A, black arrow).The full gene list is in Supplemental Table I. Functional enrich-ment analysis identified hematological system development and
function, cell growth, and regulation of immune response as thetop affected biological functions (Fig. 6B, Supplemental Fig. 2),supporting our observation that miR-223 affects the proliferativeresponses in eosinophils. Analysis of genes differentially regu-lated at day 12 of the culture (Fig. 6C) showed similar results,with “Hematological System Development and Function” beingidentified as the top affected biological pathway (Fig. 6D). Thefull gene list is provided in Supplemental Table I.
DiscussionIn this report, we show that miR-223 regulates the proliferation anddifferentiation of eosinophil progenitors. We found upregulation ofmiR-223 in an ex vivo eosinophil differentiation culture model.Targeted ablation of this single miRNA led to a major effect oneosinophil progenitor cells as miR-2232/2 cells had a markedlyincreased proliferation in response to the eosinophil growth factorIL-5. Furthermore, miR-223 deficiency led to a defect in eosino-phil maturation as indicated by a delayed upregulation of surfaceCCR3 expression.We found upregulation of the miR-223 target gene IGF1R in the
miR-2232/2 eosinophil cultures compared with miR-223+/+ cul-tures (12). The upregulation of IGF1R coincides with the onset ofthe increased proliferation seen in the miR-2232/2 eosinophilculture. We subsequently demonstrated that the increased prolif-eration in the miR-2232/2 culture could be reversed by treatmentwith an IGF1R inhibitor. Similarly, the proliferation of miR-223+/+
eosinophil progenitors could also be blocked by an IGF1R in-hibitor. This indicates that the increased proliferation seen inmiR-2232/2 eosinophil cultures has not bypassed the IGF1Rpathway. To our knowledge, these data are the first to show thatIGF1R is involved in eosinophil development. Several IGF1Rinhibitors are currently under development for the treatment ofvarious types of cancer (20). Our data indicate that the IGF1Rinhibitors could potentially also be used to treat patients witheosinophilia, such as those with hypereosinophilic syndrome (21).Furthermore, we have identified miR-223 as a regulator of eosin-ophil IGF1R levels. Although the upregulation of IGF1R likelyhas a contributory role in the increased proliferation seen in themiR-2232/2 eosinophil progenitor cultures, we do not precludethe involvement of additional pathways. In particular, our microarrayanalysis identified multiple additional growth- and proliferation-related genes differentially regulated between miR-223+/+ andmiR-2232/2 cultures. These include downregulation of NAD(P)H:quinone oxidoreductase 1 where NAD(P)H:quinone oxidoreductase1–deficient mice have been found to have a significant increasein blood granulocytes including eosinophils (22). We also noteddownregulation of inhibitor of DNA binding 2, whose knockdownhas been shown to cause increased eosinophil progenitor growthand delayed eosinophil progenitor differentiation (23).We found a delayed upregulation of CCR3 in miR-2232/2
eosinophil progenitor cultures, likely representing delayedeosinophil maturation. This is associated with the increasedexpression of IGF1R, raising the possibility that IGF1 couldnegatively regulate the expression of CCR3. We did not findsignificant differences in the mRNA expression of eosinophilgranule proteins. We measured the blood eosinophil level in vivoand did not find any difference between the miR-223+/+ and miR-2232/2 mice (Supplemental Fig. 3A). This is likely due to in vivocompensation at the stage where multipotent progenitors aredifferentiated into eosinophil lineage-committed progenitors.Indeed, when we measured the level of IL5Ra+CCR3+ eosino-phil lineage-committed progenitors in vivo, we found a decreasedlevel of eosinophil lineage-committed progenitors in the miR-2232/2 mice (Supplemental Fig. 3B). This likely compensated
FIGURE 4. Effect of IGF1R inhibitor on proliferation of eosinophil
progenitors. (A) Bone marrow–derived eosinophils from miR-223+/+ and
miR-2232/2 mice were treated with 2 mm picropodophyllin (PPP, an
IGF1R inhibitor) or an equivalent volume of DMSO. Cell proliferation is
measured by cell counting using a hemacytometer. (B) Levels of IGF1R
expression in miR-223+/+ and miR-2232/2 cells after 2-d treatment with
PPP. GAPDH is used as a loading control. (C) A dose–response study of
different concentrations of PPP on the proliferative response of eosinophil
cultures derived from miR-223+/+ and miR-2232/2 mice. n = 3 per group;
data are represented as mean 6 SEM.
