
Regulation of COX2 Expression and IL6 Release by Particulate Matter in Airway Epithelial Cells
Upload
independentCategory
view
3download
0
of June 13, 2013.This information is current as
in Human Mature B CellsIL-6 Contributes to the Expression of RAGs
Christophe JaminSophie Hillion, Maryvonne Dueymes, Pierre Youinou and
http://www.jimmunol.org/content/179/10/67902007; 179:6790-6798; ;J Immunol
Referenceshttp://www.jimmunol.org/content/179/10/6790.full#ref-list-1
, 24 of which you can access for free at: cites 50 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

IL-6 Contributes to the Expression of RAGs in HumanMature B Cells1
Sophie Hillion, Maryvonne Dueymes, Pierre Youinou,2 and Christophe Jamin
Mature B cells acquire the capacity to revise rearranged Ig V region genes in secondary lymphoid organs. In previous studies, wedemonstrated that cross-linking the BCR and the CD40 induces the expression of the RAG1 and RAG2 enzymes and, thereby,secondary rearrangements. We examine herein the mechanism that underpins RAG1 and RAG2 expression in peripheral andtonsil B cells. Coordinated engagement of the BCR and CD40 promoted the synthesis of IL-6 and, thereby, up-regulation of itsreceptor on activated B lymphocytes. Furthermore, we provide evidence that IL-6 initiates the expression of RAGs in circulatingB cells, and extends those in tonsil B cells. Thus, neutralization of IL-6 or blocking of its receptor inhibits RAG expression.Moreover, we demonstrate that IL-6 impedes BCR-mediated termination of RAG gene expression in both population of B cells.The recovered inhibition of RAG gene transcription by IL-6 receptor blockade supports the notion that once recombination islaunched, its termination is also regulated by IL-6. Taken together, these studies provide new insight into the dual role of IL-6 ininducing and terminating expression of the recombinase machinery for secondary rearrangements in mature human B cells. TheJournal of Immunology, 2007, 179: 6790–6798.
T he B cell repertoire is shaped during early ontogenesisthrough random rearrangement, replacement, and selec-tion of Ig V region genes. These events are regulated by
RAG1 and RAG2 products which help assemble V(D)J Ig genes(1) in the bone marrow (BM)3. The expression of these enzymes ismodulated by environmental factors. Thus, the early stages of Bcell differentiation (2) require IL-7, which influences Ig gene re-arrangements and regulates repertoire diversification (3). In con-trast, IL-3 and IL-6 activate RAG gene transcription following in-teractions between stromal cells and lymphoid progenitor cells (4).For RAG gene expression to occur in the latter setting, two signalsare necessary: the first is mediated by contact through the cellmembrane with stromal cells, while the second comes from solublecytokines.
Because V(D)J genes are randomly selected to participate inrearrangements, self-reactive B cells may emerge in the BM. Atleast three mechanisms prevent the immune system from retainingsuch autoreactivity. These include deletion of autoreactive B cellclones by apoptosis (5), anergy (6), and receptor editing to shift theBCR (7, 8). The latter mechanism, which is triggered as soon asimmature B cells encounter their autoantigens, requires the assis-tance of newly synthesized RAG enzymes (9). With the benefit ofsuccessful secondary rearrangements, B cells purge autoreactivity,make up a nonself-reactive BCR repertoire, switch off their V(D)J
recombination machinery, and migrate to a secondary lymphoidorgan (SLO).
Based on the association of mature B cells with a decrease in theRAG proteins and an increase in the density level of the BCR (10),secondary RAG induction was originally thought to be restricted toimmature B cells. RAG products have since been detected in ma-ture B cells stimulated in culture (11), and in germinal centers(GCs), where peripheral V(D)J recombination may consummatesomatic hypermutations in centrocytes (12). Replacing the variableregion of a rearranged Ig with another would generate BCRs withhigher affinity for the Ag (13). RAG proteins were thus detected inB cells within and outside of the GCs in mice (12, 14, 15) andhumans (16–18).
Recent evidence suggests that significant differences exist be-tween mechanisms of RAG regulation in mice and humans. Inmice, stimulation of mature splenic B cells with CD40L in thepresence of IL-7 (19), or with LPS in the presence of IL-4 (11),induces the expression of RAG genes, whereas anti-IgM fails to doso in the presence of IL-1 (15). In humans, stimulation of blood Bcells with Staphylococcus aureus Cowan I and IL-2 induces theexpression of RAGs (20), whereas stimulation with CD40L plusIL-2, IL-4, or IL-10 exerts no effects on RAG up-regulation in GCB cells (21). The stage of B cell maturation also appears influentialin the RAG response of B cells to a given stimulus.
Although external signals seem to be essential for the expressionof RAGs in mature B cells, their precise nature remains elusive.Our previously reported model revealed that, following 5 days ofculture, mature human pre-GC B cells expressed RAG proteins inresponse to CD40L and anti-IgM stimulation (18). In contrast toprevious observations in immature and mature B lymphocytes,there was no need for further introduction of cytokine into theculture medium. Prompted by this observation, we addressed theissue of whether cytokines, derived from B cells, could enableRAG products to be synthesized in mature B cells following stim-ulation with CD40 and BCR cross-linking. We found that IL-6 isproduced by costimulated B cells, contributes to the initiation andthe extension of RAG expression, and modulates the subsequentBCR-mediated down-regulation.
Laboratory of Immunology, Brest University Medical School Hospital, Brest, France
Received for publication April 4, 2007. Accepted for publication September 6, 2007.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by grants from the Communaute Urbaine de Brest, theConseil Regional de Bretagne, the Academie Nationale Francaise de Medecine, andthe Ministere de l’Enseignement Superieur et de la Recherche.2 Address correspondence and reprint requests to Prof. Pierre Youinou, Laboratory ofImmunology, Brest University Medical School Hospital, BP824, Brest, France.E-mail address: [email protected] Abbreviations used in this paper: BM, bone marrow; SLO, a secondary lymphoidorgan; GC, germinal center; LN-PCR, ligation-mediated PCR.
Copyright © 2007 by The American Association of Immunologists, Inc. 0022-1767/07/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

Materials and MethodsIsolation of B cells
Normal blood was donated by volunteers from laboratory staff. Tonsilswere obtained from children undergoing routine tonsillectomy. The tis-sues were minced up, suspended in PBS and filtered to remove tissuefragments and clumps. Tonsil cell suspensions and blood were layeredonto Ficoll-Hypaque, and centrifuged for 30 min at 450 g. Mononuclearcells were incubated with neuraminidase-treated sheep RBC for 1 h at4°C, and T cells depleted by a second 25-min round of centrifugation.
All mAbs were purchased from Beckman-Coulter, unless otherwisespecified. B cells were stained with PE-conjugated anti-CD5 and FITC- orPE-linked to cyanin 5-conjugated anti-CD19 mAbs, and CD5-negative/CD19-positive B cells sorted on an Epics Elite (Beckman Coulter) FACS.All preparations were �98% pure CD19� as determined by FACS.
The murine National Institutes of Health-3T3 fibroblasts transfectedor not with human CD40L gene were a gift from John Gordon (Depart-ment of Immunology, University of Birmingham Medical School, Bir-mingham, U.K.).
