![THE EFFECT OF SALINITY AND TEMPERATURE ON EGESTION IN MUD SNAILS (GASTROPODA: HYDROBIIDAE). A Study on Niche Overlap. [1975]](https://static.fdokumen.com/doc/165x107/6322ef93050768990e10148b/the-effect-of-salinity-and-temperature-on-egestion-in-mud-snails-gastropoda-hydrobiidae-1677547449.jpg)
Brachylaima aspersae n. sp. (Digenea: Brachylaimidae) infecting farmed snails in NW Spain:...
14
Veterinary Parasitology 175 (2011) 273–286 Contents lists available at ScienceDirect Veterinary Parasitology journal homepage: www.elsevier.com/locate/vetpar Brachylaima aspersae n. sp. (Digenea: Brachylaimidae) infecting farmed snails in NW Spain: Morphology, life cycle, pathology, and implications for heliciculture P. Segade, C. Crespo, N. García, J.M. García-Estévez, C. Arias, R. Iglesias ∗ Laboratorio de Parasitología, Facultad de Biología, Edificio de Ciencias Experimentales, Campus de Lagoas-Marcosende s/n, Universidad de Vigo, 36310 Vigo, Spain article info Article history: Received 23 June 2010 Received in revised form 11 October 2010 Accepted 12 October 2010 Keywords: Brachylaima aspersae Helix aspersa Life cycle Pathology Snail farming Heliciculture abstract The life cycle of Brachylaima aspersae n. sp. (Trematoda: Brachylaimidae) in heliciculture farms is elucidated in light of field and experimental studies. Embryonated asymmetrical eggs (33.3 m × 20.2 m) are passed in the faeces of the definitive host, the domestic mouse (Mus musculus), and are ingested by its unique first intermediate host, the helicid snail Helix aspersa aspersa. After hatching, the miracidium develops into a highly branched sporocyst in the connective tissues of the digestive gland. Microcaudate cercariae emerging from this gastropod migrate up the ureter of the second intermediate host, the snails H. a. aspersa and H. a. maxima, and develop into non-encysted metacercariae in the kidney. Following predation of infected snails, the metacercariae develop into adults preferentially in the proximal portion of the duodenum of the definitive host. The strict oioxenic character for the first intermediate host, as well as the cercarial chaetotaxy (3 C I V+1C I D, 10 C II ,5C III V, 14 C III L, 2 C III D, 16 H, 6 S I ,6S II ,6S III ,2A I L+1A I V, 1 A II L, 3 ML, 1 P I L and 3 P III L), the distinct pars prostatica, the variable appearance of testes (rounded to irregular, with smooth or slightly to moderately lobulated margins), the size of eggs, the position of acetabulum (located somewhat posterior to the anterior third of body), and the microhabitat of the adult in the final host allow differentiation of B. aspersae from other well-known species in the genus. Massive infections with sporocysts or metacercariae of this brachylaimid may induce extensive pathological changes in the organs affected. Our results confirm that control of rodents in heliciculture farms is essential to minimize the potential health risks and morbimortality associated with this newly described species. © 2010 Elsevier B.V. All rights reserved. 1. Introduction Heliciculture (snail farming) is an emerging zootech- nic activity in NW Spain, where its economic viability is being seriously threatened by severe mortalities occurring in farms based on intensive (indoor breeding and fattening) or mixed (indoor breeding and outdoor fattening) pro- duction systems. Overcrowding associated with intensive ∗ Corresponding author. Tel.: +34 986812394; fax: +34 986812565. E-mail address: [email protected] (R. Iglesias). rearing can affect the activity, growth, reproductive effi- ciency and survival of Helix aspersa not only in a direct way, as a consequence of the negative effects of excreta and mucus (Hezberg, 1965; Cameron and Carter, 1979; Chevalier, 1979; Dan and Bailey, 1982; Jess and Marks, 1995; Mayoral et al., 2004), but also indirectly by facili- tating the transmission of certain pathogens. Thus, some directly transmitted parasites, like the ciliate Tetrahymena rostrata, the mite Riccardoella limacum and the nematode Alloionema appendiculatum, which are considered poten- tially pathogenic to terrestrial gastropods (Cooper and Knowler, 1991), have been found particularly abundant in 0304-4017/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.vetpar.2010.10.026
Transcript of Brachylaima aspersae n. sp. (Digenea: Brachylaimidae) infecting farmed snails in NW Spain:...

Bsf
PLU
a
ARRA
KBHLPSH
1
nbiod
0d
Veterinary Parasitology 175 (2011) 273–286
Contents lists available at ScienceDirect
Veterinary Parasitology
journa l homepage: www.e lsev ier .com/ locate /vetpar
rachylaima aspersae n. sp. (Digenea: Brachylaimidae) infecting farmednails in NW Spain: Morphology, life cycle, pathology, and implicationsor heliciculture
. Segade, C. Crespo, N. García, J.M. García-Estévez, C. Arias, R. Iglesias ∗
aboratorio de Parasitología, Facultad de Biología, Edificio de Ciencias Experimentales, Campus de Lagoas-Marcosende s/n,niversidad de Vigo, 36310 Vigo, Spain
r t i c l e i n f o
rticle history:eceived 23 June 2010eceived in revised form 11 October 2010ccepted 12 October 2010
eywords:rachylaima aspersaeelix aspersaife cycleathologynail farmingeliciculture
a b s t r a c t
The life cycle of Brachylaima aspersae n. sp. (Trematoda: Brachylaimidae) in heliciculturefarms is elucidated in light of field and experimental studies. Embryonated asymmetricaleggs (33.3 �m × 20.2 �m) are passed in the faeces of the definitive host, the domestic mouse(Mus musculus), and are ingested by its unique first intermediate host, the helicid snail Helixaspersa aspersa. After hatching, the miracidium develops into a highly branched sporocystin the connective tissues of the digestive gland. Microcaudate cercariae emerging from thisgastropod migrate up the ureter of the second intermediate host, the snails H. a. aspersaand H. a. maxima, and develop into non-encysted metacercariae in the kidney. Followingpredation of infected snails, the metacercariae develop into adults preferentially in theproximal portion of the duodenum of the definitive host. The strict oioxenic character forthe first intermediate host, as well as the cercarial chaetotaxy (3 CIV + 1 CID, 10 CII, 5 CIIIV,14 CIIIL, 2 CIIID, 16 H, 6 SI, 6 SII, 6 SIII, 2 AIL + 1 AIV, 1 AIIL, 3 ML, 1 PIL and 3 PIIIL), the distinctpars prostatica, the variable appearance of testes (rounded to irregular, with smooth orslightly to moderately lobulated margins), the size of eggs, the position of acetabulum
(located somewhat posterior to the anterior third of body), and the microhabitat of theadult in the final host allow differentiation of B. aspersae from other well-known speciesin the genus. Massive infections with sporocysts or metacercariae of this brachylaimidmay induce extensive pathological changes in the organs affected. Our results confirm thatcontrol of rodents in heliciculture farms is essential to minimize the potential health risksy assoc
and morbimortalit. Introduction
Heliciculture (snail farming) is an emerging zootech-ic activity in NW Spain, where its economic viability is
eing seriously threatened by severe mortalities occurringn farms based on intensive (indoor breeding and fattening)r mixed (indoor breeding and outdoor fattening) pro-uction systems. Overcrowding associated with intensive
∗ Corresponding author. Tel.: +34 986812394; fax: +34 986812565.E-mail address: [email protected] (R. Iglesias).
304-4017/$ – see front matter © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.vetpar.2010.10.026
iated with this newly described species.© 2010 Elsevier B.V. All rights reserved.
rearing can affect the activity, growth, reproductive effi-ciency and survival of Helix aspersa not only in a directway, as a consequence of the negative effects of excretaand mucus (Hezberg, 1965; Cameron and Carter, 1979;Chevalier, 1979; Dan and Bailey, 1982; Jess and Marks,1995; Mayoral et al., 2004), but also indirectly by facili-tating the transmission of certain pathogens. Thus, some
directly transmitted parasites, like the ciliate Tetrahymenarostrata, the mite Riccardoella limacum and the nematodeAlloionema appendiculatum, which are considered poten-tially pathogenic to terrestrial gastropods (Cooper andKnowler, 1991), have been found particularly abundant in
Parasit
274 P. Segade et al. / VeterinaryH. aspersa snails reared under intensive conditions (Morandand Daguzan, 1986; Cabaret et al., 1988; Graham et al.,1996; Segade et al., 2009). Identifying snail parasites andtheir reservoirs/sources of infection under farming condi-tions is therefore crucial to reduce the potential adverseeffects of these agents, and minimize the potential risks forhuman health derived from the presence of certain para-sites in edible snails. In this sense, the digenetic trematodeBrachylaima cribbi, which infects H. aspersa and other heli-cid species in Australia (Butcher and Grove, 2001), has beenfound parasitizing humans in that country (Butcher et al.,1996, 1998).
Previous studies conducted in Spain have reported thepresence of Brachylaima metacercariae in H. aspersa andother commercial snails, although the specific identity ofthese trematodes was not determined (González-Morenoet al., 1994). Subsequent investigations have described twospecies, B. mascomai and B. llobregatensis, that use this snailas first and/or second intermediate host in the LlobregatDelta (Catalonia, Spain) (Gracenea and González-Moreno,2002; González-Moreno and Gracenea, 2006).
In the present work, we describe the life cycle andthe different developmental stages of a new Brachylaimaspecies infecting H. aspersa snails in heliciculture farms. Inaddition, histopathological changes associated with its lar-val stages are described. Finally, some important aspectsconcerning the epidemiology of this species under farmingconditions and its importance from veterinary and sanitaryperspectives are discussed.
2. Materials and methods
2.1. Field studies in heliciculture farms
The subspecies Helix aspersa aspersa (petit-gris) and H.a. maxima (gros-gris), which are well adapted to farming,were included in the study. Both snails exhibit a strongtendency to reproductive isolation and differ morphologi-cally (e.g. shell size, mantle edge color, and conformationof the copulatory apparatus) and allozymically (Gomot-deVaufleury and Borgo, 2001).
