
Chemometric analysis of biofluids from mice experimentally infected with Schistosoma mansoni
Upload
independentCategory
view
1download
0
A
hbSiwl(vt©
K
1
hiSciia
0d
Vaccine 25 (2007) 7217–7223
The Schistosoma bovis Sb14-3-3� recombinant protein cross-protectsagainst Schistosoma mansoni in BALB/c mice
M. Siles-Lucas a,1, N. Uribe a,b,1, J. Lopez-Aban a, B. Vicente a, A. Orfao c,J.J. Nogal-Ruiz d, A. San Feliciano e, A. Muro a,∗
a Laboratorio de Parasitologıa, Facultad de Farmacia, CIETUS, Universidad de Salamanca, Avda. Campo Charro, s/n. 37007 Salamanca, Spainb Universidad de Pamplona, Colombia
c Servicio de Citometrıa, Centro de Investigacion de Cancer, Universidad de Salamanca, Avda. Campo Charro, s/n. 37007 Salamanca, Spaind Departamento de Parasitologıa, Facultad de Farmacia, Universidad Complutense de Madrid, Avda de la Complutense, s/n. 28040 Madrid, Spain
e Departamento de Quımica Farmaceutica, CIETUS, Universidad de Salamanca, Avda. Campo Charro, s/n. 37007 Salamanca, Spain
Received 9 October 2006; received in revised form 14 May 2007; accepted 15 July 2007Available online 1 August 2007
bstract
Current control programs against schistosomiasis could be reinforced through the use of an effective vaccine. Schistosome 14-3-3 proteinsave been proposed as candidates for vaccine against the respective infections, and were seen to elicit high protection levels against Schistosomaovis in a previous work done by our group. We have therefore investigated the protective capacity of the 14-3-3 protein from S. bovis –b14� – against Schistosoma mansoni in mice. In addition, we have addressed the influence of the co-administration of three different
mmunomodulators with the 14-3-3 polypeptide. Protection was high when the Sb14� protein was combined in two independent experimentsith the AA2829 and PAL immunomodulatory molecules as regards both the reduction of worm numbers (mean: 64.8%) and egg loads in
iver (mean: 73.9%) or intestine (mean: 71.5%). In contrast, the degree of protection achieved with the Sb14�-CpG vaccine was very low
14.9% reduction in worm numbers, and 46.6% and 32% reduction in liver and intestinal egg loads). The immune responses observed in theaccinated animals showed that the production of IFN� and the absence of IL-4, accompanied by a strong humoral response, are insufficiento elicit protection against S. mansoni.2007 Elsevier Ltd. All rights reserved.
fTigc[a
eywords: Schistosoma; 14-3-3 vaccination; Immunomodulators
. Introduction
Schistosomiasis is a parasitic disease leading to chronic illealth, and it represents one of the major human health risksn certain sub-tropical and tropical areas where it is endemic.tudies on the close to sterile immunity elicited by irradiatedercariae (reviewed in [1]) and on the immune responses of
ndividuals living in endemic schistosomiasis areas, show-ng an association between antigen-specific immunity andlack of re-infection (reviewed in [2]), offer a paradigm
∗ Corresponding author. Tel.: +34 923294535; fax: +34 923294515.E-mail address: [email protected] (A. Muro).
1 These authors contributed equally to this work.
ubci
co
264-410X/$ – see front matter © 2007 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2007.07.021
or the development of a vaccine against schistosomiasis.hus, several antigens have been characterized and assayed
n vaccination trials. Nevertheless, the use of different sin-le parasite proteins for vaccination has failed to evoke theritical responses required for optimal vaccine efficacy (e.g.,3]). The failure to elicit the best protection levels could bettributed, among other reasons, to both the type of moleculesed for vaccination and the immune responses triggeredy respective antigens in specific combination with otheromponents of the vaccine, including vehicle, adjuvants and
mmunomodulators.It is well known that schistosomes have evolved in aomplex host–parasite interaction system, developing theirwn, intricate strategies to escape host immune responses.

7 accine
ItrlTtb(ta012fnpomw[af
aSicvi
2
2
(rawttSmST1aifegtttT
vtTaiitdbDmmu((wierwompTtme
wu
ucGPo
2
Iwre
cwQofAo
218 M. Siles-Lucas et al. / V
n this regard, the identification and characterization of pro-eins expressed in a stage-specific manner and playing a keyole during the life cycle of schistosomes may provide usefuleads for the development of novel intervention strategies.he 14-3-3 proteins are stage-specifically expressed, essen-
ial molecules in higher eukaryotes [4]. These molecules haveeen identified in Schistosoma mansoni and S. japonicumreviewed in [5]). The corresponding recombinant polypep-ides elicit highly variable and sub-optimal protection ratesgainst human schistosomes (27% in BALB/c mice and–45% in CBA/ca mice after intramuscular injection of a4-3-3 recombinant plasmid against S. japonicum [6,7], and5–45% in C57Bl/6J mice after subcutaneous injection of aused GST-14-3-3 protein against S. mansoni [8]) in vacci-ation trials in mice. On the other hand, the S. bovis 14-3-3rotein, recently described by us [9] and representing therthologous molecule from the above-mentioned 14-3-3 S.ansoni protein, gave rise to high protection levels in micehen combined with specific immunomodulatory molecules
10]. The insight thus gained should be useful not only forntigen selection but also for vaccine formulation prior tourther vaccination trials against human schistosomes.