Flow cytometry
We used PE-anti-CD40L (clone TRAP1) from BD Biosciences, FITC-anti-CD23 (clone 9P25) PE-anti-� (�) and FITC-anti-� (�) from Dako-Cytomation, and FITC-anti-CD19 (clone J4.119), -anti-CD10 (cloneALB1), -anti-CD44 (clone J.173), -anti-CD21 (clone BL13), FITC- andPE-anti-CD5 (clone BL1a), PE-anti-CD126 (clone M91), -anti-CD24(clone ALB9), -anti-CD62L (clone DREG56), FITC-anti-Ig, PE-linked tocyanin 5-labeled and PE-linked to cyanin 7-conjugated-anti-CD19. Intra-cellular staining of BCL-6 and IRF4 transcription factors were performedafter permeabilization of the cells with 70% methanol. Anti-BCL-6 andanti-IRF4 (Abcam) Abs were revealed with FITC-conjugated anti-rabbitAbs (Jackson ImmunoResearch Laboratories).
Cell culture
All the cells were cultured in RPMI 1640 medium supplemented with10% heat-inactivated FCS, 2 mM L-glutamine (Invitrogen Life Tech-nologies), 200U/ml penicillin, and 100 �g/ml streptomycin. To stimu-late B cells (22), we used 105 NIH-3T3 fibroblasts transfected or not,and incubated with mitomycin C (Sigma-Aldrich). Isolated B cells wereseeded at 2 � 105 cells/ml and cultured with, or without, 1 �g/mlanti-IgM Ab-coated beads for 24 h. Cells were also stimulated with 10�g/ml anti-IgM Ab-coated beads alone and, in some experiments, cul-tured in the presence of rIL-6 (ImmunoTools), anti-IL-6 Ab (Dako-Cytomation), or 40 ng/ml anti-IL-6R Ab (R&D Systems) at a concen-tration known to block IL-6 activity.
RNA isolation and RT-PCR
RNA was extracted by the RNAble kit (Eurobio), and reverse-transcribedwith Superscript II RNase H-RT according to the manufacturers’ instruc-tions (Invitrogen Life Technologies). RAG1 and RAG2 were amplified bynested RT-PCR using 1 �l of cDNA using TaqDNA polymerase (Invitro-gen Life Technologies) and the primer pairs listed in Table I. The PCRswere conducted as follows: in the first round, cDNA was amplified for 25cycles of 30 s at 94°C, 1 min at 56°C, and 1 min at 72°C, with a 10-minfinal extension at 72°C. The second round consisted of 35 cycles of 30 s at94°C, 1 min at 56°C, and 1 min at 72°C, with a 10-min final extension at72°C. There was only 1 round of 40-cycle PCR for the GAPDH RT-PCRamplification. The final products were analyzed on 2% agarose gels stainedwith ethidium bromide (18). Preliminary experiments were performed on697 pre-B cell line and HEp-2 epithelial cell line to ensure the lymphoidspecificity of the amplified products.
Single-cell protocol
Individual B cells were sorted into PCR tubes containing 10 �l RT buffer(1� first-strand buffer (Invitrogen Life Technologies), 5 �M random hex-amer, 0.5� RT buffer, 0.01 M DTT, and 0.5 mM dNTP (all from Pro-mega)), using than Epics Elite flow cytometer outfitted with an Autoclonesingle-cell deposition unit (Beckman Coulter). The mRNA conversion andRT-PCR for RAG1, RAG2, and GAPDH were performed as describedpreviously (18).
Quantitative RT-PCR
Quantitative RT-PCR was performed in 20 �l mixtures containing 50 ngcDNA template, 1� SYBR Green PCR Master mix (Applied Biosystems),and 500 nM of each primer pairs listed in Table I for IL-6 and 18S am-plification. Reactions consisted of 1 cycle at 50°C for 2 min, 1 cycle at
95°C for 10 min, followed by 40 cycles at 95°C for 15 s and at 60°C for1 min. Comparison of cycle thresholds was completed with the 2���Ct
method using 18S as internal control.
Ligation-mediated PCR (LM-PCR)
DNA was extracted from 105 cells using the DNAzol kit (Invitrogen LifeTechnologies) and 500 ng ligated to the LM-PCR linker at 20 pmol with T4DNA ligase (Promega) as previously described (18). Following two roundsof PCR using primers upstream the 5� end of either J�3 to J�7 or J�2 toJ�5 genes paired with primer encompassing the LM-PCR linker and therecombination signal sequence, PCR products were analyzed on 2% aga-rose gel stained with ethidium bromide (18). The specificity of all theamplified products were verified by sequence analysis.
Indirect immunofluorescence
Cultured blood B cells were washed in PBS, centrifuged onto glassslides, fixed in 4% paraformaldehyde, and prepared as described (18).First, cells were incubated in PBS containing 0.1% Nonidet P-40(Sigma-Aldrich), 2% nonfat dry milk, 5% FBS, and 0.02% sodiumazide, followed by incubation with Cohn fragment (Sigma-Aldrich) toavoid nonspecific fixation. Cells were left overnight with goat anti-RAG2 Ab (Santa Cruz Biotechnology), incubated with biotinylated anti-goat IgG (Jackson ImmunoResearch Laboratories) and visualized withtetramethylrhodamine isothiocyanate-streptavidin. Cells were then leftovernight with rabbit anti-RAG1 Ab (Santa Cruz Biotechnology) fol-lowed by FITC-anti-rabbit IgG (Jackson ImmunoResearch Laborato-ries). Aliquots of cells were stained without anti-RAG Abs but in thepresence of goat and rabbit Igs at the same concentrations and used ascontrols. Double stainings of either RAG protein with propidium iodidewere also performed at the same time. Cells were examined by fluo-rescence microscopy using an oil immersion lens (Zeiss). Preliminaryexperiments were performed on 697 pre-B and HEp-2 epithelial celllines to ensure the lymphoid specificity of the detected proteins.
ELISA
IL-6 was titrated in the supernatants of cultured cells by a commercialELISA using paired Abs (Beckman Coulter). Supernatant of CD40L-trans-fected fibroblasts was used as negative control.
ResultsInduction and termination of RAG gene transcription in tonsiland blood B cells
With reference to our previous demonstration that costimulationwith anti-IgM and CD40L induces the transcription of RAGs intonsil B cells (18), we analyzed the response of blood B cells tothe same stimuli. Transitional CD10high B cells, that have thecapacity to express RAG mRNA in secondary lymphoid organsof mice (23, 24), can be detected in the circulation of humans(25). In contrast to mouse B cells, however, circulating humantransitional type 1 B lymphocytes also express CD5 (25).