A total of 1105 H. a. aspersa and 805 H. a. maximaadults belonging to 13 groups were collected between 2005and 2007 from 3 mixed rearing system-based heliciculturefarms located in Galicia (NW Spain) and examined for thepresence of digenetic trematodes. The farms were locatedin Valga (42◦42′N, 8◦38′W), Serra de Outes (42◦54′N,8◦51′W), and O Rosal (41◦55′N, 8◦47′W). Excepting thesnails in groups 7, 8 and 13, which were from green-houses, all helicids examined were collected from buildingswhere reproduction is carried out at high densities andunder controlled environmental conditions. According tothe persons in charge of the farms at Valga and Serra deOutes, snails belonging to groups 1 and 6 had been sup-plied by the farm at O Rosal, while snails in the remaininggroups (2–5 and 9–12) had been purchased from differ-
ent minor producers, whose identity and location werenot released. Each gastropod was anaesthetized by injec-tion of a sterile 50 mM MgCl2 solution and examinedfor trematode infection. Organs were isolated, exam-ined externally, disgregated individually in distilled water,ology 175 (2011) 273–286
and the number of parasites released counted under thestereomicroscope.
Fourteen house mice (Mus musculus) were capturedin the farm at O Rosal, because of the high prevalenceof brachylaimid sporocysts and metacercariae observedin snails from these facilities. In addition, according tothe producers, the presence of mice was not uncommonin this farm. Mice were live-trapped with Sherman trapsbaited with rolled oats and peanut butter. After sacrifice,the intestinal tracts were isolated and fragmented in threesegments (anterior, middle and posterior), which were lon-gitudinally opened and examined for the presence of adulttrematodes under the stereomicroscope.
2.2. Life cycle experimental studies
Mature metacercariae from the kidneys of H. a. aspersaand H. a. maxima were inoculated by intra-oesophagealtube to C57BL/6J Mus musculus mice (30 metacercariaeper mouse). This inbred strain was selected on the basisof its high susceptibility to Brachylaima cribbi (Butcheret al., 2002). After detecting the presence of brachy-laimid eggs in faeces, mice were sacrificed by cervicaldislocation 21 days postinfection and their complete diges-tive tracts were examined under the stereomicroscopein order to detect adult worms. Eggs were obtained bymanual homogenization of gravid trematodes found inexperimentally and naturally infected mice, mixed withlettuce leaves homogenized in mineral water and fedto laboratory-reared parasite-free H. a. aspersa and H. a.maxima snails. Different groups of gastropods were usedfor eggs coming from adult worms infecting inbred andwild mice. Cercariae shedding was monitored periodi-cally by placing the helicids in a Petri dish containinga thin layer of mineral water and subsequent examina-tion under a stereomicroscope. Non-shedding snails weredissected and examined for the presence of sporocysts.Once cercariae began to emerge from infected snails, newlaboratory-reared parasite-free snails were infected by 2 hexposure to cercariae in the Petri dishes as well as cohabita-tion with cercariae-shedding snails. Mature metacercariaewere removed from the kidney of experimentally infectedsnails 28 weeks postinfection and given to C57BL/6J miceto maintain the parasite’s life cycle.
2.3. Light and scanning electron microscopy studies
Adults, sporocysts, cercariae and metacercariae wereobtained by dissection of naturally and experimentallyinfected hosts. All stages, including the eggs, were studiedalive in an Olympus BX41 microscope. Cercariae, metac-ercariae and adults were also fixed in Bouin’s fluid underlight coverslip pressure and stained with Semichon’s ace-tocarmine. All measurements were made according toMas-Coma et al. (1984) except for the distance betweensuckers that was taken between the centres of both sucto-
rial organs to facilitate the comparison of our species withthose described by Gracenea and González-Moreno (2002)and González-Moreno and Gracenea (2006) in Spain. Cer-carial chaetotaxy was studied after impregnation withsilver nitrate (Combes et al., 1976). For a better visualiza-
Parasit
twttwd
c0w1ae(Sgps
2
efptHh
3
s(ea
3s
3
n
TMr
P. Segade et al. / Veterinary
ion of the H cephalic papillae, some impregnated cercariaeere mounted in apical view in Kaiser’s glycerinated gela-
ine. The nomenclature of argentophilic papillae followedhat described by Bayssade-Dufour (1979). Photographsere taken using an Olympus DP11 digital camera. Allrawings were prepared with the aid of a camera lucida.
For scanning electron microscopy, cercariae, metacer-ariae and adults were fixed in 2.5% (v/v) glutaraldehyde in.1 M cacodylate buffer, pH 7.2 for 2 h at 4 ◦C. After threeashes in the same buffer, trematodes were postfixed in
% (w/v) osmium tetroxide in the same buffer for 30 mint 4 ◦C, washed thoroughly, dehydrated in a grade series ofthanol and chemically dried with hexamethyldisilazaneSigma–Aldrich Chemie, GmbH., Steinheim, Germany).pecimens were subsequently mounted on metal stubs,old coated with a sputter coater, and observed andhotographed with a Philips XL30 scanning electron micro-cope.
.4. Histopathological studies
The digestive glands and renal organs from somexperimentally infected H. a. aspersa snails were fixedor 24–48 h in 10% buffered formalin and embedded inaraffin following standard histological procedures. Sec-ions (4–5 �m) were obtained using a microtome MicromM-340E (Microm, Walldorf, Germany) and stained withaematoxylin–eosin.
. Results
Farmed snails were infected with metacercariae and/orporocysts belonging to a new brachylaimid speciesTable 1). The morphology, life cycle and infection param-ters as well as the pathology associated with the parasitere detailed below.
.1. Morphological description of Brachylaima aspersa n.
p..1.1. Eggs and miracidium (Fig. 1)Embryonated eggs (in mouse faeces and in the termi-
al uterus of adult worms) asymmetrically ovoid (one side
able 1ain characteristics of the 13 groups of snails examined for the presence of bra
ange of intensities (RI) of larval stages of B. aspersae n. sp. for each group. Parasit
Group Snail species Farm location n
1 H. a. aspersa Valga 3182 H. a. aspersa Valga 3143 H. a. aspersa Valga 504 H. a. aspersa Valga 325 H. a. aspersa Valga 506 H. a. aspersa Outes 2007 H. a. aspersa O Rosal 508 H. a. aspersa O Rosal 919 H. a. maxima Outes 20010 H. a. maxima Outes 17511 H. a. maxima Outes 20012 H. a. maxima Outes 18013 H. a. maxima O Rosal 50
ology 175 (2011) 273–286 275
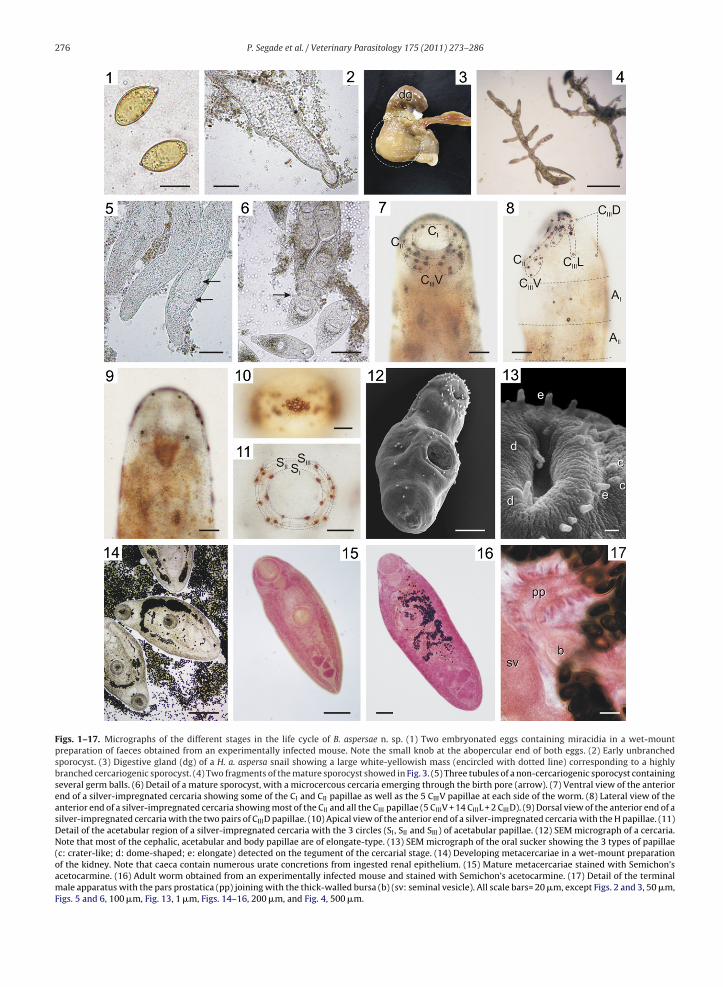
slightly flattened), 33.3 × 20.2 (31–35 × 18–25) �m in size(n = 50), with smooth light brown shell, a small abopercularknob, and well-developed miracidium.
3.1.2. Sporocyst (Figs. 2–6)Early non branched sporocysts (detected only in 2 cases)
fusiform, with smooth thick tegument and showing slowmovement in fresh preparations (Fig. 2). In heavily infectedsnails, highly branched sporocysts appear macroscopicallyas large white-yellowish masses occupying a great part ofthe digestive gland (Fig. 3). Microscopically, they exhibitprimary and secondary branched tubules containing germballs, developing and/or fully mature cercariae dependingon the stage of development (Figs. 4–6). Birth pores (exitfor cercariae) at the terminal tip of the branches (Fig. 6).
3.1.3. Cercaria (Table 2, Figs. 6–13, 18, and 23–25)The cercarial morphology (Figs. 6, 12, and 18) fol-
lows the general pattern of other brachylaimid cercariae:microcercous, elongate body lacking stylet, with tegu-ment covered by discrete conical spines, and with distinctbulbous pharynx, 2 intestinal caeca extending parallelto body margins just to the middle-posterior region ofventral sucker, postacetabular genital primordium, andstenostomate excretory system with main excretory ductsopening into lateral branches of a Y-shaped excretoryvesicle located at posterior ends. No conspicuous glandcells are visible at the end of caeca. Oral sucker slightlylarger than acetabulum. Chaetotaxy as follows: (1) cephalicpapillae (Figs. 7–10 and 23–25): 3 CIV + 1 CID pairs, withthe CID pair found deep within mouth, 10 CII pairs reg-ularly distributed in a circle around mouth, 21 CIII pairs(5 CIIIV + 14 CIIIL + 2 CIIID), and 16 H papillae arranged ina cluster on anterior end of body; (2) acetabular papillae(Figs. 11 and 23, 24): 6 SI, arranged evenly around the innermargin of the sucker, 6 SII, and 6 SIII (rarely 5 or 7) usu-ally disposed as 4 antero-lateral papillae (one pair at eachside) and 2 postero-lateral papillae (SIII circle only slightly
larger than SII); (3) body papillae (Figs. 8, 12 and 23, 24): 2AIL pairs + 1 AIV pair (sometimes only one unpaired papillapresent at one side), 1 AIIL pair, 3 ML pairs, 1 PIL pair and3 PIIIL pairs; and (4) caudal papillae: always absent. Nochaetotaxic differences were observed between naturallychylaimid parasites, with prevalences (P(%)), mean intensities (I) and/orological parameters were used according to Bush et al. (1997).
