
Cultural transmission of predator recognition in fishes: intraspecific and interspecific learning
17
Anim. Behav., 1996, 51, 185–201 Cultural transmission of predator recognition in fishes: intraspecific and interspecific learning ALICIA MATHIS, DOUGLAS P. CHIVERS & R. JAN F. SMITH Department of Biology, University of Saskatchewan (Received 2 September 1994; initial acceptance 2 January 1995; final acceptance 19 June 1995; MS. number: 7087) Abstract. Individuals that live in groups may have the opportunity to learn to recognize unfamiliar predators by observing the fright responses of experienced individuals in the group. In intraspecific trials, naive fathead minnows, Pimephales promelas, gave fright responses to chemical stimuli from predatory northern pike, Esox lucius, when paired with pike-experienced conspecifics but not when paired with pike-naive conspecifics. These pike-conditioned minnows retained the fright responses to pike odour when tested alone, indicating that learning had occurred, and transmitted their fright responses to pike-naive minnows in subsequent trials. Brook stickleback, Culaea inconstans, are found in mixed-species aggregations with fathead minnows and are also vulnerable to predation by northern pike. In a series of interspecific tests, pike-naive brook stickleback gave fright responses to chemical stimuli from northern pike when paired with pike-experienced minnows but not when paired with pike-naive minnows. Pike-conditioned stickleback also retained the fright responses when tested alone and subsequently also transmitted the fright responses to pike-naive minnows. Individuals may benefit from observations of the fright responses of conspecifics or heterospecifics by (1) being alerted to the immediate presence of unfamiliar predators and (2) learning to recognize unfamiliar predators as a potential threat. ? 1996 The Association for the Study of Animal Behaviour Individuals in either monospecific or mixed- species groups may experience a number of advantages compared with solitary individuals. One advantage may be the opportunity for cul- tural transmission of beneficial behaviour patterns from experienced group members to younger or less experienced individuals (reviewed in Mainardi 1980; Curio 1988; Mineka & Cook 1988). Cultural transmission of information used in predator rec- ognition has been reported for a number of species. For example, predator-naive individuals of several species of birds learn to recognize pred- ators by observing the mobbing responses of conspecifics (jackdaws, Corvus monedula: Lorenz 1952; European blackbirds, Turdus merula: Curio et al. 1978; zebra finches, Taeniopygia guttata: Vieth et al. 1980). Cultural transmission of pred- ator recognition has also been reported for ducks (Klopfer 1957) and for several species of primates (Herzog & Hopf 1984; review in Mineka & Cook 1988), and has been suggested for gastropod molluscs (Wood 1968; Morgan 1972). For some fishes, predator-naive individuals may be alerted to the presence of potential pred- ators via their associations with either experienced conspecifics (Verheijen 1956; Magurran & Higham 1988) or heterospecifics (Krause 1993). The short-term consequence of such information transfer may be an increased probability of survival of the current encounter with an un- familiar predator. There may also be long-term benefits if naive individuals become conditioned to recognize the predator as dangerous in future encounters. ‘Conditioned’ individuals may sub- sequently transmit the learned information to naive individuals. Only one study has reported such cultural transmission of learned fright responses for fishes. Zebra danios, Brachydanio rerio, in one tank transmitted the recognition of a synthetic chemical (morpholine) to naive conspecifics in an adjacent tank (Suboski et al. 1990). Correspondence: A. Mathis, Department of Biology, Southwest Missouri State University, Springfield, MO 65804-0095, U.S.A. D. P. Chivers and R. J. F. Smith are at the Department of Biology, University of Saskatchewan, 112 Science Place, Saskatoon, Saskatchewan S7N 5E2, Canada. 0003–3472/96/010185+17 $12.00/0 ? 1996 The Association for the Study of Animal Behaviour 185
-
Upload
missouristate -
Category
Documents
-
view
0 -
download
0
Transcript of Cultural transmission of predator recognition in fishes: intraspecific and interspecific learning

Anim. Behav., 1996, 51, 185–201
Cultural transmission of predator recognition in fishes: intraspecific andinterspecific learning
ALICIA MATHIS, DOUGLAS P. CHIVERS & R. JAN F. SMITHDepartment of Biology, University of Saskatchewan
(Received 2 September 1994; initial acceptance 2 January 1995;final acceptance 19 June 1995; MS. number: 7087)
Abstract. Individuals that live in groups may have the opportunity to learn to recognize unfamiliarpredators by observing the fright responses of experienced individuals in the group. In intraspecifictrials, naive fathead minnows, Pimephales promelas, gave fright responses to chemical stimuli frompredatory northern pike, Esox lucius, when paired with pike-experienced conspecifics but not whenpaired with pike-naive conspecifics. These pike-conditioned minnows retained the fright responses topike odour when tested alone, indicating that learning had occurred, and transmitted their frightresponses to pike-naive minnows in subsequent trials. Brook stickleback, Culaea inconstans, are foundin mixed-species aggregations with fathead minnows and are also vulnerable to predation by northernpike. In a series of interspecific tests, pike-naive brook stickleback gave fright responses to chemicalstimuli from northern pike when paired with pike-experienced minnows but not when paired withpike-naive minnows. Pike-conditioned stickleback also retained the fright responses when tested aloneand subsequently also transmitted the fright responses to pike-naive minnows. Individuals may benefitfrom observations of the fright responses of conspecifics or heterospecifics by (1) being alerted to theimmediate presence of unfamiliar predators and (2) learning to recognize unfamiliar predators as apotential threat. ? 1996 The Association for the Study of Animal Behaviour
Individuals in either monospecific or mixed-species groups may experience a number ofadvantages compared with solitary individuals.One advantage may be the opportunity for cul-tural transmission of beneficial behaviour patternsfrom experienced group members to younger orless experienced individuals (reviewed in Mainardi1980; Curio 1988; Mineka & Cook 1988). Culturaltransmission of information used in predator rec-ognition has been reported for a number ofspecies. For example, predator-naive individualsof several species of birds learn to recognize pred-ators by observing the mobbing responses ofconspecifics (jackdaws, Corvus monedula: Lorenz1952; European blackbirds, Turdus merula: Curioet al. 1978; zebra finches, Taeniopygia guttata:Vieth et al. 1980). Cultural transmission of pred-ator recognition has also been reported for ducks
(Klopfer 1957) and for several species of primates(Herzog & Hopf 1984; review in Mineka & Cook1988), and has been suggested for gastropodmolluscs (Wood 1968; Morgan 1972).For some fishes, predator-naive individuals
may be alerted to the presence of potential pred-ators via their associations with either experiencedconspecifics (Verheijen 1956; Magurran &Higham 1988) or heterospecifics (Krause 1993).The short-term consequence of such informationtransfer may be an increased probability ofsurvival of the current encounter with an un-familiar predator. There may also be long-termbenefits if naive individuals become conditionedto recognize the predator as dangerous in futureencounters. ‘Conditioned’ individuals may sub-sequently transmit the learned information tonaive individuals. Only one study has reportedsuch cultural transmission of learned frightresponses for fishes. Zebra danios, Brachydaniorerio, in one tank transmitted the recognition ofa synthetic chemical (morpholine) to naiveconspecifics in an adjacent tank (Suboski et al.1990).
Correspondence: A. Mathis, Department of Biology,Southwest Missouri State University, Springfield,MO 65804-0095, U.S.A. D. P. Chivers and R. J. F.Smith are at the Department of Biology, Universityof Saskatchewan, 112 Science Place, Saskatoon,Saskatchewan S7N 5E2, Canada.
0003–3472/96/010185+17 $12.00/0 ? 1996 The Association for the Study of Animal Behaviour
185

The purpose of this study was two-fold. Firstwe examined whether fathead minnows,Pimephales promelas, can culturally transmit therecognition of the odour of a natural predator,northern pike, Esox lucius, between conspecifics.The second goal was to determine whether cul-tural transmission of predator recognition alsocan occur between heterospecifics. Fathead min-nows and brook stickleback, Culaea inconstans,are good candidates for tests of cultural transmis-sion, because these two species are often sympatric(e.g. Robinson & Tonn 1989), occupy similarmicrohabitats (Whitaker 1968) and share com-mon predators (Becker 1983). Northern pike oftenprey upon both fathead minnows and brookstickleback, and experienced individuals of bothspecies can recognize pike as a potential predatorbased solely on chemical cues, whereas in-experienced individuals do not (Gelowitz et al.1993; Mathis et al. 1993; Chivers & Smith 1994a).For both intraspecific and interspecific tests, wedetermined whether (1) pike-naive individuals can
receive and respond to information from pike-experienced individuals, (2) the conditioned indi-viduals retain the fright response to pike whensubsequently tested alone, and (3) conditionedindividuals can transmit the learned informationto naive individuals.
