
The betagamma-crystallin superfamily contains a universal motif for binding calcium
Upload
independentCategory
view
4download
0
Molecular evidence that the genus Cadenatella Dollfus, 1946
(Digenea: Plagiorchiida) belongs in the superfamily
Haploporoidea Nicoll, 1914
Rodney A. Bray • Thomas H. Cribb •
Andrea Waeschenbach • D. Timothy J. Littlewood
Received: 17 April 2014 / Accepted: 9 June 2014
� Springer Science+Business Media Dordrecht 2014
Abstract A re-examination of published lsrDNA
sequence data of haploporid digeneans has shown that
the genus Cadenatella Dollfus, 1946, hitherto consid-
ered a lepocreadioid, is correctly placed within the
superfamily Haploporoidea Nicoll, 1914, although its
relationships within the superfamily are not resolved.
The morphological similarities and differences
between Cadenatella and other haploporoids are
discussed, and the subfamily Cadenatellinae Gibson
& Bray, 1982 is considered the best repository for
Cadenatella spp. at present.
Introduction
Molecular phylogenetics has produced some surpris-
ing results, placing species in situations where their
morphology does not, on first examination, seem to fit.
In the Digenea, for example, the genus Cableia
Sogandares-Bernal, 1959 was originally considered a
lepocreadiid (Sogandares-Bernal, 1959), later an op-
ecoelid (Yamaguti, 1971), then an enenterid (Gibson
& Bray, 1982) and finally an acanthocolpid (Bray
et al., 1996). Molecular studies, based on small subunit
nuclear ribosomal RNA gene (ssrDNA) (=18S rDNA)
and partial large subunit nuclear ribosomal RNA gene
(lsrDNA) (=28S rDNA) sequences, place this genus as
an early divergent monorchiid (Cribb et al., 2001;
Olson et al., 2003; Bray et al., 2005; unpublished). In
the study reported here, we present molecular evi-
dence that another puzzling genus, Cadenatella Doll-
fus, 1946, is not an enenterid, as has been considered
since its discovery, but a member of the Haploporoi-
dea Nicoll, 1914.
Materials and methods
Taxon choice
In this study, we have made use of the considerable
amount of published lsrDNA data available on Gen-
Bank (as at January 2013) to conduct a trematode-wide
survey of this gene. Included sequences were those of
[500 base pair length; duplicate sequences which
showed\1% sequence divergencewere excluded, thus
leaving us with information for discrete species only.
The final dataset consisted of 556 trematode sequences
representing 24 superfamilies and 97 families and
