
Cytokine mRNA profiles in pigs exposed prenatally and postnatally to Schistosoma japonicum
12
Vet. Res. 38 (2007) 25–36 25 c INRA, EDP Sciences, 2007 DOI: 10.1051/vetres:2006042 Original article Cytokine mRNA profiles in pigs exposed prenatally and postnatally to Schistosoma japonicum Michala E. T a,b , Maria V. J a , Bent A c , Peter L d , Niel O a , Isabelle P. O e * a DBL – Institute for Health Research and Development, Jaegersborg Allé 1D, 2920 Charlottenlund, Denmark b Danish Centre for Experimental Parasitology, Dyrlaegevej 100, 1870 Frederiksberg C, Denmark c Department of Veterinary Microbiology, The Royal Veterinary and Agricultural University, Stigboejlen 7, 1870 Frederiksberg C, Denmark d Department of Immunology and Biochemistry, Danish Veterinary Institute 27 Bülowsvej, 1790 Copenhagen V, Denmark e INRA, Laboratoire de Pharmacologie-Toxicologie, 180 Chemin de Tournefeuille, BP 3, 31931 Toulouse Cedex 9, France (Received 30 January 2006; accepted 12 July 2006) Abstract – The pig is a natural host for Schistosoma japonicum and a useful animal model of human infection. The aim of the present study was to assess the differences between the cytokine profiles in prenatally or postnatally S. japonicum exposed pigs. Seven prenatally exposed pigs, 7 postnatally exposed pigs and 4 uninfected control pigs were compared 27 weeks post exposure. Variables included worm burdens, tissue egg counts, liver pathology and mRNA levels of IL-2, IL-4, IL-10, IL-12, TNF-α, TGF-β1 and IFN-γ in the liver and the caecum, assessed by RT-PCR. Infection intensity and level of septal fibrosis were significantly higher in the postnatal group compared to the prenatal group (P < 0.05). A significant increase of IL-4 (P < 0.01), IL-10 (P < 0.01), IL-12 (P < 0.01) and TNF-α (P < 0.05) mRNA level was also observed in the caecum of prenatally infected animals compared to the control group (P < 0.01). The prenatal group showed higher levels of TGF-β1 in the liver compared with the postnatally infected group (P < 0.05) and the control group (P < 0.01). This suppressive immune response correlated with previously reported low hepatic pathogenesis in prenatally exposed pigs. swine / Schistosoma japonicum / cytokine / prenatal infection / postnatal infection 1. INTRODUCTION Schistosomiasis remains one of the most prevalent infections with more than 200 million people infected worldwide. The disease constitutes a major public health problem in sub-Saharan Africa, re- * Corresponding author: [email protected] gions of South America, the People’s Re- public of China, the Philippines and in mi- nor endemic foci in other Southeast Asian countries [27, 37]. Thus, there is still a crit- ical need for a further understanding of the immune responses elicited during in- fection, in order to generate insight into plausible mechanisms of immune regula- tion and treatment strategies [42]. Article available at http://www.edpsciences.org/vetres or http://dx.doi.org/10.1051/vetres:2006042 Article available at http://www.edpsciences.org/vetres or http://dx.doi.org/10.1051/vetres:2006042
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Cytokine mRNA profiles in pigs exposed prenatally and postnatally to Schistosoma japonicum

Vet. Res. 38 (2007) 25–36 25c© INRA, EDP Sciences, 2007DOI: 10.1051/vetres:2006042
Original article
Cytokine mRNA profiles in pigs exposed prenatallyand postnatally to Schistosoma japonicum
Michala E. Ta,b, Maria V. Ja, Bent Ac,Peter Ld, Niel Oa, Isabelle P. Oe*
a DBL – Institute for Health Research and Development, Jaegersborg Allé 1D,2920 Charlottenlund, Denmark
b Danish Centre for Experimental Parasitology, Dyrlaegevej 100, 1870 Frederiksberg C, Denmarkc Department of Veterinary Microbiology, The Royal Veterinary and Agricultural University,
Stigboejlen 7, 1870 Frederiksberg C, Denmarkd Department of Immunology and Biochemistry, Danish Veterinary Institute 27 Bülowsvej,
1790 Copenhagen V, Denmarke INRA, Laboratoire de Pharmacologie-Toxicologie, 180 Chemin de Tournefeuille, BP 3,
31931 Toulouse Cedex 9, France
(Received 30 January 2006; accepted 12 July 2006)
Abstract – The pig is a natural host for Schistosoma japonicum and a useful animal model ofhuman infection. The aim of the present study was to assess the differences between the cytokineprofiles in prenatally or postnatally S. japonicum exposed pigs. Seven prenatally exposed pigs,7 postnatally exposed pigs and 4 uninfected control pigs were compared 27 weeks post exposure.Variables included worm burdens, tissue egg counts, liver pathology and mRNA levels of IL-2, IL-4,IL-10, IL-12, TNF-α, TGF-β1 and IFN-γ in the liver and the caecum, assessed by RT-PCR. Infectionintensity and level of septal fibrosis were significantly higher in the postnatal group compared tothe prenatal group (P < 0.05). A significant increase of IL-4 (P < 0.01), IL-10 (P < 0.01), IL-12(P < 0.01) and TNF-α (P < 0.05) mRNA level was also observed in the caecum of prenatallyinfected animals compared to the control group (P < 0.01). The prenatal group showed higherlevels of TGF-β1 in the liver compared with the postnatally infected group (P < 0.05) and thecontrol group (P < 0.01). This suppressive immune response correlated with previously reportedlow hepatic pathogenesis in prenatally exposed pigs.
swine / Schistosoma japonicum / cytokine / prenatal infection / postnatal infection
1. INTRODUCTION
Schistosomiasis remains one of themost prevalent infections with more than200 million people infected worldwide.The disease constitutes a major publichealth problem in sub-Saharan Africa, re-
* Corresponding author:[email protected]
gions of South America, the People’s Re-public of China, the Philippines and in mi-nor endemic foci in other Southeast Asiancountries [27,37]. Thus, there is still a crit-ical need for a further understanding ofthe immune responses elicited during in-fection, in order to generate insight intoplausible mechanisms of immune regula-tion and treatment strategies [42].
