
SmCB2, a novel tegumental cathepsin B from adult Schistosoma mansoni
13
SmCB2, a novel tegumental cathepsin B from adult Schistosoma mansoni Conor R. Caffrey a, *, Jason P. Salter a , Kimberley D. Lucas a , Dustin Khiem a , Ivy Hsieh a , Kee-Chong Lim a , Andreas Ruppel b , James H. McKerrow a , Mohammed Sajid a a Tropical Disease Research Unit, Department of Pathology, Box 0511, University of California San Francisco, San Francisco, CA 94143, USA b Abteilung fu ¨r Tropenhygiene und o ¨ffentliches Gesundheitswesen, Hygiene Institut, Ruprecht-Karls-Universita ¨t Heidelberg, Im Neuenheimer Feld 324, Heidelberg 69120, Germany Received 17 October 2001; received in revised form 22 January 2002; accepted 25 January 2002 Abstract Papain-like cysteine endopeptidases have been recognized as potential targets for chemotherapy and serodiagnostic reagents in infections with the human parasitic helminth Schistosoma . A novel cathepsin B endopeptidase from adult S. mansoni has been isolated and characterized. The enzyme is termed SmCB2 to distinguish it from the first recorded schistosome cathepsin B, SmCB1, also known as Sm31. A rapid and convenient protocol involving anion exchange and affinity chromatography is described for the isolation of SmCB1 and SmCB2 from the same parasite starting material. SmCB2 has been functionally expressed in and purified from Pichia pastoris . Both native and recombinant SmCB2 migrate similarly (33 kDa) by SDS-PAGE. Both display strict acidic pH activity profiles and similar K m and k cat for dipeptidyl amidomethylcoumarin substrates. We conclude that the recombinant enzyme is properly folded. The S 2 subsite specificity of recombinant SmCB2 exhibits the preferences Phe /Leu /Val /Arg. By immunoblotting with anti-SmCB2 IgG, a 33 kDa protein was identified in soluble extracts of male schistosomes. By immunohistochemistry, SmCB2 was localized in the tegumental tubercles and parenchyma of males with less product being visualized in the parenchyma of females. The enzyme may be lysosomal and function at the host parasite-interface. # 2002 Elsevier Science B.V. All rights reserved. Keywords: Schistosoma ; Cysteine endopeptidase; SmCB2; Recombinant expression; Chemotherapy; Diagnosis 1. Introduction Helminth endoparasites of the genus Schistosoma are a major public health concern in 74 tropical and sub- tropical countries. Over 200 million people are infected with 20 million people being severely debilitated by the disease schistosomiasis [1]. Adult worms live in the mesenteric veins and portal vein leading to the liver and feed on blood. Schistosomiasis is associated with severe morbidity mainly arising from immunopathological reactions to parasite eggs in various tissues, particularly in the liver, intestinal tract and/or bladder. The characterization and isolation of schistosome proteolytic enzymes (peptidases) is driven in large part by their potential as chemotherapeutic targets. Indeed, targeting peptidases with small-molecule inhibitors has been shown to inhibit skin penetration by larval schistosomes [2], and decrease worm burden and egg production by adult Schistosoma [3]. In addition, schistosome endopeptidases have a demonstrated value as selective diagnostic markers of disease [4 /6]. Most studies concerning peptidases of the adult schistosome have dealt with cysteine endopeptidases Abbreviations: AEC, aminoethyl carbazole; BCIP, 5-bromo-4- chloro-3-indolyl-1-phosphate; EST, expressed sequence tag; NBT, nitro blue tetrazolium; NMec, 7-amido-4-methylcoumarin; PVDF, polyvinylidene difluoride; TBS-T, Tris-buffered saline-Tween 20 solution; YPD, yeast extract-peptone-dextrose; Z, benzyloxycarbonyl. Note: Nucleotide sequence data reported in this paper have been submitted to the GenBank TM , EMBL and DDBJ databases with the accession number AJ312106. * Corresponding author. Tel.: 1-415-514-3052; fax: 1-415-514- 3165. E-mail address: [email protected] (C.R. Caffrey). Molecular & Biochemical Parasitology 121 (2002) 49 /61 www.parasitology-online.com 0166-6851/02/$ - see front matter # 2002 Elsevier Science B.V. All rights reserved. PII:S0166-6851(02)00022-1
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of SmCB2, a novel tegumental cathepsin B from adult Schistosoma mansoni

SmCB2, a novel tegumental cathepsin B from adult Schistosomamansoni�
Conor R. Caffrey a,*, Jason P. Salter a, Kimberley D. Lucas a, Dustin Khiem a,Ivy Hsieh a, Kee-Chong Lim a, Andreas Ruppel b, James H. McKerrow a,
Mohammed Sajid a
a Tropical Disease Research Unit, Department of Pathology, Box 0511, University of California San Francisco, San Francisco, CA 94143, USAb Abteilung fur Tropenhygiene und offentliches Gesundheitswesen, Hygiene Institut, Ruprecht-Karls-Universitat Heidelberg, Im Neuenheimer Feld 324,
Heidelberg 69120, Germany
Received 17 October 2001; received in revised form 22 January 2002; accepted 25 January 2002
Abstract
Papain-like cysteine endopeptidases have been recognized as potential targets for chemotherapy and serodiagnostic reagents in
infections with the human parasitic helminth Schistosoma . A novel cathepsin B endopeptidase from adult S. mansoni has been
isolated and characterized. The enzyme is termed SmCB2 to distinguish it from the first recorded schistosome cathepsin B, SmCB1,
also known as Sm31. A rapid and convenient protocol involving anion exchange and affinity chromatography is described for the
isolation of SmCB1 and SmCB2 from the same parasite starting material. SmCB2 has been functionally expressed in and purified
from Pichia pastoris . Both native and recombinant SmCB2 migrate similarly (33 kDa) by SDS-PAGE. Both display strict acidic pH
activity profiles and similar Km and kcat for dipeptidyl amidomethylcoumarin substrates. We conclude that the recombinant enzyme
is properly folded. The S2 subsite specificity of recombinant SmCB2 exhibits the preferences Phe�/Leu�/Val�/Arg. By
immunoblotting with anti-SmCB2 IgG, a 33 kDa protein was identified in soluble extracts of male schistosomes. By
immunohistochemistry, SmCB2 was localized in the tegumental tubercles and parenchyma of males with less product being
visualized in the parenchyma of females. The enzyme may be lysosomal and function at the host parasite-interface. # 2002 Elsevier
Science B.V. All rights reserved.
Keywords: Schistosoma ; Cysteine endopeptidase; SmCB2; Recombinant expression; Chemotherapy; Diagnosis
1. Introduction
Helminth endoparasites of the genus Schistosoma are
a major public health concern in 74 tropical and sub-
tropical countries. Over 200 million people are infected
with 20 million people being severely debilitated by the
disease schistosomiasis [1]. Adult worms live in the
mesenteric veins and portal vein leading to the liver and
feed on blood. Schistosomiasis is associated with severe
morbidity mainly arising from immunopathological
reactions to parasite eggs in various tissues, particularly
in the liver, intestinal tract and/or bladder.
The characterization and isolation of schistosome
proteolytic enzymes (peptidases) is driven in large part
by their potential as chemotherapeutic targets. Indeed,
targeting peptidases with small-molecule inhibitors has
been shown to inhibit skin penetration by larval
schistosomes [2], and decrease worm burden and egg
production by adult Schistosoma [3]. In addition,
schistosome endopeptidases have a demonstrated value
as selective diagnostic markers of disease [4�/6].
Most studies concerning peptidases of the adult
schistosome have dealt with cysteine endopeptidases
Abbreviations: AEC, aminoethyl carbazole; BCIP, 5-bromo-4-
chloro-3-indolyl-1-phosphate; EST, expressed sequence tag; NBT,
nitro blue tetrazolium; NMec, 7-amido-4-methylcoumarin; PVDF,
polyvinylidene difluoride; TBS-T, Tris-buffered saline-Tween 20
solution; YPD, yeast extract-peptone-dextrose; Z, benzyloxycarbonyl.�
Note: Nucleotide sequence data reported in this paper have been
submitted to the GenBankTM, EMBL and DDBJ databases with the
accession number AJ312106.
* Corresponding author. Tel.: �1-415-514-3052; fax: �1-415-514-
3165.
E-mail address: [email protected] (C.R. Caffrey).
