
Preparation and Characterization of Monoclonal Antibodies to Human Neutrophil Cathepsin G,...
10
Journal of Leukocyte Biology 46:109-118 (1989) © 1989 Alan R. Lisa, Inc. Preparation and Characterization of Monoclonal Antibodies to Human Neutrophil Cathepsin G, Lactoferrin, Eosinophil Peroxidase, and Eosinophil Major Basic Protein Keith M. Skubitz, Neal P. Christiansen, and John R. Mendiola Department of Medicine, The University of Minnesota Medical School, and the Masonic Cancer Center, Minneapolis This report describes the production and characterization of five murine monoclonal antibodies that react with granule proteins of human granulocytes. Monoclonal antibody AHN-1 1 (lgG2a) reacted specifically with neutrophiI cathepsin G; no reactivity with the homologous neutrophll neutral proteases, elastase, protelnase 3, or esterase N was de- tected. Antibodies AHN-9 (IgGi) and AHN-9.1 (lgG2b) each reacted with different epitopes on human lactoferrin, but not with the homologous protein transferrin. Two IgGi anti- bodies, AHE-1 and AHE-2, reacted specifically with eosinophils; AHE-1 reacted strongly with eosinophIl peroxidase but not eoslnophii major basic protein while AHE-2 recog- nized eosinophil major basic protein but not eoslnophil peroxidase. All five antibodies could detect their respective antigens in alcohol-fixed cytospin preparations. These an- tibodies should be useful for immunolocalizatlon and quantification of their respective antigens as well as for other studies of the roles of these proteins in granulocyte function and differentiation. Key words: granulocyte, protease, immunochemical staining INTRODUCTION Human granulocytes contain a variety of biologically active molecules located within several different types of granules. These granule contents are believed to play an important role in both normal host defense as well as in the pathogenesis of a variety of disease processes [1,9,11]. The two most widely studied granules of neu- trophils are the primary and secondary granules. Primary granules contain a variety of proteases including elas- tase, cathepsin G, and the more recently described serine proteases, esterase N and proteinase 3 [1,4,14,28]. In some situations, these proteases may be released and play a role in the pathogenesis of a variety of diseases such as chronic lung disease [11,12,21,27], the adult respiratory distress syndrome [7,9,16,17], and destruc- tive joint diseases [20]. While studies of the role of neu- trophil proteases in pathologic states such as emphysema have focused on elastase, other neutrophil proteases may also play an important role and have not yet been thor- oughly evaluated. Indeed, several of these proteases are structurally related. For example, cathepsin G, elastase, and proteinase 3 share a high degree of homology in their N-terminal amino acid sequences [29,31]. Thus, assays of cathepsin G activity often detect other neutrophil pro- teases, and polyclonal antisera to cathepsin G may also react with these related proteases. The major component of secondary granules is lactoferrin, a protein that exerts a regulatory influence on myelopoiesis by feedback in- hibition, has an antibacterial action, and also can mod- ulate several immune responses [2,25,26]. Eosinophils also contain several granule proteins, including some that are specific to the eosinophil. Among these are eosinophi! peroxidase and eosinophil major basic pro- tein, which are believed to play important roles in de- fense against parasitic diseases as well as a role in the injury of normal tissue in some allergic and hypereosino- philic states [8]. This report describes the production and characterization of five murine monoclonal antibodies that react specifically with granule proteins. AHN-9 (an IgG 1) and AHN-9. 1 (an IgG2b) both react strongly with lactoferrin but not the related protein transferrin. The Received December 19, 1988; accepted February 24, 1989. Neal P. Christiansen’s current address is Department of Medicine, University of Tennessee, Memphis, TN 38163. Reprint requests: Keith Skubitz, Box 325, University Hospital, Min- neapolis. MN 55455.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Preparation and Characterization of Monoclonal Antibodies to Human Neutrophil Cathepsin G,...

Journal of Leukocyte Biology 46:109-118 (1989)
© 1989 Alan R. Lisa, Inc.
Preparation and Characterization of MonoclonalAntibodies to Human Neutrophil Cathepsin G,
Lactoferrin, Eosinophil Peroxidase, andEosinophil Major Basic Protein
Keith M. Skubitz, Neal P. Christiansen, and John R. MendiolaDepartment of Medicine, The University of Minnesota Medical School, and the Masonic
Cancer Center, Minneapolis
This report describes the production and characterization of five murine monoclonalantibodies that react with granule proteins of human granulocytes. Monoclonal antibodyAHN-1 1 (lgG2a) reacted specifically with neutrophiI cathepsin G; no reactivity with the
homologous neutrophll neutral proteases, elastase, protelnase 3, or esterase N was de-tected. Antibodies AHN-9 (IgGi) and AHN-9.1 (lgG2b) each reacted with different epitopes
on human lactoferrin, but not with the homologous protein transferrin. Two IgGi anti-bodies, AHE-1 and AHE-2, reacted specifically with eosinophils; AHE-1 reacted stronglywith eosinophIl peroxidase but not eoslnophii major basic protein while AHE-2 recog-nized eosinophil major basic protein but not eoslnophil peroxidase. All five antibodiescould detect their respective antigens in alcohol-fixed cytospin preparations. These an-tibodies should be useful for immunolocalizatlon and quantification of their respectiveantigens as well as for other studies of the roles of these proteins in granulocyte functionand differentiation.
Key words: granulocyte, protease, immunochemical staining
INTRODUCTION
Human granulocytes contain a variety of biologically
active molecules located within several different types of
granules. These granule contents are believed to play an
important role in both normal host defense as well as in
the pathogenesis of a variety of disease processes
[1,9,11]. The two most widely studied granules of neu-trophils are the primary and secondary granules. Primary
granules contain a variety of proteases including elas-
tase, cathepsin G, and the more recently described serine
proteases, esterase N and proteinase 3 [1,4,14,28]. In
some situations, these proteases may be released and
play a role in the pathogenesis of a variety of diseases
such as chronic lung disease [11,12,21,27], the adult
respiratory distress syndrome [7,9,16,17], and destruc-tive joint diseases [20]. While studies of the role of neu-
trophil proteases in pathologic states such as emphysemahave focused on elastase, other neutrophil proteases mayalso play an important role and have not yet been thor-
oughly evaluated. Indeed, several of these proteases arestructurally related. For example, cathepsin G, elastase,
and proteinase 3 share a high degree of homology in theirN-terminal amino acid sequences [29,31]. Thus, assaysof cathepsin G activity often detect other neutrophil pro-
teases, and polyclonal antisera to cathepsin G may also
react with these related proteases. The major componentof secondary granules is lactoferrin, a protein that exerts
a regulatory influence on myelopoiesis by feedback in-
hibition, has an antibacterial action, and also can mod-
ulate several immune responses [2,25,26]. Eosinophils
also contain several granule proteins, including somethat are specific to the eosinophil. Among these are
eosinophi! peroxidase and eosinophil major basic pro-tein, which are believed to play important roles in de-
fense against parasitic diseases as well as a role in theinjury of normal tissue in some allergic and hypereosino-
philic states [8]. This report describes the production and
characterization of five murine monoclonal antibodies
that react specifically with granule proteins. AHN-9 (anIgG 1) and AHN-9. 1 (an IgG2b) both react strongly withlactoferrin but not the related protein transferrin. The
Received December 19, 1988; accepted February 24, 1989.