Article available at http://www.edpsciences.org/vetres or http://dx.doi.org/10.1051/vetres:2006042Article available at http://www.edpsciences.org/vetres or http://dx.doi.org/10.1051/vetres:2006042

26 M.E. Techau et al.
In chronic infections with Schistosomamansoni and S. japonicum, parasite devel-opment and granuloma formation is be-lieved to be a T-helper (Th) lymphocytedependent process [11, 24]. In experimen-tal murine schistosome infections, it wasoriginally thought that a Th2 response,involved in the development of chronic in-fection, was the main cause of pathology,whereas a Th1 response participated inprotection against the infection [9]. How-ever, more recent studies suggest that thesituation is more complicated, showingthat schistosome infections in mice withreduced type 2 responsiveness lead to amore severe pathological outcome thanin mice capable of mounting type 2 re-sponses [14]. In agreement with the morerecent studies, immuno-epidemiologicalstudies of human populations in endemicareas have revealed a positive correla-tion between high, specific immunoglobu-lin (Ig) E levels, eosinophilia, interleukin(IL)-5 levels and resistance to reinfectionafter treatment [30,48]. However, high lev-els of gamma interferon (IFN-γ) and tumornecrosis factor (TNF) together with lowerlevels of IL-5 seem to be associated withhepatosplenic disease [33]. These resultssupport a beneficial role of Th2 responsesin both experimental murine infections andhuman schistosomiasis.
The current understanding of the im-munological basis of schistosomiasis isbased mainly on results obtained usingmurine models [7, 39]. There are however,problems in extrapolating results from thisunnatural host to humans, especially withregards to granuloma induced fibrosis for-mation in schistosomiasis patients [2, 13].The pig is a natural host for S. japon-icum [8] and experimental S. japonicuminfection has been readily established inDanish Landrace/Yorkshire/Duroc cross-bred sows, causing pathogenesis similar tothat described in humans, with the mostseverely affected organs being the intestineand the liver [17]. Willingham et al. [45]
showed that S. japonicum readily estab-lishes in the pig fetus if the sow is in-fected during mid to late pregnancy. In-terestingly, prenatally exposed pigs werelater shown to develop less severe hepaticpathology, compared to challenge controlanimals, when given a challenge infec-tion [22]. Johansen and Ørnbjerg [23] fur-ther showed that treatment with praziquan-tel, previously reported to give a 100% curerate [20], did not cure the prenatal infectionand that a challenge infection was read-ily established in these pigs. These resultsindicate that pigs might develop partial tol-erance towards S. japonicum after infectionin utero.
The prenatal S. japonicum model is wellestablished in terms of parasitological andpathological effects of the infection [45],but knowledge on induced production ofimmune mediators is very scarce [41]. Adeeper understanding of the cytokine re-sponses induced in prenatally and post-natally exposed pigs, might reveal impor-tant differences, reflecting which factorsare likely to be involved in immunopathol-ogy, given that the pathological outcomeof disease is less severe in prenatal infec-tions. Here the mRNA expression levelsfor 7 different cytokines were measured, inorder to investigate the cytokine profiles inprenatally versus postnatally S. japonicumexposed pigs.
2. MATERIALS AND METHODS
2.1. Parasites
Schistosoma japonicum of the Anhuistrain [21], were maintained by laboratorypassage in Oncomelania hupensis snailsand female NMRI mice housed at DBL –Institute for Health Research and Develop-ment. Cercariae were harvested as previ-ously described [31].

Cytokine mRNA in Schistosome infected pigs 27
2.2. Animals and experimental design
Six pathogen-free and helminth-naiveDanish Landrace/Yorkshire/Duroc cross-breed sows were used in this experiment.Ten weeks post-insemination, 3 of the sowswere infected by intramuscular injection of10 000 S. japonicum cercariae [44]. Sevenpiglets born from the infected sows wereremoved immediately after delivery, andreceived colostrum and milk from an un-infected sow, to ensure that the immunityof the sows had no influence on the im-mune responses mounted in the prenatallyexposed piglets. Among piglets born fromuninfected control sows, seven were in-fected intramuscularly with 1000 cercariae9 weeks post delivery, while 4 piglets re-mained as uninfected controls and receivedan injection of suspension medium as pre-viously described [41]. All animals werehoused at Sjaelland III and treated in ac-cordance with protocols approved by theanimal ethics committee of Denmark.
2.3. Parasitological techniques
At 27 weeks post-infection (i.e. at 21and 36 weeks of age for the prenatallyand postnatally exposed animals respec-tively), all the pigs were perfused as pre-viously described [4]. Briefly, pigs werekilled by an intravenously injected over-dose of pentobarbital (30 mg/kg). Heparinsulfate (5000 IU) was also injected in-travenously. One central longitudinal cutwas performed from the sternum to thelower abdomen and selective perfusion ofthe portal hepatic system and the intestinalmesenteric system was achieved by clamp-ing the vessels supplying blood to otherorgans and hind legs. The perfusion tubewas inserted into the aorta and sodium cit-rate containing saline was pumped throughthe portal and mesenteric vessels and col-lected at the punctured portal vein. Theperfusate was sieved and worms were col-lected [4]. Adult S. japonicum worms were
counted and classified according to sexand maturity. After perfusion, the intestinaltract of each pig was examined for resid-ual worms, and the caecal mucosae werescraped for tissue egg counts. The numberof eggs in the faeces, liver and caecum wasdetermined as previously described [41].Liver fibrosis was examined and scored asnone = 0, mild = 1, moderate = 2, or se-vere = 3 depending on the number of wormnodules, lesions and levels of interlobularconnective tissue related to fibrosis [21].