Here, we investigated the protection rates achievedgainst S. mansoni in mice after vaccination with theb14� recombinant antigen combined with three different
mmunomodulatory molecules. In addition, we discuss andompare the humoral and cell immune responses elicited afteraccination with the three different vaccine formulations usedn our experiments.
. Materials and methods
.1. Antigens and immunomodulators
Soluble somatic extracts from adult S. mansoni wormsSOSm) were obtained as previously described [11]. Theecombinant S. bovis 14-3-3� protein (Sb14�) was obtaineds described in [9]. Briefly, total RNA from adult wormsas extracted using the RNeasy kit (Quiagen) according to
he manufacturer’s instructions. First-strand cDNA was syn-hesized from S. bovis adult worms RNA using the Firsttrand cDNA Synthesis kit (Roche) as recommended by theanufacturers. cDNA was then amplified with two primers,bFwd (5′-ATGACTACGTCGTGGGTTTT) and SbRev (5′-TAGCCGTCATTTTCAACAT), designed on the S. bovis4-3-3� cDNA sequence (accession number AY615728) andmplifying the whole coding sequence for the correspond-ng protein in a PCR of 30 cycles of 94 ◦C for 40 s, 52 ◦Cor 40 s and 72 ◦C for 1 min. The PCR product was thenlectrophoresed in Tris-borate–EDTA (TBE) 1% agaroseels and visualized with ethidium bromide under a UV-
ransilluminator. The corresponding band was excised fromhe gel and purified with the Agarose Gel DNA Extrac-ion kit (Roche), following the manufacturer’s instructions.he purified PCR product was subcloned into the TOPOcTts
25 (2007) 7217–7223
ector (TOPO Cloning kit, Invitrogen) and used for theransformation of DH5� Escherichia coli cells (Invitrogen).ransformed cells were grown on LB-ampicillin (50 �g/ml)gar plates overnight at 37 ◦C. A selected colony was grownn LB-ampicillin overnight at 37 ◦C with shaking. The result-ng pelleted cells were subjected to plasmid extraction withhe NucleoSpin Plasmid kit (Macherey-Nagel). After EcoRIigestion and agarose gel purification of the correspondingand with the Agarose Gel DNA Extraction kit (Roche), theNA was ligated into the EcoRI-digested pGEX4T1 plas-id (Stratagene) with T4 DNA ligase (Roche), following theanufacturers’ instructions. The ligation reaction was then
sed for the transformation of BL21 Codon Plus E. coli cellsStratagene). Transformed cells were grown in LB-ampicillin50 �g/ml) agar plates overnight at 37 ◦C. A selected colonyas grown in LB-ampicillin overnight at 37 ◦C with shak-
ng, and the resulting cell pellet was subjected to plasmidxtraction (NucleoSpin Plasmid kit, Macherey-Nagel). Theecombinant plasmid obtained was automatically sequencedith the pGEX forward primer (Stratagene) to assess thepen reading frame of the sequence. The same transfor-ant was then used for the production of the GST-Sb14�
rotein using IPTG induction, as described elsewhere [12].he fused protein was then affinity-purified and subjected
o thrombin digestion, as described elsewhere [12]. Assess-ent of the integrity of the Sb14� protein was performed by
lectrophoresis in polyacrylamide gels, as described [13].SOSm and Sb14� protein concentrations were measured
ith the BCA assay (Pierce). Proteins were stored at −80 ◦Cntil use.
During our experiments, three different immunomod-latory molecules were used: the commercial CpG-ontaining polynucleotide (ODN 1826 [14], 5′-TCCATGAC-TTCCTGACGTT; TIB Molbiol, Germany); an extract ofolypodium leucotomos (PAL; [15]), and the syntheticallybtained amino alcohol AA2829 [16].
.2. Vaccination experiments
One hundred female BALB/c mice (8–10 weeks old,FFACredo, Spain) were used for the experiments. All miceere housed and handled under standard aseptic animal labo-
atory conditions according to the EU regulations for animalxperimentation.