Metacercariae Sporocysts
P (%) I RI P (%)
79.7 10.3 1–76 1216.6 6.3 1–33 2.218 2.8 1–10 496.9 32.1 2–97 010 3.4 1–7 080 12.8 1–57 2.596 35.8 1–94 259.3 4.3 1–25 1.1
2.5 0.7 1–2 09 3.0 1–30 08 2.3 1–7 07.3 1.1 1–36 0
38 4.8 1–23 0
276 P. Segade et al. / Veterinary Parasitology 175 (2011) 273–286
Figs. 1–17. Micrographs of the different stages in the life cycle of B. aspersae n. sp. (1) Two embryonated eggs containing miracidia in a wet-mountpreparation of faeces obtained from an experimentally infected mouse. Note the small knob at the abopercular end of both eggs. (2) Early unbranchedsporocyst. (3) Digestive gland (dg) of a H. a. aspersa snail showing a large white-yellowish mass (encircled with dotted line) corresponding to a highlybranched cercariogenic sporocyst. (4) Two fragments of the mature sporocyst showed in Fig. 3. (5) Three tubules of a non-cercariogenic sporocyst containingseveral germ balls. (6) Detail of a mature sporocyst, with a microcercous cercaria emerging through the birth pore (arrow). (7) Ventral view of the anteriorend of a silver-impregnated cercaria showing some of the CI and CII papillae as well as the 5 CIIIV papillae at each side of the worm. (8) Lateral view of theanterior end of a silver-impregnated cercaria showing most of the CII and all the CIII papillae (5 CIIIV + 14 CIIIL + 2 CIIID). (9) Dorsal view of the anterior end of asilver-impregnated cercaria with the two pairs of CIIID papillae. (10) Apical view of the anterior end of a silver-impregnated cercaria with the H papillae. (11)Detail of the acetabular region of a silver-impregnated cercaria with the 3 circles (SI, SII and SIII) of acetabular papillae. (12) SEM micrograph of a cercaria.Note that most of the cephalic, acetabular and body papillae are of elongate-type. (13) SEM micrograph of the oral sucker showing the 3 types of papillae(c: crater-like; d: dome-shaped; e: elongate) detected on the tegument of the cercarial stage. (14) Developing metacercariae in a wet-mount preparationof the kidney. Note that caeca contain numerous urate concretions from ingested renal epithelium. (15) Mature metacercariae stained with Semichon’sacetocarmine. (16) Adult worm obtained from an experimentally infected mouse and stained with Semichon’s acetocarmine. (17) Detail of the terminalmale apparatus with the pars prostatica (pp) joining with the thick-walled bursa (b) (sv: seminal vesicle). All scale bars= 20 �m, except Figs. 2 and 3, 50 �m,Figs. 5 and 6, 100 �m, Fig. 13, 1 �m, Figs. 14–16, 200 �m, and Fig. 4, 500 �m.
P. Segade et al. / Veterinary Parasitology 175 (2011) 273–286 277
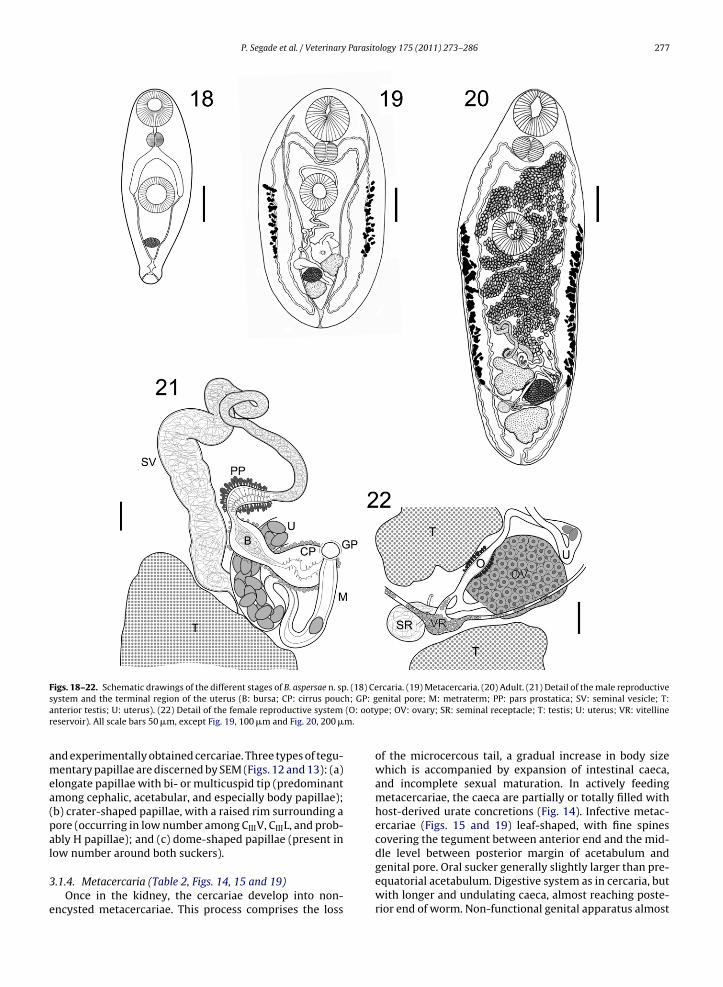
F . (18) Ces h; GP: ga (O: ootyr m.
amea(pal
3
e
igs. 18–22. Schematic drawings of the different stages of B. aspersae n. spystem and the terminal region of the uterus (B: bursa; CP: cirrus poucnterior testis; U: uterus). (22) Detail of the female reproductive systemeservoir). All scale bars 50 �m, except Fig. 19, 100 �m and Fig. 20, 200 �
nd experimentally obtained cercariae. Three types of tegu-entary papillae are discerned by SEM (Figs. 12 and 13): (a)
longate papillae with bi- or multicuspid tip (predominantmong cephalic, acetabular, and especially body papillae);b) crater-shaped papillae, with a raised rim surrounding aore (occurring in low number among CIIIV, CIIIL, and prob-bly H papillae); and (c) dome-shaped papillae (present in
ow number around both suckers)..1.4. Metacercaria (Table 2, Figs. 14, 15 and 19)Once in the kidney, the cercariae develop into non-
ncysted metacercariae. This process comprises the loss
rcaria. (19) Metacercaria. (20) Adult. (21) Detail of the male reproductiveenital pore; M: metraterm; PP: pars prostatica; SV: seminal vesicle; T:pe; OV: ovary; SR: seminal receptacle; T: testis; U: uterus; VR: vitelline
of the microcercous tail, a gradual increase in body sizewhich is accompanied by expansion of intestinal caeca,and incomplete sexual maturation. In actively feedingmetacercariae, the caeca are partially or totally filled withhost-derived urate concretions (Fig. 14). Infective metac-ercariae (Figs. 15 and 19) leaf-shaped, with fine spinescovering the tegument between anterior end and the mid-
dle level between posterior margin of acetabulum andgenital pore. Oral sucker generally slightly larger than pre-equatorial acetabulum. Digestive system as in cercaria, butwith longer and undulating caeca, almost reaching poste-rior end of worm. Non-functional genital apparatus almost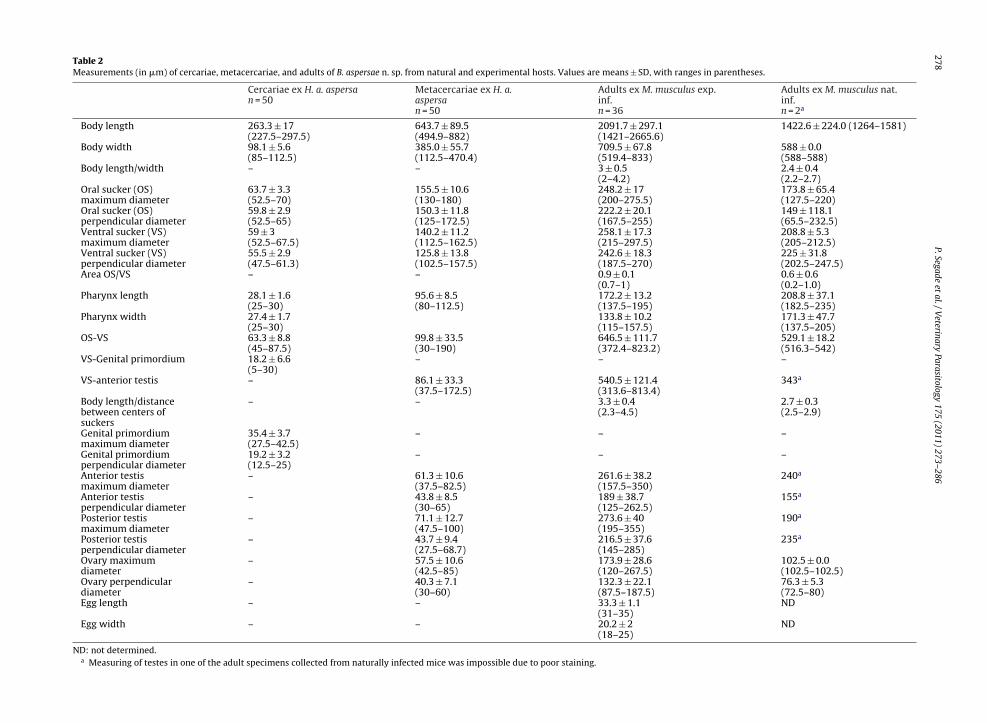
278P.Segade
etal./V
eterinaryParasitology
175 (2011) 273–286Table 2Measurements (in �m) of cercariae, metacercariae, and adults of B. aspersae n. sp. from natural and experimental hosts. Values are means ± SD, with ranges in parentheses.