2.4. Determination of cytokine mRNAexpression by semi-quantitativeRT-PCR
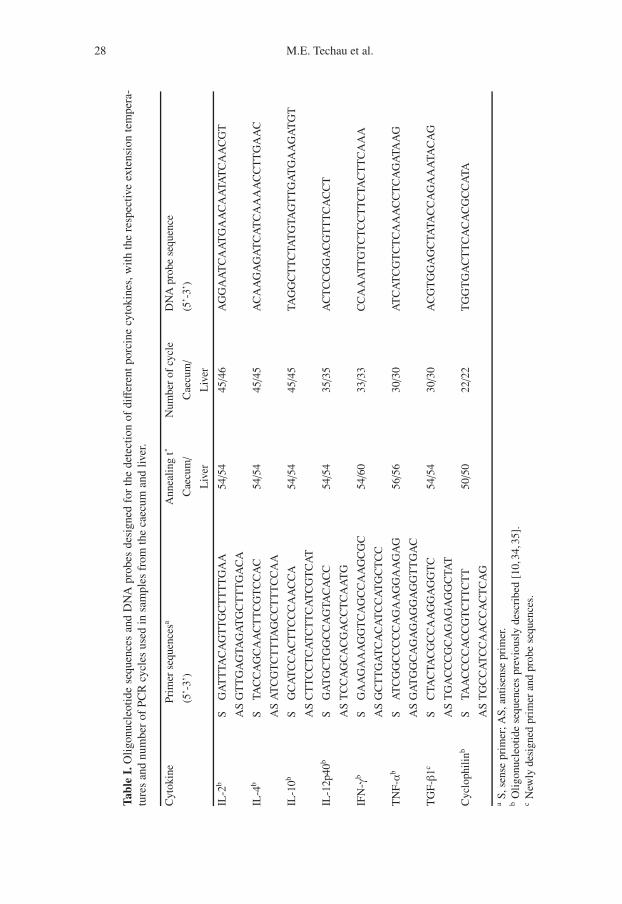
Tissue samples were taken at the au-topsy of the animals from lesion sitesin the liver and the caecum. Theywere immediately preserved in TrizolReagent (GibcoBRL, Life Technologies,Germany) and homogenized on ice us-ing a Cat homogenizer. Total RNA wasextracted as recommended by the man-ufacturer and quantitated by spectropho-tometry. Semi-quantitative determinationof IL-2, IL-4, IL-10, IL-12, TNF-α, TGF-β1 and IFN-γ levels was carried out us-ing RT-PCR as previously described, us-ing cyclophilin as the housekeeping genecontrol [34]. Briefly, mRNA was reversetranscribed with Moloney leukaemia virusreverse transcriptase (Promega, Charbon-nières, France) and amplified with DNATaq polymerase enzyme (Invitrogen, LifeTechnology, Cergy-Pontoise, France). Theprimer sequences, number of cycles andthe annealing temperature used for PCRamplification of each cytokine and cy-clophilin are summarized in Table I. Semi-quantitative analysis of PCR products wasdone by hybridization of 32P-labelled spe-cific oligonucleotide probes to the PCRproducts, which were immobilized on ni-trocellulose membranes by dot blottingas previously described [10, 35]. The
28 M.E. Techau et al.
Tabl
eI.
Oli
gonu
cleo
tide
sequ
ence
san
dD
NA
prob
esde
sign
edfo
rth
ede
tect
ion
ofdiff
eren
tpo
rcin
ecy
toki
nes,
wit
hth
ere
spec
tive
exte
nsio
nte
mpe
ra-
ture
san
dnu
mbe
rof
PC
Rcy
cles
used
insa
mpl
esfr
omth
eca
ecum
and
liver
.
Cyt
okin
ePr
imer
sequ
ence
saA
nnea
ling
t˚N
umbe
rof
cycl
eD
NA
prob
ese
quen
ce
(5’-
3’)
Cae
cum/
Cae
cum/
(5’-
3’)
Liv
erL
iver
IL-2
bS
GA
TT
TAC
AG
TT
GC
TT
TT
GA
A54/5
445/4
6A
GG
AA
TC
AA
TG
AA
CA
ATA
TC
AA
CG
T
AS
GT
TG
AG
TAG
AT
GC
TT
TG
AC
A
IL-4
bS
TAC
CA
GC
AA
CT
TC
GT
CC
AC
54/5
445/4
5A
CA
AG
AG
AT
CA
TC
AA
AA
CC
TT
GA
AC
AS
AT
CG
TC
TT
TAG
CC
TT
TC
CA
A
IL-1
0bS
GC
AT
CC
AC
TT
CC
CA
AC
CA
54/5
445/4
5TA
GG
CT
TC
TAT
GTA
GT
TG
AT
GA
AG
AT
GT
AS
CT
TC
CT
CA
TC
TT
CA
TC
GT
CA
T
IL-1
2p40
bS
GA
TG
CT
GG
CC
AG
TAC
AC
C54/5
435/3
5A
CT
CC
GG
AC
GT
TT
CA
CC
T
AS
TC
CA
GC
AC
GA
CC
TC
AA
TG
IFN
-γb
SG
AA
GA
AA
GG
TC
AG
CC
AA
GC
GC
54/6
033/3
3C
CA
AA
TT
GT
CT
CC
TT
CTA
CT
TC
AA
A
AS
GC
TT
GA
TC
AC
AT
CC
AT
GC
TC
C
TN
F-α
bS
AT
CG
GC
CC
CC
AG
AA
GG
AA
GA
G56/5
630/3
0A
TC
AT
CG
TC
TC
AA
AC
CT
CA
GA
TAA
G
AS
GA
TG
GC
AG
AG
AG
GA
GG
TT
GA
C
TG
F-β
1cS
CTA
CTA
CG
CC
AA
GG
AG
GT
C54/5
430/3
0A
CG
TG
GA
GC
TATA
CC
AG
AA
ATA
CA
G
AS
TG
AC
CC
GC
AG
AG
AG
GC
TAT
Cyc
loph
ilin
bS
TAA
CC
CC
AC
CG
TC
TT
CT
T50/5
022/2
2T
GG
TG
AC
TT
CA
CA
CG
CC
ATA
AS
TG
CC
AT
CC
AA
CC
AC
TC
AG
aS,
sens
epr
imer
;AS,
anti
sens
epr
imer
.b
Oli
gonu
cleo
tide
sequ
ence
spr
evio
usly
desc
ribe
d[1
0,34
,35]
.c
New
lyde
sign
edpr
imer
and
prob
ese
quen
ces.