EXPERIMENTAL SERIES 1:INTRASPECIFIC CULTURAL
TRANSMISSION
Overview of Experimental Design
The test for intraspecific cultural transmissionincluded four experiments (Fig. 1). Experiment 1adocumented the differential response of exper-ienced and naive fathead minnows to pike odourand tested whether naive minnows were influencedby the behaviour of their experienced or naivepartners. Differential responses by naive minnowscould be explained by social facilitation and donot necessarily demonstrate learning by the naiveminnows. Experiment 1b tested whether the
N2N1
PairedA A
No frightresponse
No frightresponse
B Alone
No frightresponse
N3
PairedC C
No frightresponse
No frightresponse
D Alone
No frightresponse
Control Experimental
EN1
PairedA A
Frightresponse
Frightresponse
B Alone
Frightresponse
N2
PairedC C
Frightresponse
Frightresponse
D Alone
Weak frightresponse
Figure 1. Flow chart illustrating the general experimental design of the intraspecific tests. Letters by arrows indicatethe four different experiments. Results of each experiment are indicated in italics. Sizes of drawings illustrate relativesize differences between minnows. N=pike-naive minnow, E=pike-experienced minnow. Minnows were tested eitherin pairs (experiments A and C) or alone (B and D).
Animal Behaviour, 51, 1186

‘conditioned’ minnows retained their response topike odour when tested alone. A positive result inthis test would indicate cultural transmission ofinformation for predator recognition betweenexperienced and naive conspecifics. The final twoexperiments tested whether minnows can subse-quently communicate information learned via cul-tural transmission to other naive conspecifics.
Collection and Maintenance
Pike-naive fathead minnows
In May 1992, we placed artificial spawningsurfaces (broken flower-pots) in Pike Lake, anoxbow lake of the South Saskatchewan River insouth-central Saskatchewan. In June, we trans-ferred the spawning surfaces containing fatheadminnow eggs to separate aquaria in the labora-tory, where they were incubated at approximately20)C. The eggs hatched in June and July, and theminnows were fed daily with commercial fish food(Tetramin liquifry, Tetramin baby food for egglayers, and eventually Tetramin flakes). In June–August we supplemented their diet with live food(including brine shrimp, Artemia fransciscana,Daphnia and other zooplankton). We maintainedthe minnows in the laboratory on a 14:10 h light:dark cycle until April 1993 when the experimentsbegan. Laboratory-reared Pike Lake minnows donot give an anti-predator response to chemicalstimuli from pike even though they come from apopulation inhabiting a lake that contains pike(Chivers & Smith 1994a). In the autumn of 1992, wealso collected pike-naive fathead minnows fromMarshy Creek in south-central Saskatchewan.Marshy Creek is within an enclosed evaporationbasin with no piscivorous fish species. Minnowsfrom Marshy Creek do not show a fright responseto chemical stimuli from pike (Mathis et al. 1993).We maintained the minnows in a large (approxi-mately 18 000 litre) outdoor pool where they werefed daily with Tetramin flakes. In the spring of1993, we transferred the minnows to the laboratorywhere they were maintained at approximately 15)Con a 14:10 h L:D cycle. We fed the fish daily asdescribed before.
Pike-experienced fathead minnows
In the autumn of 1992, we collected pike-experienced fathead minnows from Pike Lake. We
maintained the minnows in an outdoor pool(approximately 18 000 litre), where they were feddaily with Tetramin flakes. In the spring of 1993,we transferred the minnows to the laboratorywhere they were maintained at approximately15)C on a 14:10 h L:D cycle. Minnows reared inPike Lake show a fright reaction to pike odour(Mathis et al. 1993; Chivers & Smith 1993, 1994a).
Northern pike
We collected pike from Pike Lake and housedthem individually in separate compartments of a300-litre stream tank at approximately 15)C on a14:10 h L:D cycle. We fed them one or twofathead minnows every 5 days.
Pike Stimulus Preparation
Our stimulus preparation methods were ident-ical to those of Mathis et al. (1993). Prior tostimulus collection, we fed two pike (forklengths=21 and 22 cm) approximately equal vol-umes (range 3–4 ml, measured by volumetric dis-placment in water) of swordtails, Xiphophorushelleri, for each of three feedings (i.e. once every5 days). We used a swordtail diet to eliminatealarm pheromone cues associated with the pike’sdiet of fathead minnows (Mathis & Smith1993a, b). Approximately 17 h after feeding, weremoved the two pike from their holding tanks,rinsed them with dechlorinated tap water toremove any swordtail residue from the pike’sskin and then transferred them to separate,clear plastic chambers (26#8#8 cm) thateach contained 1200 ml of tap water that hadbeen run through charcoal and sodium zeolitefilters to remove chlorine and ammonia, whichwere present in the local municipal water supply.The chambers were well aerated but containedno filtration system to avoid removing activecompounds from the water. After approxi-mately 3 days, we removed the pike from thechambers, combined the stimulus water (indi-vidual pike identity does not affect predatorrecognition abilities of prey fishes; Mathis &Smith 1993a, b; Mathis et al. 1993; Gelowitzet al. 1993) and pippetted it into either 5-mlpolypropylene containers or 20-ml plastic bags,and froze it at approximately "20)C. Thestimulus water was thawed immediately prior touse.
Mathis et al.: Cultural transmission by fishes 187

Experiment 1a: Responses of Pike-naive MinnowsPaired with Pike-experienced Minnows
Methods
To test the responses of pairs of minnows tochemical stimuli from pike, we filled a series of37-litre aquaria with dechlorinated tap water. Thetanks were aerated with an airstone located at theback of each tank. The stimulus water wasinjected into the tank via clear plastic Tygontubing tied to the airline so that the end of thetubing was approximately 1 cm from the surfaceof the airstone. The inflow of air from the airstonerapidly dispersed the stimulus water. Eachaquarium contained a centrally located shelterthat consisted of a ceramic tile (9.8#20.0 cm)mounted on three cylindrical glass legs (5.5 cmlong). We placed a partial divider (1.5#9.8 cm)along the centre of the tile to reduce aggressiveinteractions when both fish were beneath theshelter.We randomly placed 30 pike-naive minnows
(X& fork length=4.7&0.51 cm; six fish fromeach of five broods) into separate testing tanks.We paired each of 15 of these pike-naive minnowswith a pike-experienced minnow (fork length=5.6&0.22 cm; experimental treatment group), andpaired each of the other 15 pike-naive minnowswith another pike-naive minnnow (fork length=5.3&0.49 cm; control treatment group). Toenable the observer to tell the minnows apart,naive test fish within each trial were the smaller ofthe pair. The small differences in size betweenminnows in a pair does not influence the interpre-tation of the results of the experiment. Bothcontrol and experimental pike-naive subjects were‘small’ and both control (other pike-naiveminnows) and experimental (pike-experiencedminnows) partners were ‘large’. Therefore, testsubjects in both treatment groups were similarlysized and were treated identically except regardingthe experience of their partners. We tested eachpair of minnows once.We conducted the observations at a mean water
temperature of 20)C on a 14:10 h L:D cycle, afterthe minnows had remained in the observationtanks undisturbed, except for daily feedings, for 3days.Immediately prior to the beginning of each
trial, we drew 60 ml of tank water through thestimulus-injection tube by a syringe and discardedit to remove any stagnant water that might have
collected in the tubing. We drew an additional60 ml of tank water into the syringe and saved it.We filled a second syringe with 20 ml of the pikestimulus water. During each trial we first observedeach pair undisturbed for 8 min and quantifiedshelter use every 15 s by recording whether eachfish of the pair was beneath the shelter. For eachtrial, we calculated a pre-stimulus index of shelteruse as the number of observations that the focalminnow was under the shelter divided by the totalnumber of observations, with a possible range of0.0 (no shelter use) to 1.0 (always under theshelter). At the end of 8 min, we slowly (approxi-mately 1 ml/s) injected the 20 ml of pike stimuluswater into the tank, and immediately thereafterinjected 60 ml of tank water to flush the pikestimulus out of the stimulus-injection tube. Obser-vations continued for an additional 8 min, and wecalculated post-stimulus indices of shelter use foreach trial. During the post-stimulus period we alsorecorded the occurrence of two additional behav-iour patterns: (1) dashing (rapid, apparently dis-oriented swimming) and (2) freezing (the fishdrops to the bottom of the tank and remainsimmobile for a minimum of 30 s). Shelter use,dashing and freezing are components of the frightreaction of fathead minnows (e.g. Lawrence &Smith 1989; Mathis & Smith 1993c; Chivers& Smith 1995a).We recorded a number of response variables
(see above paragraph) to increase our power todetect a fright response. Fathead minnowsexposed to stimuli associated with predation mayshow a number of fright responses (e.g. increasedshelter use, dashing, freezing, decreased foraging),but not all individuals show all of these behav-ioural patterns. Furthermore, the different classesof responses are not mutually exclusive. Forexample, an individual may immediately respondto the stimulus by dashing for a few seconds andthen freeze. We interpreted the appearance of anyof these behavioural patterns as an indication ofa fright response. It is not known whether thevarious response variables represent differentlevels of intensity of a fright response (e.g. isdashing a more intense response than freezing?).For each treatment group, we compared pre-stimulus versus post-stimulus indices of shelter usewith two-tailed Wilcoxon matched-pairs signed-ranks tests (Zar 1984). Because dashing andfreezing never occurred during pre-stimulusobservations, pre-stimulus versus post-stimulus
Animal Behaviour, 51, 1188
comparisons of these behavioural patterns werenot appropriate. Instead, we directly comparedthe treatment groups for differences in theproportion of post-stimulus trials in whichdashing and freezing occurred using either thechi-squared test or the Fisher exact probabilitytest (Siegel 1956). In addition to evaluating theresponses of the control and experimental sub-jects, we also conducted the same statistical testsfor the partners of the test subjects to documentthat the two treatment groups were exposed todifferent stimuli. We used a Spearman rank-ordercorrelation to determine whether the changes inshelter use by paired minnows were positivelycorrelated.