Our vaccine formulation includes four components: thearrier (Montanide® ISA763A-SEPPIC, Paris, France, plusater in a 70/30 emulsion), the adjuvant (saponin from. saponaria QuilA; Sigma), one immunomodulator (PALr AA2829) and the recombinant Sb14� protein. Thisormulation, previously assayed by us [10,15], is calledDaptation + ADjuvant (ADAD) and is administered in a setf two subcutaneous injections. The first, called “adaptation”,
ontains all the components but the recombinant protein.he second injection, administered 5 days after the adapta-ion, contains all the vaccine components. The first injectionhould stimulate and “guide” the immune system, preparing

accine
i[uttCl�PrbFdhpCf[
faGaiiippii
tcAtooitfdwsi
m
2
Tpsepo
SaadaIcs
I(dFcTs
2
nswtewEMoa
2
attambaTs
3
3
iei
M. Siles-Lucas et al. / V
t for the subsequent stimulus with parasite-specific antigens10,15]. A third immunomodulator – CpG ODN 1826 – wassed with the same vaccine formulation, but administration ofhe “adaptation” was not implemented, following its adminis-ration schedule performed by other authors (e.g., [14]). ThepG ODN 1826 should activate an innate immune response,
eading to non-specific secretion of Th1-type cytokines (IFN-and IL-12) [14]. PAL, consisting of an extract from the plant. leucotomos, has been shown to down-regulate the Th2-likeesponse induced by parasitic infections such as those causedy Trichinella spiralis [17] and Trichomonas vaginalis [18].inally, the aminoalcohol AA2829, described as a syntheticerivative from the immunomodulatory compound myriocin,as previously been tested as regards its immunomodulatoryroperties [16], and it has been shown to increase T helperD4+ and CD8+ cell numbers and nitric oxide production
rom mouse immunocompetent cells after in vitro stimulation16].
The animals were divided for two independently per-ormed experiments as follows: for experiment 1, thenimals were divided into group 1 (G1, control, n = 5),2 (infected, n = 10), G3 (Sb14� plus AA2829-immunized
nd infected, n = 10), G4 (Sb14� plus PAL-immunized andnfected, n = 10) and G5 (Sb14� plus CpG-immunized andnfected, n = 10). For experiment 2, the mice were dividednto G1 (control, n = 10), G2 (infected, n = 10), G3 (Sb14�lus AA2829-immunized and infected, n = 10), G4 (Sb14�lus PAL-immunized and infected, n = 10), G6 (AA2829-mmunized and infected, n = 5), G7 (PAL-immunized andnfected, n = 5) and G8 (CpG-immunized and infected, n = 5).
Immunizations were carried out subcutaneously withwo doses of 20 �g of QuilA (Sigma) and 25 �g of CpG-ontaining polynucleotide, 600 �g of PAL or 100 �g ofA2829 in Montanide (SEPPI, France; 70% in aqueous solu-
ion), plus 10 �g Sb14� recombinant protein when indicated,r left without immunization (G1 and G2 groups). The sec-nd immunization was carried out 2 weeks after the first. Eachmmunization was preceded by a preimmunization with allhe vaccine components but the recombinant protein, exceptor the groups receiving the CpG immunomodulator. Theoses of immunomodulators were chosen based on previousorks conducted by us and other authors [10,14,16]. The cho-
en recombinant protein dose was sufficient to elicit strongmmune responses (see below).
Infections were performed precutaneously with 150 S.ansoni cercariae, as previously described [19].
.3. Measurement of antibody and cytokine responses
Blood was taken from each animal on days 0, 28 and 70.he mice were sacrificed on day 70 (42 days post-infection,.i.) and spleen cells were recovered. For antibody mea-
urements in serum, the ELISA technique was performedssentially as described elsewhere [12]. Briefly, 96-wellolystyrene plates (Costar) were coated with 0.2 �g Sb14�r 5 �g of S. mansoni adult worm somatic antigens (SOSm).(i(g
25 (2007) 7217–7223 7219
era were then added at 1:100 dilutions, followed by theddition of peroxidase-labelled anti-mouse IgG1 or IgG2antibodies at 1:2000 dilution (Sigma). The reaction waseveloped with H2O2 and orthophenylenediamine (Sigma)nd measured at 492 on an Ear400FT ELISA reader (Labnstruments). The results are expressed as means of the opti-al density from all the animals of each group plus thetandard error.
Cell cytokine production (TNF�, IFN�, IL-2, IL-4 andL-5) was measured after in vitro stimulation of spleen cellspooled for each group; 1.5 × 105 cells/well), prepared asescribed in [20], with 2.5 �g of Sb14� for 6 h at 37 ◦C in aACSCalibur cytometer (BDB) with the BA Mouse Th1/Th2ytokine kit (BD), following the manufacturers’ instructions.he results are expressed as means of two independent mea-urements for each group.