Neal P. Christiansen’s current address is Department of Medicine,
University of Tennessee, Memphis, TN 38163.
Reprint requests: Keith Skubitz, Box 325, University Hospital, Min-neapolis. MN 55455.

110 Skubitz et al.
IgG2a monoclonal antibody AHN- 11 reacts with human
neutrophil cathepsin G but not with the other major neu-trophil neutral proteases: elastase, proteinase 3, or the
newly purified neutral protease, esterase N [23,31]. Twoeosinophil-specific monoclonal antibodies, AHE- 1
(IgGi) and AHE-2 (IgGl), react specifically with
eosinophil peroxidase and eosinophil major basic pro-
tein, respectively.
MATERIALS AND METHODSPreparation of Peripheral Blood Cells
Neutrophils, red blood cells, lymphocytes, mono-cytes, and platelets were prepared from heparinized (2units/mi) human venous blood as described [24]. Eosino-phils from a patient with the hypereosinophilic syndrome
were a gift of Dr. D. Hurd, University of Minnesota
Medical School, Minneapolis.
Immunization and Production ofHybridoma Cell Lines
Monoclonal antibodies were prepared as previously
described [24]. Briefly, for the production of antibodies
AHN-9, AHN-9.l, and AHN-ll, 8 wk-old femaleBALB/cJ mice (Jackson Laboratories, Bar Harbor, ME)
were immunized by intrapentoneal injection of 1O� hu-
man neutrophils in Freund’s complete adjuvant (GIBCO
Laboratories, Grand Island, NY) and subsequently sub-
cutaneously, at weekly intervals, with l0� neutrophils inincomplete Freund’s adjuvant (GIBCO). One week fol-
lowing the third immunization, mice received 1o� neu-trophils in phosphate-buffered saline, pH 7.2 (PBS) in-
traperitoneally. Four days later, their spleen cells were
fused with murine myeloma P3-X63-Ag8.653 cells at aratio of four spleen cells per myeloma cell, as previouslydescribed [24]. Hybridoma cell cultures secreting anti-bodies that bound to neutrophils were twice cloned in
soft agar as described [24]. Immunoglobulin isotype wasdetermined by using a commercially available isotypingkit (Bio Rad, Richmond, CA). A similar procedure wasused for the production of AHE- 1, AHE-2, and the CG
and EN series of antibodies, except that in the case ofAHE- 1, eosinophils from a patient with the hypereosino-
philic syndrome were used instead of neutrophils. Forthe production of AHE-2, 50 �g of protein purified from
these hypereosinophilic eosinophils by wheat germ af-finity chromatography [5] was used in place of io� neu-trophils for immunization, and 0.5 �ig of this protein
preparation was used in each solid-phase assay for su-pernatant screening. For the production of the CG andEN series of antibodies, 10-50 �ig of purified cathepsin
G or esterase N, respectively, was used in place of l0�neutrophils for each immunization, the first and last be-ing intraperitoneal immunizations with other immuniza-
tions being subcutaneous, and 0.5 p.g of these proteinswas used in each solid-phase antibody-binding assay for
supernatant screening.
Antibody-Binding Assays
Antibodies were assayed for cell-binding activity by asolid-phase assay, as described [24]. Briefly, target cellswere suspended at 106/ml in PBS and 0.02% NaN3, and
50i�jaliquots were applied to wells of a Linbro 96-wellmicrotiter plate and dried overnight at 23#{176}C.Two hun-
dred microliters of PBS containing 50 mg/mi BSA and0.02% NaN3 was then added to each well and incubatedfor 4 h at 23#{176}C.This solution was then removed, and 50�l of supernatant to be tested was added and incubated
for 1 h at 23#{176}C.Excess antibody was removed by wash-ing three times with 200 �i�i of ice-cold PBS containing 1mg/mi BSA and 0.1% Triton X-l0O; ‘251-labeled sheep
antimouse immunoglobulin (100,000 cpm, specific ac-
tivity 10-50 p�Ci/�g, Amersham, Arlington Heights, IL)was then added in 50 p.! of buffer containing 20 mM
Tris-HC1, pH 7.6, 1 mM EDTA, 100 mM NaC1, and 20
mg/mi BSA, and incubated for 1 h at 23#{176}C.After wash-
ing as above, 100 pA of 2 M NaOH was added and after
30 mm at 23#{176}C,the solution was transferred to glasstubes and the radioactivity was counted. Identical assay
procedures were also performed in which various puri-
fied proteins, as indicated, in 50 pA aliquots were sub-
stituted for target cells. Plasmin, elastase, lactoferrin,
transferrin, thrombin, and kallikrein were obtained from
Sigma. Human neutrophil elastase, cathepsin G, protein-
ase 3, and esterase N, purified as previously described[10,14,23,31], were gifts from Drs. B. Gray and N.Wehner, University of Minnesota, Minneapolis. Eosino-
phil peroxidase and eosinophil major basic protein were
gifts of Dr. A. Slungaard, University of Minnesota, Min-
neapolis.
Antibodies
Mouse antihuman neutrophil antiserum was preparedfrom CS7BL/6J mice as described [24]. Monoclonal an-
tibody P3, and IgG1 with no known specificity, wasobtained from the supernatant of the P3-X63-Ag8 cellline. Rabbit antimouse immunoglobulin was a gift of Dr.M. Wick, University of Minnesota Medical School. The
antihuman neutrophil elastase monocloncal antibodyAHN-lO (IgGi) has been previously described [23]. Thesource of monoclonal antibodies was either hybrid cellculture supernatants or ascites fluid diluted in PBS asindicated. All sera and ascites were heat inactivated at
56#{176}Cfor 30 mm and clarified by centrifugation at50,000g at 4#{176}Cfor 1 h before use.
Immunocytochemical Staining
Immunocytochemical studies were performed oncytospmn preparations by using the alkaline phosphatase-

4
Mnn8�l8nalAntiWin t� O�anul�vti Pr�tslna 111
,#{149}.#{149}
Fit. 1, lmmunQchQmicul �laiflIfl� 91 p9rlptIEaI �I99U c�II� �yantibody AllN-il. Cytospln preparations were reacted wIth con-trol culture supematant (panel A) or AHN-11 (panels B and C)and then staIned by using the APAAP technique as described Inthe text. The cells In panel A were from the bottom of a Ficoll-Hypaque gradient (neutrophil rich) to demonstrate lack of reac-tivity with control supernatant. All neutrophils, but not eoslno-phils, In this preparation reacted with AHN-1 1 (not Shown). Thecells In panels B and C were from the interface of a Ficoll-
mouse-anti-alkaline-phosphatase (APAAP) technique aspreviously described [6,23]. Briefly, cytospin prepara-tions were immersed in ethanol immediately after prep-
aration and stored at -20#{176}Cuntil use. The slides werethen rehydrated by dipping 40 times in each of the fol-lowing: two changes of ethanol, one change of 95% eth-
anol, one change of 70% ethanol, and three changes ofdistilled water. The slides were then blocked with 2%
normal rabbit serum in PBS for 10 mm at 23#{176}C.Appro-
priately diluted primary antibody (2 drops) was then ap-
plied and the slide was incubated at 23#{176}Cfor 2 h. The
slide was then rinsed first with PBS and thereafter with
0.05 M Tris-HC1, pH 7.6. Rabbit antimouse immuno-globulin diluted 1:40 with 0.1% BSA in PBS (35 p.1) was
then added and the slide was incubated for 20 mm at23#{176}Cin a humidified chamber. All subsequent incuba-tions were performed at 23#{176}Cin a humidified chamber.