included two Cadenatella spp. This molecular phy-
logeny was utilised, but not published in full, by
Littlewood et al. (2015) in their survey of the biodi-
versity of helminth endoparasites. It indicated, with
strong support, that theCadenatella spp. were included
within the superfamily Haploporoidea. Here, a further
R. A. Bray (&) � A. Waeschenbach � D. T. J. Littlewood
Department of Life Sciences, Natural History Museum,
Cromwell Road, London SW7 5BD, UK
e-mail: [email protected]
T. H. Cribb
School of Biological Sciences, The University of
Queensland, Brisbane, QLD 4072, Australia
123
Syst Parasitol (2014) 89:15–21
DOI 10.1007/s11230-014-9504-5
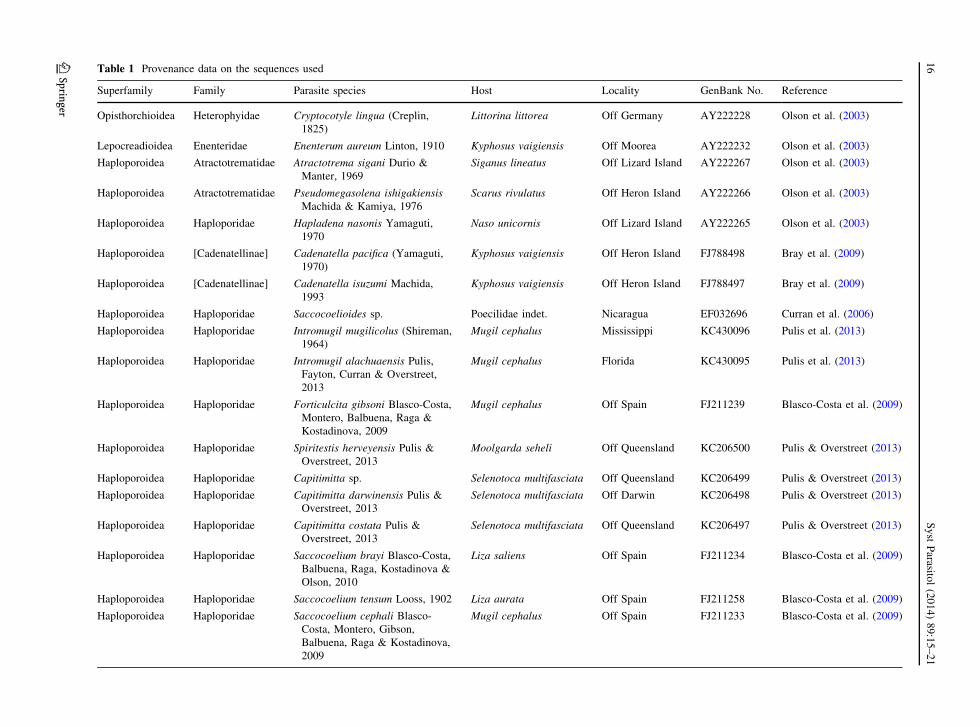
Table 1 Provenance data on the sequences used
Superfamily Family Parasite species Host Locality GenBank No. Reference
Opisthorchioidea Heterophyidae Cryptocotyle lingua (Creplin,
1825)
Littorina littorea Off Germany AY222228 Olson et al. (2003)
Lepocreadioidea Enenteridae Enenterum aureum Linton, 1910 Kyphosus vaigiensis Off Moorea AY222232 Olson et al. (2003)
Haploporoidea Atractotrematidae Atractotrema sigani Durio &
Manter, 1969
Siganus lineatus Off Lizard Island AY222267 Olson et al. (2003)
Haploporoidea Atractotrematidae Pseudomegasolena ishigakiensis
Machida & Kamiya, 1976
Scarus rivulatus Off Heron Island AY222266 Olson et al. (2003)
Haploporoidea Haploporidae Hapladena nasonis Yamaguti,
1970
Naso unicornis Off Lizard Island AY222265 Olson et al. (2003)
Haploporoidea [Cadenatellinae] Cadenatella pacifica (Yamaguti,
1970)
Kyphosus vaigiensis Off Heron Island FJ788498 Bray et al. (2009)
Haploporoidea [Cadenatellinae] Cadenatella isuzumi Machida,
1993
Kyphosus vaigiensis Off Heron Island FJ788497 Bray et al. (2009)
Haploporoidea Haploporidae Saccocoelioides sp. Poecilidae indet. Nicaragua EF032696 Curran et al. (2006)
Haploporoidea Haploporidae Intromugil mugilicolus (Shireman,
1964)
Mugil cephalus Mississippi KC430096 Pulis et al. (2013)
Haploporoidea Haploporidae Intromugil alachuaensis Pulis,
Fayton, Curran & Overstreet,
2013
Mugil cephalus Florida KC430095 Pulis et al. (2013)
Haploporoidea Haploporidae Forticulcita gibsoni Blasco-Costa,
Montero, Balbuena, Raga &
Kostadinova, 2009
Mugil cephalus Off Spain FJ211239 Blasco-Costa et al. (2009)
Haploporoidea Haploporidae Spiritestis herveyensis Pulis &
Overstreet, 2013
Moolgarda seheli Off Queensland KC206500 Pulis & Overstreet (2013)