.4. Estimation of parasite load
Parasite load was evaluated in each group as follows:ecropsied mice were perfused by intra-cardiac injection ofaline citrate, and the number of recovered S. mansoni adultorms flowing out the portal vein was recorded. In addi-
ion, the number of parasite eggs in liver and intestine wasstimated after digestion of the previously weighed organsith 30 ml 5% KOH (16 h at 37 ◦C with gentle shaking).gg counts were performed under a magnifying glass in acMaster chamber, calculating the number of eggs per gram
f liver and intestine in each animal. The results are expresseds means ± standard deviation.
.5. Statistical analysis
The overall differences among groups were compared bynalysis of variance (ANOVA) with one coefficient of varia-ion. When overall differences were detected, a post-ANOVAest, Fischer least significant differences (LSD) analysis, waspplied. Results of worm and egg recovery were reported asean ± S.D., and results of absorbances from specific anti-
odies in ELISA were reported as mean + S.E. All statisticalnalyses were considered significant at the p < 0.05 level.he analyses were performed using StarWorks and Statviewoftware packages for a Macintosh computer.
. Results
.1. Protection levels
Comparison of parasite loads between only infected andmmunized and infected mice revealed significant differ-nces between the mean S. mansoni worm and egg numbersn the Sb14�-AA2829- and Sb14�-PAL-vaccinated animals
Tables 1 and 2), resulting in a reduction of 62.2% and 67.1%n worm numbers, and of 75.4% and 70.1% in egg loadsmean values from two experiments) in the two vaccinatedroups, respectively.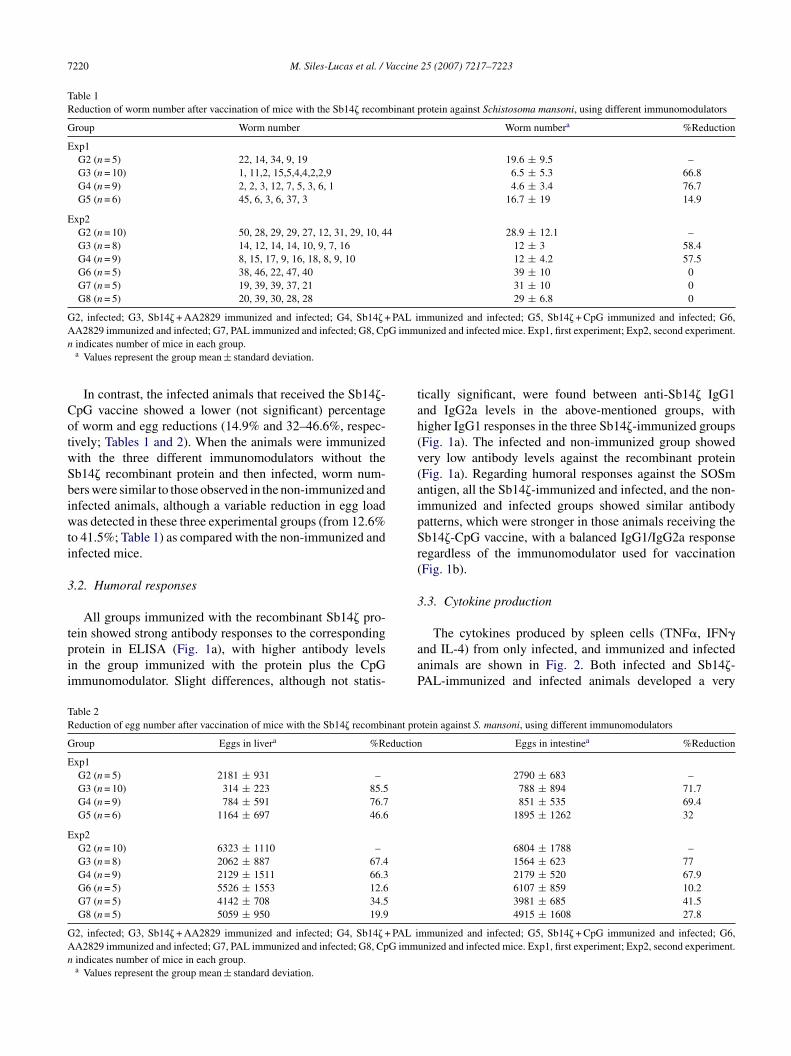
7220 M. Siles-Lucas et al. / Vaccine 25 (2007) 7217–7223
Table 1Reduction of worm number after vaccination of mice with the Sb14� recombinant protein against Schistosoma mansoni, using different immunomodulators
Group Worm number Worm numbera %Reduction
Exp1G2 (n = 5) 22, 14, 34, 9, 19 19.6 ± 9.5 –G3 (n = 10) 1, 11,2, 15,5,4,4,2,2,9 6.5 ± 5.3 66.8G4 (n = 9) 2, 2, 3, 12, 7, 5, 3, 6, 1 4.6 ± 3.4 76.7G5 (n = 6) 45, 6, 3, 6, 37, 3 16.7 ± 19 14.9
Exp2G2 (n = 10) 50, 28, 29, 29, 27, 12, 31, 29, 10, 44 28.9 ± 12.1 –G3 (n = 8) 14, 12, 14, 14, 10, 9, 7, 16 12 ± 3 58.4G4 (n = 9) 8, 15, 17, 9, 16, 18, 8, 9, 10 12 ± 4.2 57.5G6 (n = 5) 38, 46, 22, 47, 40 39 ± 10 0G7 (n = 5) 19, 39, 39, 37, 21 31 ± 10 0G8 (n = 5) 20, 39, 30, 28, 28 29 ± 6.8 0
G2, infected; G3, Sb14� + AA2829 immunized and infected; G4, Sb14� + PAL immunized and infected; G5, Sb14� + CpG immunized and infected; G6,A G immn
CotwSbiwti
3
tpii
tah(v(aipSr(
3
TR
G
E
E
GAn
A2829 immunized and infected; G7, PAL immunized and infected; G8, Cpindicates number of mice in each group.a Values represent the group mean ± standard deviation.