Molecular & Biochemical Parasitology 121 (2002) 49�/61
www.parasitology-online.com
0166-6851/02/$ - see front matter # 2002 Elsevier Science B.V. All rights reserved.
PII: S 0 1 6 6 - 6 8 5 1 ( 0 2 ) 0 0 0 2 2 - 1

related to papain (Clan CA, family C1). Genes encoding
a cathepsin B (aka Sm31) [7], two cathepsin Ls (SmCL1
and SmCL2) [8,9], and an exopeptidase, cathepsin C
[10], have been characterized. These enzymes have been
functionally expressed in different recombinant systems
[11�/15]. The open reading frames (ORF) of all of the
above cysteine peptidases encode pre-pro-proteins ana-
logous to their respective mammalian orthologs and
homology modeling has been applied to predict three
dimensional structures [16�/18].
Perhaps the best clues to a biological function of these
peptidases have come from immunolocalization experi-
ments with specific antibodies. Thus, Sm31 [19], SmCL1
[14,20] and SmCL2 [20], being localized in the schisto-
some gut, are considered to act as alimentary endopep-
tidases. A function in schistosome reproduction has also
been proposed for SmCL2 due to its presence in the
female reproductive tract [9]. Finally, a cathepsin C-like
activity has also been localized in the gut by fluorescent
histochemistry [21].
In this report, we have characterized a cDNA
sequence encoding a novel cathepsin B endopeptidase,
termed SmCB2. For the sake of clarity and the establish-
ment of a systematic nomenclature, we rename Sm31,
the first cathepsin B gene described [7], as SmCB1. This
nomenclature is comparable, therefore, with that used
for the schistosome cathepsin L endopeptidases, SmCL1
and SmCL2. We describe a technique to simultaneously
isolate SmCB1 and SmCB2 from parasite extracts.
Further, SmCB2 was functionally expressed in Pichia
pastoris and isolated, and mono-specific antibodies
raised to localize the endopeptidase in adult parasite
tissues. Interestingly, the enzyme was preferentially
expressed in male worms and was localized in the
tegument in addition to the parenchyma. Both native
and recombinant SmCB2, along with native SmCB1 and
mammalian cathepsins B and L, were compared and
contrasted in their respective abilities to degrade a range
of dipeptidyl substrates.
2. Materials and methods
2.1. Parasites
S. mansoni (Puerto Rican isolate) was maintained by
cycling through the vector snail Biomphalaria glabrata
and NMRI mice or Syrian Golden hamsters. Adult
worms were obtained by portal perfusion of mice.
Specific techniques for the maintenance of the schisto-
some life cycle and collection of parasites have been
described previously [22].
2.2. Purification of SmCB1 and SmCB2 from parasite
extracts
Adult S. mansoni worms (400�/600 pairs) weresonicated on ice in 2�/3 ml 20 mM sodium phosphate
buffer, pH 6.0 and the homogenate centrifuged for 20
min at 10 000 g . The supernatant (soluble worm extract)
was sterile-filtered (0.2 mm; Millipore, Bedford, MA)
and then cycled three times over a 3 ml bed-volume of
DE-52 anion exchanger (Whatman, Maidstone, UK)
that had been pre-equilibrated with the same buffer. The
material not adsorbed to the column (wash fraction) andcontaining SmCB2 was cycled three times over a second
3 ml bed-volume of DE-52 pre-equilibrated with the
same buffer, and then made 2 mM with respect to DTT
(Sigma, St. Louis, MO). The wash fraction was mixed
end-over-end with a 6.5 ml slurry of Ahx-Sepharose 4B
(Sigma) that had been coupled to the reversible cathe-
psin B inhibitor H-Gly-Phe-Gly-semicarbazone (Ba-
chem, Torrance, CA) as described previously [23]. Theaffinity matrix had been pre-equilibrated with 50 mM
tri-sodium citrate, pH 5.0, 1 mM di-sodium-EDTA, 2
mM DTT. Isolation of SmCB2 was then continued as
described previously for the cathepsin B-like enzyme of
Schistosoma japonicum (SjCB1) [24].
In contrast to SmCB2, SmCB1 is adsorbed to the first
DE-52 column under the conditions used. After removal
of the wash fraction, the column was washed with a 10-fold bed-volume of equilibration buffer containing 100
mM NaCl. Then, material containing SmCB1 was
eluted with equilibration buffer containing 300 mM
NaCl. The eluate, usually 6�/8 ml, was concentrated to
1�/2 ml by ultrafiltration over a YM10 membrane
(Millipore). The solution was then made 2 mM with
respect to DTT and mixed with a 3.5 ml slurry of the
cathepsin B-affinity matrix for isolation of SmCB1 aspreviously described [24]. For both SmCB1 and SmCB2,
purification was monitored by SDS-PAGE [25] through
12.5% polyacrylamide gels and silver staining [26].
Endopeptidase activity was monitored by hydrolysis of
the fluorogenic peptidyl substrate benzyloxycarbonyl-
phenylalanyl-arginine-7-amido-4-methylcoumarin [27]
(see Section 2.10). Protein concentrations were deter-
mined with the micro-adaptation of the procedure ofBradford [28].
2.3. N-terminal sequencing
Purified native SmCB1, SmCB2 or recombinant
SmCB2 (see Section 2.6) was subjected to SDS-PAGE
and transferred to PVDF membranes (Immobilon-P;
Millipore) for 45 min at 25 V using the procedures and
apparatus supplied by Invitrogen. Protein bands werevisualized in 0.1% Ponceau S (Sigma) and destained in
water. Sequencing was performed using the Edman
degradation technique by Dr. Armin Bosserhoff of the
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/6150

ZMBH, Ruprecht-Karls Universitat, Heidelberg and
Dr. Ralph Reid at the UCSF Biomolecular Resource
Center.
2.4. Sequencing of SmCB2 cDNA
Poly A�/mRNA was isolated (Amersham Pharmacia
Biotech Inc., Piscataway, NJ) from 6-week-old S.
mansoni worms of mixed sex and converted to single-
stranded cDNA for subsequent polymerase chain reac-
tions (PCR) using AMV Reverse Transcriptase as
described by the manufacturer (Life Technologies,
Inc., Rockville, MD).To obtain sufficient SmCB2 cDNA for sequencing, a
semi-nested PCR was performed using two degenerate
forward primers derived from the N-terminal sequence
of native SmCB2 (see Fig. 2). The outer and inner
forward primers were 5?-CCNAARTCNTTYGAYGC-
NAG-3? and 5?-GAYGCNAGNGTNGARTGGCC-3?,respectively, and the reverse primer was a (dT)18
oligonucleotide (Roche Molecular Biochemicals, India-napolis, IN). The first PCR (100 ml final volume)
contained 50 ng single-stranded cDNA, 10 mM Tris�/
HCl, pH 8.85, 25 mM potassium chloride, 5 mM
ammonium sulphate, 2 mM magnesium sulphate, 0.2
mM dNTP, 0.1 nmol of the outer forward and the
reverse primers and 1.0 U Pwo polymerase (Roche). The
reaction was performed in a Perkin Elmer Thermal
Cycler with 35 cycles of 94, 48 and 72 8C, each for 1 min.On completion of the first PCR, 1 ml of the reaction
product was added as template for the second reaction
(performed as described above) with the inner forward
primer and the same reverse primer. PCR products were
resolved in 1% agarose gels containing 0.5 mg ml�1
ethidium bromide, and a band of approximately 1.0 kb
(based on the expected size of the mature catalytic
domain plus a 3?-UTR sequence) excised and purifiedusing silica beads (BIO 101, Vista, CA). Purified DNA
was submitted directly for sequencing using the inner
forward degenerate primer (UCSF Biomolecular Re-
source Facility or Sequetech Corp., Mountain View,
CA). From the sequence obtained, additional forward
and reverse primers were designed and used to sequence
the entire 1.0 kb PCR product. To allow for sequencing
of the SmCB2 ORF upstream of the N-terminus of thecatalytic domain, 5?-RACE was performed according to
the manufacturer’s instructions (Clontech, Palo Alto,
CA). Sequencing of this section of the ORF was
completed using a forward primer to the N-terminal
signal sequence and a reverse primer derived from the
catalytic domain. The overall sequencing strategy en-
sured that the entire SmCB2 ORF was sequenced in
both directions. The putative cleavage point between thesignal and pro-peptides was identified using the SignalP
V1.1 World Wide Web Prediction Server at the Center
for Biological Sequence Analysis, Denmark (http://
genome.cbs.dtu.dk/services/SignalP). Networks were
trained on sequences from eukaryotes and the software
is based on the algorithm of von Heijne [29]. A multiple
sequence alignment of the primary sequence of SmCB2with SmCB1 ([7]; accession number AAA29865), SjCB1
([30]; P43157) and human cathepsin B ([31]; P07858) was
developed using the CLUSTAL W program [32] avail-
able at the UCSF Sequence Analysis Resource Facility.
