
Prosorchis palinurichthi (Digenea, Sclerodistomidae): Ultrastructure of the mature spermatozoon
6
Zoologischer Anzeiger 252 (2013) 404–409 Contents lists available at SciVerse ScienceDirect Zoologischer Anzeiger journal homepage: www.elsevier.de/jcz Prosorchis palinurichthi Kurochkin, Parukhin & Korotaeva, 1971 (Digenea, Sclerodistomidae): Ultrastructure of the mature spermatozoon Papa Ibnou Ndiaye a , Yann Quilichini b,∗ , Aminata Sène a , Rodney A. Bray c , Cheikh Tidiane Bâ a , Bernard Marchand b a Laboratory of Evolutionary Biology, Ecology and Management of Ecosystems, Faculty of Sciences and Techniques, Cheikh Anta Diop University of Dakar, BP 5055, Dakar, Senegal b CNRS UMR 6134, University of Corsica, Laboratory “Parasites and Mediterranean Ecosystems”, 20250 Corte, Corsica, France c Department of Zoology, Natural History Museum, Cromwell Road, London SW7 5BD, UK article info Article history: Received 24 August 2012 Received in revised form 7 November 2012 Accepted 7 November 2012 Available online 20 December 2012 Corresponding Editor: Martin V. Sørensen. Keywords: Platyhelminthes Trepaxonemata Hemiuroidea TEM abstract This study presents the ultrastructure of the mature spermatozoon of Prosorchis palinurichthi (Digenea: Hemiuroidea: Sclerodistomidae), parasite of the digestive tract of the teleost fish Schedophilus velaini (Sauvage, 1879) (Centrolophidae) captured in the Atlantic Ocean, near Dakar (Senegal). This is a first report of P. palinurichthi from Senegal and from this host species. The mature spermatozoon of P. palinurichthi is filiform, tapered at both ends and exhibits the general pattern described in the hemiuroids: two axonemes of the 9 + ‘1’ pattern of trepaxonematans, a nucleus, a mitochondrion, external ornamentation of the plasma membrane in the anterior region of the spermatozoon and parallel cortical microtubules disposed in one side of the spermatozoon except in the anterior spermatozoon extremity and in the mitochondrial region. However, the particularities of the spermatozoon of P. palinurichthi are the presence of cortical microtubules at the anterior extremity, a great number of cortical microtubules (up to 25) in the region II of the spermatozoon, an elongate nucleus longer than the mitochondrion and the absence of spinelike bodies. Our results are discussed in context with those of previous studies in the Digenea in general and particularly in the Hemiuroidea. © 2012 Elsevier GmbH. All rights reserved. 1. Introduction The family Sclerodistomidae is a small group of trematodes, generally parasites of marine teleosts, with only four subfam- ilies (Sclerodistominae, Kenmackenziinae, Prosogonotrematinae and Prosorchiinae) and five genera (Gibson, 2002). This family is still subject to multiple controversies (Dollfus, 1932; Yamaguti, 1934, 1958, 1971; Vigueras, 1940; Skrjabin and Guschanskaja, 1956; Gibson and Bray, 1977, 1979; Gibson, 1983). Ultrastructural data of spermatozoa have produced many use- ful characters for phylogenetic purposes in Platyhelminthes (Euzet et al., 1981; Jamieson and Daddow, 1982; Justine, 1991, 1995, 1998, 2001; Bâ and Marchand, 1995; Hoberg et al., 1997, 1999, 2001; Levron et al., 2010). To our knowledge, there are no ultrastructural data of the mature spermatozoon of a member of the Sclerodisto- midae. Only spermiogenesis was studied in Prosorchis ghanensis by Justine (1995). Here we present the first ultrastructural study of the mature spermatozoon of Prosorchis palinurichthi, a species belong- ing to Sclerodistomidae. P. palinurichthi was previously described ∗ Corresponding author. Tel.: +33 495 450 006; fax: +33 495 450 045. E-mail address: [email protected] (Y. Quilichini). as a parasite of black butterfish Hyperoglyphe pringlei (Centrolophi- dae) from the South Atlantic. Thus, our finding of P. palinurichthi in the centrolophid violet warehou Schedophilus velaini in Senegal is a new host and geographic locality record for this parasite. 2. Materials and methods Adult specimens of P. palinurichthi (Kurochkin et al., 1971) were collected from the stomach of a teleost Centrolophidae S. velaini (Sauvage, 1879) caught in the Atlantic Ocean, near Dakar (Senegal). Live worms were rinsed with a 0.9% NaCl solution and fixed in cold (4 ◦ C) 2.5% glutaraldehyde in a 0.1 M sodium cacodylate buffer at pH 7.2, rinsed in 0.1 M sodium cacodylate buffer at pH 7.2, post-fixed in cold (4 ◦ C) 1% osmium tetroxide in the same buffer for 1 h, rinsed in a 0.1 M sodium cacodylate buffer at pH 7.2, dehydrated in ethanol and propylene oxide, embedded in Spurr’s resin and polymerized at 60 ◦ C for 24 h. Ultrathin sections (60–90 nm thick) were obtained using an Ultramicrotome (Power tome PC, RMC Boeckeler ® ) with diamond knife. Sections placed on copper grids were double-stained with uranyl acetate and lead citrate. Sections placed on gold grids were stained with periodic acid, thiocarbohydrazide and silver pro- teinate to reveal the presence of glycogen (Thiéry, 1967). 0044-5231/$ – see front matter © 2012 Elsevier GmbH. All rights reserved. http://dx.doi.org/10.1016/j.jcz.2012.11.001
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Prosorchis palinurichthi (Digenea, Sclerodistomidae): Ultrastructure of the mature spermatozoon

PS
PBa
b
c
a
ARRAAC
KPTHT
1
gias11
fe2LdmJmi
0h
Zoologischer Anzeiger 252 (2013) 404–409
Contents lists available at SciVerse ScienceDirect
Zoologischer Anzeiger
journa l homepage: www.e lsev ier .de / jcz
rosorchis palinurichthi Kurochkin, Parukhin & Korotaeva, 1971 (Digenea,clerodistomidae): Ultrastructure of the mature spermatozoon
apa Ibnou Ndiayea, Yann Quilichinib,∗, Aminata Sènea, Rodney A. Brayc, Cheikh Tidiane Bâa,ernard Marchandb
Laboratory of Evolutionary Biology, Ecology and Management of Ecosystems, Faculty of Sciences and Techniques, Cheikh Anta Diop University of Dakar, BP 5055, Dakar, SenegalCNRS UMR 6134, University of Corsica, Laboratory “Parasites and Mediterranean Ecosystems”, 20250 Corte, Corsica, FranceDepartment of Zoology, Natural History Museum, Cromwell Road, London SW7 5BD, UK
r t i c l e i n f o
rticle history:eceived 24 August 2012eceived in revised form 7 November 2012ccepted 7 November 2012vailable online 20 December 2012orresponding Editor: Martin V. Sørensen.