Results
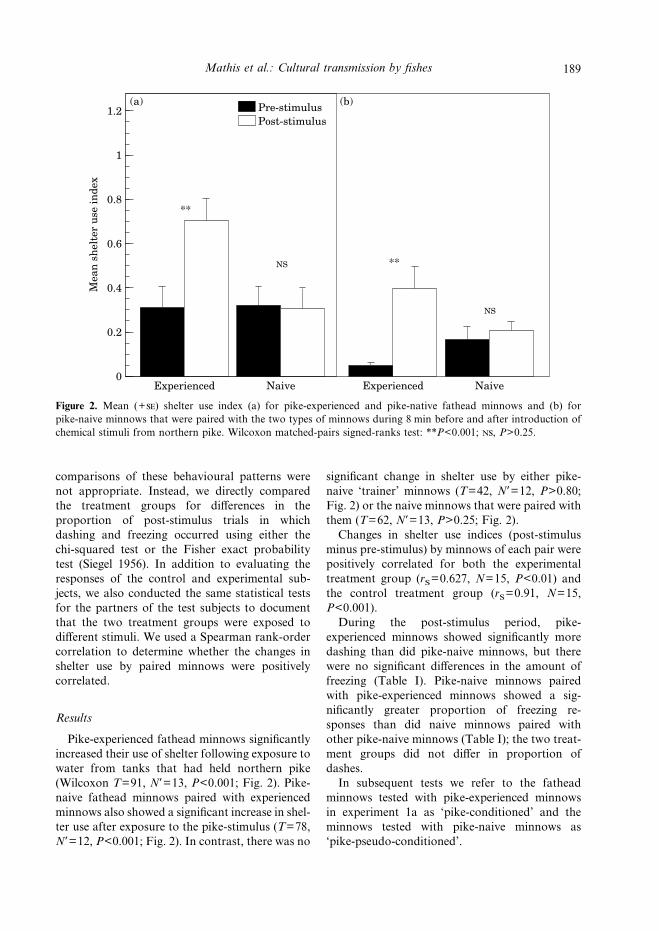
Pike-experienced fathead minnows significantlyincreased their use of shelter following exposure towater from tanks that had held northern pike(Wilcoxon T=91, N*=13, P<0.001; Fig. 2). Pike-naive fathead minnows paired with experiencedminnows also showed a significant increase in shel-ter use after exposure to the pike-stimulus (T=78,N*=12, P<0.001; Fig. 2). In contrast, there was no
significant change in shelter use by either pike-naive ‘trainer’ minnows (T=42, N*=12, P>0.80;Fig. 2) or the naive minnows that were paired withthem (T=62, N*=13, P>0.25; Fig. 2).Changes in shelter use indices (post-stimulus
minus pre-stimulus) by minnows of each pair werepositively correlated for both the experimentaltreatment group (rS=0.627, N=15, P<0.01) andthe control treatment group (rS=0.91, N=15,P<0.001).During the post-stimulus period, pike-
experienced minnows showed significantly moredashing than did pike-naive minnows, but therewere no significant differences in the amount offreezing (Table I). Pike-naive minnows pairedwith pike-experienced minnows showed a sig-nificantly greater proportion of freezing re-sponses than did naive minnows paired withother pike-naive minnows (Table I); the two treat-ment groups did not differ in proportion ofdashes.In subsequent tests we refer to the fathead
minnows tested with pike-experienced minnowsin experiment 1a as ‘pike-conditioned’ and theminnows tested with pike-naive minnows as‘pike-pseudo-conditioned’.
1.2
0Experienced
**
Mea
n s
hel
ter
use
inde
x
0.2
1
0.8
0.6
0.4
Naive
NS
Experienced
**
Naive
(a) (b)
NS
Pre-stimulusPost-stimulus
Figure 2. Mean (+) shelter use index (a) for pike-experienced and pike-native fathead minnows and (b) forpike-naive minnows that were paired with the two types of minnows during 8 min before and after introduction ofchemical stimuli from northern pike. Wilcoxon matched-pairs signed-ranks test: **P<0.001; , P>0.25.
Mathis et al.: Cultural transmission by fishes 189

Experiment 1b: Retention of the ConditionedFright Response by Fathead Minnows
Methods
The results of experiment 1a demonstratedthat pike-naive minnows show an anti-predatorresponse to chemical stimuli from pike while inthe presence of an experienced minnow perform-ing anti-predator behaviour patterns. The purposeof this experiment was to determine whether smallpike-conditioned fathead minnows from exper-iment 1a retained the fright response to chemicalstimuli from pike during subsequent tests in theabsence of large experienced conspecifics. Mathis& Smith (1993a) demonstrated that, for fatheadminnows, a fright response may be elicited by aneutral chemical stimulus (i.e. water) that is pre-sented in the same manner as a fright stimulus(chemical stimuli from pike which had eaten fat-head minnows). In that study, minnows moving toa different experimental situation eliminated thefright response to the neutral stimulus. To controlfor possible conditioning to test conditions in ourstudy, we also changed the experimental set-up inwhich the minnows were tested.One day after being used in experiment 1a, the
small pike-conditioned minnows and the small
pike-pseudo-conditioned minnows were placedindividually into Plexiglas acclimation tanks(45#45#20 cm). Water was supplied to theacclimation tanks at a constant rate of approxi-mately 250 ml/min, maintaining a water depth of4–5 cm. We fed the minnows daily as before andkept them under a 14:10 h L:D cycle. After 2 days,we transferred the minnows to separate testingtanks that were identical to the acclimationtanks except that the flow rate was increased toapproximately 450 ml/min. Testing occurred thefollowing day; 4 days passed between the timethat the minnows were tested in experiments 1aand 1b.We surrounded each testing tank with an Opto-
Varimex-Aqua tracking meter (Columbus Instru-ments) which established a grid of light beams inthe tank. The activity tracking meter was inter-faced to a computer that was preset to ‘scan’ thelight beams for breaks at intervals of 0.125 s. Thecomputer integrated and digitized informationfrom the scans to quantify the fish’s level ofactivity. A detailed description of both the accli-mation tanks and the testing apparatus is given byLemly & Smith (1986, 1987). Our system differedfrom that of Lemly & Smith’s in that outflowingwater from out tanks was discarded rather thanrecirculated. Two measures of activity that typi-cally decrease during a fright reaction by fatheadminnows (e.g. Lawrence & Smith 1989; Mathiset al. 1993) were quantified by the computer: (1)total distance travelled (cm), and (2) number ofstereotypic movements (i.e. activity in which a fishbreaks light beams without moving outside of onegrid square); many stereotypic movements wouldindicate high levels of activity even though thisactivity may not translate into linear distance.The activity meter did not recognize dashes andfreezes, so these data are not available for thesolitary trials.We conducted the experiment from an obser-
vation room that was adjacent to the test room.The observer viewed the experimental tanks viaa video monitor and injected the stimulus sol-ution into the inflow water lines that ranthrough the observation room before enteringthe test room. The design of this experiment wassimilar to that of experiment 1a. Trials lasted16 min. At the end of 8 min, the observerinjected 5 ml of pike-stimulus water from thesame source as that used in the previous exper-iment. Injections of vegetable dye into the inflow
Table I. Per cent of intraspecific trials in which minnowsshowed dashing and freezing behaviour following expo-sure to chemical stimuli from northern pike (experiment1a)
Behaviour of pike-experienced and pike-naive fatheadminnows paired with pike-naive conspecifics
Experienced(% of trials)
Naive(% of trials) P
Dashing 60 0 <0.001Freezing 0 0 —
Behaviour of pike-naive fathead minnows paired witheither pike-experienced or pike-naive conspecifics
Partner
PExperienced(% of trials)
Naive(% of trials)
Dashing 27 7 0.165Freezing 33 0 0.021
Statistical results are of Fisher exact probability tests.Percentages are based on number of trials out of 15.
Animal Behaviour, 51, 1190
lines demonstrated that it took approximately14 s for the dye to reach the experimental tanksfollowing injection.We made statistical comparisons of pre- and
post-stimulus activity using two-tailed Wilcoxonmatched-pairs signed-ranks tests (Zar 1984). Weinterpreted a significant decrease in activity as afright response; previous experiments in activitymeters demonstrated that pike-experienced (butnot pike-naive) fathead minnows responded tochemical stimuli from pike with decreased activity(Mathis et al. 1993).