Haploporoidea Haploporidae Capitimitta sp. Selenotoca multifasciata Off Queensland KC206499 Pulis & Overstreet (2013)
Haploporoidea Haploporidae Capitimitta darwinensis Pulis &
Overstreet, 2013
Selenotoca multifasciata Off Darwin KC206498 Pulis & Overstreet (2013)
Haploporoidea Haploporidae Capitimitta costata Pulis &
Overstreet, 2013
Selenotoca multifasciata Off Queensland KC206497 Pulis & Overstreet (2013)
Haploporoidea Haploporidae Saccocoelium brayi Blasco-Costa,
Balbuena, Raga, Kostadinova &
Olson, 2010
Liza saliens Off Spain FJ211234 Blasco-Costa et al. (2009)
Haploporoidea Haploporidae Saccocoelium tensum Looss, 1902 Liza aurata Off Spain FJ211258 Blasco-Costa et al. (2009)
Haploporoidea Haploporidae Saccocoelium cephali Blasco-
Costa, Montero, Gibson,
Balbuena, Raga & Kostadinova,
2009
Mugil cephalus Off Spain FJ211233 Blasco-Costa et al. (2009)
16
Syst
Parasito
l(2014)89:15–21
123
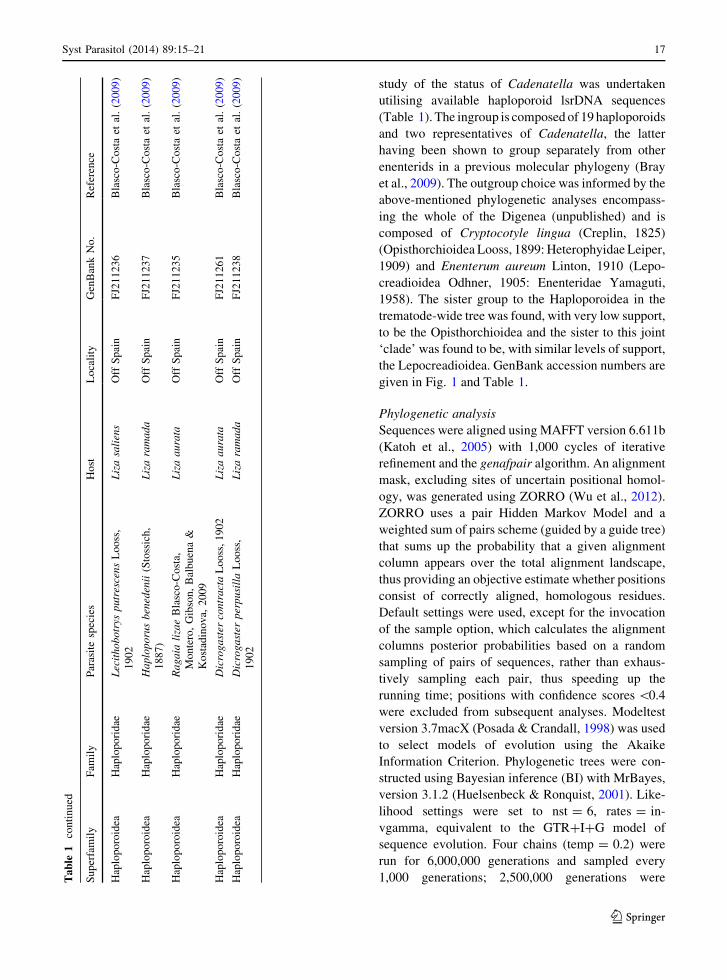
study of the status of Cadenatella was undertaken
utilising available haploporoid lsrDNA sequences
(Table 1). The ingroup is composed of 19 haploporoids
and two representatives of Cadenatella, the latter
having been shown to group separately from other
enenterids in a previous molecular phylogeny (Bray
et al., 2009). The outgroup choice was informed by the
above-mentioned phylogenetic analyses encompass-
ing the whole of the Digenea (unpublished) and is
composed of Cryptocotyle lingua (Creplin, 1825)
(Opisthorchioidea Looss, 1899:Heterophyidae Leiper,
1909) and Enenterum aureum Linton, 1910 (Lepo-
creadioidea Odhner, 1905: Enenteridae Yamaguti,
1958). The sister group to the Haploporoidea in the
trematode-wide tree was found, with very low support,
to be the Opisthorchioidea and the sister to this joint
‘clade’ was found to be, with similar levels of support,
the Lepocreadioidea. GenBank accession numbers are
given in Fig. 1 and Table 1.
Phylogenetic analysis
Sequences were aligned using MAFFT version 6.611b
(Katoh et al., 2005) with 1,000 cycles of iterative
refinement and the genafpair algorithm. An alignment
mask, excluding sites of uncertain positional homol-
ogy, was generated using ZORRO (Wu et al., 2012).
ZORRO uses a pair Hidden Markov Model and a
weighted sum of pairs scheme (guided by a guide tree)
that sums up the probability that a given alignment
column appears over the total alignment landscape,
thus providing an objective estimate whether positions
consist of correctly aligned, homologous residues.
