
Flow cytometric assay for in vivo genotoxic effects of pesticides in Green frogs (Rana clamitans)

915
J. Parasitol., 91(4), 2005, pp. 915–921q American Society of Parasitologists 2005
INTRASPECIFIC VARIATION OF HAEMATOLOECHUS FLOEDAE HARWOOD, 1932(DIGENEA: PLAGIORCHIIDAE), FROM RANA SPP. IN NORTH AND CENTRAL AMERICA
Virginia Leon-Regagnon, Sergio Guillen-Hernandez*, and Marıa Antonieta Arizmendi-EspinosaDepartamento de Zoologıa, Instituto de Biologıa, UNAM. Apdo. Postal 70-153. C.P. 04510, Mexico, D.F. Mexico. e-mail:[email protected]
ABSTRACT: Haematoloechus floedae was originally described from the lungs of Rana catesbeiana in Texas, and later reportedin Florida, in the United States. It was proposed to be synonymous with H. breviplexus, H. parviplexus, and H. varioplexus. Weobtained specimens of H. floedae from Rana brownorum and R. vaillanti in Yucatan, Mexico; R. cf. forreri and R. taylori inGuanacaste, Costa Rica; and R. catesbeiana in Georgia and California. Some specimens were processed for morphological study;sequences of the 28S of the rDNA and the mitochondrial COI were obtained from several specimens of each population.Phylogenetic analysis of molecular data indicates studied populations constitute a single taxon, different from H. varioplexus, H.breviplexus, and H. parviplexus. Some morphological characters remain constant among populations of H. floedae, and are usefulfor differentiating this species. These include the oral sucker-pharynx ratio, oral sucker-ventral sucker ratio, ovary and testesshape, extension of uterine longitudinal loops, and extension of vitelline follicles (in fully developed worms). Sequence homo-geneity among populations of H. floedae suggests a recent spread, perhaps due to the introduction of R. catesbeiana for cultureto the western United States, southeast Mexico, and Costa Rica.
Morphological variability within species of Haematoloechusspp., the frog lung flukes, has been documented (Prokopic andKrivanec, 1974; Kennedy, 1980a, 1980b), and has caused con-troversy regarding taxonomy of the group (Odening, 1960; Pro-kopic and Krivanec, 1974; Kennedy, 1981). Kennedy (1981)considered that only 6 of the 15 previously known species fromCanada and the United States were valid. This author concludedthat morphological characters, such as the shape of the ovaryand testes, arrangement of uterine loops, presence or absenceof spines on the body surface, sucker ratio, and egg size, ex-hibited intraspecific variation as the result of several factors(i.e., developmental stage, host species, crowding effect), and,therefore, are not useful for differentiating species of Haema-toloechus.
Species delimitation can be a controversial issue, especiallyin those cases in which morphological variability can confoundthe taxonomy of a group (Leon-Regagnon et al., 1999; Stocket al., 2001), or in which morphological characters are so con-servative that independent lineages cannot be differentiated(Chilton et al., 1995; Verneau et al., 1997). In these cases, DNAsequence data provide an independent source of information fordifferentiating diverging lineages, and to evaluate morphologi-cal characters that have been traditionally used in the taxonomyof the groups (Nadler, 2002).
In the present study, morphological variation of specimensof H. floedae from different localities and host species of Northand Central America are documented; molecular data are usedto corroborate whether differences among populations representintraspecific or interspecific variation, and validate the use ofsome morphological characters in the differentiation of speciesof this genus.
MATERIALS AND METHODSCollecting localities and hosts are shown on Table I. Worms collected
from freshly killed amphibians initially were placed in saline (0.65%)for 5–10 min. For the morphological study, they were fixed by suddenimmersion in hot 4% formalin and preserved in 70% alcohol. Speci-
Received 13 July 2004; revised 13 October 2004; accepted 22 October2004.
* Universidad Autonoma de Yucatan. Facultad de Medicina Veterinaria,Km 105 carretera Merida-Xmatkuil, Ap. Postal 14-6. C.P. 97000, It-zimna, Merida, Yucatan, Mexico.
mens were stained with Mayer paracarmine or Gomori trichrome, de-hydrated, cleared in methyl salicylate, and mounted in Canada balsam.Specimens were mounted permanently between coverslips and held inCobb slides. Measurements are presented as the range with means inparentheses and expressed in millimeters, unless otherwise stated. Fig-ures were drawn with the aid of a drawing tube. Voucher specimenaccession numbers are listed in Table I. The following abbreviations areused: CNHE, Coleccion Nacional de Helmintos, Instituto de Biologıa,Universidad Nacional Autonoma de Mexico; USNPC, United StatesNational Parasite Collection, Beltsville, Maryland; and HWML, HaroldW. Manter Laboratory, Lincoln, Nebraska.
Based on the phylogenetic hypothesis proposed by Leon-Regagnonand Brooks (2003) for the genus, we chose the most closely relatedspecies of Haematoloechus for reference. The following specimenswere examined for comparison: H. breviplexus Stafford, 1902 (USNPC82015 and 84806; HWML 20293, 21957, and 22691); H. danbrooksiLeon-Regagnon and Paredes-Calderon, 2002 (CNHE 4112 and 4151;USNPC 92220); H. floedae (USNPC 30879, 84804, and 091507); H.parviplexus Irwin, 1929 (USNPC 75445 and 81467; HWML 20142–43, 20753, and 21660; CNHE 4405); and H. varioplexus Stafford, 1902(USNPC 75447 and 81915; HWML 20151–20160 and 38396).