In contrast, the infected animals that received the Sb14�-pG vaccine showed a lower (not significant) percentagef worm and egg reductions (14.9% and 32–46.6%, respec-ively; Tables 1 and 2). When the animals were immunizedith the three different immunomodulators without theb14� recombinant protein and then infected, worm num-ers were similar to those observed in the non-immunized andnfected animals, although a variable reduction in egg loadas detected in these three experimental groups (from 12.6%
o 41.5%; Table 1) as compared with the non-immunized andnfected mice.
.2. Humoral responses
All groups immunized with the recombinant Sb14� pro-
ein showed strong antibody responses to the correspondingrotein in ELISA (Fig. 1a), with higher antibody levelsn the group immunized with the protein plus the CpGmmunomodulator. Slight differences, although not statis-aaP
able 2eduction of egg number after vaccination of mice with the Sb14� recombinant pro
roup Eggs in livera %Reduction
xp1G2 (n = 5) 2181 ± 931 –G3 (n = 10) 314 ± 223 85.5G4 (n = 9) 784 ± 591 76.7G5 (n = 6) 1164 ± 697 46.6
xp2G2 (n = 10) 6323 ± 1110 –G3 (n = 8) 2062 ± 887 67.4G4 (n = 9) 2129 ± 1511 66.3G6 (n = 5) 5526 ± 1553 12.6G7 (n = 5) 4142 ± 708 34.5G8 (n = 5) 5059 ± 950 19.9
2, infected; G3, Sb14� + AA2829 immunized and infected; G4, Sb14� + PAL iA2829 immunized and infected; G7, PAL immunized and infected; G8, CpG immindicates number of mice in each group.a Values represent the group mean ± standard deviation.
unized and infected mice. Exp1, first experiment; Exp2, second experiment.
ically significant, were found between anti-Sb14� IgG1nd IgG2a levels in the above-mentioned groups, withigher IgG1 responses in the three Sb14�-immunized groupsFig. 1a). The infected and non-immunized group showedery low antibody levels against the recombinant proteinFig. 1a). Regarding humoral responses against the SOSmntigen, all the Sb14�-immunized and infected, and the non-mmunized and infected groups showed similar antibodyatterns, which were stronger in those animals receiving theb14�-CpG vaccine, with a balanced IgG1/IgG2a responseegardless of the immunomodulator used for vaccinationFig. 1b).
.3. Cytokine production
The cytokines produced by spleen cells (TNF�, IFN�nd IL-4) from only infected, and immunized and infectednimals are shown in Fig. 2. Both infected and Sb14�-AL-immunized and infected animals developed a very
tein against S. mansoni, using different immunomodulators
Eggs in intestinea %Reduction
2790 ± 683 –788 ± 894 71.7851 ± 535 69.4
1895 ± 1262 32
6804 ± 1788 –1564 ± 623 772179 ± 520 67.96107 ± 859 10.23981 ± 685 41.54915 ± 1608 27.8
mmunized and infected; G5, Sb14� + CpG immunized and infected; G6,unized and infected mice. Exp1, first experiment; Exp2, second experiment.

M. Siles-Lucas et al. / Vaccine
Fig. 1. Specific serum IgG1 (grey bars) and IgG2a (striped bars) reactivitiesmeasured by ELISA against (a) the Schistosoma bovis Sb14� recombinantprotein and (b) a somatic extract from S. mansoni, in (G2) infected, (G3)Sb14� + AA2829-immunized and -infected, (G4) Sb14� + PAL-immunizedand -infected and (G5) Sb14� + CpG-immunized and -infected mice. Barsrepresent the mean optical density (OD) value from all the animals of eachgroup plus the standard error.