Table I. Sequences of oligonucleotides used as primers for theamplification of cDNA in RT-PCR and quantitative RT-PCR
Primers Oligonucleotides sequence (5�-3�)Productlength
RAG1 sense GAGAGAGCAGAGAACACACT 349 bpRAG1 antisense GCTGAGTTGGGACTGGCTTCTGACRAG1 nested sense CTGCTGAGCAAGGTACCTCAGCCAG 201 bpRAG1 nested
antisenseGAGAGGGTTTCCCCTCAAAGGAATC
RAG2 sense AGCCTTCTGCTTGCCACAGTCATAG 361 bpRAG2 antisense GAGGAGGGAGGTAGCAGGAATCCTTAGRAG2 nested sense CCCCTCTGGCCTTCAGACAAAAATC 154 bpRAG2 nested
antisenseGGGCCAGCCTTTTTGTCAAAG
IL-6 sense GACTTGCCTGGTGAAAATCATCACTG 179 bpIL-6 antisense GGGTCAGGGGTGGTTATTGCATCARN 18S sense GCCGCTAGAGGTGAAATTCTTG 108 bpARN 18S antisense CATTCTTGGCAAATGCTTTCGGAPDH sense CTTAGCACCCCTGGCCAAGG 522 bpGAPDH antisense CTTACTCCTTGGAGGCCATG
6791The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

Therefore, to ensure accurate determination of RAG expressionin circulating mature B cells, we sorted CD19�CD5� bloodlymphocytes and excluded CD19�CD5� that include transi-tional type 1 B cells. Thus, sorted cells from tonsils were foundto be CD10�/low, CD21�, CD23�, CD24�, CD62L�, BCL-6low/�, and IRF4�, and those from blood CD10�, CD44�,CD21�, CD23�, CD24�, CD62Llow, BCL-6low, and IRF4��
(Fig. 1A), confirming their mature status. B cells from tonsilscontained mRNA for RAG1 and RAG2, whereas those fromblood did not (Fig. 1B). We previously observed that 13.9 �4.2% of the CD19� CD5� tonsil B cells expressed RAG1 andRAG2 transcripts (18). Tonsil and blood B cells were then cul-tured on CD40L-transfected fibroblasts in the presence of anti-IgM Ab. After a 5-day culture (Fig. 1C), RAG1 and RAG2mRNA were maintained in B cells from tonsils, and induced inthose from blood. As shown in Fig. 2A, stimulated tonsil B cellswere found CD10�, CD44��, CD21�, CD23��, CD24low,CD62L�, BCL-6low, and IRF4��, and those from blood B cellswere found CD10�, CD44��, CD21�, CD23��, CD24�,CD62Llow/�, BCL-6low, and IRF4���. This is consistent withan activated mature status, and further indicates the lack oftransitional or immature B cells. Using single-cell RT-PCR,15.5 � 3.3% of the tonsil B cells and 8.1 � 1.9% of the bloodB cells expressed both RAG1 and RAG2 mRNA followingstimulation (Fig. 2B).
This raises the issues regarding the behavior of blood B cellsfollowing stimulation with anti-IgM Ab alone. Cross-linkingthe BCR (21, 26) on tonsil B cells down-regulated the RAGgene transcription (Fig. 1B). Similarly, blood B cells induced toexpress RAGs, and tonsil B cells sustained to express RAGs, byCD40L plus anti-IgM stimulation were impelled to down-reg-ulate RAG gene transcription by a 6-h treatment with anti-IgMalone (Fig. 1C). To determine whether RAG mRNA expressionwas associated with effective proteins at initiating rearrange-
ment, variation in the frequency of �- and �-positive B cellswere used as indicator of gene recombination process (27, 28).Induction of RAG expression following CD40L plus anti-IgMstimulation was thus associated with modifications to the �- and�-positive B cells (Fig. 2C). The �:� ratio decreased from1.86 � 0.17 to 1.34 � 0.19 in tonsil B cells, and from 1.6 �0.08 to 1.2 � 0.07 in blood B cells, suggesting that the Igrecombination process was efficiently induced. The presence ofactive Ig recombination enzyme was further demonstrated byLM-PCR on DNA isolated from CD40L and anti-IgM costimu-lated cells. DNA breaks or recombination signal sequences dueto recombinase activity were determined using primers up-stream the 5� end of each J�3 to J�7 and J�2 to J�5 genespaired with primer encompassing the LM-PCR linker and therecombination signal sequences. Specific PCR products corre-sponding to a recombination of the J�6, J�7, and J�5 geneswere found in tonsil B cells. Recombination of the J�4, J�7,J�3, and J�4 genes were also found in blood B cells (Fig. 2D).All these products were sequenced, and alignments with the 3�end upstream each recombinated gene of the genomic DNAunderscored the specificity of the products (data not shown).These data confirm that Ig recombination activity is ongoing inthis costimulation model.
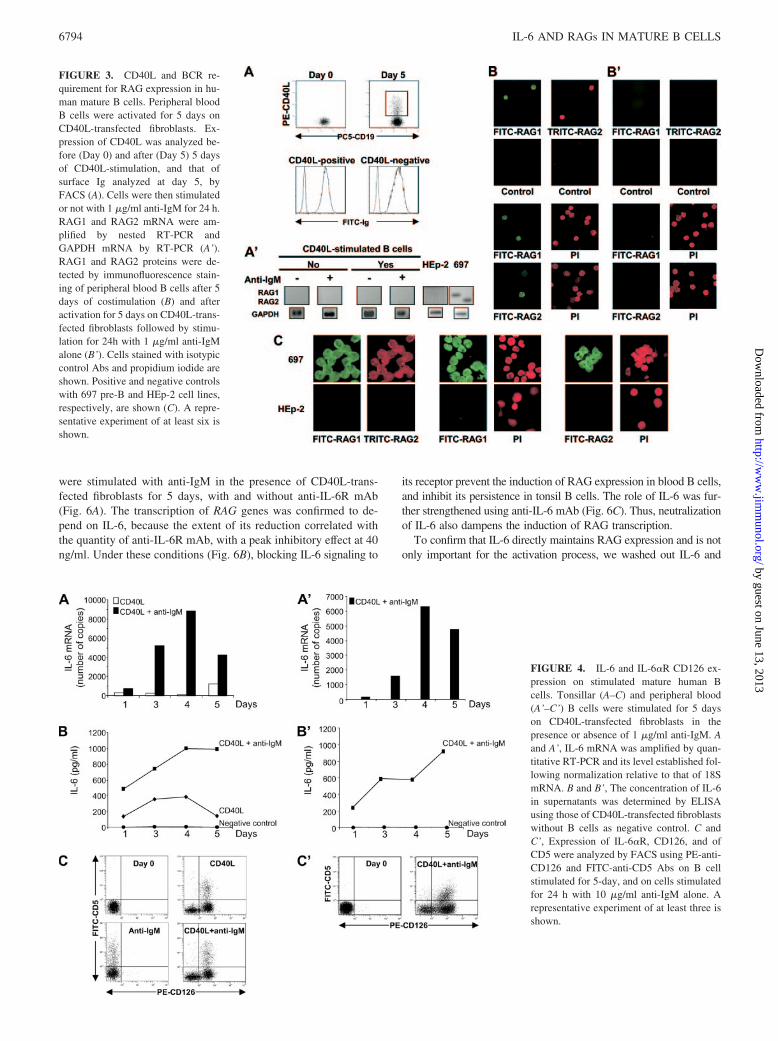
To gain insight into how costimulation triggers RAG up-reg-ulation, blood B cells were stimulated with CD40L-transfectedfibroblasts. This resulted in the up-regulation of CD40L on ma-ture Ig-positive cells (Fig. 3A) but not in the RAG expression(left panels in Fig. 3A’). Thus, preactivation with CD40L wasnot sufficient to induce RAG. After subsequent addition of anti-IgM Ab to cell culture, no RAG mRNA could be seen either(right panels in Fig. 3A’). Moreover, RAG1 and RAG2 proteinswere also induced in blood B cells costimulated with CD40L-transfected fibroblasts and anti-IgM Ab (Fig. 3B), as previouslyobserved in tonsil B cells (18), but not in cells sequentially
FIGURE 1. RAG1 and RAG2 expression in humanmature B cells. Tonsillar and peripheral blood B cellswere analyzed by FACS before stimulation to evaluatetheir maturation status using FITC-anti-CD10, -anti-CD44, -anti-CD21, -anti-CD23, -anti-BCL-6, -anti-IRF4, or PE-anti-CD24, -anti-CD62L, or –anti-CD5Abs. A representative example of three analyses isshown (A). RAG1 and RAG2 mRNA were amplified bynested RT-PCR using tonsillar and peripheral blood Bcells, before (B) and after (C) 5 days of culture onCD40L-transfected fibroblasts in the presence of 1�g/ml anti-IgM-coated beads, after 24 h of stimulationwith 10 �g/ml anti-IgM alone (B’), or after 5 days ofcostimulation followed by 6 h of culture with 10 �g/mlanti-IgM alone (C’). GAPDH mRNA was analyzed byRT-PCR. Negative and positive controls with HEp-2and 697 pre-B cell lines, respectively, are shown. A rep-resentative experiment of six is shown.