Results
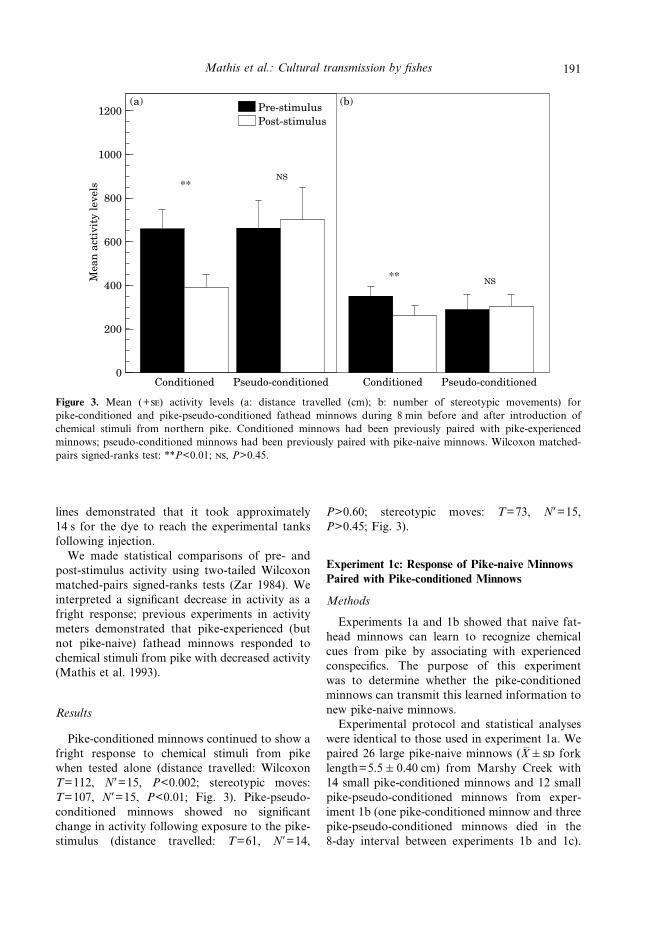
Pike-conditioned minnows continued to show afright response to chemical stimuli from pikewhen tested alone (distance travelled: WilcoxonT=112, N*=15, P<0.002; stereotypic moves:T=107, N*=15, P<0.01; Fig. 3). Pike-pseudo-conditioned minnows showed no significantchange in activity following exposure to the pike-stimulus (distance travelled: T=61, N*=14,
P>0.60; stereotypic moves: T=73, N*=15,P>0.45; Fig. 3).
Experiment 1c: Response of Pike-naive MinnowsPaired with Pike-conditioned Minnows
Methods
Experiments 1a and 1b showed that naive fat-head minnows can learn to recognize chemicalcues from pike by associating with experiencedconspecifics. The purpose of this experimentwas to determine whether the pike-conditionedminnows can transmit this learned information tonew pike-naive minnows.Experimental protocol and statistical analyses
were identical to those used in experiment 1a. Wepaired 26 large pike-naive minnows (X& forklength=5.5&0.40 cm) from Marshy Creek with14 small pike-conditioned minnows and 12 smallpike-pseudo-conditioned minnows from exper-iment 1b (one pike-conditioned minnow and threepike-pseudo-conditioned minnows died in the8-day interval between experiments 1b and 1c).
1200
0Conditioned
**
Mea
n a
ctiv
ity
leve
ls
200
1000
800
600
400
Pseudo-conditioned
NS
**
(a) (b)
NS
Pre-stimulusPost-stimulus
Conditioned Pseudo-conditioned
Figure 3. Mean (+) activity levels (a: distance travelled (cm); b: number of stereotypic movements) forpike-conditioned and pike-pseudo-conditioned fathead minnows during 8 min before and after introduction ofchemical stimuli from northern pike. Conditioned minnows had been previously paired with pike-experiencedminnows; pseudo-conditioned minnows had been previously paired with pike-naive minnows. Wilcoxon matched-pairs signed-ranks test: **P<0.01; , P>0.45.
Mathis et al.: Cultural transmission by fishes 191
We conducted tests between 1030 and 1530 hoursat approximately 20)C.
Results
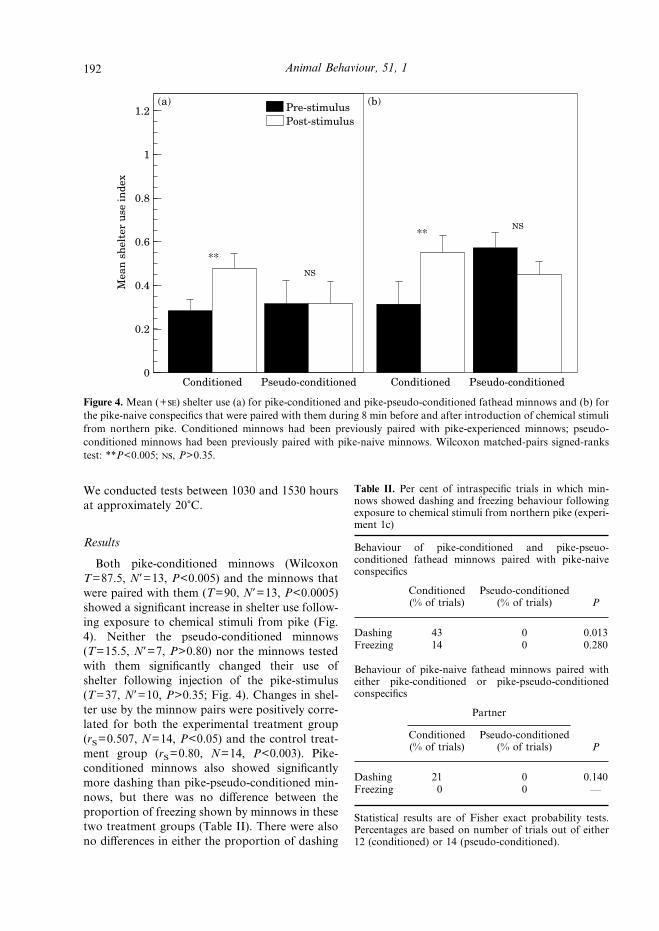
Both pike-conditioned minnows (WilcoxonT=87.5, N*=13, P<0.005) and the minnows thatwere paired with them (T=90, N*=13, P<0.0005)showed a significant increase in shelter use follow-ing exposure to chemical stimuli from pike (Fig.4). Neither the pseudo-conditioned minnows(T=15.5, N*=7, P>0.80) nor the minnows testedwith them significantly changed their use ofshelter following injection of the pike-stimulus(T=37, N*=10, P>0.35; Fig. 4). Changes in shel-ter use by the minnow pairs were positively corre-lated for both the experimental treatment group(rS=0.507, N=14, P<0.05) and the control treat-ment group (rS=0.80, N=14, P<0.003). Pike-conditioned minnows also showed significantlymore dashing than pike-pseudo-conditioned min-nows, but there was no difference between theproportion of freezing shown by minnows in thesetwo treatment groups (Table II). There were alsono differences in either the proportion of dashing
1.2
0Conditioned
**
Mea
n s
hel
ter
use
inde
x
0.2
1
0.8
0.6
0.4
Pseudo-conditioned
NS
**
(a) (b)
NS
Pre-stimulusPost-stimulus
Conditioned Pseudo-conditioned
Figure 4.Mean (+) shelter use (a) for pike-conditioned and pike-pseudo-conditioned fathead minnows and (b) forthe pike-naive conspecifics that were paired with them during 8 min before and after introduction of chemical stimulifrom northern pike. Conditioned minnows had been previously paired with pike-experienced minnows; pseudo-conditioned minnows had been previously paired with pike-naive minnows. Wilcoxon matched-pairs signed-rankstest: **P<0.005; , P>0.35.
Table II. Per cent of intraspecific trials in which min-nows showed dashing and freezing behaviour followingexposure to chemical stimuli from northern pike (experi-ment 1c)
Behaviour of pike-conditioned and pike-pseuo-conditioned fathead minnows paired with pike-naiveconspecifics
Conditioned(% of trials)
Pseudo-conditioned(% of trials) P
Dashing 43 0 0.013Freezing 14 0 0.280
Behaviour of pike-naive fathead minnows paired witheither pike-conditioned or pike-pseudo-conditionedconspecifics
Partner
PConditioned(% of trials)
Pseudo-conditioned(% of trials)
Dashing 21 0 0.140Freezing 0 0 —
Statistical results are of Fisher exact probability tests.Percentages are based on number of trials out of either12 (conditioned) or 14 (pseudo-conditioned).
Animal Behaviour, 51, 1192

or freezing shown by minnows paired with eitherconditioned and pseudo-conditioned minnows(Table II).
Experiment 1d: Retention of the ConditionedFright Response by Fathead Minnows
Methods
The results of experiment 1c indicated thatthe pike-naive fathead minnows responded topike odour with a fright response (increasedshelter use) when in the presence of pike-conditioned minnows. The purpose of thisexperiment was to determine whether the fatheadminnows from experiment 1c retained the frightresponse to chemical stimuli from pike duringsubsequent tests in the absence of the pike-conditioned minnows. The experimental protocoland statistical analyses were identical to thoseused in experiment 1b. We conducted testsbetween 0730 and 1330 hours at approximately20)C.
