
Sm21.6 a novel EF-hand family protein member located on the surface of Schistosoma mansoni adult...
9
Vaccine 27 (2009) 4127–4135 Contents lists available at ScienceDirect Vaccine journal homepage: www.elsevier.com/locate/vaccine Sm21.6 a novel EF-hand family protein member located on the surface of Schistosoma mansoni adult worm that failed to induce protection against challenge infection but reduced liver pathology Debora O. Lopes a , Leonardo F. Paiva b , Mauricio A. Martins a , Fernanda C. Cardoso a , Matheus A. Rajão a , Jean M. Pinho a , Marcelo V. Caliari c , Rodrigo Correa-Oliveira d , Samantha M. Mello b , Luciana. C.C. Leite b , Sergio C. Oliveira a,∗ a Departamento de Bioquímica e Imunologia, Universidade Federal de Minas Gerais, Avenida Antonio Carlos 6627, Pampulha, Belo Horizonte 31270-901, MG, Brazil b Centro de Biotecnologia, Instituto Butantan, Av. Vital Brasil 1500, 05503-900 São Paulo, SP, Brazil c Departmento de Patologia, Universidade Federal de Minas Gerais, Avenida Antonio Carlos 6627, Pampulha, Belo Horizonte 31270-901, MG, Brazil d Centro de Pesquisas Rene Rachou, Av. Augusto de Lima 1715, Belo Horizonte, MG 30190-002, Brazil article info Article history: Received 2 December 2008 Received in revised form 26 March 2009 Accepted 21 April 2009 Available online 14 May 2009 Keywords: Schistosoma mansoni Vaccine Sm21.6 Cytokines Recombinant protein abstract Schistosomiasis continues to be a significant public health problem that affects 200 million people world- wide. This is one of the most important parasitic diseases, and one whose effective control is unlikely in the absence of a vaccine. In this study, we have isolated a cDNA clone encoding the Schistosoma mansoni Sm21.6 protein that has 45% and 44% identity with Sm22.6 and Sj21.7 EF-hand containing antigens, respec- tively. Confocal microscopy analysis revealed that Sm21.6 is a membrane-associated protein localized on the S. mansoni adult worm. Mouse immunization with rSm21.6 induced a mixed Th1/Th2 cytokine profile and no protection against infection. However, vaccination with rSm21.6 reduced by 28% of liver granu- loma numbers, 21% of granuloma area and 34% of fibrosis. Finally, rSm21.6 was recognized by sera from individuals resistant to reinfection compared with patients susceptible to reinfection and this molecule should be further studied as potential biomarker for disease resistance. In conclusion, Sm21.6 is a new tegument protein from S. mansoni that plays an important role in reducing pathology induced by parasite infection. © 2009 Elsevier Ltd. All rights reserved. 1. Introduction Human schistosomiasis is a chronic debilitating parasitic dis- ease that represents a major health problem in endemic areas, in various parts of South America, Africa, and Southeast Asia [1]. Schistosomiasis mainly occurs in developing countries and is the most important human helminth infection in terms of global mor- tality. This parasitic disease affects more than 200 million people worldwide causing more than 250,000 deaths per year [2]. Fur- thermore, schistosomiasis causes up to 4.5 million DALY (disability adjusted life year) losses annually [3]. Current schistosomiasis con- trol strategies are mainly based on chemotherapy but in spite of decades of mass treatment, the number of infected people remains constant [4]. Extensive endemic areas and constant reinfection of individuals together with poor sanitary conditions in developing countries make drug treatment alone inefficient [5]. Many con- sider that the best long-term strategy to control schistosomiasis is ∗ Corresponding author. Fax: +55 31 34092666. E-mail address: [email protected] (S.C. Oliveira). through immunization with an anti-schistosomiasis vaccine com- bined with drug treatment [6]. A vaccine that induces even a partial reduction in worm burdens could considerably reduce pathology and limit parasite transmission [1]. In this study, we have identified the membrane protein Sm21.6 that is one member of the family of EF-hand containing para- site proteins with sequence similarity to dynein light chain (DLC), such as Sm22.6 and Sm21.7 [7]. Sm22.6 is a protein considered to be a target for the human IgE response, and it correlates with resistance to reinfection after treatment [8]. Regarding Sm21.7, it was isolated from a cDNA expression library using irradiated cercariae-vaccinated rabbit serum. This antigen was designated “vaccine dominant” because parasite-derived Sm21.7 was recog- nized preferentially by sera from vaccinated mice compared with sera from infected mice [9]. Regarding localization, both antigens were detected at the tegument surface by proteomic analysis [10]. Schistosomes have a unique outer syncytial surface, the tegument that constitutes the host–parasite interface and therefore repre- sents an obvious tissue to target for development of new control strategies [11]. Indeed, so far the two surface exposed membrane proteins identified on biotinylated adult worms, Sm29 and TSP-2, 0264-410X/$ – see front matter © 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.vaccine.2009.04.068
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Sm21.6 a novel EF-hand family protein member located on the surface of Schistosoma mansoni adult...

SSc
DJSa
b
c
d
a
ARRAA
KSVSCR
1
eiSmtwtatdcics
0d
Vaccine 27 (2009) 4127–4135
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
m21.6 a novel EF-hand family protein member located on the surface ofchistosoma mansoni adult worm that failed to induce protection againsthallenge infection but reduced liver pathology
ebora O. Lopesa , Leonardo F. Paivab , Mauricio A. Martinsa , Fernanda C. Cardosoa , Matheus A. Rajãoa ,ean M. Pinhoa, Marcelo V. Caliari c, Rodrigo Correa-Oliveirad, Samantha M. Mellob, Luciana. C.C. Leiteb,ergio C. Oliveiraa,∗
Departamento de Bioquímica e Imunologia, Universidade Federal de Minas Gerais, Avenida Antonio Carlos 6627, Pampulha, Belo Horizonte 31270-901, MG, BrazilCentro de Biotecnologia, Instituto Butantan, Av. Vital Brasil 1500, 05503-900 São Paulo, SP, BrazilDepartmento de Patologia, Universidade Federal de Minas Gerais, Avenida Antonio Carlos 6627, Pampulha, Belo Horizonte 31270-901, MG, BrazilCentro de Pesquisas Rene Rachou, Av. Augusto de Lima 1715, Belo Horizonte, MG 30190-002, Brazil
r t i c l e i n f o
rticle history:eceived 2 December 2008eceived in revised form 26 March 2009ccepted 21 April 2009vailable online 14 May 2009
a b s t r a c t
Schistosomiasis continues to be a significant public health problem that affects 200 million people world-wide. This is one of the most important parasitic diseases, and one whose effective control is unlikely inthe absence of a vaccine. In this study, we have isolated a cDNA clone encoding the Schistosoma mansoniSm21.6 protein that has 45% and 44% identity with Sm22.6 and Sj21.7 EF-hand containing antigens, respec-tively. Confocal microscopy analysis revealed that Sm21.6 is a membrane-associated protein localized on
eywords:chistosoma mansoniaccinem21.6ytokinesecombinant protein
the S. mansoni adult worm. Mouse immunization with rSm21.6 induced a mixed Th1/Th2 cytokine profileand no protection against infection. However, vaccination with rSm21.6 reduced by 28% of liver granu-loma numbers, 21% of granuloma area and 34% of fibrosis. Finally, rSm21.6 was recognized by sera fromindividuals resistant to reinfection compared with patients susceptible to reinfection and this moleculeshould be further studied as potential biomarker for disease resistance. In conclusion, Sm21.6 is a newtegument protein from S. mansoni that plays an important role in reducing pathology induced by parasite
infection.. Introduction
Human schistosomiasis is a chronic debilitating parasitic dis-ase that represents a major health problem in endemic areas,n various parts of South America, Africa, and Southeast Asia [1].chistosomiasis mainly occurs in developing countries and is theost important human helminth infection in terms of global mor-
ality. This parasitic disease affects more than 200 million peopleorldwide causing more than 250,000 deaths per year [2]. Fur-
hermore, schistosomiasis causes up to 4.5 million DALY (disabilitydjusted life year) losses annually [3]. Current schistosomiasis con-rol strategies are mainly based on chemotherapy but in spite ofecades of mass treatment, the number of infected people remains
onstant [4]. Extensive endemic areas and constant reinfection ofndividuals together with poor sanitary conditions in developingountries make drug treatment alone inefficient [5]. Many con-ider that the best long-term strategy to control schistosomiasis is∗ Corresponding author. Fax: +55 31 34092666.E-mail address: [email protected] (S.C. Oliveira).
