
Relating biochemistry and function in the myosin superfamily
7
Relating biochemistry and function in the myosin superfamily $ Enrique M De La Cruz 1 and E Michael Ostap 2 All characterized myosins share a common ATPase mechanism. However, detailed kinetic analyses suggest that modulation of the rate and equilibrium constants that define the ATPase cycle confers specific properties to these motor proteins, suiting them to specific physiological tasks. Understanding the kinetic mechanisms allows potential cellular functions of the different myosin classes and isoforms to be better defined. Addresses 1 Department of Molecular Biophysics and Biochemistry, Yale University, PO Box 208114, New Haven, CT 06520, USA e-mail: [email protected] 2 Pennsylvania Muscle Institute and Department of Physiology, University of Pennsylvania School of Medicine, B400 Richards, Philadelphia, PA 19104, USA e-mail: [email protected] Current Opinion in Cell Biology 2004, 16:61–67 This review comes from a themed issue on Cell structure and dynamics Edited by John A Cooper and Margaret A Titus 0955-0674/$ – see front matter ß 2003 Elsevier Ltd. All rights reserved. DOI 10.1016/j.ceb.2003.11.011 Abbreviations A actin M myosin P i inorganic phosphate Introduction The myosin superfamily of actin-based molecular motors consists of at least 18 members with diverse properties adapted for a variety of cellular activities [1,2]. For example, some myosins exist in large macromolecular assemblies that power contraction of muscle, while others work in small numbers to support the translocation of single organelles. Recent studies from several laboratories have probed the mechanisms of a diverse set of myosin classes and isoforms. Our goal is to provide a brief over- view of the ATPase mechanisms of a few members of the myosin superfamily and to point out how differences in biochemical rate constants provide myosins with motor properties tuned for diverse biological functions. Overview of the myosin ATPase cycle The actomyosin ATPase cycle mechanism appears to be conserved for all myosin isoforms. The reaction scheme in Figure 1 shows the mechanism of the actomyosin ATPase pathway coupled to a mechanical model [3,4]. Although it is an oversimplification of what is known about chemo- mechanical coupling, the scheme provides the basic details required for understanding the biochemical diver- sity of myosins and will be referred to throughout the kinetic discussions in this review. For more detailed discussion of chemomechanical coupling and detailed references concerning the actomyosin ATPase cycle, we recommend the excellent textbook by Howard [5] and the review by Geeves and Holmes [6]. The minimum mechanism needed to describe the myo- sin ATPase cycle consists of at least six nucleotide-linked biochemical reactions. These are described in detail in Figure 1. Intermediates are defined according to their affinities for actin. The M.ATP and M.ADP.P i states are ‘weak- binding’ or ‘pre-force generating’ intermediates that attach and detach (steps 2 and 4) from the actin filament on a submicrosecond scale, have low (micromolar) actin affinities, and do not bind stereospecifically to the actin filament [6]. The weak-binding states do not bear force, but allow myosin to ‘cock’ itself for the power-stroke while detached from actin (step 3), preventing a reverse power-stroke while attached to actin. The AM and AM.ADP states are ‘strong-binding’ intermediates that stereospecifically attach to actin filaments with high (submicromolar) affinities. The strong-binding states are the ‘force-bearing’ intermediates. The force-generating power-stroke coincides with phos- phate release at the transition from the weakly bound to strongly bound states (step 5, Figure 1). Because it con- trols the weak-to-strong transition, P i release is an ideal step for controlling myosin–actin attachment and moti- lity. The transition is regulated by the calcium regulatory system in striated muscle and by heavy-chain phosphor- ylation of some myosin-I isoforms [7]. Enzymatic adaptations of myosin classes and isoforms Two important parameters that influence the mechanical and motile properties of myosins are, first, the lifetimes (i.e. the duration of the biochemical states in units of time) of the strongly bound intermediates (AM and AMD), and, second, the duty ratio. The attachment lifetimes and duty ratios can vary considerably between myosin classes and between isoforms within a given class. The duty ratio of a molecular motor is defined as the fraction of the total ATPase cycle time that an individual $ Supplementary data associated with this article can be found, in the online version, at doi:10.1016/j.ceb.2003.11.011 www.sciencedirect.com Current Opinion in Cell Biology 2004, 16:61–67
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Relating biochemistry and function in the myosin superfamily

Relating biochemistry and function in the myosin superfamily$
Enrique M De La Cruz1 and E Michael Ostap2
All characterized myosins share a common ATPase mechanism.
However, detailed kinetic analyses suggest that modulation
of the rate and equilibrium constants that define the ATPase
cycle confers specific properties to these motor proteins, suiting
them to specific physiological tasks. Understanding the kinetic
mechanisms allows potential cellular functions of the
different myosin classes and isoforms to be better defined.
Addresses1Department of Molecular Biophysics and Biochemistry, Yale University,PO Box 208114, New Haven, CT 06520, USA
e-mail: [email protected] Muscle Institute and Department of Physiology, University
of Pennsylvania School of Medicine, B400 Richards, Philadelphia,
PA 19104, USA
e-mail: [email protected]
Current Opinion in Cell Biology 2004, 16:61–67
This review comes from a themed issue on
Cell structure and dynamicsEdited by John A Cooper and Margaret A Titus
0955-0674/$ – see front matter
� 2003 Elsevier Ltd. All rights reserved.
