
Eosinophil activation status, cytokines and liver fibrosis in Schistosoma mansoni infected patients
10
Acta Tropica 108 (2008) 150–159 Contents lists available at ScienceDirect Acta Tropica journal homepage: www.elsevier.com/locate/actatropica Eosinophil activation status, cytokines and liver fibrosis in Schistosoma mansoni infected patients Denise Silveira-Lemos a,b,c,∗ , Andr ´ ea Teixeira-Carvalho a,c , Olindo Assis Martins-Filho c , L´ ucia Fraga Alves Oliveira d , Matheus Fernandes Costa-Silva a,c , Leonardo Ferreira Matoso e , Lorena J ´ unia de Souza a , Andr ´ ea Gazzinelli e , Rodrigo Corrˆ ea-Oliveira a a Laborat´ orio de Imunologia Celular e Molecular, Centro de Pesquisas Ren´ e Rachou, FIOCRUZ, Belo Horizonte, MG, Brazil b Departamento de Bioqu´ ımica e Imunologia, Instituto de Ciˆ encias Biol´ ogicas, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil c Laborat´ orio de Biomarcadores de Diagn´ ostico e Monitorac ¸˜ ao, Centro de Pesquisas Ren´ e Rachou, FIOCRUZ, Belo Horizonte, MG, Brazil d N´ ucleo de Pesquisa em Imunologia, Faculdade de Ciˆ encias da Sa´ ude, Universidade Vale do Rio Doce, Governador Valadares, MG, Brazil e Departamento de Enfermagem, Materno-Infantil, Escola de Enfermagem, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil article info Article history: Available online 15 April 2008 Keywords: Flow cytometry Eosinophils Activation status Cytokines schistosomiasis Periportal fibrosis abstract We have been investigating whether human eosinophils play an important role in schistosomiasis mansoni morbidity. Our main focus has been on the activation-related cell surface markers (CD23/CD69/CD25/HLA- DR/CD28/CD80) and the detection of TNF-, IL-4 and IL-5 in peripheral blood eosinophils from chronic Schistosoma mansoni-infected patients. Our studies compare both, intestinal (INT) and individuals with periportal fibrosis (FIB). Our major findings, point to distinct profile of activation-related surface markers on eosinophils as the hallmark of disease morbidity during chronic S. mansoni infection. Up-regulation of several activation-related markers was observed on eosinophils from FIB group, but not INT, which include early activation markers, such as CD69 and CD23. INT displayed a distinct profile, with up-regulation of molecules related to the late activation (CD25, HLA-DR, CD28 and CD80). These results suggest that some immunoregulatory events may take place controlling the early eosinophil activation in the INT group. Higher levels of eosinophil-derived cytokines were observed in FIB, regardless the antigen stimulation in vitro. A mixed cytokine pattern, characterized by positive correlation between TNF-, IL-4 and IL-5 was observed in both INT and FIB. However, lack of correlation between the cytokine expression and the eosinophil activation status points out that even those FIB patients presenting minor increment on eosinophil activation displayed higher levels of cytokine-positive eosinophils. Indeed, the positive asso- ciation between lymphocyte-derived IL-10 and the eosinophils cytokine profile was observed exclusively in INT further emphasize our hypothesis that immunoregulatory events take place controlling disease morbidity in human schistosomiasis. The impaired IL-10-driven immunoregulatory function may play an important role on the establishment of pathology in patients bearing periportal fibrosis. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Eosinophils have been postulated to play an important protec- tive role in the immune response to Schistosoma mansoni infection (Colley, 1972; Moore et al., 1977; Butterworth et al., 1979). Such protective hypothesis was primarily based on histopathological evi- dences of the presence high number of eosinophils surrounding ∗ Corresponding author at: Laborat ´ orio de Imunologia Celular e Molecular, Centro de Pesquisas Ren ´ e Rachou, FIOCRUZ, Avenida Augusto de Lima, 1715, Barro Preto, Belo Horizonte, Minas Gerais, 30 190 002, Brazil. Tel.: +55 31 3349 7778/7777; fax: +55 31 3295 3115. E-mail address: dlemos@cpqrr.fiocruz.br (D. Silveira-Lemos). dying helminths in tissue biopsy specimens (Butterworth, 1984; Cox, 1998) and also by the in vitro eosinophils mediated killing of schistosomula in the presence of antibodies and/or complement as well as on eosinophil granule products activity (Sher et al., 1977; Ramalho-Pinto et al., 1978; Anwar et al., 1979; Butterworth et al., 1979; David et al., 1980). Additionally, several evidences have corre- lated the number of eosinophils with protective immunity during S. mansoni infection (Vadas et al., 1980; David et al., 1980; Veith and Butterworth, 1983). However, it has been proposed that more than an increase on the number of eosinophils, there is also an increase on the activity of these cells that may account for the role of eosinophils during S. mansoni, and consequently the protective importance of eosinophilia. These activities may involve not only a quantitative increase on the eosinophil numbers but also a qual- 0001-706X/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.actatropica.2008.04.006
Transcript of Eosinophil activation status, cytokines and liver fibrosis in Schistosoma mansoni infected patients

ES
DLLa
b
c
d
e
a
AA
KFEACP
1
t(pd
dBf
0d
Acta Tropica 108 (2008) 150–159
Contents lists available at ScienceDirect
Acta Tropica
journa l homepage: www.e lsev ier .com/ locate /ac ta t ropica
osinophil activation status, cytokines and liver fibrosis inchistosoma mansoni infected patients
enise Silveira-Lemosa,b,c,∗, Andrea Teixeira-Carvalhoa,c, Olindo Assis Martins-Filhoc,ucia Fraga Alves Oliveirad, Matheus Fernandes Costa-Silvaa,c, Leonardo Ferreira Matosoe,orena Junia de Souzaa, Andrea Gazzinelli e, Rodrigo Correa-Oliveiraa
Laboratorio de Imunologia Celular e Molecular, Centro de Pesquisas Rene Rachou, FIOCRUZ, Belo Horizonte, MG, BrazilDepartamento de Bioquımica e Imunologia, Instituto de Ciencias Biologicas, Universidade Federal de Minas Gerais, Belo Horizonte, MG, BrazilLaboratorio de Biomarcadores de Diagnostico e Monitoracao, Centro de Pesquisas Rene Rachou, FIOCRUZ, Belo Horizonte, MG, BrazilNucleo de Pesquisa em Imunologia, Faculdade de Ciencias da Saude, Universidade Vale do Rio Doce, Governador Valadares, MG, BrazilDepartamento de Enfermagem, Materno-Infantil, Escola de Enfermagem, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil
r t i c l e i n f o
rticle history:vailable online 15 April 2008
eywords:low cytometryosinophilsctivation statusytokines schistosomiasiseriportal fibrosis
a b s t r a c t
We have been investigating whether human eosinophils play an important role in schistosomiasis mansonimorbidity. Our main focus has been on the activation-related cell surface markers (CD23/CD69/CD25/HLA-DR/CD28/CD80) and the detection of TNF-�, IL-4 and IL-5 in peripheral blood eosinophils from chronicSchistosoma mansoni-infected patients. Our studies compare both, intestinal (INT) and individuals withperiportal fibrosis (FIB). Our major findings, point to distinct profile of activation-related surface markerson eosinophils as the hallmark of disease morbidity during chronic S. mansoni infection. Up-regulation ofseveral activation-related markers was observed on eosinophils from FIB group, but not INT, which includeearly activation markers, such as CD69 and CD23. INT displayed a distinct profile, with up-regulation ofmolecules related to the late activation (CD25, HLA-DR, CD28 and CD80). These results suggest that someimmunoregulatory events may take place controlling the early eosinophil activation in the INT group.Higher levels of eosinophil-derived cytokines were observed in FIB, regardless the antigen stimulationin vitro. A mixed cytokine pattern, characterized by positive correlation between TNF-�, IL-4 and IL-5
was observed in both INT and FIB. However, lack of correlation between the cytokine expression andthe eosinophil activation status points out that even those FIB patients presenting minor increment oneosinophil activation displayed higher levels of cytokine-positive eosinophils. Indeed, the positive asso-ciation between lymphocyte-derived IL-10 and the eosinophils cytokine profile was observed exclusivelyin INT further emphasize our hypothesis that immunoregulatory events take place controlling diseasemorbidity in human schistosomiasis. The impaired IL-10-driven immunoregulatory function may play anblish
dCs
important role on the esta
. Introduction
Eosinophils have been postulated to play an important protec-
ive role in the immune response to Schistosoma mansoni infectionColley, 1972; Moore et al., 1977; Butterworth et al., 1979). Suchrotective hypothesis was primarily based on histopathological evi-ences of the presence high number of eosinophils surrounding∗ Corresponding author at: Laboratorio de Imunologia Celular e Molecular, Centroe Pesquisas Rene Rachou, FIOCRUZ, Avenida Augusto de Lima, 1715, Barro Preto,elo Horizonte, Minas Gerais, 30 190 002, Brazil. Tel.: +55 31 3349 7778/7777;
ax: +55 31 3295 3115.E-mail address: [email protected] (D. Silveira-Lemos).