Results
Fathead minnows that were previously testedwith pike-conditioned fathead minnows showedno significant differences in activity in responseto pike odour when tested alone (distance trav-elled: X decrease=19%; Wilcoxon T=39, N*=14,P>0.40; stereotypic moves: X decrease=9.9%;T=33, N*=14, P>0.20). In control tests, min-nows previously tested with pike-pseudo-conditioned minnows also showed no significantchange in distance travelled (X increase=14.8%;T=21, N*=12, P>0.15), but showed a significantincrease in the number of stereotypic moves (Xincrease=24.7%; T=6, N*=12, P<0.01). Becauseof the significant increase in stereotypic move-ments for minnows in the control condition,and the tendency for stereotypic movementsto decrease for the minnows in the experimen-tal condition, we also directly compared thechange in stereotypic movements for minnows inthe two treatment groups. The change in stereo-typic movements was significantly different forminnows in control versus experimental treat-ments (Wilcoxon–Mann–Whitney tests; Siegel& Castellan 1988: Wx=150, m=14, N=12,P<0.05).
EXPERIMENTAL SERIES 2:INTERSPECIFIC CULTURAL
TRANSMISSION
Overview of Experimental Design
Tests for interspecific cultural transmission con-sisted of three experiments (Fig. 5) and generallyfollowed the protocol established in experimentalseries 1. In experiment 2a, we documented differ-ential responses of experienced and naive fatheadminnows to pike odour. In addition, this ex-periment tested whether the behaviour of theirexperienced or naive fathead minnow partnersinfluenced naive brook sticklebacks. Experiment2b tested whether the conditioned sticklebacksretained their response to pike odour when fat-head minnows were absent. Retention of frightresponses would indicate learning by the stickle-back and would indicate cultural transmission ofinformation for predator recognition betweenexperienced and naive heterospecifics. Experiment2c determined whether ‘conditioned’ sticklebackscan subsequently communicate culturally trans-mitted information to naive fathead minnows.
Collection and Maintenance
We collected pike-naive fathead minnows andbrook sticklebacks from Marshy Creek; like fat-head minnows, brook sticklebacks from MarshyCreek do not display fright responses to chemicalstimuli from northern pike (Gelowitz et al. 1993).We collected ‘experienced’ fathead minnows fromPike Lake which, as previously noted, do respondto chemical stimuli from northern pike with afright reaction. We maintained the minnows andsticklebacks in the laboratory in separate com-partments of a 300-litre artificial stream tankat approximately 15)C on a 14:10 h L:D cycle.Minnows were fed daily with Tetramin flakes andsticklebacks were fed daily with frozen brineshrimp, Artemia franciscana. Northern pike werecollected and maintained as described in theprevious tests.
Pike Stimulus Preparation
We prepared the pike water stimulus asdescribed for the previous tests, except that weused two different pike (fork lengths=22.5 and23.0 cm).
Mathis et al.: Cultural transmission by fishes 193

Experiment 2a: Response of Pike-naiveSticklebacks Paired with Pike-experiencedFathead Minnows
Methods
We tested stickleback/minnow pairs in 37-litreaquaria identical to those described for exper-iment 1a. We randomly paired each stickleback(X& total length=4.9&0.43 cm) with either apike-naive fathead minnow from Marshy Creek(fork length=4.9&0.34 cm) or a pike-experiencedfathead minnow from Pike Lake (fork length=4.8&0.50 cm) with 20 pairs in each treatmentgroup. Pairs remained in the tanks for 3 days priorto testing on the same photoperiod and feedingschedule as before. We conducted trials between0950 and 1730 hours at a mean water temperatureof 22)C.We conducted trials as described for experiment
1a. In addition to the measures of activityrecorded in experiment 1a, we also recorded thepresence or absence of feeding responses (nippingat either the surface of the water or the substrate)during the post-stimulus period. We classifiedsuch behaviour as feeding responses even thoughno food was introduced into the tanks because it
appears identical to the behaviour of the fishesduring feeding. We considered these responses tobe potentially important because they may indi-cate that the fishes respond to the novel chemicalas a feeding stimulant rather than as an indicationof danger.
Results
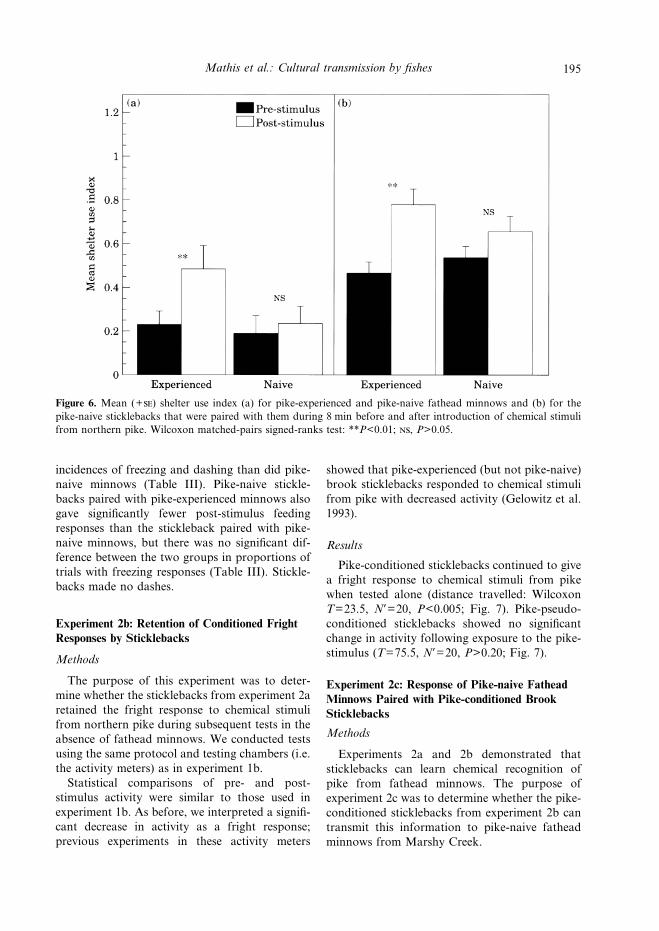
Pike-experienced fathead minnows from PikeLake significantly increased their use of shelterfollowing exposure to water from tanks that hadheld northern pike (Wilcoxon T=12, N*=14,P<0.01; Fig. 6). Pike-naive sticklebacks pairedwith pike-experienced fathead minnows also sig-nificantly increased shelter use after exposure tothe pike-stimulus (T=1, N*=11, P<0.01; Fig. 6).In contrast, there was no significant change inshelter use by either pike-naive fathead minnows(T=36.5, N*=13, P>0.50) or sticklebacks pairedwith them (T=37.4, N*=17, P>0.05) followingexposure to the pike-stimulus (Fig. 6).During the post-stimulus period, pike-
experienced fathead minnows showed significantlyfewer feeding responses and significantly more
Control Experimental
EN
PairedA A
Frightresponse
Frightresponse
B Alone
Frightresponse
N
PairedC C
Frightresponse
Frightresponse
N1N
PairedA A
No frightresponse
No frightresponse
B Alone
No frightresponse
N2
PairedC C
No frightresponse
No frightresponse
Figure 5. Flow chart illustrating the general experimental design of the interspecific tests. Letters by arrows indicatethe three different experiments. Results of each experiment are indicated in italics. N=pike-naive stickleback orfathead minnow, E=experienced minnow. Fishes were tested either in pairs (experiments A and C) or alone(experiment B).
Animal Behaviour, 51, 1194
incidences of freezing and dashing than did pike-naive minnows (Table III). Pike-naive stickle-backs paired with pike-experienced minnows alsogave significantly fewer post-stimulus feedingresponses than the stickleback paired with pike-naive minnows, but there was no significant dif-ference between the two groups in proportions oftrials with freezing responses (Table III). Stickle-backs made no dashes.
Experiment 2b: Retention of Conditioned FrightResponses by Sticklebacks
Methods
The purpose of this experiment was to deter-mine whether the sticklebacks from experiment 2aretained the fright response to chemical stimulifrom northern pike during subsequent tests in theabsence of fathead minnows. We conducted testsusing the same protocol and testing chambers (i.e.the activity meters) as in experiment 1b.Statistical comparisons of pre- and post-
stimulus activity were similar to those used inexperiment 1b. As before, we interpreted a signifi-cant decrease in activity as a fright response;previous experiments in these activity meters
showed that pike-experienced (but not pike-naive)brook sticklebacks responded to chemical stimulifrom pike with decreased activity (Gelowitz et al.1993).
