
The anthelmintic effect of plumbagin on Schistosoma mansoni
10
The anthelmintic effect of plumbagin on Schistosoma mansoni Natcha Lorsuwannarat a , Naruwan Saowakon b , Pongrama Ramasoota c , Chaitip Wanichanon a , Prasert Sobhon a,⇑ a Department of Anatomy, Faculty of Science, Mahidol University, Rama VI Road, Ratchathewi, Bangkok 10400, Thailand b Faculty of Science, Suranaree University of Technology, University Avenue, Suranaree, Muang, Nakhon Ratchasima 30000, Thailand c Department of Social and Environmental Medicine, Faculty of Tropical Medicine, Mahidol University, Ratchawithi Road, Ratchathewi, Bangkok 10400, Thailand highlights " PB exhibited more anthelmintic effect against adult Schistosoma mansoni than PZQ with IC 50 0.95 lg/ml versus 16.22 lg/ml. " Both PB and PZQ exhibited more effect on male than female parasites. " PB and PZQ caused similar sequences of tegumental damages. " Mitochondria might be the major target of PB. graphical abstract article info Article history: Received 15 August 2012 Received in revised form 28 September 2012 Accepted 2 October 2012 Available online 16 October 2012 Keywords: Schistosoma mansoni Plumbagin Anthelmintic Motility Survival Tegument abstract The anthelmintic effects of plumbagin (PB, 5-hydroxy-2-methyl-1,4-naphthoquinone) and praziquantel (PZQ) against adult Schistosoma mansoni in vitro were compared by estimating the relative motility (RM) values, survival indices (SI) and alterations of the tegument of flukes incubated in M-199 medium containing 1, 10 and 100 lg/ml of the drugs, at 1, 3, 6, 12 and 24 h. For the parasites incubated in 10 lg/ ml, the RM values of the PB-treated group decreased at a more rapid rate than the PZQ-treated group. When incubated in 100 lg/ml all PB-treated flukes appeared immobile from 1 to 24 h when 91–100% died, while in the PZQ-treated group RM values were higher than that of PB and most flukes were still alive at 1–12 h, and at 24 h only 21% of flukes were killed. Furthermore, male parasites were more affected by PZQ than females as their RM values were significantly less than that of females at all doses. While in PB treatment, males and females showed less difference in response to the drug as their RM val- ues were closer than those treated with PZQ. When observed by SEM, the tegument of untreated S. man- soni displayed no alteration, while in PB treated parasites it exhibited swelling, blebbing, loss of spines, disruption of tubercles and ridges, leading to erosion and lesion, exposure of the basal lamina, and sloughing of the tegument. PZQ induced similar tegumental changes as those observed in PB treatment but at longer incubation time and higher doses. These data indicated that PB had more anthelmintic effect on both sexes of adult S. mansoni than PZQ. Ó 2012 Elsevier Inc. All rights reserved. 1. Introduction Schistosoma mansoni is one of the major parasites causing hu- man schistosomiasis, recognized as a major global health problem with over 200 million people afflicted worldwide (World Health 0014-4894/$ - see front matter Ó 2012 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.exppara.2012.10.003 Abbreviations: DMSO, dimethyl sulphoxide; PB, 5-hydroxy-2-methyl-1,4-naph- thoquinone; PZQ, praziquantel. ⇑ Corresponding author. Fax: +66 2 3547168. E-mail addresses: [email protected] (N. Lorsuwannarat), prasert.sob@ mahidol.ac.th (P. Sobhon). Experimental Parasitology 133 (2013) 18–27 Contents lists available at SciVerse ScienceDirect Experimental Parasitology journal homepage: www.elsevier.com/locate/yexpr
Transcript of The anthelmintic effect of plumbagin on Schistosoma mansoni

Experimental Parasitology 133 (2013) 18–27
Contents lists available at SciVerse ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/locate /yexpr
The anthelmintic effect of plumbagin on Schistosoma mansoni
Natcha Lorsuwannarat a, Naruwan Saowakon b, Pongrama Ramasoota c, Chaitip Wanichanon a,Prasert Sobhon a,⇑a Department of Anatomy, Faculty of Science, Mahidol University, Rama VI Road, Ratchathewi, Bangkok 10400, Thailandb Faculty of Science, Suranaree University of Technology, University Avenue, Suranaree, Muang, Nakhon Ratchasima 30000, Thailandc Department of Social and Environmental Medicine, Faculty of Tropical Medicine, Mahidol University, Ratchawithi Road, Ratchathewi, Bangkok 10400, Thailand
h i g h l i g h t s
" PB exhibited more anthelminticeffect against adult Schistosomamansoni than PZQwith IC50 0.95 lg/ml versus16.22 lg/ml.
" Both PB and PZQ exhibited moreeffect on male than female parasites.
" PB and PZQ caused similar sequencesof tegumental damages.
" Mitochondria might be the majortarget of PB.
0014-4894/$ - see front matter � 2012 Elsevier Inc. Ahttp://dx.doi.org/10.1016/j.exppara.2012.10.003
Abbreviations: DMSO, dimethyl sulphoxide; PB, 5-thoquinone; PZQ, praziquantel.⇑ Corresponding author. Fax: +66 2 3547168.
E-mail addresses: [email protected] (N. Lomahidol.ac.th (P. Sobhon).
g r a p h i c a l a b s t r a c t
a r t i c l e i n f o
Article history:Received 15 August 2012Received in revised form 28 September2012Accepted 2 October 2012Available online 16 October 2012
Keywords:Schistosoma mansoniPlumbaginAnthelminticMotilitySurvivalTegument
a b s t r a c t
The anthelmintic effects of plumbagin (PB, 5-hydroxy-2-methyl-1,4-naphthoquinone) and praziquantel(PZQ) against adult Schistosoma mansoni in vitro were compared by estimating the relative motility(RM) values, survival indices (SI) and alterations of the tegument of flukes incubated in M-199 mediumcontaining 1, 10 and 100 lg/ml of the drugs, at 1, 3, 6, 12 and 24 h. For the parasites incubated in 10 lg/ml, the RM values of the PB-treated group decreased at a more rapid rate than the PZQ-treated group.When incubated in 100 lg/ml all PB-treated flukes appeared immobile from 1 to 24 h when 91–100%died, while in the PZQ-treated group RM values were higher than that of PB and most flukes were stillalive at 1–12 h, and at 24 h only 21% of flukes were killed. Furthermore, male parasites were moreaffected by PZQ than females as their RM values were significantly less than that of females at all doses.While in PB treatment, males and females showed less difference in response to the drug as their RM val-ues were closer than those treated with PZQ. When observed by SEM, the tegument of untreated S. man-soni displayed no alteration, while in PB treated parasites it exhibited swelling, blebbing, loss of spines,disruption of tubercles and ridges, leading to erosion and lesion, exposure of the basal lamina, andsloughing of the tegument. PZQ induced similar tegumental changes as those observed in PB treatmentbut at longer incubation time and higher doses. These data indicated that PB had more anthelmintic effecton both sexes of adult S. mansoni than PZQ.