wR1lSatioia
001-706X/$ – see front matter © 2008 Elsevier B.V. All rights reserved.oi:10.1016/j.actatropica.2008.04.006
ment of pathology in patients bearing periportal fibrosis.© 2008 Elsevier B.V. All rights reserved.
ying helminths in tissue biopsy specimens (Butterworth, 1984;ox, 1998) and also by the in vitro eosinophils mediated killing ofchistosomula in the presence of antibodies and/or complement asell as on eosinophil granule products activity (Sher et al., 1977;amalho-Pinto et al., 1978; Anwar et al., 1979; Butterworth et al.,979; David et al., 1980). Additionally, several evidences have corre-ated the number of eosinophils with protective immunity during. mansoni infection (Vadas et al., 1980; David et al., 1980; Veithnd Butterworth, 1983). However, it has been proposed that more
han an increase on the number of eosinophils, there is also anncrease on the activity of these cells that may account for the rolef eosinophils during S. mansoni, and consequently the protectivemportance of eosinophilia. These activities may involve not onlyquantitative increase on the eosinophil numbers but also a qual-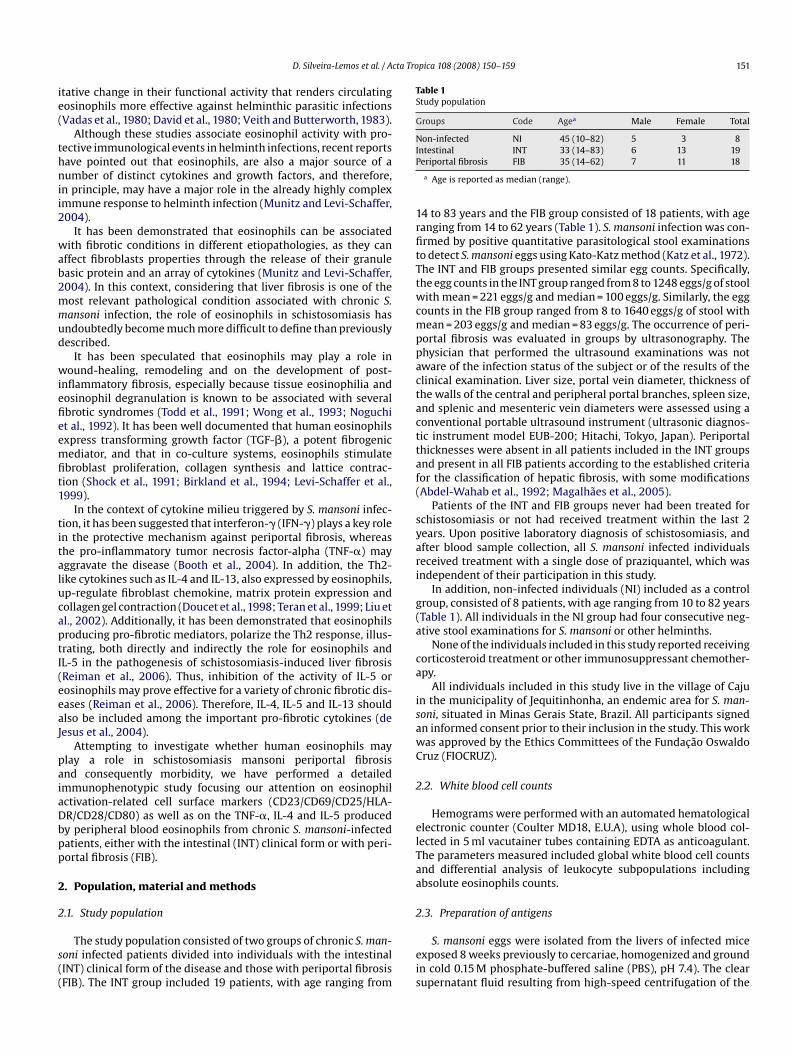
cta Tropica 108 (2008) 150–159 151
ie(
thnii2
wab2mmud
wiefieemfit1
titalucaptI(eeaJ
paiaDbpp
2
2
s((
Table 1Study population
Groups Code Agea Male Female Total
Non-infected NI 45 (10–82) 5 3 8IP
1rfitTtwcmppactacttaf(
syari
g(a
ca
isawC
2
elTaa
2
D. Silveira-Lemos et al. / A
tative change in their functional activity that renders circulatingosinophils more effective against helminthic parasitic infectionsVadas et al., 1980; David et al., 1980; Veith and Butterworth, 1983).
Although these studies associate eosinophil activity with pro-ective immunological events in helminth infections, recent reportsave pointed out that eosinophils, are also a major source of aumber of distinct cytokines and growth factors, and therefore,
n principle, may have a major role in the already highly complexmmune response to helminth infection (Munitz and Levi-Schaffer,004).
It has been demonstrated that eosinophils can be associatedith fibrotic conditions in different etiopathologies, as they can
ffect fibroblasts properties through the release of their granuleasic protein and an array of cytokines (Munitz and Levi-Schaffer,004). In this context, considering that liver fibrosis is one of theost relevant pathological condition associated with chronic S.ansoni infection, the role of eosinophils in schistosomiasis hasndoubtedly become much more difficult to define than previouslyescribed.
It has been speculated that eosinophils may play a role inound-healing, remodeling and on the development of post-
nflammatory fibrosis, especially because tissue eosinophilia andosinophil degranulation is known to be associated with severalbrotic syndromes (Todd et al., 1991; Wong et al., 1993; Noguchit al., 1992). It has been well documented that human eosinophilsxpress transforming growth factor (TGF-�), a potent fibrogenicediator, and that in co-culture systems, eosinophils stimulate
broblast proliferation, collagen synthesis and lattice contrac-ion (Shock et al., 1991; Birkland et al., 1994; Levi-Schaffer et al.,999).
In the context of cytokine milieu triggered by S. mansoni infec-ion, it has been suggested that interferon-� (IFN-�) plays a key rolen the protective mechanism against periportal fibrosis, whereashe pro-inflammatory tumor necrosis factor-alpha (TNF-�) mayggravate the disease (Booth et al., 2004). In addition, the Th2-ike cytokines such as IL-4 and IL-13, also expressed by eosinophils,p-regulate fibroblast chemokine, matrix protein expression andollagen gel contraction (Doucet et al., 1998; Teran et al., 1999; Liu etl., 2002). Additionally, it has been demonstrated that eosinophilsroducing pro-fibrotic mediators, polarize the Th2 response, illus-rating, both directly and indirectly the role for eosinophils andL-5 in the pathogenesis of schistosomiasis-induced liver fibrosisReiman et al., 2006). Thus, inhibition of the activity of IL-5 orosinophils may prove effective for a variety of chronic fibrotic dis-ases (Reiman et al., 2006). Therefore, IL-4, IL-5 and IL-13 shouldlso be included among the important pro-fibrotic cytokines (deesus et al., 2004).
Attempting to investigate whether human eosinophils maylay a role in schistosomiasis mansoni periportal fibrosisnd consequently morbidity, we have performed a detailedmmunophenotypic study focusing our attention on eosinophilctivation-related cell surface markers (CD23/CD69/CD25/HLA-R/CD28/CD80) as well as on the TNF-�, IL-4 and IL-5 producedy peripheral blood eosinophils from chronic S. mansoni-infectedatients, either with the intestinal (INT) clinical form or with peri-ortal fibrosis (FIB).
. Population, material and methods
.1. Study population
The study population consisted of two groups of chronic S. man-oni infected patients divided into individuals with the intestinalINT) clinical form of the disease and those with periportal fibrosisFIB). The INT group included 19 patients, with age ranging from
eis
ntestinal INT 33 (14–83) 6 13 19eriportal fibrosis FIB 35 (14–62) 7 11 18
a Age is reported as median (range).