eywords:
a b s t r a c t
This study presents the ultrastructure of the mature spermatozoon of Prosorchis palinurichthi (Digenea:Hemiuroidea: Sclerodistomidae), parasite of the digestive tract of the teleost fish Schedophilus velaini(Sauvage, 1879) (Centrolophidae) captured in the Atlantic Ocean, near Dakar (Senegal). This is a first reportof P. palinurichthi from Senegal and from this host species. The mature spermatozoon of P. palinurichthi isfiliform, tapered at both ends and exhibits the general pattern described in the hemiuroids: two axonemesof the 9 + ‘1’ pattern of trepaxonematans, a nucleus, a mitochondrion, external ornamentation of theplasma membrane in the anterior region of the spermatozoon and parallel cortical microtubules disposed
latyhelminthesrepaxonemataemiuroideaEM
in one side of the spermatozoon except in the anterior spermatozoon extremity and in the mitochondrialregion. However, the particularities of the spermatozoon of P. palinurichthi are the presence of corticalmicrotubules at the anterior extremity, a great number of cortical microtubules (up to 25) in the regionII of the spermatozoon, an elongate nucleus longer than the mitochondrion and the absence of spinelikebodies. Our results are discussed in context with those of previous studies in the Digenea in general andparticularly in the Hemiuroidea.
. Introduction
The family Sclerodistomidae is a small group of trematodes,enerally parasites of marine teleosts, with only four subfam-lies (Sclerodistominae, Kenmackenziinae, Prosogonotrematinaend Prosorchiinae) and five genera (Gibson, 2002). This family istill subject to multiple controversies (Dollfus, 1932; Yamaguti,934, 1958, 1971; Vigueras, 1940; Skrjabin and Guschanskaja,956; Gibson and Bray, 1977, 1979; Gibson, 1983).
Ultrastructural data of spermatozoa have produced many use-ul characters for phylogenetic purposes in Platyhelminthes (Euzett al., 1981; Jamieson and Daddow, 1982; Justine, 1991, 1995, 1998,001; Bâ and Marchand, 1995; Hoberg et al., 1997, 1999, 2001;evron et al., 2010). To our knowledge, there are no ultrastructuralata of the mature spermatozoon of a member of the Sclerodisto-idae. Only spermiogenesis was studied in Prosorchis ghanensis by
ustine (1995). Here we present the first ultrastructural study of theature spermatozoon of Prosorchis palinurichthi, a species belong-
ng to Sclerodistomidae. P. palinurichthi was previously described
∗ Corresponding author. Tel.: +33 495 450 006; fax: +33 495 450 045.E-mail address: [email protected] (Y. Quilichini).
044-5231/$ – see front matter © 2012 Elsevier GmbH. All rights reserved.ttp://dx.doi.org/10.1016/j.jcz.2012.11.001
© 2012 Elsevier GmbH. All rights reserved.
as a parasite of black butterfish Hyperoglyphe pringlei (Centrolophi-dae) from the South Atlantic. Thus, our finding of P. palinurichthi inthe centrolophid violet warehou Schedophilus velaini in Senegal isa new host and geographic locality record for this parasite.
2. Materials and methods
Adult specimens of P. palinurichthi (Kurochkin et al., 1971) werecollected from the stomach of a teleost Centrolophidae S. velaini(Sauvage, 1879) caught in the Atlantic Ocean, near Dakar (Senegal).Live worms were rinsed with a 0.9% NaCl solution and fixed in cold(4 ◦C) 2.5% glutaraldehyde in a 0.1 M sodium cacodylate buffer at pH7.2, rinsed in 0.1 M sodium cacodylate buffer at pH 7.2, post-fixed incold (4 ◦C) 1% osmium tetroxide in the same buffer for 1 h, rinsed ina 0.1 M sodium cacodylate buffer at pH 7.2, dehydrated in ethanoland propylene oxide, embedded in Spurr’s resin and polymerizedat 60 ◦C for 24 h.
