
Tissue responses exhibited by Biomphalaria alexandrina snails from different Egyptian localities...
7
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Tissue responses exhibited by Biomphalaria alexandrina snails from different Egyptian localities...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Tissue responses exhibited by Biomphalaria alexandrina snails from differentEgyptian localities following Schistosoma mansoni exposure
Azza H. Mohamed a, Ahmed T. Sharaf El-Din b,⇑, Ahmed M. Mohamed a, Mohamed R. Habib b
a Zoology Department, Faculty of Science, Menoufiya University, Shebin El-Kom, Egyptb Medical Malacology Lab., Theodor Bilharz Research Institute, P.O. Box 30, Imbaba, Egypt
a r t i c l e i n f o
Article history:Received 23 September 2010Received in revised form 1 January 2011Accepted 28 January 2011Available online 2 February 2011
Keywords:Biomphalaria alexandrinaSchistosoma mansoniResistanceSusceptibilityEncapsulation
a b s t r a c t
Snails’ susceptibilities to infection with Schistosoma mansoni were determined through observation ofinfection rates, total cercarial production and tissue responses of the first generation (F1) of Biomphalariaalexandrina snails, originally collected from different Egyptian governorates (Giza, Fayoum, Kafr El-Sheikh, Ismailia and Damietta) and responses were compared between groups. The emergence of cerca-riae for a 3-month period and the calculation of survival and infection rates, in control (Schistosome Bio-logical Supply Center; SBSC) and infected snails were evaluated. SBSC and Giza snails showed greatersusceptibilities to infection and lower mortality rates. In addition, at 6 and 72 h post-exposure to mira-cidia all the snail groups showed no difference in the anatomical locations of sporocysts. The larvae werefound in the head-foot, the mantle collar and the tentacles of the snails. Sporocysts showed normal devel-opment with low tissue reactions in SBSC and Giza snail groups infected with S. mansoni miracidia (SBSC).However, in Fayoum, Kafr El-Sheikh, Ismailia and Damietta snail groups, variable tissue responses wereobserved in which numerous hemocytes made direct contact with S. mansoni larvae forming capsules.The results suggested that, different responses of B. alexandrina snail’s hemocytes towards S. mansoniare related to the degree of susceptibility of these snails. So this is important in planning the strategyof schistosomiasis control.
� 2011 Elsevier Inc. All rights reserved.
1. Introduction
The host–parasite relationship is complex and questions remainconcerning the susceptibility of snails to infection by the respectivetrematodes and their suitability as hosts for continued parasitedevelopment. The dynamic interaction between molluscs and theirtrematode parasites leads either to a state of co-existence, in whichthe trematode thrives and produces subsequent stages of its life-cycle, or to incompatibility, where the trematode is either de-stroyed and eliminated by the host snail defensive responses orfails to develop because the host is physiologically unsuitable (Bay-ne and Yashino, 1989; van der Knaap and Loker, 1990). Successfulcolonization of a compatible snail host by a digenetic trematodemiracidium initiates a complex proliferative development programrequiring weeks to reach culmination in the form of production ofcercariae which, once started, may persist for the remainder of thelife span of the infected snail (Hanington et al., 2010).
Geographical distribution of intestinal schistosomiasis is di-rectly associated with the presence of susceptible snails of the
genus Biomphalaria and the etiological agent, S. mansoni. This trem-atode is a stenoxenic parasite, i.e., it uses specific intermediate hostspecies (Caldeira et al., 2009). However, not all Biomphalaria spe-cies are susceptible to S. mansoni. Biomphalaria susceptibility to S.mansoni infection varies among snails according to different ages,genetic variation, immune system status and geographic areas inwhich both snails and the trematode occur (Richards and Shade,1987).
During the life cycle of S. mansoni, sporocysts larval stages de-velop in the mollusc intermediate hosts. Parasites need to pene-trate into this host, develop, multiply asexually and finally leavethe host to continue their life cycle (Davies and Mckerrow, 2003;Parker et al., 2003). Parasites therefore face many challenges suchas gaining enough energy to grow and to evade the defense systemof the host (Meuleman et al., 1987; deJong-Brink et al., 2001). Inparallel, hosts have to co-evolve with their parasites to avoid beinginfected. Susceptibility or resistance to infection in planorbid snailsby S. mansoni is regulated genetically in a way that some suscepti-bility may be present in resistant snails (Richards and Merritt,1972; Carton et al., 2005).
Many studies have been done to investigate the mechanisms bywhich the snail resistance is achieved (Granath and Yashino, 1983;Loker and Bayne, 1986; Bayne and Yashino, 1989). From thesestudies, immune response of the snail intermediate host
0014-4894/$ - see front matter � 2011 Elsevier Inc. All rights reserved.doi:10.1016/j.exppara.2011.01.020
⇑ Corresponding author. Address: Department of Environmental Researches andMedical Malacology, Theodor Bilharz Research Institute, P.O. Box 30, Imbaba, Egypt.Fax: +20 2 5218 0453.
E-mail address: [email protected] (A.T. Sharaf El-Din).