The Journal of Immunology 1579
by guest on May 27, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
for the increased proliferative capacity of the miR-2232/2 eosinophilprogenitor cells.MiR-223 has been found to be overexpressed in asthma, eo-
sinophilic esophagitis, and atopic dermatitis, in which eosinophils
are implicated in the disease pathogenesis to varying degrees (24–29). We have recently found that both miR-223 and miR-21 wereupregulated in patients with eosinophilic esophagitis (28). Theyare the top two miRNAs correlated with eosinophil levels in pa-
FIGURE 5. Analysis of eosinophil progenitor culture expression of CCR3. (A) CCR3 expression at days 8, 10, and 12 of the eosinophil progenitor
culture was determined in miR-2232/2 cultures compared with miR-223+/+ cultures by quantitative RT-PCR normalized to HPRT1. n = 3 per group; data
are represented as mean 6 SEM. (B) Levels of CCR3 expression during eosinophil differentiation culture determined by FACS staining of surface CCR3
and Siglec-F levels. Mature eosinophils are identified as CCR3+Siglec-F+ cells.
1580 miR-223 REGULATES EOSINOPHIL PROGENITOR PROLIFERATION
by guest on May 27, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from
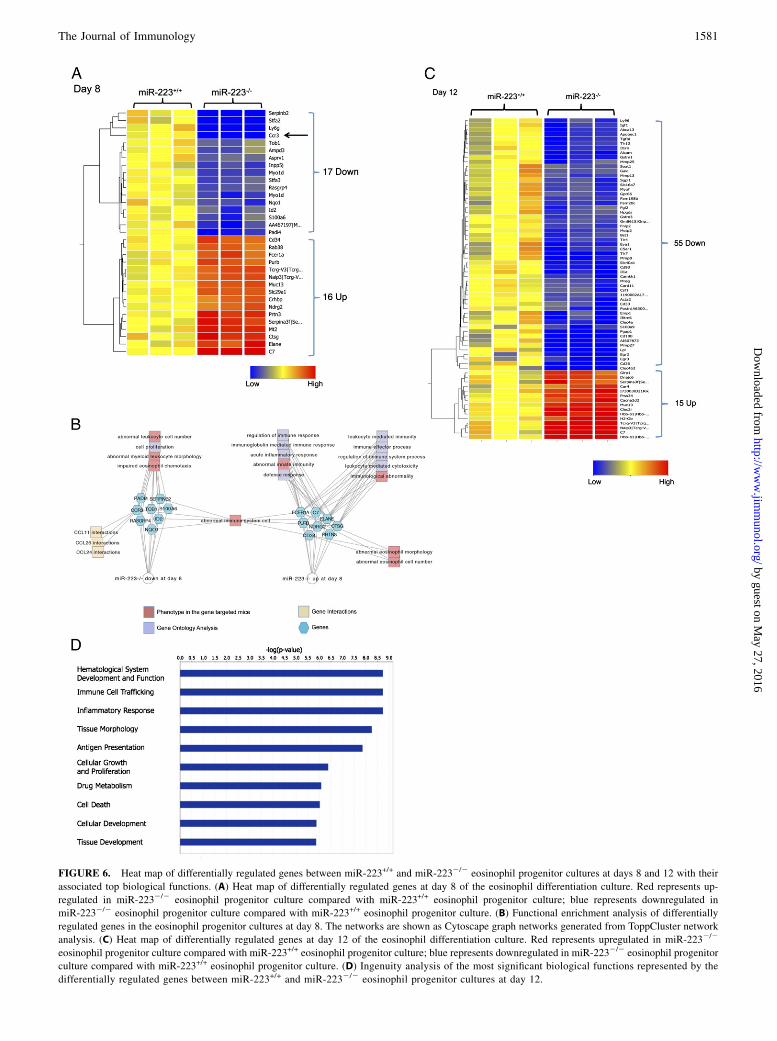
FIGURE 6. Heat map of differentially regulated genes between miR-223+/+ and miR-2232/2 eosinophil progenitor cultures at days 8 and 12 with their
associated top biological functions. (A) Heat map of differentially regulated genes at day 8 of the eosinophil differentiation culture. Red represents up-
regulated in miR-2232/2 eosinophil progenitor culture compared with miR-223+/+ eosinophil progenitor culture; blue represents downregulated in
miR-2232/2 eosinophil progenitor culture compared with miR-223+/+ eosinophil progenitor culture. (B) Functional enrichment analysis of differentially
regulated genes in the eosinophil progenitor cultures at day 8. The networks are shown as Cytoscape graph networks generated from ToppCluster network
analysis. (C) Heat map of differentially regulated genes at day 12 of the eosinophil differentiation culture. Red represents upregulated in miR-2232/2
eosinophil progenitor culture compared with miR-223+/+ eosinophil progenitor culture; blue represents downregulated in miR-2232/2 eosinophil progenitor
culture compared with miR-223+/+ eosinophil progenitor culture. (D) Ingenuity analysis of the most significant biological functions represented by the