Cytokine mRNA in Schistosome infected pigs 29
sequence of the DNA probes used for hy-bridization to the different cytokines islisted in Table I. The relative amount ofeach product was determined by measur-ing radioactivity with a Phosphor Imager(Molecular Dynamics, Sunny-vale, CA,USA). For each cytokine, the amount ofRT-PCR product was normalized to thevalues obtained for cyclophilin, as an in-ternal standard for each sample.
2.5. Statistical Analysis
Data from individual pigs are presentedas the group mean ± standard devia-tion. Statistical differences between groupmeans for worm burdens, fecal and liveregg counts and cytokine expression levelswere determined using One-way analy-sis of variance, and post hoc comparisonswere done with the Scheffe range test.Prior to analysis, caecum and liver eggcounts were transformed to log10. Corre-lations between parasitological variableswere assessed by using the Pearson corre-lation test. The ranked degree of liver grosspathology was compared between the twogroups using the Kruskal-Wallis test andMann-Whitney U-test. All analyses weredone using the program Statistical Packagefor Social Sciences1 (SPSS) version 11.0.Values of P < 0.05 were considered signif-icant.
3. RESULTS
3.1. Level of parasitic infection
Piglets were exposed prenatally or post-natally to S. japonicum infection. By exam-ination of faecal egg excretion, infectionwas demonstrated in 5 of the 7 prenatallyexposed piglets and in 6 of the 7 post-natally exposed piglets. The total number
1 www.SPSS.com.
of worms was significantly higher in thepostnatally exposed group in comparisonto the prenatally exposed group (P < 0.05)(Tab. II).
Faecal egg excretion remained lowthroughout the experiment, and due to highvariation within each group, there was nosignificant difference at any time betweenthe two groups (data not shown). At au-topsy, the number of eggs in the liver washigher in the postnatally exposed group(P < 0.01). There was no difference inthe level of portal fibrosis between the twogroups, but a significantly higher degree ofseptal fibrosis was seen in the postnatallyexposed group compared to the prenatallyexposed group (P < 0.05) (Tab. II).
3.2. Cytokine mRNA expression levelsin liver and caecum samples
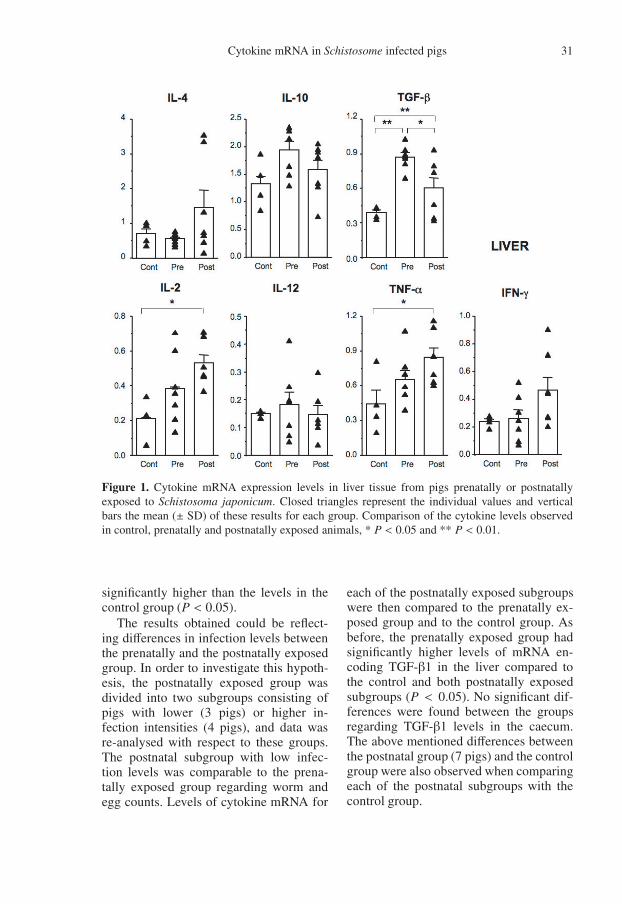
Twenty-seven weeks post infection,liver and caecum samples were collectedfor investigation of IL-2, IL-4, IL-10, IL-12, IFN-γ, TNF-α and TGF-β1 mRNAexpression levels (Figs. 1 and 2). In theliver, the TGF-β1 mRNA expression levelwas significantly higher in the prenatallyexposed group compared to the levels ob-served in both the control group (P <0.01) and the postnatally exposed group(P < 0.05). Furthermore, significantlyhigher levels of TNF-α and IL-2 mRNAwere seen in the postnatally exposed groupcompared to the control (P < 0.05), and atendency towards higher IFN-γ levels wasalso observed in the postnatally exposedgroup.