Cercariae ex H. a. aspersan = 50
Metacercariae ex H. a.aspersan = 50
Adults ex M. musculus exp.inf.n = 36
Adults ex M. musculus nat.inf.n = 2a
Body length 263.3 ± 17(227.5–297.5)
643.7 ± 89.5(494.9–882)
2091.7 ± 297.1(1421–2665.6)
1422.6 ± 224.0 (1264–1581)
Body width 98.1 ± 5.6(85–112.5)
385.0 ± 55.7(112.5–470.4)
709.5 ± 67.8(519.4–833)
588 ± 0.0(588–588)
Body length/width – – 3 ± 0.5(2–4.2)
2.4 ± 0.4(2.2–2.7)
Oral sucker (OS)maximum diameter
63.7 ± 3.3(52.5–70)
155.5 ± 10.6(130–180)
248.2 ± 17(200–275.5)
173.8 ± 65.4(127.5–220)
Oral sucker (OS)perpendicular diameter
59.8 ± 2.9(52.5–65)
150.3 ± 11.8(125–172.5)
222.2 ± 20.1(167.5–255)
149 ± 118.1(65.5–232.5)
Ventral sucker (VS)maximum diameter
59 ± 3(52.5–67.5)
140.2 ± 11.2(112.5–162.5)
258.1 ± 17.3(215–297.5)
208.8 ± 5.3(205–212.5)
Ventral sucker (VS)perpendicular diameter
55.5 ± 2.9(47.5–61.3)
125.8 ± 13.8(102.5–157.5)
242.6 ± 18.3(187.5–270)
225 ± 31.8(202.5–247.5)
Area OS/VS – – 0.9 ± 0.1(0.7–1)
0.6 ± 0.6(0.2–1.0)
Pharynx length 28.1 ± 1.6(25–30)
95.6 ± 8.5(80–112.5)
172.2 ± 13.2(137.5–195)
208.8 ± 37.1(182.5–235)
Pharynx width 27.4 ± 1.7(25–30)
133.8 ± 10.2(115–157.5)
171.3 ± 47.7(137.5–205)
OS-VS 63.3 ± 8.8(45–87.5)
99.8 ± 33.5(30–190)
646.5 ± 111.7(372.4–823.2)
529.1 ± 18.2(516.3–542)
VS-Genital primordium 18.2 ± 6.6(5–30)
– – –
VS-anterior testis – 86.1 ± 33.3(37.5–172.5)
540.5 ± 121.4(313.6–813.4)
343a
Body length/distancebetween centers ofsuckers
– – 3.3 ± 0.4(2.3–4.5)
2.7 ± 0.3(2.5–2.9)
Genital primordiummaximum diameter
35.4 ± 3.7(27.5–42.5)
– – –
Genital primordiumperpendicular diameter
19.2 ± 3.2(12.5–25)
– – –
Anterior testismaximum diameter
– 61.3 ± 10.6(37.5–82.5)
261.6 ± 38.2(157.5–350)
240a
Anterior testisperpendicular diameter
– 43.8 ± 8.5(30–65)
189 ± 38.7(125–262.5)
155a
Posterior testismaximum diameter
– 71.1 ± 12.7(47.5–100)
273.6 ± 40(195–355)
190a
Posterior testisperpendicular diameter
– 43.7 ± 9.4(27.5–68.7)
216.5 ± 37.6(145–285)
235a
Ovary maximumdiameter
– 57.5 ± 10.6(42.5–85)
173.9 ± 28.6(120–267.5)
102.5 ± 0.0(102.5–102.5)
Ovary perpendiculardiameter
– 40.3 ± 7.1(30–60)
132.3 ± 22.1(87.5–187.5)
76.3 ± 5.3(72.5–80)
Egg length – – 33.3 ± 1.1(31–35)
ND
Egg width – – 20.2 ± 2(18–25)
ND
ND: not determined.a Measuring of testes in one of the adult specimens collected from naturally infected mice was impossible due to poor staining.

P. Segade et al. / Veterinary Parasitology 175 (2011) 273–286 279
F y of B. aC II, AIII , MS
ciswmvtcVtlvtc
3
osfiiaAlItctsimph
igs. 23–25. Schematic drawings of the characteristic cercarial chaetotaxIII and H are cephalic papillae, SI, SII and SIII are acetabular papillae, AI, Acale bar= 50 �m.
ompletely developed, with testes and ovary in tandemn posterior third of body. Testes, rounded to irregular inhape, with smooth to slightly lobulated margins, some-hat larger than the ovary, generally rounded, with smoothargins and located between testes. Genital pore opens
entrally in the midline of the body, slightly before or athe level of anterior margin of first testis. Uterus intracae-al reaching anteriorly almost the intestinal bifurcation.itelline follicles extracaecal extending anteriorly along
he lateral margins of the body from the anterior-middleevel of the first testis to almost the anterior margin of theentral sucker. Terminal organs of male reproductive sys-em difficult to distinguish and surrounded by numerousells.
.1.5. Adult worms (Table 2, Figs. 16, 17 and 20–22)Morphology of adults agrees with the general pattern
f the genus Brachylaima (Pojmanska, 2002a). Species-pecific characteristics are as follows. Body elongate withne tegument spinulation extending from anterior extrem-
ty to about middle level between posterior margin ofcetabulum and genital pore. Oral sucker subterminal.cetabulum generally slightly larger than oral sucker and
ocated somewhat posterior to anterior third of body.ntestinal caeca extends parallel to lateral margins to pos-erior end of the worm forming undulations and enlarginglose to distal end. Gonads in tandem in posterior quar-er of the body with intertesticular ovary usually displaced
lightly to side (Figs. 16 and 20). Testes rounded to irregularn shape, with smooth or slightly to moderately lobulatedargins, about 1.3–1.5 times larger than ovary, and theosterior slightly larger than the anterior one. Each testisas a short vas efferens that connects anteriorly with the
spersae n. sp. (23) Ventral view. (24) Lateral view. (25) Dorsal view. CI, CII,, PI, PII, PIII are body papillae, and U correspond to caudal papillae region.
other to form a vas deferens continuing forwardly into along and convoluted external seminal vesicle filled withsperm. Seminal vesicle leads to a distinct pars prostat-ica, surrounded by numerous gland-cells, that joins witha short thick-walled bursa, sometimes containing sperm,that narrows distally prior to opening into a globular thick-walled cirrus pouch containing a relatively short unarmedcirrus (Figs. 17 and 21). Pars prostatica, bursa and cirruspouch similar in length. The two latter, especially the cirruspouch, surrounded by numerous cells which seem differentto prostatic gland-cells. Genital pore, medial or submedial,opens ventrally in posterior third of body, slightly beforeor at the level of anterior margin of first testis (Fig. 20).Extracaecal vitelline folicles parallel to caeca extendingfrom the anterior margin of acetabulum or slightly pos-terior to it to middle level of first testis (Fig. 20). Vitellineducts arising from vitelline folicles run posteriorly to joinforming a vitelline reservoir located medially or subme-dially between the ovary and the second testis. Ovarygenerally rounded with smooth margins (rarely irregularin shape with slightly lobulated margins) (Fig. 22). Oviductjoins with duct arising from seminal receptacle (locatedbeside the ovary) before converging with main vitellineduct coming from vitelline reservoir. Beyond this junction,oviduct expands slightly to form the ootype which is sur-rounded by Mehlis’ glands and connects with the uterus.Intracaecal uterus, filled with eggs and terminating at mus-cular metraterm before opening at genital pore, extends
anteriorly reaching or overlapping (rarely exceeding) theintestinal bifurcation. Laurer’s canal arises dorsally fromseminal receptacle finally opening onto dorsal surface ofthe worm. Y-shaped excretory vesicle located intracaecallyat posterior end of the body. Excretory pore terminal.
Parasit
280 P. Segade et al. / Veterinary3.2. Life cycle
Our field and experimental results demonstrate thatthe mouse, M. musculus, serves as definitive host. Preva-lences in naturally and experimentally infected mice were14.3% (2/14) and 64.1% (52/81), respectively. Adult wormsinhabit exclusively the duodenum, showing preference forthe region immediately after the pylorus. The caeca ofdigeneans contains host blood. Embryonated eggs weredetected in mouse faeces 21 days postinfection. H. a.aspersa snails, which serve as first intermediate hosts,are infected by ingesting the embryonated eggs. Onceingested, the miracidium hatches and develops into ahighly branched sporocyst in the connective tissue of thedigestive gland. In massive or long-term infections thisorgan can be almost totally invaded by the sporocyst net-work, which can also infiltrate the pulmonary, renal andeven gonadal tissues. Overall prevalence of sporocysts inthe 8 groups of petit-gris examined was 4.9%, rangingfrom 0 to 11.9% (Table 1). The prevalence in experimen-tally infected H. a. aspersa snails was much higher (57%).No sporocysts were detected in naturally or experimen-tally infected H. a. maxima snails, confirming the extremelyhigh host specificity of this brachylaimid species. Emergingof cercariae in experimentally infected gastropods started50–60 days postinfection. Cercariae enter the kidney of thesecond intermediate host via the ureter. Once in the renalsac, they feed on the renal epithelium, to develop into non-encysted fully mature metacercariae in 3–6 months. Noother organs such as pedal glands or pericardium werefound infected. Both H. a. aspersa and H. a. maxima serveas second intermediate host in heliciculture farms. Sincethe former subspecies can also act as first intermediatehost, petit-gris snails can be infected simultaneously withsporocysts and metacercariae. However, autoinfection isnot possible since cercariae are unable to infect the snailfrom which they are emerging. The life cycle is completedwhen an appropriate definitive host feeds on infected snailsand the mature metacercariae are liberated in the digestivetract.
3.3. Taxonomic summary
Type host: Mus musculus Linnaeus, 1758 (Rodentia:Muridae) as natural and experimental definitive host.
Site of infection: duodenum, with preference for theregion immediately posterior to the pylorus.
First intermediate host: Helix aspersa aspersa Müller,1774 (Gastropoda: Helicidae) as natural and experimentalhost.
Second intermediate hosts: Helix aspersa aspersa Müller,1774 and Helix aspersa maxima Taylor, 1883 (Gastropoda:Helicidae) as natural and experimental hosts.
Locality: Galicia, north western Spain.Type material: all specimens are deposited in the Inver-
tebrate Collection of Museo Nacional de Ciencias Naturales
(MNCN, Madrid, Spain). Sporocysts ex H. a. aspersa, experi-mental infection (MNCN 4.02/43); cercariae ex H. a. aspersa,experimental infection (MNCN 4.02/42); metacercariae exH. a. aspersa, natural infection (MNCN 4.02/41); adult holo-type (MNCN 4.02/38) and paratypes (MNCN 4.02/39) ex M.ology 175 (2011) 273–286
musculus C57BL/6J, experimental infection; adult paratypeex M. musculus, natural infection (MNCN 4.02/40).