differentially regulated genes between miR-223+/+ and miR-2232/2 eosinophil progenitor cultures at day 12.
The Journal of Immunology 1581
by guest on May 27, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

tient esophageal biopsies. Using systems biology analysis, wefound that miR-223 and miR-21 coregulated a set of interactingtarget genes involved in eosinophil proliferation and differentia-tion (28). MiR-21 has been known to promote cell proliferationin various cell types by downregulating a variety of proapoptoticgenes both directly and indirectly (30). The upregulation ofmiR-223 likely provides a check and balance in the system giventhe ability of miR-223 to promote eosinophil maturation. Further-more, a recent report showed that miR-21*, a complementarymiRNA of miR-21, was upregulated after GM-CSF treatmentand could inhibit the apoptosis of eosinophils (10). This indicatesthat the minor miRNAs could also have a role in regulating theproliferation of eosinophil progenitors, adding another level ofcomplexity. Future therapies targeting miRNAs, including miR-21and miR-223 and their minor miR* forms, will likely allow fine-tuning of the eosinophil level in various diseases.In summary, we have identified miR-223 as a regulator of eo-
sinophil progenitor proliferation. We found that IGF1R is upreg-ulated during eosinophil development, and miR-223 is a regulatorof IGF1R levels. Further elucidating and understanding the rolesand regulations of miRNAs during eosinophil development maylead to novel therapeutic targets for eosinophilic disorders.
AcknowledgmentsWe thank Dr. Eran Hornstein for providing the miR-223–deficient mice.
DisclosuresM.E.R. has equity interest in reslizumab through Cephalon, is a consultant
for and Chief Scientific Officer of Immune Pharmaceuticals, is on the
American Partnership for Eosinophilic Disorders Medical Advisory Board,
and is on the International Eosinophil Society’s Executive Council. The
other authors have no financial conflicts of interest.
References1. Broide, D. H., F. Finkelman, B. S. Bochner, and M. E. Rothenberg. 2011.
Advances in mechanisms of asthma, allergy, and immunology in 2010. J. AllergyClin. Immunol. 127: 689–695.
2. Venge, P. 2010. The eosinophil and airway remodelling in asthma. Clin. Respir.J. 4(Suppl. 1): 15–19.
3. Anthony, R. M., L. I. Rutitzky, J. F. Urban, Jr., M. J. Stadecker, and W. C. Gause.2007. Protective immune mechanisms in helminth infection. Nat. Rev. Immunol.7: 975–987.