4 to 83 years and the FIB group consisted of 18 patients, with ageanging from 14 to 62 years (Table 1). S. mansoni infection was con-rmed by positive quantitative parasitological stool examinationso detect S. mansoni eggs using Kato-Katz method (Katz et al., 1972).he INT and FIB groups presented similar egg counts. Specifically,he egg counts in the INT group ranged from 8 to 1248 eggs/g of stoolith mean = 221 eggs/g and median = 100 eggs/g. Similarly, the egg
ounts in the FIB group ranged from 8 to 1640 eggs/g of stool withean = 203 eggs/g and median = 83 eggs/g. The occurrence of peri-
ortal fibrosis was evaluated in groups by ultrasonography. Thehysician that performed the ultrasound examinations was notware of the infection status of the subject or of the results of thelinical examination. Liver size, portal vein diameter, thickness ofhe walls of the central and peripheral portal branches, spleen size,nd splenic and mesenteric vein diameters were assessed using aonventional portable ultrasound instrument (ultrasonic diagnos-ic instrument model EUB-200; Hitachi, Tokyo, Japan). Periportalhicknesses were absent in all patients included in the INT groupsnd present in all FIB patients according to the established criteriaor the classification of hepatic fibrosis, with some modificationsAbdel-Wahab et al., 1992; Magalhaes et al., 2005).
Patients of the INT and FIB groups never had been treated forchistosomiasis or not had received treatment within the last 2ears. Upon positive laboratory diagnosis of schistosomiasis, andfter blood sample collection, all S. mansoni infected individualseceived treatment with a single dose of praziquantel, which wasndependent of their participation in this study.
In addition, non-infected individuals (NI) included as a controlroup, consisted of 8 patients, with age ranging from 10 to 82 yearsTable 1). All individuals in the NI group had four consecutive neg-tive stool examinations for S. mansoni or other helminths.
None of the individuals included in this study reported receivingorticosteroid treatment or other immunosuppressant chemother-py.
All individuals included in this study live in the village of Cajun the municipality of Jequitinhonha, an endemic area for S. man-oni, situated in Minas Gerais State, Brazil. All participants signedn informed consent prior to their inclusion in the study. This workas approved by the Ethics Committees of the Fundacao Oswaldoruz (FIOCRUZ).
.2. White blood cell counts
Hemograms were performed with an automated hematologicallectronic counter (Coulter MD18, E.U.A), using whole blood col-ected in 5 ml vacutainer tubes containing EDTA as anticoagulant.he parameters measured included global white blood cell countsnd differential analysis of leukocyte subpopulations includingbsolute eosinophils counts.
.3. Preparation of antigens
S. mansoni eggs were isolated from the livers of infected micexposed 8 weeks previously to cercariae, homogenized and groundn cold 0.15 M phosphate-buffered saline (PBS), pH 7.4). The clearupernatant fluid resulting from high-speed centrifugation of the

1 cta Tro
hgAiwts
2
i((i
((i(r
B
2
pmibStotFlMuVs
imbvssoo(tme
2
ii1sid
iciBc3dpdv
ioiBca
osPS(tsa
awMa
filw
2
bi0briDtfgsAacaomito
52 D. Silveira-Lemos et al. / A
omogenate at 18,000 × g for 1 h was named soluble egg anti-ens (SEA) and stored at −70 ◦C until use (Gazzinelli et al., 1983).dult S. mansoni worms (males and females) were suspended in
ce-cold PBS and sonicated in ice (2× 10 min). Insoluble materialas removed by centrifugation at 18,000 × g for 1 h at 4 ◦C, and
he supernatant, named soluble worm antigen preparation (SWAP)tored at −70 ◦C until use (Gazzinelli et al., 1983).
.4. Monoclonal antibodies (mAbs)
Monoclonal antibodies (mAbs) anti-human cell surface markers,ncluding anti-CD69 (BAF2359), anti-CD25 (3G10), anti-HLA-DRTU36), anti-CD23 (M-L233), anti-CD28 (FAB342P), anti-CD80BAF140) labeled with phycoerithrin (PE) were used for ex vivo cellmmunophenotypic staining.
Anti-CD16 mAbs (B73.1) labeled with fluorescein isothiocyanateFITC), anti-TNF-� (Mab11), anti-IL-4 (MP4-25D2) and anti-IL-5TRFK5) mAbs labeled with PE were used for immunostaining forntracellular cytokines. Isotypic controls, including mouse IgG1679.1Mc7) and IgG2a (UCTH-1) were labeled with FITC and PE,espectively.
All mAbs and controls were purchased from Becton Dickinsoniosciences Pharmingen (San Diego, CA, USA).
.5. Ex vivo cell immunophenotypic staining
Fifty microliters of EDTA whole blood were incubated in theresence of 5 �l of undiluted anti-human cell surface moleculesAbs for 30 min, in the dark and at room temperature. Following
ncubation, erythrocytes lysis was performed using 2 ml of FACSrand Lysing Solution (Becton Dickinson Biosciences Pharmingen,an Diego, CA, USA), followed by incubation for 10 min, at roomemperature. Leukocyte suspension was further washed with 2 mlf phosphate-buffered saline containing 0.01% sodium azide. Prioro flow cytometric acquisition, stained cells were fixed in 200 �l ofACS fix solution (10 g/l paraformaldehyde, 10.2 g/l sodium cacody-ate, 6.65 g/l sodium chloride, Sigma Chemical Company, St. Louis,
O, USA) for at least 15 min at 4 ◦C. Data collection was performedsing a FACScalibur flow cytometer (Becton Dickinson, Mountainiew, CA, USA) and acquisition and analysis using CELLQUESTTM
oftware provided by the manufacturer.Analysis of eosinophils was performed by single color
mmunophenotyping with FL-2/PE-labeled anti-cell surfacearker mAbs. Gating strategy was based on eosinophil selection
y their autofluorescent properties on non-related FL-3 channelersus forward scatter (FSC) graphs. A confirmatory graph was alsoet up to identify the eosinophils on forward scatter (FSC) versuside scatter (SSC) dot plot distribution confined into a regionf high cell complexity. Data are presented as absolute numberf eosinophils population expressing a given surface moleculescells/mm3 of peripheral blood). The values were calculatedaking the percentage of eosinophils that express a given surface
olecules of interest multiplied by the absolute number of theosinophils obtained from the hematological analysis.
.6. Short-term whole blood in vitro culture
Five hundred microliters of peripheral blood samples collectednto Vacutainer tubes containing heparin sodium salt (Becton Dick-
nson, Mountain View, CA, USA) were dispensed into individual7 mm × 100 mm polypropylene tubes (Falcon 2059, Becton Dickin-on, Mountain View, CA, USA). Short-term cultures were performedn the absence (control cultures) or in the presence of S. mansoni-erived antigens (SEA and SWAP).twtPp
pica 108 (2008) 150–159
Analysis of cytokine profile was evaluated after short-termncubation in vitro, in the absence of exogenous stimuli (controlultures). For this purpose, 500 �l of whole blood samples werencubated in the presence of 500 �l of RPMI 1640 plus Brefeldin A,FA (Sigma Chemical Company, St. Louis, MO, USA) at a final con-entration of 10 �g/ml. Blood samples were incubated for 4 h at7 ◦C in humidified incubator in a 5% CO2 atmosphere. These con-itions were chosen considering that the detection of cytokines,articularly in the absence of exogenous stimuli, may reflect theynamic events of cytokine production by blood eosinophils inivo.
Antigen-specific stimulation was performed by previous in vitroncubation of 500 �l whole blood aliquots in the presence of (SEA)r (SWAP) at final concentration of 25 �g/ml for 1 h at 37 ◦C humid-fied incubator in a 5% CO2 atmosphere. Following the addition ofFA (Sigma Chemical Company, St. Louis, MO, USA) at a final con-entration of 10 �g/ml, blood samples were again incubated for 4 ht 37 ◦C in humidified incubator in a 5% CO2 atmosphere.
Positive control cultures were also maintained for the evaluationf the cell sample viability. For this purpose, 500 �l of whole bloodamples were incubated in the presence of 500 �l of RPMI 1640 plushorbol 12-Myristate 13-Acetate, PMA (Sigma Chemical Company,t. Louis, MO, USA) at a final concentration of 25 ng/ml, ionomycinSigma, Chemical Company, St. Louis, MO, USA) at a final concentra-ion of 1 �g/ml and BFA at a final concentration of 10 �g/ml. Bloodamples were incubated for 4 h at 37 ◦C in humidified incubator in5% CO2 atmosphere.
Prior to the immunostaining for intracellular cytokine analysis,ll cultures, including control, antigen and PMA stimulated culturesere treated with 2 mM EDTA (Sigma Chemical Company, St. Louis,O, USA) and kept at room temperature for 15 min, to stop cell
ctivation process.Cytokines patterns observed in the positive control cultures con-
rmed the cell viability of all samples, as demonstrated by highevels TNF-�+ cells in PMA-stimulated cultures when compared
ith the negative control cultures (data not shown).