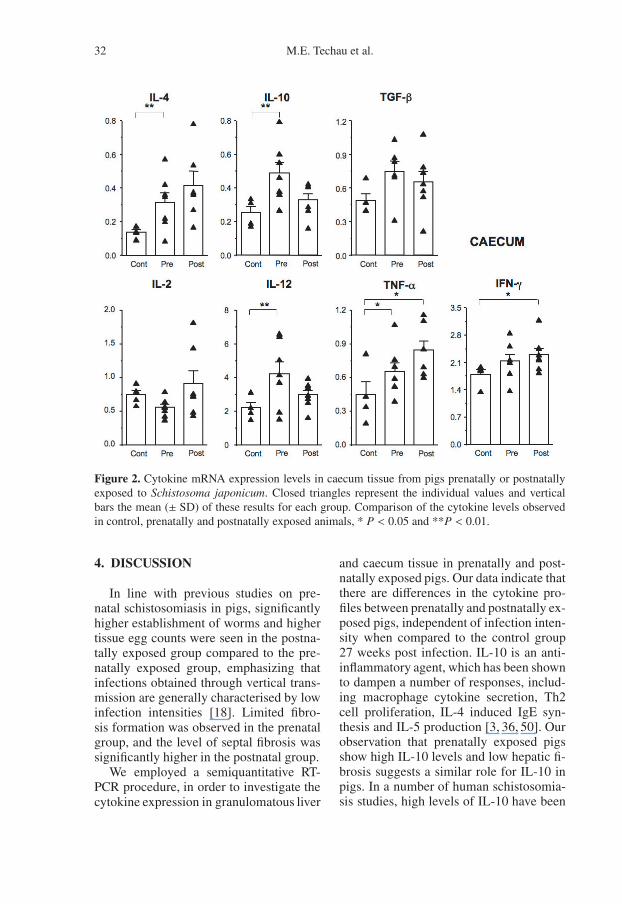
In the caecum IL-4, IL-10, IL-12 andTNF-α levels were significantly increasedin the prenatally exposed group comparedto the control group (for IL-4, IL-10, IL-12P < 0.01, and for TNF-α, P < 0.05), whilea tendency to higher TGF-β1 levels wasobserved. The levels of IFN-γ and TNF-α in the postnatally exposed group were

30 M.E. Techau et al.
Tabl
eII
.Cha
ract
eris
ticof
the
para
sitic
infe
ctio
nin
pren
atal
lyan
dpo
stna
tally
expo
sed
pigs
.
Exp
ositi
onto
the
para
site
aSt
atis
tica
leff
ect
ofth
ety
peof
expo
sure
(pre
vers
uspo
st)
Pren
atal
ly(n=
7)Po
stna
tall
y(n=
7)
Low
infe
ctio
nle
vel(
n=
3)H
igh
infe
ctio
nle
vel(
n=
4)
Tota
lwor
ms
(mea
nnu
mbe
rpe
ran
imal
)0.
7±0.8
b7.
3±7.0
20.0±1
7.5
P<
0.01
Egg
coun
t(nu
mbe
rpe
rgr
amof
tiss
ue)
Liv
er2.
9±7.6
19.7±2
0.0
233.
3±1
39.3
P<
0.01
Cae
cum
06.
7±1
1.5
28.0±3
2.2
NS
Liv
erpa
thol
ogy
(arb
itra
ryun
it)
Sept
alfib
rosi
s0.
31
1P<
0.05
Port
alfib
rosi
s0.
40.
50.
5N
Sa
Pigs
wer
epr
enat
ally
expo
sed
toS.
japo
nicu
mat
10w
eeks
gest
atio
nor
post
nata
lly
expo
sed
9w
eeks
post
deliv
ery.
bR
esul
tsar
eex
pres
sed
asm
ean±S
D.
Cytokine mRNA in Schistosome infected pigs 31
Figure 1. Cytokine mRNA expression levels in liver tissue from pigs prenatally or postnatallyexposed to Schistosoma japonicum. Closed triangles represent the individual values and verticalbars the mean (± SD) of these results for each group. Comparison of the cytokine levels observedin control, prenatally and postnatally exposed animals, * P < 0.05 and ** P < 0.01.
significantly higher than the levels in thecontrol group (P < 0.05).
The results obtained could be reflect-ing differences in infection levels betweenthe prenatally and the postnatally exposedgroup. In order to investigate this hypoth-esis, the postnatally exposed group wasdivided into two subgroups consisting ofpigs with lower (3 pigs) or higher in-fection intensities (4 pigs), and data wasre-analysed with respect to these groups.The postnatal subgroup with low infec-tion levels was comparable to the prena-tally exposed group regarding worm andegg counts. Levels of cytokine mRNA for
each of the postnatally exposed subgroupswere then compared to the prenatally ex-posed group and to the control group. Asbefore, the prenatally exposed group hadsignificantly higher levels of mRNA en-coding TGF-β1 in the liver compared tothe control and both postnatally exposedsubgroups (P < 0.05). No significant dif-ferences were found between the groupsregarding TGF-β1 levels in the caecum.The above mentioned differences betweenthe postnatal group (7 pigs) and the controlgroup were also observed when comparingeach of the postnatal subgroups with thecontrol group.
32 M.E. Techau et al.
Figure 2. Cytokine mRNA expression levels in caecum tissue from pigs prenatally or postnatallyexposed to Schistosoma japonicum. Closed triangles represent the individual values and verticalbars the mean (± SD) of these results for each group. Comparison of the cytokine levels observedin control, prenatally and postnatally exposed animals, * P < 0.05 and **P < 0.01.
4. DISCUSSION
In line with previous studies on pre-natal schistosomiasis in pigs, significantlyhigher establishment of worms and highertissue egg counts were seen in the postna-tally exposed group compared to the pre-natally exposed group, emphasizing thatinfections obtained through vertical trans-mission are generally characterised by lowinfection intensities [18]. Limited fibro-sis formation was observed in the prenatalgroup, and the level of septal fibrosis wassignificantly higher in the postnatal group.
We employed a semiquantitative RT-PCR procedure, in order to investigate thecytokine expression in granulomatous liver
and caecum tissue in prenatally and post-natally exposed pigs. Our data indicate thatthere are differences in the cytokine pro-files between prenatally and postnatally ex-posed pigs, independent of infection inten-sity when compared to the control group27 weeks post infection. IL-10 is an anti-inflammatory agent, which has been shownto dampen a number of responses, includ-ing macrophage cytokine secretion, Th2cell proliferation, IL-4 induced IgE syn-thesis and IL-5 production [3, 36, 50]. Ourobservation that prenatally exposed pigsshow high IL-10 levels and low hepatic fi-brosis suggests a similar role for IL-10 inpigs. In a number of human schistosomia-sis studies, high levels of IL-10 have been

Cytokine mRNA in Schistosome infected pigs 33
shown to be associated with reduced sus-ceptibility to infection, and to play a role inantigen-specific suppression [26,29]. TGF-β1 is also regarded as a potent immunoreg-ulatory cytokine, regulating the differenti-ation, proliferation and state of activationof a large number of immune cells, therebymodulating both cellular and humoral me-diated responses [28]. The prenatally ex-posed group had significantly higher levelsof mRNA encoding TGF-β1 in the livercompared to the control and the postna-tally exposed group (P < 0.05), whereasno significant differences were found be-tween caecum TGF-β1 levels within thethree groups. Our observation, that TGF-β1 mRNA levels were higher in prenatallyexposed pigs, showing less septal fibrosis,suggests that TGF-β1 may mediate down-regulatory responses beneficial to the pre-natally exposed pigs.