� 2012 Elsevier Inc. All rights reserved.
ll rights reserved.
hydroxy-2-methyl-1,4-naph-
rsuwannarat), prasert.sob@
1. Introduction
Schistosoma mansoni is one of the major parasites causing hu-man schistosomiasis, recognized as a major global health problemwith over 200 million people afflicted worldwide (World Health

N. Lorsuwannarat et al. / Experimental Parasitology 133 (2013) 18–27 19
Organization, 2001). The infection is a potentially life-threateningwith severe pathological manifestations in the hepato-splenic sys-tem and gastrointestinal tract (Chitsulo et al., 2000; Neghina et al.,2009). Presently, there is no satisfactory schistosomiasis vaccine,and treatments with drugs are the only effective way to controlthe infection. Praziquantel (PZQ) is a drug of choice against thisparasite, which acts by inducing a spastic contraction of its bodythrough the influx of Ca++ via the cell membrane (Becker et al.,1980). However, resistance to this drug is emerging in many areas(Fallon and Doenhoff, 1994; Pica-Mattoccia and Cioli, 2004), andthere are no other antischistosomal compounds of comparable effi-cacy currently available. Thus, there is an urgent need to find newsubstances that can be developed into alternative antischistosomaldrugs.
Plumbagin (PB, 5-hydroxy-2-methyl-1,4-naphthoquinone) is anatural yellowish quinonoid compound derived from the rootsof many plants, especially subscandent shrubs of Plumbagospecies, which is used as a traditional medicine for treatments ofdiarrhea, anasarca, skin disease, and also used as emmenagogue,appetite stimulant and antiflatulence (Satyavati et al., 1976;Wuttidhammaved, 1997). In addition, PB has been reported to haveantioxidant (Talcott et al., 1985), antimicrobial (Selma Ribeiro dePaiva et al., 2003; Sharma et al., 2009), anti-inflammatory(Reese et al., 2010) and antineoplastic effects (Kini et al.,1997; Krishnaswamy and Purushothaman, 1980). Recently, theantiparasitic activity of PB had been investigated by our group(Atjanasuppat et al., 2009) which showed that PB had the strongestactivity against Caenorhabditis elegans (IC50 at 9.71 lg/ml) andParamphistomum epiclitum (IC50 at 130 lg/ml), respectively. Wehypothesize that PB also possesses anthelmintic effect againstschistosomes of both human and animals.
In this study, we compared the in vitro anthelmintic effect of PBagainst adult S. mansoni with the currently used drug, PZQ, by esti-mating the relative motility (RM), survival indices (SI), and tegu-mental surface alterations in the treated parasites, as well asestimated the differential effects of PB on both sexes.
2. Materials and methods
2.1. Parasites
Adult S. mansoni were collected from infected ICR mice. Maleand female 8 weeks old mice, weighing approximately 38–40 g,were purchased from National Laboratory Animal Center, NakhonPathom Province, Thailand. Cercariae of S. mansoni were collectedfrom infected Biomphalaria glabrata snails after exposure to a lampfor 6 h. Each mouse was infected with approximately 300 cercariaeby tail immersion and kept at the Applied Malacology Center,Department of Social and Environmental Medicine, Faculty ofTropical Medicine, Mahidol University. Eight week after infectionthe mice were sacrificed and perfused with 0.85% NaCl solution,and the parasites were collected from the perfusate as well as fromthe minced liver. Male and female flukes were separated by beingteased apart with a paint brush and collected by a Pasteur pipette.Then they were washed several times with 0.85% NaCl solution.Only intact and actively mobile flukes were used immediately forin vitro incubation with the drugs. All the animal protocols hadbeen approved by the Animal Ethic Committee, Faculty of Science,Mahidol University.
2.2. Drugs and media
Plumbagin (5-hydroxy-2-methyl-1,4-naphthoquinone) waspurchased from Sigma Chemical Co. (St Louis, MO, USA). The stocksolution of plumbagin (PB) was made by dissolving 100 mg PB
powder in 1 ml dimethyl sulphoxide (DMSO) (Sigma Co.). The PBconcentrations used in this experiment were based on thestudy of Atjanasuppat et al. (2009), which reported half maximalinhibitory concentrations (IC50) of C. elegans and P. epiclitumin vitro between 9.71 and 130 lg/ml, respectively. A commercialanthelmintic praziquantel (PZQ) (Wormicide, Greater PharmaCo.) was used as the positive control. Stock solution was madeby dissolving 100 mg powder of PZQ in 1 ml of DMSO. TheM-199 medium (Sigma Co.) containing antibiotics (penicillin50 IU/ml, gentamycin 30 IU/ml), was mixed with the drug stocksolutions to obtain the required concentrations (1, 10 and 100 lg/ml).All chemicals used in the experiment were of analytical grade.
2.3. Assessment of the drug efficacies by relative motility (RM) valuesand survival indices (SI)
Adult flukes collected as above were randomly assigned to se-ven groups and incubated in tissue culture dishes (Nunc, Sigma–Aldrich) (P60 flukes per group). Group 1 was the negative controlwith the parasites incubated in Medium-199 containing 0.1% (v/v)DMSO. Groups 2–4 were the positive controls with the parasitesincubated in the medium contained PZQ at 1, 10 and 100 lg/ml.Groups 5–7 were the experimental groups incubated in the med-ium with PB at the same concentrations. The parasites were incu-bated for 1, 3, 6, 12 and 24 h in an incubator with 5% CO2 at 37 �C.The motility were evaluated by examining the flukes under a ste-reomicroscope and assigned the scores according to the criteriaset by Kiuchi et al. (1987), i.e., 3 = active movement of the wholebody, 2 = movement of only some parts of the body, 1 = immobile,and unstained with a vital dye (1% methylene blue in 0.85% NaClsolution), and 0 = immobile and stained with the vital dye (indicat-ing the parasites were dead).
2.3.1. Relative motility (RM) valueRM values were calculated using the formula listed below
(Kiuchi et al., 1987)
Motility index ðMIÞ ¼ R n NR N
RM value ¼MI experiementMI control
� 100
n = motility score, N = number of flukes with the score of ‘‘n’’.The negative control group where all the parasites scored 3 had
the RM value of 100, and the smaller RM values indicated strongerdrug activity.
2.3.2. Survival index (SI)Survival indices indicated the percentages of live flukes at a gi-
ven time after treatment. The flukes with motility score of 0(immobile and stained with the vital dye) were counted as dead,and those with other scores (3, 2, 1) were counted as still alive. Sur-vival indices were calculated using the formula listed below andthe survival index 0 indicated that all flukes had died.
Survival index ðSIÞ ¼ Number of live flukesNumber of all flukes
� 100
2.4. IC50 value
IC50 value is a concentration of the drug required for 50% inhi-bition of the movement after the parasites were incubatedin vitro in the drug for 24 h, determined by using an Excel add-inprogram ED50V10. The program calculated IC50 values by usingthe RM values at each drug concentration.

20 N. Lorsuwannarat et al. / Experimental Parasitology 133 (2013) 18–27
2.5. Data analysis
The means and standard deviations of RM values and survivalindices were calculated for each group. A Kruskal Wallis/Dunn’stest was used to determine the significant differences betweendrug doses in each treated groups. Comparative anthelmintic ef-fects of PZQ and PB including sex sensitivities were evaluated byMann–Whitney U test, assuming unequal variances. The statisticalanalysis was interpreted to be significantly different when p-valuewas <0.05.
2.6. Scanning electron microscopic (SEM) examination of the tegument
Ten flukes were collected from other negative control and PB-and PZQ-treated groups at each incubation times of the drug con-centrations. The parasites were fixed in 2.5% glutaraldehyde and 2%paraformaldehyde in 0.1 M phosphate buffer solution (PBS), pH 7.4at 4 �C overnight. Then, they were washed with 0.1 M PBS, pH 7.4three times for 15 min each, posted–fixed in 1% OsO4 in 0.1 M PBSfor 1 h at 4 �C, and washed with cooled distilled water three timesfor 15 min each. Subsequently, they were dehydrated through seri-al ethanol concentrations (30%, 50%, 70%, 80%, 90%, 95% and 100%)for 30 min each. Thereafter, the parasites were dried in a HitachiHCP-2 critical point drying machine using liquid CO2 as a transi-tional medium. Then, the dried specimens were mounted on alu-minum stubs and coated with platinum and palladium in an ionsputtering apparatus, Hitachi E-102, at 10–15 mA for 6 min. Theywere examined and photographed under a Hitachi scanning elec-tron microscope S-2500, operating at 15 kV.