6792 IL-6 AND RAGs IN MATURE B CELLS
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

stimulated (Fig. 3B’). Overall, these data suggest that anti-IgMshould be added simultaneously with CD40L to induce RAGexpression.
Increased synthesis of IL-6 in mature activated B cells
IL-2, IL-4, and IL-10 are not required for RAG induction inblood B cells (21). However, there is evidence that IL-6 con-tributes to the regulation of RAG genes in immature B lympho-cytes (4). This data raises the possibility that IL-6 might alsoinduce RAG expression in mature B lymphocytes. To addressthis question, first, the amount of IL-6 mRNA was measured byquantitative RT-PCR in tonsil and peripheral blood B cells (Fig.4, A and A’). The transcription of IL-6 was increased in B cellscultured with CD40L-transfected fibroblasts in the presence ofanti-IgM Ab after a 4-day incubation. Interestingly, the elevatedtranscription of IL-6 mRNA was concomitant with the sustainedRAG expression in tonsil B cells and the induction of RAGtranscription in blood B cells, as shown in Fig. 1C. In addition,IL-6 protein was detected by ELISA in the supernatant of thesecultured B cells (Fig. 4, B and B’). However, for IL-6 to befunctional its receptor must be present on the surface of matureB cells. CD126 was absent before stimulation, or after a 24-hstimulation with anti-IgM Ab, but was expressed on 47.8 �3.3% of tonsil B cells stimulated with CD40L, and 53.3 � 2.5%of those stimulated with CD40L plus anti-IgM Ab (Fig. 4C).This frequency reached 38.5 � 1.5% of the peripheral blood B
cells (Fig. 4C’). Following CD40L and anti-IgM costimulation,we observed that RAG-expressing cells were restricted to thoseturned positive for CD5 (18). Interestingly, a CD5-positive sub-population, appeared expressing CD126 and representing17.9 � 2.5% and 16.9 � 1.4% of CD40L- and CD40L plusanti-IgM Ab-stimulated tonsillar B cells, respectively. Withinthe peripheral blood B cells, this subpopulation represented18.4 � 1.5%. These data suggest that IL-6 is a candidate thatmay provide the third signal for RAG gene transcription in Bcells induced to up-regulate CD126.
To further decipher the role of IL-6, circulating blood B cellswere activated for 24 h with increasing amount of rIL-6 and forseveral days with 1000 pg/ml rIL-6. RAG expression could notbe induced in such conditions (Fig. 5A). When rIL-6 was addedto CD40L-stimulated B cells (Fig. 5B) or to anti-IgM Ab-stim-ulated B cells (Fig. 5C), no RAG induction was observed eithereven with elevated level of rIL-6. These data indicate that ex-ogenous IL-6 alone is unable to promote RAG expression, evenin CD40L-stimulated cells. Furthermore, they also highlightthat BCR signaling is absolutely essential together with CD40signals.
IL-6 regulates RAG mRNA expression in mature B cells
To verify the requirement for IL-6 in RAG expression of CD40and BCR costimulated B lymphocytes, tonsil and blood B cells
FIGURE 2. Frequency of RAG-expressing cells and enzymatic activity in human mature B cells. B cells were analyzed by FACS after 5 days of cultureon CD40L-transfected fibroblasts in the presence of 1 �g/ml anti-IgM-coated beads to determine their maturation status using FITC-anti-CD10, -anti-CD44,-anti-CD21, -anti-CD23, -anti-BCL-6, -anti-IRF4, or PE-anti-CD24, -anti-CD62L, or -anti-CD5 Abs. A representative example of three analyses is shown(A). After stimulation, individual tonsillar and peripheral blood B cells were sorted with the Autoclone single-cell deposition unit of the Epics Elite flowcytometer. Among the sorted cells, only those positive for GAPDH-mRNA were analyzed for expression of RAG1 and RAG2 mRNA by nested RT-PCR.The frequencies of single RAG1-positive and/or RAG2-positive cells are shown. Mean � SD of three experiments (B). Tonsillar and peripheral blood Bcells were analyzed by FACS before (Day 0) and after (Day 5) 5 days of CD40L and anti-IgM costimulation using FITC-anti-� and PE-anti-� Abs todetermine the proportion of �- and �-positive cells. A representative example of three analyses is shown where numbers indicate the percentages of positivecells (C). RAG1 and RAG2 enzymatic activity was also evaluated after 5 days of CD40L and anti-IgM costimulation by LM-PCR to detect the J� and J�signal breaks. LM-PCR products were visualized on ethidium bromide-stained gel. A representative experiment of two is shown. Product lengths areindicated (D).
6793The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
were stimulated with anti-IgM in the presence of CD40L-trans-fected fibroblasts for 5 days, with and without anti-IL-6R mAb(Fig. 6A). The transcription of RAG genes was confirmed to de-pend on IL-6, because the extent of its reduction correlated withthe quantity of anti-IL-6R mAb, with a peak inhibitory effect at 40ng/ml. Under these conditions (Fig. 6B), blocking IL-6 signaling to
its receptor prevent the induction of RAG expression in blood B cells,and inhibit its persistence in tonsil B cells. The role of IL-6 was fur-ther strengthened using anti-IL-6 mAb (Fig. 6C). Thus, neutralizationof IL-6 also dampens the induction of RAG transcription.