264-410X/$ – see front matter © 2009 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2009.04.068
© 2009 Elsevier Ltd. All rights reserved.
through immunization with an anti-schistosomiasis vaccine com-bined with drug treatment [6]. A vaccine that induces even a partialreduction in worm burdens could considerably reduce pathologyand limit parasite transmission [1].
In this study, we have identified the membrane protein Sm21.6that is one member of the family of EF-hand containing para-site proteins with sequence similarity to dynein light chain (DLC),such as Sm22.6 and Sm21.7 [7]. Sm22.6 is a protein consideredto be a target for the human IgE response, and it correlates withresistance to reinfection after treatment [8]. Regarding Sm21.7,it was isolated from a cDNA expression library using irradiatedcercariae-vaccinated rabbit serum. This antigen was designated“vaccine dominant” because parasite-derived Sm21.7 was recog-nized preferentially by sera from vaccinated mice compared withsera from infected mice [9]. Regarding localization, both antigenswere detected at the tegument surface by proteomic analysis [10].
Schistosomes have a unique outer syncytial surface, the tegumentthat constitutes the host–parasite interface and therefore repre-sents an obvious tissue to target for development of new controlstrategies [11]. Indeed, so far the two surface exposed membraneproteins identified on biotinylated adult worms, Sm29 and TSP-2,
4 cine 2
hmv
StttSnpvrfme
2
2
Fwiamm(ibt
2
esvsopiogccpTpmaTagntticooi
128 D.O. Lopes et al. / Vac
ave been tested as vaccines and provided protection in murineodels and are selectively recognized by naturally resistant indi-
iduals [12,13].In this present study, we investigate the vaccine potential of
m21.6 protein, a new EF-hand family protein member identified byranscriptome approach. Herein, we detected Sm21.6 transcripts inhe cDNA libraries of worm, cercariae, egg and lung-stage of Schis-osoma mansoni. Furthermore, immunolocalization revealed thatm21.6 is present on the tegument of male adult worm. Next, immu-ization of mice with rSm21.6 induced a mixed Th1/Th2 cytokinerofile but fails to engender protective immunity. However, rSm21.6accination reduced liver pathology in vaccinated mice. Finally,Sm21.6 was recognized by sera from individuals resistant to rein-ection compared with patients susceptible to reinfection and this
olecule should be further studied as potential biomarker for dis-ase resistance.
. Materials and methods
.1. Mice and parasites
Female C57BL/6 mice aged 6–8 weeks were purchased from theederal University of Minas Gerais (UFMG) animal facility. The miceere treated with anti-parasitic drugs 14 days before the exper-
ments start as previously described [14]. All protocols involvingnimals were approved by the local Ethics Committee on Ani-al Care (CETEA–UFMG no. 023/05). Cercariae of S. mansoni wasaintained routinely on Biomphalaria glabrata snails at CPqRR
Centro de Pesquisa Rene Rachou-Fiocuz) and prepared by exposingnfected snails to light for 1 h to induce shedding. Cercarial num-ers and viability were determined using a light microscope prioro infection.
.2. Study population
Peripheral blood was obtained from individuals with differ-nt genetic background living in two different endemic areas forchistosomiasis (“Melquiades” and “Côrrego do Onca”). These indi-iduals were classified in four groups, regarding their infectiontatus and the selection of subjects was performed based onlyn the criteria for inclusion and exclusion of each group inde-endent of previous knowledge of immune responses for each
ndividual. Non-infected (NI) individuals are healthy people with-ut any parasite infection or contaminated water contact. Anotherroup showed stool-negative examination after treatment and waslassified as resistant to S. mansoni reinfection (RR). Individualslassified as susceptible to S. mansoni reinfection (SR) showed stool-ositive examination following treatment (praziquantel, 40 mg/kg).he sera from RR and SR groups were obtained 6 months afterraziquantel treatment and these individuals were examined for S.ansoni infection using the Kato–Katz technique before treatment
nd 1, 6 and 12 months after treatment to check for reinfection rates.he water contact exposure was objectively evaluated by observersnd SR and RR groups had at least one contact daily. Individualsrouped as infected (INF) showed stool-positive examination ando treatment history. These infected patients had infection levelshat varied from 48 to 224 epg (egg counts per gram of faeces). Addi-ionally, RR, SR and INF patients were negative for other helminthic
nfections. These patients or their legal guardians gave informedonsent after explanation of the protocol that had been previ-usly approved by the Ethical Committee of the Federal Universityf Minas Gerais. Details regarding sex and age of the individualsncluded in this study are described in Table 1.7 (2009) 4127–4135
2.3. Isolation of Sm21.6 cDNA
The Sm21.6 DNA sequence was obtained from São Paulo S.mansoni transcriptome project by searching homologue proteinsto Sm21.6 [15]. The sequence obtained was compared with thosealready deposited in the GenBank database using the Basic LocalAlignment Search Tool (BLAST) algorithm available at the NCBIwebsite (http://www.ncbi.nlm.nih.gov/BLAST/). The amino acidsequence of Sm21.6 was obtained and the search for conserveddomains was performed using Pfam program. After that, Syfpeithialgorithm was used in the prediction of human MHC II binding epi-topes (http://www.syfpeithi.de/Scripts/MHCServer.dll). Primarystructure analysis and post-translational modification predictionof Sm21.6 were performed using Expasy (Expert Protein AnalysisSystem, http://ca.expasy.org/tools) computer program. Molecularweight (MW) and isoelectric point (pI) were calculated with theCompute pI/Mw tool (http://www.expasy.ch/tools/pi tool.html).The presence of signal peptide (SignalP 3.0, http://www.cbs.dtu.dk/services/SignalP), glycosylation sites (YinOYang, http://www.cbs.dtu.dk/services/YinOYang) and transmembrane proteintopology prediction (TMHMM, http://www.cbs.dtu.dk/services/TMHMM-2.0) was also performed by bioinformatics.
2.4. Sm21.6 cloning and sequencing
Specific oligonucleotides were designed using the S. man-soni EST assembly contig (SmAE 605436.1, http://bioinfo.iq.usp.br/schisto6/cgi-bin/ceditor/ceditor.pl). The 5′ and 3′ oligonu-cleotides, CAC CAT GTT GGA AGA ATT TAT CAA AG and TTA ACCTTT TGG TGT ACG C, were used to amplify the complete openreading frame of Sm21.6 from S. mansoni cDNA clone MA3-9999U-M275-F06 (GenBank accession CD079525). The PCR reaction wasperformed using Platinum Pfx enzyme (Invitrogen, São Paulo,Brazil), and initiated with one cycle of 5 min at 94 ◦C, followed by30 cycles of 30 s at 94 ◦C, 1 min at 55 ◦C, and 3 min at 68 ◦C. PCRproducts were purified from agarose gel electrophoresis and clonedinto pENTR/D TOPO cloning vector (Invitrogen) and sequenced toconfirm its identity.