Experimental Parasitology 127 (2011) 789–794
Contents lists available at ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/locate /yexpr

Author's personal copy
Biomphalaria glabrata is determined through complex relationshipinvolving circulating hemocytes and the early larval stages of theparasite. In resistant snails, hemocytes recognize and destroy theparasite via a cellular encapsulation response that may involveplasma activating or recognition factors, lysosomal enzymes orother cytotoxic element and phagocytosis of the damaged parasitetegument. Susceptibility generally is viewed to be the result ofhemocytes failure to recognize and/or mount an effective cytotoxicresponse against the invasive parasite larvae. Mohamed et al.(1993) reported that the natural intermediate host of S. mansoniin Egypt is completely refractory to infection with the Puerto Ricanstrain of this parasite. Although the miracidia of the parasite suc-cessfully penetrate the snail, yet they are quickly subjected tostrong tissue reactions leading into the encapsulation of the para-site larvae followed by degeneration and eventual exclusion fromthe snail tissues.
Populations of snails of the same species show different de-grees of susceptibility to infection (Paraense and Corrêa, 1963).Loker and Bayne (1982) reported that the great majority of spo-rocysts incubated in the plasma of susceptible snails and laterput into contact with amoebocytes originating from resistantsnails were destroyed. When the sporocysts were incubated inplasma from resistant snails and later exposed to amoebocytesof susceptible snails, no destruction of the larvae was noted. Sou-za et al. (1995) made a comparative study of the development ofS. mansoni during the intramolluscan phase by mean of histolog-ical sections of Biomphalaria tenagophila, Biomphalaria stramineaand B. glabrata from Brazil; they found that there were not anyobserved larvae in snails fixed 72 h after exposure. In specimensshedding cercariae, 31 days after exposure tissue reactions encap-sulating the larvae were seen in B. tenagophila and B. straminea, inthe head-foot, mantle collar and renal ducts explaining the lowerlevels of infection and average numbers of cercariae shed bythese two species.
The present work was initiated to demonstrate the differentinfectivity profiles and tissue responses generated by Biomphalariaalexandrina snails collected from different Egyptian governoratestowards S. mansoni parasite.
2. Materials and methods
2.1. Snails
The snails used were laboratory-bred B. alexandrina snails (F1)originated from five Egyptian governorates (Giza, Fayoum, KafrEl-Sheikh, Ismailia and Damietta). In addition, a sixth group of B.alexandrina snails, obtained from Schistosome Biological SupplyCenter (SBSC), Theodor Bilharz Research Institute, Giza, Egypt,was used as a reference control group.
2.2. Schistosoma mansoni
S. mansoni ova were obtained from SBSC which was origi-nally an Egyptian strain obtained from Giza Governorate andhas been routinely maintained in B. alexandrina and albino miceMus musculus CD1 strain. All experiments were done in compli-ance with the guide lines for the care and use of Laboratoryanimals.
2.3. Snail exposure to miracidia
Three replicates, each of 30 lab-bred B. alexandrina snails (4–6 mm in diameter), from each governorate offspring were individ-ually exposed to ten newly hatched S. mansoni miracidia (SBSC)suctioned by micropipette according to Théron et al. (1997).
2.4. Examination of exposed snails for cercarial shedding
Starting from the day 21 post miracidial exposure, the snailswere examined individually and repeatedly for cercarial sheddingin multidishes under artificial light for 2 h (stimulant period) and2 ml of dechlorinated tape water per snail. After initial sheddingwas observed, snails were screened individually once weekly tillthe death of snails. All snails that died during the prepatent periodwere crushed between two slides and inspected under a micro-scope for immature parasite stages (Chernin and Dunavan, 1962).The snail’s infection rate was calculated at the end of experimentby dividing the number of shedding and positive crushed snailson the number of exposed snails, the survival rate was calculatedby dividing the number of snails at first shedding on the total num-ber of exposed snails according to Yousif et al. (1998). At the firstday of detecting cercariae, positive snails were separated individu-ally in plastic cups. The produced cercariae per snail were trans-ferred to a small Petri dish by a Pasteur pipette, fixed in Bouin’ssolution and counted under a stereomicroscope. This examinationwas repeated weekly.
2.5. Histological investigations
At time intervals 6 and 72 h post miracidial exposure, five snailsfrom each snail group were carefully crushed between two glassslides, the shell fragments were removed using pointed forceps un-der a dissecting microscope. Head-foots were separated and imme-diately fixed in alcoholic Bouin’s fluid {(15 ml picric acid (saturatedaqueous solution), 5 ml of 40% formalin, and 1 ml of 100% glacialacetic acid)} for 12 h. After fixation, specimens were dehydratedin an ascending series of alcohol (70%, 80%, 90% and 100%)15 min each. The specimens were cleared in two changes of xyleneand embedded in molten paraplast at 60 �C. Serial sections werecut at 5 lm thickness using rotary microtome and stained withEhrlich’s haematoxylin and counterstained eosin (Romeis 1989).The sections were then mounted by DPX and covered by glass cov-ers. Histological sections were examined and photographed withautomatic camera using Olympus System Microscope BX2 Series(BX41, Japan) to detect any hemocytes reactions against theparasite.
