
Optimization of the sonication extraction method of Hibiscus tiliaceus L. flowers
Upload
independentCategory
view
1download
0
Interspecific hybridisation between Hibiscus syriacus,Hibiscus sinosyriacus and Hibiscus paramutabilis
Katrijn Van Laere ÆJohan M. Van Huylenbroeck ÆErik Van Bockstaele
Received: 16 June 2006 / Accepted: 29 November 2006 / Published online: 6 January 2007� Springer Science+Business Media B.V. 2007
Abstract Interspecific hybrids from crosses
between H. syriacus · H. paramutabilis and
H. syriacus · H. sinosyriacus were obtained. In both
cases unilateral incongruity was observed and
reciprocal crosses yielded no fruits. In vitro embryo
rescue, 11 weeks after pollination, increased the
germination capacity of H. syriacus · H. sinosyria-
cus embryos, while this was not the case for
H. syriacus · H. paramutabilis embryos. However,
a lot of the generated H. syriacus · H. sinosyriacus
seedlings were lost due to variegated and total
albinism. In the progeny of H. syriacus ‘Oiseau
Bleu’ · H. paramutabilis about 95% of the seedlings
had an intermediate flower and leaf morphology
compared to both parent plants. Leaves on the adult
F1 hybrids showed a polymorphism. In total four
different leaf types could be observed on the same
plant. This leaf polymorphism also was seen in the
progeny of H. syriacus ‘Melwhite’ · H. sinosyriacus
‘Lilac Queen’. In this progeny about 50% of the
seedlings had an intermediate flower and leaf
morphology compared with the parent plants. The
hybrid nature of the seedlings of both progenies was
also confirmed by AFLP analysis. Despite the low
germination vigour of the pollen of the hybrids, a
small F2 generation was obtained from H. syriacus
‘Oiseau Bleu’ · H. paramutabilis.
Keywords Embryo rescue � Hibiscus syriacus �Hibiscus sinosyriacus � Hibiscus paramutabilis �Interspecific hybrids � Leaf polymorphism
Introduction
Hibiscus belongs to the Malvaceae. It is a poly-
morphic genus of some 250 species of trees,
shrubs and herbs. Geographically Hibiscus is
mainly situated in tropical and subtropical areas,
with some species extending into the temperate
regions of the world i.e. Hibiscus syriacus L.
(althea or rose of Sharon), Hibiscus sinosyriacus
Bailey and Hibiscus paramutabilis Bailey (Bates
1965). These three species are native to China
(Bates 1965) and the similarity in their natural
distribution pattern is an indication of a similar
tolerance to environmental factors. Hibiscus
syriacus is the most popular species. About 40
different cultivars, with varying flower colour and
shape, are commonly in culture in Europe and a
lot more genotypes are present in different
K. Van Laere (&) � J. M. Van Huylenbroeck �E. Van BockstaeleApplied Genetics and Breeding, Institute forAgricultural and Fisheries Research (ILVO) – PlantUnit, Caritasstraat 21, 9090 Melle, Belgiume-mail: [email protected]
E. Van BockstaeleDepartment of Plant Production, Faculty ofBiosciences Engineering, Ghent University, CoupureLinks 653, 9000 Gent, Belgium
123
Euphytica (2007) 155:271–283
DOI 10.1007/s10681-006-9328-8

collections (Van De Laar 1997). It is a deciduous
shrub with more or less distinctly trilobate leaves.
Flowering is from the end of July till the first part
of October (Kim and Lee 1991). Basic chromo-
some number of Hibiscus syriacus is x = 20 and
most cultivars are tetraploid, 2n = 4x = 80 (Skov-
sted 1941). However, in literature frequently the
term diploid is used instead of tetraploid. Breeding
work resulted also in hexaploid (so called triploid)
and octaploid (so called tetraploid) cultivars,
which in general have larger flowers and a
longer flowering period (Egolf 1971, 1981; Van
Huylenbroeck et al. 2000). Hibiscus sinosyriacus,
2n = 4x = 80 (Skovsted 1941), has broader
leaves compared to Hibiscus syriacus. The leaves
have short triangular lobes and the involucratal
bracts outside the calyx are as long as the calyx or
even longer (Bates 1965). Hibiscus paramutabilis
has big round-shaped leaves and the flowers are
larger compared to H. syriacus (Bates 1965). The
chromosome number is 2n = 82 (Niimoto 1966).
Both, Hibiscus sinosyriacus and Hibiscus para-
mutabilis, are more vigorous in growth compared
to Hibiscus syriacus. The genetic relationships
among these three species and some morpho-
logical characteristics were published before
by Van Huylenbroeck et al. (2000). Also close
relationships with a few other Chinese species,
Hibiscus mutabilis and Hibiscus indicus, are
suggested (Bates 1965).
In the breeding of ornamentals, the role of
interspecific hybridisation in creating more vari-
ation is very important. When using interspecific
hybrids in backcrosses and in new cross combi-
nations hybrid vigour can be captured and traits
that do not occur within a single species can be
combined (Van Tuyl and De Jeu 1997). Inter-
specific and intergeneric hybrids can extend not
only the qualitative traits but also the quantitative
traits of the parent species. The transfer of alien
genes by sexual means requires both the produc-
tion of F1 hybrids and the subsequent fertility of
these hybrids. The difficulty of creating inter-
specific hybrids increases along with the phylo-
genetic distance between the parents (Sharma
1995). Pre- and postfertilisation barriers prevent
the production of F1 hybrid plants (Hogenboom
1973). Nevertheless, the introduction of specific
techniques such as use of mentor pollen and
grafting of the style has opened opportunities to
overcome prefertilisation barriers. Other in vitro
techniques in the form of embryo rescue and
ploidy manipulation can be used to deal with
postfertilisation barriers. All these techniques
have greatly facilitated interspecific hybridisation
in a wide variety of plant species (Eeckhaut et al.
2006).
A lot of breeding work is done in Hibiscus
syriacus. Interspecific hybridisation between
H. syriacus, H. sinosyriacus and H. paramutabilis
was reported (Kyung and Kim 2001a, b; Kyung
et al. 2001a, b). But the best hybridisation
strategy and the inheritance of morphological
traits remains unclear. Embryo rescue in intra-
specific Hibiscus syriacus crosses has already
successfully been performed (Kim et al. 1996a,
b; Paek et al. 1989a, b). Paek et al. (1989c)
reported the establishment of somatic hybridi-
sation of Hibiscus syriacus protoplasts with other
Hibiscus protoplasts, but the regeneration of
the hybridised protoplasts remains impossible.