Etymology: the specific name is derived from the sub-specific epithet of its unique first intermediate host, thesnail H. a. aspersa.
3.4. Histopathological findings
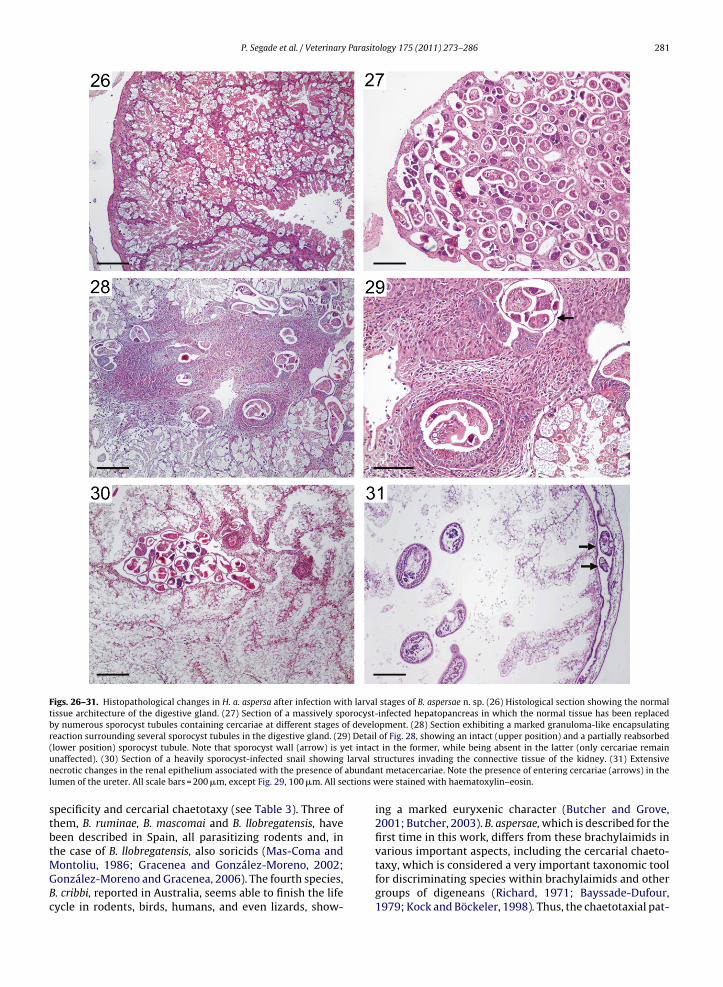
In heavily infected H. a. aspersa snails a great part ofthe digestive gland was extensively replaced by highlybranched sporocysts, with loss of the normal tissue archi-tecture in the areas directly affected by the parasite(Figs. 26 and 27), and a reduction in the lumen of adja-cent tubules resulting from the compressive effect ofthe larval stage. In some individuals, severe diffuse orfocal granuloma-like encapsulating lesions with abundantamoebocytes were observed surrounding intact or partiallydisintegrated structures of the parasite (Figs. 28 and 29).No encapsulating responses were observed in the kid-ney, although this organ was only occasionally infectedby sporocysts (Fig. 30). Regarding the metacercarial stage,extensive necrotic changes were observed in the renalepithelium of hosts which were infected by numerousactively feeding parasites (Fig. 31).
4. Discussion
The parasite described in the present work belongs tothe large genus Brachylaima Dujardin, 1843, which com-prises numerous species that complete their life cycles inmammals and birds throughout the world (Ubelaker andDailey, 1966; Yamaguti, 1971). This is, however, a veryproblematic and confused genus that includes many poorlyknown species, for which the description of the adult stageis the only information available (Yamaguti, 1971). Unfor-tunately, identifying new Brachylaima species only on thebasis of the morphoanatomical and morphometric char-acteristics of the adult stage may not provide enoughinformation to distinguish all combinations of species,since (1) the sympatry of two or more hardly distinguish-able species is not uncommon (Mas-Coma and Montoliu,1986) and (2) considerable allometric variation associatedwith adult growth and maturation has been demonstratedin certain species (Cribb, 1990, 1992). Faced with thesedifficulties, most authors currently agree that establishingnew Brachylaima species should be supported not only by adetailed description of all developmental stages, includingthe cercarial chaetotaxy, but also by additional informa-tion concerning the life cycle, host specificities, site(s) ofinfection, and geographical distribution of the digenean(Mas-Coma and Montoliu, 1986; Butcher and Grove, 2001;Gracenea and González-Moreno, 2002; González-Morenoand Gracenea, 2006). The range of final hosts is, however,another problematic aspect not only because current regu-lations make it difficult to conduct experimental infectionswith a broad range of parasite-free mammals and birds, but
also because the capture of naturally infected hosts shouldbe minimized to avoid negative effects on biodiversity.In spite of the numerous Brachylaima species relatedin the literature, only four have been described in depthto date, including information about their life cycle, host
P. Segade et al. / Veterinary Parasitology 175 (2011) 273–286 281
Figs. 26–31. Histopathological changes in H. a. aspersa after infection with larval stages of B. aspersae n. sp. (26) Histological section showing the normaltissue architecture of the digestive gland. (27) Section of a massively sporocyst-infected hepatopancreas in which the normal tissue has been replacedby numerous sporocyst tubules containing cercariae at different stages of development. (28) Section exhibiting a marked granuloma-like encapsulatingreaction surrounding several sporocyst tubules in the digestive gland. (29) Detail of Fig. 28, showing an intact (upper position) and a partially reabsorbed( et intacu larval sn abundanl ctions w
stbtMGBc
lower position) sporocyst tubule. Note that sporocyst wall (arrow) is ynaffected). (30) Section of a heavily sporocyst-infected snail showingecrotic changes in the renal epithelium associated with the presence of
umen of the ureter. All scale bars = 200 �m, except Fig. 29, 100 �m. All se
pecificity and cercarial chaetotaxy (see Table 3). Three ofhem, B. ruminae, B. mascomai and B. llobregatensis, haveeen described in Spain, all parasitizing rodents and, in
he case of B. llobregatensis, also soricids (Mas-Coma andontoliu, 1986; Gracenea and González-Moreno, 2002;onzález-Moreno and Gracenea, 2006). The fourth species,. cribbi, reported in Australia, seems able to finish the lifeycle in rodents, birds, humans, and even lizards, show-
t in the former, while being absent in the latter (only cercariae remaintructures invading the connective tissue of the kidney. (31) Extensivet metacercariae. Note the presence of entering cercariae (arrows) in theere stained with haematoxylin–eosin.
ing a marked euryxenic character (Butcher and Grove,2001; Butcher, 2003). B. aspersae, which is described for thefirst time in this work, differs from these brachylaimids in
various important aspects, including the cercarial chaeto-taxy, which is considered a very important taxonomic toolfor discriminating species within brachylaimids and othergroups of digeneans (Richard, 1971; Bayssade-Dufour,1979; Kock and Böckeler, 1998). Thus, the chaetotaxial pat-
Parasit
282 P. Segade et al. / Veterinarytern of B. ruminae includes 22 CIII, 18 H, 1 PIIL, 4 PIIIL, and3–4 SIII, among other papillae (Mas-Coma and Montoliu,1986). This pattern contrasts with the 21 CIII, 16 H, 0 PIIL,3 PIIIL, and 6 SIII papillae detected in B. aspersae. In addi-tion, B. ruminae exhibits a strict specificity for the subulinidsnail Rumina decollata as first intermediate host, its testesare oval and with smooth margins, its eggs are smaller(27.2–29.5 �m × 15.5–16 �m), and its acetabulum is moreanterior (the body length/distance between the centres ofsuckers ratio is about 4.5) (Mas-Coma and Montoliu, 1986).Similarly, the species B. mascomai, described by Graceneaand González-Moreno (2002), differs by having 13–15 CIII,1AIL and 1AIIIL, 1 PIIL, 4 PIIIL, 8–10 SII, and 4–5 SIII cercarialpapillae, smaller eggs (25.4 �m × 12.7 �m), more anterioracetabulum (the body length/distance between the cen-tres of suckers ratio is about 5.6), and non-lobulated ovaltestes, and by its unique first intermediate host, the helicidPseudotachea splendida. B. llobregatensis, like our species,exhibits a strict specificity for H. aspersa as first inter-mediate host. Nevertheless, no information was given byGonzález-Moreno and Gracenea (2006) about which sub-species, H. a. aspersa or H. a. maxima, was found infectedin their study. In any case, B. llobregatensis can be differ-entiated from B. aspersae by its cercarial chaetotaxy (5–6CI, 8–9 CII, 12–14 CIII, 1AIL, 1AIIIL, and 7–8 SIII papillae),the non-lobulated oval testes, and the adult microhabitatwhich seems clearly restricted to the middle portion of theintestine (González-Moreno and Gracenea, 2006).
Although B. cribbi has been only reported to date inAustralia, Cribb (1990) and Butcher and Grove (2001) havesuggested a possible European origin for this species. How-ever, the cercarial chaetotaxy (especially 5 CI, 11 CII, 19 CIII,1 PIIL, 1 PIIID, 12–14 SII and 0 SIII papillae), the broad range offirst intermediate hosts, represented by the helicid speciesTheba pisana, Cernuella virgata, Cochlicella barbara and C.acuta, the slightly smaller eggs (29.1 �m × 16.6 �m), thepresence of vitelline folicles never exceeding the poste-rior margin of the acetabulum, the slenderness of the adultstage (length/width ratio = 7.4), the more anterior posi-tion of the acetabulum (the body length/distance betweenthe centres of suckers ratio is about 5), and, finally, thenon-lobulated oval testes, allow this species to be easilydistinguished from B. aspersae (Butcher and Grove, 2001;Butcher, 2003).
In addition to these considerations, it must be under-lined that the male reproductive system of B. aspersae ischaracterized by the presence of a distinct pars prostatica.From the taxonomical point of view, the presence/absenceof this organ in digeneans belonging to the genus Brachy-laima is controversial. Thus, its presence is mentioned (as“prostatic complex”) in the generic description given byYamaguti (1971), but it is undetectable in all members ofthe family Brachylaimidae according to the taxonomic keygiven by Pojmanska (2002b). This confused situation is alsofound in the descriptions of the fourth species cited above.While no prostatic complex is described for B. ruminae, B.
mascomai, and B. llobregatensis (Mas-Coma and Montoliu,1986; Gracenea and González-Moreno, 2002; González-Moreno and Gracenea, 2006),’a short’ pars prostatica ismentioned in the description given by Butcher and Grove(2001) for B. cribbi. In light of these comments, this aspectology 175 (2011) 273–286
should be taken into consideration when new taxonomickeys concerning brachylaimid digeneans are developed.