Results
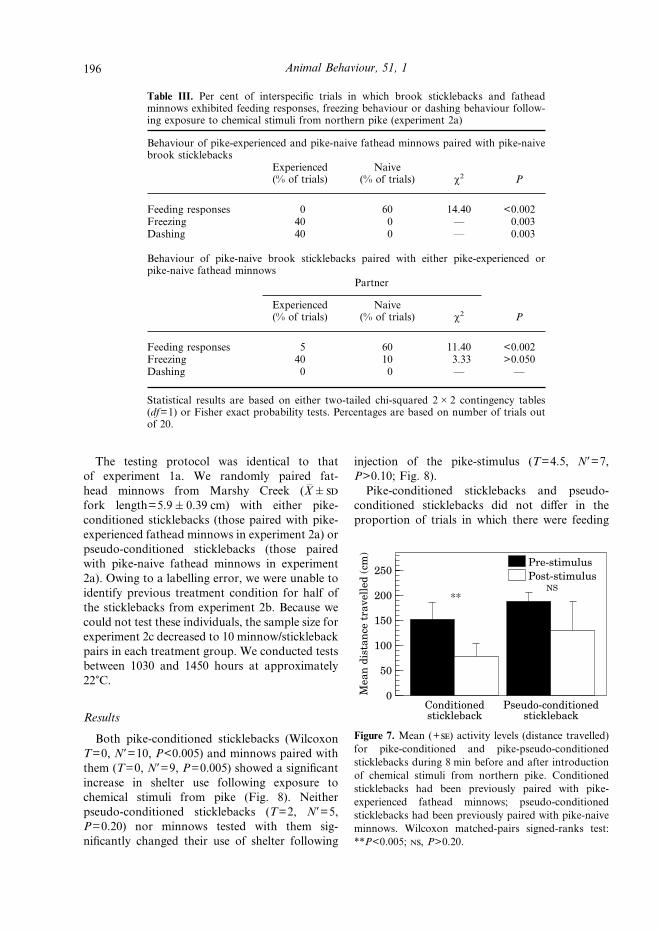
Pike-conditioned sticklebacks continued to givea fright response to chemical stimuli from pikewhen tested alone (distance travelled: WilcoxonT=23.5, N*=20, P<0.005; Fig. 7). Pike-pseudo-conditioned sticklebacks showed no significantchange in activity following exposure to the pike-stimulus (T=75.5, N*=20, P>0.20; Fig. 7).
Experiment 2c: Response of Pike-naive FatheadMinnows Paired with Pike-conditioned BrookSticklebacks
Methods
Experiments 2a and 2b demonstrated thatsticklebacks can learn chemical recognition ofpike from fathead minnows. The purpose ofexperiment 2c was to determine whether the pike-conditioned sticklebacks from experiment 2b cantransmit this information to pike-naive fatheadminnows from Marshy Creek.
Figure 6. Mean (+) shelter use index (a) for pike-experienced and pike-naive fathead minnows and (b) for thepike-naive sticklebacks that were paired with them during 8 min before and after introduction of chemical stimulifrom northern pike. Wilcoxon matched-pairs signed-ranks test: **P<0.01; , P>0.05.
Mathis et al.: Cultural transmission by fishes 195
The testing protocol was identical to thatof experiment 1a. We randomly paired fat-head minnows from Marshy Creek (X&
fork length=5.9&0.39 cm) with either pike-conditioned sticklebacks (those paired with pike-experienced fathead minnows in experiment 2a) orpseudo-conditioned sticklebacks (those pairedwith pike-naive fathead minnows in experiment2a). Owing to a labelling error, we were unable toidentify previous treatment condition for half ofthe sticklebacks from experiment 2b. Because wecould not test these individuals, the sample size forexperiment 2c decreased to 10 minnow/sticklebackpairs in each treatment group. We conducted testsbetween 1030 and 1450 hours at approximately22)C.
Results
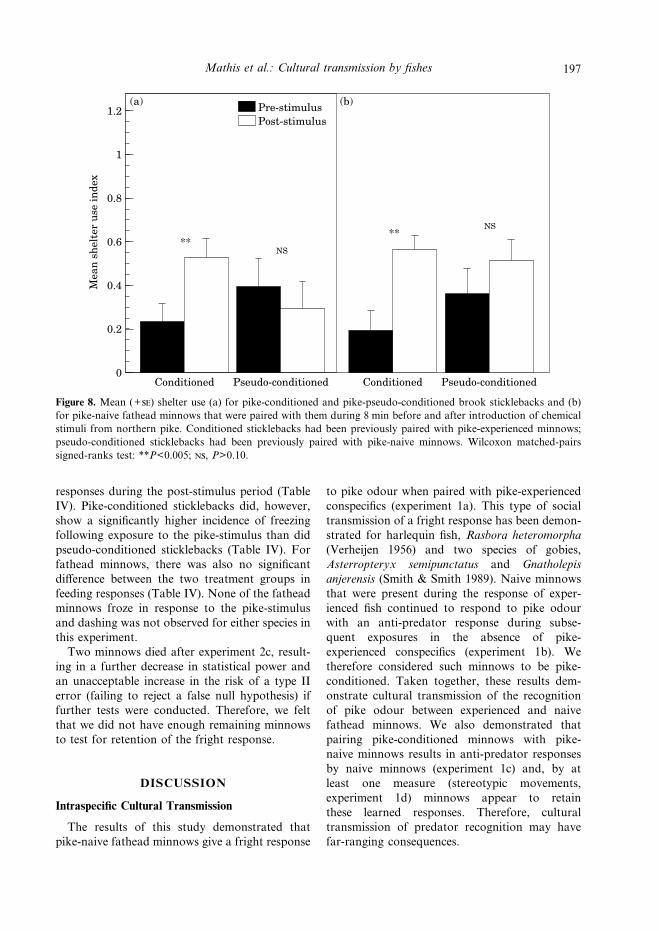
Both pike-conditioned sticklebacks (WilcoxonT=0, N*=10, P<0.005) and minnows paired withthem (T=0, N*=9, P=0.005) showed a significantincrease in shelter use following exposure tochemical stimuli from pike (Fig. 8). Neitherpseudo-conditioned sticklebacks (T=2, N*=5,P=0.20) nor minnows tested with them sig-nificantly changed their use of shelter following
injection of the pike-stimulus (T=4.5, N*=7,P>0.10; Fig. 8).Pike-conditioned sticklebacks and pseudo-
conditioned sticklebacks did not differ in theproportion of trials in which there were feeding
Table III. Per cent of interspecific trials in which brook sticklebacks and fatheadminnows exhibited feeding responses, freezing behaviour or dashing behaviour follow-ing exposure to chemical stimuli from northern pike (experiment 2a)
Behaviour of pike-experienced and pike-naive fathead minnows paired with pike-naivebrook sticklebacks
Experienced(% of trials)
Naive(% of trials) ÷2 P
Feeding responses 0 60 14.40 <0.002Freezing 40 0 — 0.003Dashing 40 0 — 0.003
Behaviour of pike-naive brook sticklebacks paired with either pike-experienced orpike-naive fathead minnows
Partner
PExperienced(% of trials)
Naive(% of trials) ÷2
Feeding responses 5 60 11.40 <0.002Freezing 40 10 3.33 >0.050Dashing 0 0 — —
Statistical results are based on either two-tailed chi-squared 2#2 contingency tables(df=1) or Fisher exact probability tests. Percentages are based on number of trials outof 20.
250
0Conditionedstickleback
**
Mea
n d
ista
nce
tra
vell
ed (
cm)
200
150
100
50
Pseudo-conditionedstickleback
NS
Pre-stimulusPost-stimulus
Figure 7. Mean (+) activity levels (distance travelled)for pike-conditioned and pike-pseudo-conditionedsticklebacks during 8 min before and after introductionof chemical stimuli from northern pike. Conditionedsticklebacks had been previously paired with pike-experienced fathead minnows; pseudo-conditionedsticklebacks had been previously paired with pike-naiveminnows. Wilcoxon matched-pairs signed-ranks test:**P<0.005; , P>0.20.
Animal Behaviour, 51, 1196
responses during the post-stimulus period (TableIV). Pike-conditioned sticklebacks did, however,show a significantly higher incidence of freezingfollowing exposure to the pike-stimulus than didpseudo-conditioned sticklebacks (Table IV). Forfathead minnows, there was also no significantdifference between the two treatment groups infeeding responses (Table IV). None of the fatheadminnows froze in response to the pike-stimulusand dashing was not observed for either species inthis experiment.Two minnows died after experiment 2c, result-
ing in a further decrease in statistical power andan unacceptable increase in the risk of a type IIerror (failing to reject a false null hypothesis) iffurther tests were conducted. Therefore, we feltthat we did not have enough remaining minnowsto test for retention of the fright response.