2.5. Design of a vector construct for expression of
SmCB2 in P. pastoris
The nucleotide sequence encoding the pro-domain ofSmCB2 was included in the expression vector construct
as it was found previously for cysteine-class cathepsins
that the pro-region is necessary for proper protein
folding [33]. Pro-SmCB2 was amplified by PCR from
single stranded cDNA derived from adult worms
(above). PCR primers were: forward, 5?- ATACTCGA-
GAAAAGAGATGCTAGACGACATAAACGTATG-
3? which incorporated an Xho I (Roche) site immedi-ately 5? of the P. pastoris Kex 2 endopeptidase recogni-
tion site (both sites are underlined); and reverse 5?-AATGCGGCCGCCTAGTTTTTTATTTTTGGTAT-
TCCAGCAT-3?, which incorporated a transcription
termination codon immediately 3? of a Not I site (both
sites are underlined). Amplification reactions were as
described in Section 2.4 except that the annealing
temperature was 55 8C. The approximately 1.2 kbPCR product, purified from agarose gels, was restriction
digested overnight at 37 8C with Xho I and Not I, re-
purified and ligated into the expression vector pPIC ZaA (Invitrogen Corp., San Diego, CA) that had been
similarly restriction digested and purified. The resulting
construct places the pro-SmCB2 gene downstream and
in frame with the a-mating factor of P. pastoris , thereby
targeting SmCB2 for secretion. Sufficient recombinantplasmid was produced in and purified from E. coli
(DH5a strain; Life Technologies, Inc., Rockville, MD)
for transformation of P. pastoris (see below).
2.6. Expression and purification of recombinant SmCB2
Procedures to express SmCB2 were as detailed by the
manufacturer (Invitrogen). Briefly, to allow for homo-logous incorporation of the recombinant plasmid into
the AOX 1 locus of the P. pastoris genome, the pro-
SmCB2-pPIC Za A plasmid construct (10 mg) was
linearized with Sac I overnight at 37 8C and purified.
Electrocompetent X33 strain of P. pastoris (Invitrogen)
was electroporated at 1.5 kV and 129 ohms in electro-
poration cuvettes (2 mm gap; BTX, San Diego, CA).
Pichia colonies growing at 30 8C under zeocin-selection(100 mg ml�1; Invitrogen) on yeast extract-peptone-
dextrose (YPD)-agar plates were picked for expansion
first in 10 ml and then in 500 ml of YPD medium
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/61 51

containing the antibiotic. Expression of recombinant
protein was induced by incubation of P. pastoris for 24�/
48 h at 30 8C in buffered minimal medium con-
taining 1% methanol as the sole carbon source. En-dopeptidase activity in the medium (1�/5 ml) was
monitored by hydrolysis of Z-Phe-Arg-NMec (see
Section 2.10).
Media (200�/300 ml) from P. pastoris that had been
induced were lyophilized and stored in a desiccator at
4 8C until use. Lyophilized samples were resuspended in
50 mM sodium acetate, pH 4.5 (10% of the original
volume) and complete equilibration with this buffer wasaccomplished using PD10 columns (Amersham Phar-
macia Biotech Inc.). The solution was loaded onto a
Mono S HR 5/5 cation exchange column, which had
been pre-equilibrated in 50 mM sodium acetate, pH 4.5,
and protein resolved by use of a 0�/1 M NaCl gradient.
Endopeptidase activity associated with SmCB2 eluted at
500 mM NaCl. Fractions (1 ml) were stored at �/80 8Cuntil use. Purification was monitored by SDS-PAGEthrough 4�/12% polyacrylamide gels (Invitrogen). After
electrophoresis, gels were stained in 40% methanol, 10%
acetic acid containing 0.5% Coomassie Brilliant Blue R-
250 (BIO-RAD, Hercules, CA) and destained in the
same solution, but omitting Coomassie Blue.
2.7. Production of antiserum to recombinant SmCB2 and
purification of IgG
Antiserum was raised in a New Zealand white rabbit
by Covance, Richmond, CA. Prior to the first injection
of enzyme, pre-immune serum was collected and stored
at �/20 8C. SmCB2 (200 mg) was deglycosylated with
500 U PNGase F for 4 h at 37 8C as described by the
manufacturer (New England Biolabs, Beverly, MA) and
resolved by SDS-PAGE. The Coomassie stained band
was excised and mixed with an approximately equalvolume of adjuvant. Freund’s Complete Adjuvant was
used for the primary injection, whereas Freund’s In-
complete Adjuvant was used for the second and third
injections on days 18 and 36, respectively. The rabbit
was exsanguinated at 54 days and serum stored at
�/20 8C. IgG was isolated from both pre-immune and
hyper-immune sera using Protein G-Sepharose (1 ml) as
described [34] and stored at �/20 8C.
2.8. Immunoblotting
Adult male (100) and female (40) S. mansoni worms
were sonicated over an ice bath in 0.5 ml 50 mM sodium
phosphate buffer, pH 7.0. After centrifugation at 9000 g
for 5 min at 4 8C, the supernatant was either used
immediately or stored at �/70 8C. Worm supernatant(100 mg) or pure recombinant SmCB2 (5 mg) were
resolved by SDS-PAGE (4�/12% gradient gels) and
transferred to PVDF membrane as described above.
Following blocking of the membrane for 2 h at room
temperature in TBS-T (100 mM Tris�/HCl, 100 mM
NaCl, pH 8.0, 0.05% Tween 20), the membrane was
incubated for 1 h with preimmune or anti-SmCB2 IgG
at a dilution of 1:3000 in TBS-T. The membrane was
then washed 5�/5 min in TBS-T and incubated for 1 h
in TBS-T containing alkaline phosphatase-conjugated
goat anti-rabbit IgG diluted 1:4000 (2.5 mg; Life
Technologies). After the same washing steps, blots
were developed in 100 mM Tris�/HCl, 100 mM NaCl,
5 mM MgCl2, pH 9.0 containing 0.33% nitro blue
tetrazolium (NBT; Promega, Madison, WI) and 0.66%
5-bromo-4-chloro-3-indolyl-1-phosphate (BCIP; Pro-
mega). Development was stopped by incubating the
membrane in TBS containing 0.005 M di-sodium
EDTA.
2.9. Immunohistochemistry
Adult S. mansoni worms were washed once in PBS
and fixed in 2% paraformaldehyde/0.1% glutaraldehyde/
PBS for 2�/4 h at room temperature. The schistosome
material was allowed to stand in PBS overnight at 4 8C.
The embedding and immunolabeling procedures have
been previously described [35]. Briefly, parasites were
dehydrated through a graded series of acetone solutions,
embedded in JB-4 (Polyscience, Warrington, PA) and
sectioned at 2 mm with a Sorvall JB-4 microtome.
Sections were placed onto glass slides, air-dried over-
night and incubated in 0.25% trypsin (Zymed, South
San Francisco, CA) at 37 8C for 10 min. Sections were
washed 3�/5 min in wash buffer (PBS containing 0.05%
Tween-20 and 0.1% BSA). Potential endogenous perox-
idase activity was quenched by incubating sections for
10 min at room temperature in 1% H2O2/0.1% NaN3 in
the blocking buffer that is part of the Vectastain Elite
ABC Kit (Vector Labs, Burlingame, CA). Sections were
incubated in blocking buffer alone for a further 30 min
and then incubated overnight at 4 8C with a 1:100
dilution (10 mg) of purified rabbit pre-immune or anti-
SmCB2 IgG in blocking buffer. After the above series of
washes, sections were incubated for 30 min at room
temperature with a 1:200 dilution of biotinylated goat
anti-rabbit IgG (Vectastain Elite ABC Kit). After
washing, sections were incubated for 30 min in the
Vectastain ABC Reagent (horseradish peroxidase H
conjugated to avidin DH), washed again and incubated
for 5 min in AEC (aminoethyl carbazole) Single Solu-
tion (Zymed). Following a further series of washes, the
sections were stained with Gill’s Hematoxylin solution
No. 3 (Sigma), washed in water and air-dried. Sections
were mounted in Aquamount (Lerner Labs, Pittsburgh,
PA).