Also some interspecific breeding work involving
more tropical species of Hibiscus is published.
Tachibana (1958) and Kuwada (1964) made
successful crosses between H. mutabilis and
H. moscheutos, but the obtained F1 seedlings
were sterile. Attempts to create interspecific
hybrids between H. syriacus and H. rosa-sinensis,
especially to introduce new flower colours and
forms, were not successful so far (Yu et al. 1976;
Paek et al. 1989c).
In this study, the objective was to evaluate the
possibilities of interspecific hybridisation within
frost resistant Hibiscus. More specific, the aim
was to introgress increased growth vigour
into Hibiscus syriacus by interspecific hybridi-
sation with Hibiscus paramutabilis and Hibiscus
sinosyriacus. The hybrid character of the obtained
seedlings was analysed by morphological para-
meters and AFLP analysis.
Materials and methods
Plant material
Different cultivars of Hibiscus syriacus [tetraploid
‘Oiseau Bleu’, ‘Melwhite’, ‘Freedom’ and
272 Euphytica (2007) 155:271–283
123

octoploid ‘Purple CV2’, ‘Red Heart CV’ (Shim
et al. 1993)] were used to make reciprocal crosses
with H. paramutabilis and H. sinosyriacus [tetra-
ploid ‘Autumn Surprise’, ‘Lilac Queen’ (Bean
1973) and ‘Melmauve’].
Interspecific crosses
Hand pollination, preceeded by emasculation,
was done from the end of July till the end of
September between 2000 and 2005, under
controlled environmental conditions in an insect
free plastic greenhouse. Pollination was
performed by rubbing anthers with releasing
pollen over a susceptible stigma of a fully
opened flower. Whenever not impeded by
various reasons, also the reciprocal crosses were
performed. An overview of the crosses is given
in Table 1.
One F1 seedling of H. syriacus ‘Red Heart
CV’ · H. paramutabilis and about 250 F1 seedlings
of H. syriacus ‘Oiseau Bleu’ · H. paramutabilis
flowered in 2004 and 2005. An F2 generation was
made of these F1 plants by self-pollination and by
back crossing of the F1 plants to the female parent
H. syriacus.
In vitro ‘embryo rescue’
Ten to eleven weeks after pollination, swollen
fruits were harvested. The seeds were sterilised in
a 10% NaOCl solution with a drop of Teepol and
rinsed three times in autoclaved water. The
embryos could be easily isolated from the ovules
and were transferred to an in vitro germination
medium containing half strength MS medium
(Murashige and Skoog 1962), 30 g l–1 sucrose and
7 g l–1 agar (pH 5.8). The media were autoclaved
(121�C, 30 min). The embryos were cultured in
petri dishes (diameter 5.5 cm, 10 ml medium
per dish, sealed with LDPE foil). After the
formation of the first true leaves, the germinated
embryos were transferred to jars containing
100 ml medium per jar. Cultures were maintained
at 23 ± 2�C under a 16 h photoperiod at
40 lMol m–2 s–1 PAR, supplied by cool white
fluorescent lamps (OSRAM L36W/31). Seedlings
were transferred to fresh medium every 6 weeks.
Three to four months after embryos were placed
on the in vitro medium, the obtained seedlings
were transferred to the greenhouse in 77-well
trays filled with peat mixture (Organic matter
20%, dry matter 25%, 1.5 kg m-3 fertiliser:
12N:14P:24K + trace elements). They were kept
in a fog unit during 3 weeks for acclimatisation.
In vivo sowing
Seeds were harvested when fully matured. They
were sown directly in 77-well trays filled with peat
mixture with the same composition as described
above.
Morphological characteristics
The progenies obtained from crosses made in
2003 were morphologically screened in the field.
Of the F1 seedlings that had intermediate
characteristics compared to the parent plants,
leaf parameters, flower colour and fertility were
determined.
Leaf parameters of these hybrid seedlings and
their parent plants were determined as described
by Van Huylenbroeck et al. (2000). The following
parameters were measured on fully developed
leaves: angle of the leaf basis (a), leaf length over
width (L/B), the relative width of the mid lob
(M/B), relative length of the side lobes (C/L) and
the relative depth of indentation of the lobes
(A/L) (see Fig. 1).
Fig. 1 Schematic overview of the measured Hibiscus leafparameters. The parameters were measured on fullydeveloped leaves: angle of the leaf basis (a), leaf lengthover width (L/B), the relative width of the mid lob (M/B),relative length of the side lobes (C/L) and the relativedepth of indentation of the lobes (A/L)
Euphytica (2007) 155:271–283 273
123

Flower colour of the plants was determined
using the Royal Horticultural Colour Chart (RHS
colours).
The fertility of both parental plants and the
F1 progenies was determined by analysing the
seed formation on the plants and the pollen
germination vigour. Therefore, pollen was
collected and incubated overnight (dark) in
a liquid medium according to Tian and Russell
(1997) with slight modifications (100 g l–1
sucrose, 100 mg l–1 H3BO3, 100 mg l–1 CaCl2,
200 mg l–1 MgSO4 � 7H2O, 100 mg l–1 KH2PO4,
pH 5.4). After incubation, the pollen was trans-
ferred to a glass slide and observed with a light
microscope (LEICA DM IRB). When the pollen
had formed a pollen tube that was twice the
diameter of the pollen, it was considered as
vigorous germinating.
DNA extraction and AFLP
Young leaf material of 27 generated H. syriacus
‘Oiseau Bleu’ · H. paramutabilis seedlings, eight
H. syriacus ‘Melwhite’ · H. sinosyriacus ‘Lilac
Queen’ and their parent plants was harvested
for AFLP analysis. DNA extraction for AFLP
analysis was based on the CTAB method by
Doyle and Doyle (1987). AFLP reactions were
run on an ABI 3130 DNA sequencer using
the commercially available kit for fluorescent
fragment detection (Perkin-Elmer 1995). EcoRI
and MseI were used for DNA digestion. Selective
amplification was done using two fluorescent
labelled EcoRI-MseI primer combinations
with six selective bases: E-AAC + M-CAT and
E-ACA + M-CTG. The number of markers
uniquely present in each of the parent plants
was checked for segregation in the offspring.
Statistical calculations and data analysis
Statistical calculations on the measurements of
the leaf indices were done using SPSS 11.5 for
Windows (one way ANOVA). If significant dif-
ferences occurred, means were separated by the
LSD (P = 0.05) method.