DOI 10.1016/j.ceb.2003.11.011
AbbreviationsA actin
M myosin
Pi inorganic phosphate
IntroductionThe myosin superfamily of actin-based molecular motors
consists of at least 18 members with diverse properties
adapted for a variety of cellular activities [1,2]. For
example, some myosins exist in large macromolecular
assemblies that power contraction of muscle, while others
work in small numbers to support the translocation of
single organelles. Recent studies from several laboratories
have probed the mechanisms of a diverse set of myosin
classes and isoforms. Our goal is to provide a brief over-
view of the ATPase mechanisms of a few members of the
myosin superfamily and to point out how differences in
biochemical rate constants provide myosins with motor
properties tuned for diverse biological functions.
Overview of the myosin ATPase cycleThe actomyosin ATPase cycle mechanism appears to be
conserved for all myosin isoforms. The reaction scheme in
Figure 1 shows the mechanism of the actomyosin ATPase
pathway coupled to a mechanical model [3,4]. Although it
is an oversimplification of what is known about chemo-
mechanical coupling, the scheme provides the basic
details required for understanding the biochemical diver-
sity of myosins and will be referred to throughout the
kinetic discussions in this review. For more detailed
discussion of chemomechanical coupling and detailed
references concerning the actomyosin ATPase cycle,
we recommend the excellent textbook by Howard [5]
and the review by Geeves and Holmes [6].
The minimum mechanism needed to describe the myo-
sin ATPase cycle consists of at least six nucleotide-linked
biochemical reactions. These are described in detail in
Figure 1.
Intermediates are defined according to their affinities
for actin. The M.ATP and M.ADP.Pi states are ‘weak-
binding’ or ‘pre-force generating’ intermediates that
attach and detach (steps 2 and 4) from the actin filament
on a submicrosecond scale, have low (micromolar) actin
affinities, and do not bind stereospecifically to the actin
filament [6]. The weak-binding states do not bear force,
but allow myosin to ‘cock’ itself for the power-stroke
while detached from actin (step 3), preventing a reverse
power-stroke while attached to actin. The AM and
AM.ADP states are ‘strong-binding’ intermediates that
stereospecifically attach to actin filaments with high
(submicromolar) affinities. The strong-binding states
are the ‘force-bearing’ intermediates.
The force-generating power-stroke coincides with phos-
phate release at the transition from the weakly bound to
strongly bound states (step 5, Figure 1). Because it con-
trols the weak-to-strong transition, Pi release is an ideal
step for controlling myosin–actin attachment and moti-
lity. The transition is regulated by the calcium regulatory
system in striated muscle and by heavy-chain phosphor-
ylation of some myosin-I isoforms [7].
Enzymatic adaptations of myosin classesand isoformsTwo important parameters that influence the mechanical
and motile properties of myosins are, first, the lifetimes
(i.e. the duration of the biochemical states in units of
time) of the strongly bound intermediates (AM and
AMD), and, second, the duty ratio. The attachment
lifetimes and duty ratios can vary considerably between
myosin classes and between isoforms within a given class.
The duty ratio of a molecular motor is defined as the
fraction of the total ATPase cycle time that an individual
$ Supplementary data associated with this article can be found, in the
online version, at doi:10.1016/j.ceb.2003.11.011
www.sciencedirect.com Current Opinion in Cell Biology 2004, 16:61–67

motor is attached to its track [5]. For a population of
motors, the duty ratio is the steady-state fraction of
attached molecules. The attached motors can be in the
pre- [8–11] or post-power-stroke states. When discussing
myosins, the duty ratio usually refers to the fraction of
time spent in the strongly bound states. For example, low-
duty-ratio myosins spend a large proportion of time in the
M.ATP and M.ADP.Pi states, whereas high-duty-ratio
myosins occupy the AM and AMD states. In simple kinetic
terms, one would expect steps 4 or 5 (Figure 1) to be rate
limiting (i.e. the slowest step or ‘bottleneck’ in the ATPase
cycle) for low-duty-ratio myosins, and steps 1 or 6
(Figure 1) to be rate limiting for high-duty-ratio myosins
(see animation in supplementary material). The duty ratio
of characterized myosins depends on the rates into and out
of the strong-binding states, and therefore depends on the
actin and nucleotide concentrations [12,13�].
Myosin-I
Myosin-I isoforms are the single-headed, membrane-asso-
ciated members of the myosin superfamily found in most
eukaryotic cells. They play essential roles in membrane
dynamics, cytoskeletal structure, mechanical signal-trans-
duction and endosome processing [1]. The myosin-I
family can be divided into two major subclasses on the
basis of motor domain sequences (Figure 2a). Subclass 1
contains the ‘amoeboid’ or long-tailed myosin-I proteins.
Subclass 2 contains the ‘short-tailed’ myosins, including
the well-characterized myo1a (brush border myosin-I) and
the more widely expressed myo1b and myo1c.