To confirm that IL-6 directly maintains RAG expression and is notonly important for the activation process, we washed out IL-6 and
FIGURE 3. CD40L and BCR re-quirement for RAG expression in hu-man mature B cells. Peripheral bloodB cells were activated for 5 days onCD40L-transfected fibroblasts. Ex-pression of CD40L was analyzed be-fore (Day 0) and after (Day 5) 5 daysof CD40L-stimulation, and that ofsurface Ig analyzed at day 5, byFACS (A). Cells were then stimulatedor not with 1 �g/ml anti-IgM for 24 h.RAG1 and RAG2 mRNA were am-plified by nested RT-PCR andGAPDH mRNA by RT-PCR (A’).RAG1 and RAG2 proteins were de-tected by immunofluorescence stain-ing of peripheral blood B cells after 5days of costimulation (B) and afteractivation for 5 days on CD40L-trans-fected fibroblasts followed by stimu-lation for 24h with 1 �g/ml anti-IgMalone (B’). Cells stained with isotypiccontrol Abs and propidium iodide areshown. Positive and negative controlswith 697 pre-B and HEp-2 cell lines,respectively, are shown (C). A repre-sentative experiment of at least six isshown.
FIGURE 4. IL-6 and IL-6�R CD126 ex-pression on stimulated mature human Bcells. Tonsillar (A–C) and peripheral blood(A’–C’) B cells were stimulated for 5 dayson CD40L-transfected fibroblasts in thepresence or absence of 1 �g/ml anti-IgM. Aand A’, IL-6 mRNA was amplified by quan-titative RT-PCR and its level established fol-lowing normalization relative to that of 18SmRNA. B and B’, The concentration of IL-6in supernatants was determined by ELISAusing those of CD40L-transfected fibroblastswithout B cells as negative control. C andC’, Expression of IL-6�R, CD126, and ofCD5 were analyzed by FACS using PE-anti-CD126 and FITC-anti-CD5 Abs on B cellstimulated for 5-day, and on cells stimulatedfor 24 h with 10 �g/ml anti-IgM alone. Arepresentative experiment of at least three isshown.
6794 IL-6 AND RAGs IN MATURE B CELLS
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

BCR cross-linking one day before the endpoint cultures of tonsil Bcells (day 4), and then added back IL-6 on these cells further culturedon CD40L-transfected fibroblasts or in medium alone. With the per-sistence of CD40L signals, RAG transcripts were maintained inde-pendently on the addition of rIL-6 (Fig. 7A), likely due to the endog-
enous secretion of IL-6 by these cells (Fig. 4B). However, when theywere further cultured in medium alone, RAG transcription was sus-tained only in the presence of rIL-6 (Fig. 7B). These data indicate that,following their induction, RAG expression is maintained due to thecontribution of IL-6.
FIGURE 5. Effect of rIL-6 on RAG expression in mature human B cells. RAG1 and RAG2 mRNA were amplified by nested RT-PCR in peripheral bloodB cells following 24 h of stimulation with a range of rIL-6 concentrations (0, 10, 100, and 1000 pg/ml), and in blood B cells cultured with 1000 pg/mlrIL-6 for 5 days (A). Tonsillar and peripheral blood B cells were stimulated for 5 days on CD40L-transfected fibroblasts (B), or for 24 h with 10 �g/mlanti-IgM (C), with a range of rIL-6 concentrations (0, 10, 100, and 1000 pg/ml). RAG1 and RAG2 mRNA were amplified by nested RT-PCR and that ofGAPDH by RT-PCR. A representative example of three experiments is shown.
FIGURE 6. IL-6-dependent induction of RAG in mature human B cells. RAG1 and RAG2 mRNA were amplified by nested RT-PCR in tonsillar andperipheral B cells following 5 days of culture on CD40L-transfected fibroblasts in the presence of 1 �g/ml anti-IgM Abs, with a range of anti-IL-6R Abconcentrations, (0, 4, 40, or 400 ng/ml) (A), before and after 5 days of stimulation on CD40L-transfected fibroblasts with 1 �g/ml anti-IgM Abs in thepresence or absence of 40 ng/ml anti-IL-6R Abs (B), or with a range of anti-IL-6 Ab concentrations (0, 1, 10, and 100 ng/ml) (C). A representative exampleof four experiments is shown.
6795The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

To determine whether IL-6 could also have an effect on therepression of RAG transcription, we measured RAG transcriptsin anti-IgM-stimulated B cells cultured on CD40L-transfectedfibroblasts. Both RAG1 and RAG2 mRNA were expressed intonsil and blood B cells after a 5-day culture. The conditionedsupernatants were collected, and the cells washed in RPMI1640. Tonsil as well as blood B cells were then seeded into 3flasks. All cells were cultured with anti-IgM Ab for 6 h. Thefirst aliquots of tonsil B cells and blood B cells were placed intofresh medium (left column in Fig. 8), the second aliquots werecultured in conditioned supernatants (medium column in Fig. 8),and the third cultured in conditioned supernatants with anti-IL-6R mAb (right column in Fig. 8). Repression of RAG tran-scription was achieved by anti-IgM in the first aliquots, im-peded in the second aliquots by the presence of IL-6 inconditioned supernatants, and recovered in the third aliquots byanti-IL-6R-mAb-induced blockade of IL-6 in the conditionedsupernatants. These results demonstrate that IL-6 has the ca-pacity to modulate the RAG gene transcription by inhibiting itsrepression.
DiscussionThe expression of RAG is tightly regulated in immature, and ma-ture B cells (2–4, 11, 12). We previously noted that, althoughcytokines are required for RAG up-regulation, the provision ofcytokines is dispensable in our model of RAG induction in humanmature B cells, in which the cells are costimulated by CD40L andanti-IgM Ab for up to 5 days (18). We hypothesized that, follow-ing 5 days of costimulation, some cytokines produced from theactivated B cells acted in an autocrine manner to induce RAGexpression. To test this hypothesis, we assessed the presence ofcytokines in the culture medium, and searched for their receptorson activated B cells. The results revealed that, following stimula-tion, tonsil and blood B cells behaved similarly by secreting acytokine required to induce the expression of RAGs, and to sub-sequently control their down-regulation.
Several investigations have demonstrated that, in SLOs, RAG-positive B cells could be immature transitional type 2 B cells (15,23, 24). Given that transitional type 1 B cells can be detected in thecirculation of human (25), circulating RAG-positive B cells couldbe precursor cells recently emigrated from the BM. However, webelieve this is unlikely in our study. Indeed, human circulatingtransitional type 1 B cells belong to the CD5-positive subpopula-tion (25) that have been excluded from the analysis. Moreover,stimulation of immature B cells with anti-IgM Abs induced RAGexpression (29), which is not the case in our sensitive RT-PCRassay (Fig. 1B). Furthermore, stimulation of splenic mouse B cellswith anti-CD40 plus IL-7 (19) induces IL-7R, which ultimatelyleads to the transcription of RAG genes. Despite the presence ofmRNA for CD127 in resting B cells, we were unable to detectCD127-positive B cells by FACS, and activation of B cells withCD40L and/or anti-IgM Ab, in the presence of IL-7 did not lead toIL-7R up-regulation on the membrane (data not shown). BecauseIL-7R is expressed on human pre-B lymphocytes but not on ma-ture lymphocytes (30, 31), the lack of IL-7R expression is a furtherargument indicating that immature B cells are absent from thepurified CD19�CD5� blood B cells in our study and that inductionof RAG cannot be ascribed to the presence of these cells either inblood, or in tonsil B cells (18), following CD40L and anti-IgMcostimulation. On top of that, these data indicate that human ma-ture B cells do not require IL-7 to transcribe RAG genes, in con-trast to murine B cells (19), and that secondary V(D)J rearrange-ments are, thus, likely to be regulated by other cytokines.