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/6152
2.10. Endopeptidase activity measured with fluorogenic
substrates: pH optimum and S2 subsite specificity
Routine assays used Z-Phe-Arg-NMec and initialrates of activity were measured at room temperature in
black 96-well microtiter plates using a Labsystems
Fluoroskan II fluorometer at excitation and emission
wavelengths of 350 and 460 nm, respectively. To activate
enzyme activity, either native SmCB1 or SmCB2 (both
0.5 nM), or recombinant SmCB2 (5 nM) was preincu-
bated for 5 min in 100 ml 100 mM sodium citrate, pH
5.5, containing 2 mM DTT. Substrate (100 ml; 40 mM),in the same buffer, was added and the reaction
continued for 5 min. Control reactions contained buffer
in place of enzyme. Activity was expressed either as
nmol NMec released min�1 mg�1 (purification of
native enzymes) or mol NMec released min�1 mol
enzyme�1 (recombinant SmCB2) using a standard curve
of free NMec prepared in ethanol and further diluted in
water. The concentration of active enzyme used inassays was determined by active site titration with E-
64 (Sigma), a stoichiometric irreversible inhibitor of
papain-like cysteine endopeptidases [36].
The pH optimum of native and recombinant SmCB2
was measured with Z-Phe-Arg-NMec, suitable for the
assay of both cathepsins B and L, and Z-Arg-Arg-
NMec, the cathepsin B-selective substrate [27]. A 100
mM citrate�/phosphate buffer over the pH range of 3.0�/
8.0 was used. All buffers contained 300 mM NaCl to
minimize variations in ionic strength. Otherwise assays
were performed as above. To probe the S2 subsite
specificity of SmCB2 (nomenclature of Schechter and
Berger [37]), four substrates varying in the P2 amino acid
(Phe, Leu, Val or Arg) were used (Z-Leu-Arg-NMec and
Z-Val-Arg-NMec were kindly provided by Dr. Dieter
Bromme, Mount Sinai School of Medicine, New York).Given that the choice of substrate can itself influence pH
activity profiles [38], the initial rates of hydrolysis were
measured between pH 4.0 and 7.0.
2.11. Determination of Km and kcat values with NMec
substrates
For determination of the Michaelis constant, Km, a
range of substrate concentrations between 10 and 300mM (solubility limit), and between 50 and 1400 mM was
employed for Z-Phe-Arg-NMec and Z-Arg-Arg-NMec,
respectively. Otherwise, the design of the assays was as
described above. Km and Vmax values were measured by
the direct liner plot method [39]. The catalytic rate
constant kcat was determined from Vmax/e , where e
represents the active enzyme concentration, as measured
by titration with E-64. For comparative purposes underour experimental conditions, kinetic constants for
bovine spleen cathepsin B (Sigma) and rat-liver cathe-
psin L (kindly provided by Dr. Heidrun Kirschke,
Institut fur Physiologische Chemie, Martin-Luther Uni-
versitat, Halle, Germany) were also determined.
3. Results
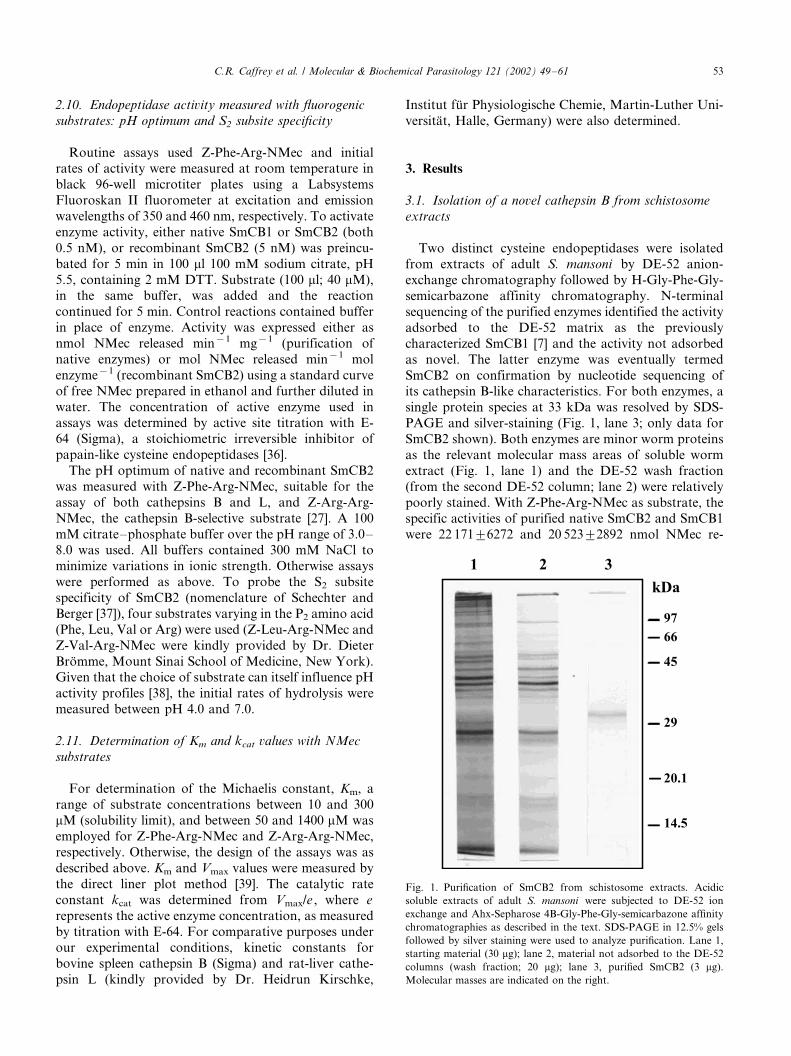
3.1. Isolation of a novel cathepsin B from schistosome
extracts
Two distinct cysteine endopeptidases were isolatedfrom extracts of adult S. mansoni by DE-52 anion-
exchange chromatography followed by H-Gly-Phe-Gly-
semicarbazone affinity chromatography. N-terminal
sequencing of the purified enzymes identified the activity
adsorbed to the DE-52 matrix as the previously
characterized SmCB1 [7] and the activity not adsorbed
as novel. The latter enzyme was eventually termed
SmCB2 on confirmation by nucleotide sequencing ofits cathepsin B-like characteristics. For both enzymes, a
single protein species at 33 kDa was resolved by SDS-
PAGE and silver-staining (Fig. 1, lane 3; only data for
SmCB2 shown). Both enzymes are minor worm proteins
as the relevant molecular mass areas of soluble worm
extract (Fig. 1, lane 1) and the DE-52 wash fraction
(from the second DE-52 column; lane 2) were relatively
poorly stained. With Z-Phe-Arg-NMec as substrate, thespecific activities of purified native SmCB2 and SmCB1
were 22 1719/6272 and 20 5239/2892 nmol NMec re-
Fig. 1. Purification of SmCB2 from schistosome extracts. Acidic
soluble extracts of adult S. mansoni were subjected to DE-52 ion
exchange and Ahx-Sepharose 4B-Gly-Phe-Gly-semicarbazone affinity
chromatographies as described in the text. SDS-PAGE in 12.5% gels
followed by silver staining were used to analyze purification. Lane 1,
starting material (30 mg); lane 2, material not adsorbed to the DE-52
columns (wash fraction; 20 mg); lane 3, purified SmCB2 (3 mg).
Molecular masses are indicated on the right.
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/61 53

leased min�1 mg�1, respectively, compared to the
endopeptidolytic activity of soluble worm extract, which
was 1789/97 nmol NMec released min�1 mg�1 (average
from three purifications). Thus, the fold purification ofSmCB2 and SmCB1 was 142 and 115, respectively. Final
activity yields of both enzymes were between 3 and 8%
of the starting material.