On the AFLP data multivariate analysis was
performed using Simple Matching Similarity
indices in SPSS 11.5 for Windows. A phyloge-
netic tree (dendrogram) was constructed using
clustering with the Unweighted Pair Group
Method with Arithmatic Mean (UPGMA)
(Sneath and Sokal 1973).
Results
In total over the different years, 590 H. syriacus
· H.paramutabilis crosses were made. The crosses
resulted in 263 fruits (44.6% successful pollina-
tions) which contained on average 13.8 seeds
(Table 1). However, in the reciprocal cross with
H. paramutabilis used as female parent, no fruits
were obtained. The success of a cross also was
cultivar dependent. When using H. syriacus ‘Red
Heart CV’ as female parent, only in 5.5% of the
pollinations fruits were obtained.
For the cross H. syriacus · H. sinosyriacus, in
total 824 syriacus flowers were pollinated. Only 27
fruits were obtained (3.3% successful pollinations).
The fruits contained on average 8.7 seeds. Also here
unilateral incongruity was observed. No fruits were
yielded in the H. sinosyriacus · H. syriacus crosses
(Table 1).
In the control crosses of H. syriacus · H. syriacus
17 fruits were obtained (9.7% successful pollinations)
which contained on average 18.6 seeds.
In vitro embryo rescue versus in vivo sowing
A comparison in efficiency between in vitro
‘embryo rescue’ and in vivo sowing was made for
each cross combination (Tables 2, 3). In general
in vivo sowing resulted in more plantlets for
H. syriacus · H. paramutabilis crosses. The
germination rate of the seeds was 42.8% in vitro
versus 80.7% in vivo. However, for the cross
H. syriacus · H. sinosyriacus the in vitro germi-
nation rate of the embryos was 87.4% while the
in vivo germination rate was 52.0%. For
the control cross of H. syriacus · H. syriacus
no difference in germination capacity was
observed between both sowing methods. Germi-
nation of the seeds was 78.2% in vitro and
73.1% in vivo.
A lot of the in vitro obtained seedlings were
lost during acclimatisation due to total and
274 Euphytica (2007) 155:271–283
123

variegated albinism and to growth aberrations. Of
the in vitro obtained H. syriacus · H. paramuta-
bilis seedlings 83.6% could be planted in the field
versus 91.2% of the in vivo seedlings. For the H.
syriacus · H. sinosyriacus seedlings this was
34.5% versus 75.6% respectively. In the control
cross finally 73.3% of the in vitro obtained
seedlings and 88.4% of the in vivo seedlings were
planted on the field (Tables 2, 3).
In general no genotype effect was present for
germination and acclimatisation rates.
Morphological analysis
The seedlings obtained from crosses made in 2003
were morphologically screened in the field. Only in
the progenies of H. syriacus ‘Oiseau Bleu’ ·H. paramutabilis and H. syriacus ‘Melwhite’ ·H. sinosyriacus ‘Lilac Queen’ seedlings with inter-
mediate characteristics were found (Table 4). Of
the F1 progeny from H. syriacus ‘Oiseau Bleu’ ·H. paramutabilis 95.3% grew very vigorous and
seemed to have intermediate characteristics com-
Table 1 Overview of the different interspecific crosses, obtained fruits and seeds between Hibiscus syriacus (H.s.), Hibiscussinosyriacus (H.si.) and Hibiscus paramutabilis during different years
Yearofcross
Parentage No. ofpollinatedflowers
No. offruits
Successfulpollinations(%)
No. ofseeds
No. ofseeds perfruit$ #
2000 H.s. ‘Oiseau Bleu’ H. paramutabilis 41 7 17.0 45 6.4H.s. ‘Red Heart CV’ H. paramutabilis 48 5 10.4 26 5.2H. paramutabilis H.s.’Oiseau Bleu’ 2 –a 0.0 – –H. paramutabilis H.s. ‘Red Heart CV’ 5 – 0.0 – –
2003 H.s. ‘Oiseau Bleu’ H. paramutabilis 62 38 61.2 766 20.12004 H.s. ‘Melwhite’ H. paramutabilis 63 50 79.4 485 9.7
H.s. ‘Oiseau Bleu’ H. paramutabilis 99 79 79.8 877 11.1H.s. ‘Purple CV2’ H. paramutabilis 47 27 57.4 300 11.1H.s. ‘Red Heart CV’ H. paramutabilis 42 – 0.0 – –H. paramutabilis H.s. ‘Melwhite’ 29 – 0.0 – –H. paramutabilis H.s. ‘Oiseau Bleu’ 10 – 0.0 – –H. paramutabilis H.s. ‘Purple CV2’ 8 – 0.0 – –H. paramutabilis H.s. ‘Red Heart CV’ 7 – 0.0 – –
2005 H.s. ‘Freedom’ H. paramutabilis 188 57 30.3 1145 20.12003 H.s. ‘Oiseau Bleu’ H.si. ‘Melmauve’ 80 4 5.0 29 7.3
H.s. ‘Oiseau Bleu’ H.si. ‘Lilac Queen’ 115 2 1.7 24 12H.s. ‘Melwhite’ H.si. ‘Melmauve’ 60 – 0.0 – –H.s. ‘Melwhite’ H.si. ‘Lilac Queen’ 158 12 2.5 82 6.8H.s. ‘Red Heart CV’ H.si. ‘Lilac Queen’ 75 – 0.0 – –H.s. ‘Purple CV2’ H.si. ‘Lilac Queen’ 88 7 8.0 94 13.4
2004 H.s. ‘Melwhite’ H.si. ‘Autumn Surprise’ 11 – 0.0 – –H.s. ‘Purple CV2’ H.si. ‘Autumn Surprise’ 14 2 14.3 8 4H.s. ‘Melwhite’ H.si. ‘Lilac Queen’ 52 – 0.0 – –H.s. ‘Oiseau Bleu’ H.si. ‘Lilac Queen’ 61 – 0.0 – –H.s. ‘Purple CV2’ H.si. ‘Lilac Queen’ 69 – 0.0 – –H.s. ‘Red Heart CV’ H.si. ‘Lilac Queen’ 41 – 0.0 – –H.si. ‘Autumn Surprise’ H.s. ‘Melwhite’ 34 – 0.0 – –H.si. ‘Autumn Surprise’ H.s. ‘Oiseau Bleu’ 37 – 0.0 – –H.si. ‘Autumn Surprise’ H.s. ‘Purple CV2’ 39 – 0.0 – –H.si. ‘Autumn Surprise’ H.s. ‘Red Heart CV’ 56 – 0.0 – –H.si. ‘Lilac Queen’ H.s. ‘Melwhite’ 10 – 0.0 – –H.si. ‘Lilac Queen’ H.s. ‘Oiseau Bleu’ 3 – 0.0 – –H.si. ‘Lilac Queen’ H.s. ‘Purple CV2’ 38 – 0.0 – –H.si. ‘Lilac Queen’ H.s. ‘Red Heart CV’ 47 – 0.0 – –
2003 H.s. ‘Oiseau Bleu’ H.s. ‘Melwhite’ 61 7 11.5 150 21.4H.s. ‘Purple CV2’ H.s. ‘Melwhite’ 88 6 6.8 115 19.1
2004 H. s. ‘Oiseau Bleu’ H. s. ‘Melwhite’ 27 4 14.8 51 12.8
a No fruit set was observed
Euphytica (2007) 155:271–283 275
123
pared to the parent plants. Morphology of the
other 4.7% of the progeny was the same as H.