Fig. 2. In vitro cytokine production of Sb14�-stimulated spleen cellsfrom (G2) infected, (G3) Sb14� + AA2829-immunized and -infected, (G4)Sb14� + PAL-immunized and -infected and (G5) Sb14� + CpG-immunizedabm
stl4paSmg
4
caei
ttohnttaTceb
tsgb[cpauhriibbtbpmbBmSwatcptsfmdprtsTb
nd -infected mice. TNF� (white bars), IFN� (grey bars) and IL-4 (stripedars). Bars represent the means of two independently performed experi-ents, except for group 5.
imilar cytokine response, with detectable levels of thehree above-mentioned molecules, although IL-4 levels wereower in the immunized group (p < 0.02; Fig. 2). No IL-
was detected in animals immunized with the proteinlus AA2829 or CpG. Regarding IFN� production in thebove-mentioned groups, this was not detectable in theb14�-AA2829-vaccinated and infected group, and reachedaximal levels in the Sb14�-CpG-immunized and infected
roup (Fig. 2).
. Discussion
Chronic schistosomiasis in human patients and animals,
aused by different species of the genus Schistosoma, is char-cterized by the formation of granulomas around parasiteggs, which become trapped in different organs: mainly liver,ntestine and bladder. The subsequent organ damage can leadubtw
25 (2007) 7217–7223 7221
o high morbidity rates in infected individuals. Praziquan-el treatment against schistosomes during chronic stages isnly effective against adult worms. Thus, enormous effortsave been made in vaccine development in order to elimi-ate schistosomes before egg production starts. Despite this,he results obtained have so far been disappointing [1] dueo the highly variable and often sub-optimal protection levelsttained using different parasite molecules for vaccination.hese discouraging results could be due to both the moleculeshosen and/or to the need to add other vaccine components –.g., adjuvants, immunomodulators or vehicles – to achieveetter protection levels.
Previous studies have demonstrated the potential pro-ective role that immunity against a specific antigen fromchistosomes – the 14-3-3 protein – could play. This anti-en has been described as one of the molecules recognizedy antibodies from irradiated cercariae-vaccinated animals21]. Furthermore, the use of homologous 14-3-3-based vac-ines against S. japonicum and S. mansoni afford varyingrotection levels in mice (from 0% to 45%; [6–8]), usingnaked DNA vaccine and the protein plus complete Fre-
nds’ adjuvant, respectively. We have recently shown that theomologous protein in S. bovis, the Sb14� protein [9], givesise to high protection levels against respective infectionsn mice (from 40% to 61%) when combined with specificmmunomodulatory molecules [10]. These differences coulde in part attributed to slight immunostimulatory differencesetween the 14-3-3 proteins from human and animal schis-osomes, and also to the immunomodulatory molecules usedy us. This protein has also been shown to be crucial forarasite survival in an Echinococcus multilocularis murineodel, in which vaccination with the corresponding recom-
inant protein resulted in high protection levels (97%, [22]).ased on such findings, we tackled the S. bovis 14-3-3olecule (Sb14�) as a potential vaccine candidate against
. mansoni. Three different immunomodulatory moleculesere studied: CpG, PAL and AA2827. The commercially
vailable CpG polynucleotides have been shown to exertheir immunomodulatory activity due to their un-methylatedharacter. CpG dinucleotide un-methylation is a frequenthenomenon in bacterial genomes, leading to its recogni-ion by the immune system of higher eukaryotes (e.g., [23]),timulating a Th1-type immune response. This was alsoound during our experiments, since Sb14�-CpG-vaccinatedice showed high IFN� levels and the absence of IL-4 pro-
uction. Sb14�-CpG-vaccinated animals also exhibited theroduction of TNF� and a balanced IgG1/IgG2a humoralesponse. Th1-type adjuvants have been shown to improvehe protection levels obtained with single proteins againstchistosomes (e.g., [24–26]). However, in our experimentsh1-type responses did not result in the best protection levelut in a very low reduction in the parasite load (14.9%). The
se of the same vaccine formulation against S. bovis has alsoeen seen to elicit very low protection levels [10]. Similar tohese findings, Zhang et al. [6] obtained low protection rateshen a recombinant plasmid containing the S. japonicum 14-
7 accine
3o–
maTsoottIptng
Poomt
oittnl
sdhncpl
A
tCt((R(
R
[
[
[
[
[
[
[
[
[
[
[
222 M. Siles-Lucas et al. / V
-3 coding sequence, among others, was used, either aloner in combination with IL-12 – a Th1-type response induceragainst S. japonicum in mice.In contrast, both the Sb14�-AA2829- and PAL-immunized
ice presented high protection rates (62–67%), despite thebsence of IFN� or the presence of IL-4, a cytokine-triggeringh2-type responses in the supernatant of Sb14�-stimulatedpleen cells from these animals. Similarly, we have previ-usly described that the use of the Sb14� together with PALr with the synthetic immunomodulator AA0029 – similaro AA2829 – to vaccinate against S. bovis in mice gives riseo protection levels comparable to those reported here [10].n this regard, several authors have suggested that an optimalrotective response against schistosomes could be achievedhrough a multifaceted immune response, in which a combi-ation of Th1 and Th2-type cytokines and the presence of aiven proportion of Treg cells is needed (e.g., [27]).