3.2. Confirmation that SmCB2 is a novel schistosome
cathepsin B
The first 13 N-terminal amino acid residues of native
SmCB2 were sequenced (Fig. 2, underlined). Thisfacilitated the design of a semi-nested PCR using
degenerate forward primers (based on the amino acid
sequence with the dashed line in Fig. 2) and a (dT)18
reverse primer to isolate the appropriate DNA sequence
from adult S. mansoni single-stranded cDNA. A 1.0 kb
product was amplified. Sequencing of this product
identified a novel endopeptidase distinct from SmCB1
(see below). 5?-RACE amplified the remaining SmCB2sequence upstream of the N-terminus of the mature
catalytic domain. The ORF of SmCB2 is made up of
1041 nucleotides with a 285 base 3?-UTR and encodes a
pre-pro-protein of 347 amino acids with a calculated
molecular mass of 39 017 Da (Fig. 2). A predicted
hydrophobic leader sequence of 23 amino acids is
upstream of a typical cathepsin B pro-peptide of 70
residues. The 254 amino acid residue catalytic domain ofSmCB2 is 49, 51 and 52% identical, and 57, 59 and 61%
similar to SmCB1, SjCB1 and human cathepsin B,
respectively (Fig. 3). SmCB2 possesses the conserved
active site residues Cys29, His200, and Asn220 (number-
ing system as depicted in Fig. 3) of papain-like cysteine
endopeptidases. In addition, SmCB2 contains all those
residues of the papain superfamily endopeptidases that
interact with the main chain of the bound substrate,namely, Gln23, Gly73, Gly74, and Trp222 [40]. All 12
cysteine residues that form the 6 intramolecular disulfide
bridges within the catalytic domain are conserved
between the schistosome and human enzymes [41].
Characteristic of the cathepsin B group, SmCB2 has
an occluding loop (residues 108 through 119), which is
responsible for dipeptidyl peptidase activity [41]. Two
potential Asn-linked glycosylation sites are present inthe pro-peptide and catalytic domain, at positions p27
and 95, respectively.
3.3. Expression of recombinant active SmCB2 in P.
pastoris
SmCB2 was expressed by P. pastoris as an active
endopeptidase. Yields of purified enzyme were in therange of 40�/60 mg l�1 of culture medium, and on a
weight basis, tests with the stoichiometric irreversible
inhibitor E-64 indicated that the enzyme was 50% active.
By SDS-PAGE, the media from induced P. pastoris
cultures contained a predominant protein of 33 kDa and
a second band at 43 kDa (Fig. 4, lane 1). A single
purification step with Mono S cation exchange chroma-tography was sufficient to isolate SmCB2 as judged by
overloading SDS gels with column fractions containing
the greatest hydrolytic activity against Z-Phe-Arg-NMec
(Fig. 4, lane 2). The purified enzyme migrated with an
apparent mass of 33 kDa, but on treatment with the
glycosidase PNGase F, the molecular mass was approxi-
mately 31 kDa (data not shown), thus indicating that
the recombinant protein is N-glycosylated. N-terminalsequencing of the recombinant enzyme yielded G Y I S
D E, i.e. most of the pro-domain had been removed to
within six amino acids of the N-terminus of the catalytic
domain.
3.4. Identification of SmCB2 in adult schistosome
extracts
By immunoblotting, rabbit anti-SmCB2 IgG reactedwith its recombinant target (Fig. 5, lane 2), whereas
preimmune IgG did not (lane 4). When tested with
parasite extracts of male worms, the antibody reacted
with bands of 33 and 40 kDa (Fig. 5, lane 1). Preimmune
IgG did not react (lane 3). No reaction was visible with
female worm extracts (not shown), and this was in spite
of varying conditions of protein loading and antibody
dilutions.
3.5. Immunolocalization of SmCB2 in adult male and
female S. mansoni
Immunohistochemical studies with anti-SmCB2 IgG
localized the enzyme in tissues of both sexes, but the
reaction was more marked in sections of male worms. In
males, reactivity was detected in the dorsal and lateraltubercles of the tegument; otherwise the tegument was
not stained (Fig. 6A). Labeling was distributed in a
punctate manner throughout the parenchyma below the
muscle layers. Labeling was not observed in the testes
(not shown), gastrodermis or gut lumen. In females, less
labeling was evident compared to males and was found
in the parenchyma below the muscle layers (Fig. 6C).
Also, the reaction was occasionally found in theparenchyma between the vitellaria. No reaction was
detected in the tegument, vitellaria, gastrodermis or in
the gut lumen. For both sexes, no reaction was observed
with pre-immune IgG (Fig. 6B and D).
3.6. Acidic pH optima, S2 subsite specificity and kinetics
of substrate hydrolysis
Both native and recombinant SmCB2 degraded Z-
Phe-Arg-NMec, the cathepsin L/B substrate and Z-Arg-
Arg-NMec, the cathepsin B-selective substrate [27]. For
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/6154
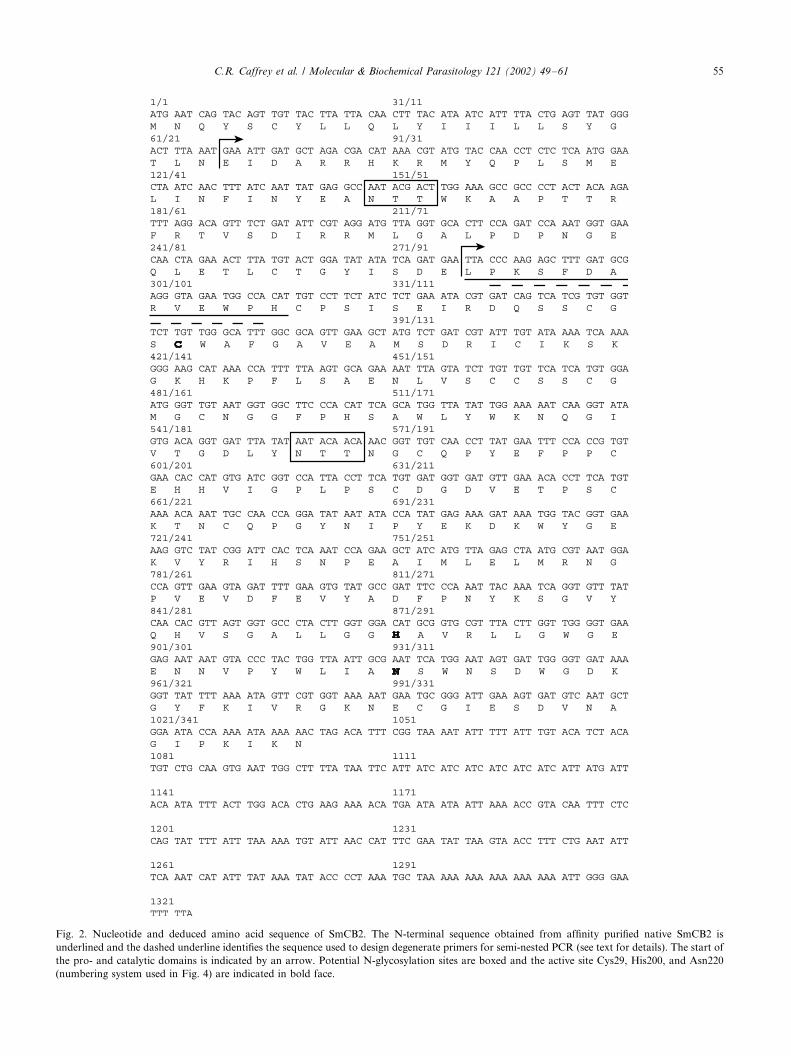
Fig. 2. Nucleotide and deduced amino acid sequence of SmCB2. The N-terminal sequence obtained from affinity purified native SmCB2 is
underlined and the dashed underline identifies the sequence used to design degenerate primers for semi-nested PCR (see text for details). The start of
the pro- and catalytic domains is indicated by an arrow. Potential N-glycosylation sites are boxed and the active site Cys29, His200, and Asn220
(numbering system used in Fig. 4) are indicated in bold face.