.7. Immunostaining for intracellular cytokine
Following the short term in vitro stimulation, cultured wholelood samples were washed with 6 ml of FACS buffer contain-
ng PBS supplemented with 0.5% Bovine Serum Albumin—BSA and.1% sodium azide (Sigma Chemical Company, St. Louis, MO, USA),y centrifugation at 600 × g at room temperature for 7 min. Afteresuspension in 1 ml of FACS buffer, 400 �l aliquots were dispensednto one 12 mm × 75 mm polystyrene tube (Falcon no. 2052, Bectonickinson, Mountain View, CA, USA) and stained with the manufac-
ures recommended amount of mAbs anti-CD16-FITC in the dark,or 30 min at room temperature. Stained samples were treated byentle vortexing with 2 ml of FACS Lysing Solution (Becton Dickin-on, San Jose, CA, USA) and re-incubated for an additional 10 min.fter erythrocyte lyses, the samples were centrifuged at 600 × gt room temperature for 7 min, the supernatant discarded and theell pellet resuspended and incubated with 2 ml of FACS perme-bilizing solution, containing FACS buffer supplemented with 0.5%f saponin (Sigma Chemical Company, St. Louis, MO, USA) andaintained in the dark for 10 min at room temperature. Follow-
ng incubation, the samples were centrifuged at 600 × g at roomemperature for 7 min, the supernatant gently decanted and 3 mlf FACS buffer added to the resuspended pellet. After centrifugation,
he pellet was resuspended and 30 �l aliquots of the cell suspensionere distributed in 96 wells U bottom microplate (Falcon, Bec-on Dickinson, Mountain View, CA, USA) and stained with 20 �l ofE-labeled anti-cytokine mAb (anti-TNF-�, anti-IL-4 and anti-IL-5)reviously diluted 1:50 in sterile FACS permeabilizing solution and

cta Tro
ttpwTc
ifssivrioeeaa
2
sPv
dtu(p
3
3im
patFiilwa
Fiws(lsb
D. Silveira-Lemos et al. / A
he cells were incubated in the dark for 30 min at room tempera-ure. After incubation, the cells were washed with 150 �l of FACSermeabilizing solution followed by 200 �l of FACS buffer. Afterashing, stained cells were fixed in 200 �l of FACS fix solution.
he samples were stored at 4 ◦C in the dark and analyzed by flowytometry within 24 h.
Cytokine-expressing eosinophils were identified by dual colormmunophenotyping with FL1/FITC-labeled anti-human cell sur-ace marker CD16 and FL-2/PE-labeled anti-cytokine mAbs. Gatingtrategy was based on eosinophil selection by their high sidecatter properties besides being negative for CD16 immunostain-ng. Eosinophils expressing cytokines were quantified using FSCersus FL2/anti-cytokine-PE dot plots, by setting quadrants to seg-egate FL2 positive and negative cells based on the negative controlmmunostaining. Results were expressed as the absolute numberf eosinophils expressing a given cytokine (cells/mm3 of periph-ral blood). The values were calculated taking the percentage ofosinophils that express the cytokine of interest multiplied by thebsolute number of the eosinophils obtained from hematologicalnalysis.
.8. Statistical analysis
Differences between groups were first evaluated by MINITAB®
tatistical software release 13.20 for Windows (State College,A, USA) to test three hypotheses: independence, normality andariance of data sets. Considering its non-parametric nature, all
Ctoat
ig. 1. Absolute counts (top panel) and activation marker expression (middle and bottom pncluding intestinal (INT: = 19) and those with periportal fibrosis (FIB: = 18) asas essentially based on their autofluorescent cells using non-related FL-3 channel versu
ingle color immunophenotyping. The results are expressed in box-plot format highlighttop panel), absolute numbers of CD69+, CD25+ and HLA-DR+ (middle panels) as well asower hinge (defined as the 25th percentile) to the upper hinge (the 75th percentile) andhown as a line across the box. Therefore 1/4 of the distribution is between this line andy connecting lines.
pica 108 (2008) 150–159 153
ata sets were analyzed by Kruskal–Wallis followed by Dunnsest with further analysis by Spearman’s rank correlation testsing the Graphpad PRISM® software release 4.02 for WindowsSan Diego, CA, USA). Significance was defined in both cases at< 0.05.
. Results
.1. Distinct up-regulation of activation-related surface markersn eosinophils is a hallmark of disease morbidity during chronic S.
ansoni infection
Ex vivo analysis of peripheral blood eosinophil counts (topanel) and their activation status (middle and bottom panels)re shown in Fig. 1. Although eosinophil counts increase selec-ively in the INT group, all S. mansoni infected patients (INT andIB) displayed higher number of circulating eosinophils express-ng activation-related surface markers as compared to NI. Its interesting to note that, despite the basal levels of circu-ating eosinophil observed in patients with periportal fibrosis,
e observed an up-regulation of all activation-related markersnalyzed, even the early activation markers, such as CD69 and
D23, suggesting active involvement of these cells in the schis-osomal fibrosis process (Fig. 1). On the other hand, analysisf eosinophils from INT demonstrated a distinct expression onctivation-related marker, with up-regulation of molecules relatedo the late activation status (CD25, HLA-DR, CD28 and CD80), sug-anels) of peripheral blood eosinophils from Schistosoma mansoni infected patients,compared to non-infected individuals (NI: = 8). The selection of eosinophilss forward scatter (FSC) and the analysis of activated eosinophils was carry out by
ing the gap of 50% of data set measurement for the absolute counts of eosinophilsCD23+, CD28+ and CD80+ eosinophils (bottom panels). The box stretches from the
therefore contains the middle half of the score in the distribution. The median isthe bottom or the top of the box. Significant differences at p < 0.05 are highlighted

154 D. Silveira-Lemos et al. / Acta Tropica 108 (2008) 150–159
Fig. 2. Intracellular levels of TNF-� (left panels), IL-4 (center panels) and IL-5 (right panels) of peripheral blood eosinophils from S. mansoni infected patients, includingintestinal (INT: = 19) and those with periportal fibrosis (FIB: = 18) as compared to non-infected individuals (NI: = 8), following short-term in vitro controlculture of whole blood samples (top panel) and after short-term stimulation with soluble egg S. mansoni antigen, SEA (middle panel) or soluble worm antigen preparationfrom S. mansoni, SWAP (bottom panel). The selection of eosinophils was essentially based on autofluorescent cells using non-related FL-3 channel versus forward scatter (FSC)and the analysis of cytokine-positive eosinophils was carry out by single color immunophenotyping. The results are expressed in box-plot format highlighting the gap of 50%o e box( ibutiob 0.05 a
gt(
stwfibda(
3o
csvaTsrthS
Fili
itawDeetpcclms
cs
f data set measurement for the absolute counts of cytokine positive eosinophils. Ththe 75th percentile) and therefore contains the middle half of the score in the distretween this line and the bottom or the top of the box. Significant differences at p <
esting that immunoregulatory events may take place controllinghe early eosinophil activation in these asymptomatic patientsFig. 1).
Considering that egg burdens are a proxy for in vivo SEA expo-ure, further analysis was performed in order to evaluate whetherhe levels of activation markers was somehow related to egg countsithin each disease groups. Even thought the egg count did not dif-
er between INT and FIB group as reported in material and methods,t would be still possible that the levels of activation markers maye related to egg count within each disease groups. Data analysisid not show any significant correlation between the egg countsnd the eosinophil activation status of both INT and FIB patientsdata not shown).
.2. High expression of cytokine by eosinophils is selectivelybserved in patients with periportal fibrosis
Analysis of cytokine expression by circulating eosinophils wasarried out after in vitro short-term incubation of whole bloodamples in the absence (control cultures) to access the basal exivo cytokine profile and in the presence of S. mansoni-derivedntigens to evaluate the inducible cytokine pattern. Analysis ofNF-�, IL-4 and IL-5 are shown in Fig. 2. Data analysis demon-
trated an increment of all these cytokines in the FIB group,egardless the culture condition. Specifically, at the basal level,he cytokine profile observed in the FIB group was significantlyigher when compared to both NI and INT. Interestingly, uponEA stimulation, higher levels of cytokines were observed inc�Ftp
stretches from the lower hinge (defined as the 25th percentile) to the upper hingen. The median is shown as a line across the box. Therefore 1/4 of the distribution isre highlighted by connecting lines.