The slide was then rinsed with 0.05 M Tris-HC1, pH 7.6,
and 35 p.1 of APAAP complex (Dako, Santa Barbara,
CA) diluted 1:25 with 0.05 M Tris-HC1, pH 7.6, wasadded. The slide was then incubated for 40 mm and
washed, and 35 p.1 of rabbit antimouse immunoglobulin(1:40) was added. After a 20 mm incubation, the slidewas washed and 35 pA of APAAP complex (1:25) wasadded, and the slide was incubated for another 20 mm.
The slide was then rinsed and freshly prepared substrate[2.6 mg Naphthol ASMX phosphate (Sigma), 200 p.1
N,N-dimethylformamide (Sigma), 9.8 ml of 0.5 M Tris-
HC1, pH 8.2, 2.4 mg Levamisole (Sigma), and 20 mg ofFast Red TR (Sigma)] was added. After incubation for15 mm, the slide was washed with water and counter-
stained with Harris Hematoxylin for 60 sec, washed with
MypaqusWraUlant(lymphocytemUmonocytsrich)toallowox.aminatlon of cells other than neutrophils and eosinophlls. AHN-11 staIned neutrophlls (arrows) but not lymphocytes, mono-cytes, eosInophils, or platelets. Since this exposure wasdesigned to optimally demonstrate the APAAP reaction prod-uct, the morphology of the neutrophils (identified by arrows inpanels B and C) is not clear; appropriate illumination demon-strated the characteristic nuclear structure of neutrophils. Barrepresents 20 jim.
water for 5 mm, dipped in saturated Li2CO3 five times,
and washed in water for 2 mm. Coverslips were then
mounted by using Glycergel (Dako) and the preparations
were examined by light microscopy.
RESULTSCharacterization of Antibody AHN-1 1
Antibody AHN-l 1 bound to normal human neutro-
phils by solid-phase radioimmunoassay but not to plate-
lets or erythrocytes (data not shown). Immunocytochem-ical studies demonstrated that monoclonal antibody
AHN-l 1 reacted strongly with neutrophils but did not
react with other peripheral blood cells, including plate-
lets, lymphocytes, or eosinophils (Fig. 1). Faint immu-nostaining of some monocytes was occasionally de-
tected. No binding of AHN-ll to the surface of livenormal human peripheral blood cells could be detectedby using flow cytofluorimetry, implying that the antigenwas not expressed on the surface of the cells. AHN- 11
was an IgG2a molecule, as determined by the use of
class-specific antisera in the solid-phase immunoassay.In an effort to characterize the antigens recognized bythese antibodies, fractions of primary granules, second-
ary granules, plasma membranes, and cytoplasm wereprepared from neutrophils by the technique of Borre-
gaard [3] and used as antigens in the solid-phase-bindingassay. AHN-l 1 reacted nearly exclusively with the pn-mary granule fraction as determined by the solid-phase-
binding assay (data not shown). These results led us totest the reactivity of AHN- 11 with elastase and cathepsin
G, the two major proteases of primary granules. AHN- 11
10000
8000
6000
4000
2000
0
C.,C,
CI,a.a
-Ca
C.)
a- a�aa auJ a
auJ
0auJ
aC C C C
E0a E � a
a � 0 -
a - .- - 0� a .� - -
oa
I VVV�J
8000
6000
4000
2000
B
U
0N
zuJ
N NN
ZLu
zLu
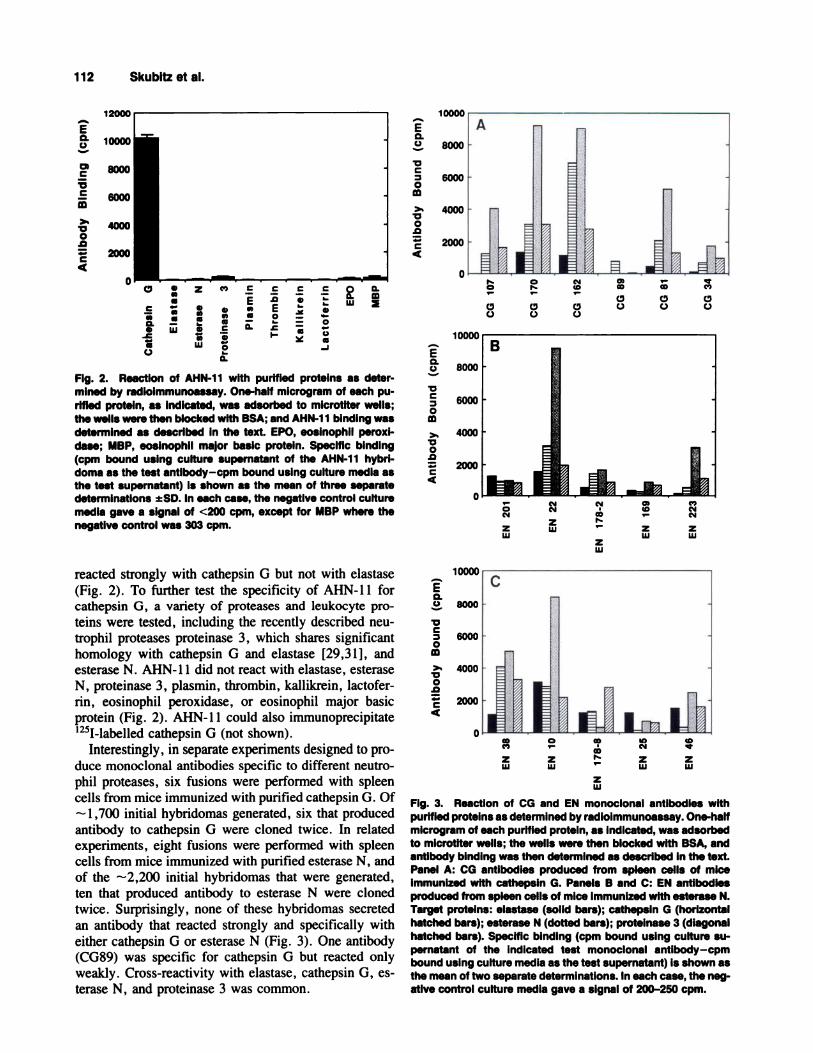
Fig. 2. ReactIon of AHN-1 1 with purified proteins as deter-mined by radioimmunoassay. One-half microgram of each pu-rified protein, as indicated, was adsorbed to microtiter wells;the wells were then blocked with BSA; and AHN-1 1 bindIng wasdetermined as described in the text. EPO, eosinophii peroxl-dase; MBP, eoslnophil major basic protein. Specific binding(cpm bound using culture supernatant of the AHN-11 hybri-doma as the test antlbody-cpm bound using culture media asthe test supernatant) is shown as the mean of three separatedeterminations ±SD. in each case, the negative control culturemedia gave a signal of <200 cpm, except for MBP where thenegative control was 303 cpm.