Two or 3 specimens from each locality and host species were usedfor molecular work. They were identified in vivo and preserved in 100%ethanol. Standard phenol extraction methods were used to recover DNAfrom individual specimens. Laboratory protocols followed those de-scribed by Palumbi (1996) and Hillis et al. (1996). Polymerase chainreaction (PCR) was used for amplifying the 59 end of the 28S ribosomalRNA gene and partial COI mitochondrial gene. Amplification and se-quencing were performed using the primers 28Sy 59-cta acc agg att ccctca gta acg gcg agt-39 (forward) and 28Sz 59-aga ctc ctt ggt ccg tgt ttcaag ac-39 (reverse) for the 28S; and COIF 59-ttt ttt ggg gat cct gag gtttat-39 (forward) and COIR 59-taa aga aag aac ata atg aaa atg-39 (re-verse). Amplification programs consisted of 1 min at 94 C followed by35 cycles of 30 sec at 92 C, 30 sec at 50 C, and 1 min at 72 C; followedby 4 min at 72 C for final elongation for the 28S gene; and 5 min at95 C followed by 35 cycles of 30 sec at 95 C, 45 sec at 50 C, and 1.5min at 72 C, followed by 10 min at 72 C for final elongation for theCOI gene. PCR products were sequenced directly on an ABI Prism 310automated DNA sequencer using Thermo Sequenase radiolabeled ter-minator cycle sequencing kits (Amersham Life Science, Inc., Cleveland,Ohio). GenBank accession numbers are listed in Table I.
We obtained more than 1 COI sequence from each population of H.floedae to evaluate genetic divergence among and within populations;only 1 sequence of each population was included in the combined par-simony analysis. Sequences were aligned visually using the computerprogram Bioedit (Hall, 1999). Uncorrected distance matrices were ob-tained for the pairs of examined sequences, and phylogenetic trees wereconstructed using PAUP (version 4.0b10 Swofford, 2002). Unweightedparsimony analyses using a branch-and-bound search were performedconsidering character states as unordered, and gaps as missing data forseparate and combined matrices. In each case, a nonparametric boot-
916 THE JOURNAL OF PARASITOLOGY, VOL. 91, NO. 4, AUGUST 2005
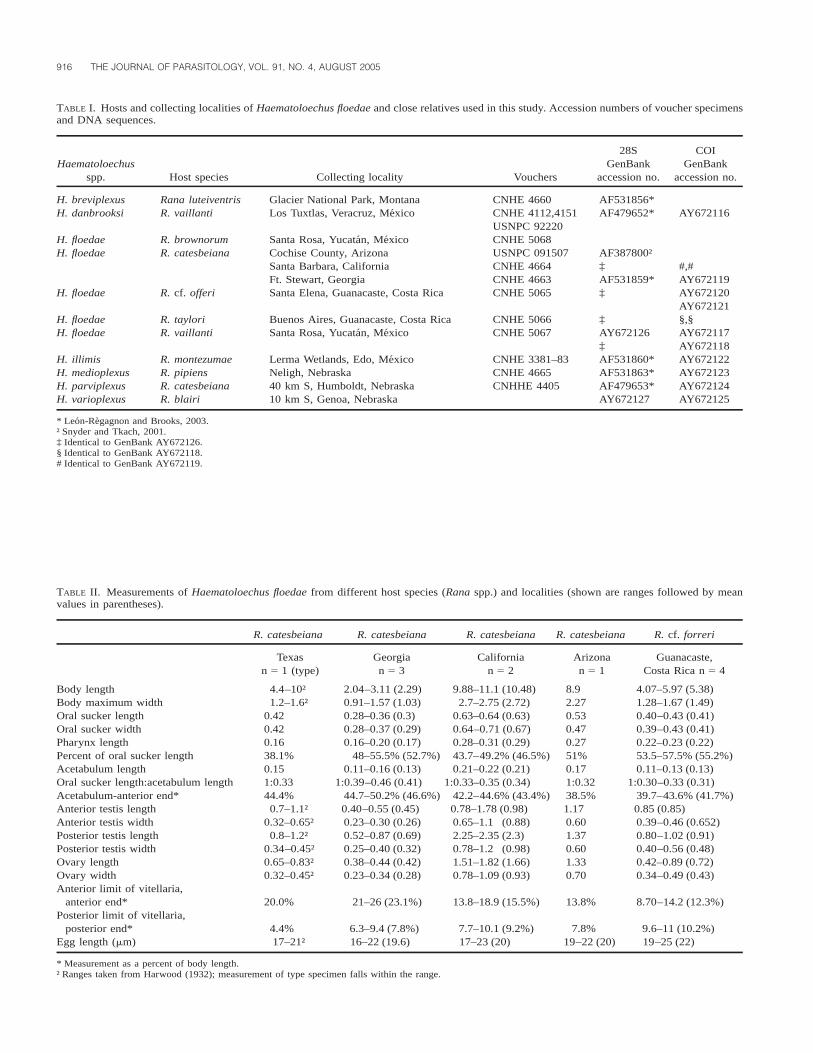
TABLE I. Hosts and collecting localities of Haematoloechus floedae and close relatives used in this study. Accession numbers of voucher specimensand DNA sequences.