Ultrathin sections (60–90 nm thick) were obtained using anUltramicrotome (Power tome PC, RMC Boeckeler®) with diamond
knife. Sections placed on copper grids were double-stained withuranyl acetate and lead citrate. Sections placed on gold grids werestained with periodic acid, thiocarbohydrazide and silver pro-teinate to reveal the presence of glycogen (Thiéry, 1967).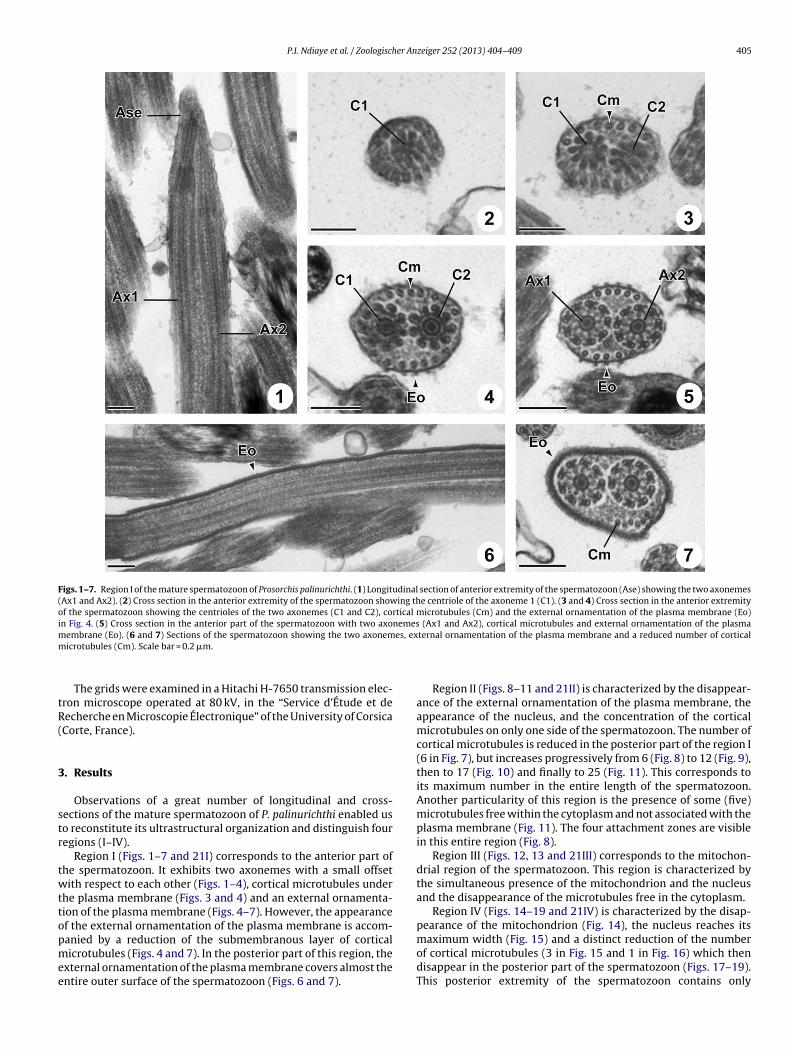
P.I. Ndiaye et al. / Zoologischer Anzeiger 252 (2013) 404–409 405
Figs. 1–7. Region I of the mature spermatozoon of Prosorchis palinurichthi. (1) Longitudinal section of anterior extremity of the spermatozoon (Ase) showing the two axonemes(Ax1 and Ax2). (2) Cross section in the anterior extremity of the spermatozoon showing the centriole of the axoneme 1 (C1). (3 and 4) Cross section in the anterior extremityof the spermatozoon showing the centrioles of the two axonemes (C1 and C2), cortical microtubules (Cm) and the external ornamentation of the plasma membrane (Eo)in Fig. 4. (5) Cross section in the anterior part of the spermatozoon with two axonemes (Ax1 and Ax2), cortical microtubules and external ornamentation of the plasmam es, exm
tR(
3
str
twttopmee
embrane (Eo). (6 and 7) Sections of the spermatozoon showing the two axonemicrotubules (Cm). Scale bar = 0.2 �m.
The grids were examined in a Hitachi H-7650 transmission elec-ron microscope operated at 80 kV, in the “Service d’Étude et deecherche en Microscopie Électronique” of the University of CorsicaCorte, France).
. Results
Observations of a great number of longitudinal and cross-ections of the mature spermatozoon of P. palinurichthi enabled uso reconstitute its ultrastructural organization and distinguish fouregions (I–IV).
Region I (Figs. 1–7 and 21I) corresponds to the anterior part ofhe spermatozoon. It exhibits two axonemes with a small offsetith respect to each other (Figs. 1–4), cortical microtubules under
he plasma membrane (Figs. 3 and 4) and an external ornamenta-ion of the plasma membrane (Figs. 4–7). However, the appearancef the external ornamentation of the plasma membrane is accom-
anied by a reduction of the submembranous layer of corticalicrotubules (Figs. 4 and 7). In the posterior part of this region, thexternal ornamentation of the plasma membrane covers almost thentire outer surface of the spermatozoon (Figs. 6 and 7).
ternal ornamentation of the plasma membrane and a reduced number of cortical
Region II (Figs. 8–11 and 21II) is characterized by the disappear-ance of the external ornamentation of the plasma membrane, theappearance of the nucleus, and the concentration of the corticalmicrotubules on only one side of the spermatozoon. The number ofcortical microtubules is reduced in the posterior part of the region I(6 in Fig. 7), but increases progressively from 6 (Fig. 8) to 12 (Fig. 9),then to 17 (Fig. 10) and finally to 25 (Fig. 11). This corresponds toits maximum number in the entire length of the spermatozoon.Another particularity of this region is the presence of some (five)microtubules free within the cytoplasm and not associated with theplasma membrane (Fig. 11). The four attachment zones are visiblein this entire region (Fig. 8).
Region III (Figs. 12, 13 and 21III) corresponds to the mitochon-drial region of the spermatozoon. This region is characterized bythe simultaneous presence of the mitochondrion and the nucleusand the disappearance of the microtubules free in the cytoplasm.