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/61 55

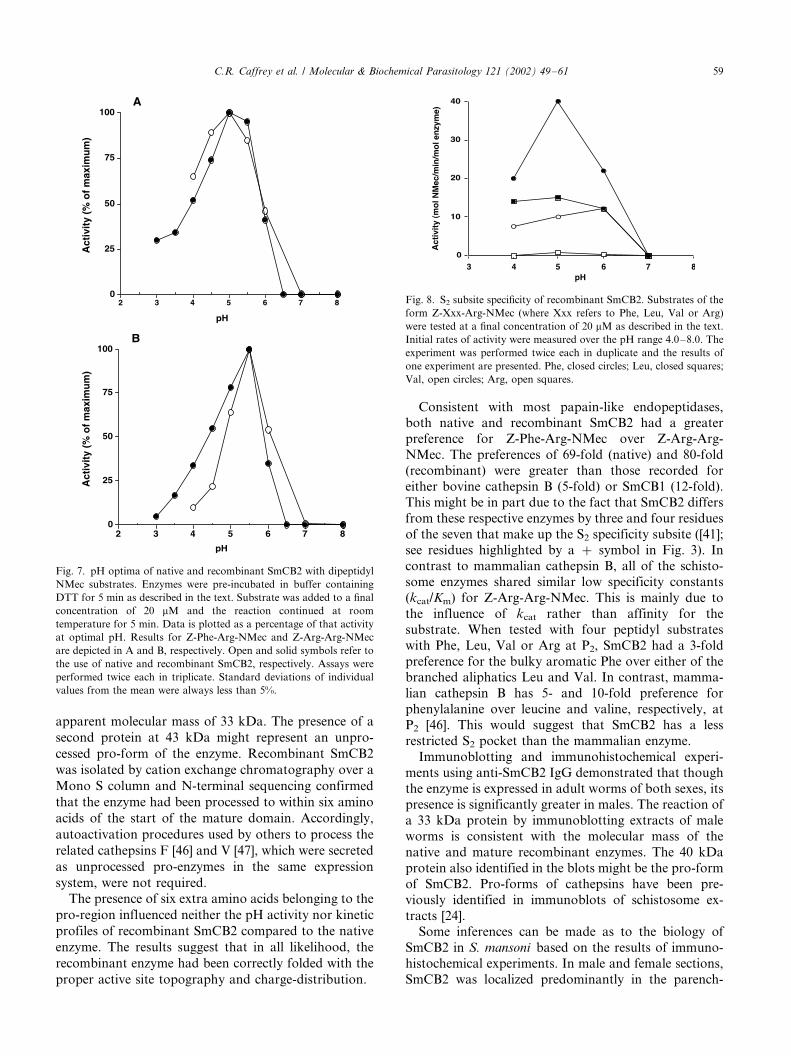
both forms of SmCB2, the pH optima for hydrolysis of
Z-Phe-Arg-NMec (Fig. 7A) and Z-Arg-Arg-NMec (Fig.
7B) were 5.0 and 5.5, respectively. For native SmCB1,
the pH optima for hydrolysis of Z-Phe-Arg-NMec and
Z-Arg-Arg-NMec were 4.5 and 5.5, respectively (not
shown). At pH values at and above 6.5 with either
Fig. 3. Multiple sequence alignment of SmCB2 with other cathepsin B enzymes. Alignments were made with SmCB1 ([7]; accession number
AAA29865), SjCB1 ([30]; P43157) and human cathepsin B ([31]; P07858). Above the sequences, numbers prefixed with a ‘‘p’’ refer to the pro-domain
of the enzyme. Below the sequences, identities across all four sequences are indicated by asterisks, one or more conservative substitutions are
indicated by colons and, where non-conservative changes are present, points are used. The catalytic triad residues (Cys29, His200 and Asn220) are
indicated in bold face. Those residues making up the S2 specificity subsite [41] are indicated with � symbols in bold face.
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/6156

substrate, no hydrolysis was detected for SmCB2 or
SmCB1.
For papain-like endopeptidases, such as SmCB2, the
S2 subsite of the active site cleft defines the primary
specificity towards peptidyl substrates. Four dipeptidyl
NMec substrates, each varying in the P2 amino acid
(Phe, Leu, Val or Arg), were employed to probe the S2
specificity of recombinant SmCB2. At optimal pH, the
substrate with Phe at P2 was degraded 3-fold quicker
than those in which P2 was Leu or Val (Fig. 8). Arg at P2
was least preferred (see also kinetic results below).
Both native and recombinant SmCB2 had similar Km
and kcat values when tested with Z-Phe-Arg-NMec or Z-
Arg-Arg-NMec (Table 1). SmCB2 had a greater speci-
ficity constant (kcat/Km values) for Z-Phe-Arg-NMec
over Z-Arg-Arg-NMec. The preference for the substrate
with Phe at P2 was 69- and 80-fold for native and
recombinant SmCB2, respectively. These values are
significantly greater than those recorded for native
SmCB1 (12-fold) and bovine spleen cathepsin B (5-
fold; Table 1). The kinetic constants measured here for
mammalian cathepsins B and L are consistent with
those obtained by others [27,42].
4. Discussion
We have isolated, functionally expressed and char-
acterized a novel cathepsin B endopeptidase from the
parasitic helminth S. mansoni . The enzyme was tissue-
localized in sections of adult worms. We have termed the
endopeptidase SmCB2, as it is the second cathepsin B ofS. mansoni for which a gene has been identified, the first
being Sm31 [7]. For this report, we have consolidated
the nomenclature regarding schistosome cathepsin Bs,
analogous to that of schistosome cathepsin L enzymes
[14,18,20]. Thus, we now describe Sm31 as SmCB1.
SmCB2 was originally purified from acidic, soluble
worm extracts by a combination of DE-52 anion-
exchange and inhibitor-affinity chromatographies. Thepurification scheme is quicker and involves less handling
compared to previous purifications of schistosome
cathepsins [43�/45]. It is especially convenient as the
Fig. 4. Recombinant expression and purification of active SmCB2.
Concentrated induced culture medium (see text) from P. pastoris
transformed with pro-SmCB2-pPIC Za A was equilibrated with 50
mM sodium acetate buffer, pH 4.5 and loaded onto a Mono S HR 5-5
cation exchange column. Protein was eluted in a 0�/1 M NaCl gradient
and fractions containing highest endopeptidolytic activity (with Z-Phe-
Arg-NMec as substrate) were pooled. A 4�/12% gradient SDS gel
stained with Coomassie Blue is shown. Lane 1, concentrated Pichia
culture medium (10 mg); lane 2, purified SmCB2 pool (70 mg).
Molecular masses are indicated on the right.
Fig. 5. Reaction of monospecific IgG with recombinant SmCB2 and
adult male S. mansoni extracts. Worm supernatant (100 mg; lanes 1 and
3) or pure recombinant SmCB2 (5 mg; lanes 2 and 4) were resolved by
SDS-PAGE (4�/12% gradient gels) and transferred to PVDF mem-
brane. Blots were probed with IgG purified from either a rabbit serum
raised to deglycosylated recombinant SmCB2 (lanes 1 and 2) or pre-
immune serum (lanes 3 and 4). Note the reaction in worm extracts of
protein species of 35 and 43 kDa (lane 1), presumed to represent the
fully processed and zymogen forms of SmCB2, respectively.
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/61 57

same batch of starting material can be used to purify
both SmCB1 and SmCB2. Though sufficient native
enzyme could be purified for limited biochemical
analyses, to perform more extensive analyses, including
crystallography, it is necessary to have a recombinant
source of SmCB2. Accordingly, the information gained
from the N-terminal sequence of purified native SmCB2
facilitated the design of a semi-nested PCR strategy to
amplify, sequence and eventually clone SmCB2 for
expression in P. pastoris .
Sequencing of the ORF of SmCB2 was accomplished
directly from gel-purified PCR products. No ambigu-
ities arose regarding nucleotide identities during the
course of sequencing indicating that the PCR product
contained a single DNA sequence. The sequence of
SmCB2 is clearly related to other cathepsin B enzymes
from both schistosomes and mammals. Most notable is
the presence in SmCB2 of a section encoding the
occluding loop (residues 108�/119) which restricts access
of potential substrates to the S? subsites of the active site
cleft and confers the peptidyl dipeptidase activity unique
to cathepsin B among the cysteine-class cathepsins [41].
Using the entire SmCB2 ORF, a standard nucleotide-
nucleotide BLAST (blastn) analysis limited to the S.
mansoni EST database returned 14 partial but exactly
matching sequences. This would suggest that, thus far,
no isoforms of SmCB2 exist. Also, as the ESTs
identified were derived from both male and female S.
mansoni cDNA libraries, the analysis confirms the
presence by immunohistochemistry of SmCB2 in both
sexes.
The expression and purification of active recombinant
SmCB2 using P. pastoris was straightforward. SmCB2
was the major protein secreted by P. pastoris with an
Fig. 6. Immunolocalization of SmCB2 in adult male and female S. mansoni . Experiments using JB-4-embedded sections of male (A and B) and
female (C and D) worms were performed with IgG purified from a rabbit antiserum to deglycosylated recombinant SmCB2 (A and C) or preimmune
serum (B and D). Secondary antibody was biotinylated goat anti-rabbit IgG which was followed by incubation with horseradish peroxidase H
conjugated to avidin DH. Development of the reaction causes precipitation of the red substrate aminoethyl carbazole at those sites where SmCB2 is
present. Sections were counter-stained with Gill’s Hematoxylin solution No. 3. Reaction product was seen in the parenchyma (p) and tegumental
tubercles (tl) of male worms, and in the parenchyma of females. Note the greater presence of SmCB2 in male worms. g, gastrodermis; gl, gut lumen;
m, muscle; v, vitellaria. Magnification �250.