2.6. Statistical analysis
Data for the number of cercariae produced were square roottransformed before statistical analyses to satisfy the distributionalassumptions of the test. ANOVA were performed on cercarial shed-ding per week as a repetition factor and parasitic infection as thetreatment factor (Sokal and Rohlf, 1995). Experimental infectionrates of every two snail groups were compared by means of2 � 2 contingency tables, using the chi-square (v2) test. Significantdifferences were considered at p 6 0.05. Data were expressed asmean ± standard error of the mean (SEM).
3. Results
3.1. Survival rate
The survival rate of different snail groups exposed to S. mansonimiracidia (SBSC strain), at first cercarial shedding, was highest inGiza group (87%), while the lowest one was recorded in Ismailiagroup being 40%. In between these two values there were variablepercentages of survival rates recorded for the other snail groups;85% (Fayoum), 60% (SBSC), 50% (Kafr El-Sheikh) and 43% for Dam-ietta snail group (Fig. 1). The differences observed in the survivalrates among the five snail groups were significantly increased in
790 A.H. Mohamed et al. / Experimental Parasitology 127 (2011) 789–794

Author's personal copy
Giza and Fayoum snail groups, while it was significantly decreasedin Ismailia and Damietta groups compared to SBSC snail group.
3.2. Infection rate
The highest infection rate was that of SBSC group with infectionrate 50.3%. On the other hand, B. alexandrina snail groups fromIsmailia and Kafr El-Sheikh showed an equal infection rate (20%),while a moderately higher percent (33.3%) was obtained withDamietta group (Fig. 1). The differences in the infection rates ofthe five snail groups compared to that of SBSC group, were de-creased significantly (p < 0.001) in all groups except in Giza group(43.3%) which showed no significant difference.
3.3. Mean total number of cercariae per snail
A marked variation in the cercarial outputs was observedamong the six B. alexandrina snail groups. The highest mean totalnumber of cercariae per snail was 5198.17 ± 2486.1 in Giza group,while the lowest one was 1175.33 ± 626.99 in Kafr El-Sheikh groupwhich are significantly different (p < 0.01) compared to SBSC B.alexandrina snail group (2575.85 ± 1699.63 cercariae per snail). InFayoum and Ismailia snail groups, the mean total number of cerca-riae per snail were 1267.50 ± 1120.50 and 2495.17 ± 1221.80,respectively for the two snail groups.
3.4. Histological observations
The present results showed that, at each interval post-exposure(6 and 72 h), all B. alexandrina snail groups showed no difference inthe anatomical locations of miracidia and sporocysts, the larvaewere found in the head-foot, the mantle collar and the tentaclesof the snails.