IB in comparison to INT, re-emphasizing that the SEA-inducedmmune response is a relevant event associated with the estab-ishment/maintenance of distinct clinical status during S. mansoninfection (Fig. 2).
As previously mentioned, as the egg counts might represent anndicative biomarker for in vivo SEA exposure, it would be feasibleo hypothesize that despite no differences between the egg countmong INT and FIB, the levels of in vitro SEA-induced cytokineould be related to the egg counts within each disease groups.ata analysis pointed out that in vitro SEA-induced IL-5 synthesis byosinophils was the single parameter positively correlated with theggs counts but restricted to the INT group. No significant correla-ion between egg counts and the other eosinophil-derived cytokinerofiling were found in the INT group. Additionally, no significantorrelation was observed between egg counts and eosinophil intra-ellular cytokine levels in the FIB group, suggesting that higherevels of cytokines were observed in FIB group may represent a
ore complex mechanism regardless the egg counts (data nothown).
On the other hand, SWAP stimulation was the only experimentalondition able to trigger higher cytokines (TNF-� and IL-5) expres-ion in INT when compared to NI (Fig. 2).
Interestingly, correlation analysis demonstrated that a mixed
ytokine pattern, characterized by the parallel increment of TNF-+ with IL-4+ and IL-5+ eosinophils was observed in both INT andIB (Fig. 3). Then, despite the higher cytokine levels observed selec-ively in FIB, both groups of patients presented a similar mixedattern of eosinophil-derived cytokines.
D. Silveira-Lemos et al. / Acta Tropica 108 (2008) 150–159 155
Fig. 3. Correlation between the absolute numbers of TNF-�+ eosinophils with IL-4+ and IL-5+ eosinophils in peripheral blood from S. mansoni infected patients includingintestinal (INT: = 19) and those with periportal fibrosis (FIB: = 18), following in vitro short-term control culture of whole blood samples (top panel) and aftershort-term stimulation with soluble egg S. mansoni antigen, SEA (middle panel) or soluble worm antigen preparation from S. mansoni, SWAP (bottom panel). The results aree for eaa sidere
3o
tipt1(tote
mis
ecaacreTilofi
pdc
pbffitotFiast
4
aIiei
xpressed as scattering of paired absolute numbers of cytokine-positive eosinophilss well as curve fit regression are shown on graphs. Significant differences were con
.3. Decoding the mixed eosinophil-derived cytokine patternbserved in INT and FIB
Aiming to decode whether the similar mixed cytokine pat-ern observed in both group would be associated with a distinctmmunological network, we intended to further characterizeotential correlation between the eosinophil cytokine profiles withheir ex vivo activation status (“Mechanism 1”) as well as the IL-0 produced by circulating T-lymphocytes in the control culture“Mechanism 2”). These conditions were chosen considering thathe ex vivo analysis of activation status as well as the detectionf cytokine, particularly in the absence of exogenous stimuli (con-rol cultures) may reflect the dynamic events taking place in bloodosinophils in vivo.
“Mechanism 1”: Lack of association between activation-relatedarkers and cytokine expression by eosinophils suggest an
mpairment of immunomodulatory activity during chronic fibroticchistosomiais.
Analysis of the correlation indexes regarding the ex vivoxpression of cell activation markers and the basal levels ofytokine-expressing eosinophils observed in the control culturesre shown in Table 2. Data analysis showed that despite a directssociation between these parameters in the INT group, particularlyoncerning the associations with the IL-4 synthesis, the lack of cor-elation between the expression of activation markers and cytokinexpression by eosinophils was the hallmark of the FIB group.
hese data pointed out to the fact that even FIB patients present-ng minor increment on the eosinophil activation display higherevels of cytokine-positive eosinophils, suggesting an impairmentf immunoregulatory mechanisms in patients bearing periportalbrosis.1tdSc
ch patient in the INT and FIB groups. Spearman correlation indexes (r and p values)d at p < 0.05.
“Mechanism 2”: IL-10 produced by lymphocytes emerges as autative immunoregulatory mechanism to control the eosinophil-erived cytokines and assure asymptomatic clinical status duringhronic S. mansoni infection.
Considering that IL-10 derived from T-lymphocytes has beenointed out as a putative immunoregulatory event controlling mor-idity during chronic human schistosomiasis mansoni, we haveurther investigated whether the selective eosinophil cytokine pro-le observed for INT and FIB may reflect a distinct association withhe level of IL-10+ lymphocytes (Fig. 4). For this purposes the levelsf cytokine-positive eosinophil and IL-10+ lymphocytes observed inhe control cultures were paired for each individual in the INT andIB groups. Data analysis pointed out that only INT presented a pos-tive correlation between the lymphocyte-derived IL-10 synthesisnd the profile of eosinophil cytokine synthesis, further empha-izing our hypothesis that impaired immunoregulatory responseakes place in patients bearing periportal fibrosis (Fig. 4).
. Discussion
Eosinophils are pro-inflammatory cells associated with bothllergic diseases and parasitic infections (Thorne and Mazza, 1991).n the latter it has been shown to be a hallmark of the helminthsnfections. Although in parasitic infections, elevated eosinophil lev-ls have been associated with protective response to helminthsnfections (Colley, 1972; Moore et al., 1977; Butterworth et al.,
979; Thorne and Mazza, 1991), recent studies have pointed outhat under specific conditions, eosinophil derived factors can causeamage and contribute to the tissue pathology (Munitz and Levi-chaffer, 2004). A particular property of eosinophils is that theyan, like macrophages, exist in different states of activation and
156 D. Silveira-Lemos et al. / Acta Tropica 108 (2008) 150–159
Table 2Correlation indexes between eosinophil phenotypic features and cytokine profile in Schistosoma mansoni chronic infected patients
Phenotypic features Clinical groups
Intestinal Periportal fibrosis
TNF-� IL-4 IL-5 TNF-� IL-4 IL-5
CD69 r – – – – – –p – – – – – –
CD25 r 0.7083 0.8225 0.6559 – – –p 0.0015 <0.0001 0.0050 – – –
HLA-DR r 0.5996 0.7211 0.5025 – – –p 0.0085 0.0005 0.0398 – – –
CD23 r 0.6760 0.7523 0.7231 – – –p 0.0021 0.0003 0.0035 – – –
CD28 r 0.6509 0.7947 – – – –p 0.0025 0.0001 – – – –
C 21
–
bga(iomisi
totlh(
FiTS
D80 r 0.6018 0.784p 0.0064 <0.000
: non-significant correlation indexes.
e a source of a significant number of different cytokines androwth factors and therefore display both pro-inflammatory ornti-inflammatory activities as well as immunoregulatory onesMunitz and Levi-Schaffer, 2004). These properties may havemportant consequences for the eosinophils in either protectiver pathological events during human schistosomiasis. This is even
ore important when we take into consideration that eosinophilias a prominent feature of the innate cellular response in S. man-oni infection. Infection with this trematode worm causes dramaticncreases in levels of eosinophils in peripheral blood throughout
pcth
ig. 4. Correlation between the absolute numbers of TNF-�+ (left panel) and IL-4+ (righnfected patients, including intestinal (INT: = 19) and those with periportal fibrosis (Fhe results are expressed as scattering of paired absolute numbers of cytokine-positive epearman correlation indexes (r and p values) as well as curve fit regression are shown on
– – – –– – – –
he disease onset and progression as well as in the granulomas,bserved in host tissues, induced by the eggs of the parasite duringhe acute infection (Moore et al., 1977; Colley, 1972). Therefore, bothocal and systemic eosinophilia occurs in schistosomiasis, whichas been considered a potent protective event during infectionThorne and Mazza, 1991). However, the lack of correlation between
rotective mechanisms and the level of peripheral blood eosinophilounts observed by Butterworth and Richardson (1985) may reflecthat the role of eosinophils in the immunological network duringuman schistosomiasis may involve higher complexity than pre-t panel) eosinophils with IL-10+ lymphocytes in peripheral blood from S. mansoniIB: �= 18), following short-term in vitro control culture of whole blood samples.
osinophils for each patient in the INT (top panels) and FIB groups (bottom panels).graphs. Significant differences were considered at p < 0.05.

cta Tro
vefStilfdt
eotapabmeffocl
wqtsoaitbbBsuigetbaseesda(vm
aegreoti
t
qcsmceug(scyabnirmaspiMiitecnuIioiidtIhoaedpfied
bptbcfic
tab
D. Silveira-Lemos et al. / A
iously expected (Thorne and Mazza, 1991). As recently shown,osinophils are associated with several fibrotic conditions with dif-erent etiopathologies (Munitz and Levi-Schaffer, 2004). In human. mansoni infection, it is an even more pertinent problem to inves-igate due to all the conditions described above. Therefore, we havenvestigated whether these cells may be involved on the immuno-ogical processes that develop during the chronic phase infection,ocusing mainly on whether these cells may play a role during theevelopment/maintenance of liver fibrosis besides its relevant pro-ective function in schistosomiasis.