Default settings were used, except for the invocation
of the sample option, which calculates the alignment
columns posterior probabilities based on a random
sampling of pairs of sequences, rather than exhaus-
tively sampling each pair, thus speeding up the
running time; positions with confidence scores\0.4
were excluded from subsequent analyses. Modeltest
version 3.7macX (Posada & Crandall, 1998) was used
to select models of evolution using the Akaike
Information Criterion. Phylogenetic trees were con-
structed using Bayesian inference (BI) with MrBayes,
version 3.1.2 (Huelsenbeck & Ronquist, 2001). Like-
lihood settings were set to nst = 6, rates = in-
vgamma, equivalent to the GTR?I?G model of
sequence evolution. Four chains (temp = 0.2) were
run for 6,000,000 generations and sampled every
1,000 generations; 2,500,000 generations wereTable
1continued
Superfamily
Fam
ily
Parasitespecies
Host
Locality
GenBankNo.
Reference
Haploporoidea
Haploporidae
LecithobotrysputrescensLooss,
1902
Lizasaliens
OffSpain
FJ211236
Blasco-Costaet
al.(2009)
Haploporoidea
Haploporidae
Haploporusbenedenii(Stossich,
1887)
Lizaramada
OffSpain
FJ211237
Blasco-Costaet
al.(2009)
Haploporoidea
Haploporidae
Ragaia
lizaeBlasco-Costa,
Montero,Gibson,Balbuena&
Kostadinova,
2009
Lizaaurata
OffSpain
FJ211235
Blasco-Costaet
al.( 2009)
Haploporoidea
Haploporidae
Dicrogastercontracta
Looss,1902
Lizaaurata
OffSpain
FJ211261
Blasco-Costaet
al.( 2009)
Haploporoidea
Haploporidae
DicrogasterperpusillaLooss,
1902
Lizaramada
OffSpain
FJ211238
Blasco-Costaet
al.(2009)
Syst Parasitol (2014) 89:15–21 17
123

discarded as burn-in, at which point the average
standard deviation of split frequencies were\0.01.
Results
The lsrDNA tree (Fig. 1) shows the superfamily
Haploporoidea Nicoll, 1914, including the families
Atractotrematidae Yamaguti, 1939 and Haploporidae
Nicoll, 1914 and the Cadenatella spp. as a well-
supported monophyletic group; the topology is iden-
tical to that obtained in the lsrDNA analysis by
Littlewood et al. (2015), apart from the addition of the
recently published sequences of the haploporids
Intromugil mugilicolus (Shireman, 1964), I. alachua-
ensis Pulis, Fayton, Curran & Overstreet, 2013,
Spiritestis herveyensis Pulis & Overstreet, 2013, an
unnamed species of Capitimitta Pulis & Overstreet,
2013, C. darwinensis Pulis & Overstreet, 2013 and
C. costata Pulis & Overstreet, 2013, which had not
been included by Littlewood et al. (2015). The
Atractotrematidae, represented here by just two spe-
cies, is also well supported as monophyletic. The two
species of Cadenatella are monophyletic, forming a
group for which the subfamily name Cadenatellinae
Gibson & Bray, 1982 is used here, it being in our
opinion premature to decide on the hierarchical status
of this clade. These results strongly indicate that
Cadenatella is not an enenterid, or indeed a lepoc-
readioid, as previously thought.
The Haploporidae is less well supported in that
Hapladena Linton, 1910 is not clustered with the
remainder of the Haploporidae. Within the Haplopo-
ridae, there are two well-supported groups: a sister
Fig. 1 Bayesian inference analysis of published haploporoid lsrDNA sequences, constructed using MrBayes under the GTR?I?G
model. Analyses were run for 6,000,000 generations with 2,500,000 generations discarded as burn-in. The branch-length scale-bar
indicates the number of substitutions per site
18 Syst Parasitol (2014) 89:15–21
123

group relationship between Saccocoelioides Szidat,
1954 and IntromugilOverstreet & Curran, 2005, and a
clade in which species of Saccocoelium Looss, 1902
form the sister group to an unresolved assemblage of
(Lecithobotrys Looss, 1902, Haploporus Looss,
1902), Ragaia lizae Blasco-Costa, Montero, Gibson,
Balbuena & Kostadinova, 2009 and species of
Dicrogaster Looss, 1902. However, the interrelation-
ships between those two lineages, Forticulcita gibsoni
Blasco-Costa, Montero, Balbuena, Raga & Kostadi-
nova, 2009, Spiritestis herveyensis and Capitimitta
spp. are unresolved.