All characterized myosin-I isoforms are low-duty-ratio
motors (i.e. they spend a small fraction of their catalytic
cycle strongly bound to actin), and the weak-binding
states have a very low actin affinity [14,15,16�]. There-
fore, small numbers of myosin-I motors cannot support
contraction or movements of cargo over long distances.
Comparative kinetic studies of myosin-I isoforms have
identified important differences between subclasses. The
rate constants of most characterized subclass-2 isoforms
are slower than subclass-1 isoforms [14,15,16�,17,18],
resulting in all biochemical intermediates having signifi-
cantly longer lifetimes.
By examining the lifetimes of the strong-binding states
(Figure 2b), we gain important insights into the physio-
logical function and regulation of myosin-I proteins. The
long lifetimes of the force-bearing intermediates of sub-
class-2 myosins are likely to be crucial for structural or
tension-maintenance roles [15,17]. For example, the
force-generating properties of these motors may function
Figure 1
AMAM.ADPAM.ATP
M.ATP
AM.ADP.Pi
M.ADP.Pi
A = Actin
M = Myosin
Weak-binding states
Strong-binding states
AM
(5)Powerstroke
ADPPi
Rate-limiting step for low-duty-ratio myosins
Rate-limiting step for high-duty-ratio myosins
ATP
(1)
(2)
(3)
(4)
(6)
Current Opinion in Cell Biology
The actomyosin ATPase cycle, identifying the weak- and strong-binding states of myosin. In the absence of ATP, myosin binds tightly to actin.
ATP binding (step 1) induces a conformational change in myosin that weakens its actin affinity and causes myosin to detach from actin (step 2).
A second conformational change allows hydrolysis of ATP to ADP and inorganic phosphate (Pi) (step 3); the hydrolysis products remain bound to
myosin. The M.ADP.Pi state rebinds to the actin filament (step 4) and a force-generating power-stroke accompanies phosphate release (step 5).
ADP is released (step 6) and the cycle repeats upon ATP binding. The actin-detached M and M.ADP states are omitted for clarity and to illustrate
that these states are not populated during enzymatic cycling in the presence of saturating actin and ATP.
62 Cell structure and dynamics
Current Opinion in Cell Biology 2004, 16:61–67 www.sciencedirect.com

in putting tension-sensitive calcium channels under ten-
sion [19��] and in defining the structure and organization
of cellular organelles [20]. The short lifetimes of the
strong-binding states of subclass-1 isoforms may favor
their playing a role in rapid contractile events where large
numbers of myosins are required, as in the rapid contrac-
tion of actin-rich membrane protrusions [21,22].
Myosin-II
Myosin-II isoforms are the major contractile proteins in
muscle and also play several crucial roles in non-muscle
contractility. Myosin-II molecules contain two motor
domains and assemble into bipolar filaments. Like the
myosin-I family, phylogenetic analysis of the motor
domains of the myosin-II family shows a division of
isoforms into distinct subclasses that cross phylogenetic
boundaries. The myosin-II family can be divided into
non-muscle cytoplasmic, cardiac muscle, smooth muscle,
and skeletal muscle subclasses, each with multiple iso-
forms [2,23].
All characterized muscle myosin-II isoforms have com-
parable unitary forces and displacements — a single
skeletal muscle myosin-II molecule generates the same
force and displacement as a single smooth muscle myo-
sin-II molecule [24]. However, the kinetic intermediate
lifetimes and duty ratios of muscle isoforms show a great
deal of variation that results in important mechanical
differences, accounting for their functional diversity.
For example, the duty ratio and the lifetime of the
strong-binding states of smooth muscle myosin are sig-
nificantly longer than those of skeletal muscle isoforms
[24,25]. In muscle tissue, these kinetic differences are
likely to be responsible for smooth muscle having slower
rates of contraction and producing higher forces than
skeletal muscle [24].
Recent investigations of non-muscle myosin-IIb demon-
strate that it is a relatively high-duty-ratio motor under
physiological actin and nucleotide concentrations (myosin
is attached to actin for �20–50% of its ATPase cycle time)
Figure 2
0
100
200
300
400
500
Subclass-1Myosin-I(Myo1e)
Subclass-2Myosin-I(Myo1b)
Myosin-V
Life
time
of s
tron
g-bi
ndin
g st
ates
(m
s)
Human Myo1aMouse Myo1aRat Myo1aChick Myo1aMouse Myo1bRat Myo1bHuman Myo1cMouse Myo1cDicty MyoEAcan Myo1aHuman Myo1eRat Myo1eAcan Myo1bDicty Myo1bAcan Myo1c
Sub
clas
s-2
Sub
clas
s-1
Subclass-1Myosin-I(Myo1e)
Subclass-2Myosin-I(Myo1b)
Myosin-V
Fraction of ATPasecycle spent inweak–bindingstates
Fraction of ATPasecycle spent instrong–bindingstates
Myosin-VI
(b)
(c)
(a)
Current Opinion in Cell Biology
Relationship between myosin duty ratio and lifetime of the strong-binding state. (a) Phylogenetic analysis of several members of the myosin-I
family showing subclass divisions. (b) Lifetimes of the AM.ADP strong-binding states of myosins -I, -V, and -VI [15,16�,37,43]. Notice that the lifetime
of the strong-binding state of myo1b (a low-duty-ratio motor) is significantly longer than that of processive myosin-V. (c) Pie charts showing the
relative amounts of time per ATPase cycle that myosin-I proteins (low-duty-ratio motors) and myosin-V (a high-duty-ratio motor) spend in the
weak- and strong-binding states in the presence of 50 mM actin.