Fig. 1. (A) Relative motility (RM) values of adult S. mansoni (including both sexes) treate(B) Survival indices (SI) of adult S. mansoni (including both sexes) treated with praziqua
3. Results
3.1. RM and SI values of the flukes treated with PB and PZQ
As shown in Fig. 1A and B, all flukes (including both males andfemales) in the negative control group exhibited active movement,and were alive throughout the experimental period (RM = 100,SI = 100). In the group treated with 1 lg/ml PB, at 1 h all the flukesexhibited active movement and appeared normal as in the negativecontrol group (RM = 99, SI = 100). At 3–6 h the flukes exhibiteddecreased RM values (reduced motility) that were higher thanPZQ-treated group, but between 6 and 24 h the RM values of thePB-treated group were lower than that of the PZQ-treated group.
In 10 lg/ml, the RM values of PB-treated flukes decreased at amore rapid rate than the flukes treated with PZQ. The RM valuesof the PB-treated flukes were reduced by 50% at 1 h (RM = 50)and 99% at 24 h (RM = 1), while flukes treated with PZQ showed37% reduction of RM values at 1 h (RM = 63), and 47% reductionat 24 h (RM = 53). At 6 h incubation, 100% of PB-treated flukes be-came completely immobile (36% still alive, 66% were killed), and96% of them died at 24 h (SI = 4). In comparison, when incubatedin 10 lg/ml PZQ 58% of the flukes were immobilized at 6 h butall of them were still alive (SI = 100). At 24 h, only 15% of thePZQ-treated flukes were dead (SI = 85).
In 100 lg/ml, all PB-treated flukes ceased to be mobile at 1 h,while the RM values dropped rapidly to 3 and were stable between1 and 12 h; then the RM value became 0 at 24 h. Survival index ofthe flukes in these groups was reduced from 100% to approxi-mately 9% and appeared to be stable from 1 to 12 h, until at 24 h
d with praziquantel (PZQ) and plumbagin (PB) at various concentrations and times.ntel (PZQ) and plumbagin (PB) at various concentrations and times.

Fig. 2. (A) Comparative anthelmintic effects of PZQ and PB on S. mansoni at threedoses of treatments based on relative motility (RM) values. The RM values from alltime points of each dose were pooled and expressed as mean ± SE. Significantdifference between each group with the negative control is indicated by ‘‘a’’,between the consecutive doses of the same drug is indicated by ‘‘b’’ and ‘‘c’’(compared with 1 and 10 lg/ml, respectively), and between the same doses of PZQand PB indicated by ‘‘d’’, (p < 0.05). (B) Comparative anthelmintic effects of PZQ andPB on S. mansoni based on survival indices (SI). The SI values from all time points ofeach dose were pooled and expressed as mean ± SE. Significant difference betweeneach group with the negative control is indicated by ‘‘a’’, between the consecutivedoses of the same drug is indicated by ‘‘b’’ and ‘‘c’’ (compared with 1 and 10 lg/ml,respectively), and between the same dose of PZQ and PB indicated by ‘‘d’’, (p < 0.05).
N. Lorsuwannarat et al. / Experimental Parasitology 133 (2013) 18–27 21
when all flukes took up the vital dye which indicated that theywere dead (SI = 0). When incubated in PZQ at 100 lg/ml, the flukes’RM values and motility were decreased throughout the experimen-tal period as compared to negative control group (RM values re-duced from 62 at 1 h to 42 at 24 h). Although some flukes wereimmobile during these incubation times, most of them were stillalive and unstained by vital dye. At 24 h incubation only 21% ofthe PZQ-treated flukes were killed (SI = 79).
3.2. Comparison of the anthelmintic effects of praziquantel (PZQ) andplumbagin (PB)
When comparing the effects of varying doses of PZQ, by poolingRM and SI values from all time points of each dose (Fig. 2A and B),the RM values were decreased to 60, 59 and 53 for doses 1, 10 and100 lg/ml, respectively, which were significantly reduced from thenegative control group. However, the differences between thethree doses were not significant. As well for SI there was no signif-icant difference from the negative control among the three doses.The IC50 of PZQ for the parasites (pooled both sexes) at 24 h wascalculated to be 16.22 lg/ml.
When comparing the effects of varying doses of PB, RM valueswere significantly declined in comparison to the negative control,to 74, 19 and 2 for the doses 1, 10 and 100 lg/ml, respectively.Fig. 2A clearly exhibited a greater and significant decrease of motil-ity at higher doses of PB. The survival indices were also significantly
decreased from negative control group when treated with 10 and100 lg/ml of PB at 46% and 7%, respectively, whereas the decreasewas not significantly at 1 lg/ml, (Fig. 2B). The IC50 of PB for bothsexes of the parasites at 24 h was calculated to be 0.95 lg/ml.
When comparing the anthelmintic effects between the twodrugs (Fig. 2A and B), PB showed a higher and significant differentRM value than PZQ at dose 1 lg/ml (p < 0.05), but the SI valueswere not significantly different from each other and from the neg-ative control. The RM and SI values of PB were much lower and sig-nificantly different from PZQ and the negative control at doses 10and 100 lg/ml (p < 0.05).
3.3. Sex-related sensitivities to the drugs
The relative sensitivities of adult female and male flukes to PZQand PB were estimated. The paired male and female were sepa-rated by using a paint brush and collected by a Pasteur pipette.They were placed in the same tissue culture dish containing med-ium M-199 with the drugs at the same concentrations (1, 10 and100 lg/ml). Then the RM and SI values were evaluated, and sensi-tivities to the drugs were compared between the two sexes (Fig. 3Aand B).
For both males and females, PB caused significantly lower RMand SI values than PZQ and negative control at the doses 10 and100 lg/ml (p < 0.05). When compared between sexes, PB causedsignificantly lower RM and SI values in males than females onlyat dose 100 lg/ml (p < 0.05). In contrast, in the PZQ treatment,males showed significantly lower RM values than females at alldoses (p < 0.05), but no significantly difference in SI values fromfemales.
3.4. Changes of tegumental surface as observed by SEM
3.4.1. Control parasitesThe body of the adult fluke can be divided into three parts based
on characteristics of the surface features: (1) the anterior partstarting from the oral sucker to the area behind ventral sucker;(2) the middle part covering the largest area of body; and (3) theposterior part showing fewer tubercles in male or crowded spinesin female. Overall the surface morphology of the males and femaleparasites are quite different as have been previously described(Becker et al., 1980; Xiao et al., 2000). However, as the base linesfor comparing the drugs’ effects we had observed and, here, de-scribed the key surface features of the two sexes in the negativecontrol group (Fig. 4). The adult male is larger and shorter thanthe female and has a gynaecophoric canal at the ventral surface,in which the female is lodged during copulation (Fig. 4A). The high-er magnifications (insets) are representative images of the surfacesfrom the head and middle regions of the male’s body. There arenumerous large tubercles with spines on the middle part of thebody (Fig. 4B), and they become fewer at the posterior end(Fig. 4C). At high magnification, the surface showed spiny tuberclesand ridges between the tubercles which have a pitted appearanceand bear sensory papillae (Fig. 4B). In adult female, the surface ap-pears smooth and less prominent than that of male due to the lackof spiny tubercles as represent by the higher magnification (inset).(Fig. 4D). Ridges were separated by grooves and they appeared pit-ted (Fig. 4E, inset ii). There are also numerous sensory papillae withcilia on the ridges (Fig 4E, inset i). Numerous spines are presentonly at the posterior end (Fig. 4F). After 24 h incubation in themedium without drugs, the tubercles, spines and tegumentalridges of the flukes appeared intact.