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/6158
apparent molecular mass of 33 kDa. The presence of a
second protein at 43 kDa might represent an unpro-
cessed pro-form of the enzyme. Recombinant SmCB2
was isolated by cation exchange chromatography over a
Mono S column and N-terminal sequencing confirmed
that the enzyme had been processed to within six amino
acids of the start of the mature domain. Accordingly,
autoactivation procedures used by others to process the
related cathepsins F [46] and V [47], which were secreted
as unprocessed pro-enzymes in the same expression
system, were not required.
The presence of six extra amino acids belonging to the
pro-region influenced neither the pH activity nor kinetic
profiles of recombinant SmCB2 compared to the native
enzyme. The results suggest that in all likelihood, the
recombinant enzyme had been correctly folded with the
proper active site topography and charge-distribution.
Consistent with most papain-like endopeptidases,both native and recombinant SmCB2 had a greater
preference for Z-Phe-Arg-NMec over Z-Arg-Arg-
NMec. The preferences of 69-fold (native) and 80-fold
(recombinant) were greater than those recorded for
either bovine cathepsin B (5-fold) or SmCB1 (12-fold).
This might be in part due to the fact that SmCB2 differs
from these respective enzymes by three and four residues
of the seven that make up the S2 specificity subsite ([41];see residues highlighted by a �/ symbol in Fig. 3). In
contrast to mammalian cathepsin B, all of the schisto-
some enzymes shared similar low specificity constants
(kcat/Km) for Z-Arg-Arg-NMec. This is mainly due to
the influence of kcat rather than affinity for the
substrate. When tested with four peptidyl substrates
with Phe, Leu, Val or Arg at P2, SmCB2 had a 3-fold
preference for the bulky aromatic Phe over either of thebranched aliphatics Leu and Val. In contrast, mamma-
lian cathepsin B has 5- and 10-fold preference for
phenylalanine over leucine and valine, respectively, at
P2 [46]. This would suggest that SmCB2 has a less
restricted S2 pocket than the mammalian enzyme.
Immunoblotting and immunohistochemical experi-
ments using anti-SmCB2 IgG demonstrated that though
the enzyme is expressed in adult worms of both sexes, itspresence is significantly greater in males. The reaction of
a 33 kDa protein by immunoblotting extracts of male
worms is consistent with the molecular mass of the
native and mature recombinant enzymes. The 40 kDa
protein also identified in the blots might be the pro-form
of SmCB2. Pro-forms of cathepsins have been pre-
viously identified in immunoblots of schistosome ex-
tracts [24].Some inferences can be made as to the biology of
SmCB2 in S. mansoni based on the results of immuno-
histochemical experiments. In male and female sections,
SmCB2 was localized predominantly in the parench-
Fig. 7. pH optima of native and recombinant SmCB2 with dipeptidyl
NMec substrates. Enzymes were pre-incubated in buffer containing
DTT for 5 min as described in the text. Substrate was added to a final
concentration of 20 mM and the reaction continued at room
temperature for 5 min. Data is plotted as a percentage of that activity
at optimal pH. Results for Z-Phe-Arg-NMec and Z-Arg-Arg-NMec
are depicted in A and B, respectively. Open and solid symbols refer to
the use of native and recombinant SmCB2, respectively. Assays were
performed twice each in triplicate. Standard deviations of individual
values from the mean were always less than 5%.
Fig. 8. S2 subsite specificity of recombinant SmCB2. Substrates of the
form Z-Xxx-Arg-NMec (where Xxx refers to Phe, Leu, Val or Arg)
were tested at a final concentration of 20 mM as described in the text.
Initial rates of activity were measured over the pH range 4.0�/8.0. The
experiment was performed twice each in duplicate and the results of
one experiment are presented. Phe, closed circles; Leu, closed squares;
Val, open circles; Arg, open squares.
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/61 59

ymous tissues. The antibody reactivity was not evenly
distributed, but was found in discrete areas suggesting a
localization of action. The enzyme may well be present
in the lysosomes or another acidified compartment as
suggested by the strict acidic pH profile of the enzyme
recorded here using dipeptidyl substrates. Of particularnote is the absence of SmCB2 from the gastrodermis and
lumen of the parasite gut. Accordingly, the enzyme is
unlikely to participate directly in the digestion of
ingested nutrients, a function that has been postulated
for the gut-localized endopeptidases SmCB1 [3,7,24],
SmCL1 [14] and SmCL2 [20]. Interestingly, the dorsal
and lateral tubercles of the male were heavily stained,
suggesting that SmCB2 functions at the host�/parasiteinterface. The enzyme may participate in the turnover of
tegumental proteins, the degradation of endocytosed
proteins and/or operate in a protective capacity.
Acknowledgements
Supported by a TDRU grant (AI35707) from the
NIAID and the Sandler Family Foundation.
References
[1] WHO. Schistosomiasis. Fact Sheet no. 115 (http://www.who.int/
inf-fs/en/fact115.html); 1996.
[2] Lim KC, Sun E, Bahgat M, et al. Blockage of skin invasion by
schistosome cercariae by serine protease inhibitors. Am J Trop
Med Hyg 1999;60:487�/92.
[3] Wasilewski MM, Lim KC, Phillips J, McKerrow JH. Cysteine
protease inhibitors block schistosome hemoglobin degradation in
vitro and decrease worm burden and egg production in vivo. Mol
Biochem Parasitol 1996;81:179�/89.
[4] Ruppel A, Shi YE, Wei DX, Diesfeld HJ. Sera of Schistosoma
japonicum -infected patients cross-react with diagnostic 31/32 kD
proteins of S. mansoni . Clin Exp Immunol 1987;69:291�/8.
[5] Li YL, Idris MA, Corachan M, Han JJ, Kirschfink M, Ruppel A.
Circulating antigens in schistosomiasis: detection of 31/32-kDa
proteins in sera from patients infected with Schistosoma japoni-
cum , S. mansoni , S. haematobium , or S. intercalatum . Parasitol
Res 1996;82:14�/8.
[6] Grogan J, Rotmans P, Ghoneim H, Deelder A, Yazdanbakhsh
M, Klinkert M-Q. Recognition of Schistosoma mansoni cathe-
psins B and L by human IgG1 and IgG4 antibodies. Parasite
Immunol 1997;19:215�/20.
[7] Klinkert M-Q, Felleisen R, Link G, Ruppel A, Beck E. Primary
structures of Sm31/32 diagnostic proteins of Schistosoma mansoni
and their identification as proteases. Mol Biochem Parasitol
1989;33:113�/22.
[8] Smith AM, Dalton JP, Clough KA, et al. Adult Schistosoma
mansoni express cathepsin L proteinase activity. Mol Biochem
Parasitol 1994;67:11�/9.
[9] Michel A, Ghoneim H, Resto M, Klinkert M-Q, Kunz W.
Sequence, characterization and localization of a cysteine protei-
nase cathepsin L in Schistosoma mansoni . Mol Biochem Parasitol
1995;73:7�/18.
[10] Hola-Jamriska L, Tort JF, Dalton JP, et al. Cathepsin C from
Schistosoma japonicum -cDNA encoding the preproenzyme
and its phylogenetic relationships. Eur J Biochem
1998;255:527�/34.
[11] Gotz B, Felleisen R, Shaw E, Klinkert M-Q. Expression of an
active cathepsin B-like protein Sm31 from Schistosoma mansoni in
insect cells. Trop Med Parasitol 1992;43:282�/4.
[12] Gotz B, Klinkert M-Q. Expression and characterization of a
cathepsin B-like enzyme (Sm31) and a proposed ‘haemoglobinase’
(Sm32) from Schistosoma mansoni . Biochem J 1993;290:801�/6.
[13] Lipps G, Fullkrug R, Beck E. Cathepsin B of Schistosoma
mansoni . Purification and activation of the recombinant proen-
zyme secreted by Saccharomyces cerevisiae . J Biol Chem
1996;271:1717�/25.
[14] Brady CP, Dowd AJ, Brindley PJ, Ryan T, Day SR, Dalton JP.