Relating biochemistry and function in the myosin superfamily De La Cruz and Ostap 63
www.sciencedirect.com Current Opinion in Cell Biology 2004, 16:61–67

[13�,26�]. In this way it differs significantly from non-
muscle myosin-IIa [27] and muscle myosin isoforms,
which possess low duty ratios (myosin is attached to actin
for �10% of its ATPase cycle time). Thus, non-muscle
myosin-IIb appears to be adapted for cellular roles requir-
ing tension maintenance, whereas non-muscle myosin-IIa
is better suited to more rapid sliding. Kovacs et al. [27]
suggest that cellular myosin filaments may be composed
of multiple isoforms, which raises the possibility of
further kinetic ‘fine-tuning’ by the regulation of myosin
filament content.
When operating under high forces (as in a muscle under-
going an isometric contraction), low-duty-ratio myosin-II
isoforms may become high duty-ratio motors; in other
words, high forces may slow the rate at which intermedi-
ates leave the strong binding states [5,6]. However, the
effect of force on the kinetic and molecular mechanical
cycles is not understood and is the focus of intensive
study.
Myosin-V
Myosin-V transports vesicles along actin filament tracks
over long distances as a single, two-headed molecule [28–
31,32��,33��,34�,35�,36]. Detailed kinetic analysis of sin-
gle-headed myosin-V [37–40] demonstrate that ATP
binding (step 1, Figure 1), dissociation from actin (step
2, Figure 1), hydrolysis off actin (step 3, Figure 1) and Pi
release on actin (steps 4–5, Figure 1) are rapid (�200-
800 s�1) and essentially irreversible, whereas ADP release
(step 6, Figure 1) is slow (�16 s�1) and rate-limiting. As a
result, the predominant steady-state intermediate in the
presence of physiological ATP concentrations is the
strongly bound AM.ADP state, making myosin-V a
high-duty-ratio motor.
It is important to recognize that both the duty ratio and
attachment lifetimes reflect enzymatic adaptations
required for specific biological functions, and that a high
duty ratio does not necessarily indicate that the actin-
attached intermediates have long lifetimes (see the ani-
mation in supplementary material for an illustration of
this principle). For example, myosin-V has a much greater
duty ratio than subclass-2 myosin-I proteins (Figure 2c),
yet the lifetime of the strong binding states of myosin-V
are significantly shorter than those of the low-duty-ratio
myosin-I isoform (Figure 2b). Myosin-V is better suited to
rapid transport, whereas it appears that subclass-2 myosin-
I proteins are designed for slow transport in actin mesh-
works or for tension maintenance [15,17].
Myosin-VI
Myosin-VI is apparently a two-headed molecule that does
not associate into filaments and that may play a role in
endocytosis and membrane trafficking [41]. It was the first
myosin discovered to move toward the pointed end of
actin filaments [42,43]. Like myosin-V, myosin-VI is a
high-duty-ratio motor as a result of its slow rate-limiting
ADP release [44]. Additionally, ATP binding is slow and
weak, resulting in a population of nucleotide-free AM
(rigor) state intermediates under physiological conditions,
which further increases the duty ratio [44].
Myosin kinetics and processivityA high duty ratio is necessary for continuous movement
by myosin motors. Myosin-I and most characterized
myosin-II isoforms are low-duty-ratio motors and must
work in ensembles of many motors (which generates a
high effective duty ratio) to sustain continuous sliding
(Figure 3a). This requirement has been demonstrated invitro by examining sliding as a function of protein
density [45–47]. Locally high concentrations of myo-
sin-II are created in cells by the assembly of myosin into
filaments and by incorporation of the filaments into
actin-containing structures such as stress fibers or sar-
comeres. Myosin-I isoforms are restricted to membranes
and the actin-rich cell cortex, but it is not known if
accessory proteins play a role in concentration of these
motors.
Myosin-V from vertebrates [28–31,32��,33��,34�,35�,36],
but not yeast [48], and myosin-VI [49,50] are highly
processive, which is generally agreed to mean that an
individual molecule can take multiple steps, each
coupled to a cycle of ATP hydrolysis, along an actin
filament track before dissociating. Both heads are
required for processivity. Each motor of a processive
two-headed myosin (in which the motor domains are
not independently processive) must have a high duty
ratio (>0.5) to increase the probability that at least one
motor domain is always attached to actin, or the motor and
transported cargo will diffuse away from its track. In
addition, there must be some form of communication
between the motor domains: at a minimum to bias the
position of lead head attachment, but perhaps coordinat-
ing the catalytic cycles.
Many models have been proposed to account for the
motility of two-headed processive myosins [28–31,32��,34�,35�,36,51,52,53��,54,55]. Most implicate coordination
of the ATPase cycle kinetics of the two heads. Although
head–head coordination is not a requirement for proces-
sivity, it has been observed for double-headed myosin-VI
[44] and has been shown to be Ca2þ-dependent [56].