2.5. Sm21.6 transcript analysis in parasite life-stage cDNAlibraries
In order to identify Sm21.6 mRNA transcripts in the cDNAlibraries from specific human developmental stages of S. mansoni,PCR was performed using Sm21.6 specific primers. Adult worm,lung-stage, egg and cercariae cDNA libraries were gifted by theMinas Gerais Genome Network. PCR parameters using 1 �L (100 ng)of each library as template were as follows: denaturation for 30 sat 94 ◦C, annealing for 30 s at 57 ◦C and extension for 1 min at 72 ◦C(35 cycles). Agarose gel (1%) eletrophoresis and ethidium bromidestaining were performed to visualize the Sm21.6 sequence amplifiedfrom each library.
2.6. Expression and purification of recombinant Sm21.6
To produce a recombinant (r) Sm21.6, the full-length DNAsequence was directionally cloned by recombination into pDEST17(to produce a protein that contains an N-terminal hexahistidinetag). For protein expression, transformed BL21(DE3) cells (Invit-rogen) were grown in 600 mL LB plus ampicillin (1 mg/mL) untilOD600 = 0.6. Isopropyl-�-d-thiogalactopyranoside (IPTG, Invitro-gen) was added to the culture to a final concentration of 1 mM,
and cells were incubated for 3–4 h at 37 ◦C. Cells were harvestedby centrifugation and resuspended in 50 mL of lysis buffer (10 mMNaHPO4 pH 7.5, 150 mM NaCl, 1× Protease Inhibitor Cocktail (PIC)(Sigma, St. Louis, MO)). The cell suspension was passed three times(1500 psi) through a French press and the crude homogenate was
D.O. Lopes et al. / Vaccine 27 (2009) 4127–4135 4129
Table 1Study population.
Group Infection status Description Age (mean ± S.D.) Sex (M/F)
Infected (I) n = 7 Infected with S.mansoni
Individuals living in an endemic area for schistomiasiswith stool-positive examinations
17.1 ± 14.9 5/2
Susceptible toreinfection (SR) n = 9
Infected with S.mansoni
Individuals living in a endemic area for schistosomiasisthat present stool-positive examination afterpraziquantel treatment
22.8 ± 12.1 3/6
Resistant to reinfection(RR) n = 9
Not infected Individuals living in a endemic area for schistosomiasisthat although water contact, present stool-negative
fter pr
18.8 ± 9.3 5/4
N out ois that
ctbfiubtpb–bbtteNvrFotu
2m
eawiSSauPfiawiu
2
av(obwP
examination aon-infected (NI) n = 8 Not infected Individuals living
schistosomiastheir lives
entrifuged at 20,000 × g for 30 min at 4 ◦C. For protein purification,he inclusion bodies of rSm21.6 were rinsed twice with washinguffer (10 mM NaHPO4 pH 7.5, 500 mM NaCl, 1 M Urea, 1× PIC), andnally resuspended in solubilization buffer (wash buffer plus 8 Mrea, 10 mM imidazole). The recombinant protein was then purifiedy metal affinity chromatography after on-column refolding usinghe Akta Prime system (GE Healthcare, São Paulo, Brazil). Briefly, therotein concentration was adjusted to 0.5 �g/�L with solubilizationuffer and the sample loaded onto a Ni2+-NTA column (HisTrap HPGE Healthcare, 5 mL bed volume) pre-equilibrated with the sameuffer. The column was washed with 12 bed volumes of the equili-ration buffer until the UV absorbance baseline was reached. Then,he refolding of the bound protein was performed on-column byhe use of a linear urea gradient from 8 to 0 M, starting with thequilibration buffer and finishing with a buffer containing 10 mMaHPO4, pH 7.5, 500 mM NaCl, 10 mM imidazole. The total gradientolume was 25 mL and the flow rate was 1.0 mL/min. The refoldedSm21.6 was eluted with 10–500 mM imidazole linear gradient.ractions encompassing the main peak were pooled and the purityf the preparation assessed by SDS-PAGE. The buffer was exchangedo PBS (pH 7.4) using a desalting column (GE Healthcare) before these of this antigen.
.7. Immunolocalization of Sm21.6 in male adult worm of S.ansoni
Adult worms used in confocal microscopy studies were recov-red from perfused mice, as previously described [12]. For sectionsssays, 7 �m slices were deparaffinized with xylol series. Parasitesere blocked with 1% BSA in PBST (Tween 20, 0.05%) for 1 h and
ncubated with anti-rSm21.6 serum diluted 1:20 in blocking buffer.erum from non-immunized mice was used as a negative control.amples were washed three times with PBST and incubated withnti-mouse IgG antibody conjugated to Alexa Fluor 594 (Molec-lar Probes, CA, USA) diluted 1:100 in blocking buffer containinghalloidin Alexa Fluor 488 (Molecular Probes) to stain actin micro-laments. The samples were washed four times and mounted inntifade reagent (Prolong gold–Molecular Probes). The parasitesere visualized in a Zeiss 510 Meta confocal microscope using an
mmersion objective. All the parameters and microscope settingssed were maintained throughout the process.
.8. Measurement of specific anti-Sm21.6 antibodies
ELISA was performed to confirm the titer of specific IgG, IgG1nd IgG2a anti-Sm21.6 in the serum of immunized animals as pre-iously described [16]. Briefly, 96 well flat-bottom microtiter plates
Nunc, Rochester, NY) were coated overnight at 4 ◦C with 100 �Lf rSm21.6 at a concentration of 5 �g/mL in a 0.1 M carbonateicarbonate buffer (pH 9.6) per well. The plate was then washedith 0.05% Tween 20 and blocked with bovine fetal serum 10% inBS for 2 h at room temperature. Further, the plates were washed
aziquantel treatmentf endemic area fornever present this disease in
28.3 ± 4.5 3/6
three times with PBS plus 0.05% Tween 20 (PBST). One hundredmicroliters of each serum diluted 1:100 in PBST were added perwell and incubated for 1 h at room temperature. Plate-bound anti-body was detected by peroxidase-conjugated anti-mouse IgG, IgG1and IgG2a (Southern Biotechnology, Birmingham, AL) diluted inPBST 1:10000, 1:5000 and 1:2000, respectively. After 1 h at 37 ◦C,the plate was washed and orthophenylenediamine (Sigma) plus0.05% hydrogen peroxide in phosphate citrate buffer (pH 5) wasadded (100 �L/well). This mixture was then incubated for 30 minat room temperature, and the reaction was stopped by addictionof 2 N H2SO4 (50 �L/well). Absorbance was read at 492 nm usinga microplate reader (Bio-Rad, Hercules, CA, USA). Animals thatreceived PBS were used as negative control.
2.9. Cytokine analysis
Cytokine experiments were performed using splenocyte cul-tures from individual mice immunized with rSm21.6 plus CFA/IFA(n = 5 for each group). Splenocytes were isolated from maceratedspleens of individual mice 10 days after the third immunizationand washed twice with sterile PBS. After washing, the cells wereadjusted to 1 × 106 cells per well for IL-4, IL-10, IFN-� and TNF-�assays in RPMI 1640 medium (Gibco, CA, USA) supplemented with10% FBS, 100 U/mL penicillin G sodium, 100 �g/mL streptomycinsulfate, 250 ng/mL amphotericin B. Splenocytes were maintained inculture with medium alone or stimulated with rSm21.6 (25 �g/mL)or concanavalin A (ConA) (5 �g/mL) as previously described [16].The 96-well plates (Nunc) were maintained in an incubator at37 ◦C with 5% CO2. For cytokine assays polymyxin B (30 �g/mL)was added to the cultures and this treatment completely abrogatethe cytokine response to LPS as previously described [17]. Culturesupernatants were collected after 24 h of ConA stimulation, 48 hof rSm21.6 stimulation for IL-4 and TNF-� analysis and 72 h ofrSm21.6 stimulation for IL-10 and IFN-�. The assays for measure-ment of IL-4, IL-10, IFN-� and TNF-� were performed using theDuoset ELISA kit (R&D Diagnostic, Minneapolis, MN) according tothe manufacturer’s directions.