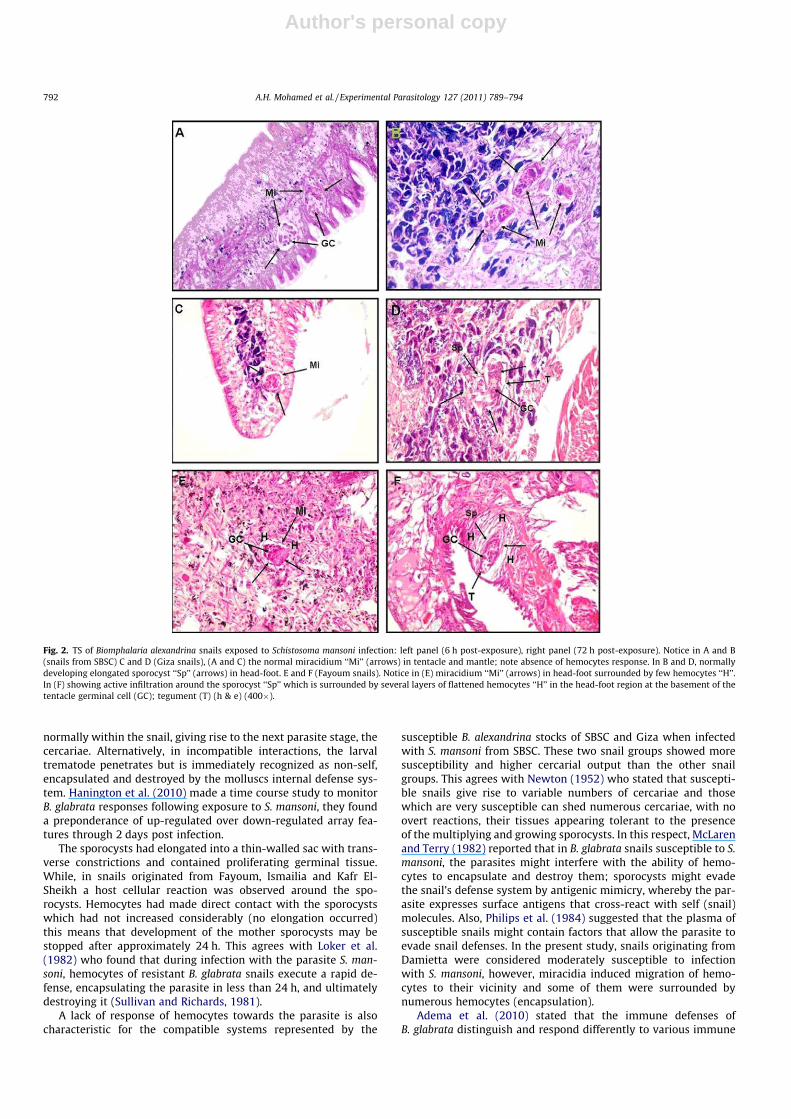
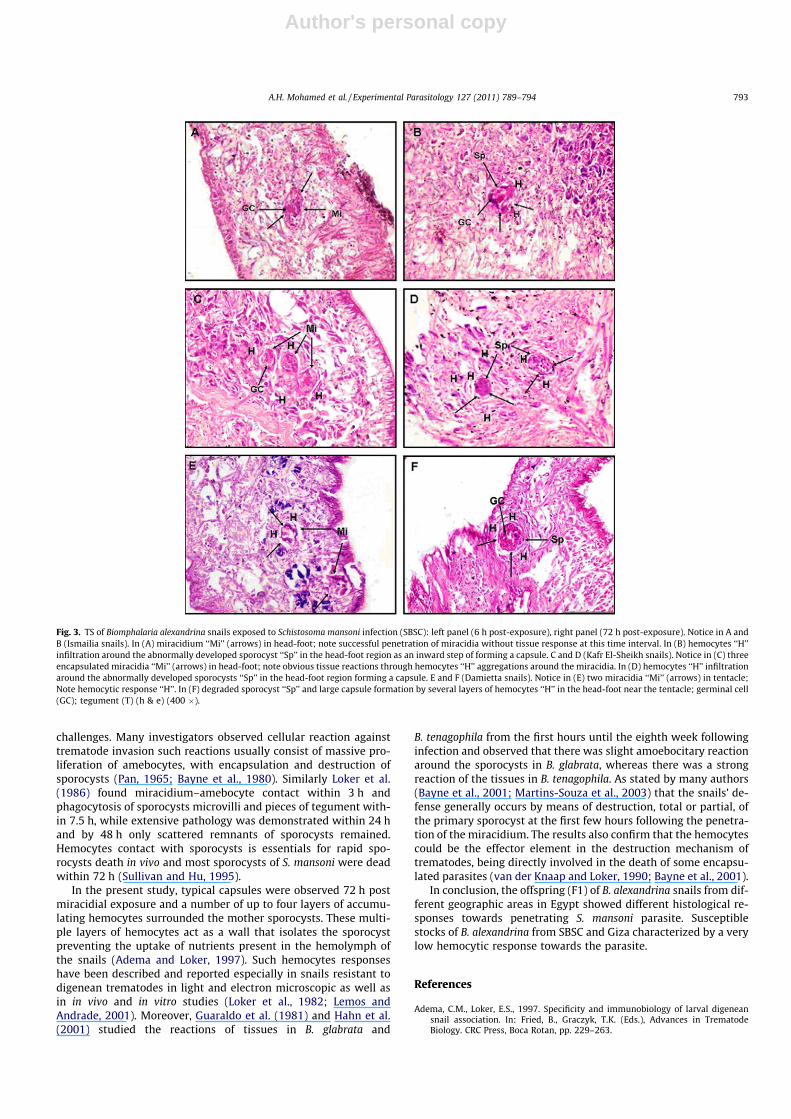
Six-hour post-exposure, some miracidia developed apparentlynormally, while others underwent encapsulation, the penetratingmiracidia were surrounded by numerous hemocytes in snails orig-inated from Fayoum (Fig. 2E), Ismailia (Fig. 3A), Kafr El-Sheikh(Fig. 3C) and Damietta (Fig. 3E). While no cellular reactions wasusually observed and miracidia showed normal development insnails of SBSC (Fig. 2A) and snails from Giza (Fig. 2C). These twosnail groups showed a lower hemocytes response to penetratingmiracidia. Some miracidia had already induced migration of hemo-cytes to their vicinity as in Damietta snails although of its moder-ate susceptibility to S. mansoni infection (Fig. 3E).
Seventy-two hours post-exposure, mother sporocysts were ob-served in various stages of developmental or deterioration in tissuesections of the different B. alexandrina snail groups investigated. Insnails obtained from SBSC and Giza, S. mansoni mother sporocysts
showed normal development, most germinal cells stained nor-mally had characteristic nucleoli which seemed to be proliferated.There was no contact of the sporocysts surface with hemocytes. Nohost cellular response was usually observed around sporocysts.Sporocysts had elongated into a thin-walled sac with transverseconstrictions and contained proliferating germinal tissue (Fig. 2Band D). In snails originated from Fayoum, Ismailia, Kafr El-Sheikhand Damietta, a host cellular reaction was observed around thesporocysts. Hemocytes had made direct contact with the spo-rocysts and usually formed capsules. Capsules were spherical, asseen in Damietta snails (Fig. 3F), or oval shape (Kafr El-Sheikhsnails, Fig. 3D).
4. Discussion
The present results showed clear differences in the degree ofsusceptibility of snail populations originating from some localitiesin Egypt to infection with S. mansoni strain from SBSC. B. alexand-rina from SBSC and Giza exhibited the highest degrees of suscepti-bility amongst snail populations studied during the presentinvestigation. The infection rates were 50.3% and 43.3%, respec-tively. These variations in susceptibility agree with Frandsen(1979) who recorded that B. alexandrina snails from various local-ities in Egypt showed different susceptibility rates to a strain of S.mansoni. Compared to the present results, Bakry (2009) reportedthat B. alexandrina snails from Damietta were less susceptible toinfection with an Egyptian strain of S. mansoni (Giza) than B. alex-andrina from Fayoum and Giza. This overlap may be partiallyattributed to that the snails used in the present study were the firstgeneration and not the field ones.