Haematoloechusspp. Host species Collecting locality Vouchers
28SGenBank
accession no.
COIGenBank
accession no.
H. breviplexusH. danbrooksi
Rana luteiventrisR. vaillanti
Glacier National Park, MontanaLos Tuxtlas, Veracruz, Mexico
CNHE 4660CNHE 4112,4151USNPC 92220
AF531856*AF479652* AY672116
H. floedaeH. floedae
R. brownorumR. catesbeiana
Santa Rosa, Yucatan, MexicoCochise County, ArizonaSanta Barbara, CaliforniaFt. Stewart, Georgia
CNHE 5068USNPC 091507CNHE 4664CNHE 4663
AF387800†‡AF531859*
#,#AY672119
H. floedae
H. floedae
R. cf. offeri
R. taylori
Santa Elena, Guanacaste, Costa Rica
Buenos Aires, Guanacaste, Costa Rica
CNHE 5065
CNHE 5066
‡
‡
AY672120AY672121§,§
H. floedae
H. illimisH. medioplexusH. parviplexusH. varioplexus
R. vaillanti
R. montezumaeR. pipiensR. catesbeianaR. blairi
Santa Rosa, Yucatan, Mexico
Lerma Wetlands, Edo, MexicoNeligh, Nebraska40 km S, Humboldt, Nebraska10 km S, Genoa, Nebraska
CNHE 5067
CNHE 3381–83CNHE 4665CNHHE 4405
AY672126‡AF531860*AF531863*AF479653*AY672127
AY672117AY672118AY672122AY672123AY672124AY672125
* Leon-Regagnon and Brooks, 2003.† Snyder and Tkach, 2001.‡ Identical to GenBank AY672126.§ Identical to GenBank AY672118.# Identical to GenBank AY672119.
TABLE II. Measurements of Haematoloechus floedae from different host species (Rana spp.) and localities (shown are ranges followed by meanvalues in parentheses).
R. catesbeiana R. catesbeiana R. catesbeiana R. catesbeiana R. cf. forreri
Texasn 5 1 (type)
Georgian 5 3
Californian 5 2
Arizonan 5 1
Guanacaste,Costa Rica n 5 4
Body lengthBody maximum widthOral sucker length
4.4–10†1.2–1.6†
0.42
2.04–3.11 (2.29)0.91–1.57 (1.03)0.28–0.36 (0.3)
9.88–11.1 (10.48)2.7–2.75 (2.72)
0.63–0.64 (0.63)
8.92.270.53
4.07–5.97 (5.38)1.28–1.67 (1.49)0.40–0.43 (0.41)
Oral sucker widthPharynx lengthPercent of oral sucker lengthAcetabulum lengthOral sucker length:acetabulum lengthAcetabulum-anterior end*
0.420.1638.1%0.151:0.3344.4%
0.28–0.37 (0.29)0.16–0.20 (0.17)
48–55.5% (52.7%)0.11–0.16 (0.13)
1:0.39–0.46 (0.41)44.7–50.2% (46.6%)
0.64–0.71 (0.67)0.28–0.31 (0.29)43.7–49.2% (46.5%)0.21–0.22 (0.21)
1:0.33–0.35 (0.34)42.2–44.6% (43.4%)
0.470.2751%0.171:0.3238.5%
0.39–0.43 (0.41)0.22–0.23 (0.22)53.5–57.5% (55.2%)0.11–0.13 (0.13)
1:0.30–0.33 (0.31)39.7–43.6% (41.7%)
Anterior testis length 0.7–1.1† 0.40–0.55 (0.45) 0.78–1.78 (0.98) 1.17 0.85 (0.85)Anterior testis widthPosterior testis lengthPosterior testis widthOvary lengthOvary width
0.32–0.65†0.8–1.2†
0.34–0.45†0.65–0.83†0.32–0.45†
0.23–0.30 (0.26)0.52–0.87 (0.69)0.25–0.40 (0.32)0.38–0.44 (0.42)0.23–0.34 (0.28)
0.65–1.1 (0.88)2.25–2.35 (2.3)0.78–1.2 (0.98)1.51–1.82 (1.66)0.78–1.09 (0.93)
0.601.370.601.330.70
0.39–0.46 (0.652)0.80–1.02 (0.91)0.40–0.56 (0.48)0.42–0.89 (0.72)0.34–0.49 (0.43)
Anterior limit of vitellaria,anterior end* 20.0% 21–26 (23.1%) 13.8–18.9 (15.5%) 13.8% 8.70–14.2 (12.3%)
Posterior limit of vitellaria,posterior end* 4.4% 6.3–9.4 (7.8%) 7.7–10.1 (9.2%) 7.8% 9.6–11 (10.2%)
Egg length (mm) 17–21† 16–22 (19.6) 17–23 (20) 19–22 (20) 19–25 (22)
* Measurement as a percent of body length.† Ranges taken from Harwood (1932); measurement of type specimen falls within the range.

LEON-REGAGNON ET AL.—HAEMATOLOECHUS FLOEDAE IN NORTH AND CENTRAL AMERICA 917
TABLE II. Extended.