reacted strongly with cathepsin G but not with elastase
(Fig. 2). To further test the specificity of AHN-l 1 forcathepsin G, a variety of proteases and leukocyte pro-
teins were tested, including the recently described neu-
trophil proteases proteinase 3, which shares significanthomology with cathepsin G and elastase [29,31], and
esterase N. AHN-l 1 did not react with elastase, esteraseN, proteinase 3, plasmin, thrombin, kallikrein, lactofer-rim, eosinophil peroxidase, or eosinophil major basicprotein (Fig. 2). AHN-l 1 could also immunoprecipitate
‘251-labelled cathepsin G (not shown).Interestingly, in separate experiments designed to pro-
duce monoclonal antibodies specific to different neutro-phil proteases, six fusions were performed with spleen
cells from mice immunized with purified cathepsin G. Of
1,700 initial hybridomas generated, six that producedantibody to cathepsin G were cloned twice. In related
experiments, eight fusions were performed with spleencells from mice immunized with purified esterase N, and
of the -2,200 initial hybridomas that were generated,ten that produced antibody to esterase N were clonedtwice. Surprisingly, none of these hybridomas secreted
an antibody that reacted strongly and specifically witheither cathepsin G or esterase N (Fig. 3). One antibody
(CG89) was specific for cathepsin G but reacted onlyweakly. Cross-reactivity with elastase, cathepsin 0, es-terase N, and proteinase 3 was common.
0C)
z zLu Lu
U)N
zLu
Co
zLu
Co
Co
PS
zLu
FIg. 3. Reaction of CG and EN monoclonal antibodies withpurified proteins as determined by radloimmunoassay. One-halfmicrogram of each purified protein, as indicated, was adsorbedto microtlter wells; the wells were then blocked with BSA, andantibody binding was then determined as described in the text.Panel A: CG antibodies produced from spleen cells of mIceimmunized with cathepsin G. Panels B and C: EN antibodiesproduced from spleen cells of mica immunized with esterase N.Target proteins: elastase (solid bars); cathepsin G (horizontalhatched bars); esterase N (dotted bars); proteinase 3 (diagonalhatched bars). Specific binding (cpm bound using culture su-pernatant of the indicated test monoclonal antibody-cpmbound using culture media as the test supernatant) is shown asthe mean of two separate determinations. In each case, the neg-ative control culture media gave a signal of 200-250 cpm.
112 Skubitz et al.
E0.C.)
C
C
0.0
C
4
E0.C,)
C
0
�00
.0
C4
E0.C,)
C
0
�00
.0
C4
EaC)
�0
C
0
�00
.0
C4
C’)CO N
N
z zLu Lu

A
Ia
Monoclonal Antibodies to Granulocyte Proteins 113
I
.,y.
..-.. .‘ . S
$
;,
C I
Fig. 4. ImmunochemIcal staIning of peripheral blood cells byantibodies AHN-9 and AHN 9.1. Cytoapln preparations were re-acted with antIbodies AHN-9 (panels A and B) or AHN-9.1(panels C and D), and then stained by using the APAAP tech-nique as described In the text. The cells in panels A-C werefrom the Interface of a FIcoll-Hypaque gradient (lymphocyte andmonocyte rIch) to allow examination of cells other than neutro-phils and eoslnophlls. The cells In panel D were from the bottom
Characterization of AntibodiesAHN-9 and AHN-9.1
Antibodies AHN-9 and AHN-9. 1 both bound to nor-
mal human neutrophils by solid-phase radioimmumoas-say but not to platelets or erythrocytes (data not shown).
hnmunocytochemical studies demonstrated that both ofthese monoclonal antibodies reacted strongly with neu-
trophils but did not react with other peripheral bloodcells, including platelets, lymphocytes, monocytes, or
eosinophils (Fig. 4). Usually, no binding of these anti-bodies to the surface of live normal human peripheralblood cells could be detected by using flow cytofluorim-
etry, implying that the antigen was not expressed on thesurface of the cells; however, weak binding to the sur-face of 5-40% of neutrophils and 40-70% of monocytes
was observed in some cell preparations. This may havebeen due to adsorption of antigen released by neutrophils
during the cell preparation process. In most experiments,
AHN-9 and AHN-9. 1 did not immunoprecipitate any ra-
.#{149} w..
�0#{149}’�
Dsof a Ficoll-Hypaque gradient (neutrophil rich). Both antIbodIes
stained neutrophils but not lymphocytes, monocytes, eoslno-phils, or platelets. Since this exposure was designed to opti-mally demonstrate the APAAP reaction product, the morphol-ogy of the neutrophils (identifIed by arrows in panels A-C) isnot clear; appropriate Illumination demonstrated the character-istic nuclear structure of neutrophils. Bar represents 20 �m.
diolabeled proteins from extracts of neutrophils labeledat the cell surface with 1251 using lactoperoxidase. Incontrast, when cells were solubilized before 1251 label-
ing, both antibodies specifically and reproducibly immu-
noprecipitated a ‘251-labeled protein of -75-80 kd (notshown). AHN-9 was of the IgG 1 subtype, and AHN-9.
was an IgG2b, as determined by the use of class-specificantisera in the solid-phase immunoassay. Both AHN-9and AHN-9. 1 reacted almost exclusively with the sec-ondary granule fraction (data not shown). These results
led us to test their reactivity with lactoferrin, the majorconstituent of secondary granules. AHN-9 and AHN-9. 1
both reacted strongly with lactoferrin while other an-
tineutrophil monoclonal antibodies did not. To furthertest the specificity of these antibodies for lactoferrin, avariety of other leukocyte proteins were also tested, in-cluding transferrin, which shares significant homology
with lactoferrin [2], three neutrophil primary granuleproteins, and two eosmnophil granule proteins. AHN-9
C �
- ‘- C
o a a.- C aC) aa �. a
.J I- C.)