In addition, the protection levels in Sb14�-AA2829- andAL-immunized mice reported here are higher than thosebtained by other authors using the parasite 14-3-3 cDNAr the 14-3-3 recombinant protein against S. japonicum or S.ansoni in mice [6–8]. These differences could be attributed
o the use of different vaccine formulations.Regarding the differences in protection achieved during
ur experiments when either the CpG or the PAL and AA2829mmunomodulator was used could be due to the absence ofhe “adaptation” step in the CpG vaccination schedule. Thus,he adaptation step could induce early signals of innate immu-ity necessary for the achievement of optimal protectionevels, although this should be further investigated.
In summary, we have demonstrated that the use of apecific schistosome vaccine candidate in combination withifferent immunomodulators and the characterization of bothumoral and cell immune responses elicited by those combi-ations seem to be essential before ruling out the usefulness ofertain molecules. Additionally, we have shown that the bestrotection levels against these parasites are not necessarilyinked to a Th1-type immune response.
cknowledgements
MSL acknowledges financial support from the Minis-erio de Educacion y Ciencia (MEC), Spain (“Ramon yajal” contract). NU acknowledges financial support from
he Carolina Foundation (Spain) and University of PamplonaColombia). This work was supported by the MEC, SpainProjects AGL2002-00644 and AGL2005-02168) and theed de Investigacion de Centros en Enfermedades Tropicales
RICET, FIS, Spain).
eferences
[1] Hewitson JP, Hamblin PA, Mountford AP. Immunity inducedby the radiation-attenuated schistosome vaccine. Parasite Immunol2005;27(7–8):271–80.
[
25 (2007) 7217–7223
[2] Bergquist R, Al-Sherbiny M, Barakat R, Olds R. Blueprint for schisto-somiasis vaccine development. Acta Tropica 2002;82(2):183–92.
[3] Ganley-Leal LM, Guarner J, Todd CW, Da’Dara AA, Freeman Jr GL,Boyer AE, et al. Comparison of Schistosoma mansoni irradiated cer-cariae and Sm23 DNA vaccines. Parasite Immunol 2005;27(9):341–9.
[4] Aitken A. 14-3-3 proteins: a historic overview. Semin Cancer Biol2006;16(3):162–72.
[5] Siles-Lucas M, Gottstein B. The 14-3-3 protein: a key molecule inparasites as in other organisms. Trends Parasitol 2003;19(12):575–81.
[6] Zhang Y, Taylor MG, Johansen MV, Bickle QD. Vaccination of micewith a cocktail DNA vaccine induces a Th1-type immune response andpartial protection against Schistosoma japonicum infection. Vaccine2001;20(5–6):724–30.
[7] Li DF, Chen YS, Zu Y, Shen JL. Construction of a novel Schistosomajaponicum DNA vaccine pBK-Sj14-3-3 and studies on its immuno-protection in mice. Zhonghua Yu Fang Yi Xue Za Zhi 2004;38(3):193–5.
[8] Schechtman D, Tarrab-Hazdai R, Arnon R. The 14-3-3 proteinas a vaccine candidate against schistosomiasis. Parasite Immunol2001;23(4):213–7.
[9] Uribe N, Muro A, Vieira C, Lopez-Aban J, del Olmo E, Suarez L, etal. Genetic and immunological characterization of the 14-3-3� proteinfrom Schistosoma bovis. J Parasitol, in press.
10] Uribe N, Siles-Lucas M, Lopez-Aban J, Esteban A, Suarez L, Martinez-Fernandez A, et al. The Sb14-3-3� recombinant protein protects againstSchistosoma bovis in BALB/c mice. Vaccine 2007;25(23):4533–9.
11] Pardo J, Carranza C, Turrientes MC, Perez Arellano JL, Lopez VelezR, Ramajo V, et al. Utility of Schistosoma bovis adult worm antigensfor diagnosis of human schistosomiasis by enzyme-linked immunosor-bent assay and electroimmunotransfer blot techniques. Clin Diagn LabImmunol 2004;11(6):1165–70.