R. taylori R. brownorum R. vaillanti
Guanacaste,Costa Rica n 5 7
Yucatan, Mexicon 5 1
Yucatan, Mexicon 5 5
5.85–6.9 (6.58)1.7–2.55 (2.12)
0.44–0.57 (0.48)
4.051.250.36
3.25–9.72 (45.59)0.72–2.47 (1.35)0.28–0.54 (0.36)
0.42–0.52 (0.47)0.21–0.27 (0.24)43.7–60.6% (49.8%)0.13–0.18 (0.16)
1:0.28–0.36 (0.31)39.7–44.4% (42.5%)
0.360.1849.9%0.141:0.3844.8%
0.29–0.49 (0.37)0.14–0.23 (0.18)42.7–60.6% (49.2%)0.12–0.14 (0.13)
1:0.37–0.38 (0.38)45.9–46.3% (46.1%)
0.96–1.6 (1.09) 0.50 0.50–1.12 (0.67)0.52–0.9 (0.69)1.23–1.81 (1.56)0.51–0.82 (0.67)0.92–1.26 (1.09)0.45–0.76 (0.65)
0.350.760.330.730.55
0.20–0.49 (0.38)0.70–1.32 (0.96)0.41–0.56 (0.472)0.60–1.54 (0.91)0.30–0.88 (0.51)
12.1–21.5% (16.1%) 15% 12.1–20.3% (16.8%)
3.6–11.6% (7.5%) 7.9–9.9% (8.9%)24–27 (23.9) 14.8–17.3 (15.6) 14.8–22.2 (20.1)
strap with 1,000 replicates, as well as decay indices, were calculated toevaluate the stability of branches in the resulting topologies. We usedthe methodology proposed by Wiens (1998) to combine the data sets.
RESULTS
Morphological data
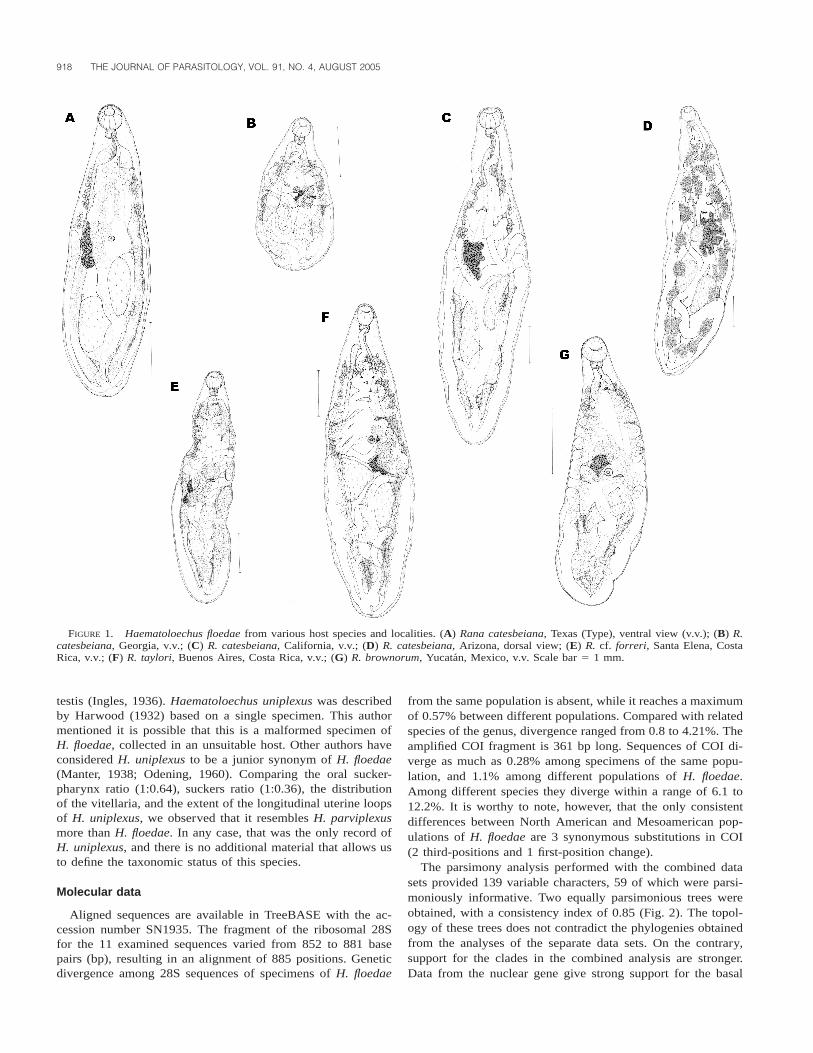
We observed a wide amount of variation in the general shapeand size of the body among the studied populations of H. floe-dae (Fig. 1; Table II). Specimens collected in Georgia werevery small, and the shape of the body was pyriform (Fig. 1B).Vitelline follicles in the posterior region were fewer in numberthan in worms from other localities, and vitellaria in the op-posite side of the ovary reached only the posterior end of thebody, and did not invade the ovarian side. Apparently, theywere not fully developed worms. Specimens from Costa Rica(Fig. 1E, F) and Yucatan (Fig. 1G) were smaller than those fromCalifornia and Arizona (Fig. 1C, D), but all of them fell withinthe range originally described for the species (Harwood, 1932)(Fig. 1A). We found that morphological traits that remainedconstant among studied populations were the oral sucker-phar-ynx ratio, oral sucker-ventral sucker ratio, ovary and testesshape, extension of the vitelline follicles (in fully developedworms), and extension of uterine longitudinal loops.