aaaaa
UI
C
.0
E0
I-
C
Eaaa
zaaaaa
Lu
0aLu
C
a
a
114 Skubitz et al.
Ea.C,)
ciC
�0C
>5
�00
.0
C4
‘- ‘- � .0 � E w a. �o a 0. � - -
- C w �
_I I- (�) w
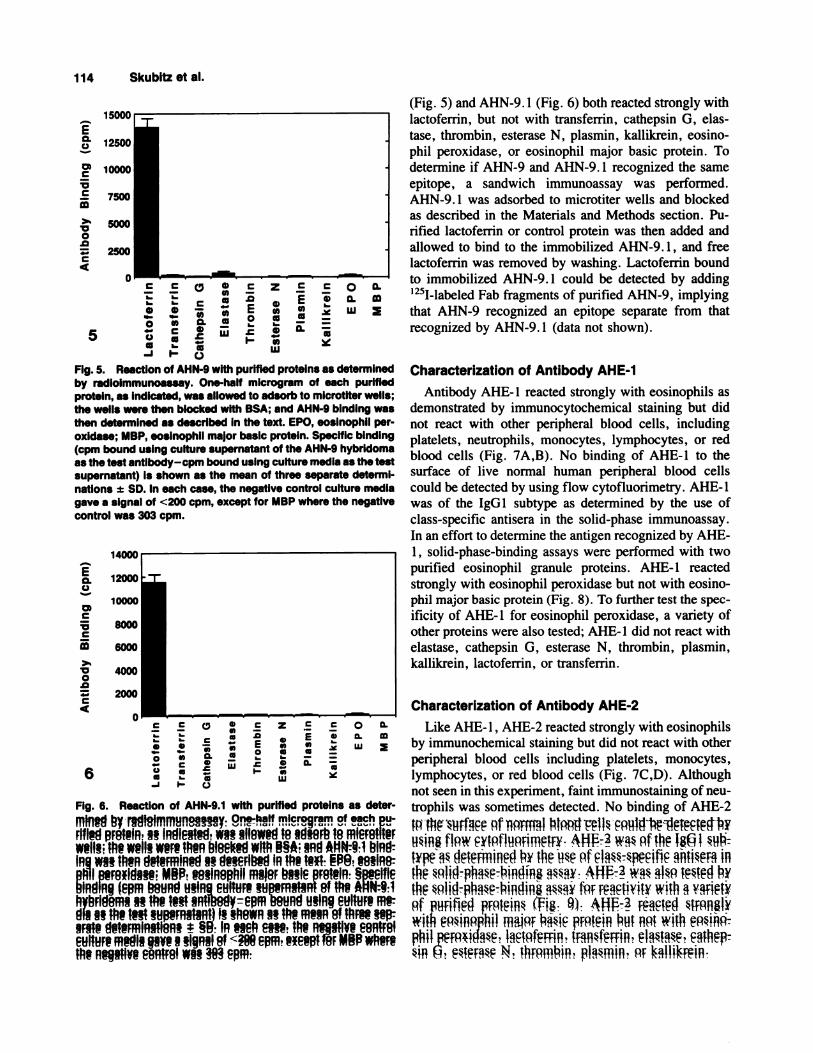
Fig. 5. ReactIon of AHN-9 with purified proteins as determinedby radlolmmunoassay. One-half microgram of each purifiedprotein, as indicated, was allowed to adsorb to microtlter wells;the wells were then blocked with BSA; and AHN-9 binding wasthen determined as described in the text. EPO, eosInophil per-oxidase; MBP, eosinophil major basic protein. Specific binding(cpm bound using culture supernatant of the AHN-9 hybridomaas the test antlbody-cpm bound using culture medIa as the testsupernatant) Is shown as the mean of three separate determi-nations ± SD. In each case, the negative control culture mediagave a signal of <200 cpm, except for MBP where the negativecontrol was 303 cpm.
E0.0
ciC
C
m>5
V0
.0
C4
6
Fig. 6. Reaction of AHN-9.1 with purified proteins as deter-mined by radiolmmunoassay. One-half microgram of each pu-rIfied proteIn, as Indicated, was allowed to adsorb to microtiterwells; the wells were then blocked with BSA; and AHN-9.1 bind-ing was then determined as described in the text. EPO, eosino-phil peroxldase; MBP, eosinophil major basic protein. Specificbinding (cpm bound using culture supernatant of the AHN-9.1hybridoma as the test antibody-cpm bound using culture me-dia as the test supernatant) is shown as the mean of three sep-arate determinations ± SD. In each case, the negative controlculture media gave a signal of <200 cpm, except for MBP wherethe negative control was 303 cpm.
(Fig. 5) and AHN-9. 1 (Fig. 6) both reacted strongly with
lactoferrin, but not with transferrin, cathepsin G, elas-
tase, thrombin, esterase N, plasmin, kallikrein, eosino-phil peroxidase, or eosinophil major basic protein. To
determine if AHN-9 and AHN-9. 1 recognized the same
epitope, a sandwich immunoassay was performed.AHN-9. 1 was adsorbed to microtiter wells and blocked
as described in the Materials and Methods section. Pu-rified lactoferrin or control protein was then added and
allowed to bind to the immobilized AHN-9. 1, and freelactoferrin was removed by washing. Lactoferrin boundto immobilized AHN-9. 1 could be detected by adding
‘251-labeled Fab fragments of purified AHN-9, implying
that AHN-9 recognized an epitope separate from that
recognized by AHN-9. 1 (data not shown).
Characterization of Antibody AHE-1
Antibody AHE- 1 reacted strongly with eosinophils asdemonstrated by immunocytochemical staining but did
not react with other peripheral blood cells, includingplatelets, neutrophils, monocytes, lymphocytes, or red
blood cells (Fig. 7A,B). No binding of AHE-l to thesurface of live normal human peripheral blood cells
could be detected by using flow cytofluorimetry. AHE- 1was of the IgGl subtype as determined by the use of
class-specific antisera in the solid-phase immunoassay.In an effort to determine the antigen recognized by AHE-
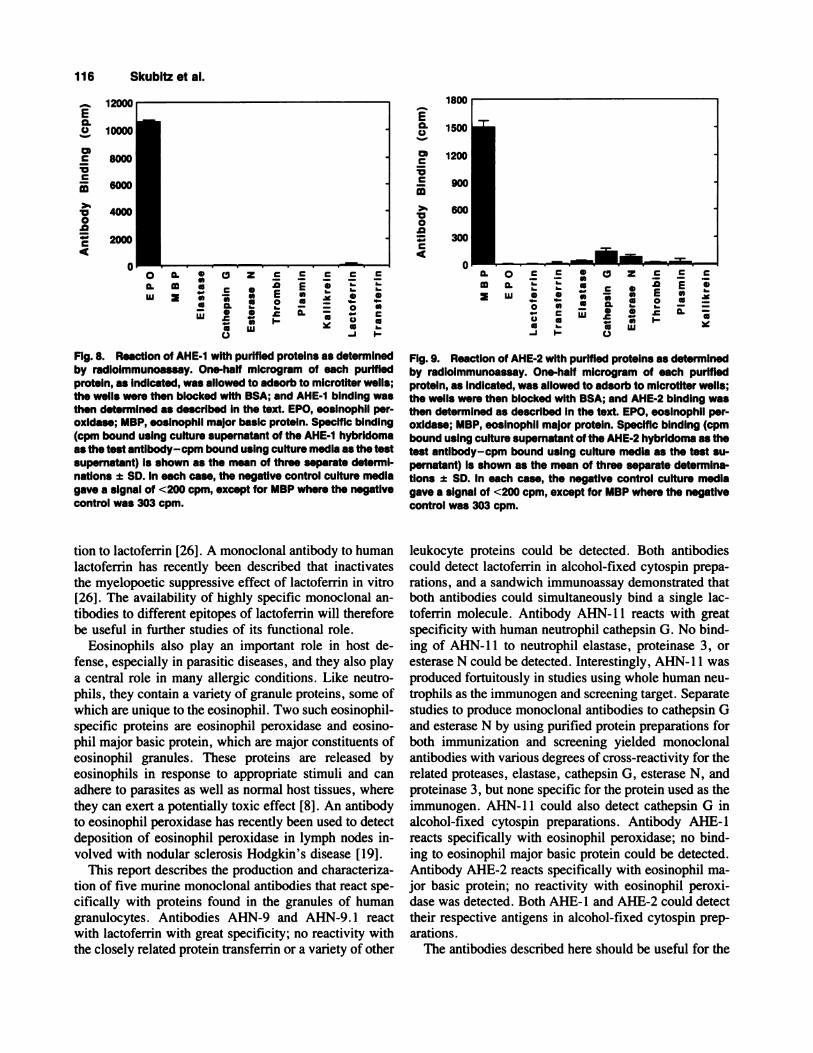
1, solid-phase-binding assays were performed with twopurified eosmnophil granule proteins. AHE- 1 reacted
strongly with eosinophil peroxidase but not with eosino-phil major basic protein (Fig. 8). To further test the spec-
ificity of AHE-l for eosinophil peroxidase, a variety ofother proteins were also tested; AHE- 1 did not react with
elastase, cathepsin 0, esterase N, thrombin, plasmin,
kallikrein, lactoferrin, or transferrin.