2.10. Challenge infection and worm burden recovery
Fifteen days after the last boost, mice were challenged throughpercutaneous exposure of abdomen skin for 1 h in water con-taining 100 cercariae (LE strain). Forty-five days after challenge,adult worms were perfused by portal veins [18]. Two independentexperiments were performed to determine protection levels. Theprotection was calculated comparing the number of worm recov-
ered from each group of immunization with its respective controls,using the formula:PL = WRCG − WREGWRCG
× 100

4 cine 2
wg
2
mcerwpGZipqcobgiwsJwtpw�
2
B[aotpbrt1pa1aimtAH
2
u
3
3
S
130 D.O. Lopes et al. / Vac
here PL = protection level, WRCG = worms recovered from controlroup, and WREG = worms recovered from experimental group.
.11. Hepatic granuloma analysis
Liver sections from mice (10/group) of control and experi-ental group immunized with rSm22.4 and infected with 100
ercariae were collected 45 days post-infection to evaluate theffect of immunization in granuloma formation. The livers sectionsemoved from the central part of the left lateral lobe were fixedith 10% buffered formaldehyde in PBS. Histological sections wereerformed using microtome (4 mm) and stained in a slide withomory’s trichromic. The granulomas were counted in Axiolab Carleiss microscopy using 10× objective lens. All slides were digital-zed by scanner HP scanjet 2400, in the resolution of 300 dpi. Theixels of each histological section were fully screened, with subse-uent creation of a binary image and calculating the total area of theut. The area of the lower cutoff was used as a minimum standardf tissue to be statistically analyzed. The results were expressedy the number of granuloma per area of liver (mm2). The area ofranulomas was obtained through the KS300 software containedn Carl Zeiss image analyzer. Fifteen granulomas from each mouse
ith a single well-defined egg were randomly chosen at a micro-cope with 10× objective lens and scanned through microcameraVC TK-1270/RGB. Using a digital pad, the total area of granulomasas measured and the results were expressed in square microme-
ers (�m2). To calculate the area of fibrosis in the granulomas, theixels of the collagen areas were selected through the real imageith subsequent creation of a binary image and obtain the area inm2.
.12. Measurement of human IgG anti-Sm21.6
Sera of schistosomiasis patients living in endemic areas inrazil were tested by ELISA as previously described by our group19] to measure the levels of IgG to rSm21.6 antigen. For thisssay, 96-well flat-bottom microtiter plates (Nunc) were coatedvernight at 4 ◦C with 100 �l of rSm21.6–6×HIS at a concentra-ion of 5 �g/mL in 0.1 M carbonate bicarbonate buffer (pH 9.6)er well. The plates were then washed with 0.05% Tween 20 andlocked with 10% bovine fetal serum in PBS (pH 7.4) for 2 h atoom temperature. Subsequently, the plates were washed threeimes with PBS plus 0.05% Tween 20 (PBST). Serum samples diluted:50 (IgG) in PBST (100 �L/well) were added in duplicate and thelates incubated for 1 h at room temperature. Peroxidase-labellednti-human IgG (Sigma, St Louis, MO) was added at dilutions of:10,000 (100 �L/well). After 1 h at 37 ◦C, the plates were washednd orthophenylenediamine (Sigma) plus 0.05% hydrogen peroxiden phosphate citrate buffer (pH 5) was added (100 �L/well). This
ixture was than incubated for 30 min at room temperature, andhe reaction was stopped by addition of 2 N H2SO4 (50 �L/well).bsorbance was read at 492 nm using a microplate reader (Bio-Rad,ercules. CA, USA).
.13. Statistical analysis
Statistical analysis was performed with Student’s t-test andsing a computer software package GraphPad Prism(La Jolla, CA).
. Results
.1. Cloning and molecular characterization of S. mansoni Sm21.6
The full-length sequence of the S. mansoni cDNA encodingm21.6 was obtained by PCR amplification from a S. mansoni tran-
7 (2009) 4127–4135
scriptome cDNA clone with specific oligonucleotides that weredesigned using the corresponding EST contig (SmAE 605436.1).The resulting full-length cDNA displayed an ORF of 552 bp, encod-ing a protein of 183 amino acids with a predicted molecular massof approximately 21.6 kDa and an isoelectric point of 5.18. BlastPcomparisons of the deduced S. mansoni protein sequence to Gen-Bank showed that the best match (E-value = 1 × 10−80) was to anunknown S. japonicum protein (SJCHGC01853), with 77% identityand 86% similarity, probably the Sj21.6 ortholog. This was followedby S. mansoni tegument antigen Sm22.6 (AAA29856) and the S.japonicum 21.7 kDa antigen (AAD13338) with 45% and 44% identity,respectively.
3.2. Bioinformatic analysis of Sm21.6
Bioinformatic analysis of Sm21.6 primary sequence showed thatthis protein did not possess a signal peptide and transmembranedomain and it was predicted as a soluble protein. Three glycosy-lation sites are presented in the mature sequence on the Thr83,Thr85 and Thr126 (data not show). The analysis performed revealedthat all aligned proteins shown in Fig. 1, have an EF-hand motifpresent in several antigens associated with the tegument. Sm22.6and Sj21.7 have two EF-hand motifs while Sm21.6 and the S. japon-icum unknown protein has only one (Fig. 1). Moreover, we detectedthe presence of a dynein domain in Sm21.6 as observed in the otherproteins aligned. Additionally, prediction of human MHC class IIepitopes revealed that Sm21.6 possess four promiscuous peptidesthat have high binding score for HLA-DRB1 0101, HLA-DRB1 0301,HLA-DRB1 0401, HLA-DRB1 0701, HLA-DRB1 1101 and HLA-DRB11501 alleles (data not show). The profile of predicted HLA peptidesis more similar between Sm21.6 and the S. japonicum unknownprotein when compared Sm21.6 with Sm22.6 or Sj21.7 (Fig. 1). TheHLA epitopes are more concentrated at the center to the C-terminalregion of Sm21.6.
3.3. Sm21.6 transcripts are detected in cDNA libraries of allparasite stages
Sm21.6 transcripts with 552 bp were detected in all stages of S.mansoni life cycle tested: adult worm, cercariae, egg and lung-stagecDNA libraries (Fig. 2). Since the lungs seem to be the major site ofparasite elimination in the irradiated cercariae model of vaccina-tion; we hypothesized that the use of Sm21.6 as a vaccine candidatecould be an interesting approach in the control of schistosomiasis[20].
3.4. Production of recombinant Sm21.6
The gene was cloned into pDEST17 expression vector, and theprotein was expressed in E. coli BL21 (DE3) strain. Protein extract oftransformed bacteria showed a band at ∼22 kDa when induced withIPTG (Fig. 3A, lane 2). The bacteria were then lysed by a French pressand the lysate separated into soluble and insoluble fractions. Theinclusion bodies were shown to contain the majority of the recom-binant protein, which was mostly solubilized by extraction with8 M urea (Fig. 3B, lane 3). The protein was bound to a nickel-chargedcolumn under denaturing conditions, and purified after on-columnrefolding by affinity chromatography through an imidazole linear
gradient from 10 to 500 mM (data not shown). Eluted fractions con-taining rSm21.6 were pooled, and protein yield after purificationwas estimated to be around 12.0 mg of rSm21.6/L (Fig. 3B, lane 5).After buffer exchange the protein was used in vaccination experi-ments.