We favor an alternating-site ‘hand-over-hand’ model [44]
for processive stepping in the presence of saturating ATP
(Figure 3b). The model predicts that myosin dwells
predominantly in a state with the trailing head strongly
bound to actin in the AM.ADP state, and the leading head
in equilibrium between a detached and an ‘engaged’
ADP.Pi state. The engaged head is bound to actin but
has not undergone its power-stroke. The trailing head
impedes the leading head from binding actin strongly and
64 Cell structure and dynamics
Current Opinion in Cell Biology 2004, 16:61–67 www.sciencedirect.com
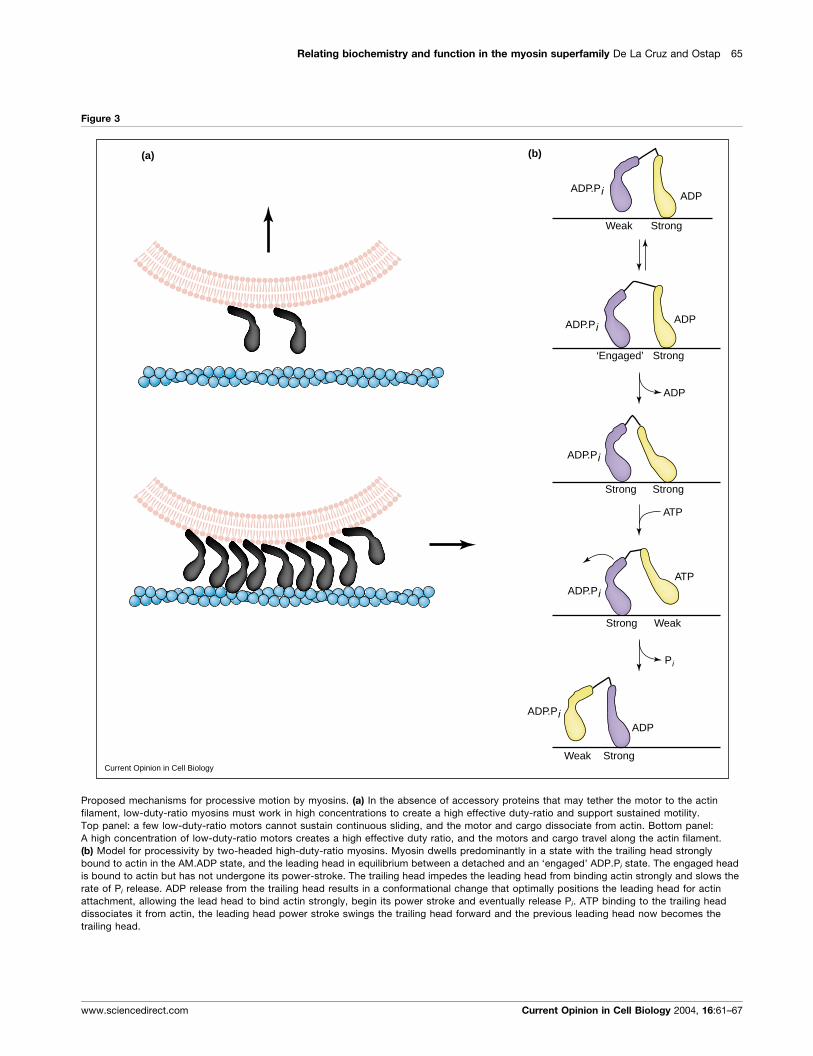
Figure 3
ADP.Pi ADP
ADP
ADP.Pi
ADP.Pi
ATP
ADP.Pi
Pi
ATP
ADP.PiADP
ADP
Weak Strong
WeakStrong
Strong Strong
‘Engaged’ Strong
Weak Strong
(a) (b)
Current Opinion in Cell Biology
Proposed mechanisms for processive motion by myosins. (a) In the absence of accessory proteins that may tether the motor to the actin
filament, low-duty-ratio myosins must work in high concentrations to create a high effective duty-ratio and support sustained motility.
Top panel: a few low-duty-ratio motors cannot sustain continuous sliding, and the motor and cargo dissociate from actin. Bottom panel:
A high concentration of low-duty-ratio motors creates a high effective duty ratio, and the motors and cargo travel along the actin filament.
(b) Model for processivity by two-headed high-duty-ratio myosins. Myosin dwells predominantly in a state with the trailing head stronglybound to actin in the AM.ADP state, and the leading head in equilibrium between a detached and an ‘engaged’ ADP.Pi state. The engaged head
is bound to actin but has not undergone its power-stroke. The trailing head impedes the leading head from binding actin strongly and slows the
rate of Pi release. ADP release from the trailing head results in a conformational change that optimally positions the leading head for actin
attachment, allowing the lead head to bind actin strongly, begin its power stroke and eventually release Pi. ATP binding to the trailing head
dissociates it from actin, the leading head power stroke swings the trailing head forward and the previous leading head now becomes the
trailing head.