syriacus and these seedlings were considered to be
the result of a self-pollination of H. syriacus. Of the
F1 progeny from H. syriacus ‘Melwhite’ · H.
sinosyriacus ‘Lilac Queen’ 52.9% of the seedlings
had intermediate characteristics compared to the
parent plants. These seedlings also grow very
vigorous. The other 47.1% of the seedlings looked
morphologically like the mother parent H. syriacus
‘Melwhite’ and were the result of a self-pollination
of H. syriacus.
Leaf morphology of the H. syriacus ‘Oiseau
Bleu’ · H. paramutabilis seedlings (Table 5)
changed since F1 hybrids became older. The
1-year-old hybrids all showed a uniform inter-
mediate leaf type 1 with L/B comparable to
H. paramutabilis and C/L the same as H. syriacus
(Table 5, Fig. 2). The leaf angle and the relative
width of the mid lob (M/B ratio) were higher than
both parent plants. The indentation of the lobes
(A/L) was intermediate. When the hybrid seed-
lings were 2-years-old, two types of leaves could
be distinguished on the same plant. The first type
had a same M/B, A/L and C/L ratio compared
with the leaves of a 1-year-old seedling. They had
a larger L/B ratio and a smaller leaf angle. The
second leaf type was more indented than the first
one as was expressed by a lower A/L and M/B
ratio. Hybrids that were 3-years-old showed four
different types of leaves (Table 5, Fig. 2). Type 1
and type 2 leaves were statistically the same as
type 1 and type 2 leaves of a 2-year-old seedling.
Type 3 leaves were again more indented than
type 2 leaves as shown in the A/L, C/L and M/B
ratios. The 4th leaf type was asymmetric, as it was
indented only at one side (see Fig. 2). The side
that was not indented expressed a same A/L and
C/L ratio as type 1 leaves, while the A/L ratio
of the indented side was lower than for H.
syriacus. Also on the 3-year-old hybrid seedlings
of H. syriacus ‘Melwhite’ · H. sinosyriacus ‘Lilac
Queen’ the same four different types of leaves
could be distinguished on the same plant (Fig. 2).
Type 1 leaves had an L/B ratio, leaf angle and
indentation comparable with H. sinosyriacus, the
M/B ratio was intermediate. Indentation in-
creased in type 2 and type 3 leaves, while the
4th leaf type was indented only at one side.
Compared to both parent plants, the F1 hybrid
seedlings of H. syriacus ‘Oiseau Bleu’ · H. para-
mutabilis had bigger flowers, while the flower colour
was intermediate (Table 6). Flower colours of all
the F1 hybrids belonged, according to the RHS-
charts, to the purple group. Within this group, three
subgroups could be observed: 48.5% of the seed-
lings belonged to RHS purple group 76A, 24.5% to
RHS purple group 76B and 27.0% to RHS purple
group 76D (Table 6). So far, only seven F1 hybrid
seedlings of H. syriacus ‘Melwhite’ · H. sinosyria-
cus ‘Lilac Queen’ flowered. Flowers of these hybrids
were also bigger compared to their parents and
flower colours were intermediate (Table 7).
Molecular analysis
For the AFLP analysis of H. syriacus ‘Oiseau
Bleu’ · H. paramutabilis, leaf material of 27 F1
seedlings was harvested. Among them, two plants
(OISPAR-3 and OISPAR-20) that looked mor-
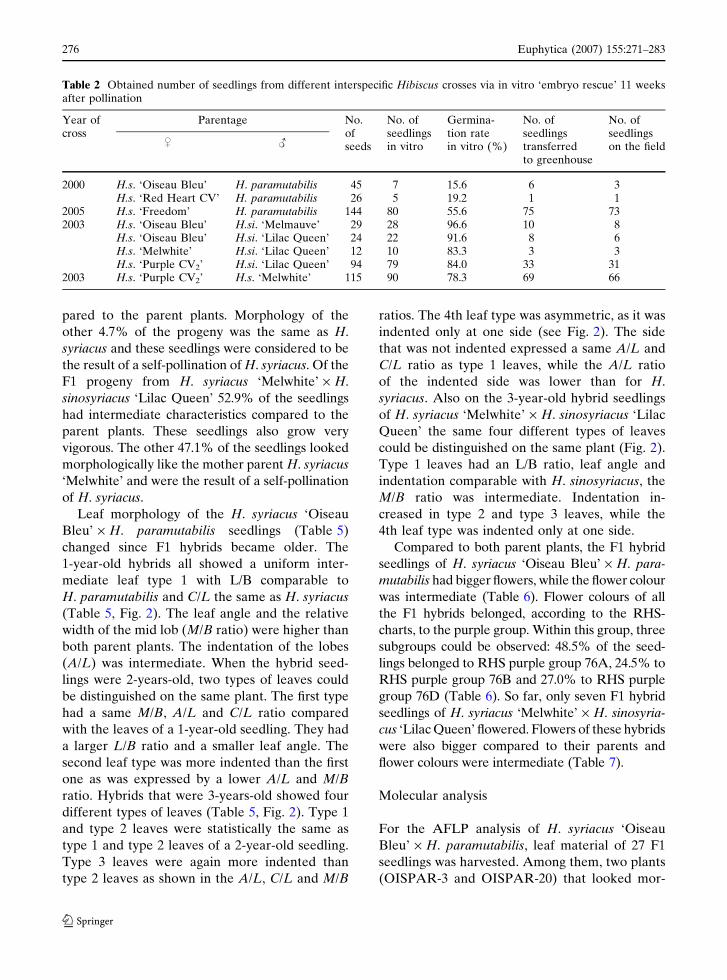
Table 2 Obtained number of seedlings from different interspecific Hibiscus crosses via in vitro ‘embryo rescue’ 11 weeksafter pollination
Year ofcross
Parentage No.ofseeds
No. ofseedlingsin vitro
Germina-tion ratein vitro (%)
No. ofseedlingstransferredto greenhouse
No. ofseedlingson the field$ #
2000 H.s. ‘Oiseau Bleu’ H. paramutabilis 45 7 15.6 6 3H.s. ‘Red Heart CV’ H. paramutabilis 26 5 19.2 1 1
2005 H.s. ‘Freedom’ H. paramutabilis 144 80 55.6 75 732003 H.s. ‘Oiseau Bleu’ H.si. ‘Melmauve’ 29 28 96.6 10 8
H.s. ‘Oiseau Bleu’ H.si. ‘Lilac Queen’ 24 22 91.6 8 6H.s. ‘Melwhite’ H.si. ‘Lilac Queen’ 12 10 83.3 3 3H.s. ‘Purple CV2’ H.si. ‘Lilac Queen’ 94 79 84.0 33 31
2003 H.s. ‘Purple CV2’ H.s. ‘Melwhite’ 115 90 78.3 69 66
276 Euphytica (2007) 155:271–283
123

phologically like the mother parent H. syriacus
were sampled. The other 25 plants had intermedi-