As commented above, the comparison of B. aspersaewith poorly described Brachylaima species reported inEurope and North Africa is extremely difficult and prob-ably unreliable. In spite of this, some of these digeneansexhibit certain characteristics that allow differentiationfrom the present species. Thus, the acetabulum of B.simoni, which was originally reported infecting the sori-cid Crocidura russula in Ibiza (Balearic Islands, Spain), istwice larger than the oral sucker (Feliu et al., 1986). Inaddition, its testes are subspheric to oval with smoothmargins, and its eggs are smaller than those of B. asper-sae (26–27.5 �m × 14.5–16 �m). B. recurvum, for whichB. mesostomum and B. columbae have been postulated assynonyms (Macchioni, 1970), seems to be able to use sev-eral gastropod snails as first intermediate hosts (Yamaguti,1971). In addition, metacercariae of this species are not freein the kidney of second intermediate hosts, but encystedin the pedal glands (Timon-David, 1954, 1959), and theacetabulum is clearly more anterior than in B. aspersae(the body length/distance between the centres of suck-ers ratio is about 8; Joyeux and Foley, 1930). B. fuscatum,which is probably synonym of the questioned species B.nicolli (Dollfus, 1954; Macchioni, 1970), has been foundin the large intestine of birds, but never in experimen-tally and naturally infected mammals (Macchioni, 1968;Joyeux et al., 1934; Timon-David, 1965). Furthermore,sporocysts of this brachylaimid develop in the helicidsnail T. pisana (=Helix (Euparypha) pisana) (Joyeux et al.,1934). The microhabitat of B. fulvus and B. oesophagei isrestricted to the oesophagus and stomach of soricids, whilethe uterus of these species does not reach the anteriormargin of the acetabulum (Pojmanska, 1961; Lewis, 1969;Jourdane, 1971; Ubelaker and Dailey, 1966). Sporocystsof B. erinacei, for which B. caudatum is considered a syn-onym (Yamaguti, 1971), are able to develop in severalsnails belonging to the genera Helicella (=Xerophila), Helix,Arion and Succinea, while adults of this species lacks aspinous tegument and can measure up to 7 mm in length(Simón-Vicente, 1955a,b; Yamaguti, 1971; Butcher andGrove, 2001). Finally, according to Balozet (1937a,b), B. suisuses Xerophila snails as first intermediate hosts while thepreferred microhabitats for metacercariae and adults arethe pedal glands and the large intestine, respectively. Theresults obtained by this author have been questioned, how-ever, by Mas-Coma and Montoliu (1986).
Since no other hosts apart from the mouse M. mus-culus and the snails H. a. aspersa and H. a. maxima wereincluded in this study, we cannot exclude the possibilitythat other mammals, birds and terrestrial gastropods mayalso serve as final and second intermediate hosts for B.aspersae. In fact, this situation probably occurs since thefour Brachylaima species for which the entire life cyclehas been well documented are able to infect more thanone definitive or second intermediate host (Mas-Coma and
Montoliu, 1986; Butcher and Grove, 2001; Gracenea andGonzález-Moreno, 2002; Butcher, 2003; González-Morenoand Gracenea, 2006). Contrarily, a strict oioxenic specificityfor the first intermediate host is typically observed in thesedigeneans, except for B. cribbi which has been reported par-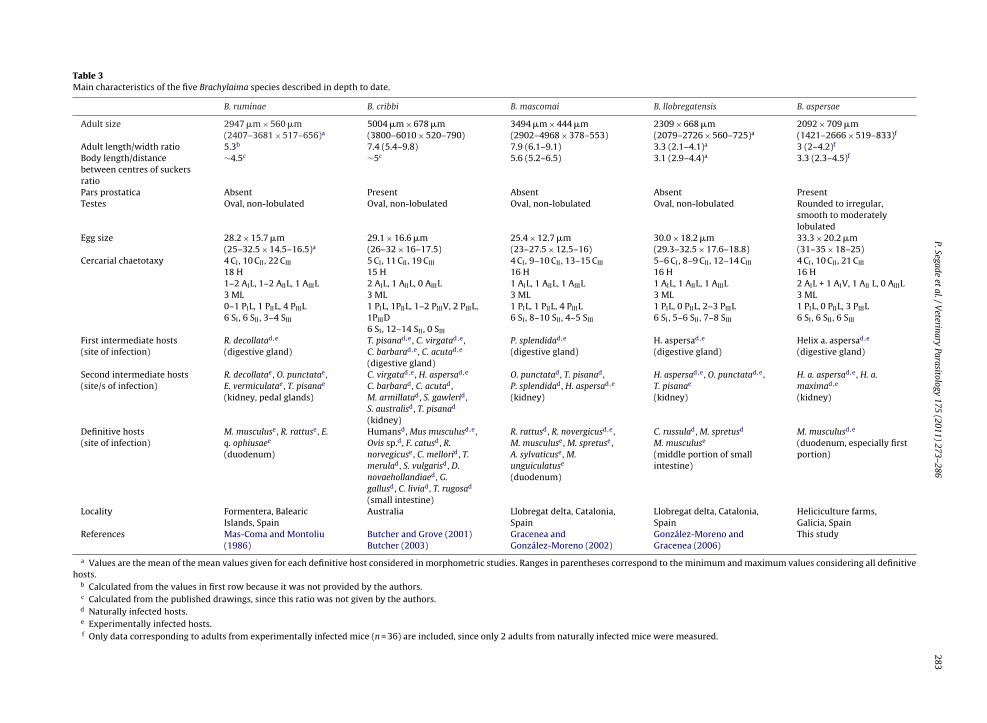
P.Segadeet
al./Veterinary
Parasitology175 (2011) 273–286
283
Table 3Main characteristics of the five Brachylaima species described in depth to date.
B. ruminae B. cribbi B. mascomai B. llobregatensis B. aspersae
Adult size 2947 �m × 560 �m(2407–3681 × 517–656)a
5004 �m × 678 �m(3800–6010 × 520–790)
3494 �m × 444 �m(2902–4968 × 378–553)
2309 × 668 �m(2079–2726 × 560–725)a
2092 × 709 �m(1421–2666 × 519–833)f
Adult length/width ratio 5.3b 7.4 (5.4–9.8) 7.9 (6.1–9.1) 3.3 (2.1–4.1)a 3 (2–4.2)f
Body length/distancebetween centres of suckersratio
∼4.5c ∼5c 5.6 (5.2–6.5) 3.1 (2.9–4.4)a 3.3 (2.3–4.5)f
Pars prostatica Absent Present Absent Absent PresentTestes Oval, non-lobulated Oval, non-lobulated Oval, non-lobulated Oval, non-lobulated Rounded to irregular,
smooth to moderatelylobulated
Egg size 28.2 × 15.7 �m(25–32.5 × 14.5–16.5)a
29.1 × 16.6 �m(26–32 × 16–17.5)
25.4 × 12.7 �m(23–27.5 × 12.5–16)
30.0 × 18.2 �m(29.3–32.5 × 17.6–18.8)
33.3 × 20.2 �m(31–35 × 18–25)
Cercarial chaetotaxy 4 CI, 10 CII, 22 CIII
18 H1–2 AIL, 1–2 AIIL, 1 AIIIL3 ML0–1 PIL, 1 PIIL, 4 PIIIL6 SI, 6 SII, 3–4 SIII
5 CI, 11 CII, 19 CIII
15 H2 AIL, 1 AIIL, 0 AIIIL3 ML1 PIL, 1PIIL, 1–2 PIIIV, 2 PIIIL,1PIIID6 SI, 12–14 SII, 0 SIII
4 CI, 9–10 CII, 13–15 CIII
16 H1 AIL, 1 AIIL, 1 AIIIL3 ML1 PIL, 1 PIIL, 4 PIIIL6 SI, 8–10 SII, 4–5 SIII
5–6 CI, 8–9 CII, 12–14 CIII
16 H1 AIL, 1 AIIL, 1 AIIIL3 ML1 PIL, 0 PIIL, 2–3 PIIIL6 SI, 5–6 SII, 7–8 SIII
4 CI, 10 CII, 21 CIII
16 H2 AIL + 1 AIV, 1 AII L, 0 AIIIL3 ML1 PIL, 0 PIIL, 3 PIIIL6 SI, 6 SII, 6 SIII
First intermediate hosts(site of infection)
R. decollatad,e
(digestive gland)T. pisanad,e, C. virgatad,e,C. barbarad,e, C. acutad,e
(digestive gland)
P. splendidad,e
(digestive gland)H. aspersad,e
(digestive gland)Helix a. aspersad,e
(digestive gland)
Second intermediate hosts(site/s of infection)
R. decollatae, O. punctatae,E. vermiculatae, T. pisanae
(kidney, pedal glands)
C. virgatad,e, H. aspersad,e
C. barbarad, C. acutad,M. armillatad, S. gawlerid,S. australisd, T. pisanad
(kidney)
O. punctatad, T. pisanad,P. splendidad, H. aspersad,e
(kidney)
H. aspersad,e, O. punctatad,e,T. pisanae
(kidney)
H. a. aspersad,e, H. a.maximad,e
(kidney)
Definitive hosts(site of infection)
M. musculuse, R. rattuse, E.q. ophiusaee
(duodenum)
Humansd, Mus musculusd,e,Ovis sp.d, F. catusd, R.norvegicuse, C. mellorid, T.merulad, S. vulgarisd, D.novaehollandiaed, G.gallusd, C. liviad, T. rugosad
(small intestine)
R. rattusd, R. novergicusd,e,M. musculuse, M. spretuse,A. sylvaticuse, M.unguiculatuse
(duodenum)
C. russulad, M. spretusd
M. musculuse
(middle portion of smallintestine)
M. musculusd,e
(duodenum, especially firstportion)
Locality Formentera, BalearicIslands, Spain
Australia Llobregat delta, Catalonia,Spain
Llobregat delta, Catalonia,Spain
Heliciculture farms,Galicia, Spain
References Mas-Coma and Montoliu(1986)
Butcher and Grove (2001)Butcher (2003)
Gracenea andGonzález-Moreno (2002)
González-Moreno andGracenea (2006)
This study
a Values are the mean of the mean values given for each definitive host considered in morphometric studies. Ranges in parentheses correspond to the minimum and maximum values considering all definitivehosts.
b Calculated from the values in first row because it was not provided by the authors.c Calculated from the published drawings, since this ratio was not given by the authors.d Naturally infected hosts.e Experimentally infected hosts.f Only data corresponding to adults from experimentally infected mice (n = 36) are included, since only 2 adults from naturally infected mice were measured.