Relating biochemistry and function in the myosin superfamily De La Cruz and Ostap 65
www.sciencedirect.com Current Opinion in Cell Biology 2004, 16:61–67

slows the rate of Pi release. ADP release from the trailing
head results in a conformational change that optimally
positions the leading head for actin attachment, allowing
the lead head to bind actin strongly, begin its power
stroke, and eventually release Pi. The conformational
change can be a rotation of the lever arm [31,33��,34�]or, with myosin VI, a more complex, larger-scale rearran-
gement that has yet to be determined [49,55]. At low ATP
concentrations (at which some structural experiments
have been performed), it has been proposed that the lead
head may release its products before the trailing head
dissociates [53��]. ATP binding to the trailing head dis-
sociates it from actin, whereupon the leading-head power
stroke swings the trailing head forward and the previous
leading head now becomes the trailing head. A defining
feature of this model is that the trailing head restricts the
lead head from progressing through the cycle.
Conclusions and future directionsThough the ATPase cycles of myosin motors are con-
served, the kinetics of the individual reactions defining
the cycle can vary dramatically and are optimized for
specific biological functions. It is generally believed that
the tail domains of myosin motors provide the source of
functional diversity. However, detailed kinetic analysis
demonstrates that modulation of the rate and equilibrium
constants defining the ATPase cycle can confer unique
and specific properties on a motor that allow it to accom-
plish a specific physiological task. Understanding the
kinetic mechanisms allows us to better define the poten-
tial cellular functions of the different myosin classes and
isoforms.
Future kinetic studies need to address the molecular
mechanisms of ATPase cycle regulation by associated
light chains [12,57–60], actin-binding proteins such as
tropomyosin, and post-translational modifications. It is
also critical to understand how external forces and loads
influence the rate constants of the ATPase cycle.
AcknowledgementsWe thank members of the De La Cruz and Ostap laboratories for helpfuldiscussions, and Lee Ligon and Chris Yengo for comments on themanuscript. EDLC is supported by grants from NSF (MCB-0216834)and a Scientist Development Grant from AHA. EMO is supported bygrants from NIH (GM57247) and an Established Investigator Awardfrom AHA.
References and recommended readingPapers of particular interest, published within the annual period ofreview, have been highlighted as:
� of special interest��of outstanding interest
1. Sokac AM, Bement WM: Regulation and expression of metazoanunconventional myosins. Int Rev Cytol 2000, 200:197-304.
2. Berg JS, Powell BC, Cheney RE: A millennial myosin census. MolBiol Cell 2001, 12:780-794.
3. Lymn RW, Taylor EW: Mechanism of adenosine triphosphatehydrolysis by actomyosin. Biochemistry 1971, 10:4617-4624.
4. Eisenberg E, Greene LE: The relation of muscle biochemistryto muscle physiology. Annu Rev Physiol 1980, 42:293-309.
5. Howard J: Mechanics of Motor Proteins and the Cytoskeleton.Sunderland, MA: Sinauer Assoc; 2001.
6. Geeves MA, Holmes KC: Structural mechanism of musclecontraction. Annu Rev Biochem 1999, 68:687-728.
7. Ostap EM, Lin T, Rosenfeld SS, Tang N: Mechanism of regulationof Acanthamoeba myosin-IC by heavy-chain phosphorylation.Biochemistry 2002, 41:12450-12456.
8. Okada Y, Hirokawa N: Mechanism of the single-headedprocessivity: diffusional anchoring between the K-loop ofkinesin and the C terminus of tubulin. Proc Natl Acad Sci U S A2000, 97:640-645.
9. Okada Y, Higuchi H, Hirokawa N: Processivity of the single-headed kinesin KIF1A through biased binding to tubulin.Nature 2003, 424:574-577.
10. Post PL, Tyska MJ, O’Connell CB, Johung K, Hayward A,Mooseker M: Myosin-IXb is a single-headed and processivemotor. J Biol Chem 2002, 277:11679-11683.
11. Inoue A, Saita J, Ikebe R, Ikebe M: Myosin-IXb is a single-headedminus-end-directed processive motor. Nat Cell Biol 2002,4:302-306.
12. De La Cruz EM, Wells AL, Sweeney HL, Ostap EM: Actin and lightchain isoform dependence of myosin V kinetics. Biochemistry2000, 39:14196-14202.
13.�
Wang F, Kovacs M, Hu A, Limouze J, Harvey EV, Sellers JR:Kinetic mechanism of non-muscle myosin IIB: functionaladaptations for tension generation and maintenance.J Biol Chem 2003, 278:27439-27448.
This paper and [26�] are the first to demonstrate that some myosin-IIisoforms are high-duty-ratio motors in solution.
14. Ostap EM, Pollard TD: Biochemical kinetic characterizationof the Acanthamoeba myosin-I ATPase. J Cell Biol 1996,132:1053-1060.
15. Coluccio LM, Geeves MA: Transient kinetic analysis of the130-kDa myosin I (MYR-1 gene product) from rat liver. A myosinI designed for maintenance of tension? J Biol Chem 1999,274:21575-21580.
16.�
El Mezgueldi M, Tang N, Rosenfeld SS, Ostap EM: The kineticmechanism of myo1e (human myosin-IC). J Biol Chem 2002,277:21514-21521.
The authors propose that the different myosin-I subclasses have differentbiochemical properties.
17. Jontes JD, Milligan RA, Pollard TD, Ostap EM: Kineticcharacterization of brush border myosin-I ATPase. Proc NatlAcad Sci U S A 1997, 94:14332-14337.