Recombinant expression and localization of Schistosoma mansoni
cathepsin L1 support its role in the degradation of host
hemoglobin. Infect Immun 1999;67:368�/74.
[15] Hola-Jamriska L, King LT, Dalton JP, Mann VH, Aaskov JG,
Brindley PJ. Functional expression of dipeptidyl peptidase I
(Cathepsin C) of the oriental blood fluke Schistosoma japonicum
in Trichoplusia ni insect cells. Protein Expr Purif 2000;19:384�/92.
[16] Klinkert M-Q, Cioli D, Shaw E, Turk V, Bode W, Butler R.
Sequence and structure similarities of cathepsin B from the
parasite Schistosoma mansoni . FEBS Lett 1994;351:397�/400.
[17] Butler R, Michel A, Kunz W, Klinkert M-Q. Sequence of
Schistosoma mansoni cathepsin C and its structural comparison
with papain and cathepsins B and L of the parasite. Prot Pept Lett
1995;2:313�/20.
Table 1
Comparison of kinetic values for native and recombinant SmCB2, native SmCB1, bovine-spleen cathepsin B and rat-liver cathepsin L
Enzyme Substrate Difference in kcat/Km values (-fold)
Z-Phe-Arg-NMec Z-Arg-Arg-NMec
Km (mM) kcat (s�1) kcat/Km (mM�1 s�1) Km (mM) kcat (s�1) kcat/Km (mM�1 s�1)
SmCB2 0.056 35 625 1.0 9 9 69
rSmCB2 0.063 70 1111 0.65 9 14 80
SmCB1 0.055 11 196 1.4 23 17 12
Cat Ba 0.25 122 488 1.3 121 93 5
Cat L 0.008 43 5326 n.h.b
a Assays were performed as described in the text, except for bovine cathepsin B (assays at pH 6.0 with Z-Arg-Arg-NMec) and SmCB1 (assays at pH
4.5 with Z-Phe Arg-NMec). For each enzyme, experiments were performed twice, each in duplicate. The results of one experiment for each enzyme
are shown here.b n.h., not hydrolyzed.
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/6160

[18] Brady CP, Brinkworth RI, Dalton JP, Dowd AJ, Verity CK,
Brindley PJ. Molecular modeling and substrate specificity of
discrete cruzipain-like and cathepsin L-like cysteine proteinases of
the human blood fluke Schistosoma mansoni . Arch Biochem
Biophys 2000;380:46�/55.
[19] Ruppel A, Breternitz U, Burger R. Diagnostic Mr 31,000
Schistosoma mansoni proteins: requirement of infection, but not
of immunization, and use of the ‘miniblot’ technique for the
production of monoclonal antibodies. J Helminthol 1987;61:11�/
9.
[20] Bogitsh BJ, Dalton JP, Brady CP, Brindley PJ. Gut-associated
immunolocalization of the Schistosoma mansoni cysteine pro-
teases, SmCL1 and SmCL2. J Parasitol 2001;87:237�/41.
[21] Bogitsh BJ, Dresden MH. Fluorescent histochemistry of acid
proteases in adult Schistosoma mansoni and S. japonicum . J
Parasitol 1983;69:106�/10.
[22] Ruppel A, Shi YE, Moloney NA. Schistosoma mansoni and S.
japonicum : comparison of levels of ultraviolet irradiation for
vaccination of mice with cercariae. Parasitology 1990;101:23�/6.
[23] Rich DH, Brown MA, Barrett AJ. Purification of cathepsin B by
a new form of affinity chromatography. Biochem J 1986;235:731�/
4.
[24] Caffrey CR, Ruppel A. Affinity isolation and characterization of
the cathepsin B-like proteinase Sj31 from Schistosoma japonicum .
J Parasitol 1997;83:1112�/8.
[25] Laemmli UK. Cleavage of structural proteins during the assembly
of the head of bacteriophage T4. Nature 1970;227:680�/5.
[26] Merril CR. Gel-staining techniques. Methods Enzymol
1990;182:477�/88.
[27] Barrett AJ, Kirschke H. Cathepsin B, Cathepsin H, and
Cathepsin L. Methods Enzymol 1981;80:535�/61.
[28] Bradford MM. A rapid and sensitive method for the quantitation
of microgram quantities of protein utilizing the principle of
protein-dye binding. Anal Biochem 1976;72:248�/54.
[29] Nielsen H, Brunak S, von Heijne G. Machine learning approaches
to the prediction of signal peptides and other protein sorting
signals. Prot Eng 1999;12:3�/9.
[30] Merckelbach A, Hasse S, Dell R, Eschlbeck A, Ruppel A. cDNA
sequences of Schistosoma japonicum coding for two cathepsin B-
like proteins and Sj32. Trop Med Parasitol 1994;45:193�/8.
[31] Chan SJ, San Segundo B, McCormick MB, Steiner DF. Nucleo-
tide and predicted amino acid sequences of cloned human and
mouse preprocathepsin B cDNAs. Proc Natl Acad Sci USA
1986;83:7721�/5.
[32] Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: Improv-
ing the sensitivity of progressive multiple sequence alignment
through sequence weighting, positions-specific gap penalties and
weight matrix choice. Nucleic Acids Res 1994;22:4673�/80.
[33] Tao K, Stearns N, Dong J, Wu Q, Sahagian GG. The proregion
of cathepsin L is required for proper folding, stability, and ER
exit. Arch Biochem Biophys 1994;311:19�/27.
[34] Harlow E, Lane D. Using Antibodies. A Laboratory Manual.
New York: Cold Spring Harbor Press, 1999.
[35] Yezzi MJ, Hsieh IE, Caughey GH. Mast cell and neutrophil
expression of dog mast cell protease-3. A novel tryptase-related
serine protease. J Immunol 1994;152:3064�/72.
[36] Barrett AJ, Kembhavi AA, Brown MA, et al. L-trans -Epoxysuc-
cinyl-leucylamido(4-guanidino)butane (E-64) and its analogues as
inhibitors of cysteine proteinases including cathepsins B, H and L.
Biochem J 1982;201:89�/98.
[37] Schechter I, Berger A. On the size of the active site in proteases. I.
Papain. Biochem Biophys Res Commun 1967;27:157�/62.
[38] Khouri HE, Plouffe C, Hasnain S, Hirama T, Storer AC, Menard
R. A model to explain the pH-dependent specificity of cathepsin
B-catalyzed hydrolyses. Biochem J 1991;275:751�/7.
[39] Eisenthal R, Cornish-Bowden A. The direct linear plot. A new
graphical procedure for estimating enzyme kinetic parameters.
Biochem J 1974;139:715�/20.
[40] Turk B, Turk D, Turk V. Lysosomal cysteine proteases: more
than scavengers. Biochim Biophys Acta 2000;1477:98�/111.
[41] Musil D, Zucic D, Turk D, et al. The refined 2.15 A X-ray crystal
structure of human liver cathepsin B: the structural basis for its
specificity. EMBO J 1991;10:2321�/30.
[42] Hasnain S, Hirama T, Tam A, Mort JS. Characterization of
recombinant rat cathepsin B and nonglycosylated mutants
expressed in yeast. New insights into the pH dependence of
cathepsin B-catalyzed hydrolyses. J Biol Chem 1992;267:4713�/21.
[43] Chappell CL, Dresden MH. Purification of cysteine proteinases
from adult Schistosoma mansoni . Arch Biochem Biophys
1987;256:560�/8.
[44] Lindquist RN, Senft AW, Petitt M, McKerrow J. Schistosoma
mansoni : purification and characterization of the major
acidic proteinase from adult worms. Exp Parasitol 1986;61:398�/
404.
[45] Ghoneim H, Klinkert MQ. Biochemical properties of purified
cathepsin B from Schistosoma mansoni . Int J Parasitol
1995;25:1515�/9.
[46] Wang B, Shi GP, Yao PM, Li Z, Chapman HA, Bromme D.
Human cathepsin F. Molecular cloning, functional expression,
tissue localization, and enzymatic characterization. J Biol Chem
1998;273:32000�/8.
[47] Bromme D, Li Z, Barnes M, Mehler E. Human cathepsin V.
Functional expression, tissue localization, electrostatic surface
potential, enzymatic characterization, and chromosomal localiza-
tion. Biochemistry 1999;38:2377�/85.
C.R. Caffrey et al. / Molecular & Biochemical Parasitology 121 (2002) 49�/61 61




