We have recently addressed the activation status of circulatingosinophils from INT and demonstrated an increased frequencyf activated eosinophils with lower expression of early activa-ion marker CD69 in INT, suggesting a minor contribution of earlyctivation events during chronic asymptomatic disease. We havereviously proposed that early eosinophil activation events may becompartmentalized phenomenon, not detectable in peripheral
lood where these cells may be at late stages of activation beingore prone to migrate to the inflammatory focus (Silveira-Lemos
t al., 2006). Aiming to investigate the hypothesis that eosinophilsrom INT and FIB may differ on their activation status, we have per-ormed a parallel investigation to characterize the ex vivo profilef activation-related cell surface markers as well as the patterns ofytokines in peripheral blood eosinophils after short-term stimu-ation with S. mansoni-derived antigens.
Herein, our data demonstrated that higher eosinophils countsere observed selectively in INT, whereas FIB presented basal fre-
uency of circulating eosinophils, compatible to that observed inhe control group. In this context, it is important to mention thateveral studies have presented evidences that the effectivenessf eosinophils in the protective mechanisms during schistosomi-sis may not directly correlate with the number of cells presentn the peripheral blood (Butterworth and Richardson, 1985) andherefore the analysis of the activation status of these cells maye more relevant than the single analysis on increase/decrease oflood cell counts (Vadas et al., 1980; David et al., 1980; Veith andutterworth, 1983). Our major findings, regarding the activationtatus of eosinophils from INT and FIB group, points out that thep-regulation of activation-related surface markers on eosinophils
s the hallmark of chronic S. mansoni infection. These results sug-est that the enhanced of expression of CD28 by peripheral bloodosinophils from S. mansoni infected patients may be linked tohe well-known Type-2 response in helminthic infection. It haseen reported that, as mature lymphocytes, successful eosinophilctivation also requires cell/cell interaction mediated by the co-timulatory molecules via the CD28/B7-1/B7-2 pathway (Woerlyt al., 1999; Shi, 2004). Moreover, it has been demonstrated inxperimental schistosomiasis that CD28 may play a important rolehifting the immune response toward a Type-2 profile, since CD28-eficient mice (−/−) generated diminished egg antigen-driven IL-4nd IL-5 production compared to CD28-expressing (+/+) littermatesKing et al., 1996). Together, these findings indicated that CD28 acti-ation might contributes for the generation of a Th2 response to S.ansoni infection.
Interestingly, we have observed a distinct profile of eosinophilctivation between INT and FIB, with an up-regulation of CD69, anarly activation cell marker status, observed selectively in FIB, sug-esting that in INT patients immunoregulatory events that downegulate the expression of this cell marker may occur controllingarly eosinophil activation. These findings re-enforce our previ-
us observation that circulating eosinophils from INT may be athe late stages of activation being more prone to migrate to thenflammatory focus (Silveira-Lemos et al., 2006).Another relevant issue investigated was whether the pro-ective/pathological role of eosinophils may involve not only a
TthF2
pica 108 (2008) 150–159 157
ualitative change in their activation state but actually signifi-ant alteration on their functional capacity to produce and releaseoluble factors that render circulating eosinophils effective inediating the control or induction of pathological conditions asso-
iated with S. mansoni infection. It has been demonstrated thatosinophils from helminth-infected patients exhibit, in addition top-regulation of surface activation molecules, enhanced release ofranule proteins, cytokines and other mediators of inflammationBochner, 2000). Indeed, eosinophils are a prominent cell withinchistosome granulomas and are known to produce a variety ofytokines (Weller, 1994). Of special interest in this study is the anal-sis of the production of cytokines by circulating eosinophils. Thenalysis of TNF-�, IL-4 and IL-5 profiles following short-term incu-ation with S. mansoni soluble antigens demonstrated that higherumber of cytokine-positive eosinophils was observed selectively
n the FIB group. These findings suggest that besides the up-egulation of TNF-�, a Type-2 response mediated by IL-4 and IL-5ay also be significantly important on the development of hep-
tic fibrosis and chronic morbidity in FIB. Several investigators havehown that a shift from a Type-1 to a Type-2 cytokine patterns hasrofound effects on the ability of the host to respond to S. mansoni
nfection (Araujo et al., 1996; Malaquias et al., 1997; Pearce andacdonald, 2002). It has been recognized for quite some time that
t is the egg stage of the schistosome that is responsible for induc-ng the Type-2 response during infection. By contrast, the wormhemselves seem to be poor inducers of a Type-2 response (Pearcet al., 1991; Grzych et al., 1991). Analysis of the SEA/SWAP inducedytokine ratio confirmed these reports, demonstrating that despiteo differences between INT and FIB in comparison to SWAP stim-lated cultures, SEA induced preferentially IL-5 (data not shown).
nterestingly, the SEA/SWAP TNF-� ratio was significantly highern FIB when compared to INT (data not shown) suggesting a lackf SEA-induced modulatory events in this group. Besides thosenvestigation showing that TNF-� plays a central role in promot-ng periportal fibrosis during S. mansoni infection, it has also beenemonstrated that pathological fibrosis can also be associated withhe chronic up-regulated production of Type-2 cytokines, IL-4 andL-13 in response to the parasite eggs in mice (Wynn, 2004) andumans (Silveira et al., 2004). IL-4 is important for the initiationf Type-2 cytokine responses during schistosomiasis (Chensue etl., 1992), while IL-13 is the key mediator of fibrosis (Chiaramontet al., 1999, 2003; Silveira et al., 2004). Additionally, it has beenemonstrated that eosinophils producing pro-fibrotic mediatorsolarize the Th2 response illustrating both direct and indirect rolesor eosinophils and IL-5 in the pathogenesis of schistosomiasis-nduced liver fibrosis. Therefore, inhibiting the activity of IL-5 orosinophils may prove effective for a variety of chronic fibroticiseases (Reiman et al., 2006).
Despite the higher cytokine levels observed selectively in FIB,oth groups of patients (INT and FIB) presented a similar mixedattern of eosinophil-derived cytokines. Aiming to decode whetherhe similar mixed cytokine pattern observed in both groups woulde associated with a distinct immunological network, we furtherharacterized the correlation between the eosinophil cytokine pro-les with the IL-10 produced by circulating T-lymphocytes in theontrol culture.
It has been proposed that both systemic and granuloma-ous eosinophilia appear to be T-lymphocyte dependent (Bastennd Beeson, 1970). Indeed, eosinophil production is controlledy T-lymphocyte-derived factors (IL-3, GM-CSF and IL-5). Since
-lymphocyte-derived IL-10 has been pointed out as an impor-ant immunoregulatory component controlling morbidity duringuman chronic schistosomiasis mansoni (Malaquias et al., 1997;alcao et al., 1998; de Jesus et al., 2004; Teixeira-Carvalho et al.,008), attempts to correlate the eosinophil cytokine profile with the
1 cta Tro
lddoeip
w(smpHihdptaa
sosfihpocTcamoonotmIeltrad
rbImteifi
A
Rs(t
LB
R
A
A
A
A
B
B
B
B
B
B
B
C
C
C
C
C
C
D
d
D
F
G
58 D. Silveira-Lemos et al. / A
evels of IL-10+ lymphocytes in INT and FIB were carried out. Ourata points out to a strong association between the lymphocyte-erived IL-10 and the lower cytokine profile from eosinophilsbserved in INT. The lack of correlation between these two param-ters in the FIB group further emphasizes our hypothesis thatmpaired immunoregulatory events occur in patients bearing peri-ortal fibrosis.
It has been previously suggested that risk of fibrosis is associatedith low production of IL-10 in cases of S. mansoni-derived fibrosis
Booth et al., 2004). Study of Alves-Oliveira et al. (2006) demon-trated that during chronic schistosomiasis, infected patients withoderate/severe fibrosis presented lower levels of IL-10 when com-
ared to asymptomatic schistosome infected subjects. Moreover,offmann et al. (2000) established in experimental S. mansoni
nfection that mice deficient in IL-10 display increased risk forepatic fibrosis. In addition, others studies by our group haveemonstrated that IL-10 is an important cytokine to reduce theathology of acute schistosomiasis (Falcao et al., 1998), regulatinghe immune response and possibly controlling morbidity in chronicsymptomatic patients (Malaquias et al., 1997; Correa-Oliveira etl., 1998; de Jesus et al., 2004).