Haematoloechus floedae was described from Rana catesbei-ana and R. clamitans from Houston, Texas (Harwood, 1932).According to the original description, this species most closelyresembles H. parviplexus, but they differ in the size of the ac-etabulum compared to the oral sucker, which is smaller in H.parviplexus (1:0.25 vs. 1:0.31–0.41 in H. floedae); the tegumentis heavily spined in H. parviplexus, but smooth in H. floedae.Additionally, we noticed that the pharynx, compared to the oral
sucker, is larger in H. parviplexus (1:0.66) than in H. floedae(1:0.46–0.55), and uterine longitudinal loops in H. parviplexusreach only to the posterior testis, while they extend to the ovaryin H. floedae. It also resembles H. breviplexus Stafford, 1902,and was declared a junior synonym of this species (Odening,1960). These 2 species differ in the shape of testes, being el-liptical or slightly lobed in H. floedae and deeply lobed in H.breviplexus. The acetabulum, compared to the oral sucker, islarger in H. breviplexus (1:0.5 vs. 1:0.31–0.41 in H. floedae).The longitudinal uterine loops run more anteriorly in H. floedae,reaching the level of the anterior testis and sometimes even thelevel of the ovary, while in H. breviplexus they reach only tothe level of the posterior testis. In Cort’s (1915) redescriptionof H. breviplexus, it is said that longitudinal uterine loops reachthe level of the posterior testis, and in some specimens theyreach the level of the ovary. This author also stated that thesize of oral sucker, pharynx, and acetabulum and, therefore, theratio between these structures, is subject to wide individual var-iation and is of little value in specific diagnosis. These state-ments are contrary to what we observed in the present study.In his redescription of H. breviplexus, Cort (1915) includedspecimens collected in Oklahoma, in the southern UnitedStates; it is possible that some specimens of Cort’s redescriptionof H. breviplexus were in fact H. floedae, which had not yetbeen described. Haematoloechus floedae has been identified asH. breviplexus on other occasions (i.e., specimens with acces-sion numbers USNPC 84804 and 091507 were originally iden-tified as H. breviplexus, and later reassigned to H. floedae [V.Leon-Regagnon, unpubl. obs.; Leon-Regagnon and Brooks,2003]).
The distribution of the vitellaria is also different between H.parviplexus and H. breviplexus. In H. floedae, vitelline follicleson the ovarian side reach the level of the posterior testis, whileon the opposite side, they reach the posterior end and extendupward on the ovarian side of the body. In H. breviplexus andH. parviplexus, vitellaria also extend asymetrically on bothsides of the body but, on the opposite side of the ovary, theyreach the posterior end, not invading the ovarian side of thebody.
More recently, Kennedy (1981) proposed the synonymy ofH. floedae, H. parviplexus, H. buttensis Ingles, 1936; H. simi-liplexus Stafford, 1902; and H. uniplexus Harwood, 1932 withH. varioplexus, considering of no taxonomic value the differ-ences in the shape of the ovary and testes, spination of the body,length of the longitudinal uterine loops, and other morphometrictraits. Differences with H. parviplexus were already mentionedabove. Haematoloechus similiplexus has been considered as ajunior synonym of H. varioplexus by several authors (Caballeroand Sokoloff, 1934; Manter, 1938; Odening, 1960), and we con-cur. Haematoloechus floedae can be easily distinguished fromH. varioplexus by the size of the acetabulum, compared to thesize of the oral sucker, which is much larger in H. varioplexus(1:0.86 vs. 1:0.31–0.41 in H. floedae). The ovary is rounded orslightly lobed in H. varioplexus, while it is deeply lobed in H.floedae. Longitudinal uterine loops are shorter in H. varioplex-us, reaching the level of the posterior testis. Haematoloechusbuttensis differs from H. floedae in having a larger acetabulumcompared to the oral sucker (1:0.7), in the shape of the ovary(which is kidney-shaped and never lobed), and in the longitu-dinal uterine loops, which reach only the level of the posterior
918 THE JOURNAL OF PARASITOLOGY, VOL. 91, NO. 4, AUGUST 2005
FIGURE 1. Haematoloechus floedae from various host species and localities. (A) Rana catesbeiana, Texas (Type), ventral view (v.v.); (B) R.catesbeiana, Georgia, v.v.; (C) R. catesbeiana, California, v.v.; (D) R. catesbeiana, Arizona, dorsal view; (E) R. cf. forreri, Santa Elena, CostaRica, v.v.; (F) R. taylori, Buenos Aires, Costa Rica, v.v.; (G) R. brownorum, Yucatan, Mexico, v.v. Scale bar 5 1 mm.
testis (Ingles, 1936). Haematoloechus uniplexus was describedby Harwood (1932) based on a single specimen. This authormentioned it is possible that this is a malformed specimen ofH. floedae, collected in an unsuitable host. Other authors haveconsidered H. uniplexus to be a junior synonym of H. floedae(Manter, 1938; Odening, 1960). Comparing the oral sucker-pharynx ratio (1:0.64), suckers ratio (1:0.36), the distributionof the vitellaria, and the extent of the longitudinal uterine loopsof H. uniplexus, we observed that it resembles H. parviplexusmore than H. floedae. In any case, that was the only record ofH. uniplexus, and there is no additional material that allows usto define the taxonomic status of this species.