Given its high level in SLOs (32) and its involvement in RAGexpression in BM progenitor cells (4), IL-6 appears to be a crediblecandidate for RAG regulation in peripheral B cells. The experi-ments describe in the current study show that activation of matureB cells by CD40L and anti-IgM increases the level of mRNA forIL-6 and the amount of IL-6 protein. For IL-6 to be functionallyeffective, its receptor must be expressed at detectable levels on thecell surface. Consistent with previous reports (33), we noted thatactivated, but not resting, B cells harbor CD126 on their surface.How IL-6 and IL-6R expression are regulated following CD40 andIgM coligation is still an open question. Because engagement ofthe BCR alone takes 30 h to induce apoptosis (34, 35), there islittle time to evaluate IL-6 synthesis and IL-6R up-regulation.However, it appears that a 24-h stimulation of B cells had noeffects on the expression of IL-6Rs. In contrast, engagement ofCD40 alone was able to induce detectable, albeit a modest increasein IL-6 production. In accord with previous reports (36), this isassociated with efficient up-regulation of CD126. Coligated CD40and BCR act synergistically to increase the production of IL-6 andexpression of its receptor. To demonstrate the direct role of IL-6 inthe induction of RAGs, rIL-6 was added to cultured B cells. RAG1
FIGURE 7. Role of IL-6 on sustained RAG expression in mature hu-man B cells. Tonsillar B cells were cultured for 4 days on CD40L-transfected fibroblasts in the presence of 1 �g/ml anti-IgM to induceRAG1 and RAG2 expression. Cells were collected to wash out IL-6 andBCR cross-linking. These cells were further cultured for 24 h onCD40L-transfected fibroblasts (A), or in medium alone (B), with orwithout 1000 pg/ml rIL-6. RAG1 and RAG2 mRNA were amplified bynested RT-PCR and that of GAPDH by RT-PCR. A representative ex-ample of 3 experiments is shown.
FIGURE 8. Influence of IL-6 on RAG repression in mature human Bcells. Tonsillar and peripheral blood B cells were cultured for 5 days onCD40L-transfected fibroblasts in the presence of 1 �g/ml anti-IgM to in-duce RAG1 and RAG2 expression. Cells and culture supernatants werecollected. The cells were then stimulated for 6 h with 10 �g/ml anti-IgMalone, with the culture supernatant only, and in the presence of 40 ng/mlanti-IL-6R Abs. RAG1 and RAG2 mRNA were amplified by nested RT-PCR, and that of GAPDH by RT-PCR. A representative example of threeexperiments is shown.
6796 IL-6 AND RAGs IN MATURE B CELLS
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

and RAG2 expression was not detected in IL-6-activated cells, andnot up-regulated either in CD40L- or in anti-IgM- and IL-6 co-stimulated B cells. These findings establish that IL-6 is unable todirectly regulate RAG induction even in CD40L-activated B cellsor anti-IgM-stimulated B cells. However, when IL-6 was neutral-ized by anti-IL-6 mAb, or IL-6 binding to its receptor inhibited byanti-IL-6R mAb, up-regulation of RAGs induced by CD40L plusanti-IgM could be inhibited. This indicates that RAG expressiondepends on IL-6 signaling in CD40L-activated mature B cells pro-vided that BCR signaling pathways are also stimulated. Interest-ingly, following removal of inducing signals, IL-6 was also able tomaintain RAG expression of stimulated B cells when cultured inmedium without activating signals. This suggests that IL-6 partic-ipates to the induction of RAG expressions, and also contributes totheir sustained up-regulation. However, it remains unclear howdoes IL-6 up-regulate RAG expression: IL-6 may possibly activatetranscription factors involved in RAG gene transcription (37). Al-ternatively, IL-6 may activate transcription factors that induce acis-element to counteract a distant silencer that, normally, re-presses the transcription of RAGs (38). The physiological settingthat triggers such RAG gene transcription might be direct cognateinteraction between B and T cells via CD40 after BCR engage-ment. This interaction could permit the Ag-specific B cells tosecrete and respond to IL-6 in an autocrine manner. Furthermore,following IL-6 synthesis by follicular dendritic cells, T cells orstromal cells, IL-6 may also operate in a paracrine fashion (32).Secondary V(D)J recombination may thus modify specificity ofthe BCR and allow the tuning of a more efficient immune response.These events may affect a subpopulation of B cells, because onlypart of the cells was induced to express CD126. In this context, itis interesting that expression of the RAG products is associatedwith the expression of the CD5 marker (18). The phenotypic anal-ysis confirmed that CD5-positive B cells are among those inducedto express CD126. It is, therefore, likely that RAG expressionmight be confined to the CD126-positive subpopulation of acti-vated B cells expressing the CD5 marker. The CD5-positive Bcells are mainly located in the follicular mantle zone of the GCs inSLOs (39). They may thus recombinate their Ig genes before theyenter the GCs. CD5 has a negative function in the BCR signaling(40). Therefore, one may hypothesized that B cells with BCR af-finity not strong enough to allow migration in the GCs and theensuing maturation, would receive BCR signals triggering CD5expression which will elevate the BCR threshold. Together withCD40 ligation, BCR engagement will induces IL-6 secretion. Thisthird signal would then promote V(D)J rearrangements. The rele-vance of RAG induction may be to raise the BCR affinity up to thelevel required for maturation (41).
The promotion of the recombinase machinery for secondary IgVgene rearrangements by IL-6 as a result of RAG expressionprompted us to determine whether IL-6 was also involved in thetermination of the receptor revision. Following a 5-day costimu-lation with CD40L and anti-IgM, human B cells transcribed RAG1and RAG2, and up-regulated CD126 on their surface. The pheno-type changes indicate that RAG-expressing mature B cells shouldrespond to IL-6 stimulation. We first confirmed that a strong BCR-mediated signal alone restrains RAG expression (21, 26). This wasreversed by keeping the cells in culture medium. Surprisingly,when IL-6 binding was blocked with anti-IL-6R, the BCR-inducedrepression of RAG expression was recovered. These experimentsfurther demonstrate that IL-6 controls BCR-mediated down-regu-lation of RAGs. IL-6 is, thus, not only necessary for the inductionof RAG genes but is also influential in its suppression. The re-maining bands for RAG1 observed when IL-6 is absent or blocked(Fig. 8) suggest that IL-6 mainly affect RAG2 mRNA expression.
IL-6 is able to induce G1 arrest (42). The possibility thereforeexists that this effect may participate in the variation of RAG2mRNA regulation. Finally, the current study indicates that IL-6 iskey in the regulation of RAG expression in human mature B cells.This is intriguing, considering that IL-6 is overexpressed in auto-immune disease (43–45). High levels of IL-6 in patients with au-toimmune diseases could impede BCR engagement to switch offRAG expression, and may thus participate in uncontrolled BCRrevision (46–48) leading to the production of high-affinity auto-Abs (49, 50).