3.4.2. Tegument damages following the treatments with PZQ and PBThe tegumental surface alterations and damages due to the
treatments by PZQ and PB exhibited the same sequences but
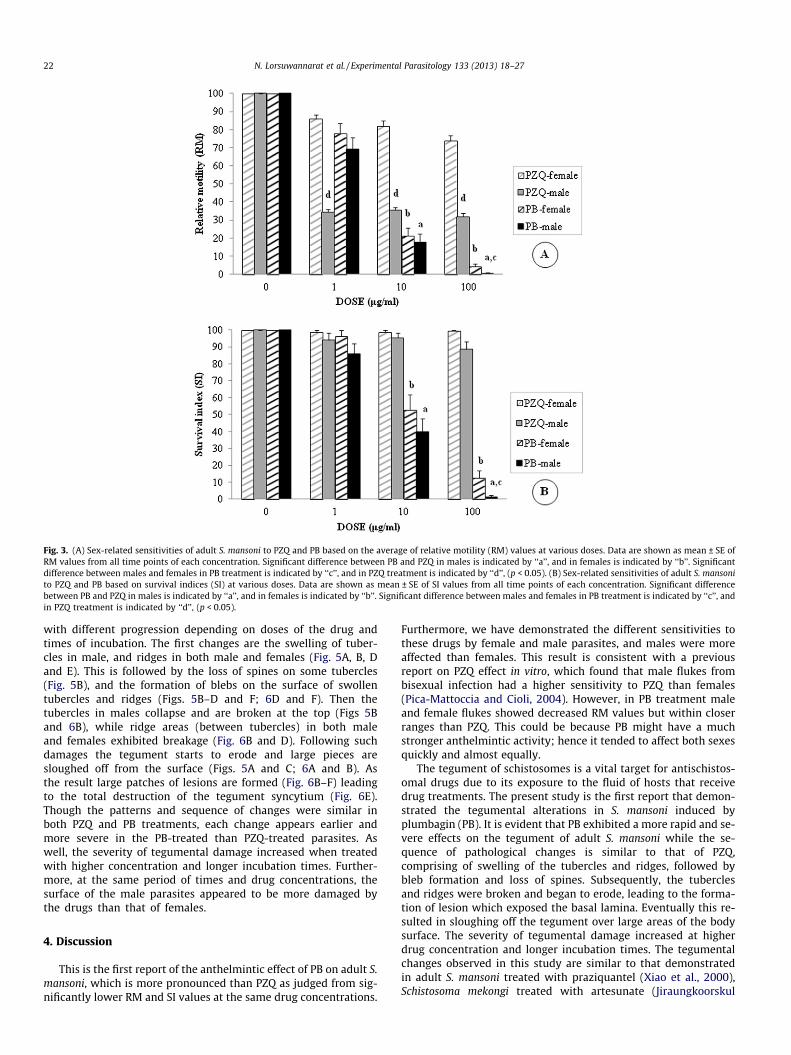
Fig. 3. (A) Sex-related sensitivities of adult S. mansoni to PZQ and PB based on the average of relative motility (RM) values at various doses. Data are shown as mean ± SE ofRM values from all time points of each concentration. Significant difference between PB and PZQ in males is indicated by ‘‘a’’, and in females is indicated by ‘‘b’’. Significantdifference between males and females in PB treatment is indicated by ‘‘c’’, and in PZQ treatment is indicated by ‘‘d’’, (p < 0.05). (B) Sex-related sensitivities of adult S. mansonito PZQ and PB based on survival indices (SI) at various doses. Data are shown as mean ± SE of SI values from all time points of each concentration. Significant differencebetween PB and PZQ in males is indicated by ‘‘a’’, and in females is indicated by ‘‘b’’. Significant difference between males and females in PB treatment is indicated by ‘‘c’’, andin PZQ treatment is indicated by ‘‘d’’, (p < 0.05).
22 N. Lorsuwannarat et al. / Experimental Parasitology 133 (2013) 18–27
with different progression depending on doses of the drug andtimes of incubation. The first changes are the swelling of tuber-cles in male, and ridges in both male and females (Fig. 5A, B, Dand E). This is followed by the loss of spines on some tubercles(Fig. 5B), and the formation of blebs on the surface of swollentubercles and ridges (Figs. 5B–D and F; 6D and F). Then thetubercles in males collapse and are broken at the top (Figs 5Band 6B), while ridge areas (between tubercles) in both maleand females exhibited breakage (Fig. 6B and D). Following suchdamages the tegument starts to erode and large pieces aresloughed off from the surface (Figs. 5A and C; 6A and B). Asthe result large patches of lesions are formed (Fig. 6B–F) leadingto the total destruction of the tegument syncytium (Fig. 6E).Though the patterns and sequence of changes were similar inboth PZQ and PB treatments, each change appears earlier andmore severe in the PB-treated than PZQ-treated parasites. Aswell, the severity of tegumental damage increased when treatedwith higher concentration and longer incubation times. Further-more, at the same period of times and drug concentrations, thesurface of the male parasites appeared to be more damaged bythe drugs than that of females.
4. Discussion
This is the first report of the anthelmintic effect of PB on adult S.mansoni, which is more pronounced than PZQ as judged from sig-nificantly lower RM and SI values at the same drug concentrations.
Furthermore, we have demonstrated the different sensitivities tothese drugs by female and male parasites, and males were moreaffected than females. This result is consistent with a previousreport on PZQ effect in vitro, which found that male flukes frombisexual infection had a higher sensitivity to PZQ than females(Pica-Mattoccia and Cioli, 2004). However, in PB treatment maleand female flukes showed decreased RM values but within closerranges than PZQ. This could be because PB might have a muchstronger anthelmintic activity; hence it tended to affect both sexesquickly and almost equally.