4. Iwasaki, H., S. Mizuno, R. Mayfield, H. Shigematsu, Y. Arinobu, B. Seed,M. F. Gurish, K. Takatsu, and K. Akashi. 2005. Identification of eosinophillineage-committed progenitors in the murine bone marrow. J. Exp. Med. 201:1891–1897.
5. Hogan, S. P., H. F. Rosenberg, R. Moqbel, S. Phipps, P. S. Foster, P. Lacy,A. B. Kay, and M. E. Rothenberg. 2008. Eosinophils: biological properties androle in health and disease. Clin. Exp. Allergy 38: 709–750.
6. Dyer, K. D., J. M. Moser, M. Czapiga, S. J. Siegel, C. M. Percopo, andH. F. Rosenberg. 2008. Functionally competent eosinophils differentiated ex vivoin high purity from normal mouse bone marrow. J. Immunol. 181: 4004–4009.
7. Petriv, O. I., F. Kuchenbauer, A. D. Delaney, V. Lecault, A. White, D. Kent,L. Marmolejo, M. Heuser, T. Berg, M. Copley, et al. 2010. ComprehensivemicroRNA expression profiling of the hematopoietic hierarchy. Proc. Natl. Acad.Sci. USA 107: 15443–15448.
8. Navarro, F., and J. Lieberman. 2010. Small RNAs guide hematopoietic celldifferentiation and function. J. Immunol. 184: 5939–5947.
9. Rothenberg, M. E., and S. P. Hogan. 2006. The eosinophil. Annu. Rev. Immunol.24: 147–174.
10. Wong, C. K., K. M. Lau, I. H. Chan, S. Hu, Y. Y. Lam, A. O. Choi, and C. W.Lam. MicroRNA-21* regulates the prosurvival effect of GM-CSF on humaneosinophils. Immunobiology. In press.
11. Fukao, T., Y. Fukuda, K. Kiga, J. Sharif, K. Hino, Y. Enomoto, A. Kawamura,K. Nakamura, T. Takeuchi, and M. Tanabe. 2007. An evolutionarily conservedmechanism for microRNA-223 expression revealed by microRNA gene profil-ing. Cell 129: 617–631.
12. Johnnidis, J. B., M. H. Harris, R. T. Wheeler, S. Stehling-Sun, M. H. Lam,O. Kirak, T. R. Brummelkamp, M. D. Fleming, and F. D. Camargo. 2008.Regulation of progenitor cell proliferation and granulocyte function by micro-RNA-223. Nature 451: 1125–1129.
13. Livak, K. J., and T. D. Schmittgen. 2001. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(-Delta Delta C(T)) method.Methods 25: 402–408.
14. Lim, E. J., T. X. Lu, C. Blanchard, and M. E. Rothenberg. 2011. Epigeneticregulation of the IL-13-induced human eotaxin-3 gene by CREB-bindingprotein-mediated histone 3 acetylation. J. Biol. Chem. 286: 13193–13204.
15. Yin, S., A. Girnita, T. Stromberg, Z. Khan, S. Andersson, H. Zheng, C. Ericsson,M. Axelson, M. Nister, O. Larsson, et al. 2010. Targeting the insulin-like growthfactor-1 receptor by picropodophyllin as a treatment option for glioblastoma.Neuro-oncol. 12: 19–27.
16. Chen, J., E. E. Bardes, B. J. Aronow, and A. G. Jegga. 2009. ToppGene Suite forgene list enrichment analysis and candidate gene prioritization. Nucleic AcidsRes. 37(Web Server issue): W305–W311.
17. Kaimal, V., E. E. Bardes, S. C. Tabar, A. G. Jegga, and B. J. Aronow. 2010.ToppCluster: a multiple gene list feature analyzer for comparative enrichmentclustering and network-based dissection of biological systems. Nucleic AcidsRes. 38(Web Server issue): W96–W102.
18. Smith, T. J. 2010. Insulin-like growth factor-I regulation of immune function: a po-tential therapeutic target in autoimmune diseases? Pharmacol. Rev. 62: 199–236.