AcknowledgmentsWe acknowledge Prof. Rizgar A. Mageed for his valuable editorial assis-tance. Thanks are also due to Simone Forest and Cindy Sene for secretarialassistance.
DisclosuresThe authors have no financial conflict of interest.
References1. Oettinger, M. A., D. G. Schatz, C. Gorka, and D. Baltimore. 1990. RAG-1 and
RAG-2, adjacent genes that synergistically activate VDJ recombination. Science248: 1517–1523.
2. Faust, E. A., D. C. Saffran, D. Toksoz, D. A. Williams, and O. N. Witte. 1993.Distinctive growth requirements and gene expression patterns distinguish pro-genitor B cells from pre-B cells. J. Exp. Med. 177: 915–923.
3. Billips, L. G., C. A. Nunez, F. E. Bertrand III, A. K. Stankovic, G. L. Gartland,P. D. Burrows, and M. D. Cooper. 1995. Immunoglobulin recombinase geneactivity is modulated reciprocally by interleukin 7 and CD19 in B cell progeni-tors. J. Exp. Med. 182: 973–982.
4. Muraguchi, A., H. Tagoh, T. Kitagawa, T. Nagata, and H. Kishi. 1998. Stromalcells and cytokines in the induction of recombination activating gene (RAG)expression in a human lymphoid progenitor cell. Leuk. Lymphoma 30: 73–85.
5. Nemazee, D., and K. Buerki. 1989. Clonal deletion of autoreactive B lympho-cytes in bone marrow chimeras. Proc. Natl. Acad. Sci. USA 86: 8039–8043.
6. Goodnow, C. C., J. Crosbie, S. Adelstein, T. B. Lavoie, S. J. Smith-Gill,R. A. Brink, H. Pritchard-Briscoe, J. S. Wotherspoon, R. H. Loblay, andK. Raphael. 1988. Altered immunoglobulin expression and functional silencingof self-reactive B lymphocytes in transgenic mice. Nature 334: 676–682.
7. Gay, D., T. Saunders, S. Camper, and M. Weigert. 1993. Receptor editing: anapproach by autoreactive B cells to escape tolerance. J. Exp. Med. 177:999–1008.
8. Tiegs, S. L., D. M. Russel, and D. Nemazee. 1993. Receptor editing in self-reactive bone marrow B cells. J. Exp. Med. 177: 1009–1020.
9. Hillion, S., C. Rochas, P. Youinou, and C. Jamin. 2005. Expression and reex-pression of recombination activating genes: relevance to the development of au-toimmune states. Ann. NY Acad. Sci. 1050: 10–18.
10. Jankovic, M., R. Casellas, N. Yannoutsos, H. Wardemann, and M. C. Nussenz-weig. 2004. RAGs and regulation of autoantibodies. Annu. Rev. Immunol. 22:485–501.
11. Hikida, M., M. Mori, T. Takai, K. Tomochika, K. Hamatani, and H. Ohmori.1996. Reexpression of RAG-1 and RAG-2 genes in activated mature mouse Bcells. Science 274: 2092–2094.
12. Han, S., B. Zheng, D. G. Schatz, E. Spanopoulou, and G. Kelsoe. 1996. Neotenyin lymphocytes: Rag1 and Rag2 expression in germinal center B cells. Science274: 2094–2097.
13. Meffre, E., R. Casellas, and M. C. Nussenzweig. 2000. Antibody regulation of Bcell development. Nat. Immunol. 1: 379–385.
14. Hikida, M., M. Mori, T. Kawabata, T. Takai, and H. Ohmori. 1997. Character-ization of B cells expressing recombination activating genes in germinal centersof immunized mouse lymph nodes. J. Immunol. 158: 2509–2512.
15. Igarashi, H., N. Kuwata, K. Kiyota, K. Sumita, T. Suda, S., Ono, S. R. Bauer, andN. Sakaguchi. 2001. Localization of recombination activating gene 1/green flu-orescent protein (RAG1/GFP) expression in secondary lymphoid organs afterimmunization with T-dependent antigens in rag1/gfp knockin mice. Blood 97:2680–2687.
16. Girschick, H. J., A. C. Grammer, T. Nanki, M. Mayo, and P. E. Lipsky. 2001.RAG1 and RAG2 expression by B cell subsets from human tonsil and peripheralblood. J. Immunol. 166: 377–386.
17. Giachino, C., Padovan, and E. A. Lanzavecchia. 1998. Re-expression of RAG-1and RAG-2 genes and evidence for secondary rearrangements in human germinalcenter B lymphocytes. Eur. J. Immunol. 28: 3506–3513.
18. Hillion, S., A. Saraux, P. Youinou, and C. Jamin. 2005. Expression of RAGs inperipheral B cells outside germinal centers is associated with the expression ofCD5. J. Immunol. 174: 5553–5561.
19. Hikida, M., Y. Nakayama, Y. Yamashita, Y. Kumazawa, S. I. Nishikawa, andH. Ohmori. 1998. Expression of recombination activating genes in germinal cen-ter B cells: involvement of interleukin 7 (IL-7) and the IL-7 receptor. J. Exp. Med.188: 365–372.
20. Nagafuchi, H., H. Yoshikawa, Y. Takeba, K. Nara, K. Miura, M. S. Kurokawa,and N. Suzuki. 2004. Recombination activating genes (RAG) induce secondary
6797The Journal of Immunology
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from

Ig gene rearrangement in and subsequent apoptosis of human peripheral bloodcirculating B lymphocytes. Clin. Exp. Immunol. 136: 76–84.
21. Meffre, E., F. Papavasiliou, P. Cohen, O. de Bouteiller, D. Bell, H. Karasuyama,C. Schiff, J. Banchereau, Y. J. Liu, and M. C. Nussenzweig. 1998. Antigen re-ceptor engagement turns off the VDJ recombination machinery in human tonsil Bcells. J. Exp. Med. 188: 765–772.
22. Schultze, J., L. Michalak, S. Seamon, M. J. Dranoff, G. Jung, K. J. Daley,J. C. Delgado, J. G. Gribben, and L. M. Nadler. 1997. CD40-activated human Bcells: an alternative source of highly efficient antigen presenting cells to generateautologous antigen-specific T cells for adoptive immunotherapy. J. Clin. Invest.100: 2757–2765.
23. Gartner, F., F. W. Alt, R. J. Monroe, and K. J. Seidl. 2000. Antigen-independentappearance of recombination activating gene (RAG)-positive bone marrow Bcells in the spleens of immunized mice. J. Exp. Med. 192: 1745–1754.
24. Yu, W., H. Nagaoka, M. Jankovic, Z. Misulovin, H. Suh, A. Rolink, F. Melchers,E. Meffre, and M. C. Nussenzweig. 1999. Continued RAG expression in latestages of B cell development and no apparent re-induction after immunization.Nature 400: 682–687.
25. Sims, G. P., R. Ettinger, Y. Shirota, C. H. Yarboro, G. G. Illei, and P. E. Lipsky.2005. Identification and characterization of circulating human transitional B cells.Blood 105: 4390–4398.
26. Hertz, M., V. Kouskoff, T. Nakamura, and D. Nemazee. 1998. VDJ recombinaseinduction in splenic B lymphocytes is inhibited by antigen-receptor signalling.Nature 394: 292–295.