Taken together, despite the limitations regarding the cross-ectional design of this study and the relatively small sample size,ur major findings pointed out that eosinophils in chronic humanchistosomiasis, might contribute to the development of hepaticbrosis and chronic morbidity infection. Our data demonstratedigher activation status of circulating eosinophils in FIB as com-ared to INT. Higher levels of eosinophil-derived cytokines werebserved in FIB, regardless the antigen stimulation in vitro. A mixedytokine pattern, characterized by positive correlation betweenNF-�, IL-4 and IL-5 was observed in both INT and FIB. Lack oforrelation between the cytokine expression and the eosinophilctivation status points out that even those FIB patients presentinginor increment on eosinophil activation displayed higher levels
f cytokine-positive eosinophils. These results re-enforce previ-us evidences that peripheral blood eosinophilia by itself doesot reflect the qualitative change in eosinophil functional activityr state of pathology development in S. mansoni infection. Fur-hermore, our results show disrupted modulatory mechanisms,
ediated by damaged relationship between T-lymphocyte-derivedL-10 and the up-regulated levels of cytokines produced byosinophil, which may influence the development/maintenance ofiver fibrosis during chronic infection with S. mansoni. It is impor-ant to mention that we were aware of possible study limitationsegarding potential confounders (such as alcoholism and/or hep-titis B and C infection) which have been used as exclusion criteriauring patient recruitment.
In conclusion, our major findings, pointed out a distinct up-egulation of activation-related surface markers on eosinophilsesides higher levels of cytokines, including TNF-�, IL-4 and
L-5, as the hallmark of periportal fibrosis during chronic S.ansoni infection. Additionally, the lack of association between
he lymphocyte-derived IL-10 and the cytokine profile fromosinophils, further emphasized our hypothesis that impairedmmunoregulatory events take place in patients bearing periportalbrosis.
cknowledgments
This work was supported by Centro de Pesquisas Reneachou, Fundacao Oswaldo Cruz (CPqRR/FIOCRUZ-Brazil), Con-elho Nacional de Desenvolvimento Cientıfico e TecnologicoCNPq-Brazil) and National Institutes of Health (PA-98-078). Wehank Pollyanna Castro e Silva and the technical support from the
G
pica 108 (2008) 150–159
aboratorio de Imunologia Celular e Molecular at CPqRR/FIOCRUZ,razil.
eferences
bdel-Wahab, M.F., Esmat, G., Farrag, A., El-Boraey, Y.A., Strickland, G.T., 1992. Grad-ing of hepatic schistosomiasis by the use of ultrasonography. Am. J. Trop. Med.Hyg. 46, 403–408.
lves-Oliveira, L.F., Moreno, E.C., Gazzinelli, G., Martins-Filho, O.A., Silveira, A.M.S.,Gazzinelli, A., Malaquias, L.C.C., LoVerde, P., Leite, P.M., Correa-Oliveira, R., 2006.Cytokine production associated with periportal fibrosis during chronic Schisto-somiasis mansoni in humans. Infect. Immun. 74, 1215–1221.
nwar, A.R.E., Smithers, S.R., Kay, A.B., 1979. Killing of schistosomula of Schistosomamansoni coated with antibody and/or complement by human leukocytes in vitro:requirements for complement in preferential killing by eosinophils. J. Immunol.122, 628–637.
raujo, M.I., de Jesus, A.R., Bacellar, O., Sabin, E., Pearce, E., Carvalho, E.M., 1996. Evi-dence of a T helper type 2 activation in human schistosomiasis. Eur. J. Immunol.26, 1399–1403.
asten, A., Beeson, P.B., 1970. Mechanism of eosinophilia. II. Role of the lymphocyte.J. Exp. Med. 131, 1288–1305.
irkland, T.P., Cheavens, M.D., Pincus, S.H., 1994. Human eosinophils stimulate DNAsynthesis and matrix production in dermal fibroblasts. Arch. Dermatol. Res. 286,312–318.
ochner, B.S., 2000. Systemic activation of basophils and eosinophils: markers andconsequences. J. Allergy Clin. Immunol. 106, S292–S302.
ooth, M., Mwatha, J.K., Joseph, S., Jones, F.M., Kadzo, H., Ireri, E., Kazibwe, F.,Kemijumbi, J., Kariuki, C., Kimani, G., Ouma, J.H., Kabatereine, N.B., Birgitte, J.,Vennervald, B.J., Dunne, D.W., 2004. Periportal fibrosis in human Schistosomamansoni infection is associated with low IL-10, low IFN-�, high TNF-�, or lowRANTES, depending on age and gender. J. Immunol. 172, 1295–1303.
utterworth, A.E., Wassom, D.L., Gleich, G.J., Loegering, D.A., David, J.R., 1979. Dam-age to schistosomula of Schistosoma mansoni induced directly by eosinophilmajor basic protein. J. Immunol. 122, 221–229.
utterworth, A.E., 1984. Cell-mediated damage to helminths. Adv. Parasitol. 23,143–235.
utterworth, A.E., Richardson, B.A., 1985. Factors affecting the levels of antibody-and complement-dependent eosinophil-mediated damage to schistosomula ofSchistosoma mansoni in vitro. Parasite Immunol. 7, 119–131.
hensue, S.W., Terebuh, P.D., Warmington, K.S., Hershey, S.D., Evanoff, H.L., Kunkel,S.L., Higashi, G.I., 1992. Role of IL-4 and IFN-gamma in Schistosoma mansoniegg-induced hypersensitivity granuloma formation. Orchestration, relative con-tribution, and relationship to macrophage function. J. Immunol. 148, 900–906.
hiaramonte, M.G., Donaldson, D.D., Cheever, A.W., Wynn, T.A., 1999. IL-13 inhibitorblocks the development of hepatic fibrosis during a T-helper type 2-dominatedinflammatory response. J. Clin. Invest. 104, 777–785.
hiaramonte, M.G., Mentink-Kane, M., Jacobson, B.A., Cheever, A.W., Whitters, M.J.,Goad, M.E., Wong, A., Collins, M., Donaldson, D.D., Grusby, M.J., Wynn, T.A., 2003.Regulation and function of the interleukin 13 receptor alpha 2 during a T helpercell type 2-dominant immune response. J. Exp. Med. Mar. 197, 687–701.
olley, D.G., 1972. Schistosoma mansoni: eosinophilia and the development of lym-phocyte blastogenesis in response to soluble egg antigen in inbred mice. Exp.Parasitol. 32, 520–526.
orrea-Oliveira, R., Malaquias, L.C.C., Falcao, P.L., Viana, I.R., Bahia-Oliveira, L.M., Sil-veira, A.M.S., Fraga, L.A., Prata, A., Coffman, R.L., Lambertucci, J.R., Cunha-Melo,J.R., Martins-Filho, O.A., Wilson, R.A., Gazzinelli, G., 1998. Cytokines as determi-nants of resistance and pathology in human Schistosoma mansoni infection. Braz.J. Med. Biol. Res. 31, 171–177.
ox, F., 1998. Immune effectors mechanisms in parasitic infections. Parasitol. Today14, 504.
avid, J.R., Butterworth, A.E., Vadas, M.A., 1980. Mechanism of interaction mediatingkilling of Schistosoma mansoni by human eosinophils. Am. J. Trop. Med. Hyg. 29,842–848.
e Jesus, A.R., Magalhaes, A., Miranda, D.G., Miranda, R.G., Araujo, M.I., Jesus, A.A.,Silva, A., Santana, L.B., Pearce, E., Carvalho, E.M., 2004. Association of type 2cytokines with hepatic fibrosis in human Schistosoma mansoni infection. Infect.Immun. 72, 3391–3397.
oucet, C., Brouty-Boye, D., Pottin-Clemenceau, C., Jasmin, C., Canonica, G.W.,Azzarone, B., 1998. IL-4 and IL-13 specifically increase adhesion molecule andinflammatory cytokine expression in human lung fibroblasts. Int. Immunol. 10,1421–1433.
alcao, P.L., Malaquias, L.C.C., Martins-Filho, O.A., Silveira, A.M.S., Passos, M., Prata,A., Gazzinelli, G., Coffman, R.L., Correa-Oliveira, R., 1998. Human schistosomiasismansoni: IL-10 modulates the in vitro granuloma formation. Parasite Immunol.20, 447–454.
azzinelli, G., Katz, N., Rocha, R.S., Colley, D.G., 1983. Immune response dur-ing human schistosomiasis mansoni. X. Production and standardization of
an antigen-induced mitogenic activity by peripheral blood mononuclearcells from treated, but not active cases of schistosomiasis. J. Immunol. 130,2891–2895.rzych, J.M., Pearce, E., Cheever, A., Caulada, Z.A., Caspar, P., Heiny, S., Lewis, F., Sher,A., 1991. Egg deposition is the major stimulus for the production of Th2 cytokinesin murine schistosomiasis mansoni. J. Immunol. 146, 1322–1327.