Region IV (Figs. 14–19 and 21IV) is characterized by the disap-pearance of the mitochondrion (Fig. 14), the nucleus reaches its
maximum width (Fig. 15) and a distinct reduction of the numberof cortical microtubules (3 in Fig. 15 and 1 in Fig. 16) which thendisappear in the posterior part of the spermatozoon (Figs. 17–19).This posterior extremity of the spermatozoon contains only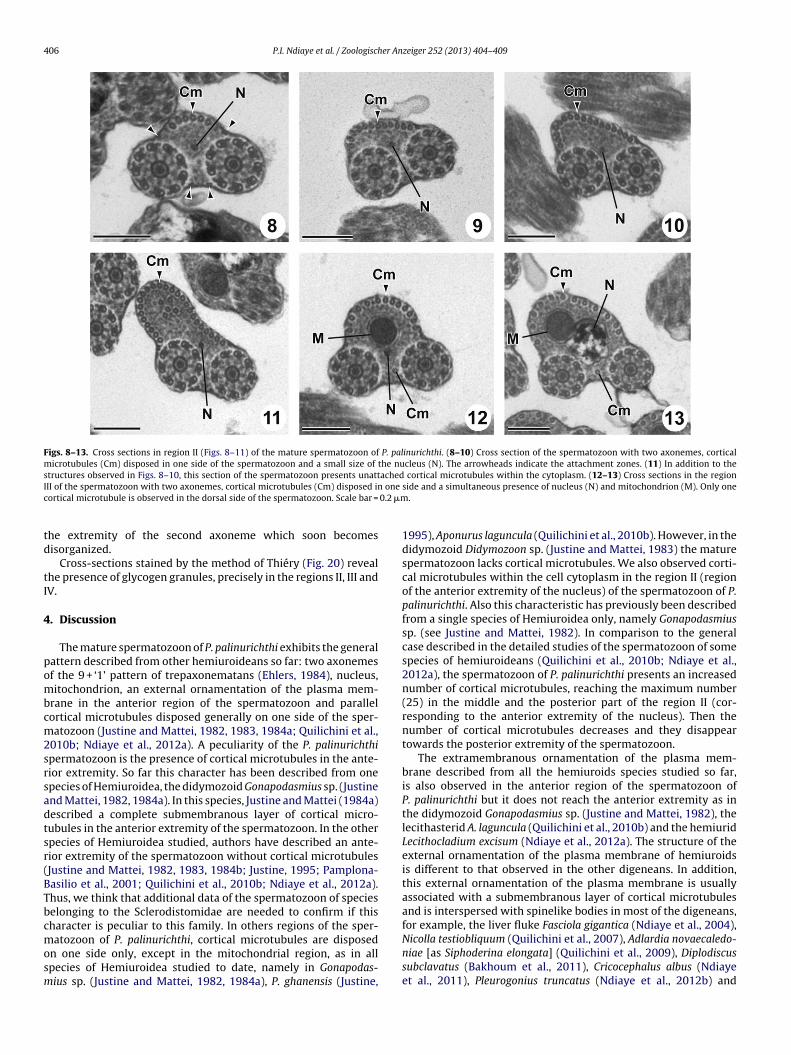
406 P.I. Ndiaye et al. / Zoologischer Anzeiger 252 (2013) 404–409
Figs. 8–13. Cross sections in region II (Figs. 8–11) of the mature spermatozoon of P. palinurichthi. (8–10) Cross section of the spermatozoon with two axonemes, corticalmicrotubules (Cm) disposed in one side of the spermatozoon and a small size of the nucleus (N). The arrowheads indicate the attachment zones. (11) In addition to thestructures observed in Figs. 8–10, this section of the spermatozoon presents unattached cortical microtubules within the cytoplasm. (12–13) Cross sections in the regionI onec 0.2 �m
td
tI
4
pombcm2srsadtsr(BTbcmosm
II of the spermatozoon with two axonemes, cortical microtubules (Cm) disposed inortical microtubule is observed in the dorsal side of the spermatozoon. Scale bar =
he extremity of the second axoneme which soon becomesisorganized.
Cross-sections stained by the method of Thiéry (Fig. 20) revealhe presence of glycogen granules, precisely in the regions II, III andV.
. Discussion
The mature spermatozoon of P. palinurichthi exhibits the generalattern described from other hemiuroideans so far: two axonemesf the 9 + ‘1’ pattern of trepaxonematans (Ehlers, 1984), nucleus,itochondrion, an external ornamentation of the plasma mem-
rane in the anterior region of the spermatozoon and parallelortical microtubules disposed generally on one side of the sper-atozoon (Justine and Mattei, 1982, 1983, 1984a; Quilichini et al.,
010b; Ndiaye et al., 2012a). A peculiarity of the P. palinurichthipermatozoon is the presence of cortical microtubules in the ante-ior extremity. So far this character has been described from onepecies of Hemiuroidea, the didymozoid Gonapodasmius sp. (Justinend Mattei, 1982, 1984a). In this species, Justine and Mattei (1984a)escribed a complete submembranous layer of cortical micro-ubules in the anterior extremity of the spermatozoon. In the otherpecies of Hemiuroidea studied, authors have described an ante-ior extremity of the spermatozoon without cortical microtubulesJustine and Mattei, 1982, 1983, 1984b; Justine, 1995; Pamplona-asilio et al., 2001; Quilichini et al., 2010b; Ndiaye et al., 2012a).hus, we think that additional data of the spermatozoon of specieselonging to the Sclerodistomidae are needed to confirm if thisharacter is peculiar to this family. In others regions of the sper-
atozoon of P. palinurichthi, cortical microtubules are disposedn one side only, except in the mitochondrial region, as in allpecies of Hemiuroidea studied to date, namely in Gonapodas-ius sp. (Justine and Mattei, 1982, 1984a), P. ghanensis (Justine,
side and a simultaneous presence of nucleus (N) and mitochondrion (M). Only one.