27. Li, Y., H. Li, and M. Weigert. 2002. Autoreactive B cells in the marginal zonethat express dual receptors. J. Exp. Med. 195: 181–188.
28. Diamant, E., Z. Keren, and D. Melamed. 2005. CD19 regulates positive selectionand maturation in B lymphopoiesis: lack of CD19 imposes developmental arrestof immature B cells and consequential stimulation of receptor editing. Blood 105:3247–3254.
29. Hertz, M., and D. Nemazee. 1997. BCR ligation induces receptor editing inIgM�IgD� bone marrow B cells in vitro. Immunity 6: 429–436.
30. Armitage, R. J., S. F. Ziegler, M. P. Beckmann, R. L. Idzerda, L. S. Park, andW. C. Fanslow. 1991. Expression of receptors for interleukin 4 and interleukin 7on human T cells. Adv. Exp. Med. Biol. 292: 121–130.
31. Ryan, D. H., B. L. Nuccie, I. Ritterman, J. L. Liesveld, C. N. Abboud, andR. A. Insel. 1997. Expression of interleukin-7 receptor by lineage-negative hu-man bone marrow progenitors with enhanced lymphoid proliferative potentialand B-lineage differentiation capacity. Blood 89: 929–940.
32. Skibinski, G., A. Skibinska, M. Deckers, and K. James. 1998. Tonsil stromal-celllines expressing FDC-like properties: isolation, characterization, and interactionwith B lymphocytes. Dev. Immunol. 6: 273–284.
33. Hirata, Y., T. Taga, M. Hibi, N. Nakano, T. Hirano, and T. Kishimoto. 1989.Characterization of IL-6 receptor expression by monoclonal and polyclonal an-tibodies. J. Immunol. 143: 2900–2906.
34. Chaouchi, N., A. Vazquez, P. Galanaud, and C. Leprince. 1995. B cell antigenreceptor-mediated apoptosis. Importance of accessory molecules CD19 andCD22, and of surface IgM cross-linking. J. Immunol. 154: 3094–3104.
35. Pers, J. O., C. Jamin, R. Le Corre, P. M. Lydyard, and P. Youinou. 1998. Ligationof CD5 on resting B cells, but not on resting T cells, results in apoptosis. Eur.J. Immunol. 28: 4170–4176.
36. Baccam, M., S. Y. Woo, C. Winson, and G. A. Bishop. 2003. CD40-mediatedtranscriptional regulation of the IL-6 gene in B lymphocytes: involvement ofNF-kappa B, AP-1, and C/EBP. J. Immunol. 170: 3099–3108.
37. Monroe, R. J., F. Chen, R. Ferrini, L. Davidson, and F. W. Alt. 1999. RAG2 isregulated differentially in B and T cells by elements 5� of the promoter. Proc.Natl. Acad. Sci. USA 96: 12713–12718.
38. Yannoutsos, N., V. Barreto, Z. Misulovin, A. Gazumyan, W. Yu, N. Rajewsky,B. R. Peixoto, T. Eisenreich, and M.C. Nussenzweig. 2004. A cis element in therecombination activating gene locus regulates gene expression by counteractinga distant silencer. Nat. Immunol. 5: 443–450.
39. Dono, M., V. L. Burgio, C. Tacchetti, A. Favre, A. Augliera, S. Zupo,G. Taborelli, N. Chiorazzi, C. E. Grossi, and M. Ferrarini. 1996. Subepithelial Bcells in the human palatine tonsil, I: morphologic, cytochemical and phenotypiccharacterization. Eur. J. Immunol. 26: 2035–2042.
40. Bikah, G., J. Carey, J. R. Ciallella, A. Tarakhovsky, and S. Bondada. 1996.CD5-mediated negative regulation of antigen receptor-induced growth signals inB-1 B cells. Science 274: 1906–1909.
41. Magari, M., T. Sawatari, Y. Kawano, M. Cascalho, M. Wabl, N. Kanayama,M. Hikida, and H. Ohmori. 2002. Contribution of light chain rearrangement inperipheral B cells to the generation of high-affinity antibodies. Eur. J. Immunol.32: 957–966.
42. Mori, S., K. Murakami-Mori, and B. Bonavida. 1999. Interleukin-6 induces G1
arrest through induction of p27(Kip1), a cyclin-dependent kinase inhibitor, andneuron-like morphology in LNCaP prostate tumor cells. Biochem. Biophys. Res.Commun. 257: 609–614.
43. Stuart, R. A., A. J. Littlewood, P. J. Maddison, and N. D. Hall. 1995. Elevatedserum interleukin-6 levels associated with active disease in systemic connectivetissue disorders. Clin. Exp. Rheumatol. 13: 17–22.
44. Hirano, T., T. Matsuda, M. Turner, N. Miyasaka, G. Buchan, B. Tang, K. Sato,M. Shimizu, R. Maini, and M. Feldmann. 1988. Excessive production of inter-leukin 6/B cell stimulatory factor-2 in rheumatoid arthritis. Eur. J. Immunol. 18:1797–1801.
45. Grisius, M. M., D. K. Bermudez, and P. C. Fox. 1997. Salivary and serum in-terleukin 6 in primary Sjogren’s syndrome. J. Rheumatol. 24: 1089–1091.
46. Girschick, H. J., A. C. Grammer, T. Nanki, E. Vazquez, and P. E. Lipsky. 2002.Expression of recombination activating genes 1 and 2 in peripheral B cells ofpatients with systemic lupus erythematosus. Arthritis Rheum. 46: 1255–1263.
47. Zhang, Z., X. Wu, B. H. Limbaugh, and S. L. Bridges, Jr. 2001. Expression ofrecombination-activating genes and terminal deoxynucleotidyl transferase andsecondary rearrangement of immunoglobulin kappa light chains in rheumatoidarthritis synovial tissue. Arthritis Rheum. 44: 2275–2284.
48. Hillion, S., S. Garaud, V. Devauchelle, A. Bordron, C. Berthou, P. Youinou, andC. Jamin. 2007. Interleukin-6 is responsible for aberrant B-cell receptor-mediatedregulation of RAG expression in systemic lupus erythematosus. Immunology 122:371–380.
49. Itoh, K., E. Meffre, E. Albesiano, A. Farber, D. Dines, P. Stein, S. E. Asnis,R.A. Furie, R. I. Jain, and N. Chiorazzi. 2000. Immunoglobulin heavy chainvariable region gene replacement as a mechanism for receptor revision in rheu-matoid arthritis synovial tissue B lymphocytes. J. Exp. Med. 192: 1151–1164.
50. Armengol, M. P., M. Juan, A. Lucas-Martin, M. T. Fernandez-Figueras,D. Jaraquemada, T. Gallart, and R. Pujol-Borrell. 2001. Thyroid autoimmunedisease: demonstration of thyroid antigen-specific B cells and recombination-activating gene expression in chemokine-containing active intrathyroidal germi-nal centers. Am. J. Pathol. 159: 861–873.
6798 IL-6 AND RAGs IN MATURE B CELLS
by guest on June 13, 2013http://w
ww
.jimm
unol.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