Molecular data
Aligned sequences are available in TreeBASE with the ac-cession number SN1935. The fragment of the ribosomal 28Sfor the 11 examined sequences varied from 852 to 881 basepairs (bp), resulting in an alignment of 885 positions. Geneticdivergence among 28S sequences of specimens of H. floedae
from the same population is absent, while it reaches a maximumof 0.57% between different populations. Compared with relatedspecies of the genus, divergence ranged from 0.8 to 4.21%. Theamplified COI fragment is 361 bp long. Sequences of COI di-verge as much as 0.28% among specimens of the same popu-lation, and 1.1% among different populations of H. floedae.Among different species they diverge within a range of 6.1 to12.2%. It is worthy to note, however, that the only consistentdifferences between North American and Mesoamerican pop-ulations of H. floedae are 3 synonymous substitutions in COI(2 third-positions and 1 first-position change).
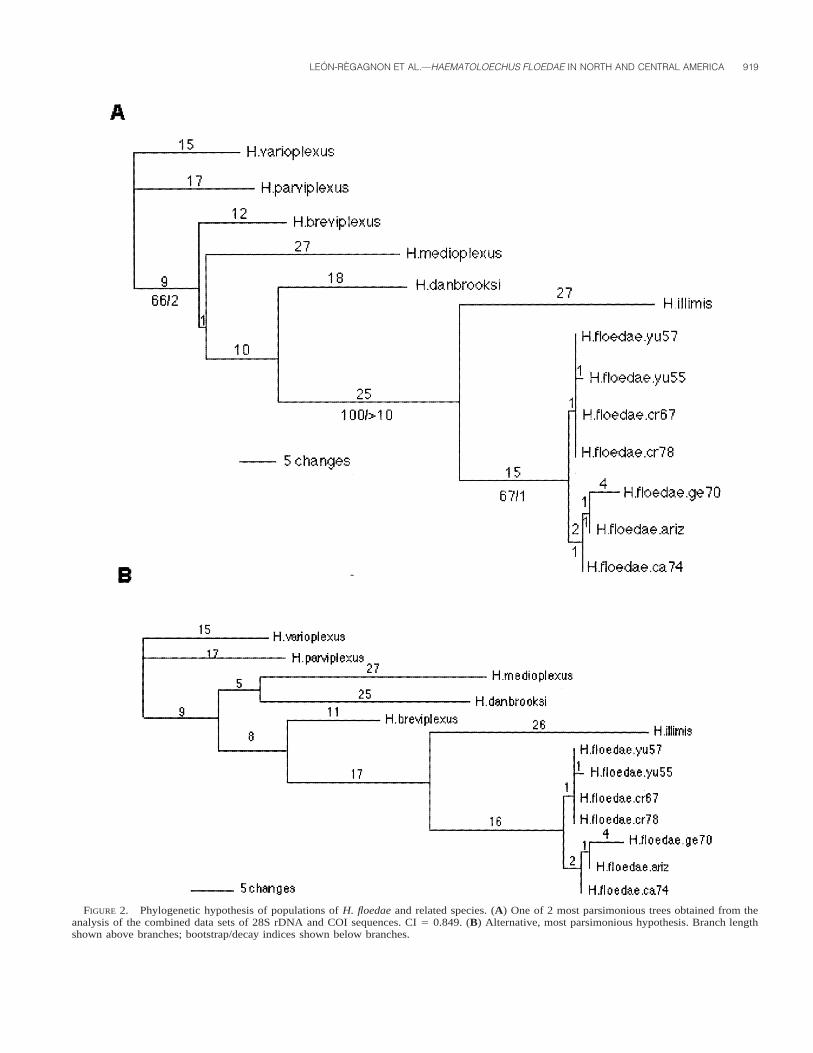
The parsimony analysis performed with the combined datasets provided 139 variable characters, 59 of which were parsi-moniously informative. Two equally parsimonious trees wereobtained, with a consistency index of 0.85 (Fig. 2). The topol-ogy of these trees does not contradict the phylogenies obtainedfrom the analyses of the separate data sets. On the contrary,support for the clades in the combined analysis are stronger.Data from the nuclear gene give strong support for the basal
LEON-REGAGNON ET AL.—HAEMATOLOECHUS FLOEDAE IN NORTH AND CENTRAL AMERICA 919
FIGURE 2. Phylogenetic hypothesis of populations of H. floedae and related species. (A) One of 2 most parsimonious trees obtained from theanalysis of the combined data sets of 28S rDNA and COI sequences. CI 5 0.849. (B) Alternative, most parsimonious hypothesis. Branch lengthshown above branches; bootstrap/decay indices shown below branches.