D.O. Lopes et al. / Vaccine 27 (2009) 4127–4135 4131
Fig. 1. In silico analysis of S. mansoni EF-hand domain containing proteins. The full amino acid sequences of Sm21.6, Sm22.6 (AAY57921.1), Sj21.7 (AAD13338.1) and S.japonicum unknown protein (AAW25529.1) are aligned to show homologous domains (Cluacid position of each protein. EF-hand motifs are boxed in light grey while dynein domaiSyfpeithi computer program and they are underlined.
Fig. 2. PCR analysis of cDNA libraries using specific primers for the completes((
3t
f
trol group and presented the highest level at 45 days after the first
Facfli
equence of Sm21.6. Lanes: (1) ladder (bp); (2) negative control; (3) egg cDNA library;4) lung-stage cDNA library; (5) adult worm cDNA library; (6) cercariae cDNA library;7) positive control.
.5. Sm21.6 is a membrane-associated protein located on the
egument of S. mansoni adult wormA mouse serum raised against rSm21.6 revealed through con-ocal microscopy that Sm21.6 is located at the surface of the adult
ig. 3. Expression and purification of Sm21.6 in Escherichia coli. (A) SDS-PAGE (12%) profileclone before and after induction with 1 mM IPTG, respectively. (B) Inclusion bodies wereolumn and purified after on-column refolding; lanes (1) soluble fraction after lysis; (2) wow through; (5) pooled fractions of rSm21.6–6×HIS-tag fusion protein eluted after Ni2+ c
ndicate the rSm21.6.
stal W). Numbers after each row indicate the residue position relative to the aminons are boxed in dark grey (Pfam). HLA-DRB binding peptides were predicted using
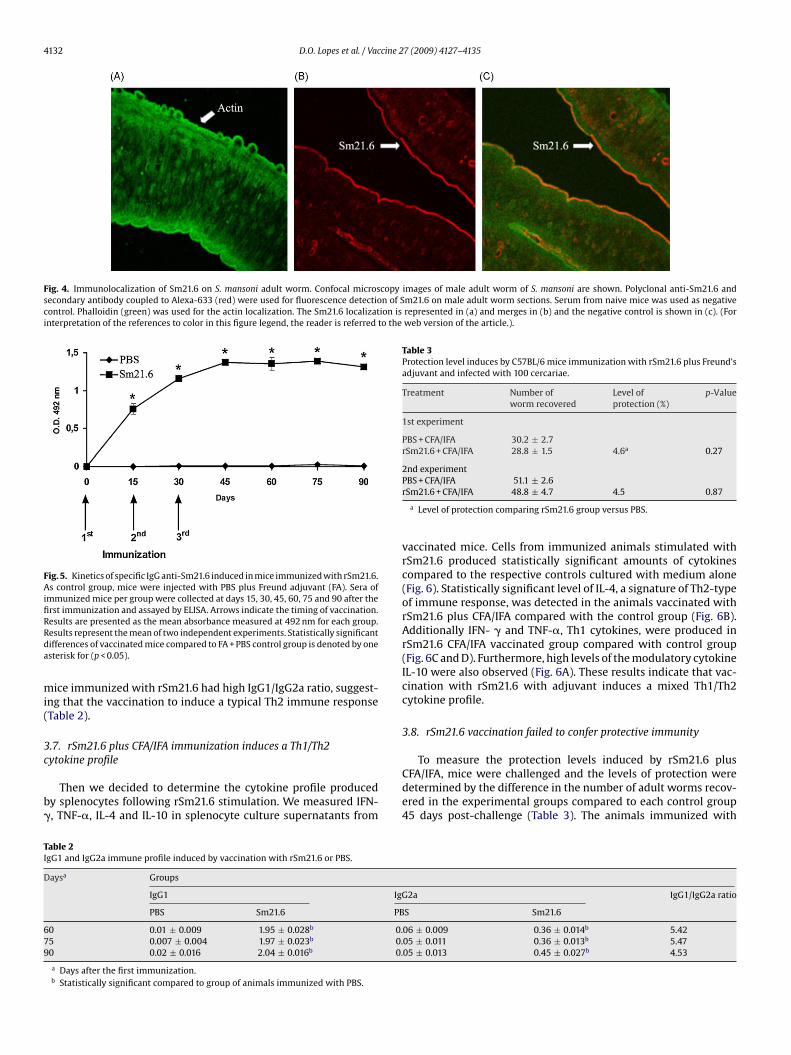
worm stage of S. mansoni (Fig. 4B and C). This membrane-associatedantigen appears to be highly expressed on the surface of adult wormand is not found in internal tissues of the parasite body. No anti-Sm21.6 antibody staining was observed in sections incubated withpre-immune mouse serum (Fig. 4A).
3.6. Analysis of IgG isotype responses to rSm21.6 followingimmunization
To investigate the presence of specific anti-Sm21.6 IgG antibod-ies, sera from vaccinated mice were tested by ELISA. Experimentalgroup vaccinated with rSm21.6 showed high levels of IgG thatappeared 15 days after the first immunization compared to the con-
immunization (Fig. 5). To determine the type of immune responseinduced after vaccination, the subclasses IgG1 and IgG2a werealso analyzed. The levels of specific anti-rSm21.6 IgG1 and IgG2aat 60, 75 and 90 days after the first immunization revealed that
of E. coli (BL21DE3) transformed with the pDEST17-Sm21.6. Lanes 1 and 2 representextracted with 8 M urea and the denatured protein (lane 3) was bind to Ni2+ charged
ashed fraction from the inclusion bodies before solubilization with 8 M urea; (4),hromatography. Positions of molecular mass standards (kDa) are indicated. Arrows
4132 D.O. Lopes et al. / Vaccine 27 (2009) 4127–4135
Fig. 4. Immunolocalization of Sm21.6 on S. mansoni adult worm. Confocal microscopy images of male adult worm of S. mansoni are shown. Polyclonal anti-Sm21.6 andsecondary antibody coupled to Alexa-633 (red) were used for fluorescence detection of Sm21.6 on male adult worm sections. Serum from naive mice was used as negativecontrol. Phalloidin (green) was used for the actin localization. The Sm21.6 localization is represented in (a) and merges in (b) and the negative control is shown in (c). (Forinterpretation of the references to color in this figure legend, the reader is referred to the web version of the article.).
Fig. 5. Kinetics of specific IgG anti-Sm21.6 induced in mice immunized with rSm21.6.As control group, mice were injected with PBS plus Freund adjuvant (FA). Sera ofimmunized mice per group were collected at days 15, 30, 45, 60, 75 and 90 after thefirst immunization and assayed by ELISA. Arrows indicate the timing of vaccination.RRda
mi(
3c
b�
Table 3Protection level induces by C57BL/6 mice immunization with rSm21.6 plus Freund’sadjuvant and infected with 100 cercariae.
Treatment Number of Level of p-Valueworm recovered protection (%)
1st experiment
PBS + CFA/IFA 30.2 ± 2.7rSm21.6 + CFA/IFA 28.8 ± 1.5 4.6a 0.27
2nd experiment
TI
D
679
esults are presented as the mean absorbance measured at 492 nm for each group.esults represent the mean of two independent experiments. Statistically significantifferences of vaccinated mice compared to FA + PBS control group is denoted by onesterisk for (p < 0.05).
ice immunized with rSm21.6 had high IgG1/IgG2a ratio, suggest-ng that the vaccination to induce a typical Th2 immune responseTable 2).
.7. rSm21.6 plus CFA/IFA immunization induces a Th1/Th2
ytokine profileThen we decided to determine the cytokine profile producedy splenocytes following rSm21.6 stimulation. We measured IFN-, TNF-�, IL-4 and IL-10 in splenocyte culture supernatants from
able 2gG1 and IgG2a immune profile induced by vaccination with rSm21.6 or PBS.
aysa Groups
IgG1 IgG
PBS Sm21.6 PB
0 0.01 ± 0.009 1.95 ± 0.028b 0.5 0.007 ± 0.004 1.97 ± 0.023b 0.0 0.02 ± 0.016 2.04 ± 0.016b 0.
a Days after the first immunization.b Statistically significant compared to group of animals immunized with PBS.