In the postnatally exposed pigs, signif-icantly higher levels of TNF-α, togetherwith a tendency to higher levels of IL-2 andIL-4, indicate the existence of a dynamicand not yet polarized Th2 response [40].Apart from being the principal mediatorof acute inflammatory responses, TNF-αis important for the induction of fibroblastmitosis [19]. Interestingly, TNF-α treat-ment has been shown to restore granulomaformation in S. mansoni infected SCIDmice, and furthermore to lead to a dose-dependent increase in the total number ofparasite eggs [1]. Our results suggest thatTNF-α may also play a role in hepaticfibrosis development in pigs postnatally in-fected with S. japonicum.
Blockade of IL-2 responses in S. japon-icum infected mice, leads to reducedhepatic fibrosis, an effect thought to becaused by the down-regulation of Th2 re-sponses [5]. Moreover, anti-IL-4 treatmentof S. mansoni or S. japonicum infectedmice has been shown to result in decreasedfibrosis formation and reduced granulomasize [6, 9]. These results emphasise theimportance of a Th2 response in murine
schistosome fibrosis and granuloma for-mation [38, 39]. Our observations are inline with these results, indicating that highIL-2 and IL-4 responses in postnatally ex-posed pigs, may mediate fibrosis forma-tion. IFN-γ responses have been shown tobe prominent in early disease, while down-regulated in chronic human schistosomeinfections [15, 32]. In our experiment, lev-els of IFN-γ mRNA did not differ betweeninfected and control pigs, but some varia-tion in the expression level of this cytokinewas observed.
The cytokine profiles in the caecum dis-played similar differences between prena-tally or postnatally exposed pigs, as ob-served for the liver. A high IL-10/IL-4ratio in prenatally exposed pigs togetherwith high TGF-β1 levels seem to domi-nate, while postnatally exposed pigs showhigher expression of IL-2 and IL-4. Intesti-nal tissue damage, due to toxic products re-leased from developing schistosome eggs,is often combined with secondary bacte-rial infection [37], which might explain thehigher IL-12 and TNF-α levels seen in thecaecum of prenatally exposed pigs.
Interleukin (IL)-13 is a key inducerof several type-2 cytokine-dependentpathologies [46]. Recently, this cytokinehas been described to regulate tissueremodelling and fibrosis formation inthe murine schistosomiasis model and isregarded as a central mediator of chronicinfection-induced liver pathology [46, 47].At the time of this study, antibody towardsporcine IL-13 and porcine IL-13 sequencewere not available and hence IL-13 levelscould not be determined. In future ex-periments, however, it will be of interestto characterize the expression patternof IL-13 in the porcine schistosomiasismodel.
In line with a previous study [26], weshow that postnatally exposed pigs seemto mount a Th2 like response, with highIL-2 and IL-4 levels locally in lesion sitesof the liver and caecum. Th2 responses,

34 M.E. Techau et al.
although related to hepatic fibrosis, seemto be associated with reduced susceptibil-ity and host protection against overt humanhepatosplenic disease due to inflammatoryresponses [25,30,33]. Indeed, also porcineIL-4 has been shown to play an impor-tant role in the regulation of macrophagemediated inflammatory responses [49]. Inprenatally exposed pigs, a high IL-10/IL-4ratio, together with high levels of TGF-βseem to dominate, suggesting that thesepigs mount a more Th3 like response [16,43]. Such IL-10 and TGF-β1 dominatedresponses have been shown to dominateand mediate antigen-specific cellular hy-poresponsiveness in other chronic humanhelminth infections [12,26]. This indicatesthat the partial tolerance induced in pre-natally exposed pigs [22] might be me-diated by antigen-specific hyporesponsive-ness, resulting in lower levels of hepaticfibrosis.
ACKNOWLEDGEMENTS
Special thanks to Ionelia Taranu and DanielaMarin (Laboratory of Pharmacology and Tox-icology, INRA, Toulouse, France), to HeidiM. Nielsen (DBL – Institute for Health Re-search and Development, Denmark) and toBrian Lassen and Pernille Bach (The Royal Vet-erinary and Agricultural University, Denmark)for technical assistance. Advice on statisticalanalysis from Henry Madsen was greatly appre-ciated. This study was financially supported byThe Danish National Research Foundation.
REFERENCES
[1] Amiri P., Locksley R.M., Parslow T.G.,Sadick M., Rector E., Ritter D., McKerrowJ.H., Tumour necrosis factor alpha restoresgranulomas and induces parasite egg-layingin schistosome-infected SCID mice, Nature(1992) 356:604–607.
[2] Andrade Z.A., Cheever A.W., Characte-rization of the murine model of schistosomalhepatic periportal fibrosis, Int. J. Exp. Pathol.(1993) 74:195–202.
[3] Bogdan C., Nathan C., Modulation ofmacrophage function by transforminggrowth factor beta, interleukin-4, andinterleukin-10, Ann. NY Acad. Sci. (1993)685:713–739.
[4] Bogh H.O., Willingham A.L., JohansenM.V., Eriksen L., Christensen N.O.,Recovery of Schistosoma japonicum fromexperimentally infected pigs by perfusion ofliver and mesenteric veins, Acta Vet. Scand.(1997) 38:147–155.
[5] Cheever A.W., Xu Y., Sher A., FinkelmanF.D., Cox T.M., Macedonia J.G.,Schistosoma japonicum-infected mice showreduced hepatic fibrosis and eosinophiliaand selective inhibition of interleukin-5secretion by CD4+ cells after treatment withanti-interleukin-2 antibodies, Infect. Immun.(1993) 61:1288–1292.