The highest infection rate exhibited by the snails of SBSC(50.3%) and Giza (43.3%) reflect higher susceptibility to schisto-some infection, since the source of both snail and parasite consid-ered the same. This is agreeing with the theory of local adaptationof the parasite to its snail host (Gandon et al., 1996; Lively, 1999).
The present results indicated that the first generation (F1) ofdifferent B. alexandrina snail groups collected from different geo-graphic areas in Egypt acquired infection with S. mansoni but thesnails exhibited different histological responses towards penetrat-ing S. mansoni parasite, this is in accordance with Théron et al.(1997) who demonstrated that for the same species/specieshost–parasite couple the intraspecific differences occur betweentwo geographical combinations. In the present study, different cel-lular responses were observed in B. alexandrina snails of Fayoum,Ismailia and Kafr El-Sheikh (low susceptible) and even in moder-ately susceptible snails from Damietta.
Miracidia and mother sporocysts were found in the head-foot,tentacles and mantle collar in all B. alexandrina groups after 6and 72 h of exposure to S. mansoni (SBSC). This is in accordancewith the majority of previous observations on Biomphalaria snailsinfected with S. mansoni (Pan, 1965, 1980; Meuleman et al.,1987; Mohamed, 1998). However, Théron et al. (1997) demon-strated experimentally that, distribution patterns of schistosomelarvae among the snail host population may differ depending uponthe host–parasite combination.
Seventy-two hours post-exposure, sporocysts were observed invarious stages of developmental or deterioration in tissue sectionsof the different B. alexandrina groups investigated. With compati-ble B. alexandrina snail hosts obtained from SBSC and Giza, S. man-soni mother sporocysts showed a normal development followingthe usual scheme mentioned by Pan (1965) and there was no con-tact of the sporocysts surface with hemocytes. In the same context,Théron and Coustau (2005) stated that in natural populations somesnail/schistosome combinations are compatible and others are not.In compatible interactions, the parasite penetrates and develops
0102030405060708090
100
SheikhSBSC Giza Fayoum Kafr El- Ismailia Damietta
Snail origin
perc
enta
ge
Survival Infection
Fig. 1. The survival and infection rates of six Biomphalaria alexandrina snail groupsfrom SBSC and five Egyptian governorates infected with Schistosoma mansonimiracidia from SBSC.
A.H. Mohamed et al. / Experimental Parasitology 127 (2011) 789–794 791
Author's personal copy
normally within the snail, giving rise to the next parasite stage, thecercariae. Alternatively, in incompatible interactions, the larvaltrematode penetrates but is immediately recognized as non-self,encapsulated and destroyed by the molluscs internal defense sys-tem. Hanington et al. (2010) made a time course study to monitorB. glabrata responses following exposure to S. mansoni, they founda preponderance of up-regulated over down-regulated array fea-tures through 2 days post infection.
The sporocysts had elongated into a thin-walled sac with trans-verse constrictions and contained proliferating germinal tissue.While, in snails originated from Fayoum, Ismailia and Kafr El-Sheikh a host cellular reaction was observed around the spo-rocysts. Hemocytes had made direct contact with the sporocystswhich had not increased considerably (no elongation occurred)this means that development of the mother sporocysts may bestopped after approximately 24 h. This agrees with Loker et al.(1982) who found that during infection with the parasite S. man-soni, hemocytes of resistant B. glabrata snails execute a rapid de-fense, encapsulating the parasite in less than 24 h, and ultimatelydestroying it (Sullivan and Richards, 1981).
A lack of response of hemocytes towards the parasite is alsocharacteristic for the compatible systems represented by the
susceptible B. alexandrina stocks of SBSC and Giza when infectedwith S. mansoni from SBSC. These two snail groups showed moresusceptibility and higher cercarial output than the other snailgroups. This agrees with Newton (1952) who stated that suscepti-ble snails give rise to variable numbers of cercariae and thosewhich are very susceptible can shed numerous cercariae, with noovert reactions, their tissues appearing tolerant to the presenceof the multiplying and growing sporocysts. In this respect, McLarenand Terry (1982) reported that in B. glabrata snails susceptible to S.mansoni, the parasites might interfere with the ability of hemo-cytes to encapsulate and destroy them; sporocysts might evadethe snail’s defense system by antigenic mimicry, whereby the par-asite expresses surface antigens that cross-react with self (snail)molecules. Also, Philips et al. (1984) suggested that the plasma ofsusceptible snails might contain factors that allow the parasite toevade snail defenses. In the present study, snails originating fromDamietta were considered moderately susceptible to infectionwith S. mansoni, however, miracidia induced migration of hemo-cytes to their vicinity and some of them were surrounded bynumerous hemocytes (encapsulation).