DISCUSSION
Intraspecific Cultural Transmission
The results of this study demonstrated thatpike-naive fathead minnows give a fright response
to pike odour when paired with pike-experiencedconspecifics (experiment 1a). This type of socialtransmission of a fright response has been demon-strated for harlequin fish, Rasbora heteromorpha(Verheijen 1956) and two species of gobies,Asterropteryx semipunctatus and Gnatholepisanjerensis (Smith & Smith 1989). Naive minnowsthat were present during the response of exper-ienced fish continued to respond to pike odourwith an anti-predator response during subse-quent exposures in the absence of pike-experienced conspecifics (experiment 1b). Wetherefore considered such minnows to be pike-conditioned. Taken together, these results dem-onstrate cultural transmission of the recognitionof pike odour between experienced and naivefathead minnows. We also demonstrated thatpairing pike-conditioned minnows with pike-naive minnows results in anti-predator responsesby naive minnows (experiment 1c) and, by atleast one measure (stereotypic movements,experiment 1d) minnows appear to retainthese learned responses. Therefore, culturaltransmission of predator recognition may havefar-ranging consequences.
1.2
0Conditioned
**
Mea
n s
hel
ter
use
inde
x
0.2
1
0.8
0.6
0.4
Pseudo-conditioned
NS
**
(a) (b)
NS
Pre-stimulusPost-stimulus
Conditioned Pseudo-conditioned
Figure 8. Mean (+) shelter use (a) for pike-conditioned and pike-pseudo-conditioned brook sticklebacks and (b)for pike-naive fathead minnows that were paired with them during 8 min before and after introduction of chemicalstimuli from northern pike. Conditioned sticklebacks had been previously paired with pike-experienced minnows;pseudo-conditioned sticklebacks had been previously paired with pike-naive minnows. Wilcoxon matched-pairssigned-ranks test: **P<0.005; , P>0.10.
Mathis et al.: Cultural transmission by fishes 197
The nature of the fright response may change asthe sequence of testing progresses. In the firstlearning trials (experiment 1a), naive minnowspaired with experienced minnows showed signifi-cantly more freezing, but not more dashing, thancontrols (Table I), but the same minnows showedthe opposite responses in experiment 1c (Table II).We do not know whether this change in behaviourreflects differences in response intensity. Compar-ing the responses (activity levels) of solitary indi-viduals, however, shows that the intensity of theresponse at the first transmission (experiment 1b)is stronger than that observed at the secondtransmission (experiment 1d). Although we sus-pect that the success of the transmission by ourminnows should eventually be eliminated, Curioet al. (1978) demonstrated that a conditionedmobbing response by blackbirds was passed alonga chain of six birds without any detectabledecrease in response intensity. The apparentrapidity of response attenuation between the firstand second transmission in our study may be
confounded by the difference in size betweenthe larger pike-naive minnows and the smallerpike-conditioned minnows. Given the general cor-relation between body size and age, and thereforeexperience, large minnows may be somewhat lessapt to learn from small minnows than from largeminnows (Dugatkin & Godin 1993).Previous studies of ostariophysan fishes have
shown that an alarm pheromone, or Schreckstoff,is given off when a predator mechanically dam-ages the skin of a prey fish (reviewed in Smith1992). Alarm pheromone released by an injuredminnow can condition predator-naive minnows torespond to chemical (Göz 1941; Magurran 1989;Chivers & Smith 1994a) and visual (Chivers &Smith 1994b) stimuli from predators. In additionto this mechanism of learned predator recog-nition, Mathis & Smith (1993a, b) showed thatalarm substance ingested by predators chemicallylabels them as predators that can then be recog-nized by naive prey. These mechanisms of pred-ator recognition, together with the mechanism ofcultural transmission demonstrated by this study,may have profound implications for animals innatural habitats. Chivers & Smith (1995b) intro-duced 10 northern pike into ponds containingapproximately 20 000 naive minnows; the min-nows learned to respond to pike odour with afright reaction within 14 days. Such a frightresponse may result in an increased probabilityof escape (Mathis & Smith 1993c). The frightresponse may also warn other prey in the vicinity(Verheijen 1956; Smith & Smith 1989; this study)and may reduce predator success and thus dis-courage future foraging in the area (Trivers 1971).Other naive individuals may also learn to recog-nize predators by being present during the frightresponse. Finally, the fright response of the indi-vidual may act directly upon the predator toinform it that its presence has been detected,making it unlikely that an attack will be successful(Högstedt 1983).
Interspecific Cultural Transmission
The results of this study show that inexper-ienced brook sticklebacks and fathead minnowsmay benefit from mixed-species associations bygaining information from heterospecifics concern-ing recognition of potentially dangerous pred-ators. For brook sticklebacks, the benefits may betwo-fold, because individuals not only are alerted
Table IV. Per cent of interspecific trials in which brooksticklebacks and fathead minnows showed feedingresponses, freezing behaviour or dashing behaviour fol-lowing exposure to chemical stimuli from northern pike(experiment 2c)
Behaviour of pike-conditioned and pike-pseudo-conditioned brook sticklebacks paired with pike-naivefathead minows
Conditioned(% of trials)
Pseudo-conditioned(% of trials) P
Feeding responses 30 40 >0.950Freezing 80 20 0.023Dashing 0 0 —
Behaviour of pike-naive fathead minnows paired witheither pike-conditioned or pike-pseudo-conditionedbrook sticklebacks
Partner
PConditioned(% of trials)
Pseudo-conditioned(% of trials)
Feeding responses 10 50 0.140Freezing 0 0 —Dashing 0 0 —
Statistical results are based on two-tailed Fisher exactprobability tests. Percentages are based on number oftrials out of 10.
Animal Behaviour, 51, 1198

to an immediate threat (experiment 2a), but mayalso learn to recognize the predator as dangerousin subsequent encounters (experiment 2b). In ourstudy, fright responses to conditioned stimuli wereretained without reinforcement for at least twoexposures after the initial learning trials, lastingfor a period of approximately 1 month. Pike-naivefathead minnows showed fright responses tochemical stimuli from pike when in the presence ofpike-conditioned sticklebacks, indicating that theinformation transfer from stickleback to minnowcan also occur (experiment 2c).Predator-naive individuals in this study gained
information concerning recognition of predatorsfrom heterospecifics. It is not known, however,whether the opportunity for predator-related cul-tural transmission is greater in mixed-speciesgroups than in monospecific groups. Althoughbrook sticklebacks and fathead minnows oftenoccupy the same microhabitat, their niches donot completely overlap. Therefore, individuals inmixed-species aggregations may be able to capi-talize on a wider range of experiences by groupmembers than individuals in strictly monospecificgroups. That is, there may be a higher probabilitythat at least one individual will be familiar with agiven predator in mixed-species shoals than insingle-species shoals.Mixed-species aggregations may also offer
other benefits over single-species groups, includ-ing increased foraging efficiency due to eithersocial learning (Krebs 1973), flushing of prey byheterospecifics (Siegfried 1971) or shared vigilance(FitzGibbon 1990), decreased competition forfood while still providing group-related protectionfrom predation (Moynihan 1962), and earlywarning of danger (Thompson & Barnard 1983;Smith & Smith 1989; Krause 1993; Mathis &Smith 1993d). Less preferred prey may benefit byassociating with prey that are more preferred ormore easily captured (FitzGibbon 1990). Thislatter benefit may apply to brook stickleback/fathead minnow shoals because the spines ofbrook sticklebacks make them less vulnerable tosome fish predators (Reist 1980a), but morevulnerable to some invertebrate predators (Reist1980b).Associating with heterospecifics instead of con-
specifics may also carry costs. One benefit ofgroup living is the confusion effect, wherebypredators find it difficult to select an individualtarget; this effect is reduced if individuals in the
group differ in size and/or appearance (Magurran1990). Furthermore, individuals that look differ-ent from other group members may be the mostvulnerable target for predators (Mueller 1971).Single-species shoals may also be more effective atpredator-evasion manoeuvres than are mixed-species groups (Allan & Pitcher 1986).The dynamics of mixed-species aggregations are
poorly understood for most taxa, and, for fishes,the costs and benefits associated with multi-species groups have only recently begun to beaddressed (Pitcher & Parrish 1993). Our studysuggests that interspecific cultural transmission ofinformation concerning predator recognition maybe an important variable in determining the adap-tive significance of mixed-species aggregations.
ACKNOWLEDGMENTS
We thank Cherie Gelowitz for assistance withcollection of the fishes. We also thank BrianWisenden, Grant Brown and Debra Lancaster forproviding helpful suggestions concerning themanuscript. Financial support was provided bythe Natural Sciences and Engineering ResearchCouncil of Canada and the University ofSaskatchewan.
REFERENCES
Allan, J. R. & Pitcher, T. J. 1986. Species segre-gation during predator evasion in cyprinid fish shoals.Freshwat. Biol., 16, 653–659.
Becker, G. C. 1983. Fishes of Wisconsin. Madison,Wisconsin: University of Wisconsin Press.
Chivers, D. P. & Smith, R. J. F. 1993. The role ofolfaction in chemosensory-based predator recognitionin the fathead minnow, Pimephales promelas. J. chem.Ecol., 19, 623–633.
Chivers, D. P. & Smith, R. J. F. 1994a. The role ofexperience and chemical alarm signalling in predatorrecognition by fathead minnows (Pimephalespromelas). J. Fish Biol., 44, 273–285.
Chivers, D. P. & Smith, R. J. F. 1994b. Fatheadminnows, Pimephales promelas, acquire predator rec-ognition when alarm substance is associated with thesight of unfamiliar fish. Anim. Behav., 48, 597–605.