PBS + CFA/IFA 51.1 ± 2.6rSm21.6 + CFA/IFA 48.8 ± 4.7 4.5 0.87
a Level of protection comparing rSm21.6 group versus PBS.
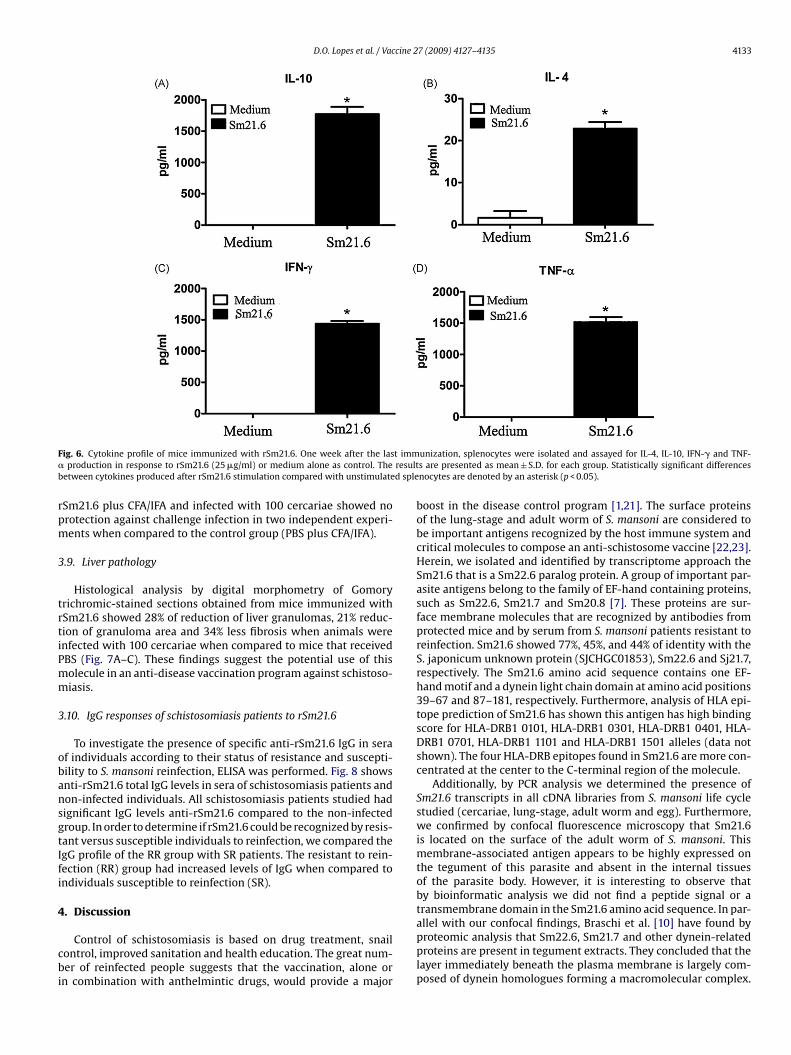
vaccinated mice. Cells from immunized animals stimulated withrSm21.6 produced statistically significant amounts of cytokinescompared to the respective controls cultured with medium alone(Fig. 6). Statistically significant level of IL-4, a signature of Th2-typeof immune response, was detected in the animals vaccinated withrSm21.6 plus CFA/IFA compared with the control group (Fig. 6B).Additionally IFN- � and TNF-�, Th1 cytokines, were produced inrSm21.6 CFA/IFA vaccinated group compared with control group(Fig. 6C and D). Furthermore, high levels of the modulatory cytokineIL-10 were also observed (Fig. 6A). These results indicate that vac-cination with rSm21.6 with adjuvant induces a mixed Th1/Th2cytokine profile.
3.8. rSm21.6 vaccination failed to confer protective immunity
To measure the protection levels induced by rSm21.6 plusCFA/IFA, mice were challenged and the levels of protection weredetermined by the difference in the number of adult worms recov-ered in the experimental groups compared to each control group45 days post-challenge (Table 3). The animals immunized with
2a IgG1/IgG2a ratio
S Sm21.6
06 ± 0.009 0.36 ± 0.014b 5.4205 ± 0.011 0.36 ± 0.013b 5.4705 ± 0.013 0.45 ± 0.027b 4.53
D.O. Lopes et al. / Vaccine 27 (2009) 4127–4135 4133
F t imm� resultb d sple
rpm
3
trtiPmm
3
obansgtIfi
4
cbi
ig. 6. Cytokine profile of mice immunized with rSm21.6. One week after the lasproduction in response to rSm21.6 (25 �g/ml) or medium alone as control. The
etween cytokines produced after rSm21.6 stimulation compared with unstimulate
Sm21.6 plus CFA/IFA and infected with 100 cercariae showed norotection against challenge infection in two independent experi-ents when compared to the control group (PBS plus CFA/IFA).
.9. Liver pathology
Histological analysis by digital morphometry of Gomoryrichromic-stained sections obtained from mice immunized withSm21.6 showed 28% of reduction of liver granulomas, 21% reduc-ion of granuloma area and 34% less fibrosis when animals werenfected with 100 cercariae when compared to mice that receivedBS (Fig. 7A–C). These findings suggest the potential use of thisolecule in an anti-disease vaccination program against schistoso-iasis.
.10. IgG responses of schistosomiasis patients to rSm21.6
To investigate the presence of specific anti-rSm21.6 IgG in seraf individuals according to their status of resistance and suscepti-ility to S. mansoni reinfection, ELISA was performed. Fig. 8 showsnti-rSm21.6 total IgG levels in sera of schistosomiasis patients andon-infected individuals. All schistosomiasis patients studied hadignificant IgG levels anti-rSm21.6 compared to the non-infectedroup. In order to determine if rSm21.6 could be recognized by resis-ant versus susceptible individuals to reinfection, we compared thegG profile of the RR group with SR patients. The resistant to rein-ection (RR) group had increased levels of IgG when compared tondividuals susceptible to reinfection (SR).
. Discussion
Control of schistosomiasis is based on drug treatment, snailontrol, improved sanitation and health education. The great num-er of reinfected people suggests that the vaccination, alone or
n combination with anthelmintic drugs, would provide a major
unization, splenocytes were isolated and assayed for IL-4, IL-10, IFN-� and TNF-s are presented as mean ± S.D. for each group. Statistically significant differencesnocytes are denoted by an asterisk (p < 0.05).
boost in the disease control program [1,21]. The surface proteinsof the lung-stage and adult worm of S. mansoni are considered tobe important antigens recognized by the host immune system andcritical molecules to compose an anti-schistosome vaccine [22,23].Herein, we isolated and identified by transcriptome approach theSm21.6 that is a Sm22.6 paralog protein. A group of important par-asite antigens belong to the family of EF-hand containing proteins,such as Sm22.6, Sm21.7 and Sm20.8 [7]. These proteins are sur-face membrane molecules that are recognized by antibodies fromprotected mice and by serum from S. mansoni patients resistant toreinfection. Sm21.6 showed 77%, 45%, and 44% of identity with theS. japonicum unknown protein (SJCHGC01853), Sm22.6 and Sj21.7,respectively. The Sm21.6 amino acid sequence contains one EF-hand motif and a dynein light chain domain at amino acid positions39–67 and 87–181, respectively. Furthermore, analysis of HLA epi-tope prediction of Sm21.6 has shown this antigen has high bindingscore for HLA-DRB1 0101, HLA-DRB1 0301, HLA-DRB1 0401, HLA-DRB1 0701, HLA-DRB1 1101 and HLA-DRB1 1501 alleles (data notshown). The four HLA-DRB epitopes found in Sm21.6 are more con-centrated at the center to the C-terminal region of the molecule.