19. Vasilcanu, R., D. Vasilcanu, L. Rosengren, N. Natalishvili, B. Sehat, S. Yin,A. Girnita, M. Axelson, L. Girnita, and O. Larsson. 2008. Picropodophyllininduces downregulation of the insulin-like growth factor 1 receptor: potentialmechanistic involvement of Mdm2 and beta-arrestin1. Oncogene 27: 1629–1638.
20. Yee, D. 2012. Insulin-like growth factor receptor inhibitors: baby or the bath-water? J. Natl. Cancer Inst. 104: 975–981.
21. Arefi, M., J. L. Garcıa, M. M. Briz, F. de Arriba, J. N. Rodrıguez, G. Martın-Nunez, J. Martınez, J. Lopez, J. G. Suarez, M. J. Moreno, et al. 2012. Responseto imatinib mesylate in patients with hypereosinophilic syndrome. Int. J. Hem-atol. 96: 320–326.
22. Long, D. J., II, A. Gaikwad, A. Multani, S. Pathak, C. A. Montgomery,F. J. Gonzalez, and A. K. Jaiswal. 2002. Disruption of the NAD(P)H:quinoneoxidoreductase 1 (NQO1) gene in mice causes myelogenous hyperplasia. CancerRes. 62: 3030–3036.
23. Buitenhuis, M., H. W. van Deutekom, L. P. Verhagen, A. Castor, S. E. Jacobsen,J. W. Lammers, L. Koenderman, and P. J. Coffer. 2005. Differential regulation ofgranulopoiesis by the basic helix-loop-helix transcriptional inhibitors Id1 andId2. Blood 105: 4272–4281.
24. Lu, T. X., A. Munitz, and M. E. Rothenberg. 2009. MicroRNA-21 is up-regulated in allergic airway inflammation and regulates IL-12p35 expression.J. Immunol. 182: 4994–5002.
25. Garbacki, N., E. Di Valentin, V. A. Huynh-Thu, P. Geurts, A. Irrthum, C. Crahay,T. Arnould, C. Deroanne, J. Piette, D. Cataldo, and A. Colige. 2011. MicroRNAsprofiling in murine models of acute and chronic asthma: a relationship withmRNAs targets. PLoS ONE 6: e16509.
26. Sonkoly, E., P. Janson, M. L. Majuri, T. Savinko, N. Fyhrquist, L. Eidsmo,N. Xu, F. Meisgen, T. Wei, M. Bradley, et al. 2010. MiR-155 is overexpressed inpatients with atopic dermatitis and modulates T-cell proliferative responses bytargeting cytotoxic T lymphocyte-associated antigen 4. J. Allergy Clin. Immunol.126: 581–589.e1–e20.
27. Mattes, J., A. Collison, M. Plank, S. Phipps, and P. S. Foster. 2009. Antagonismof microRNA-126 suppresses the effector function of TH2 cells and the devel-opment of allergic airways disease. Proc. Natl. Acad. Sci. USA 106: 18704–18709.
28. Lu, T. X., J. D. Sherrill, T. Wen, A. J. Plassard, J. A. Besse, J. P. Abonia, J. P.Franciosi, P. E. Putnam, M. Eby, L. J. Martin, et al. 2012. MicroRNA signaturein patients with eosinophilic esophagitis, reversibility with glucocorticoids, andassessment as disease biomarkers. J. Allergy Clin. Immunol. 129: 1064–1075.e9.
29. Lu, S., V. A. Mukkada, S. Mangray, K. Cleveland, N. Shillingford, C. Schorl,A. S. Brodsky, and M. B. Resnick. 2012. MicroRNA profiling in mucosal bi-opsies of eosinophilic esophagitis patients pre and post treatment with steroidsand relationship with mRNA targets. PLoS ONE 7: e40676.
30. Buscaglia, L. E., and Y. Li. 2011. Apoptosis and the target genes of microRNA-21. Chin. J. Cancer 30: 371–380.
1582 miR-223 REGULATES EOSINOPHIL PROGENITOR PROLIFERATION
by guest on May 27, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from




