ate morphological characteristics as described
above. The two primers generated in total 311
marker bands. The two crossing parents appeared
to be very polymorphic. Hibiscus paramutabilis
showed 80 completely unique markers, while
H. syriacus ‘Oiseau Bleu’ could be characterised
by 75 unique markers (Table 8). Of the 27 tested F1
seedlings 25 showed an equal share of markers that
could be traced back to each of the crossing parents
indicating the true hybrid nature of the offspring
(Table 8). In the other two seedlings (OISPAR-3
and OISPAR-20) almost no unique markers of the
male parent inherited, proving that those two
seedlings were self-pollinations of H. syriacus.
Figure 3 gives an overall view of the genetic
relationships between the plants analysed by
AFLP markers. The group of hybrids clearly was
genetically different from both H. syriacus and
Table 3 Obtained number of seedlings from different interspecific Hibiscus crosses after in vivo sowing
Year ofcross
Parentage No. ofseeds
No. ofseedlingsin vivo
Germinationratein vivo (%)
No. ofseedlingson the field$ #
2003 H.s. ‘Oiseau Bleu’ H. paramutabilis 766 660 86.2 6412004 H.s. ‘Melwhite’ H. paramutabilis 485 334 68.9 226
H.s. ‘Oiseau Bleu’ H. paramutabilis 877 705 80.4 592H.s. ‘Purple CV2’ H. paramutabilis 300 298 99.3 298
2005 H.s. ‘Freedom’ H. paramutabilis 1,001 772 77.1 7692003 H.s. ‘Melwhite’ H.si. ‘Lilac Queen’ 70 40 57.1 312004 H.s. ‘Purple CV2’ H.si. ‘Autumn Surprise’ 8 1 12.5 –2003 H.s. ‘Oiseau Bleu’ H.s. ‘Melwhite’ 150 136 90.6 1262004 H.s. ‘Oiseau Bleu’ H.s. ‘Melwhite’ 51 11 21.6 4
Table 4 Generalmorphological analysis ofthe F1 seedlings obtainedafter interspecific crossesin 2003
Parentage No. of plantson the field
No. of plants withintermediatecharacteristics
No. of plantswithH. syriacuscharacteristics
$ #
H. s. ‘Oiseau Bleu’ H. paramutabilis 641 611 30H. s. ‘Oiseau Bleu’ H. si. ‘Melmauve’ 8 0 8H. s. ‘Oiseau Bleu’ H. si. ‘Lilac Queen’ 6 0 6H. s. ‘Melwhite’ H. si. ‘Lilac Queen’ 34 18 16H. s. ‘Purple CV2’ H. si. ‘Lilac Queen’ 31 0 31
Table 5 Leaf indices for H. syriacus ‘Oiseau Bleu’,H. paramutabilis and H. syriacus ‘Oiseau Bleu’ · H. para-mutabilis F1 hybrids (crosses in 2003) for different plant
ages and leaf types (n = 10). (C1, A1: measured at mostindented site ; C2, A2: measured at less indented site)
Leaftype L/B a M/B C1/L A1/L C2/L A2/L
H. syriacus ‘Oiseau Bleu’ 1.41 e z 112 a 0.27 b 0.55 ab 0.40 c 0.56 bc 0.41 bH. paramutabilis 0.88 ab 242 d 0.43 d 0.70 d 0.64 f 0.71 d 0.65 gF1 (1-year-old) Type 1 0.92 bc 274 e 0.50 ef 0.51 a 0.49 de 0.52 ab 0.49 deF1 (2-year-old) Type 1 1.04 d 190 b 0.52 f 0.56 b 0.53 e 0.55 bc 0.53 f
Type 2 0.92 bc 196 bc 0.35 c 0.62 c 0.47 d 0.59 c 0.47 cdF1 (3-year-old) Type 1 0.98 cd 201 bc 0.47 de 0.55 ab 0.53 e 0.56 bc 0.53 ef
Type 2 0.91 b 212 c 0.34 c 0.56 b 0.41 c 0.57 c 0.43 bcType 3 0.82 a 236 d 0.18 a 0.48 a 0.25 a 0.49 a 0.24 aType 4 0.92 bc 192 b 0.38 c 0.61 c 0.32 b 0.58 c 0.53 ef
z Means indicated by the same symbol are not statistically different (LSD 0.05)
Euphytica (2007) 155:271–283 277
123

H. paramutabilis. The hybrids were more related to
each other than to one of the crossing parents. Only
the two hybrids that were considered to be the
result of a self-pollination of H. syriacus, were
indeed clustered close to H. syriacus ‘Oiseau Bleu’.
For the AFLP analysis of H. syriacus
‘Melwhite’ · H. sinosyriacus ‘Lilac Queen’, leaf
material of eight F1 seedlings was harvested
from which two plants (MELLILAC-2 and
MELLILAC-6) looked morphologically like the
mother parent H. syriacus. Also here, in the
intermediate looking seedlings enough unique
markers could be found from both parent plants
(Table 9). This indicated the true hybrid nature of
these plants. The two seedlings MELLILAC-2 and
MELLILAC-6 had no unique markers of the male
parent inherited, as was expected. Clustering of the
seedlings as presented in a dendrogram (Fig. 4)
demonstrated also the hybrid and self-pollinated
origin of the seedlings.