18. Veigel C, Coluccio LM, Jontes JD, Sparrow JC, Milligan RA,Molloy JE: The motor protein myosin-I produces its workingstroke in two steps. Nature 1999, 398:463-465.
19.��
Holt JR, Gillespie SK, Provance DW, Shah K, Shokat KM, Corey DP,Mercer JA, Gillespie PG: A chemical-genetic strategy implicatesmyosin-1c in adaptation in hair cells. Cell 2002, 108:371-381.
The investigators utilize a novel strategy to demonstrate that myosin-Iplays a role in the mechanical adaptation of hair cell signaling. This is oneof the first demonstrations of a molecular role for a myosin-I isoform.
20. Raposo G, Cordonnier MN, Tenza D, Menichi B, Durrbach A,Louvard D, Coudrier E: Association of myosin Ia with endosomesand lysosomes in mammalian cells. Mol Biol Cell 1999,10:1477-1494.
21. Stoffler HE, Ruppert C, Reinhard J, Bahler M: A novel mammalianmyosin I from rat with an SH3 domain localizes to Con-A-inducible, F-actin-rich structures at cell–cell contacts. J CellBiol 1995, 129:819-830.
22. Ostap EM, Maupin P, Doberstein SK, Baines IC, Korn ED,Pollard TD: Dynamic localization of myosin-I to endocyticstructures in Acanthamoeba. Cell Motil Cytoskeleton 2003,54:29-40.
23. Sellers JR: Myosins. Oxford: Oxford University Press; 1999.
66 Cell structure and dynamics
Current Opinion in Cell Biology 2004, 16:61–67 www.sciencedirect.com

24. Guilford WH, Dupuis DE, Kennedy G, Wu J, Patlak JB,Warshaw DM: Smooth muscle and skeletal muscle myosinsproduce similar unitary forces and displacements in the lasertrap. Biophys J 1997, 72:1006-1021.
25. Marston SB, Taylor EW: Comparison of the myosin andactomyosin ATPase mechanisms of the four types ofvertebrate muscles. J Mol Biol 1980, 139:573-600.
26.�
Rosenfeld SS, Xing J, Cheng LQ, Sweeney HL: Myosin IIb isunconventionally conventional. J Biol Chem 2003,278:27449-27455.
See annotation to [13�].
27. Kovacs M, Wang F, Hu A, Zhang Y, Sellers JR: Functionaldivergence of human cytoplasmic myosin II: kineticcharacterization of the non-muscle IIA isoform. J Biol Chem2003, 278:38132-38140.
28. Mehta AD, Rock RS, Rief M, Spudich JA, Mooseker MS,Cheney RE: Myosin-V is a processive actin-based motor.Nature 1999, 400:590-593.
29. Sakamoto T, Amitani I, Yokota E, Ando T: Direct observation ofprocessive movement by individual myosin V molecules.Biochem Biophys Res Commun 2000, 272:586-590.
30. Rief M, Rock RS, Mehta AD, Mooseker MS, Cheney RE,Spudich JA: Myosin-V stepping kinetics: a molecular modelfor processivity. Proc Natl Acad Sci U S A 2000, 97:9482-9486.
31. Moore JR, Krementsova EB, Trybus KM, Warshaw DM: Myosin Vexhibits a high duty cycle and large unitary displacement.J Cell Biol 2001, 155:625-635.
32.��
Veigel C, Wang F, Bartoo ML, Sellers JR, Molloy JE: The gated gaitof the processive molecular motor, myosin V. Nat Cell Biol 2002,4:59-65.
The authors examine the mechanism of myosin-V processivity, withemphasis on understanding how the biochemical and mechanical statesof the two heads in a molecule are coupled.
33.��
Forkey JN, Quinlan ME, Shaw MA, Corrie JE, Goldman YE:Three-dimensional structural dynamics of myosin V by single-molecule fluorescence polarization. Nature 2003, 422:399-404.
This technical tour-de-force demonstrates directly the lever arm rotationof a single myosin-V molecule as it processes along an actin filament. Theresults support a hand-over-hand model for myosin-V stepping.
34.�
Purcell TJ, Morris C, Spudich JA, Sweeney HL: Role of the leverarm in the processive stepping of myosin V. Proc Natl AcadSci U S A 2002, 99:14159-14164.
With [35�], this paper demonstrates the importance of the lever arm lengthin myosin-V step size and processivity.
35.�
Sakamoto T, Wang F, Schmitz S, Xu Y, Xu Q, Molloy JE, Veigel C,Sellers JR: Neck length and processivity of myosin V. J BiolChem 2003, 278:29201-29207.
See annotation to [34�].
36. Yildiz A, Forkey JN, McKinney SA, Ha T, Goldman YE, Selvin PR:Myosin V walks hand-over-hand: single fluorophore imagingwith 1.5-nm localization. Science 2003, 300:2061-2065.
37. De La Cruz EM, Wells AL, Rosenfeld SS, Ostap EM, Sweeney HL:The kinetic mechanism of myosin V. Proc Natl Acad Sci U S A1999, 96:13726-13731.