920 THE JOURNAL OF PARASITOLOGY, VOL. 91, NO. 4, AUGUST 2005
nodes, while the COI data set gives additional support for themonophyly of H. floedae and resolves relationships among pop-ulations of this species. In the trees obtained from the combinedanalysis, populations of H. floedae group in 2 clades, one in-cluding populations from Costa Rica and Yucatan, and a secondincluding populations from the United States. These clades havelow bootstrap and decay support, as well as little genetic di-vergence (Fig. 2). The clade including the populations of H.floedae is supported by high bootstrap and decay values, andshow large genetic distance from other related species of thegenus (Fig. 2).
According to the tree-based species delimitation approach forDNA sequence data proposed by Wiens and Penkrot (2002),species can be identified as being those well-supported basallineages in a haplotype phylogeny that are concordant with theirgeography. Following such criteria and with the genetic dis-tances observed in this study, populations from Yucatan, CostaRica, California, Georgia, and Arizona, despite their morpho-logical differences, belong to a single species, clearly differentfrom other species of the genus. In every case, populationscluster together with high branch support values, and are sep-arated from other species by a deep genetic divergence (Fig.2).
DISCUSSION
Recent molecular studies gave some support to the validityof H. floedae and H. parviplexus (Leon-Regagnon and Brooks,2003), also validating some morphological characters as the ar-rangement of the uterine loops, the shape of the reproductiveorgans, and the relative size of other structures such as thepharynx and suckers (Leon-Regagnon et al., 2001; Leon-Re-gagnon and Paredes-Calderon, 2002).
In the present paper, we give additional support to those stud-ies, finding that some morphological characters that are invari-able among distant populations of H. floedae, and also amongspecimens of other similar species (i.e., H. breviplexus, H. par-viplexus, and H. varioplexus). These characters can, therefore,be used to diagnosis species in this genus (i.e., the oral sucker-pharynx ratio, oral sucker-ventral sucker ratio, ovary and testesshape, extension of vitelline follicles [in fully developedworms], and extension of uterine longitudinal loops). Morpho-logical variation in the specimens found in Yucatan, Costa Rica,and the United States may be related to the developmental stageof the worms, and the use of different host (final or interme-diate) species in each locality.
The most common trait in Haematoloechus species is to havea restricted geographical distribution (Leon-Regagnon, 2003).Apparently, H. breviplexus is restricted to the northeastern Unit-ed States and eastern Canada, while H. parviplexus has beenfound only in the North central and eastern parts of the UnitedStates. Haematoloechus floedae, on the contrary, shows a widegeographical distribution ranging from South Carolina to Cali-fornia in the southern United States, and southern Mexico toCosta Rica. We think that the wide distribution of this speciesis due to the distribution of its original host species, R. cates-beiana. This frog species is widely distributed in eastern NorthAmerica, but it has been introduced to the West, where it isnow common. Equally, this species has been introduced to Cen-tral and South America. Its presence in Costa Rica and Yucatan
can be related with its culture and commercialization in the1960s and 1990s, respectively. Individuals of this species havesporadically been found in the wild in both countries, but thereis no record of established wild populations (Flores-Nava;Chaves and Puschendorf, pers. comm.). We were not able tofind specimens of R. catesbeiana in nature for helminthologicalexamination in these localities, but the genetic homogeneity in-dicates that the occurrence of H. floedae in native frog speciesfrom southern Mexico and Costa Rica could be due to a recentintroduction of the bull frog and the establishment of the par-asite life cycle in nature.
ACKNOWLEDGMENTS
We deeply acknowledge Dan Brooks, Valerie McKenzie, DavidGreen, and Anindo Choudhury for donation of specimens, and GregoryArjona and Isabel Jimenez for their help in the collection. We are alsograteful to the scientific and technical staff of the ACG for support ofthis study, in particular Elda Araya, Roger Blanco, Carolina Cano, Ma-ria Marta Chavarrıa, Felipe Chavarrıa, Roberto Espinoza, Dunia Garcia,Guillermo Jimenez, Elba Lopez, Sigifredo Marın, Alejandro Masis, Ca-lixto Moraga, Fredy Quesada, and Petrona Rios. Thanks also to DanJanzen and Winnie Hallwachs, scientific advisers to the ACG, for theirsupport. Special thanks to Laura Marquez Valdelamar (IBUNAM) forher help in the sequencing of samples; Luis Garcıa (CNHE), Erik Hob-erg, and Pat Pillitt (USNPC), and Scott Gardner and Skip Sterner(HWML) for the loan of specimens. This study was partially funded byCONACYT project J27985-N to V.L.R. and National Science Founda-tion grant DEB0102383 to Jonathan Campbell (University of Texas),V.L.R., S.G.H., and M.A.A.E.
LITERATURE CITED
CABALLERO Y C., E., AND D. SOKOLOFF. 1934. Segunda contribucion alconocimiento de la parasitologıa de Rana montezumae con un re-sumen, descripcion de una nueva especie y clave del genero Hae-matoloechus. Anales del Instituto de Biologıa, Universidad Na-cional Autonoma de Mexico 5: 5–40.
CHILTON, N. B., R. B. GASSER, AND I. BEVERIDGE. 1995. Differences ina ribosomal DNA sequence of morphologically indistinguishablespecies within the Hypodontus macropi complex (Nematoda:Strongyloidea). International Journal for Parasitology 25: 647–651.
CORT, W. W. 1915. North American frog lung flukes. Transactions ofthe American Microscopical Society 34: 203–243.