Characterization of Antibody AHE-2
Like AHE- 1, AHE-2 reacted strongly with eosmnophilsby immunochemical staining but did not react with otherperipheral blood cells including platelets, monocytes,lymphocytes, or red blood cells (Fig. 7C,D). Althoughnot seen in this experiment, faint immunostaining of neu-
trophils was sometimes detected. No binding of AHE-2
to the surface of normal blood cells could be detected byusing flow cytofluonmetry. AHE-2 was of the IgG 1 sub-
type as determined by the use of class-specific antisera inthe solid-phase-binding assay. AHE-2 was also tested bythe solid-phase-binding assay for reactivity with a variety
of purified proteins (Fig. 9). AHE-2 reacted stronglywith eosmnophil major basic protein but not with eosino-
phil peroxidase, lactoferrin, transferrin, elastase, cathep-sin G, esterase N, thrombin, plasmin, or kallikrein.
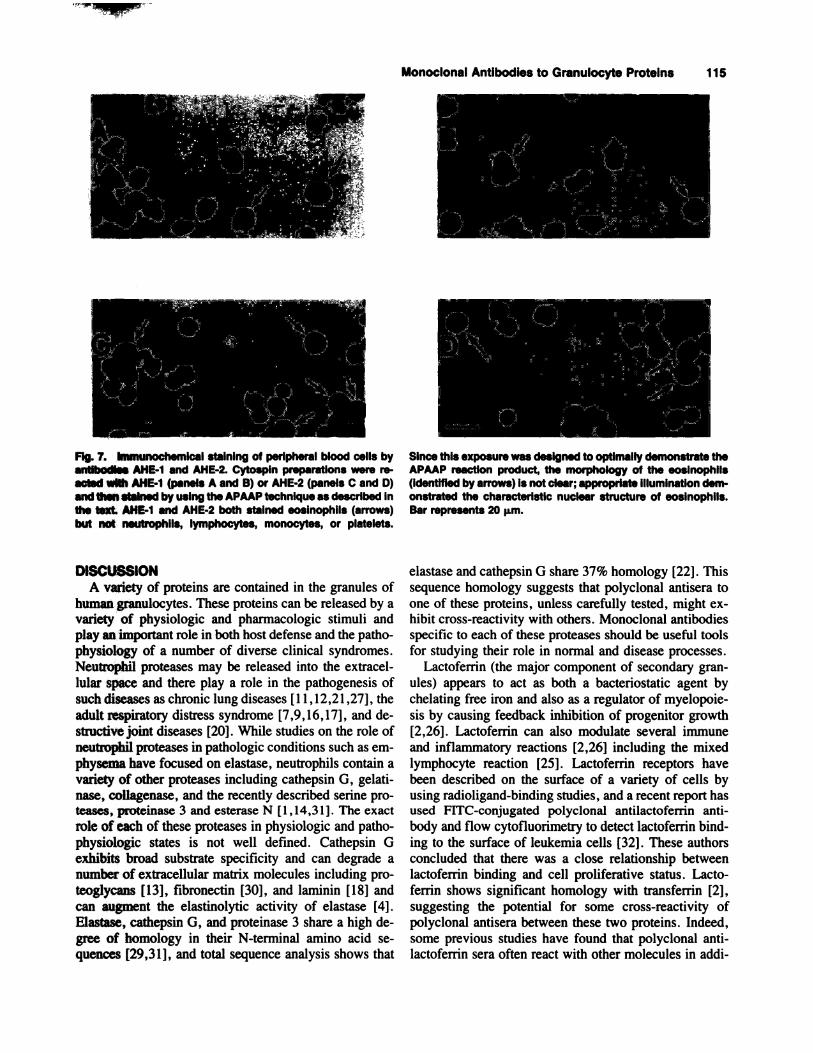
“I
A.4. B-�
. �
Since this exposure was designed to optimally demonstrate theAPAAP reaction product, the morphology of the eosinophlls(identified by arrows) is not clear; appropriate illumination dem-onstrated the characteristic nuclear structure of eosinophils.Bar represents 20 �m.
Monoclonal Antibodies to Granulocyte Proteins 115
4
.�
‘LIFIg. 7. Immunochemical staining of peripheral blood cells byantibodies AHE-1 and AHE-2. Cytospln preparations were re-acted with AHE-1 (panels A and B) or AHE-2 (panels C and D)and then stained by using the APAAP technique as described inthe text AHE-1 and AHE-2 both stained eosinophils (arrows)but not neutrophlls, lymphocytes, monocytes, or platelets.
DISCUSSIONA variety of proteins are contained in the granules of
human granulocytes. These proteins can be released by avariety of physiologic and pharmacologic stimuli andplay an important role in both host defense and the patho-physiology of a number of diverse clinical syndromes.
Neutrophil proteases may be released into the extracel-lular space and there play a role in the pathogenesis of
such diseases as chronic lung diseases [11 ,l2,21 ,27], the
adult respiratory distress syndrome [7,9,16,17], and de-structive joint diseases [20]. While studies on the role ofneutrophil proteases in pathologic conditions such as em-
physema have focused on elastase, neutrophils contain avariety of other proteases including cathepsin G, gelati-nase, collagenase, and the recently described serine pro-teases, proteinase 3 and esterase N [1,14,31]. The exact
role of each of these proteases in physiologic and patho-
physiologic states is not well defined. Cathepsin 0exhibits broad substrate specificity and can degrade anumber of extracellular matrix molecules including pro-
teoglycans [13], fibronectin [30], and laminin [18] andcan augment the elastinolytic activity of elastase [4].