Discussion
The Haploporoidea
The monophyly of the Haploporoidea, including
Cadenatella, is well supported, thus supplying strong
evidence of the superfamilial position of Cadenatella.
The haploporid subfamily Haploporinae is well sup-
ported (including the genera Saccocoelium, Lecitho-
botrys, Haploporus, Ragaia Blasco-Costa, Montero,
Gibson, Balbuena & Kostadinova, 2009 and Dicrog-
aster), but the subfamily status of other haploporids is
less clear. Blasco-Costa et al. (2009) erected the
subfamily Forticulcitinae Blasco-Costa, Balbuena,
Kostadinova & Olson, 2009 for Forticulcita Over-
street, 1982 and retained Saccocoelioides within the
Chalcinotrematinae Overstreet & Curran 2005 fol-
lowing Overstreet & Curran (2005). Pulis & Over-
street (2013) produced sequences of Spiritestis
Nagaty, 1948 and Capitimitta, which we have
included in our analysis, but their relationships within
the family are not resolved. The sequences of Introm-
ugil spp. produced by Pulis et al. (2013) cluster with
Saccocoelioides sp. adding weight to their statement
that Intromugil ‘‘will be more closely allied with the
chalcinotrematines’’ although no molecular data are
yet available for Chalcinotrema Freitas, 1947.
Cadenatella
Dollfus (1946) originally erected Cadenatella as a
subgenus of Enenterum Linton, 1910 and in the same
paper erected another subgenus, Jeancadenatia Doll-
fus, 1946. Manter (1947) considered that both sub-
genera probably warranted generic rank and Nagaty
(1948) used both names at the generic level. Nahhas &
Cable (1964) synonymised Jeancadenatia with Ca-
denatella, a situation followed by many subsequent
workers. Recent revisions have all considered Caden-
atella an enenterid because of the general morpho-
logical similarity and especially, superficially, of the
oral sucker lobation and the restriction of Cadenatella
to hosts of the genus Kyphosus which is the most
important host taxon for enenterids (Brooks et al.,
2000; Bray & Cribb, 2001, 2002). Molecular evidence
in Bray et al. (2009) showed that Cadenatella was not
a lepocreadioid, and, therefore, not an enenterid. The
sample size used in their analysis did not produce
convincing evidence of the status of Cadenatella.
Jones (2005) and Overstreet & Curran (2005)
considered that members of the Haploporoidea are
characterised by the presence of a sac enclosing the
terminal genitalia, considered a ‘hermaphroditic sac’.
Cadenatella lacks a hermaphroditic sac, but the exact
condition of the terminal genitalia is variably
described. Dollfus (1946) described the male terminal
genitalia of the type-species C. cadenatiDollfus, 1946
as ‘‘La poche de cirre occupe l’espace entre la
bifurcation intestinale et bord anterieur de l’acetabu-
lum, il y a une vesicule seminale interne et des cellules
prostatiques; il ne semble exist de vesicule seminale
externe.’’ The other species described by Dollfus
(1946), C. brumpti (Dollfus, 1946), originally placed
in the subgenus Jeancadenatia, has ‘‘La poche de
cirre, extremement petit et mal distincte, est situee au
contact du bord anterieur gauche de l’acetabulum’’.
Manter (1949) described the terminal genitalia of
C. americana Manter, 1949 as ‘‘Cirrus sac thin-
walled, small and very inconspicuous’’ and described
an external seminal vesicle. Nahhas & Cable (1964)
stated that ‘‘reexamination of the holotype reveals that
the pars prostatica probably was misinterpreted as a
cirrus sac’’. Overstreet (1969) examined further spec-
imens and stated that a ‘‘sectioned specimen does not
have a cirrus sac, confirming what Nahhas and Cable
(1964: 192) believed. A thin membrane, however,
appears to surround the vesicle in some whole-
mounts’’. Winter (1957) described a large cirrus-sac
in C. dohenyi (Winter, 1957), and presented a micro-
photograph which shows a large, sperm-filled sac (?