Adema et al. (2010) stated that the immune defenses ofB. glabrata distinguish and respond differently to various immune
Fig. 2. TS of Biomphalaria alexandrina snails exposed to Schistosoma mansoni infection: left panel (6 h post-exposure), right panel (72 h post-exposure). Notice in A and B(snails from SBSC) C and D (Giza snails), (A and C) the normal miracidium ‘‘Mi’’ (arrows) in tentacle and mantle; note absence of hemocytes response. In B and D, normallydeveloping elongated sporocyst ‘‘Sp’’ (arrows) in head-foot. E and F (Fayoum snails). Notice in (E) miracidium ‘‘Mi’’ (arrows) in head-foot surrounded by few hemocytes ‘‘H’’.In (F) showing active infiltration around the sporocyst ‘‘Sp’’ which is surrounded by several layers of flattened hemocytes ‘‘H’’ in the head-foot region at the basement of thetentacle germinal cell (GC); tegument (T) (h & e) (400�).
792 A.H. Mohamed et al. / Experimental Parasitology 127 (2011) 789–794
Author's personal copy
challenges. Many investigators observed cellular reaction againsttrematode invasion such reactions usually consist of massive pro-liferation of amebocytes, with encapsulation and destruction ofsporocysts (Pan, 1965; Bayne et al., 1980). Similarly Loker et al.(1986) found miracidium–amebocyte contact within 3 h andphagocytosis of sporocysts microvilli and pieces of tegument with-in 7.5 h, while extensive pathology was demonstrated within 24 hand by 48 h only scattered remnants of sporocysts remained.Hemocytes contact with sporocysts is essentials for rapid spo-rocysts death in vivo and most sporocysts of S. mansoni were deadwithin 72 h (Sullivan and Hu, 1995).
In the present study, typical capsules were observed 72 h postmiracidial exposure and a number of up to four layers of accumu-lating hemocytes surrounded the mother sporocysts. These multi-ple layers of hemocytes act as a wall that isolates the sporocystpreventing the uptake of nutrients present in the hemolymph ofthe snails (Adema and Loker, 1997). Such hemocytes responseshave been described and reported especially in snails resistant todigenean trematodes in light and electron microscopic as well asin in vivo and in vitro studies (Loker et al., 1982; Lemos andAndrade, 2001). Moreover, Guaraldo et al. (1981) and Hahn et al.(2001) studied the reactions of tissues in B. glabrata and
B. tenagophila from the first hours until the eighth week followinginfection and observed that there was slight amoebocitary reactionaround the sporocysts in B. glabrata, whereas there was a strongreaction of the tissues in B. tenagophila. As stated by many authors(Bayne et al., 2001; Martins-Souza et al., 2003) that the snails’ de-fense generally occurs by means of destruction, total or partial, ofthe primary sporocyst at the first few hours following the penetra-tion of the miracidium. The results also confirm that the hemocytescould be the effector element in the destruction mechanism oftrematodes, being directly involved in the death of some encapsu-lated parasites (van der Knaap and Loker, 1990; Bayne et al., 2001).
In conclusion, the offspring (F1) of B. alexandrina snails from dif-ferent geographic areas in Egypt showed different histological re-sponses towards penetrating S. mansoni parasite. Susceptiblestocks of B. alexandrina from SBSC and Giza characterized by a verylow hemocytic response towards the parasite.
References
Adema, C.M., Loker, E.S., 1997. Specificity and immunobiology of larval digeneansnail association. In: Fried, B., Graczyk, T.K. (Eds.), Advances in TrematodeBiology. CRC Press, Boca Rotan, pp. 229–263.
Fig. 3. TS of Biomphalaria alexandrina snails exposed to Schistosoma mansoni infection (SBSC): left panel (6 h post-exposure), right panel (72 h post-exposure). Notice in A andB (Ismailia snails). In (A) miracidium ‘‘Mi’’ (arrows) in head-foot; note successful penetration of miracidia without tissue response at this time interval. In (B) hemocytes ‘‘H’’infiltration around the abnormally developed sporocyst ‘‘Sp’’ in the head-foot region as an inward step of forming a capsule. C and D (Kafr El-Sheikh snails). Notice in (C) threeencapsulated miracidia ‘‘Mi’’ (arrows) in head-foot; note obvious tissue reactions through hemocytes ‘‘H’’ aggregations around the miracidia. In (D) hemocytes ‘‘H’’ infiltrationaround the abnormally developed sporocysts ‘‘Sp’’ in the head-foot region forming a capsule. E and F (Damietta snails). Notice in (E) two miracidia ‘‘Mi’’ (arrows) in tentacle;Note hemocytic response ‘‘H’’. In (F) degraded sporocyst ‘‘Sp’’ and large capsule formation by several layers of hemocytes ‘‘H’’ in the head-foot near the tentacle; germinal cell(GC); tegument (T) (h & e) (400 �).
A.H. Mohamed et al. / Experimental Parasitology 127 (2011) 789–794 793

Author's personal copy
Adema, C.M., Hanington, P.C., Lun, C., Rosenberg, G.H., Aragon, A.D., Stout, B.A.,Lennard Richard, M.L., Gross, P.S., Loker, E.S., 2010. Differential transcriptomicresponses of Biomphalaria glabrata Gastropoda, Mollusca to bacteria andmetazoan parasites, Schistosoma mansoni and Echinostoma paraensei digenea,platyhelminthes. Molecular Immunology 47, 849–860.