F2 generation of H. syriacus ·H. paramutabilis
Only 1.9% of the pollen of the H. syriacus ‘Oiseau
Bleu’ · H. paramutabilis hybrids germinated in
vitro, while H. syriacus pollen had an in vitro
germination rate of 56.0% and H. paramutabilis
43.0%. Despite this low germination vigour of
the hybrid pollen, a small F2 generation was
obtained from both H. syriacus ‘Oiseau Bleu’ ·H. paramutabilis hybrids and H. syriacus ‘Red
Heart CV’ · H. paramutabilis hybrids. After in
vitro ‘embryo rescue’, three F2 seedlings resulting
from self-crosses of H. syriacus ‘Red Heart
CV’ · H. paramutabilis and four F2 seedlings
of H. syriacus ‘Oiseau Bleu’ · H. paramutabilis
could be acclimatised. Also two F2 plants from
backcrosses of H. syriacus ‘Red Heart CV’ · (H.
syriacus ‘Red Heart CV’ · H. paramutabilis) were
obtained (Table 10).
Fig. 2 Leaf morphology of the four different leaf types ofthe 3-year-old H. syriacus ‘Oiseau Bleu’ · H. paramutabilisand H. syriacus ‘Melwhite’ · H. sinosyriacus ‘Lilac Queen’hybrids compared to the leaf morphology of the parent plants
Table 6 Flower characteristics of H. syriacus ‘Oiseau Bleu’, H. paramutabilis and the H. syriacus ‘Oiseau Bleu’ · H.paramutabilis F1 hybrids obtained after interspecific crosses in 2003
Petala Basal spot on thepetals
Percentage ofseedlings
Diameterflowers (cm)b
Type offlower
H. syriacus ‘Oiseau Bleu’ Violet–purple group, 93B Red–purple group, 71A 10.5 a SingleH. paramutabilis 56D (white) Red group, 45A 14.0 b SingleF1 Purple group, 76A Red–purple group, 60A 48.5 16.0 c Single
Purple group, 76B Red–purple group, 60B 24.5 16.0 c SinglePurple group, 76D Red–purple group, 60B 27.0 16.0 c single
a Colour was determined by using the Royal Horticultural Society Colour Chartb Means indicated by the same symbol are not statistically different (LSD 0.05)
278 Euphytica (2007) 155:271–283
123

Discussion
Our objective was to evaluate the possibilities
of interspecific hybridisation within frost
resistant Hibiscus. Interspecific hybrids between
H. syriacus, H. paramutabilis and H. sinosyria-
cus were obtained. However crossing compati-
bility was unilateral and only successful when
H. syriacus was used as the mother parent. The
success rate of a cross was also cultivar depen-
dent. In our study, H. syriacus ‘Red Heart CV’
was less successful. Differences in compatibili-
ties in crosses among different Hibiscus species
were also reported by Kyung and Kim (2001a,
b). Unilateral incongruity has been described
in the H. mutabilis · H. moscheutos crosses
Table 7 Flower characteristics of H. syriacus ‘Melwhite’, H. sinosyriacus ‘Lilac Queen’ and the H. syriacus ‘Melwhite’ · H.sinosyriacus ‘Lilac Queen’ F1 hybrids obtained after interspecific crosses made in 2003
Petalsa Basal spot on the petals No.ofseedlings
Diameterflowers(cm)b
Type offlower
H. syriacus ‘Melwhite’ Purple group 76A–76D Red–purple group 60A 8 a SingleH. sinosyriacus ‘Lilac Queen’ 56D (White) / 11 b SingleF1 Purple group, 76A Red–purple group, 60A 3/7 15.2 c Single
Purple group, 76B Red–purple group, 60B 3/7 15.2 c SinglePurple group, 76D Red–purple group, 60B 1/7 15.2 c single
a Colour was determined by using the Royal Horticultural Society Colour Chartb Means indicated by the same symbol are not statistically different (LSD 0.05)
Table 8 Distribution ofAFLP––markers that areunique to one of thecrossing parents afterinterspecific H. syriacus‘Oiseau Bleu’ · H.paramutabilis crossesmade in 2003
Plant No. of markers segregating for H.syriacus parent
No. of markers segregating forH. paramutabilis parent
Unique to H. s.parent
In offspringplant
Unique to H. p.parent
In offspringplant
OISPAR-1 75 60 80 57OISPAR-2 75 60 80 60OISPAR-3 75 69 80 3OISPAR-4 75 60 80 61OISPAR-5 75 55 80 63OISPAR-6 75 57 80 63OISPAR-7 75 60 80 57OISPAR-8 75 59 80 61OISPAR-9 75 57 80 54OISPAR-10 75 55 80 54OISPAR-11 75 58 80 55OISPAR-12 75 54 80 64OISPAR-13 75 57 80 55OISPAR-14 75 57 80 64OISPAR-15 75 54 80 58OISPAR-16 75 57 80 63OISPAR-17 75 56 80 61OISPAR-18 75 54 80 65OISPAR-19 75 58 80 64OISPAR-20 75 72 80 8OISPAR-21 75 58 80 63OISPAR-22 75 57 80 61OISPAR-23 75 55 80 59OISPAR-24 75 56 80 59OISPAR-25 75 56 80 50OISPAR-26 75 57 80 55OISPAR-27 75 58 80 63
Euphytica (2007) 155:271–283 279
123

Fig. 3 Dendrogramshowing the geneticrelatedness of 27 H.syriacus ‘OiseauBleu’ · H. paramutabilisseedlings and the parentplants tested by AFLP
Table 9 Distribution ofAFLP––markers that areunique to one of the crossingparents after interspecificH. syriacus‘Melwhite’ · H. sinosyriacus‘Lilac Queen’ crosses madein 2003
Plant No. of markers segregating forH. syriacus parent
No. of markers segregating forH. sinosyriacus parent
Unique toH. s. parent
In offspring plant Unique toH. si. parent
In offspring plant
MELLILAC-1 47 31 73 55MELLILAC-2 47 43 73 1MELLILAC-3 47 30 73 52MELLILAC-4 47 33 73 51MELLILAC-5 47 35 73 55MELLILAC-6 47 40 73 0MELLILAC-7 47 37 73 52MELLILAC-8 47 31 73 53
280 Euphytica (2007) 155:271–283
123

of Kuwada (1964) and Tachibana (1958) and
in many other species, like Lilium (Van Tuyl et al.