seminal vesicle) in the posterior forebody. Yamaguti
(1970) stated that C. pacifica (Yamaguti, 1970) had
‘‘no cirrus-sac’’ a situation confirmed by Machida
(1993) and Bray & Cribb (2001). According to
Syst Parasitol (2014) 89:15–21 19
123

Hafeezullah (1980), C. dollfusi (Hafeezullah, 1980)
has a ‘‘short cirrus sac … enclosing short tubular
internal seminal vesicle’’. Machida (1993) apparently
found the cirrus-sac absent in C. isuzumi Machida,
1993 and Bray & Cribb (2001) confirmed this finding.
Brooks et al. (2000) re-examined specimens of
C. americana, C. dohenyi, C. kyphosi Nahhas &
Cable, 1964, C. floridae Overstreet, 1969, C. isuzumi
and C. pacifica and found the cirrus-sac missing in all
but C. dohenyi, which they considered along with
C. brumpti and C. dollfusi, to have a cirrus-sac
‘‘partially enclosing the seminal vesicle’’. In no case
was a hermaphroditic sac described. Gibson & Bray
(1982) erected the enenterid subfamily Cadenatelli-
nae, to contain Cadenatella (and its now recognised
synonym Jeancadenatia Dollfus, 1946), based, inter
alia, on the lack of a cirrus-sac.
Nahhas & Cable (1964) commented ‘‘it may be
noted that the genera Enenterum, Cadenatella, and
Jeancadenatia which generally are placed in the
Lepocreadiidae, have many features in common with
those of families included in the Haploporoidea and
ultimately may be transferred to that superfamily’’.
The major characteristic features of Cadenatella are
the elaborately lobed oral sucker, the uroproct and the
single testis. Comparison with the superfamily diag-
nosis of the Haploporoidea in Jones (2005) reinforces
the view that most features of Cadenatella are shared
with members of this superfamily. We know that
complex lobation of the oral sucker occurs in the
Haploporoidea, e.g. in Waretrema Srivastava, 1937,
Spiritestis Nagaty, 1948, Capitimitta Pulis & Over-
street, 2013. Pulis & Overstreet (2013) illustrated
examples from these genera, both with microphoto-
graphs and line drawings. An uroproct is shared with
the genusMyoderaMontgomery, 1957 (see Overstreet
& Curran, 2005), whose species are also reported in
kyphosids (Montgomery, 1957; Sogandares-Bernal,
1959). A single testis is also the usual condition in the
Haploporidae. In fact, Overstreet & Curran (2005)
considered it possible that the three genera whose
members exhibit two testes, Vitellibaculum Mont-
gomery, 1957, Metamegasolena Yamaguti, 1970 and
Megasolena Linton, 1910, ‘‘do not even belong in the
family Haploporidae’’. All of the haploporoid species
used in the current study have a single testis.
The evidence from lsrDNA is, therefore, strong that
Cadenatella has been misplaced by previous workers.
The topology of the tree (Fig. 1) suggests that the
terminal genitalia of Cadenatella are derived from the
‘hermaphroditic sac’ by loss of the wall. The lobation
of the oral sucker is not homologous with that of
species of Enenterum and, in detail, the lobation
appears rather distinct (see Bray & Cribb, 2001,
figures 2, 7 vs 14, 15, 18, 19). The current placement
of Cadenatella in the haploporoid tree is poorly
resolved. Convincing relationships within the Hap-
loporoidea await the molecular study of many more
haploporoids, but it is probable that the subfamily
Cadenatellinae will be recognised at the family level.
In retrospect, the confusion relating to the position of
Cadenatella can be seen as having arisen from a case
of convergent evolution driven, apparently, by the
nature of the habitat provided by the kyphosid gut. We
note too that identity as a haploporoid, which implies a
likely two-host life cycle in which the cercariae encyst
in the open (Bartoli & Gibson, 2007), is entirely
consistent with the herbivorous diet of species of
Kyphosus.
Acknowledgements We would like to thank Peter Foster
(NHM) for assistance in compiling the original data set drawn
from GenBank.