Parasit
284 P. Segade et al. / Veterinaryasitizing several snails species as sporocyst (Butcher andGrove, 2001; Butcher, 2003). The fact that sporocysts of B.aspersae are able to develop in the subspecies H. a. aspersa,but not in H. a. maxima, confirms that this species is strictlyoioxenic agreeing with the latter statement.
Although new studies should be conducted in order toidentify the full range of hosts of this brachylaimid, ourresults indicate that, in the absence of appropriate controlmeasures, mice are enough to maintain the life cycle of B.aspersae in the farms dedicated to the production of petit-gris snails, since this subspecies can act as first and secondintermediate host. In this regards, mice were frequentlyseen at the three farms included in the present study and inone of them, the rodents were even observed in the bags ofcommercial feed. Mice may therefore spread the digeneaninfection not only by depositing their contaminated faecesin outdoor pens or greenhouses where fattening of snailsis carried out, but also by contaminating the snail feed. Onthe other hand, in two of the farms studied, intensive mat-ing is accomplished in climate-controlled buildings. Duringthis phase, the density of snails is maintained high to facil-itate mating and the humidity is controlled by periodicalspraying of water. Since these conditions facilitate the sur-vival and effective transmission of cercariae, especial careshould be taken to avoid the entry of rodents and to min-imize the introduction of sporocyst-infected breeders inthese buildings.
Since H. a. maxima cannot act as first intermediate hostfor B. aspersae, the infection of farmed gros-gris snails bymetacercariae of this species can only be explained by thesimultaneous rearing of the two H. aspersa subspecies inthe farms studied. In fact, as was expected, the prevalenceand intensities of metacercariae were especially high in thefarm at O Rosal, where both subspecies were maintainedtogether in the same pens.
Flukes are considered a potential source of morbid-ity and mortality for captive snails (Cooper and Knowler,1991; Cuéllar and Cuéllar, 2003; Fontanillas and García-Cuenca, 2005; Arrébola and Munoz-Araujo, 2007). In thecase of B. aspersae, the presence of large sporocysts andnumerous metacercariae in heavily infected snails couldhave important consequences for their survival, especiallyunder intensive breeding conditions, where overcrowding,and probably reproductive stress, can have additional dele-terious effects for gastropods (Daguzan and Verly, 1989;Blanc and Attia, 1992; Jess y Marks, 1995; Dupont-Nivetet al., 2000; García et al., 2006). Although the associationbetween the presence of brachylaimid metacercariae andhost mortality is not clear, there is no doubt that these larvalstages, which feed on the renal epithelium, cause necroticchanges in this organ (this study; Foster, 1958; Lewis, 1969;Rashed et al., 1995). Numerous authors have also reportedthe extensive pathologic changes associated with the pres-ence of sporocysts in several aquatic snails (Reader, 1971;Huffman and Fried, 1985; Crews and Esch, 1987). To ourknowledge, however, no studies have described to date
the pathological consequences derived from the presenceof highly branched brachylaimid sporocysts. In this sense,we have observed that a great part of the hepatopancreascan be replaced by the parasite. Since this organ is respon-sible for the production of digestive enzymes, nutrientology 175 (2011) 273–286
absorption and digestion, endocytosis of food substances,food storage and excretion (Owen, 1996), massive infec-tions with sporocysts may have an important impact onsnail morbimortality by altering its normal functions. Insupport of this idea, we observed that most of the heav-ily sporocyst-infected snails died during our experimentalinfection studies (data not shown).
Regardless of its pathological effects, the presence ofbrachylaimids in edible snails is important from the san-itary perspective, since the zoonotic potential of thesedigeneans has been demonstrated in Australia, where threecases of human infection, probably linked to the inges-tion of raw infected snails, have been reported (Butcheret al., 1996, 1998). Although the zoonotic transmission ofbrachylaimids in Europe is highly improbable since edi-ble snails are always cooked, this aspect should be takenin mind if some culinary practices involving undercook-ing snails are introduced. Apart from this, ingestion ofsnails can induce asthma and other allergic manifesta-tions in humans, especially in house-dust-mite-sensitizedpatients (Server, 1999; Martins et al., 2005). Initially, itwas thought that the mite R. limacum, which lives in themantle cavity of certain edible snails including H. aspersa(Morand and Daguzan, 1986; Graham et al., 1996; Iglesiaset al., 2004), could be the source of allergens responsiblefor these reactions. However, current evidences suggestthat hypersensitivity is related to certain snail components,which remain currently unknown (Vuitton et al., 1998).Since some helminths are well-known sources of allergens(Sereda et al., 2008), the possibility that the anaphylacticsymptoms following snail consumption are actually relatedto allergenic substances of brachylaimids or other snail-infecting parasites cannot be ruled out. In view of theseconsiderations and taking into account the most recent reg-ulations on food hygiene and safety, snail farmers shouldincrease their efforts to avoid the infection of edible snailswith potentially zoonotic and pathogenic parasites like B.aspersae.
Acknowledgements
This work was supported by grantPGIDIT04RAG310015PR (Xunta de Galicia). We wishto express our gratitude to J. Méndez and I. Pazos fromC.A.C.T.I. (Centro de Apoio Científico e Tecnolóxico á Inves-tigación; Universidade de Vigo) for technical assistance inSEM studies.
References
Arrébola, J.R., Munoz-Araujo, B., 2007. Patologías y mortalidad en loscaracoles terrestres. In: Asociación Nacional de cría y engorde delcaracol. Helicicultura sotenible. Ministerio de Agricultura, Pesca y Ali-mentación, Madrid, 115 pp.
Balozet, L., 1937a. Brachylaemus suis Mihi 1936, Trématode de lıintestindu porc. Rôle pathogène et cycle évolutif. Arch. Inst. Pasteur Tunis 26,36–67.
Balozet, L., 1937b. Cycle évolutif de Brachylaemus suis. C. R. Acad. Sci. Paris
204, 622–624.Bayssade-Dufour, C., 1979. L’appareil sensorial des cercaires et la sys-tématique des Trématodes Digénétiques. Mém. Mus. Hist. Nat. 113,1–81.
Blanc, A., Attia, J., 1992. Effect of population density on growth of landsnail Helix aspersa maxima. J. Appl. Anim. Res. 2, 73–80.

Parasit
B
B
B
B
B
B
C
C
C
C
C
C
C
C
C
D
D
D
D
F
F
F
G
G
G
G
P. Segade et al. / Veterinary
ush, A.O., Lafferty, K.D., Lotz, J.M., Shostak, A.W., 1997. Parasitology meetsecology on its own terms: Margolis et al. revisited. J. Parasitol. 83,575–583.
utcher, A.R., Graham, A.T., Norton, R.E., Kirk, M.D., Cribb, T.H., Forsyth,J.R.L., Knight, B., Cameron, S., 1996. Locally acquired Brachylaima sp.(Digenea: Brachylaimidae) intestinal fluke infection in two South Aus-tralian infants. Med. J. Aust. 164, 475–478.
utcher, A.R., Parasuramar, P., Thompson, ChS., Grove, D., 1998. First reportof the isolation of an adult worm of the genus Brachylaima (Dige-nea: Brachylaimidae), from the gastrointestinal tract of a human. Int.J. Parasitol. 28, 607–610.
utcher, A.R., Grove, D.I., 2001. Description of the life-cycle stages ofBrachylaima cribbi n. sp. (Digenea: Brachylaimidae) derived from eggsrecovered from human faeces in Australia. Syst. Parasitol. 49, 211–221.
utcher, A.R., Palethorpe, H., Grove, D.I., 2002. The susceptibility on inbredmice to infection with Brachylaima cribbi (Digenea: Brachylaimidae).Parasitol. Int. 51, 259–263.
utcher, A.R., 2003. Brachylaima cribbi n. sp. (Digenea: Brachylaimidae):taxonomy, life-cycle kinetics and infections in animals and humans.Doctoral Thesis, University of Adelaide, South Australia, 205 pp.
abaret, J., Morand, S., Aubert, C., Yvore, P., 1988. Snail farming: a surveyof breeding management, hygiene and parasitism of the garden snail,Helix aspersa Müller. J. Molluscan Stud. 54, 209–214.
ameron, R.A.D., Carter, M.A., 1979. Intra- and interspecific effects of pop-ulation density on growth and activity in some helicid land snails(Gastropoda: Pulmonata). J. Anim. Ecol. 48, 237–246.
hevalier, H., 1979. Les escargots. Un élevage d’Avenir. Dargaud, Neuilly-Sur-Seine, 96 pp.
ombes, C., Bayssade-Dufour, C., Cassone, J., 1976. Sur l’impregnation et lemontage des cercaires pour l’étude chétotaxique. Ann. Parasitol. Hum.Comp. 51, 399–400.
ooper, J.E., Knowler, C., 1991. Snails and snail farming: an introductionfor the veterinary profession. Vet. Rec. 129, 541–549.
rews, A.Y., Esch, G., 1987. Histopathology of larval trematode infectionsin the freshwater snail, Helisoma anceps. J. Invertebr. Pathol. 49, 76–82.
ribb, T.H., 1990. Introduction of a Brachylaima species (Digenea: Brachy-laimidae) to Australia. Int. J. Parasitol. 20, 789–796.
ribb, T.H., 1992. The Brachylaimidae (Trematoda: Digenea) of Australiannative mammals and birds, including descriptions of Dasyurotrema n.g. and four new species of Brachylaima. Syst. Parasitol. 22, 45–72.
uéllar, R., Cuéllar, M.C., 2003. Producción de caracoles. Bases fisiológicas.sistemas de producción y patología, 2a edición. Ediciones Mundi-Prensa, Madrid, 173 pp.
aguzan, J., Verly, D., 1989. Etude experimentale del’effet de la densite surla reproduction de l’escargot petit-gris (Helix aspersa Müller). Haliotis19, 105–115.