1995), Aponurus laguncula (Quilichini et al., 2010b). However, in thedidymozoid Didymozoon sp. (Justine and Mattei, 1983) the maturespermatozoon lacks cortical microtubules. We also observed corti-cal microtubules within the cell cytoplasm in the region II (regionof the anterior extremity of the nucleus) of the spermatozoon of P.palinurichthi. Also this characteristic has previously been describedfrom a single species of Hemiuroidea only, namely Gonapodasmiussp. (see Justine and Mattei, 1982). In comparison to the generalcase described in the detailed studies of the spermatozoon of somespecies of hemiuroideans (Quilichini et al., 2010b; Ndiaye et al.,2012a), the spermatozoon of P. palinurichthi presents an increasednumber of cortical microtubules, reaching the maximum number(25) in the middle and the posterior part of the region II (cor-responding to the anterior extremity of the nucleus). Then thenumber of cortical microtubules decreases and they disappeartowards the posterior extremity of the spermatozoon.
The extramembranous ornamentation of the plasma mem-brane described from all the hemiuroids species studied so far,is also observed in the anterior region of the spermatozoon ofP. palinurichthi but it does not reach the anterior extremity as inthe didymozoid Gonapodasmius sp. (Justine and Mattei, 1982), thelecithasterid A. laguncula (Quilichini et al., 2010b) and the hemiuridLecithocladium excisum (Ndiaye et al., 2012a). The structure of theexternal ornamentation of the plasma membrane of hemiuroidsis different to that observed in the other digeneans. In addition,this external ornamentation of the plasma membrane is usuallyassociated with a submembranous layer of cortical microtubulesand is interspersed with spinelike bodies in most of the digeneans,for example, the liver fluke Fasciola gigantica (Ndiaye et al., 2004),
Nicolla testiobliquum (Quilichini et al., 2007), Adlardia novaecaledo-niae [as Siphoderina elongata] (Quilichini et al., 2009), Diplodiscussubclavatus (Bakhoum et al., 2011), Cricocephalus albus (Ndiayeet al., 2011), Pleurogonius truncatus (Ndiaye et al., 2012b) and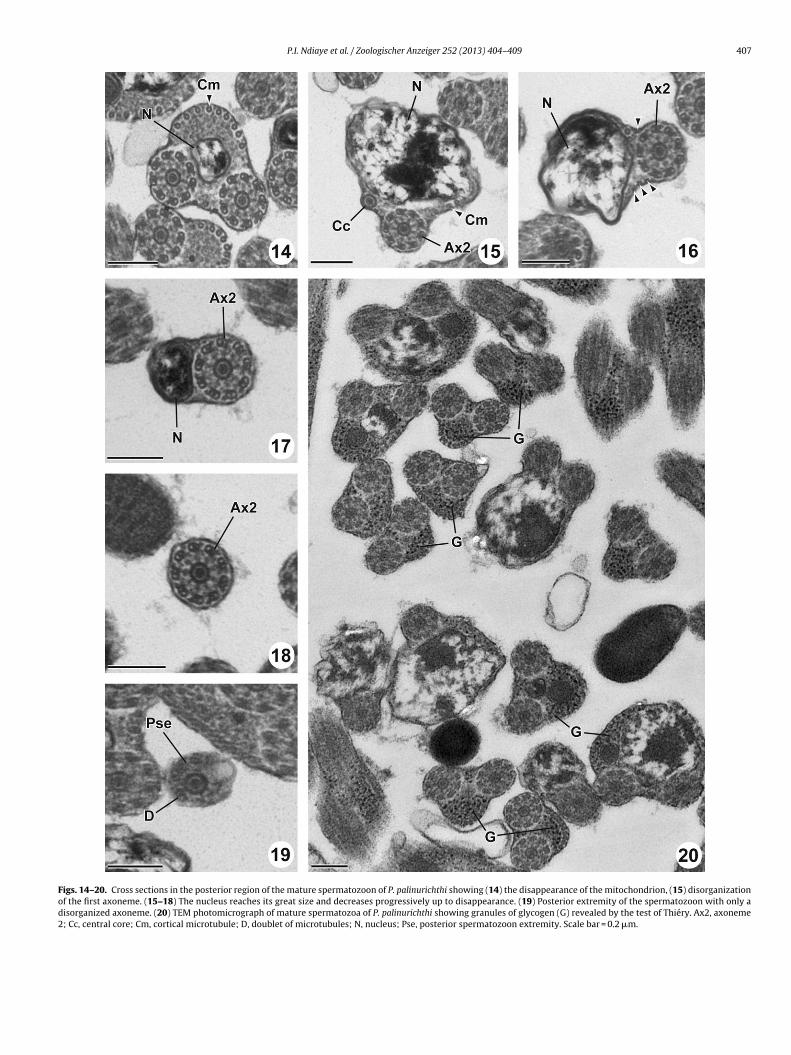
P.I. Ndiaye et al. / Zoologischer Anzeiger 252 (2013) 404–409 407
Figs. 14–20. Cross sections in the posterior region of the mature spermatozoon of P. palinurichthi showing (14) the disappearance of the mitochondrion, (15) disorganizationof the first axoneme. (15–18) The nucleus reaches its great size and decreases progressively up to disappearance. (19) Posterior extremity of the spermatozoon with only adisorganized axoneme. (20) TEM photomicrograph of mature spermatozoa of P. palinurichthi showing granules of glycogen (G) revealed by the test of Thiéry. Ax2, axoneme2; Cc, central core; Cm, cortical microtubule; D, doublet of microtubules; N, nucleus; Pse, posterior spermatozoon extremity. Scale bar = 0.2 �m.