Additionally, by PCR analysis we determined the presence ofSm21.6 transcripts in all cDNA libraries from S. mansoni life cyclestudied (cercariae, lung-stage, adult worm and egg). Furthermore,we confirmed by confocal fluorescence microscopy that Sm21.6is located on the surface of the adult worm of S. mansoni. Thismembrane-associated antigen appears to be highly expressed onthe tegument of this parasite and absent in the internal tissuesof the parasite body. However, it is interesting to observe thatby bioinformatic analysis we did not find a peptide signal or atransmembrane domain in the Sm21.6 amino acid sequence. In par-
allel with our confocal findings, Braschi et al. [10] have found byproteomic analysis that Sm22.6, Sm21.7 and other dynein-relatedproteins are present in tegument extracts. They concluded that thelayer immediately beneath the plasma membrane is largely com-posed of dynein homologues forming a macromolecular complex.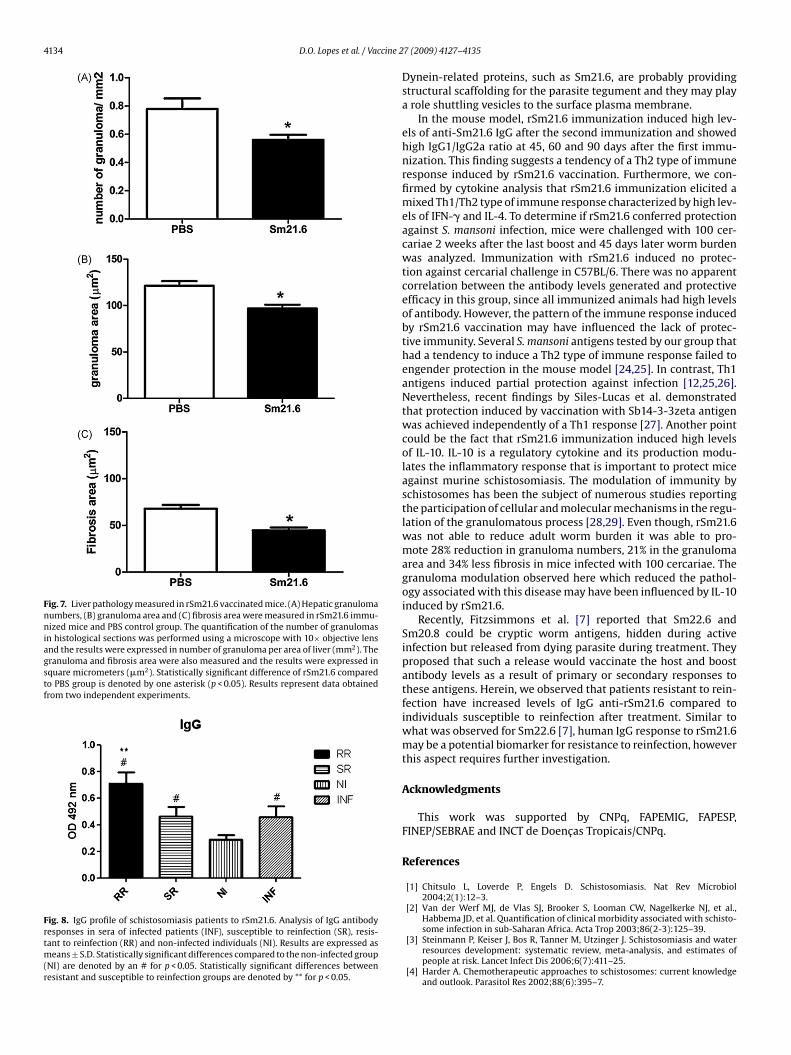
4134 D.O. Lopes et al. / Vaccine 2
Fig. 7. Liver pathology measured in rSm21.6 vaccinated mice. (A) Hepatic granulomanumbers, (B) granuloma area and (C) fibrosis area were measured in rSm21.6 immu-nized mice and PBS control group. The quantification of the number of granulomasin histological sections was performed using a microscope with 10× objective lensand the results were expressed in number of granuloma per area of liver (mm2). Thegranuloma and fibrosis area were also measured and the results were expressed insquare micrometers (�m2). Statistically significant difference of rSm21.6 comparedto PBS group is denoted by one asterisk (p < 0.05). Results represent data obtainedfrom two independent experiments.
Fig. 8. IgG profile of schistosomiasis patients to rSm21.6. Analysis of IgG antibodyresponses in sera of infected patients (INF), susceptible to reinfection (SR), resis-tant to reinfection (RR) and non-infected individuals (NI). Results are expressed asmeans ± S.D. Statistically significant differences compared to the non-infected group(NI) are denoted by an # for p < 0.05. Statistically significant differences betweenresistant and susceptible to reinfection groups are denoted by ** for p < 0.05.
7 (2009) 4127–4135
Dynein-related proteins, such as Sm21.6, are probably providingstructural scaffolding for the parasite tegument and they may playa role shuttling vesicles to the surface plasma membrane.
In the mouse model, rSm21.6 immunization induced high lev-els of anti-Sm21.6 IgG after the second immunization and showedhigh IgG1/IgG2a ratio at 45, 60 and 90 days after the first immu-nization. This finding suggests a tendency of a Th2 type of immuneresponse induced by rSm21.6 vaccination. Furthermore, we con-firmed by cytokine analysis that rSm21.6 immunization elicited amixed Th1/Th2 type of immune response characterized by high lev-els of IFN-� and IL-4. To determine if rSm21.6 conferred protectionagainst S. mansoni infection, mice were challenged with 100 cer-cariae 2 weeks after the last boost and 45 days later worm burdenwas analyzed. Immunization with rSm21.6 induced no protec-tion against cercarial challenge in C57BL/6. There was no apparentcorrelation between the antibody levels generated and protectiveefficacy in this group, since all immunized animals had high levelsof antibody. However, the pattern of the immune response inducedby rSm21.6 vaccination may have influenced the lack of protec-tive immunity. Several S. mansoni antigens tested by our group thathad a tendency to induce a Th2 type of immune response failed toengender protection in the mouse model [24,25]. In contrast, Th1antigens induced partial protection against infection [12,25,26].Nevertheless, recent findings by Siles-Lucas et al. demonstratedthat protection induced by vaccination with Sb14-3-3zeta antigenwas achieved independently of a Th1 response [27]. Another pointcould be the fact that rSm21.6 immunization induced high levelsof IL-10. IL-10 is a regulatory cytokine and its production modu-lates the inflammatory response that is important to protect miceagainst murine schistosomiasis. The modulation of immunity byschistosomes has been the subject of numerous studies reportingthe participation of cellular and molecular mechanisms in the regu-lation of the granulomatous process [28,29]. Even though, rSm21.6was not able to reduce adult worm burden it was able to pro-mote 28% reduction in granuloma numbers, 21% in the granulomaarea and 34% less fibrosis in mice infected with 100 cercariae. Thegranuloma modulation observed here which reduced the pathol-ogy associated with this disease may have been influenced by IL-10induced by rSm21.6.
Recently, Fitzsimmons et al. [7] reported that Sm22.6 andSm20.8 could be cryptic worm antigens, hidden during activeinfection but released from dying parasite during treatment. Theyproposed that such a release would vaccinate the host and boostantibody levels as a result of primary or secondary responses tothese antigens. Herein, we observed that patients resistant to rein-fection have increased levels of IgG anti-rSm21.6 compared toindividuals susceptible to reinfection after treatment. Similar towhat was observed for Sm22.6 [7], human IgG response to rSm21.6may be a potential biomarker for resistance to reinfection, howeverthis aspect requires further investigation.