Elastase, cathepsin G, and proteinase 3 share a high de-
gree of homology in their N-terminal amino acid se-quences [29,31], and total sequence analysis shows that
elastase and cathepsin 0 share 37% homology [22]. This
sequence homology suggests that polyclonal antisera toone of these proteins, unless carefully tested, might ex-hibit cross-reactivity with others. Monoclonal antibodiesspecific to each of these proteases should be useful tools
for studying their role in normal and disease processes.Lactoferrin (the major component of secondary gran-
ules) appears to act as both a bacteriostatic agent by
chelating free iron and also as a regulator of myelopoie-sis by causing feedback inhibition of progenitor growth[2,26]. Lactoferrin can also modulate several immune
and inflammatory reactions [2,26] including the mixed
lymphocyte reaction [25]. Lactoferrin receptors havebeen described on the surface of a variety of cells by
using radioligand-binding studies, and a recent report hasused FITC-conjugated polyclonal antilactoferrin anti-
body and flow cytofluorimetry to detect lactoferrin bind-ing to the surface of leukemia cells [32]. These authors
concluded that there was a close relationship betweenlactoferrin binding and cell proliferative status. Lacto-
ferrin shows significant homology with transfernn [2],suggesting the potential for some cross-reactivity ofpolyclonal antisera between these two proteins. Indeed,some previous studies have found that polyclonal anti-lactoferrin sera often react with other molecules in addi-
E0.0
ciC
VC
m>5
V0
.0
C4
E0.C.)
ciC
VC
>5V0
.0
C4
0a.Lu
a aaaaa
Lu
0 Z .EC a
- a Ea a .C
�PS
C C C
E a
- - o aa - - C
� aa -
_i PS
a o� a� UI
�! �0a C
aa�o a � a.
a .- a-J PS C.)
zaaaaaw
C C
.� EE a0
I-
C
a
a
116 Skubitz et al.
Fig. 8. ReactIon of AHE-1 with purified proteins as determinedby radloimmunoassay. One-half microgram of each purifiedprotein, as indicated, was allowed to adsorb to microtiter wells;the wells were then blocked with BSA; and AHE-1 binding wasthen determined as described in the text. EPO, eosinophil per-oxldase; MBP, eosinophil major basic protein. Specific binding(cpm bound using culture supernatant of the AHE-1 hybridomaas the test antibody-cpm bound using culture media as the testsupernatant) is shown as the mean of three separate determI-nations ± SD. In each case, the negative control culture mediagave a signal of <200 cpm, except for MBP where the negativecontrol was 303 cpm.
tion to lactoferrin [261. A monoclonal antibody to human
lactoferrin has recently been described that inactivatesthe myelopoetic suppressive effect of lactoferrin in vitro[26]. The availability of highly specific monoclonal an-
tibodies to different epitopes of lactoferrin will therefore
be useful in further studies of its functional role.
Eosinophils also play an important role in host de-fense, especially in parasitic diseases, and they also play
a central role in many allergic conditions. Like neutro-phils, they contain a variety of granule proteins, some ofwhich are unique to the eosinophil. Two such eosinophil-
specific proteins are eosinophil peroxidase and eosino-phil major basic protein, which are major constituents ofeosinophil granules. These proteins are released by
eosinophils in response to appropriate stimuli and canadhere to parasites as well as normal host tissues, wherethey can exert a potentially toxic effect [8]. An antibodyto eosinophil peroxidase has recently been used to detect
deposition of eosinophil peroxidase in lymph nodes in-
volved with nodular sclerosis Hodgkin’s disease [19].This report describes the production and characteriza-
tion of five murine monoclonal antibodies that react spe-
cifically with proteins found in the granules of humangranulocytes. Antibodies AHN-9 and AHN-9. 1 reactwith lactoferrin with great specificity; no reactivity withthe closely related protein transferrin or a variety of other
Fig. 9. Reaction of AHE-2 with purified proteins as determinedby radioimmunoassay. One-half microgram of each purifiedprotein, as indicated, was allowed to adsorb to microtiter wells;the wells were then blocked with BSA; and AHE-2 binding wasthen determined as described in the text. EPO, eosinophil per-oxidase; MBP, eosinophil major protein. Specific binding (cpmbound using culture supernatant of the AHE-2 hybridoma as thetest antibody-cpm bound using culture media as the test su-pernatant) is shown as the mean of three separate determina-tions ± SD. in each case, the negative control culture mediagave a signal of <200 cpm, except for MBP where the negativecontrol was 303 cpm.
leukocyte proteins could be detected. Both antibodies
could detect lactoferrin in alcohol-fixed cytospin prepa-rations, and a sandwich immunoassay demonstrated thatboth antibodies could simultaneously bind a single lac-toferrin molecule. Antibody AHN-l 1 reacts with great
specificity with human neutrophil cathepsin G. No bind-ing of AHN- 11 to neutrophil elastase, proteinase 3, or
esterase N could be detected. Interestingly, AHN- 11 was
produced fortuitously in studies using whole human neu-trophils as the immunogen and screening target. Separate
studies to produce monoclonal antibodies to cathepsin 0and esterase N by using purified protein preparations for
both immunization and screening yielded monoclonalantibodies with various degrees of cross-reactivity for therelated proteases, elastase, cathepsin 0, esterase N, andproteinase 3, but none specific for the protein used as the
immunogen. AHN-l 1 could also detect cathepsin G in
alcohol-fixed cytospin preparations. Antibody AHE- 1
reacts specifically with eosinophil peroxidase; no bind-
ing to eosinophil major basic protein could be detected.Antibody AHE-2 reacts specifically with eosinophil ma-jor basic protein; no reactivity with eosinophil peroxi-dase was detected. Both AHE-1 and AHE-2 could detect
their respective antigens in alcohol-fixed cytospin prep-
arations.The antibodies described here should be useful for the

Monoclonal Antibodies to Granulocyte Proteins 117
immunolocalization of their respective antigens in tissue
specimens, for quantification of cell surface binding, andfor the identification of cell lineage in the MAC tech-nique of chromosome analysis [151. In addition, theywill serve as a stable source of characterized antibody for
quantitative identification of lactofemn, cathepsin 0,
eosmophil peroxidase, and eosinophil major basic pro-tein, as well as for other studies of the roles of theseproteins in granulocyte function and differentiation.
ACKNOWLEDGMENTS
We thank Dr. D. Hurd for providing eosinophils, Drs.
B. Gray and N. Wehner for providing purified cathepsin
0, esterase N, and proteinase 3, Dr. D. Northfelt forassistance with preliminary experiments, Dr. A. Slun-
gaard for providing purified eosinophil peroxidase andeosinophil major basic protein, Dr. M. Wick for assis-
tance in developing the APAAP technique, Dr. A. Sku-bitz for helpful discussion and a critical review of the
manuscript, and M. Peterson for preparing the manu-
script. This investigation was supported in part by Na-tional Institutes of Health grant RO 1-CA-36248, the Leu-
kemia Task Force, and the Masonic Memorial HospitalFund, Inc.
REFERENCES
1. Baggiolini, M., and Dewald, B. The neutrophil. Int. Arch. Al-lergy App!. Immunol. 76,13, 1985.
2. Birgens, H. The biological significance of lactoferrin in haema-
tology. Scand. J. Haematol. 33,225, 1984.3. Borregaard, N., Heiple, J.M., Simons, E.R., and Clark, R.A.
Subcellular localization of the b-cytochrome component of the
human neutrophil microbicidal oxidase: Translocation during ac-tivation. J. Cell Biol. 97,52, 1983.