38. Trybus KM, Krementsova E, Freyzon Y: Kinetic characterizationof a monomeric unconventional myosin V construct. J BiolChem 1999, 274:27448-27456.
39. Wang F, Chen L, Arcucci O, Harvey EV, Bowers B, Xu Y,Hammer JA III, Sellers JR: Effect of ADP and ionic strength on thekinetic and motile properties of recombinant mouse myosin V.J Biol Chem 2000, 275:4329-4335.
40. Yengo CM, De la Cruz EM, Safer D, Ostap EM, Sweeney HL:Kinetic characterization of the weak binding states of myosinV. Biochemistry 2002, 41:8508-8517.
41. Buss F, Luzio JP, Kendrick-Jones J: Myosin-VI, and actin motorfor membrane traffic and cell migration. Traffic 2002, 3:851-858.
42. Wells AL, Lin AW, Chen LQ, Safer D, Cain SM, Hasson T,Carragher BO, Milligan RA, Sweeney HL: Myosin VI is anactin-based motor that moves backwards. Nature 1999,401:505-508.
43. Homma K, Yoshimura M, Saito J, Ikebe R, Ikebe M: The core of themotor domain determines the direction of myosin movement.Nature 2001, 412:831-834.
44. De La Cruz EM, Ostap EM, Sweeney HL: Kinetic mechanism andregulation of myosin VI. J Biol Chem 2001, 276:32373-32381.
45. Uyeda TQ, Kron SJ, Spudich JA: Myosin step size. Estimationfrom slow sliding movement of actin over low densities ofheavy meromyosin. J Mol Biol 1990, 214:699-710.
46. Harada Y, Sakurada K, Aoki T, Thomas DD, Yanagida T:Mechanochemical coupling in actomyosin energy transductionstudied by in vitro movement assay. J Mol Biol 1990, 216:49-68.
47. Finer JT, Simmons RM, Spudich JA: Single myosin moleculemechanics: piconewton forces and nanometre steps. Nature1994, 368:113-119.
48. Reck-Peterson SL, Tyska MJ, Novick PJ, Mooseker MS: The yeastclass V myosins, Myo2p and Myo4p, are nonprocessive actin-based motors. J Cell Biol 2001, 153:1121-1126.
49. Rock RS, Rice SE, Wells AL, Purcell TJ, Spudich JA, Sweeney HL:Myosin VI is a processive motor with a large step size.Proc Natl Acad Sci U S A 2001, 98:13655-13659.
50. Nishikawa S, Homma K, Komori Y, Iwaki M, Wazawa T,Hikikoshi Iwane A, Saito J, Ikebe R, Katayama E, Yanagida T, IkebeM: Class VI myosin moves processively along actin filamentsbackward with large steps. Biochem Biophys Res Commun 2002,290:311-317.
51. Walker ML, Burgess SA, Sellers JR, Wang F, Hammer JA III,Trinick J, Knight PJ: Two-headed binding of a processive myosinto F-actin. Nature 2000, 405:804-807.
52. Kolomeisky AB, Fisher ME: A simple kinetic model describes theprocessivity of Myosin-V. Biophys J 2003, 84:1642-1650.
53.��
Burgess S, Walker M, Wang F, Sellers JR, White HD, Knight PJ,Trinick J: The prepower stroke conformation of myosin V.J Cell Biol 2002, 159:983-991.
This paper uses single particle image processing to investigate thestructure of the myosin-V motor in the presence and absence of nucleo-tides. The authors conclude that stepping is produced by changes in thelever arm angle brought about by converter movement plus thermalenergy.
54. Ali MY, Uemura S, Adachi K, Itoh H, Kinosita K Jr, Ishiwata S:Myosin V is a left-handed spiral motor on the right-handedactin helix. Nat Struct Biol 2002, 9:464-467.
55. Tanaka H, Homma K, Iwane AH, Katayama E, Ikebe R, Saito J,Yanagida T, Ikebe M: The motor domain determines the largestep of myosin-V. Nature 2002, 415:192-195.
56. Morris CA, Wells AL, Yang Z, Chen LQ, Baldacchino CV,Sweeney HL: Calcium functionally uncouples the heads ofmyosin VI. J Biol Chem 2003, 278:23324-23330.
57. Homma K, Saito J, Ikebe R, Ikebe M: Ca2R-dependent regulationof the motor activity of myosin V. J Biol Chem 2000,275:34766-34771.
58. Yoshimura M, Homma K, Saito J, Inoue A, Ikebe R, Ikebe M:Dual regulation of mammalian myosin VI motor function.J Biol Chem 2001, 276:39600-39607.
59. Martin SR, Bayley PM: Regulatory implications of a novel modeof interaction of calmodulin with a double IQ-motif targetsequence from murine dilute myosin V. Protein Sci 2002,11:2909-2923.
60. Terrak M, Wu G, Stafford WF, Lu RC, Dominguez R: Two distinctmyosin light chain structures are induced by specific variationswithin the bound IQ motifs — functional implications.EMBO J 2003, 22:362-371.
Relating biochemistry and function in the myosin superfamily De La Cruz and Ostap 67
www.sciencedirect.com Current Opinion in Cell Biology 2004, 16:61–67




