1991) and Bromeliaceae (Vervaeke et al. 2001).
In our study in vitro embryo rescue was
compared with direct in vivo sowing of the seeds
in the greenhouse, since it was possible for the
fruits to stay on the plant until they where fully
matured. For the embryo rescue, the embryos
were isolated 10–11 weeks after pollination. They
had developed by then to the torpedo stage or in
some cases even to the cotyledonairy stage and
could easily be isolated from the ovules. So it was
possible to perform embryo culture instead of
ovule culture. Embryo rescue in H. syriacus was
already successfully performed following intra-
specific tetraploid (so called diploid) · octoploid
(so called tetraploid) pollination (Kim et al.
1996a, b). Embryo culture also was applied in
many other ornamental species where pollinated
flowers could remain on the plant for a notable
time as in Alstroemeria (Buitendijk et al. 1992)
and Lilium (Van Tuyl et al. 1991). In our study
both in vitro embryo rescue and in vivo sowing
were successful, since for the two methods F1
seedlings were obtained. Although higher amount
of seedlings from H. syriacus · H. sinosyriacus
crosses could be generated in vitro, a lot of them
were lost during acclimatisation due to albinism
and growth aberrations. This phenomenon of
chlorosis is frequently observed in interspecific
crosses as in Rhododendron (Michishita et al.
2002) and Zantedeschia (Yao and Cohen 2000). A
logical approach towards this problem would be
to perform the reciprocal cross. Our experimental
setup already anticipated to this problem since
reciprocal crosses were performed whenever
possible. However, those were hampered by
unilateral incongruity.
Morphological screening of the F1 seedlings on
the field together with AFLP analysis revealed that
most of the in vitro and in vivo H. syriacus ‘Oiseau
Bleu’ · H. paramutabilis seedlings were true
interspecific hybrids. But of the progeny obtained
from H. syriacus ‘Melwhite’ · H. sinosyriacus ‘Lilac
Queen’ crosses only about 50% of the obtained
seedlings were true hybrids as proven by morpho-
logical and genetic analysis. The other 50% of the
seedlings as well as all acclimatised seedlings from
other H. syriacus · H. sinosyriacus cross
combinations were self-pollinations of H. syriacus.
Fig. 4 Dendrogramshowing the geneticrelatedness of eight H.syriacus ‘Melwhite’ · H.sinosyriacus ‘Lilac Queen’seedlings and the parentplants tested by AFLP
Table 10 Self-crosses and backcrosses of the H. syriacus · H. paramutabilis F1 hybrids and the generated F2 seedlings.(REDPAR: H.s. ‘Red Heart CV’ · H. paramutabilis; OISPAR: H.s. ‘Oiseau Bleu’ · H. paramutabilis)
Cross No. ofpollinations
No. offruits
Success ofpollination (%)
No. ofseeds
Seedsper fruit
No. ofseedlings invitro
No. of seedlings ingreenhouse
REDPAR · REDPAR 274 1 0.36 14 14.0 4 3OISPAR · OISPAR 708 15 2.12 29 1.9 5 4‘Oiseau
Bleu’ · OISPAR337 1 0.30 2 2.0 – –
‘Red HeartCV’ · REDPAR
249 3 1.20 21 7.0 2 2
Euphytica (2007) 155:271–283 281
123

Probably these self-pollinations were the green
vigorous growing seedlings in vitro, while the
aberrant (albino) plantlets which could not be
acclimatised might have been hybrids.
In both H. syriacus · H. paramutabilis and
H. syriacus · H. sinosyriacus, true hybrids had
larger flowers compared to their parent plants and
flower colour was intermediate. On adult plants of
both hybrid progenies four different leaf types could
be observed completely at random. Leaf dimor-
phisms were described before for some other plant
genera. Juvenile plants or parts of plants of Hedera
helix bear alternately arranged, palmately lobed
leaves while the mature leaf is entirely ovate or
rhombic (Metcalfe 2005; Wareing and Frydman
1976). Similar observations of different foliage types
are reported in many species of Populus (Critchfield
1960) and also in Morus Alba (Everett 1960). It is
not clear whether or not this phenomenon on the
Hibiscus F1 seedlings can be explained by the
change from juvenile to adult phase. Changes in leaf
shape have never been seen in seedlings of the
parental species H. syriacus, H. sinosyriacus
or H. paramutabilis. So most probably the leaf
polymorphisms in the hybrids are caused by genetic
modifications after interspecific hybridisation.
The obtained hybrid plants were almost sterile.
According to Heslop-Harrison (1999) errors in
chromosome segregation at meiosis may cause F1
sterility. It is a well-known phenomenon
after interspecific hybridisation. Despite the low
fertility of the H. syriacus · H. paramutabilis
hybrids, a small F2 progeny could be obtained
so far. However, a larger F2 progeny is needed to
study segregation of leaf types and flower colours.
Breeding efforts to introgress new features in
Hibiscus cultivars is continued.
Acknowledgements The authors wish to thank the staffof the biotech lab for their support and skilful assistance.This work is financed by BEST-Select and IWT-Flanders(VIS-CO 020802).