cta Tro
H
K
K
L
L
M
M
M
M
N
P
P
R
R
S
S
S
S
S
T
T
T
T
W
W
W
W
V
D. Silveira-Lemos et al. / A
offmann, K.F., Cheever, A.W., Wynn, T.A., 2000. IL-10 and the dangers of immunepolarization: excessive type 1 and type 2 cytokine responses induce distinctforms of lethal immunopathology in murine schistosomiasis. J. Immunol. 164,6406–6416.
atz, N., Chaves, A., Pellegrino, J., 1972. A simple device for quantitative stool thicksmear technique in Schistosoma mansoni. Rev. Inst. Med. Trop. Sao Paulo 14,397–400.
ing, C.L., Xianli, J., June, C.H., Abe, R., Lee, K.P., 1996. CD28-deficient mice generatean impaired Th2 response to Schistosoma mansoni infection. Eur. J. Immunol. 26,2448–2455.
evi-Schaffer, F., Garbuzenko, E., Rubin, A., Reich, R., Pickholtz, D., Gillery, P., 1999.Human eosinophils regulate human lung- and skin-derived fibroblast propertiesin vitro: a role for transforming growth factor beta (TGF-beta). Proc. Natl. Acad.Sci. U.S.A. 96, 9660–9665.
iu, X., Kohyama, T., Wang, H., 2002. Th2 cytokine regulation of type I collagen gelcontraction mediated by human lung mesenchymal cells. Am. J. Physiol. LungCell. Mol. Physiol. 282, L1049–L1056.
agalhaes, T.V., Gazzinelli, G., Alvarez, M.C., Lima e Silva, F.C., Fraga, L.A., Silveira,A.M., Gazzinelli, A., Bethony, J., LoVerde, P., Caldas, I.R., Correa-Oliveira, R., Prata,A., 2005. Comparative clinical and ultrasound study of egg-negative and egg-positive individuals from Schistosoma mansoni low morbidity endemic areas,and hospitalized patients with hepatosplenic disease. Rev. Soc. Bras. Med. Trop.38, 33–37.
alaquias, L.C.C., Falcao, P.L., Silveira, A.M.S., Gazzinelli, G., Prata, A., Coffmann, R.L.,Pizziolo, V.R., Correa-Oliveira, R., 1997. Cytokine regulation of human immuneresponse to Schistosoma mansoni: analysis of the role of IL-4, IL-5 and IL-10 onperipheral blood mononuclear cell responses. Scand. J. Immunol. 46, 393–398.
oore, D.L., Grove, D.I., Warren, K.S., 1977. The Schistosoma mansoni egg granuloma:quantitation of cell populations. J. Pathol. 121, 41–50.
unitz, A., Levi-Schaffer, F., 2004. Eosinophils: “new” roles for “old” cells. Allergy59, 268–275.
oguchi, H., Kephart, G.M., Colby, T.V., Gleich, G.J., 1992. Tissue eosinophilia andeosinophil degranulation in syndromes associated with fibrosis. Am. J. Pathol.140, 521–528.
earce, E.J., Macdonald, A.S., 2002. The immunobiology of schistosomiasis. Nat. Rev.Immunol. 7, 499–511.
earce, E.J., Casper, P., Grzych, J.M., Lewis, F.A., Sher, A., 1991. Downregulation ofTh1 cytokine production accompanies induction of Th2 responses by a parasitichelminth, Schistosoma mansoni. J. Exp. Med. 173, 159–166.
amalho-Pinto, F.J., McLaren, D.J., Smithers, S.R., 1978. Complement-mediatedkilling of schistosomula of Schistosoma mansoni by rat eosinophils in vitro. J.
Exp. Med. 147, 147–156.eiman, R.M., Thompson, R.W., Feng, C.G., Hari, D., Knight, R., Cheever, A.W., Rosen-berg, H.F., Wynn, T.A., 2006. Interleukin-5 (IL-5) augments the progression ofliver fibrosis by regulating IL-13 activity. Infect. Immun. 74, 1471–1479.
her, A., Butterworth, A.E., Colley, D.G., Cook, A.J., Freeman, G.L.R., Jordan, P., 1977.Immune responses during human schistosomiasis mansoni. II. Occurrence of
V
pica 108 (2008) 150–159 159
eosinophil-dependent cytotoxic antibodies in relation to intensity and durationof infection. Am. J. Trop. Med. Hyg. 26, 909–916.
hi, H.Z., 2004. Eosinophils function as antigen-presenting cells. J. Leukoc. Biol. 76,520–527.
hock, A., Rabe, K.F., Dent, G., 1991. Eosinophils adhere to and stimulate replicationof lung fibroblasts in vitro. Clin. Exp. Immunol. 86, 185–190.
ilveira, A.M., Gazzinelli, G., Alves-Oliveira, L.F., Bethony, J., Gazzinelli, A., Carvalho-Queiroz, C., Alvarez, M.C., Lima-Silva, F.C., Prata, A., LoVerde, P.T., Correa-Oliveira,R., 2004. Human schistosomiasis mansoni: intensity of infection differentiallyaffects the production of interleukin-10, interferon-gamma and interleukin-13by soluble egg antigen or adult worm antigen stimulated cultures. Trans. R. Soc.Trop. Med. Hyg. 98, 514–519.
ilveira-Lemos, D., Teixeira-Carvalho, A., Martins-Filho, O.A., Alves Oliveira, L.F.,Correa-Oliveira, R., 2006. High expression of co-stimulatory and adhesionmolecules are observed on eosinophils during human Schistosoma mansoniinfection. Mem. Inst. Oswaldo Cruz 101, 345–351.
eixeira-Carvalho, A., Martins-Filho, O.A., Peruhype-Magalhães, V., Silveira-Lemos,D., Malaquias, L.C.C., Alves Oliveira, L.F., Silveira, A.M.S., Gazzinelli, A., Alvarez,M.C.B., Gazzinelli, G., Corrêa-Oliveira, R., 2008. Cytokines, chemokine receptors.CD4+CD25HIGH+ T-cells and clinical forms of human schistosomiasis. Acta Trop.108, 139–149.
eran, L.M., Mochizuki, M., Bartels, J., 1999. Th1- and Th2-type cytokines regulate theexpression and production of eotaxin and RANTES by human lung fibroblasts.Am. J. Respir. Cell Mol. Biol. 20, 777–786.
horne, K.J.I., Mazza, G., 1991. Eosinophilia, activated eosinophils and human schis-tosomiasis. J. Cell Sci. 98, 265–270.
odd, R., Donoff, B.R., Chiang, T., 1991. The eosinophil as a cellular source of trans-forming growth factor alpha in healing cutaneous wounds. Am. J. Pathol. 138,1307–1313.
eller, P.F., 1994. Eosinophils: structure and functions. Curr. Opin. Immunol. 6,85–90.
oerly, G., Roger, N., Loiseau, S., Dombrowicz, D., Capron, A., Capron, M., 1999.Expression of CD28 and CD86 by human eosinophils and role in the secre-tion of type 1 cytokines (interleukin 2 and interferon gamma): inhibition byimmunoglobulin a complexes. J. Exp. Med. 190, 487–495.
ong, D.T., Donoff, R.B., Yang, J., 1993. Sequential expression of transforming growthfactors alpha and beta 1 by eosinophils during cutaneous wound healing in thehamster. Am. J. Pathol. 143, 130–142.
ynn, T.A., 2004. Fibrotic disease and the Th1/Th2 paradigm. Nat. Rev. Immunol. 8,583–594.
adas, M.A., David, J.R., Butterworth, A.E., Houba, V., Sturrock, R.F., David, L., Herson,
R., Siongok, T.A., Kimani, R., 1980. Functional studies on purified eosinophilsand neutrophils from patients with Schistosoma mansoni infections. Clin. Exp.Immunol. 39, 683–694.eith, M.C., Butterworth, A.E., 1983. Enhancement of human eosinophil-mediatedkilling of Schistosoma mansoni larvae by mononuclear cell products in vitro. J.Exp. Med. 157, 1828–1843.




