408 P.I. Ndiaye et al. / Zoologischer Anzeiger 252 (2013) 404–409
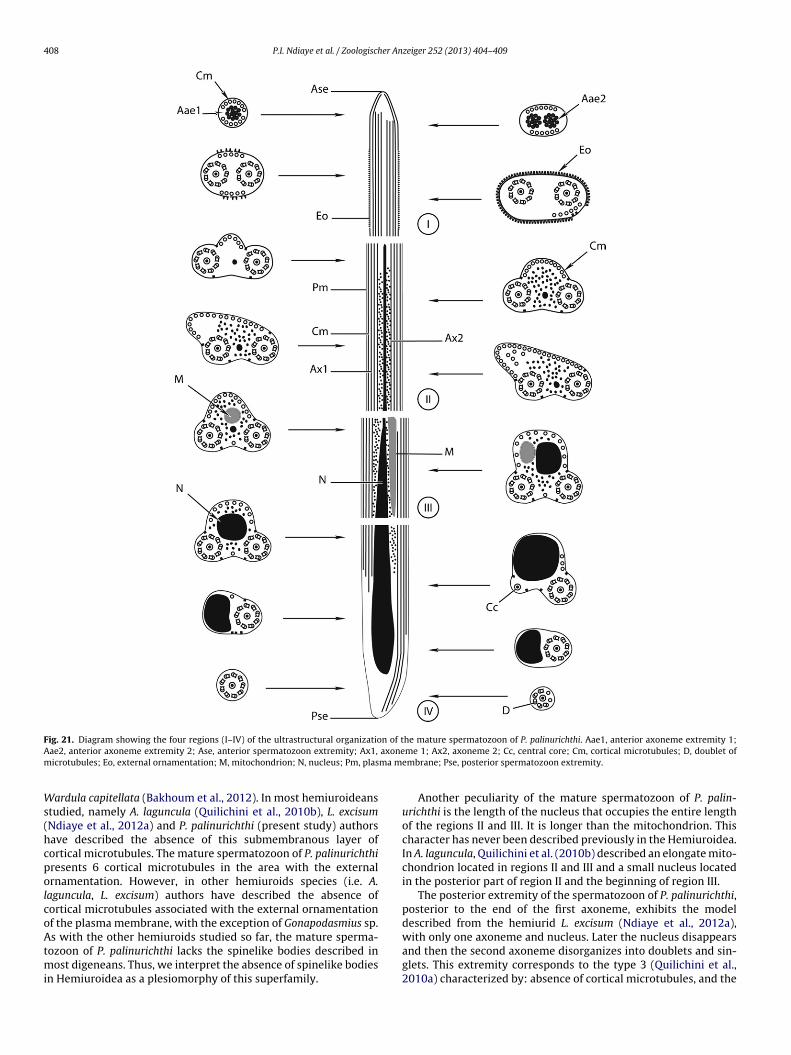
F n of tA axonem ma m
Ws(hcpolcoAtmi
ig. 21. Diagram showing the four regions (I–IV) of the ultrastructural organizatioae2, anterior axoneme extremity 2; Ase, anterior spermatozoon extremity; Ax1,icrotubules; Eo, external ornamentation; M, mitochondrion; N, nucleus; Pm, plas
ardula capitellata (Bakhoum et al., 2012). In most hemiuroideanstudied, namely A. laguncula (Quilichini et al., 2010b), L. excisumNdiaye et al., 2012a) and P. palinurichthi (present study) authorsave described the absence of this submembranous layer ofortical microtubules. The mature spermatozoon of P. palinurichthiresents 6 cortical microtubules in the area with the externalrnamentation. However, in other hemiuroids species (i.e. A.
aguncula, L. excisum) authors have described the absence ofortical microtubules associated with the external ornamentationf the plasma membrane, with the exception of Gonapodasmius sp.
s with the other hemiuroids studied so far, the mature sperma-ozoon of P. palinurichthi lacks the spinelike bodies described inost digeneans. Thus, we interpret the absence of spinelike bodies
n Hemiuroidea as a plesiomorphy of this superfamily.
he mature spermatozoon of P. palinurichthi. Aae1, anterior axoneme extremity 1;me 1; Ax2, axoneme 2; Cc, central core; Cm, cortical microtubules; D, doublet of
embrane; Pse, posterior spermatozoon extremity.
Another peculiarity of the mature spermatozoon of P. palin-urichthi is the length of the nucleus that occupies the entire lengthof the regions II and III. It is longer than the mitochondrion. Thischaracter has never been described previously in the Hemiuroidea.In A. laguncula, Quilichini et al. (2010b) described an elongate mito-chondrion located in regions II and III and a small nucleus locatedin the posterior part of region II and the beginning of region III.
The posterior extremity of the spermatozoon of P. palinurichthi,posterior to the end of the first axoneme, exhibits the modeldescribed from the hemiurid L. excisum (Ndiaye et al., 2012a),
with only one axoneme and nucleus. Later the nucleus disappearsand then the second axoneme disorganizes into doublets and sin-glets. This extremity corresponds to the type 3 (Quilichini et al.,2010a) characterized by: absence of cortical microtubules, and the
er An
seAssatgr
utv
A
m
R
B
B
B
D
E
E
G
G
G
G
H
H
H
J
J
J
fishes, I. Japanese Journal of Zoology 5, 249–541.