Bakry, F.A., 2009. Genetic variation between Biomphalaria alexandrina andBiomphalaria glabrata snails and their infection with Egyptian strain ofSchistosoma mansoni. Journal of Applied Sciences Researches 5, 1142–1148.
Bayne, C.J., Yashino, T.P., 1989. Determination of compatibility in mollusca-trematode parasitism. American Zoologist 29, 399–406.
Bayne, C.J., Buckely, P.M., Dewan, P.C., 1980. Schistosoma mansoni cytotoxicity ofhemocytes from susceptible snail hosts for sporocysts in plasma from resistantBiomphalaria glabrata. Experimental Parasitology 50, 409–416.
Bayne, C.J., Hahn, U.K., Bender, R.C., 2001. Mechanisms of molluscan host resistanceand of parasite strategies for survival. Parasitology 123, 159–167.
Caldeira, R.L., Jannotti-Passos, L.K., Carvalho, O.S., 2009. Molecular epidemiology ofBrazilian Biomphalaria: a review of the identification of species and thedetection of infected snails. Acta Tropica 111, 1–6.
Carton, Y., Nappi, A.J., Poirie, M., 2005. Genetics of anti-parasite resistance ininvertebrates. Developmental and Comparative Immunology 29, 9–32.
Chernin, E., Dunavan, C.A., 1962. The influence of host–parasite dispersion upon thecapacity of Schistosoma mansoni miracidia to infection Australorbis glabratus.American Journal of Tropical Medicine and Hygiene 11, 455–471.
Davies, S.J., Mckerrow, J.H., 2003. Developmental plasticity in schistosomes andother helminths. International Journal of Parasitology 33, 1277–1284.
deJong-Brink, M., Bergamin-Sassen, M., Solis Soto, M., 2001. Multiple strategies ofschistosomes to meet their requirements in the intermediate snail host.Parasitology 123, 129–141.
Frandsen, F., 1979. Studies on the relationship between Schistosoma and theirintermediate hosts. III. The genus Biomphalaria and Schistosoma mansoni fromEgypt, Kenya, Uganda, West Indies and Zaire two different strains from Katangaand Kinshasha. Journal of Helminthology 53, 321–348.
Gandon, S., Capowiez, Y., Dubois, Y., Michalakis, Y., Olivieri, I., 1996. Localadaptation and gene – for gene coevolution in a metapopulation model.Proceedings of the Royal Society of London Series B 263, 1003–1009.
Granath, W.O., Yashino, T.P., 1983. Lysosomal enzymes activities in susceptible andrefractory snail groups of Biomphalaria glabrata during the course of infectionwith Schistosoma mansoni. Journal of Parasitology 69, 1018–1026.
Guaraldo, A.M., Magalhães, L.A., Rangel, H.A., Pareja, G., 1981. Evolução dosesporocistos de Schistosoma mansoni Sambon, 1907, em Biomphalaria glabrataSay, 1818 e Biomphalaria tenagophila D’Orbigny, 1835. Revista de Saúde Pública15, 436–448.
Hahn, U.K., Bender, R.C., Bayne, C.J., 2001. Killing of Schistosoma mansoni sporocystsby hemocytes from resistant Biomphalaria glabrata: role of reactive oxygenspecies. Journal of Parasitology 87, 292–299.
Hanington, P.C., Lun, C., Adema, C.M., Loker, E.S., 2010. Time series analysis of thetranscriptional responses of Biomphalaria glabrata throughout the course ofintramolluscan development of Schistosoma mansoni and Echinostoma paraensei.International Journal of Parasitology 40, 819–831.
Lemos, Q.T., Andrade, Z.A., 2001. Sequential histological changes in Biomphalariaglabrata during the course of Schistosoma mansoni infection. Memorias doInstituto Oswaldo Cruz 96, 719–721.
Lively, C.M., 1999. Migration, virulence, and the geographic mosaic of adaptation byparasites. American Naturalist 153, 34–47.
Loker, E.S., Bayne, C.J., 1982. In vitro encounters between Schistosoma mansoniprimary sporocysts and hemolymph components of susceptible and resistantsnail groups of Biomphalaria glabrata. American Journal of Tropical Medicineand Hygiene 31, 999–1005.
Loker, E.S., Bayne, C.J., 1986. Immunity to trematode larvae in the snailBiomphalaria. In: Lackie, A.M. (Ed.), Immune Mechanisms in VertebrateVectors. Oxford University Press, New York, pp. 199–220.
Loker, E.S., Bayne, C.J., Yui, M.A., 1986. Echinostoma paraensei: hemocytes ofBiomphalaria glabrata as targets of Echinostoma mediate interference with hostsnail’s resistance to Schistosoma mansoni. Experimental Parasitology 62, 149–154.
Loker, E.S., Bayne, C.J., Buckley, P.M., Kruse, K.T., 1982. Ultrastructure ofencapsulation of Schistosoma mansoni mother sporocysts by hemocytes of
juveniles of the 10-R2 snail group of Biomphalaria glabrata. Journal ofParasitology 68, 84–94.
Martins-Souza, R.L., Pereira, C.A., Coelho, P.M., Negrão-Corrêa, D., 2003. Silicatreatment increases the susceptibility of the Cabo Frio snail group ofBiomphalaria tenagophila to Schistosoma mansoni infection but does not alterthe natural resistance of the Taim snail group. Parasitology Research 91, 500–507.