References
Bartoli, P., & Gibson, D. I. (2007). Synopsis of the life cycles of
Digenea (Platyhelminthes) from lagoons of the northern
coast of the western Mediterranean. Journal of Natural
History, 41, 1553–1570.
Blasco-Costa, I., Balbuena, J. A., Kostadinova, A., & Olson, P.
D. (2009). Interrelationships of the Haploporinae (Dige-
nea: Haploporidae): A molecular test of the taxonomic
framework based on morphology. Parasitology Interna-
tional, 58, 263–269.
Bray, R. A., & Cribb, T. H. (2001). A review of the family
Enenteridae Yamaguti, 1958 (Digenea), with descriptions
of species from Australian waters, including Koseiria
huxleyi n. sp. Systematic Parasitology, 48, 1–29.
Bray, R. A., & Cribb, T. H. (2002). Further observations on the
Enenteridae Yamaguti, 1958 (Digenea: Lepocreadioidea)
of the Indo-West Pacific region, including a new species
fromWestern Australia. Acta Parasitologica, 47, 208–223.
Bray, R. A., Cribb, T. H., & Barker, S. C. (1996).Cableia pudica
n. sp. (Digenea: Acanthocolpidae) from monacanthid
fishes of the southern Great Barrier Reef, Australia. Par-
asite, 3, 49–54.
Bray, R. A., Waeschenbach, A., Cribb, T. H., Weedall, G. D.,
Dyal, P., & Littlewood, D. T. J. (2009). The phylogeny of
the Lepocreadioidea (Platyhelminthes: Digenea) inferred
from nuclear and mitochondrial genes: implications for
their systematics and evolution. Acta Parasitologica, 54,
310–329.
20 Syst Parasitol (2014) 89:15–21
123

Bray, R. A., Webster, B. L., Bartoli, P., & Littlewood, D. T. J.
(2005). Relationships within the Acanthocolpidae Luhe,
1906 and their place among the Digenea. Acta Parasito-
logica, 50, 281–291.
Brooks, D. R., Perez-Ponce de Leon, G., & Leon-Regagnon, V.
(2000). Phylogenetic analysis of the Enenterinae (Digenea,
Lepocreadiidae) and discussion of the evolution of the
digenean fauna of kyphosid fishes. Zoologica Scripta, 29,
237–246.
Cribb, T. H., Bray, R. A., Littlewood, D. T. J., Pichelin, S., &
Herniou, E. A. (2001). The Digenea. In: Littlewood,
D. T. J. & Bray, R. A. (Eds) Interrelationships of the
Platyhelminthes. London: Taylor & Francis, pp. 168–185.
Curran, S. S., Tkach, V. V., & Overstreet, R. M. (2006). A
review of Polylekithum Arnold, 1934 and its familial
affinities using morphological and molecular data, with
description of Polylekithum catahoulensis sp. nov. Acta
Parasitologica, 51, 238–248.
Dollfus, R. P. (1946). Sur trois especes de distomes, dont une a
17 ventouses (Enenterum (Jeancadenatia) brumpti n. sp.)
parasites du poisson marin Kyphosus sectatrix (L.). An-
nales de Parasitologie Humaine et Comparee, 21,
119–128.
Gibson, D. I., & Bray, R. A. (1982). A study and reorganization
of Plagioporus Stafford, 1904 (Digenea: Opecoelidae) and
related genera, with special reference to forms from
European Atlantic waters. Journal of Natural History, 16,
529–559.
Hafeezullah, M. (1980). Two digenetic trematodes of a marine
fish, Kyphosus cinerascens (Forskal) from the Gulf of
Mannar with a note on the systematic positions of the
genera Enenterum Linton, 1910, Cadenatella Dollfus,
1946 and Jeancadenatia Dollfus, 1946. Bulletin of the
Zoological Survey of India, 2, 145–151.
Huelsenbeck, J. P., & Ronquist, F. (2001). MRBAYES:
Bayesian inference of phylogenetic trees. Bioinformatics,
17, 754–755.
Jones, A. (2005). Superfamily Haploporoidea Nicoll, 1914. In:
Jones, A., Bray, R. A., & Gibson, D. I. (Eds) Keys to the
Trematoda, Volume 2. Wallingford: CABI Publishing and
the Natural History Museum, pp. 127–128.