The tegument of schistosomes is a vital target for antischistos-omal drugs due to its exposure to the fluid of hosts that receivedrug treatments. The present study is the first report that demon-strated the tegumental alterations in S. mansoni induced byplumbagin (PB). It is evident that PB exhibited a more rapid and se-vere effects on the tegument of adult S. mansoni while the se-quence of pathological changes is similar to that of PZQ,comprising of swelling of the tubercles and ridges, followed bybleb formation and loss of spines. Subsequently, the tuberclesand ridges were broken and began to erode, leading to the forma-tion of lesion which exposed the basal lamina. Eventually this re-sulted in sloughing off the tegument over large areas of the bodysurface. The severity of tegumental damage increased at higherdrug concentration and longer incubation times. The tegumentalchanges observed in this study are similar to that demonstratedin adult S. mansoni treated with praziquantel (Xiao et al., 2000),Schistosoma mekongi treated with artesunate (Jiraungkoorskul
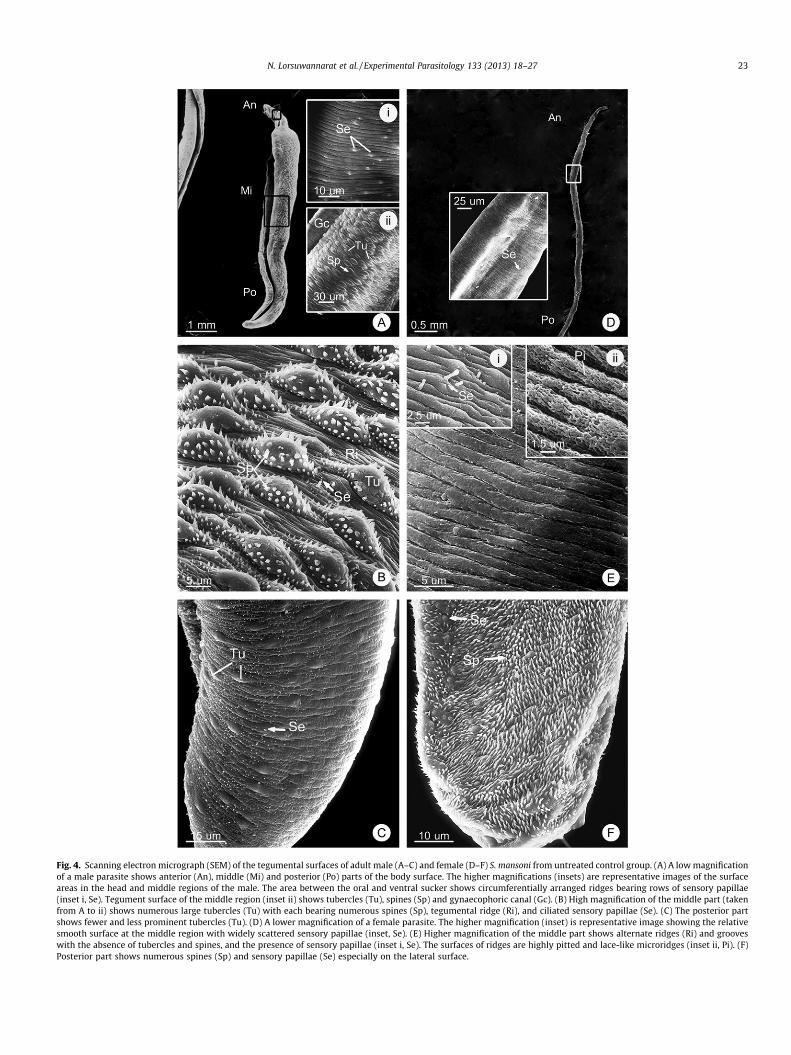
Fig. 4. Scanning electron micrograph (SEM) of the tegumental surfaces of adult male (A–C) and female (D–F) S. mansoni from untreated control group. (A) A low magnificationof a male parasite shows anterior (An), middle (Mi) and posterior (Po) parts of the body surface. The higher magnifications (insets) are representative images of the surfaceareas in the head and middle regions of the male. The area between the oral and ventral sucker shows circumferentially arranged ridges bearing rows of sensory papillae(inset i, Se). Tegument surface of the middle region (inset ii) shows tubercles (Tu), spines (Sp) and gynaecophoric canal (Gc). (B) High magnification of the middle part (takenfrom A to ii) shows numerous large tubercles (Tu) with each bearing numerous spines (Sp), tegumental ridge (Ri), and ciliated sensory papillae (Se). (C) The posterior partshows fewer and less prominent tubercles (Tu). (D) A lower magnification of a female parasite. The higher magnification (inset) is representative image showing the relativesmooth surface at the middle region with widely scattered sensory papillae (inset, Se). (E) Higher magnification of the middle part shows alternate ridges (Ri) and grooveswith the absence of tubercles and spines, and the presence of sensory papillae (inset i, Se). The surfaces of ridges are highly pitted and lace-like microridges (inset ii, Pi). (F)Posterior part shows numerous spines (Sp) and sensory papillae (Se) especially on the lateral surface.
N. Lorsuwannarat et al. / Experimental Parasitology 133 (2013) 18–27 23
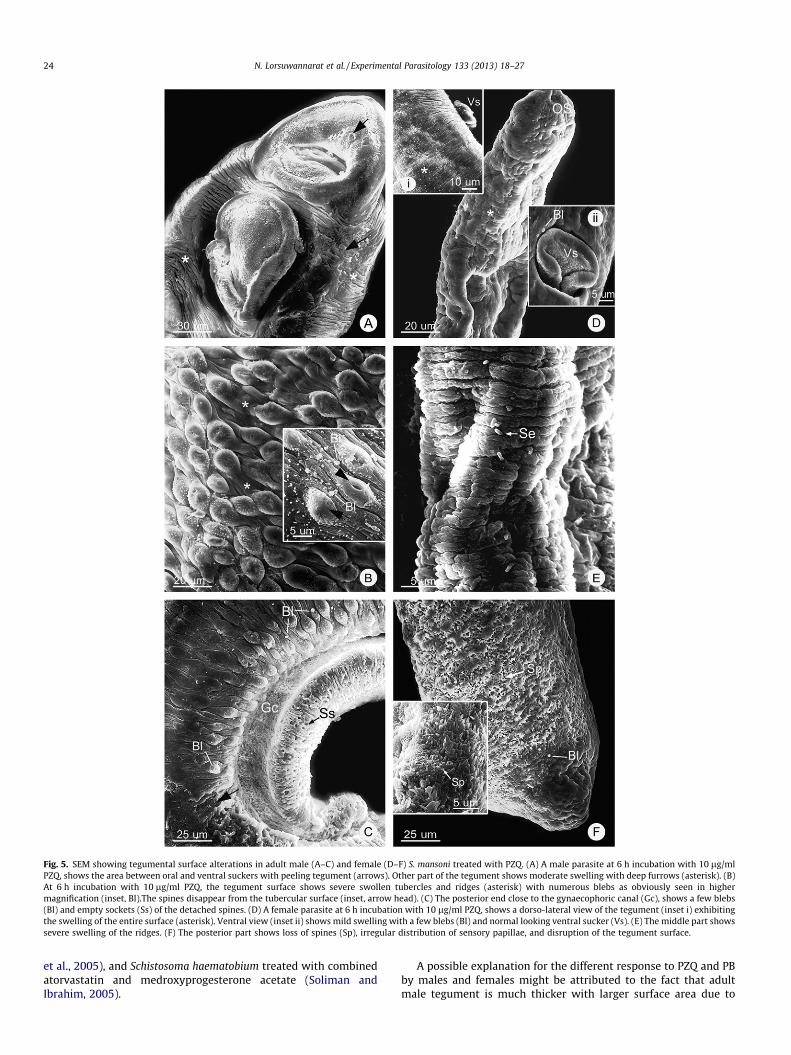
Fig. 5. SEM showing tegumental surface alterations in adult male (A–C) and female (D–F) S. mansoni treated with PZQ. (A) A male parasite at 6 h incubation with 10 lg/mlPZQ, shows the area between oral and ventral suckers with peeling tegument (arrows). Other part of the tegument shows moderate swelling with deep furrows (asterisk). (B)At 6 h incubation with 10 lg/ml PZQ, the tegument surface shows severe swollen tubercles and ridges (asterisk) with numerous blebs as obviously seen in highermagnification (inset, Bl).The spines disappear from the tubercular surface (inset, arrow head). (C) The posterior end close to the gynaecophoric canal (Gc), shows a few blebs(Bl) and empty sockets (Ss) of the detached spines. (D) A female parasite at 6 h incubation with 10 lg/ml PZQ, shows a dorso-lateral view of the tegument (inset i) exhibitingthe swelling of the entire surface (asterisk). Ventral view (inset ii) shows mild swelling with a few blebs (Bl) and normal looking ventral sucker (Vs). (E) The middle part showssevere swelling of the ridges. (F) The posterior part shows loss of spines (Sp), irregular distribution of sensory papillae, and disruption of the tegument surface.
24 N. Lorsuwannarat et al. / Experimental Parasitology 133 (2013) 18–27
et al., 2005), and Schistosoma haematobium treated with combinedatorvastatin and medroxyprogesterone acetate (Soliman andIbrahim, 2005).