12] Siles-Lucas M, Felleisen RS, Hemphill A, Wilson W, Gottstein B.Stage-specific expression of the 14-3-3 gene in Echinococcus multi-locularis. Mol Biochem Parasitol 1998;91(2):281–93.
13] Siles-Lucas M, Nunes CP, Zaha A. Comparative analysis of the 14-3-3gene and its expression in Echinococcus granulosus and Echinococcusmultilocularis metacestodes. Parasitology 2001;122(Pt 3):281–7.
14] Kumar S, Jones TR, Oakley MS, Zheng H, Kuppusamy SP, Taye A, etal. CpG oligodeoxynucleotide and Montanide ISA 51 adjuvant com-bination enhanced the protective efficacy of a subunit malaria vaccine.Infect Immun 2004;72:949–57.
15] Martinez-Fernandez AR, Nogal-Ruiz JJ, Lopez-Aban J, RamajoV, Oleaga A, Manga-Gonzalez Y, et al. Vaccination of miceand sheep with Fh12 FABP from Fasciola hepatica usingthe new adjuvant/immunomodulator system ADAD. Vet Parasitol2004;126(3):287–98.
16] Olmo ED, Plaza A, Muro A, Martinez-Fernandez AR, Nogal-Ruiz JJ,Lopez-Perez JL, et al. Synthesis and evaluation of some lipidic aminoal-cohols and diamines as immunomodulators. Bioorg Med Chem Lett2006;16(23):6091–5.
17] Dea-Ayuela M, Rodero M, Rodriguez-Bueno R, Bolas-Fernandez F,Martinez-Fernandez AR. Modulation by Anapsos (Polypodium leuco-tomos extract) of the antibody responses against the nematode parasiteTrichinella spiralis. Phytother Res 1999;13:566–70.
18] Nogal-Ruiz JJ, Gomez-Barrio A, Escario JA, Martinez-Fernandez AR.Effect of Anapsos in a murine model of experimental trichomoniasis.Parasite 2003;10:303–8.
19] Abane JL, Oleaga A, Ramajo V, Casanueva P, Arellano JL, Hillyer GV,et al. Vaccination of mice against Schistosoma bovis with a recombi-nant fatty acid binding protein from Fasciola hepatica. Vet Parasitol2000;91(1–2):33–42.
20] Bosze S, Caccamo N, Majer Z, Mezo G, Dieli F, Hudecz F. In vitro T-
cell immunogenicity of oligopeptides derived from the region 92–110of the 16-kDa protein of Mycobacterium tuberculosis. Biopolymers2004;76:467–76.21] Zhang Y, Taylor MG, McCrossan MV, Bickle QD. Molecularcloning and characterization of a novel Schistosoma japonicum “irra-

accine
[
[
[
[
[
M. Siles-Lucas et al. / V
diated vaccine-specific” antigen, Sj14-3-3. Mol Biochem Parasitol1999;103(1):25–34.
22] Siles-Lucas M, Merli M, Mackenstedt U, Gottstein B. The Echinococ-cus multilocularis 14-3-3 protein protects mice against primarybut not secondary alveolar echinococcosis. Vaccine 2003;21(5–6):431–9.
23] Krieg AM, Love-Homan L, Yi AK, Harty JT. CpG DNA inducessustained IL-12 expression in vivo and resistance to Listeria mono-
cytogenes challenge. J Immunol 1998;161(5):2428–34.24] Da’Dara AA, Skelly PJ, Walker CM, Harn DA. A DNA-prime/protein-boost vaccination regimen enhances Th2 immune responses but notprotection following Schistosoma mansoni infection. Parasite Immunol2003;25(8–9):429–37.
[
25 (2007) 7217–7223 7223
25] Fonseca CT, Brito CF, Alves JB, Oliveira SC. IL-12 enhancesprotective immunity in mice engendered by immunization withrecombinant 14 kDa Schistosoma mansoni fatty acid-binding proteinthrough an IFN-gamma and TNF-alpha dependent pathway. Vaccine2004;22(3–4):503–10.
26] Siddiqui AA, Pinkston JR, Quinlin ML, Kavikondala V, Rewers-Felkins KA, Phillips T, et al. Characterization of protective immunityinduced against Schistosoma mansoni via DNA priming with the large
subunit of calpain (Sm-p80) in the presence of genetic adjuvants. Par-asite 2005;12(1):3–8.27] Maizels RM, Yazdanbakhsh M. Immune regulation by helminthparasites: cellular and molecular mechanisms. Nat Rev Immunol2003;3(9):733–44.
Copyright © 2022 FDOKUMEN




