Katoh, K., Kuma, K.-I., Toh, H., & Miyata, T. (2005). MAFFT
version 5: Improvement in accuracy of multiple sequence
alignment. Nucleic Acids Research, 33, 511–518.
Littlewood, D. T. J., Bray, R. A., & Waeschenbach, A. (2015).
Phylogenetic patterns of diversity in cestodes and trema-
todes. In: Morand, S., Krasnov, B. & Littlewood, D. T. J.
(Eds) Parasite Diversity and Diversification: Evolutionary
Ecology meets Phylogenetics. Cambridge: Cambridge
University Press (in press).
Machida, M. (1993). Trematodes from kyphosid fishes in Jap-
anese and adjacent waters. Bulletin of the National Science
Museum, Tokyo. Series A. Zoology, 19, 27–36.
Manter, H.W. (1947). The digenetic trematodes of marine fishes
of Tortugas, Florida. American Midland Naturalist, 38,
257–416.
Manter, H. W. (1949). An additional trematode from Tortugas,
Florida, and new name for Opisthoporus Manter, 1947,
preoccupied. American Midland Naturalist, 41, 432–435.
Montgomery, W. R. (1957). Studies on digenetic trematodes
from marine fishes of La Jolla, California. Transactions of
the American Microscopical Society, 76, 13–36.
Nagaty, H. F. (1948). Trematodes of fishes from the Red Sea.
Part 4. On some new and known forms with a single testis.
Journal of Parasitology, 34, 355–363.
Nahhas, F. M., & Cable, R. M. (1964). Digenetic and aspidog-
astrid trematodes from marine fishes of Curacao and
Jamaica. Tulane Studies in Zoology, 11, 169–228.
Olson, P. D., Cribb, T. H., Tkach, V. V., Bray, R. A., & Lit-
tlewood, D. T. J. (2003). Phylogeny and classification of
the Digenea (Platyhelminthes: Trematoda). International
Journal for Parasitology, 33, 733–755.
Overstreet, R. M. (1969). Digenetic trematodes of marine teleost
fishes from Biscayne Bay, Florida. Tulane Studies in
Zoology and Botany, 15, 119–176.
Overstreet, R. M., & Curran, S. S. (2005). Family Haploporidae
Nicoll, 1914. In: Jones, A., Bray, R. A., & Gibson, D. I.
(Eds)Keys to the Trematoda, Volume 2.Wallingford: CABI
Publishing and the Natural History Museum, pp. 129–165.
Posada, D., & Crandall, K. A. (1998). MODELTEST: Testing
the model of DNA substitution. Bioinformatics, 14,
817–818.
Pulis, E. E., Fayton, T. J., Curran, S. S., & Overstreet, R. M.
(2013). A new species of Intromugil (Digenea: Haplopo-
ridae) and redescription of Intromugil mugilicolus. Journal
of Parasitology, 99, 501–508.
Pulis, E. E., & Overstreet, R. M. (2013). Review of haploporid
(Trematoda) genera with ornate muscularisation in the
region of the oral sucker, including four new species and a
new genus. Systematic Parasitology, 84, 167–191.
Sogandares-Bernal, F. (1959). Digenetic trematodes of marine
fishes from the Gulf of Panama and Bimini, British West
Indies. Tulane Studies in Zoology, 7, 69–117.
Winter, H. A. (1957). Trematodos de peces marinos de aguas
Mexicanas. XII. Dos generos de digeneos (Lepocreadii-
dae), incluyendo una nueva especie procedente de Ky-
phosus elegans (Peters) de las Islas Tres Marias, en el
Oceano Pacifico. Anales del Instituto de Biologıa. Uni-
versidad de Mexico, 27, 403–413.
Wu, M., Chatterji, S., & Eisen, J. A. (2012). Accounting for
alignment uncertainty in phylogenomics. PLOS One, 7,
e30288.
Yamaguti, S. (1970). Digenetic trematodes of Hawaiian fishes.
Tokyo: Keigaku, 436 pp.
Yamaguti, S. (1971). Synopsis of digenetic trematodes of ver-
tebrates. Tokyo: Keigaku, Vol. I, 1074 pp.; Vol. II, 349 pp.
Syst Parasitol (2014) 89:15–21 21
123
Copyright © 2022 FDOKUMEN




