A possible explanation for the different response to PZQ and PBby males and females might be attributed to the fact that adultmale tegument is much thicker with larger surface area due to
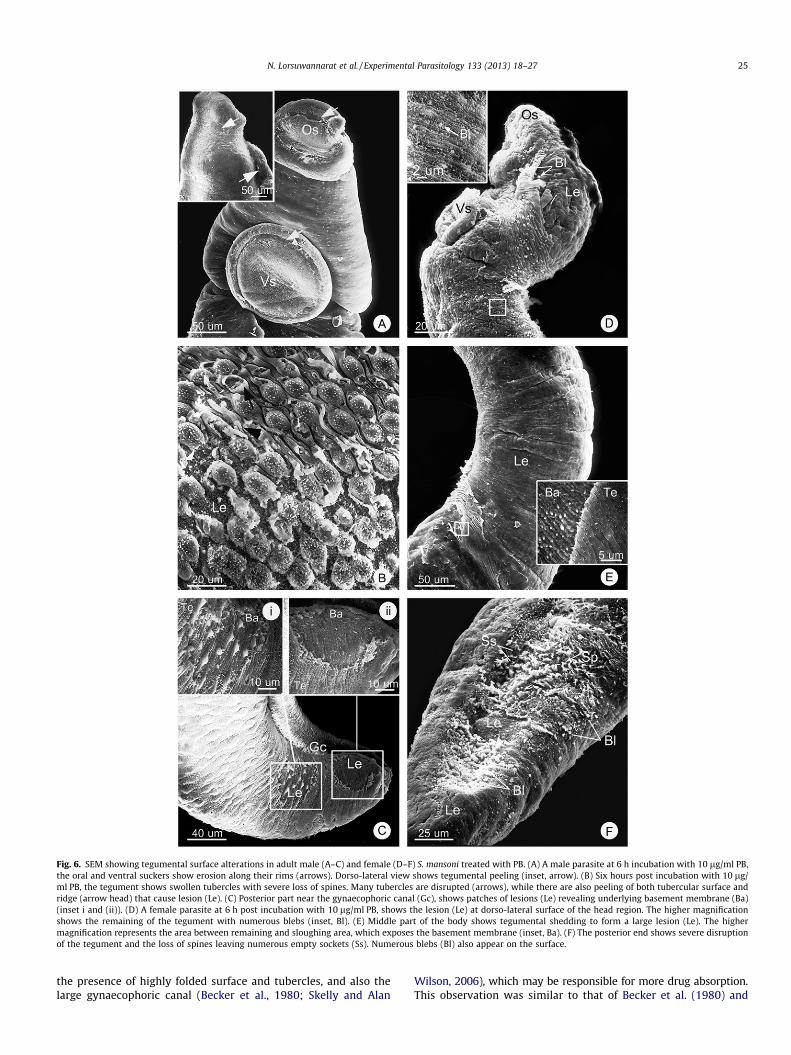
Fig. 6. SEM showing tegumental surface alterations in adult male (A–C) and female (D–F) S. mansoni treated with PB. (A) A male parasite at 6 h incubation with 10 lg/ml PB,the oral and ventral suckers show erosion along their rims (arrows). Dorso-lateral view shows tegumental peeling (inset, arrow). (B) Six hours post incubation with 10 lg/ml PB, the tegument shows swollen tubercles with severe loss of spines. Many tubercles are disrupted (arrows), while there are also peeling of both tubercular surface andridge (arrow head) that cause lesion (Le). (C) Posterior part near the gynaecophoric canal (Gc), shows patches of lesions (Le) revealing underlying basement membrane (Ba)(inset i and (ii)). (D) A female parasite at 6 h post incubation with 10 lg/ml PB, shows the lesion (Le) at dorso-lateral surface of the head region. The higher magnificationshows the remaining of the tegument with numerous blebs (inset, Bl). (E) Middle part of the body shows tegumental shedding to form a large lesion (Le). The highermagnification represents the area between remaining and sloughing area, which exposes the basement membrane (inset, Ba). (F) The posterior end shows severe disruptionof the tegument and the loss of spines leaving numerous empty sockets (Ss). Numerous blebs (Bl) also appear on the surface.
N. Lorsuwannarat et al. / Experimental Parasitology 133 (2013) 18–27 25
the presence of highly folded surface and tubercles, and also thelarge gynaecophoric canal (Becker et al., 1980; Skelly and Alan
Wilson, 2006), which may be responsible for more drug absorption.This observation was similar to that of Becker et al. (1980) and

26 N. Lorsuwannarat et al. / Experimental Parasitology 133 (2013) 18–27
Pica-Mattoccia and Cioli (2004), who suggested that the lower drugsensitivity in female is due to intrinsic physiological property and/or differences in tegument morphology between the two sexes.
Presently, PZQ is the recommended drug for treatments of alltypes of schistosomes. This drug affects the permeability of thetegument surface membrane by increasing calcium influx, causingimpairment of the tegument structure and function, and severespasm and paralysis of the underlying muscles, that eventuallyresult in death of the parasites (Becker et al., 1980; Pax et al.,1978; Shaw and Erasmus, 1983; Sobhon and Upatham, 1990).The efficacy of PZQ is dependent on many factors such as the timeof infection, sexes of the worms, their paired or unpaired condition,and the degree of drug resistance (Cioli and Pica-Mattoccia, 2003;Gryseels et al., 2001; Jiraungkoorskul et al., 2005).
Plumbagin (PB), derived from plants in Plumbagineae and Dro-seraceae families, was reported long ago to have medicinal proper-ties in treating dyspepsia, piles, diarrhea, tuberculosis, leprosy andskin diseases (Nadkarni and Nadkarni, 1954; Satyavati et al., 1976;Wuttidhammaved, 1997). More recent investigations indicatedthat PB has a broad range of biological activities, including beingantioxidant (Talcott et al., 1985), anticancer (Krishnaswamy andPurushothaman, 1980), anti-inflammatory (Reese et al., 2010),antifungal (Dzoyem et al., 2007), and antibacterial properties(Sharma et al., 2009). These activities arise mainly from its effectson multiple signalling, its redox reaction generating reactiveoxygen species (ROS), and its ability to chelate with trace metalspresent in biological system (Padhye et al., 2010). However, infor-mation on its mechanism of schistosomicidal activity is still notunderstood.
Many adult helminthes living in human hosts cannot obtain en-ough oxygen, and thus develop a special cellular respiration byemploying the NADH-fumarate reductase system for anaerobic en-ergy metabolism, and using rhodoquinone instead of ubiquinone(CoQ) (Masuma et al., 2009; Shiomi et al., 2007). However, schisto-somes are believed to be one of few parasitic helminths that pro-duce at least one-third of their energy by aerobic process, usingCoQ as electron carriers between respiratory complexes (Kitaet al., 2002; Masuma et al., 2009; van Oordt et al., 1985). Therefore,it is possible that PB schistosomicidal action is linked to its abilityto compete with CoQ in electron transport chain. It has been shownthat hydroxy-naphthoquinones compounds, including PB, can actas CoQ antagonists by being electrons acceptor in competion withCoQ in the electron transport chain (Babula et al., 2007; Srinivaset al., 2004).