4. Boudier, C., Holle, C., and Bieth, J.G. Stimulation of the elas-
tolytic activity of leukocyte elastase by leukocyte cathepsin G. J.Biol. Chem. 256,10256, 1981.
5. Christiansen, N.P., and Skubtiz, K.M. Identification of the majorlectin-binding surface proteins of human neutrophils and alveolar
macrophages. Blood 71,1624, 1982.6. Cordell, J.L., Falini, B., Erber, W.N., Ghosh, AK., Abdulaziz,
Z., MacDonald, S., Pulford, K.A.F., Stein, H., and Mason, D.J.hnmunoenzymatic labeling of monoclonal antibodies using im-mune complexes of alkaline phosphatase and monoclonal anti-
alkaline phosphatase (APAAP complexes). J. Histochem. Cy-tochem. 32,219, 1984.
7. Emmett, M., Fowler, A.A., Hyers, T.M., and Crowle, A.J.
Crossed immunoelectrophoretic analysis of ARDS lavage pro-
teins. Proc. Soc. Exp. Biol. Med. 184,83, 1987.
8. Gleich, G.J., and Adolphson, C.R. The eosinophilic leukocyte:Structure and function. Adv. Immunol. 39,177, 1986.
9. Henson, P.M., and Johnston, R.B., Jr. Tissue injury in inflam-mation. J. Clin. Invest. 79,669, 1987.
10. Hovde, C.J., and Gray, B.H. Characterization of a protein from
normal human polymorphonuclear leukocytes with bactericidalactivity against pseudomonas aeruginosa. Infect. Immun. 54,142,
1986.
11. Janoff, A. Elastase in tissue injury. Annu. Rev. Med. 36,207,
1985.
12. Janoff, A., Sloan, B., Weinbaum, G., Damiano, V., Sandhaus,
R.A., Elias, J., and Kimbel, P. Experimental emphysema in-duced with purified human neutrophil elastase: Tissue localiza-
tion of the instilledprotease. Am. Rev. Respir. Dis. 115,461,
1977.
13. Kaiser, H., Greenwald, R.A., Feinstein, G., and Janoff, A. Deg-radation of cartilage proteoglycan by human leukocyte granule
neutral protease; a model of joint injury. II.Degradation of iso-
lated bovine nasal cartilage proteoglycan. J. Clin. Invest. 57,625,1976.
14. Kao, R.C.-C., Wehner, N.G., Skubitz, K.M., Gray, B.H., andHoidal, J.R. Proteinase 3: A distinct polymorphonuclear leuko-
cyte protease which produced emphysema. J. Clin. Invest. 82,1963, 1988.
15. Knuutila, S., Elonen, E., Teerenhovi, L., Rossi, L., Leskinen,R., Bloomfield, C.D., and de Ia Chapelle, A. Trisomy 12 in B
cells of patients with B-cell chronic lymphotyctic leukemia. N.
EngI. J. Med. 314,865, 1986.
16. Lee, C.T., Fein, AM., Lippman, M., Holtzman, H., Kimbel,
P., and Weinbaum, G. Elastolytic activity in pulmonary lavagefluid from patients with adult respiratory-distress syndrome. N.
EngI. J. Med. 304,192, 1981.
17. McGuire, W.W., Spragg, R.G., Cohen, A.B., and Cochrane,C.G. Studies on the pathogenesis of the adult respiratory distress
syndrome. J. Clin. Invest. 69,543, 1982.
18. Rao, C.N., Margulies, J.M.K., Goldfarb, F.H., Madri, J.A.,Woodley, D.T., and Liotta, L.A. Differential proteolytic suscep-
tibility of laminin alpha and beta subunits. Arch. Biochem. Bio-
phys. 219,65, 1982.19. Samoszuk, M., Sholly, S., and Epstein, A.L. Eosinophil perox-
idase is detectable with a monoclonal antibody in collagen bands
of nodular sclerosis Hodgkin’s disease. Lab. Invest. 56,394,
1987.
20. Schneider, M., Lohmann, J., Krummenerl, T., and Gerlach, U.
Concentration of fibronectin and granulocyte elastase in plasma of
patients with systemic connective-tissue diseases. Int. J. TissueReact. 9,335, 1987.
21. Senior, R.M., Tegner, H., Kuhn, C., Ohlsson, K., StarcherB.C., and Pierce, J.A. The induction of pulmonary emphysemawith human leukocyte elastase. Am. Rev. Respir. Dis. 116,469,
1977.
22. Sinha, S., Watorek, W., Karr, S., Giles, J., Bode, W., andTravis, J. Primary structure of human neutrophil elastase. Proc.Natl. Acad. Sci. USA 84,2228, 1987.
23. Skubitz, K.M., Wehner, N.G., and Gray, B.H. Preparation andcharacterization of a monoclonal antibody that inhibits human
neutrophil elastase activity. J. Leukocyte Biol. 44,158, 1988.
24. Skubitz, K.M., Zhen, Y., and August, J.T. A human granulo-
cyte-specific antigen characterized by use of monoclonal antibod-ies. Blood 61,19, 1983.
25. Slater, K., and Fletcher, J. Lactoferrin derived from neutrophilsinhibits the mixed lymphocyte reaction. Blood 69,1328, 1987.
26. Sledge, OW., Jr., Bicknell, D.C., Hams, E.L., Zegarra, G.,and Broxmeyer, H.E. Monoclonal antibody (112C) to human lac-
toferrin inactivates the myelopoietic suppressive effect of human
lactoferrin in vitro. Exp. Hematol. 14,333, 1986.
27. Snider, G.L., Lucey, E.C., Christensen, T.G., Stone, P.J., Ca-lore, J.D., Cantanese, A., and Franzblau, C. Emphysema and
bronchial secretory cell metaplasia induced in hamsters by human
neutrophil products. Am. Rev. Respir. Dis. 129,155, 1984.
28. Tanaka, T., Minematsu, Y., Reilly, C.F., Travis, J., and Powers,J.C. Human leukocyte cathepsin G: Subsite mapping with 4-ni-

118 Skubitz et al.
troanilides,chemical modification, and effect of possible cofac-tors. Biochemistry 24,2040, 1985.
29. Travis, J.,Giles, P.J.,Porcelli,L., Reilly, C.F., Baugh, R., and
Powers, J. Human leucocyte elastase and cathepsin 0: Structural
and functional characteristics. Excerpta Med. 75,51, 1980.
30. Vartio, T., Seppa, H., and Vaheri, A. Susceptibility of soluble
and matrix fibronectins to degradation by tissue proteinase, mastcell chymase and cathepsin G.J. Biol. Chem. 256,471, 1981.
31. Wehner, N. Biochemistry and Function of Proteases Isolated
From the Cytoplasmic Granules of Polymorphonuclear Leuko-
cytes. Ph.D. Thesis, University of Minnesota, Minneapolis,1987.
32. Yamada, Y., Amagasaki, T., Jacobsen, D.W., and Green, R.
Lactoferrin binding by leukemia cell lines. Blood 70,264, 1987.




