P.I. Ndiaye et al. / Zoologisch
equence – posterior extremity of the nucleus then the posteriorxtremity of the second axoneme. In the Lecithasteridae species. laguncula (Quilichini et al., 2010b), the posterior part of thepermatozoon is different. Indeed in A. laguncula this part of thepermatozoon exhibits a mitochondrion associated with the twoxonemes which then become disorganized progressively towardshe posterior end of the spermatozoon. Only the doublets and sin-lets resulting from the disorganization of the second axonemeeach the posterior end of the spermatozoon.
Thus, we think that more studies in all the families of hemi-roids are needed to elucidate the models of the spermatozoon inhe Hemiuroidea and the characters of the spermatozoa which arealuable for phylogenetic utility.
cknowledgements
Authors are thank to Online Access to Research in the Environ-ent (OARE) for facilitate us access to party of the bibliography.
eferences
â, C.T., Marchand, B., 1995. Spermiogenesis, spermatozoa and phyletic affinitiesin the Cestoda. Mémoires du Muséum National d’Histoire Naturelle. Serie B.Botanique. Paris 166, 87–95.
akhoum, A.J.S., Ndiaye, P.I., Sène, A., Bâ, C.T., Miquel, J., 2012. Spermiogenesis andultrastructure of the spermatozoon of Wardula capitellata (Digenea, Mesometri-dae), an intestinal parasite of the sparid teleost Sarpa salpa in Senegal. ActaParasitologica 57, 34–45.
akhoum, A.J.S., Torres, J., Shimalov, V.V., Bâ, C.T., Miquel, J., 2011. Spermiogene-sis and spermatozoon ultrastructure of Diplodiscus subclavatus (Pallas, 1760)(Paramphistomoidea, Diplodiscidae), an intestinal fluke of the pool frog Ranalessonae. Parasitology International 60, 64–74.
ollfus, R.-P., 1932. Trématodes. Résultats scientifiques du voyage aux Indes Orien-tales Néerlandaises de LL. AA. RR. Le Prince et la Princesse Léopold de Belgique.Bulletin de Musee royal d’Histoire naturelle de Belgique 2, 1–18.
hlers, U., 1984. Phylogenetisches System der Plathelminthes. Verhandlungen desnaturwissenschaftlichen Vereins Hamburg 27, 291–294.
uzet, L., Swiderski, Z., Mokhtar-Maamouri, F., 1981. Ultrastructure comparée duspermatozoïde des Cestodes. Relations avec la phylogénèse. Annales de Para-sitologie Humaine et Comparee 56, 247–259.
ibson, D.I., 1983. Kenmackenzia gen. nov. and Kenmackenziinae subfam. nov. (Dige-nea, Sclerodistomidae): new taxa to accommodate the giant trematode Distomagigas Nardo. Journal of Natural History 17, 189–202.
ibson, D.I., 2002. Family Sclerodistomidae Odhner, 1927. In: Gibson, D.I., Jones, A.,Bray, R.A. (Eds.), Keys to the Trematoda, vol. 1. CAB International and The NaturalHistory Museum, London, pp. 401–406.
ibson, D.I., Bray, R.A., 1977. The Azygiidae, Hirudinellidae, Ptychogonimidae, Scle-rodistomidae and Syncoeliidae (Digenea) of fishes from the north-east Atlantic.British Museum (Natural History): Zoology 32, 167–245.
ibson, D.I., Bray, R.A., 1979. The Hemiuroidea: terminology, systematics and evo-lution. British Museum (Natural History): Zoology 36, 35–146.
oberg, E.P., Jones, A., Bray, R.A., 1999. Phylogenetic analysis among the familiesof the Cyclophyllidea (Eucestoda) based on comparative morphology, with newhypotheses for co-evolution in vertebrates. Systematic Parasitology 42, 51–73.
oberg, E.P., Mariaux, J., Brooks, D.R., 2001. Phylogeny among orders of theEucestoda (Cercomeromorphae): integrating morphology, molecules and totalevidence. In: Littlewood, D.T.J., Bray, R.A. (Eds.), Interrelationships of the Platy-helminthes. Taylor and Francis, London, pp. 112–126.
oberg, E.P., Mariaux, J., Justine, J.-L., Brooks, D.R., Weekes, P.J., 1997. Phylogenyof the orders of the Eucestoda (Cercomeromorphae) based on comparativemorphology: historical perspectives and a new working hypothesis. Journal ofParasitology 83, 1128–1147.
amieson, B.G.M., Daddow, L.M., 1982. The ultrastructure of the spermatozoon ofNeochasmus sp. (Cryptogonimidae, Digenea, Trematoda) and its phylogeneticsignificance. International Journal for Parasitology 12, 547–559.
ustine, J.-L., 1991. Spermatozoa as a tool for taxonomy of species and supraspecific
taxa in the Platyhelminthes. In: Bacceti, B. (Ed.), Comparative Spermatology 20Years After. Raven Press, New York, pp. 981–984.ustine, J.-L., 1995. Spermatozoal ultrastructure and phylogeny of the parasiticPlatyhelminthes. Mémoires du Muséum National d’Histoire Naturelle. Serie B.Botanique. Paris 166, 55–86.
zeiger 252 (2013) 404–409 409
Justine, J.-L., 1998. Spermatozoa as phylogenetic characters for the Eucestoda. Jour-nal of Parasitology 84, 385–408.
Justine, J.-L., 2001. Spermatozoa as phylogenetic characters for the Platyhelminthes.In: Littlewood, D.T.J., Bray, R.A. (Eds.), Interrelationships of the Platyhelminthes.Taylor and Francis, London, pp. 231–238.
Justine, J.-L., Mattei, X., 1982. Étude ultrastructurale de la spermiogenèse et du sper-matozoïde d’un Plathelminthe: Gonapodasmius (Trematoda: Didymozoidae).Journal of Ultrastructure Research 79, 350–365.