Chivers, D. P. & Smith, R. J. F. 1995a. Fatheadminnows, Pimephales promelas, learn to recognizechemical stimuli from high risk habitats by the pres-ence of alarm substance. Behav. Ecol., 6, 155–158.
Chivers, D. P. & Smith, R. J. F. 1995b. Free-livingfathead minnows (Pimephales promelas) rapidly learn
Mathis et al.: Cultural transmission by fishes 199

to recognize pike (Esox lucius) as predators. J. FishBiol., 46, 949–954.
Curio, E. 1988. Cultural transmission of enemy recog-nition in birds. In: Social Learning: Psychological andBiological Perspectives. (Ed. by T. R. Zentall & B. G.Galef, Jr), pp. 75–98. Hillsdale, New Jersey: LawrenceErlbaum Associates.
Curio, E., Ernst, U. & Vieth, W. 1978. The adaptivesignificance of avian mobbing. II. Cultural trans-mission of enemy recognition in blackbirds: effective-ness and some constraints. Z. Tierpsychol., 48,184–202.
Dugatkin, L. A. & Godin, J.-G. J. 1993. Female matecopying in the guppy (Poecilia reticulata): age-dependent effects. Behav. Ecol., 4, 289–292.
FitzGibbon, C. D. 1990. Mixed-species grouping inThomson’s and Grant’s gazelles: the antipredatorbenefits. Anim. Behav., 39, 1116–1126.
Gelowitz, C. M., Mathis, A. & Smith, R. J. F. 1993.Chemosensory recognition of northern pike (Esoxlucius) by brook stickleback (Culaea inconstans):population differences and the influence of predatordiet. Behaviour, 127, 105–118.
Göz, H. 1941. Ûber den Art- und Individualgeruch beiFischen. Z. vergl. Physiol., 29, 1–45.
Herzog, M. & Hopf, S. 1984. Behavioural responsesto species specific warning calls in infant squirrelmonkeys reared in social isolation. Am. J. Primatol., 7,99–106.
Högstedt, G. 1983. Adaptation unto death: function offear screams. Am. Nat., 121, 562–570.
Klopfer, P. H. 1957. An experiment on emphatic learn-ing in ducks. Am. Nat., 91, 61–63.
Krause, J. 1993. Transmission of fright reaction betweendifferent species of fish. Behaviour, 127, 37–48.
Krebs, J. R. 1973. Social learning and the significance ofmixed-species flocks of chickadees (Parus spp.). Can.J. Zool., 51, 1275–1288.
Lawrence, B. J. & Smith, R. J. F. 1989. Behavioralresponse of solitary fathead minnows, Pimephalespromelas, to alarm substance. J. chem. Ecol., 15,209–219.
Lemly, A. D. & Smith, R. J. F. 1986. A behavioral assayfor assessing effects of pollutants on fish chemorecep-tion. Ecotoxicol. environ. Saf., 11, 210–218.
Lemly, A. D. & Smith, R. J. F. 1987. Effects of chronicexposure to acidified water on chemoreception offeeding stimuli in fathead minnows (Pimephalespromelas): mechanisms and ecological implications.Environ. toxicol. Chem., 6, 225–238.
Lorenz, K. 1952. King Solomon’s Ring. New York:Crowell.
Magurran, A. E. 1989. Acquired recognition of predatorodour in the European minnow, Phoxinus phoxinus.Ethology, 82, 216–223.
Magurran, A. E. 1990. The adaptive significance ofschooling as an anti-predator defence in fish. AnnlsZool. Fenn., 27, 51–66.
Magurran, A. E. & Higham, A. 1988. Informationtransfer across fish shoals under predator threat.Ethology, 78, 153–158.
Mainardi, D. 1980. Tradition and the social transmissionof behaviour in animals. In: Sociobiology: Beyond
Nature/Nurture? (Ed. by G. W. Barlow & J. Silverberg),pp. 227–255. Boulder, Colorado: Westview Press.
Mathis, A., Chivers, D. P. & Smith, R. J. F. 1993.Population differences in responses of fathead min-nows (Pimephales promelas) to visual and chemicalstimuli from predators. Ethology, 93, 31–40.
Mathis, A. & Smith, R. J. F. 1993a. Fathead minnows(Pimephales promelas) learn to recognize pike (Esoxlucius) as predators on the basis of chemical stimulifrom minnows in the pike’s diet. Anim. Behav., 46,645–656.
Mathis, A. & Smith, R. J. F. 1993b. Chemical labellingof northern pike, Esox lucius, by the alarm pherom-one of fathead minnows, Pimephales promelas.J. chem. Ecol., 19, 1967–1979.
Mathis, A. & Smith, R. J. F. 1993c. Chemical alarmsignals increase the survival time of fathead minnows,Pimephales promelas, during encounters with northernpike, Esox lucius. Behav. Ecol., 4, 260–265.
Mathis, A. & Smith, R. J. F. 1993d. Intraspecific andcross-superorder responses to chemical alarm signalsby brook stickleback. Ecology, 74, 2395–2404.
Mineka, S. & Cook, M. 1988. Social Learning: Psycho-logical and Biological Perspectives (Ed. by T. R.Zentall & B. G. Galef, Jr), pp. 51–74. Hillsdale, NewJersey: Lawrence Erlbaum Associates.
Morgan, P. R. 1972. The influence of prey availabilityon the distribution and predatory behaviour ofNucella lapillus (L.). J. Anim. Ecol., 41, 257–274.
Moynihan, M. 1962. The organisation and probableevolution of some mixed species flocks of neotropicalbirds. Smithson. misc. Collns, 143, 1–140.
Mueller, H. C. 1971. Oddity and specific searchingimage more important than conspicuousness in preyselection. Nature, Lond., 233, 345–346.
Pitcher, T. J. & Parrish, J. K. 1993. Functions ofshoaling behaviour in teleosts. In: Behaviour ofTeleost Fishes. 2nd edn (Ed. by T. J. Pitcher),pp. 363–439. London: Chapman & Hall.
Reist, J. D. 1980a. Selective predation upon pelvicphenotypes of brook stickleback, Culaea inconstans,by northern pike, Esox lucius. Can. J. Zool., 58,1245–1252.
Reist, J. D. 1980b. Predation upon pelvic phenotypesof brook stickleback, Culea inconstans, by selectedinvertebrates. Can. J. Zool., 58, 1253–1258.
Robinson, C. L. K. & Tonn, W. M. 1989. Influence ofenvironmental factors and piscivory in structuring fishassemblages of small Alberta lakes. Can. J. Fish.Aquat. Sci., 46, 81–89.
Siegel, S. 1956. Nonparametric Statistics for the Behav-ioural Sciences. New York: McGraw-Hill.
Siegel, S. & Castellan, N. J. 1988. Nonparametric Statis-tics for the Behavioral Sciences. 2nd edn. New York:McGraw-Hill.
Siegfried, W. R. 1971. Feeding association betweenPodiceps ruficallis and Anas smithii. Ibis, 113, 236–238.
Smith, R. J. F. 1992. Alarm signals in fishes. Rev. FishBiol. Fish., 2, 33–63.
Smith, R. J. F. & Smith, M. J. 1989. Predator-recognition behaviour in two species of gobiid fishes,
Animal Behaviour, 51, 1200

Asterropteryx semipunctatus and Gnatholepsis anjeren-sis. Ethology, 83, 19–30.
Suboski, M. D., Bain, S., Carty, A. E., McQuoid, L. M.,Seelen, M. I. & Seifert, M. 1990. Alarm reaction inacquisition and social transmission of simulated-predator recognition by zebra danio fish (Brachydaniorerio). J. comp. Psychol., 104, 101–112.
Thompson, D. B. A. & Barnard, C. J. 1983. Anti-predator responses in mixed-species associations oflapwings, golden plovers and black-headed gulls.Anim. Behav., 31, 581–593.
Trivers, R. L. 1971. The evolution of reciprocal altruism.Q. Rev. Biol., 46, 35–57.
Verheijen, F. J. 1956. Transmission of a fright reactionamongst a school of fish and the underlying sensorymechanisms. Experientia, 12, 202–204.
Vieth, W., Curio, E. & Ernst, U. 1980. The adap-tive significance of avian mobbing. III. Culturaltransmission of enemy recognition in blackbirds:cross-species tutoring and properties of learning.Anim. Behav., 28, 1217–1229.
Whitaker, J. W. 1968. Habitat selection and competitionin Saskatchewan stickleback (Gasterosteidae) duringthe ice-free season. M.Sc. thesis, University ofSaskatchewan.
Wood, L. 1968. Physiological and ecological aspects ofprey selection by the marine gastropod Urosalpinxcinerea (Prosobranchia: Muricidae). Malacologia, 6,267–320.
Zar, J. H. 1984. Biostatistical Analysis. 2nd edn.Englewood Cliffs, New Jersey: Prentice-Hall.
Mathis et al.: Cultural transmission by fishes 201




