Acknowledgments
This work was supported by CNPq, FAPEMIG, FAPESP,FINEP/SEBRAE and INCT de Doencas Tropicais/CNPq.
References
[1] Chitsulo L, Loverde P, Engels D. Schistosomiasis. Nat Rev Microbiol2004;2(1):12–3.
[2] Van der Werf MJ, de Vlas SJ, Brooker S, Looman CW, Nagelkerke NJ, et al.,Habbema JD, et al. Quantification of clinical morbidity associated with schisto-
some infection in sub-Saharan Africa. Acta Trop 2003;86(2-3):125–39.[3] Steinmann P, Keiser J, Bos R, Tanner M, Utzinger J. Schistosomiasis and waterresources development: systematic review, meta-analysis, and estimates ofpeople at risk. Lancet Infect Dis 2006;6(7):411–25.
[4] Harder A. Chemotherapeutic approaches to schistosomes: current knowledgeand outlook. Parasitol Res 2002;88(6):395–7.

cine 2
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
D.O. Lopes et al. / Vac
[5] Bergquist NR. Schistosomiasis vaccine development: progress and prospects.Mem Inst Oswaldo Cruz 1998;93(1):95–101.
[6] Bergquist NR. Schistosomiasis: from risk assessment to control. Trends Parasitol2002;18(7):309–14.
[7] Fitzsimmons CM, McBeath R, Joseph S, Jones FM, Walter K, Hoffmann KF, etal. Factors affecting human IgE and IgG responses to allergen-like Schistosomamansoni antigens: molecular structure and patterns of in vivo exposure. IntArch Allergy Immunol 2007;142(1):40–50.
[8] Fitzsimmons CM, Stewart TJ, Hoffmann KF, Grogan JL, Yazdanbakhsh M, DunneDW. Human IgE response to the Schistosoma haematobium 22 6 kDa antigen.Parasite Immunol 2004;26(8-9):371–6.
[9] Francis P, Bickle Q. Cloning of a 21 7-kDa vaccine-dominant antigen gene ofSchistosoma mansoni reveals an EF hand-like motif. Mol Biochem Parasitol1992;50(2):215–24.
10] Braschi S, Curwen RS, Ashton PD, Verjovski-Almeida S, Wilson A. The tegu-ment surface membranes of the human blood parasite Schistosoma mansoni:a proteomic analysis after differential extraction. Proteomics 2006;6(5):1471–82.
11] Loukas A, Tran M, Pearson MS. Schistosome membrane proteins as vaccines. IntJ Parasitol 2007;37(3-4):257–63.
12] Cardoso FC, Macedo GC, Gava E, Kitten GT, Mati VL, de Melo AL, et al. Schis-tosoma mansoni tegument protein Sm29 is able to induce a Th1-type ofimmune response and protection against parasite infection. PLoS Negl TropDis 2008;2(10):e308.
13] Tran MH, Pearson MS, Bethony JM, Smyth DJ, Jones MK, Duke M, et al.Tetraspanins on the surface of Schistosoma mansoni are protective antigensagainst schistosomiasis. Nat Med 2006;12(7):835–40.
14] Pacífico LG, Marinho FA, Fonseca CT, Barsante MM, Pinho V, Sales-Junior PA,et al. Schistosoma mansoni antigens modulate experimental allergic asthma ina murine model: a major role for CD4+ CD25+ Foxp3+ T cells independent ofinterleukin-10. Infect Immun 2009;77(1):98–107.
15] Verjovski-Almeida S, DeMarco R, Martins EAL, Guimarães PEM, Ojopi EPB,Paquola ACM, et al. Transcriptome analysis of the acoelomate human parasiteSchistosoma mansoni. Nat Genet 2003;35(2):148–57.
16] Fonseca CT, Pacífico LG, Barsante MM, Rassi T, Cassali GD, Oliveira SC. Co-administration of plasmid expressing IL-12 with 14-kDa Schistosoma mansoni
fatty acid-binding protein cDNA alters immune response profiles and failsto enhance protection induced by Sm14 DNA vaccine alone. Microbes Infect2006;8(9-10):2509–16.17] Cardoso LS, Araujo MI, Goes AM, Pacifico LG, Oliveira RR, Oliveira SC. PolymyxinB as inhibitor of LPS contamination of Schistosoma mansoni recombinant pro-teins in human cytokine analysis. Microb Cell Fact 2007;6:1.
[
7 (2009) 4127–4135 4135
18] Pacheco LG, Zucconi E, Mati VL, Garcia RM, Miyoshi A, Oliveira SC, et al. Oraladministration of a live Aro attenuated Salmonella vaccine strain expressing 14-kDa Schistosoma mansoni fatty acid-binding protein induced partial protectionagainst experimental schistosomiasis. Acta Trop 2005;95(2):132–42.
19] Cardoso FC, Pacífico RNA, Mortara RA, Oliveira SC. Human antibody responsesof patients living in endemic areas for schistosomiasis to the tegumen-tal protein Sm29 identified through genomic studies. Clin Exp Immunol2006;144(3):382–91.
20] Oliveira SC, Fonseca CT, Cardoso FC, Farias LP, Leite LC. Recent advances invaccine research against schistosomiasis in Brazil. Acta Trop 2008;108:256–62.
21] Hotez PJ, Bethony JM, Oliveira SC, Brindley PJ, Loukas A. Multivalentanthelminthic vaccine to prevent hookworm and schistosomiasis. Expert RevVaccines 2008;7(6):745–52.
22] Simpson AJ. Schistosome surface antigens: developmental expression andimmunological function. Parasitol Today 1990;6(2):40–5.
23] Wilson RA, Langermans JA, van Dam GJ, Vervenne RA, Hall SL, Borges WC, etal. Elimination of Schistosoma mansoni Adult worms by rhesus macaques: basisfor a therapeutic vaccine? PLoS Negl Trop Dis 2008;2(9):e290.
24] Pacífico LG, Fonseca CT, Barsante MM, Cardoso LS, Araújo MI, Oliveira SC. Alu-minum hydroxide associated to Schistosoma mansoni 22 6 kDa protein abrogatespartial protection against experimental infection but not alter interleukin-10production. Mem Inst Oswaldo Cruz 2006;101(1):365–8.
25] Garcia TC, Fonseca CT, Pacifico LG, Durães F, do V, Marinho FA, et al. Pep-tides containing T cell epitopes, derived from Sm14, but not from paramyosin,induce a Th1 type of immune response, reduction in liver pathology andpartial protection against Schistosoma mansoni infection in mice. Acta Trop2008;106(3):162–7.
26] Fonseca CT, Brito CF, Alves JB, Oliveira SC. IL-12 enhances protective immunityin mice engendered by immunization with recombinant 14 kDa Schistosomamansoni fatty acid-binding protein through an IFN-gamma and TNF-alphadependent pathway. Vaccine 2004;22(3-4):503–10.
27] Siles-Lucas M, Uribe N, López-Abán J, Vicente B, Orfao A, Nogal-Ruiz JJ, et al. TheSchistosoma bovis Sb14-3-3zeta recombinant protein cross-protects againstSchistosoma mansoni in BALB/c mice. Vaccine 2007;25(41):7217–23.
28] Bazzone LE, Smith PM, Rutitzky LI, Shainheit MG, Urban JF, Setiawan T, etal. Coinfection with the intestinal nematode Heligmosomoides polygyrus
markedly reduces hepatic egg-induced immunopathology and proinflamma-tory cytokines in mouse models of severe schistosomiasis. Infect Immun2008;76(11):5164–72.29] Layland LE, Wagner H, da Costa CU. Lack of antigen-specific Th1 responsealters granuloma formation and composition in Schistosoma mansoni-infectedMyD88-/- mice. Eur J Immunol 2005;35(11):3248–57.




