McLaren, D.J., Terry, R.J., 1982. The protective role of acquired host antigens duringschistosome maturation. Parasite Immunology 4, 129–148.
Meuleman, E.A., Bayne, C.J., van Der Knaap, W.P., 1987. Immunological aspects ofsnail–trematode interactions. Progress in Clinical and Biological Research 233,113–127.
Mohamed, A.H., 1998. Characterization of the Hemocytes of Biomphalaria glabrata,Say 1818 Gastropoda: Planorbidae in Interaction with two Snail Groups ofSchistosoma mansoni Sambon 1907 Trematoda. Ph.D. Thesis. Faculty of Biology,Tübingen University, Germany.
Mohamed, S.H., Saoud, M.F., Rivera, E.R., Bruce, J.I., 1993. Granulocytes andhyalinocytes in Biomphalaria alexandrina resisting infection with the PuertoRican snail group of Schistosoma mansoni. Abstract SRP Conference Cairo Egypt17, 165–178.
Newton, W.L., 1952. The comparative tissue reaction of two snail groups ofAustralorbis glabratus to infection with Schistosoma mansoni. Journal ofParasitology 38, 362–366.
Pan, C.T., 1965. Studies on the host–parasite relationship between Schistosomamansoni and the snail Australorbis glabratus. American Journal of TropicalMedicine and Hygiene 14, 931–976.
Pan, C.T., 1980. The fine structure of miracidium of Schistosoma mansoni. Journal ofInvertebrate Pathology 36, 307–372.
Paraense, W.L., Corrêa, L.R., 1963. Variation in susceptibility of population ofAustralorbis glabratus to a strain of Schistosoma mansoni. Revista do Instituto deMedicina. Tropical de Sao Paulo 5, 15–22.
Parker, G.A., Chubb, J.C., Ball, M.A., Roberts, G.N., 2003. Evolution of complexlifecycles in helminth parasites. Nature 425, 480–484.
Philips, T.L., Shoulberg, N., Gherson, J., 1984. Role of cellular and humoralcomponents in the encapsulation response of Biomphalaria glabrata toSchistosoma mansoni sporocysts in vitro. Recognition Proteins, Recept Probes:Invertebrates, 17–29.
Richards, C.S., Merritt, J.W., 1972. Genetic factors in the susceptibility for juvenileBiomphalaria glabrata to Schistosoma mansoni infection. American Journal ofTropical Medicine and Hygiene 21, 425–434.
Richards, C.S., Shade, P.C., 1987. The genetic variation of compatibility inBiomphalaria glabrata and Schistosoma mansoni. Journal of Parasitology 3,1146–1151.
Romeis, B., 1989. Mikroskopische Technik. Auflage, Urban and Schwarzenb-erg,Munchen – Wien-Baltimore 17, 235–236.
Sokal, R.R., Rohlf, F.J., 1995. Biometry: The Principles and Practice of Statistics inBiological Research, third ed. W.H. Freeman and Co., New York. 887pp. ISBN: 0-7167-2411-1.
Souza, C.P., Cunha, R.C., Andrade, Z.A., 1995. Development of Schistosoma mansoni inBiomphalaria tenagophila, Biomphalaria straminea and Biomphalaria glabrata.Revista do Instituto de Medicina. Tropical de Sao Paulo 37, 201–206.
Sullivan, J.T., Hu, P.C., 1995. A method for immunoisolation of sporocysts ofSchistosoma mansoni in non-susceptible snails. Journal of Parasitology 81, 1029–1031.
Sullivan, J.T., Richards, C.S., 1981. Schistosoma mansoni, NIH-SM-PR-PR snail groupin susceptible and non-susceptible stocks of Biomphalaria glabrata: comparativehistology. Journal of Parasitology 67, 702–708.
Théron, A., Coustau, C., 2005. Are Biomphalaria snails resistant to Schistosomamansoni? Journal of Helminthology 79, 187–191.
Théron, A., Pages, J.R., Rognon, A., 1997. Schistosoma mansoni: distribution patternsof miracidia among Biomphalaria glabrata snail as related to host susceptibilityand sporocyst regulatory processes. Experimental Parasitology 85, 1–9.
van der Knaap, W.P., Loker, E.S., 1990. Immune mechanisms in trematode–snailinteractions. Parasitology Today 6, 175–182.
Yousif, F., Ibrahim, A., El-Bardicy, S.N., 1998. Compatibility of Biomphalariaalexandrina, Biomphalaria glabrata and a hybrid of both to seven strains ofSchistosoma mansoni from Egypt. Journal of the Egyptian Society of Parasitology28, 863–881.
794 A.H. Mohamed et al. / Experimental Parasitology 127 (2011) 789–794


















![THE EFFECT OF SALINITY AND TEMPERATURE ON EGESTION IN MUD SNAILS (GASTROPODA: HYDROBIIDAE). A Study on Niche Overlap. [1975]](https://static.fdokumen.com/doc/165x107/6322ef93050768990e10148b/the-effect-of-salinity-and-temperature-on-egestion-in-mud-snails-gastropoda-hydrobiidae-1677547449.jpg)