There is also a possibility that PB antischistosome effect may berelated to its chemical structure as 5-hydroxy-2-methyl-1,4-naph-thoquinone, which is similar to atovaquone (2-(trans-4-(4-chloro-phenyl)-cyclohexyl)-3-hydroxy-1,4-naphthoquinone), a drug usedfor treating pneumocystis pneumonia, toxoplasmosis, and malaria(Davies et al., 1993). As Fry and Pudney (1992) found that atovaqu-one is a mitochondria-targeted antimalaria that acts as a specificinhibitor of the mitochondrial enzyme in the electron transportchain by competitive binding to the ubiquinol oxidation site ofthe mitochondrial cytochrome bc1 complex (complex III). (Hudsonet al., 1985; Kessl et al., 2007; Trumpower and Haggerty, 1980). Inaddition, PB could inhibit other mitochondrial enzymes includingdihydroorotate dehydrogenase (DHO-DH), succinate dehydroge-nase (complex II) and NADH oxidase as reported in the case of ma-laria and yeast cells, which in turn inhibit the normal electronstransport process (Fry and Pudney, 1992; Roberts et al., 1978).Consequently, the drastic decrease in ATP production may resultin the death of the flukes (Rohrer et al., 1986).
The tegument and associated structures (e.g. tegument cells) ofadult S. mansoni contain numerous mitochondria, which are com-monly distributed throughout the entire tegument (Hockley andMcLaren, 1973; Morris and Threadgold, 1968), and thus could be
the major target of PB. The disruption of regular ATP productionin these mitochondria might affect the energy-dependent Na+/K+-ATPase ion pumps at the apical plasma membrane, resulting inthe defective osmoregulartory system (Pax et al., 1987). Thus, largeinflux of Na+ and water into the syncytium due to the osmoticimbalance could lead to the swelling and blebbing, followed laterby disruption, erosion and lesion of the tegument. Similar changeshave been observed in adult Fasciola hepatica after treatment withclorsulon (Meaney et al., 2004), and in 3-week-old juvenile Fasciolagigantica after treatment with artesunate (Tansatit et al., 2012).
Alternatively, there has been a suggestion that PB and its ana-logs may cause cytotoxicity through semiquinone radical produc-tion and ROS generation. The increasing of ROS and H2O2
subsequently deplete glutathione reserve by oxidizing GSH toGSSG, thus affecting the parasites’ ability to protect themselvesfrom free radicals (Brophy and Barrett, 1990; McTigue et al.,1995; Padhye et al., 2010). Consequently, the depletion of GSH re-serve might be responsible for their death from PB treatment. Fur-ther study on the efficiencies of PB treatment in vivo, as well aspossible side effects to hosts are being investigated, as the data ob-tained in this study indicate a high therapeutic potential of PB asanother antischistosome drug.
Acknowledgments
This research was supported by Grants from Faculty of Science,Mahidol University to Prasert Sobhon, and the Thailand ResearchFund-Mahidol RGJ Ph.D. Scholarship to Natcha Lorsuwannarat.We would also like to thank Supatee Kitkha, Pichai Jirawattana-pong and Center of Nanoimaging (CNI) for their technicalassistances.
References
Atjanasuppat, K., Wongkham, W., Meepowpan, P., Kittakoop, P., Sobhon, P., Bartlett,A., Whitfield, P.J., 2009. In vitro screening for anthelmintic and antitumouractivity of ethnomedicinal plants from Thailand. J. Ethnopharmacol. 123, 475–482.
Babula, P., Adam, V., Havel, L., Kizek, R., 2007. Naphthoquinones and theirpharmacological properties. Ceská a Slovenská farmacie: casopis Ceskéfarmaceutické spolecnosti a Slovenské farmaceutické spolecnosti 56, 114–120.
Becker, B., Mehlhorn, H., Andrews, P., Thomas, H., Eckert, J., 1980. Light and electronmicroscopic studies on the effect of praziquantel on Schistosoma mansoni,Dicrocoelium dendriticum, and Fasciola hepatica (Trematoda) in vitro. Z.Parasitenkd. 63, 113–128.
Brophy, P.M., Barrett, J., 1990. Glutathione transferase in helminths. Parasitology100, 345–349.
Chitsulo, L., Engels, D., Montresor, A., Savioli, L., 2000. The global status ofschistosomiasis and its control. Acta Trop. 77, 41–51.
Cioli, D., Pica-Mattoccia, L., 2003. Praziquantel. Parasitol. Res. 90 (Suppl. 1), S3–S9.Davies, C.S., Pudney, M., Nicholas, J.C., Sinden, R.E., 1993. The novel
hydroxynaphthoquinone 566C80 inhibits the development of liver stages ofPlasmodium berghei cultured in vitro. Parasitology 106 (Pt 1), 1–6.
Dzoyem, J.P., Tangmouo, J.G., Lontsi, D., Etoa, F.X., Lohoue, P.J., 2007. In vitroantifungal activity of extract and plumbagin from the stem bark of Diospyroscrassiflora Hiern (Ebenaceae). Phytother. Res. 21, 671–674.
Fallon, P.G., Doenhoff, M.J., 1994. Drug-resistant schistosomiasis: resistance topraziquantel and oxamniquine induced in Schistosoma mansoni in mice is drugspecific. Am. J. Trop. Med. Hyg. 51, 83–88.
Fry, M., Pudney, M., 1992. Site of action of the antimalarialhydroxynaphthoquinone, 2-[trans-4-(40-chlorophenyl) cyclohexyl]-3-hydroxy-1,4-naphthoquinone (566C80). Biochem. Pharmacol. 43, 1545–1553.
Gryseels, B., Mbaye, A., De Vlas, S.J., Stelma, F.F., Guisse, F., Van Lieshout, L., Faye, D.,Diop, M., Ly, A., Tchuem-Tchuente, L.A., Engels, D., Polman, K., 2001. Are poorresponses to praziquantel for the treatment of Schistosoma mansoni infections inSenegal due to resistance? An overview of the evidence. Trop. Med. Int. Health6, 864–873.
Hockley, D.J., McLaren, D.J., 1973. Schistosoma mansoni: changes in the outermembrane of the tegument during development from cercaria to adult worm.Int. J. Parasitol. 3, 13–25.
Hudson, A.T., Randall, A.W., Fry, M., Ginger, C.D., Hill, B., Latter, V.S., McHardy, N.,Williams, R.B., 1985. Novel anti-malarial hydroxynaphthoquinones with potentbroad spectrum anti-protozoal activity. Parasitology 90 (Pt 1), 45–55.
Jiraungkoorskul, W., Sahaphong, S., Sobhon, P., Riengrojpitak, S., Kangwanrangsan,N., 2005. Effects of praziquantel and artesunate on the tegument of adultSchistosoma mekongi harboured in mice. Parasitol. Int. 54, 177–183.

N. Lorsuwannarat et al. / Experimental Parasitology 133 (2013) 18–27 27
Kessl, J.J., Moskalev, N.V., Gribble, G.W., Nasr, M., Meshnick, S.R., Trumpower, B.L.,2007. Parameters determining the relative efficacy of hydroxy-naphthoquinoneinhibitors of the cytochrome bc1 complex. Biochim. Biophys. Acta 1767, 319–326.
Kini, D.P., Pandey, S., Shenoy, B.D., Singh, U.V., Udupa, N., Umadevi, P., Kamath, R.,Nagarajkumari, , Ramanarayan, K., 1997. Antitumor and antifertility activities ofplumbagin controlled release formulations. Indian J. Exp. Biol. 35, 374–379.
Kita, K., Hirawake, H., Miyadera, H., Amino, H., Takeo, S., 2002. Role of complex II inanaerobic respiration of the parasite mitochondria from Ascaris suum andPlasmodium falciparum. Biochim. Biophys. Acta 1553, 123–139.
Kiuchi, F., Miyashita, N., Tsuda, Y., Kondo, K., Yoshimura, H., 1987. Studies on crudedrugs effective on visceral larva migrans. I. Identification of larvicidal principlesin betel nuts. Chem. Pharm. Bull. 35, 2880–2886.