HALL, T. A. 1999. Bioedit: A user-friendly biological sequence align-ment editor and analysis program for Windows 95/98/NT. NucleicAcids Symposium Series 41: 95–98.
HARWOOD, P. D. 1932. The helminths parasitic in the Amphibia andReptilia in Houston, Texas and vicinity. Proceedings of the U.S.National Museum 81: 1–71.
HILLIS, D. M., B. K. MABLE, AND C. MORITZ. 1996. Nucleic acids IV:Sequencing and cloning. In Molecular systematics, D. M. Hillis, C.Moritz, and B. K. Mable (eds.). Sinauer, Sunderland, Massachu-setts, p. 321–383.
INGLES, L. G. 1932. Four new species of Haematoloechus (Trematoda)from Rana aurora draytoni from California. University of Califor-nia Publications in Zoology 37: 189–202.
———. 1936. Worm parasites of California amphibia. Transactions ofthe American Microscopical Society 53: 73–92.
KENNEDY, M. J. 1980a. Host-induced variations in Haematoloechus but-tensis (Trematoda: Haematoloechidae). Canadian Journal of Zool-ogy 58: 427–442.
———. 1980b. Geographical variation in some representatives of Hae-matoloechus, Looss, 1899 (Trematoda: Haematoloechidae) fromCanada and the United States. Canadian Journal of Zoology 58:1151–1167.
———. 1981. A revision of species of the genus HaematoloechusLooss, 1899 (Trematoda: Haematoloechidae) from Canada and theUnited States. Canadian Journal of Zoology 59: 1836–1846.
LEON-REGAGNON, V. 2003. Incorporating morphological and molecular

LEON-REGAGNON ET AL.—HAEMATOLOECHUS FLOEDAE IN NORTH AND CENTRAL AMERICA 921
data in biodiversity inventories. Parasites of leopard frogs. Journalof Parasitology 89: S141–S148.
———, AND D. R. BROOKS. 2003. Molecular phylogeny of Haemato-loechus Looss, 1899 (Digenea: Plagiorchiidae), with emphasis onNorth American species. Journal of Parasitology 89: 1206–1211.
———, ———, AND G. PEREZ-PONCE DE LEON. 1999. Differentiationof Mexican species of Haematoloechus Looss, 1899 (Digenea: Pla-giorchiformes): Molecular and morphological evidence. Journal ofParasitology 85: 935–946.
———, ———, AND D. A. ZELMER. 2001. Morphological and molec-ular description of Haematoloechus meridionalis n. sp. (Digenea:Plagiorchioidea: Haematoloechidae) from Rana vaillanti Brocchiof Guanacaste, Costa Rica. Journal of Parasitology 87: 1423–1427.
———, AND E. L. PAREDES-CALDERON. 2002. Haematoloechus dan-brooksi n. sp. (Digenea: Plagiorchioidea), from Rana vaillanti ofLos Tuxtlas, Veracruz, Mexico. Journal of Parasitology 88: 1215–1221.
MANTER, H. W. 1938. A collection of trematodes from Florida amphibia.Transactions of the American Microscopical Society 57: 26–38.
NADLER, S. A. 2002. Species delimitation and nematode biodiversity:Phylogenies rule. Nematology 4: 615–625.
ODENING, K. 1960. Plagiorchiidae III. (Haematoloechinae) und Om-phalometrinae. In Das Tierreich. Eine Zusammenstellung undKennzeichnung der rezenten Tierformen, R. Mertens and W. Hen-nig (eds.). Walter de Gruyter and Co., Berlin, Germany, p. 1–75.
PALUMBI, S. R. 1996. Nucleic acids II: The polymerase chain reaction.In Molecular systematics, D. M. Hillis, C. Moritz, and B. K. Mable(eds.). Sinauer, Sunderland, Massachusetts, p. 205–247.
PROKOPIC, J., AND K. KRIVANEK. 1974. Trematodes of the genus Hae-matoloechus Looss, and their variability. Helminthologia 15: 779–802.
SNYDER, S., AND V. TKACH. 2001. Phylogenetic and biogeographicalrelationships among some holarctic frog lung flukes (Digenea: Hae-matoloechidae). Journal of Parasitology 87: 1433–1440.
STOCK, S. P., J. HENG, D. J. HUNT, A. P. REID, X. SHEN, AND H. Y. CHOO.2001. Redescription of Steinernema longicaudum Shen and Wang(Nematoda: Steinernematidae); geographic distribution and phe-notypic variation between allopatric populations. Journal of Hel-minthology 75: 81–92.
SWOFFORD, D. L. 2002. PAUP*. Phylogenetic analysis using parsimony(*and other methods). Version 4. Sinauer Associates, Sunderland,Massachusetts.
VERNEAU, O., F. RENAUD, AND F. CATZEFLIS. 1997. Evolutionary rela-tionships of sibling tapeworm species (Cestoda) parasitizing teleostfishes. Molecular Biology and Evolution 14: 630–636.
WEINS, J. J. 1998. Combining data sets with different phylogenetic his-tories. Systematic Biology 47: 568–581.
———, AND T. A. PENKROT. 2002. Delimiting species using DNA andmorphological variation and discordant species limits in spiny liz-ards (Sceloporus). Systematic Biology 51: 69–91.
Copyright © 2022 FDOKUMEN




