[6] Cheever A.W., Finkelman F.D., Cox T.M.,Anti-interleukin-4 treatment diminishes se-cretion of Th2 cytokines and inhibits hepaticfibrosis in murine schistosomiasis Japonica,Parasite Immunol. (1995) 17:103–109.
[7] Cheever A.W., Yap G.S., Immunologic basisof disease and disease regulation in schisto-somiasis, Chem. Immunol. (1997) 66:159–176.
[8] Chen M.G., Schistosoma japonicum and S.japonicum-like infections – Epidemiology,clinical and pathological aspects, in: JordanP., Webbe G., Sturrock R.F. (Eds.), Humanschistosomiasis, CAB International, Oxon,1993, pp. 242–243.
[9] Chensue S.W., Terebuh P.D., WarmingtonK.S., Hershey S.D., Evanoff H.L., KunkelS.L., Higashi G.I., Role of IL-4 andIFN-gamma in Schistosoma mansoni egg-induced hypersensitivity granuloma forma-tion. Orchestration, relative contribution,and relationship to macrophage function, J.Immunol. (1992) 148:900–906.
[10] Darwich L., Pié S., Rovira A., SegalésJ., Domingo M., Oswald I.P., Mateu E.,Cytokine mRNA expression profiles in lym-phoid tissues of pigs naturally affectedby postweaning multisystemic wasting syn-drome, J. Gen. Virol. (2003) 84:2117–2125.
[11] Davies S., Grogan J., Blank R., Lim K.,Locksley R., McKerrow J., Modulation ofblood fluke development in the liver byhepatic CD4+ lymphocytes, Science (2001)294:1358–1361.
[12] Doetze A., Satoguina J., Burchard G., RauT., Loliger C., Fleischer B., Hoerauf A.,Antigen-specific cellular hyporesponsive-ness in a chronic human helminth infection is

Cytokine mRNA in Schistosome infected pigs 35
mediated by T(h)3/T(r)1-type cytokines IL-10 and transforming growth factor-beta butnot by a T(h)1 to T(h)2 shift, Int. Immunol.(2000) 12:623–630.
[13] Fallon P.G., Immunopathology of schistoso-miasis: a cautionary tale of mice and men,Immunol. Today (2000) 21:29–35.
[14] Fallon P.G., Richardson E.J., McKenzie G.J.,McKenzie A.N., Schistosome infection oftransgenic mice defines distinct and contrast-ing pathogenic roles for IL-4 and IL-13: IL-13 is a profibrotic agent, J. Immunol. (2000)164:2585–2591.
[15] Grogan J.L., Kremsner P.G., DeelderA.M., Yazdanbakhsh M., Antigen-specificproliferation and interferon-gamma andinterleukin-5 production are down-regulatedduring Schistosoma haematobium infection,J. Infect. Dis. (1998) 177:1433–1437.
[16] Groux H., O’Garra A., Bigler M., RouleauM., Antonenko S., de Vries J.E., RoncaroloM.G., A CD4+ T-cell subset inhibits antigen-specific T-cell responses and prevents colitis,Nature (1997) 389:737–742.
[17] Hurst M.H., Shi Y.E., Lindberg R.,Pathology and course of natural Schistosomajaponicum infection in pigs: results of a fieldstudy in Hubei province, China, Ann. Trop.Med. Parasitol. (2000) 94:461–477.
[18] Iburg T., Balemba O.B., Danizer V., LeifssonP.S., Johansen M.V., Pathogenesis of con-genital infection with Schistosoma japon-icum in pigs, J. Parasitol. (2002) 88:1021–1024.
[19] Jaattela M., Biologic activities and mech-anisms of action of tumor necrosis factor-alpha/ cachectin, Lab. Invest. (1991) 64:724–742.
[20] Johansen M.V., Effect of praziquantel treat-ment on experimental porcine Schistosomajaponicum infection, Parasitology (1998)116:519–524.
[21] Johansen M.V., Bogh H.O., Nansen P.,Christensen N.O., Schistosoma japonicuminfection in the pig as a model for humanschistosomiasis Japonica, Acta Trop. (2000)76:85–99.
[22] Johansen M.V., Iburg T., Bogh H.O.,Christensen N.O., Postnatal challengeinfections of congenitally Schistosomajaponicum-infected piglets, J. Parasitol.(2001) 87:813–815.
[23] Johansen M.V., Ørnbjerg N., PrenatalSchistosoma japonicum infection in piglets:effect of repeated exposure of the damson treatment efficacy and susceptibility to
challenge infections, J. Parasitol. (2004)90:392–396.
[24] Kaplan M., Whitfield J., Boros D.,Grusby M., Th2 cells are required forthe Schistosoma mansoni egg-inducedgranulomatous response, J. Immunol. (1998)160:1850–1856.
[25] Karanja D.M., Hightower A.W., ColleyD.G., Mwinzi P.N., Galil K., Andove J.,Secor W.E., Resistance to reinfection withSchistosoma mansoni in occupationally ex-posed adults and effect of HIV-1 co-infectionon susceptibility to schistosomiasis: a longi-tudinal study, Lancet (2002) 360:592–596.
[26] King C.L., Medhat A., Malhotra I., NafehM., Helmy A., Khaudary J., Ibrahim S., ElSherbiny M., Zaky S., Stupi R.J., BrustoskiK., Shehata M., Shata T., Cytokine con-trol of parasite-specific anergy in humanurinary schistosomiasis. IL-10 modulateslymphocyte reactivity, J. Immunol. (1996)156:4715–4721.
[27] King C., Dickman K., Tisch D.,Reassessment of the cost of chronichelmintic infection: a meta-analysis ofdisability-related outcomes in endemicschistosomiasis, Lancet (2005) 365:1561–1569.
[28] Letterio J.L., Roberts A.B., TGF-b: a criti-cal modulator of immune cell function, Clin.Immunol. Immunopathol. (1997) 84:244–250.