an, N., Bailey, S.E.R., 1982. Growth, mortality and feeding rates of thesnail Helix aspersa at different population densities in the laboratoryand the depression of activity of helicid snails by other individuals ortheir mucus. J. Molluscan Stud. 48, 257–265.
ollfus, R.Ph., 1954. Un Brachylaema (Trématodes, distomes) de pigeondomestique. Arch. Inst. Pasteur Maroc. 4, 603–611.
upont-Nivet, M., Coste, V., Coinon, P., Bonnet, J., Blanc, J., 2000. Rearingdensity effect on the production performance of the edible snail Helixaspersa Müller in indoor rearing. Ann. Zootech. 49, 447–456.
eliu, C., Mas-Coma, S., Montoliu, I., Gállego, J., 1986. Brachylaima simoni sp.n. (Trematoda: Brachylaimidae) a parasite of the shrew Crocidura rus-sula (Hermann, 1780) (Insectivora: Soricidae) from the Pityusic islandof Ibiza (Balearics). Helminthologia 23, 65–72.
ontanillas, J.C., García-Cuenca, I., 2005. El caracol y la helicicultura, 2a
edición. Ediciones Mundi-Prensa, Madrid, 136 pp.oster, R., 1958. The effects of trematode metacercariae (Brachylaemi-
dae) on the slugs Milax sowerbii Férussac and Agriolimax reticulatusMüller. Parasitology 48, 261–268.
arcía, A., Perea, J.M., Mayoral, A., Acero, R., Martos, J., Gómez, G., Pena, F.,2006. Laboratory rearing conditions for improved growth of juvenileHelix aspersa Müller snails. Lab. Anim. 40, 309–316.
omot-de Vaufleury, A., Borgo, R., 2001. Experimental hybridizationbetween two sub-species of snails (Helix aspersa aspersa and Helixaspersa maxima): consequences for fertility, survival and growth.Invertebr. Repr. Dev. 40, 217–226.
onzález-Moreno, O., Gracenea, M., Montoliu, I., Villa, M., 1994. Presencia
de digénidos Brachylaimidae en el Delta del Llobregat (Barcelona):Estadios larvarios parásitos de gasterópodos terrestres. Iberus 12,49–57.onzález-Moreno, O., Gracenea, M., 2006. Life cycle and description of anew species of brachylaimid (Trematoda: Digenea) in Spain. J. Para-sitol. 92, 1305–1312.
ology 175 (2011) 273–286 285
Gracenea, M., González-Moreno, O., 2002. Life cycle of Brachylaima mas-comai n. sp. (Trematoda: Brachylaimidae), a parasite of rats in theLlobregat Delta (Spain). J. Parasitol. 88, 124–133.
Graham, F.J., Runham, N.W., Ford, J.B., 1996. Long-term effects of Riccar-doella limacum living in the lung of Helix aspersa. In: Henderson, I.F.(Ed.), Slug and snail pests in Agriculture. BCPC Symposium Proceed-ings, 66, pp. 359–364.
Hezberg, F., 1965. Crowding as a factor in growth and reproduction of Helixaspersa Müller. Am. Zool. 5, 254–262.
Huffman, J.E., Fried, B., 1985. Histopathological and histochemical effectsof larval trematodes in Goniobasis viginica (Gastropoda; Pleuroceri-dae). Veliger 27, 273–281.
Iglesias, R., Rivera, P., Souto, O., Segade, P., Rodríguez-Domínguez, H.,Crespo-González, C., García-Estévez, J.M., 2004. Parasites infecting thebrown garden snail Helix aspersa aspersa in heliciculture farms of Gali-cia. IX European Multicolloquium of Parasitology. Valencia, p. 610.
Jess, S., Marks, R.J., 1995. Population density effects on growth in cultureof the edible snail Helix aspersa var. maxima. J. Molluscan Stud. 61,313–323.
Jourdane, J., 1971. Helminth parasites des micromammifères desPyrénées-Orientales. II. Les plathelmintes de Soricinae. Ann. Parasitol.Hum. Comp. 46, 553–574.
Joyeux, C., Foley, H., 1930. Les helminthes de Meriones shawi Rozet dansle nord de l’Algerie. Bull. Soc. Zool. Fr. 55, 355–374.
Joyeux, Ch., Baer, J.G., Timon-David, J., 1934. Recherches sur les trématodesdu genre Brachylemus Dujardin (synHarmostomum Braun). Bull. BiolFr. Belg. 68, 385–418.
Kock, S., Böckeler, W., 1998. Observations on cercarial chaetotaxy as ameans for the identification of European species of TrichobilharziaSkrjabin & Zakharow, 1920 (Digenea: Schistosomatidae). Syst. Para-sitol. 43, 159–166.
Lewis, J.W., 1969. Studies on the life history of Brachylaimus oesophageiShaldybin, 1953 (Digenea: Brachylaimidae). J. Helminthol. 43, 79–98.
Macchioni, G., 1968. Segnalazione di nuovi molluschi ospiti intermedidi Brachylaima fuscata (Rudolphi, 1819). Atti. Soc. Ital. Sci. Vet. 22,746–749.
Macchioni, G., 1970. Variazioni intraspecifiche di Brachylaema fuscata(Rudolphi, 1819). Caratteri tassonomicamente validi per la diagnosidi specie. Acta Med. Vet. (Napoli). 16, 45–71.
Martins, L.M.L., Peltre, G., Da Costa Faro, C.J.F., Pires, E.M., Da Cruz Inácio,F.F., 2005. The Helix aspersa (brown garden snail) allergen repertoire.Int. Arch. Allergy Immunol. 136, 7–15.
Mas-Coma, S., Montoliu, I., Valero, M.A., 1984. Methodologie d’étude mor-phométrique de la variabilité intraspécifique chez les digènes de lafamille Brachylaimidae Joyeux et Foley, 1930. Bull. Soc. Neuchât. Sci.Nat. 107, 185–195.
Mas-Coma, S., Montoliu, I., 1986. The life cycle of Brachylaima ruminae n.sp. (Trematoda: Brachylaimidae), a parasite of rodents. Z. Parasitenkd72, 739–753.
Mayoral, A.G., García, A., Perea, J., Martín, R., Martos, J., Acero, R., Pena, F.,2004. Efecto de la densidad de población sobre el tamano del caracolHelix aspersa Müller. Arch. Zootech. 53, 379–382.
Morand, S., Daguzan, J., 1986. Contribution al’etude du parasitisme del’escargot petit-gris (Helix aspersa Müller): premiers resultats con-cernant l’acarien Riccardoella limacum (Schrank) et le nematodeAlloionema appendiculatum (Schneider). Haliotis 15, 31–39.
Owen, G., 1996. Digestion. In: Wilbur, K.M., Yonge, C.M. (Eds.), Physiologyof Mollusca, vol. 2. Academic Press, New York, pp. 53–96.
Pojmanska, T., 1961. Investigations on the occurrence and biology oftrematodes of Sorex araneus araneus L. in Bialowieza National Park.Acta Parasitol. Pol. 9, 305–330.
Pojmanska, T., 2002a. Family Brachylaimaidae Joyeux and Foley, 1930. In:Gibson, D.I., Jones, A., Bray, R.A. (Eds.), Keys to the Trematoda. CABInternational and Natural History Museum, London, pp. 37–45.
Pojmanska, T., 2002b. Superfamily Brachylaimoidea Joyeux and Foley,1930. In: Gibson, D.I., Jones, A., Bray, R.A. (Eds.), Keys to the Trematoda.CAB International and Natural History Museum, London, pp. 31–36.
Rashed, A.A., Wanas, M.Q.A., Al Shareef, A.M.F., Al Attar, N.M.O., Sabry, A.A.,Morsy, T.A., 1995. The histopathological effect of metacercariae of thegenus Brachylaima on the land snail, Monacha obstructa. J. Egypt. Soc.Parasitol. 25, 535–542.
Reader, T.A.J., 1971. The pathological effects of sporocysts, rediae andmetacercariae on the digestive gland of Bithynia tentaculata (Mollusca:
Gastropoda). Parasitology 63, 483–489.Richard, J., 1971. La chétotaxie des cercaires: valeur systématique et phylé-tique. Mem. Mus. Natl. Hist. Nat. Paris 67, 1–179.
Segade, P., Kher, C.P., Lynn, D.H., Iglesias, R., 2009. Morphological andmolecular characterization of renal ciliates infecting farmed snails inSpain. Parasitology 136, 771–782.

Parasit
286 P. Segade et al. / VeterinarySereda, M.J., Hartmann, S., Lucius, R., 2008. Helminths and allergy: theexample of tropomyosin. Trends Parasitol. 24, 272–278.
Server, M.T., 1999. Reactividad cruzada entre ácaros y caracol. Alergol.Inmunol. Clin. 14, 79–82.
Simón-Vicente, F., 1955a. Brachylaemus en infestación experimental y nat-ural. Rev. Iber. Parasitol. 15, 301–304.
Simón-Vicente, F., 1955b. Nota previa sobre el ciclo experimental de un
Brachylaemus. Rev. Iber. Parasitol. 15, 607–610.Timon-David, J., 1954. Kystes á Brachylaemus chez Cyclostoma elegansMüller Développment expérimental du parasite. C. R. Soc. Biol. 148,707–710.
Timon-David, J., 1959. Recherches sur les kystes a Brachylaemus ducyclostome. Ann. Parasitol. Hum. Comp. 34, 271–287.
ology 175 (2011) 273–286
Timon-David, J., 1965. Infestation expérimentale dıune Héllicelle par huitespèces de Trématodes digénétiques appartenant à quatre famillesdifférentes. Ann. Parasitol. Hum. Comp. 40, 149–154.
Ubelaker, J.E., Dailey, M.D., 1966. Taxonomy of the genus BrachylaimaDujardin (Trematoda: Digenea) with description of B. chiapensis sp.n. from Peromyscus guatemalensis in Mexico. J. Parasitol. 52, 1062–1065.
Vuitton, D.A., Rancé, F., Paquin, M.L., Adessi, B., Vigan, M., Gomot, A., Dutau,G., 1998. Cross-reactivity between terrestrial snails (Helix species) andhouse-dust mite (Dermatophagoides pteronyssinus). I. In vivo study.Allergy 53, 144–150.
Yamaguti, S., 1971. Synopsis of digenetic trematodes of vertebrates, vol. I.Keigaku Pub. Co., Tokyo, Japan, p. 1074.




