References
Bates DM (1965) Notes on the cultivated Malvaceae. 1.Hibiscus. Baileya 13:57–130
Bean WJ (1973) Trees and shrubs hardy in the British isles(8th edition revised volume II). John Murray,London, UK
Buitendijk JH, Ramanna MS, Jacobsen E (1992) Micro-propagation ability: towards a selection criteria inAlstroemeria breeding. Acta Hortic 325:493–498
Critchfield WB (1960) Leaf dimorphism in Populustrichocarpa. Am J Bot 47:699–711
Doyle J, Doyle J (1987) Genomic plant DNA preparationfrom fresh tissue - CTAB method. Phytochem Bull19:11
Eeckhaut T, Van Laere K, De Riek J, Van HuylenbroeckJ (2006) Overcoming interspecific barriers in orna-mental plant breeding. In: Taxeira da Silva JA (ed)Floriculture, ornamental and plant biotechnology:advances and topical issues, 1st edn. Global ScienceBooks, London, UK. pp 540–551
Egolf DR (1971) Hibicsus syriacus ‘Diana’ a new cultivar(Malvaceae). Baileya 17:75–78
Egolf DR (1981) Meet ‘Helene’ a new Hibiscus syriacus.Am. Nurseryman 154:11–67
Everett TH (1960) New illustrated encyclopedia of gar-dening, vol. 7. Greystone Press
Heslop-Harrison Y (1999) Aspects of the cell biology ofpollination and wide hybridisation. In: Cresti M, CaiG, Moscatelli A (eds) Fertilisation in higher plants(molecular and cytological aspects). Springer, Berlin,Germany, pp 139–144
Hogenboom N (1973) A model for incongruity in intimatepartner relationships. Euphytica 22:219–233
Kim JH, Lee KC (1991) Studies on the flower colourvariation in Hibiscus syriacus L. I. Spectral propertiesof fresh petals and flower colour classification. JKorean Soc Hortic Sci 32:103–111
Kim J, Ham D, Lim H, Lee K (1996a) Cross compatibility,seed germinability, and embryo rescue following2x · 4x crosses in Hibiscus syriacus. J Korean SocHortic Sci 37:713–718
Kim J, Ham D, Lim H, Lee K (1996b) Development ofimmature embryo and abnormal endosperm afterreciprocal crosses between diploids and tetraploids inHibiscus syriacus. J Korean Soc Hortic Sci 37:462–467
Kuwada H (1964) The newly artificially raised amphidip-loid plant named ‘Ai-Fuyo’ (Hibiscus muta-moscheu-tos), obtained from the progeny of H. mutabilis · H.moscheutos (Studies on interspecific and intergenerichybridisation in Malvaceae VII). Jap J Breed 14:27–32
Kyung HY, Kim JH (2001a) Effects of flowering season onthe compatibilities of interspecific single, three wayand double crosses among Hibiscus syriacus, H.sinosyriacus and interspecific hybrids (H. syria-cus · H. paramutabilis). J Korean Soc Hortic Sci42:568–574
Kyung HY, Kim JH (2001b) Relationship between pistilreceptivity and cross compatibility in interspecificsingle, three way and double crosses among Hibiscussyriacus, H. sinosyriacus and interspecific hybrids (H.syriacus · H. paramutabilis). J Korean Soc Hortic Sci42:561–567
Kyung HY, Park SM, Kim JH (2001a) Effects of bud andold-flower pollination on the interspecific single andthree way cross compatibilities between cultivargroups of Hibiscus syriacus and H. sinosyriacus.J Korean Soc Hortic Sci 42:575–580
282 Euphytica (2007) 155:271–283
123

Kyung HY, Park SM, Kim JH (2001b) Embryo abortionand germination inhibition in interspecific single,three way and double crosses among Hibiscus syria-cus, H. sinosyriacus and interspecific hybrids (H.syriacus · H. paramutabilis). J Korean Soc HorticSci 42:737–742
Metcalfe DJ (2005) Biological flora of the British isles:Hedera helix L. J Ecol 93:632–648
Michishita A, Ureshino K, Miyajima I (2002) Plastome-genome incompatibility of Rhododendron serpyllifo-lium (A. Gray) Miq. to evergreen azalea speciesbelonging to series Kaempferia. J Jap Soc Hort Sci71:375–381
Murashige T, Skoog F (1962) A revised medium for rapidgrowth and bioassays with tobacco culture. PhysiolPlant 15:473–497
Niimoto DH (1966) Chromosome numbers of some Hibis-cus species and other Malvaceae. Baileya 14:29–34
Paek K, Hwang J, Jong S, Park S (1989a) In vitropollination, ovule culture of Hibiscus syriacus anddifferentiation of adventious bud and somatic embryofrom callus. J Korean Soc Hortic Sci 30:145–156
Paek K, Hwang J, Jong S, Park S (1989b) Plant regener-ation in tissue cultures initiated from immatureembryos of Hibiscus syriacus. J Korean Soc HorticSci 16:93–104
Paek K, Hwang J, Jong S, Park S (1989c) Somatichybridisation by protoplast fusion in Hibiscus syriacusand Hibiscus rosachinensis. Kor J Breed 21:95–102
Perkin-Elmer (1995) AFLPTM Plant Mapping Kit: ProtocolSharma H (1995) How wide can a wide cross be?
Euphytica 82:43–64Shim KK, Kim KH, Ha YM (1993) Characteristics of
triploid cultivars ‘Diana’ and ‘Helene’ in Hibiscussyriacus L. J Korean Soc Hortic Sci 34:54–67
Skovsted A (1941) Chromosome numbers in the Malva-ceae II. Comptes rendus des traveaux du laboratoireCarlberg. Serie Physiologique 23:195–242
Sneath P, Sokal R (1973) Numerical taxonomy: theprinciples and practice of numerical classification.W.H. Freeman, San Francisco
Tachibana Y (1958) An interspecific hybrid of H. mutabilisL. and H. moscheutos L. (Studies on Hibiscus III).J Hort Ass Japan 27:201–206
Tian HQ, Russell SD (1997) Micromanipulation of maleand female gametes of Nicotiana tabacum: I. Isolationof gametes. Plant Cell Rep 16:555–560
Van De Laar HJ (1997) Hibiscus syriacus. Dendroflora34:43–60
Van Huylenbroeck JM, De Riek J, De Loose M (2000)Genetic relationships among Hibiscus syriacus, Hibis-cus sinosyriacus and Hibiscus paramutabilis revealedby AFLP, morphology and ploidy analysis. GenetResour Crop Evol 47:335–343
Van Tuyl J, De Jeu M (1997) Methods for overcominginterspecific crossing barriers. In: Shivanna K, Sawh-ney V (eds) Pollen biotechnology for crop productionand improvement. University Press, Cambridge, UK.pp 273–292
Van Tuyl JM, Van Dien MP, Van Creij MGM, VanKleinwee TCM, Franken J, Bino RJ (1991) Applica-tion of in vitro pollination, ovary culture, ovuleculture and embryo rescue for overcoming incongru-ity barriers in interspecific Lilium crosses. Plant Sci74:115–126
Vervaeke I, Parton E, Maene L, Deroose R, De Proft M(2001) Prefertilisation barriers between differentBromeliaceae. Euphytica 118:91–97
Wareing PF, Frydman VM (1976) General aspects ofphase change with special reference to Hedera helix L.Acta Hortic 56:57–69
Yao JL, Cohen D (2000) Multiple gene control ofplastome-genome incompatibility and plastid DNAinheritance in interspecific hybrids of Zantedeschia.Theor Appl Genet 101:400–406
Yu TY, Yeam DY, Kim Y (1976) A study on the breedingof Hibiscus syriacus L. on hybridisation amongintroduced tetraploids and H. rosa-sinensis. J KoreanSoc Hortic Sci 17:107–112
Euphytica (2007) 155:271–283 283
123
Copyright © 2022 FDOKUMEN




