[29] McManus D.P., Ross A.G., Sleigh A.C.,Williams G.M., Yang W., Li Y.S., LiY., Acosta L., Waine G.J., Production ofinterleukin-10 by peripheral blood mononu-clear cells from residents of a marshland areain China endemic for Schistosoma japon-icum, Parasitol. Int. (1999) 48:169–177.
[30] Medhat A., Shehata M., Bucci K., MohamedS., Dief A.D., Badary S., Galal H., NafehM., King C.L., Increased interleukin-4 andinterleukin-5 production in response toSchistosoma haematobium adult worm anti-gens correlates with lack of reinfection aftertreatment, J. Infect. Dis. (1998) 178:512–519.
[31] Moloney N.A., Webbe G., A rapid methodfor the infection of laboratory mice withSchistosoma japonicum, Trans. R. Soc. Trop.Med. Hyg. (1982) 76:200–203.
[32] Montenegro S.M., Miranda P., MahantyS., Abath F.G., Teixeira K.M., CoutinhoE.M., Brinkman J., Goncalves I., DominguesL.A., Domingues A.L., Sher A., Wynn T.A.,Cytokine production in acute versus chronic

36 M.E. Techau et al.
human schistosomiasis mansoni: the cross-regulatory role of interferon-gamma andinterleukin-10 in the responses of peripheralblood mononuclear cells and splenocytesto parasite antigens, J. Infect. Dis. (1999)179:1502–1514.
[33] Mwatha J.K., Kimani G., Kamau T., MbuguaG.G., Ouma J.H., Mumo J., Fulford A.J.,Jones F.M., Butterworth A.E., Roberts M.B.,Dunne D.W., High levels of TNF, solu-ble TNF receptors, soluble ICAM-1, andIFN-gamma, but low levels of IL-5, are as-sociated with hepatosplenic disease in hu-man schistosomiasis mansoni, J. Immunol.(1998) 160:1992–1999.
[34] Oswald I.P., Dozois C.M., Barlagne R.,Fournout S., Johansen M.V., Bogh H.O.,Cytokine mRNA expression in pigs infectedwith Schistosoma japonicum, Parasitology(2001) 122:299–307.
[35] Pié S., Lallès J.P., Blazy F., Laffitte J., SèveB., Oswald I.P., Weaning is associated withan up-regulation of expression of inflamma-tory cytokines in the intestine of piglets, J.Nutr. (2004) 134:641–647.
[36] Punnonen J., Malefyt de Waal R., vanVlasselaer P., Gauchat J.F., de Vries J.E.,IL-10 and viral IL-10 prevent IL-4-inducedIgE synthesis by inhibiting the accessory cellfunction of monocytes, J. Immunol. (1993)151:1280–1209.
[37] Ross A., Sleigh A., Li Y., Davis G.,Williams G., Jiang Z., Feng Z., McManusD., Schistosomiasis in the People’s Republicof China: prospects and challenges for the21st Century, Clin. Microbiol. Rev. (2001)14:270–295.
[38] Rumbley C.A., Zekavat S.A., Sugaya H.,Perrin P.J., Ramadan M.A., Phillips S.M.,The schistosome granuloma: characteriza-tion of lymphocyte migration, activation,and cytokine production, J. Immunol. (1998)161:4129–4137.
[39] Rumbley C.A., Sugaya H., Zekavat S.A.,El Refaei M., Perrin P.J., Phillips S.M.,Activated eosinophils are the major Sourceof Th2-associated cytokines in the schis-tosome granuloma, J. Immunol. (1999)162:1003–1009.
[40] Stadecker M.J., The regulatory role of theantigen-presenting cell in the development of
hepatic immunopathology during infectionwith Schistosoma mansoni, Pathobiology(1999) 67:269–272.
[41] Techau M.E., Johansen M.V., Lind P.,Ornbjerg N., The effect of colostrum onpigs pre-natally or post-natally exposed toSchistosoma japonicum, Parasitology (2004)129:597–604.
[42] Vennervald B., Dunne D., Morbidity inschistosomiasis: an update, Curr. Opin.Infect. Dis. (2004) 5:439–447.
[43] Weiner H.L., Oral tolerance: immune mech-anisms and treatment of autoimmune dis-eases, Immunol. Today (1997) 18:335–343.
[44] Willingham A.L., Bogh H.O., VennervaldB.J., Johansen M.V., Eriksen L., ChristensenN.O., Nansen P., Worm establishment andegg production of Schistosoma japonicum inpigs infected by percutaneous methods or in-tramuscular injection, Vet. Parasitol. (1996)61:157–163.
[45] Willingham A.L., Johansen M.V., BoghH.O., Ito A., Andreassen J., Lindberg R.,Christensen N.O., Nansen P., Congenitaltransmission of Schistosoma japonicum inpigs, Am. J. Trop. Med. Hyg. (1999) 60:311–312.
[46] Wynn T.A., IL-13 effector functions, Annu.Rev. Immunol. (2003) 21:425–456.
[47] Wynn T.A., Thompson R.W., Cheever A.W.,Mentink-Kane M.M., Immunopathogenesisof schistosomiasis, Immunol. Rev. (2004)201:156–167.
[48] Zhang Z., Wu H., Chen S., Hu L., Xie Z., QiuY., Su C., Cao J.P., Wu Y., Zhang S., Wu G.,Association between IgE antibody againstsoluble egg antigen and resistance to reinfec-tion with Schistosoma japonicum, Trans. R.Soc. Trop. Med. Hyg. (1997) 91:606–608.
[49] Zhou Y., Lin G., Baarsch M.J., ScamurraR.W., Murtaugh M.P., Interleukin-4 sup-presses inflammatory cytokine gene tran-scription in porcine macrophages, J. Leukoc.Biol. (1994) 56:507–513.
[50] Zuany-Amorim C., Creminon C., NeversM.C., Nahori M.A., Vargaftig B.B., PretolaniM., Modulation by IL-10 of antigen-inducedIL-5 generation, and CD4+ T lympho-cyte and eosinophil infiltration into themouse peritoneal cavity, J. Immunol. (1996)157:377–384.




