Krishnaswamy, M., Purushothaman, K.K., 1980. Plumbagin: a study of its anticancer,antibacterial and antifungal properties. Indian J. Exp. Biol. 18, 876–877.
Masuma, R., Shiomi, K., Omura, S., 2009. Helminth electron transport inhibitorsproduced by fungi. In: Anke, T., Weber, D. (Eds.), Physiology and Genetics.Springer Berlin Heidelberg, pp. 247–271.
McTigue, M.A., Williams, D.R., Tainer, J.A., 1995. Crystal structures of a schistosomaldrug and vaccine target: glutathione S-transferase from Schistosoma japonicaand its complex with the leading antischistosomal drug praziquantel. J. Mol.Biol. 246, 21–27.
Meaney, M., Fairweather, I., Brennan, G.P., Forbes, A.B., 2004. Transmission electronmicroscope study of the ultrastructural changes induced in the tegument andgut of Fasciola hepatica following in vivo drug treatment with clorsulon.Parasitol. Res. 92, 232–241.
Morris, G.P., Threadgold, L.T., 1968. Ultrastructure of the tegument of adultSchistosoma mansoni. J. Parasitol. 54, 15–27.
Nadkarni, K.M., Nadkarni, A.K., 1954. Indian Materia Medica with Ayurvedic, Unani-Tibbi, Siddha, Allopathic, Homeopathic, Naturopathic & Home Remedies,Appendices & Indexes, third ed. Popular Book Depot, Bombay.
Neghina, R., Neghina, A.M., Merkler, C., Marincu, I., Moldovan, R., Iacobiciu, I., 2009.Intestinal schistosomiasis, importation of a neglected tropical disease inRomania: case report of a traveler to endemic regions. Travel Med. Infect. Dis.7, 49–51.
Padhye, S., Dandawate, P., Yusufi, M., Ahmad, A., Sarkar, F.H., 2010. Perspectives onmedicinal properties of plumbagin and its analogs. Med. Res. Rev..
Pax, R., Bennett, J.L., Fetterer, R., 1978. A benzodiazepine derivative andpraziquantel: effects on musculature of Schistosoma mansoni and Schistosomajaponicum. Naunyn-Schmiedebergs Arch. Pharmacol. 304, 309–315.
Pax, R.A., Chen, G.Z., Bennett, J.L., 1987. Schistosoma mansoni: measurement of Na+
ion activity in the tegument and the extracellular spaces using ion-selectivemicroelectrodes. Exp. Parasitol. 64, 219–227.
Pica-Mattoccia, L., Cioli, D., 2004. Sex- and stage-related sensitivity of Schistosomamansoni to in vivo and in vitro praziquantel treatment. Int. J. Parasitol. 34, 527–533.
Reese, S., Vidyasagar, A., Jacobson, L., Acun, Z., Esnault, S., Hullett, D., Malter, J.S.,Djamali, A., 2010. The Pin 1 inhibitor juglone attenuates kidney fibrogenesis viaPin 1-independent mechanisms in the unilateral ureteral occlusion model.Fibrogenesis Tissue Repair 3, 1.
Roberts, H., Choo, W.M., Smith, S.C., Marzuki, S., Linnane, A.W., Porter, T.H., Folkers,K., 1978. The site of inhibition of mitochondrial electron transfer by coenzyme Qanalogues. Arch. Biochem. Biophys. 191, 306–315.
Rohrer, S.P., Saz, H.J., Nowak, T., 1986. 31P NMR studies of the metabolisms of theparasitic helminths Ascaris suum and Fasciola hepatica. Arch. Biochem. Biophys.248, 200–209.
Satyavati, G.V., Raina, M.K., Sharma, M., Seshadri, T.R., Gupta, A.K., Tandon, N.,Dwarakanath, C., 1976. Medicinal Plants of India, vol. 1–2. Indian Council ofMedical Research, New Delhi.
Selma Ribeiro de Paiva, M.R.F., Aragão T.V., Kaplan, M.A.C., 2003. Antimicrobialactivity in vitro of plumbagin isolated from Plumbago species. 98, 959–961.
Sharma, S., Sharma, B.K., Prabhakar, Y.S., 2009. Juglone derivatives as antitubercularagents: a rationale for the activity profile. Eur. J. Med. Chem. 44, 2847–2853.
Shaw, M.K., Erasmus, D.A., 1983. Schistosoma mansoni: dose-related tegumentalsurface changes after in vivo treatment with praziquantel. Z. Parasitenkd. 69,643–653.
Shiomi, K., Hatano, H., Morimoto, H., Ui, H., Sakamoto, K., Kita, K., Tomoda, H., Lee,E.W., Heo, T.R., Kawagishi, H., Omura, S., 2007. Decursin and decursinol angelateselectively inhibit NADH-fumarate reductase of Ascaris suum. Planta Med. 73,1478–1481.
Skelly, P.J., Alan Wilson, R., 2006. Making sense of the schistosome surface. Adv.Parasitol. 63, 185–284.
Sobhon, P., Upatham, E.S., 1990. Snail Hosts, Life-Cycle, and Tegumental Structure ofOriental Schistosomes. UNDP/World Bank/WHO Special Programme forResearch and Training in Tropical Diseases, Bangkok, pp. 204–232.
Soliman, M.F., Ibrahim, M.M., 2005. Antischistosomal action of atorvastatin aloneand concurrently with medroxyprogesterone acetate on Schistosomahaematobium harboured in hamster: surface ultrastructure and parasitologicalstudy. Acta Trop. 93, 1–9.
Srinivas, P., Gopinath, G., Banerji, A., Dinakar, A., Srinivas, G., 2004. Plumbagininduces reactive oxygen species, which mediate apoptosis in human cervicalcancer cells. Mol. Carcinog. 40, 201–211.
Talcott, R.E., Smith, M.T., Giannini, D.D., 1985. Inhibition of microsomal lipidperoxidation by naphthoquinones: structure–activity relationships andpossible mechanisms of action. Arch. Biochem. Biophys. 241, 88–94.
Tansatit, T., Sahaphong, S., Riengrojpitak, S., Viyanant, V., Sobhon, P., 2012. Fasciolagigantica: the in vitro effects of artesunate as compared to triclabendazole onthe 3-weeks-old juvenile. Exp. Parasitol. 131, 8–19.
Trumpower, B.L., Haggerty, J.G., 1980. Inhibition of electron transfer in thecytochrome b–c, segment of the mitochondrial respiratory chain by asynthetic analogue of ubiquinone. J. Bioenerg. Biomembr. 12, 151–164.
van Oordt, B.E., van den Heuvel, J.M., Tielens, A.G., van den Bergh, S.G., 1985. Theenergy production of the adult Schistosoma mansoni is for a large part aerobic.Mol. Biochem. Parasitol. 16, 117–126.
World Health Organization, 2001. Strategy Development and Monitoring forParasitic Diseases and Vector Control Team. Report of the WHO InformalConsultation on Schistosomiasis in Low Transmission Areas: Control Strategiesand Criteria for Elimination, London, 10–13 April 2000. World HealthOrganization, Geneva, 51pp.
Wuttidhammaved, W., 1997. Thai Traditional Medicine, revised ed. O-dien StorePrinting, Bangkok, p. 339.
Xiao, S., Shen, B., Chollet, J., Utzinger, J., Tanner, M., 2000. Tegumental changes inadult Schistosoma mansoni harbored in mice treated with artemether. J.Parasitol. 86, 1125–1132.




