Justine, J.-L., Mattei, X., 1983. A spermatozoon with two 9 + 0 axonemes in a parasiticflatworm, Didymozoon (Digenea: Didymozoidae). Journal of SubmicroscopicCytology 15, 1101–1105.
Justine, J.-L., Mattei, X., 1984a. Ultrastructural observations on the spermatozoon,ovocyte and fertilization process in Gonapodasmius, a gonochoristic Trematode(Trematoda: Digenea: Didymozoidae). Acta Zoologica (Stockholm, Sweden) 65,171–177.
Justine, J.-L., Mattei, X., 1984b. Atypical spermiogenesis in a parasitic flatworm,Didymozoon (Trematoda: Digenea: Didymozoidae). Journal of UltrastructureResearch 87, 106–111.
Kurochkin, Y.V., Parukhin, A.M., Korotaeva, V.D., 1971. New representatives andcomposition of the subfamily Prosorchinae Yamaguti, 1934 (Trematoda, Din-uridae). Parazitologiya 5, 212–221.
Levron, C., Miquel, J., Oros, M., Scholz, T., 2010. Spermatozoa of tapeworms (Platy-helminthes, Eucestoda): advances in ultrastructural and phylogenetic studies.Biological Reviews 85, 523–543.
Ndiaye, P.I., Diagne, P.Mb., Sène, A., Miquel, J., 2012a. Ultrastructure of the sperma-tozoon of the digenean Lecithocladium excisum (Rudolphi, 1819) (Hemiuroidea:Hemiuridae), a parasite of marine teleosts in Senegal. Folia Parasitologica 59,173–178.
Ndiaye, P.I., Miquel, J., Bâ, C.T., Marchand, B., 2004. Spermiogenesis and ultrastruc-ture of the spermatozoon of the liver fluke Fasciola gigantica Cobbold, 1856(Digenea, Fasciolidae), a parasite of cattle in Senegal. Journal of Parasitology90, 30–40.
Ndiaye, P.I., Quilichini, Y., Sène, A., Tkach, V.V., Bâ, C.T., Marchand, B., 2012b. Ultra-structural study of the male gamete of Pleurogonius truncatus Prudhoe, 1944(Platyhelminthes, Digenea, Pronocephalidae) parasite of Eretmochelys imbricata(Linnaeus, 1766). Comptes Rendus Biologies 335, 239–246.
Ndiaye, P.I., Quilichini, Y., Sène, A., Bâ, C.T., Marchand, B., 2011. Ultrastructure of thespermatozoon of the digenean Cricocephalus albus (Kuhl & van Hasselt, 1822)Loss, 1899 (Platyhelminthe, Pronocephaloidea, Pronocephalidae) parasite of the“hawksbill sea turtle” Eretmochelis imbricata (Linnaeus, 1766) in Senegal. Zool-ogischer Anzeiger 250, 215–222.
Pamplona-Basilio, M.C., Baptista-Farias, M.F.D., Kohn, A., 2001. Spermatogenesis andspermiogenesis in Didymocystis wedli Ariola, 1902 (Didymozoidae, Digenea).Memorias do Instituto Oswaldo Cruz 96, 1153–1159.
Quilichini, Y., Foata, J., Marchand, B., 2007. Ultrastructural study of the spermatozoonof Nicolla testiobliquum (Digenea, Opecoelidae) parasite of brown trout Salmotrutta (Pisces, Teleostei). Parasitology Research 101, 1295–1301.
Quilichini, Y., Foata, J., Justine, J.-L., Bray, R.A., Marchand, B., 2009. Sperm ultrastruc-ture of the digenean Siphoderina elongata (Platyhelminthes, Cryptogonimidae)intestinal parasite of Nemipterus furcosus (Pisces, Teleostei). ParasitologyResearch 105, 87–95.
Quilichini, Y., Foata, J., Justine, J.-L., Bray, R.A., Marchand, B., 2010a. Ultrastructuralstudy of the spermatozoon of Heterolebes maculosus (Digenea, Opisthole-betidae), a parasite of the porcupinefish Diodon hystrix (Pisces, Teleostei).Parasitology International 59, 427–434.
Quilichini, Y., Foata, J., Justine, J.-L., Bray, R.A., Marchand, B., 2010b. Sperma-tozoon ultrastructure of Aponurus laguncula (Digenea: Lecithasteridae), aparasite of Aluterus monoceros (Pisces, Teleostei). Parasitology International 59,22–28.
Sauvage, H.E., 1879. Mémoire sur la faune ichthyologique de l’île St. Paul. Archivesde zoologie expérimentale générale 8, 1–46.
Skrjabin, K.I., Guschanskaja, L.K., 1956. System of trematodes of the suborderHemiurata (Markevitsch, 1951) Skrjabin et Guschanskaja, 1954. Trudy Gelmintologichesko laboratorii 8, 144–158.
Thiéry, J.P., 1967. Mise en évidence des polysaccharides sur coupes fines en micro-scopie électronique. Journal of Microscopy 6, 987–1018.
Vigueras, I.P., 1940. Prosogonotrematidae n. fam. Y Prosogonotrema bilabiatumn. gen., n. sp. (Trematoda, Distoma) parásito de Ocyurus chrysurus (Bloch)(Pisces). Memorias de la Sociedad Cubana de Historia Natural “Felipe Poey” 14,249–252.
Yamaguti, S., 1934. Studies on the helminth fauna of Japan. Part 2. Trematodes of
Yamaguti, S., 1958. Systema Helminthum. Vol. 1. The Digenetic Trematodes of Ver-tebrates. Interscience, New York.
Yamaguti, S., 1971. Synopsis of the Digenetic Trematodes of Vertebrates. KeigakuPublishing Company, Tokyo.




















