
PARASITIC FLATWORMS (PLATYHELMINTHES: MONOGENEA, DIGENEA, CESTODA) OF FISHES FROM THE ADRIATIC SEA
Upload
independentCategory
view
2download
0
Molecules, morphology and morphometrics of Cainocreadium labracisand Cainocreadium dentecis n. sp.
(Digenea: Opecoelidae) parasitic in marine ®shesq
O. Joussona,b,*, P. Bartolib
aDeÂpartement de Zoologie et Biologie Animale, Universite de GeneÁve,154 route de Malagnou, CH-1224 CheÃne-Bougeries/Geneva, SwitzerlandbCentre d'OceÂanologie de Marseille, UMR CNRS 6540, F-13288 Marseille, France
Received 11 December 2000; received in revised form 14 February 2001; accepted 14 February 2001
Abstract
Molecular, morphological and morphometric analyses were conducted on several samples of Cainocreadium labracis (Opecoelidae), a
trematode parasitic in marine teleosts. The samples were isolated from several specimens of Dicentrarchus labrax, the type host, and Dentex
dentex. The molecular analysis of complete Internal Transcribed Spacer sequences of ribosomal DNA revealed that specimens isolated from
each host species form two well-de®ned groups, whose sequence divergence reaches 7.5%. The morphological study showed that the two
groups can be distinguished by several characters, including the level of maximum body breadth, the relative position of the testes, the shape
of the cirrus pouch, and the extent of the uterus. Multivariate analyses of morphometrics demonstrated consistency of most of the characters
for discriminating the two groups. Our results show that C. labracis specimens isolated from D. labrax and D. dentex represent clearly
distinct entities from molecular, morphological and statistical points of view, which has enabled us to describe a new species, Cainocreadium
dentecis n. sp. q 2001 Australian Society for Parasitology Inc. Published by Elsevier Science Ltd. All rights reserved.
Keywords: Cainocreadium labracis; Cainocreadium dentecis n. sp; Molecules; Morphology; Morphometrics; Host speci®city
1. Introduction
A key problem in evolutionary studies of parasitic organ-
isms lies in the knowledge of their degree of host-speci®-
city, which can be de®ned by the number of host species a
parasite is able to infect (Combes, 1995). Among the
Digenea, the miracidial stage that infects the molluscan
®rst host usually shows a strict speci®city (Gibson and
Bray, 1994; Nunez and De Jong-Brink, 1997). The speci®-
city of the developmental stages infecting second and de®-
nitive host may be less strict. In such cases it is considered
that speci®city is mainly driven by factors associated with
host ecology, such as habitat use or feeding preferences
(Adamson and Caira, 1994; Jousson et al., 2000).
The Digenea lack conserved and hard structures, which
makes the identi®cation of sibling species problematic. As
a consequence, discrete morphological differences observed
between specimens collected from different hosts species
have often been attributed to phenotypic plasticity (Bartoli
et al., 1989a). Given the dif®culties of closely-related species
identi®cation, species diversity and the degree of host-speci-
®city may have been underestimated among these parasites.
Recently, molecular data such as Internal Transcribed
Spacer (ITS) ribosomal DNA sequences have been used to
distinguish among closely related digenean species (Luton
et al., 1992; Adlard et al., 1993; DespreÁs et al., 1995; Jous-
son et al., 1998; Jousson and Bartoli, 2000). Comparative
analyses of morphometrics have been used in studies
attempting the identi®cation of closely related nematodes
(Hugot et al., 1995; Stock and Kaya, 1996) and trematodes
(Fried et al., 1997; Kostadinova et al., 2000) species.
In the present work, we have chosen as a model a well
known species of the family Opecoelidae Ozaki, 1925:
Cainocreadium labracis (Dujardin, 1845) Nicoll, 1909.
This species has been reported from Dicentrarchus labrax
(Moronidae) (in Maillard, 1971), Dicentrarchus punctatus
(in Fischthal, 1982), Dentex dentex (Sparidae) (in Stossich,
1905; Janiszewska, 1953; Bartoli et al., 1989b), Platichthys
¯esus (Pleuronectidae), Scophthalmus rhombus (Scophthal-
midae) (in Stossich, 1905), Labrus bergylta (Labridae) (in
Linstow, 1878), and Gaidropsarus mediterraneus (Gadidae)
(in Osmanov, 1940). We investigated the digenean fauna of
International Journal for Parasitology 31 (2001) 706±714
0020-7519/01/$20.00 q 2001 Australian Society for Parasitology Inc. Published by Elsevier Science Ltd. All rights reserved.
PII: S0020-7519(01)00180-1
www.parasitology-online.com
q DNA sequences analysed in this study have been deposited in the
EMBL / GenBank database under accession numbers AJ241795,
AJ241806 and AJ241808.
* Corresponding author. Tel.: 141-22-3498644; fax: 141-22-3492647.
E-mail address: [email protected] (O. Jousson).
D. labrax (the type host of C. labracis), D. dentex, L.
bergylta, and G. mediterraneus. Cainocreadium labracis
specimens were found only in D. labrax and D. dentex.
To test the hypothesis of the existence of a host-associated
species complex within C. labracis, we obtained morpholo-
gical data, as well as ITS ribosomal DNA sequences of
several C. labracis specimens isolated from the two de®ni-
tive host species. These ITS sequences from adults were
compared with opecoelid cercariae sequences obtained in
a previous study (Jousson et al., 1999). Additionally, a
comparative morphometric analysis was conducted on para-
sites isolated from the two host species in order to examine
the degree of the variability in the metrical characteristics of
the adults and to assess their value in discriminating species.
2. Material and methods
2.1. Sample collection
Living adult specimens of C. labraciswere isolated from
the digestive tract of several individuals of D. labrax (40
parasite specimens) and D. dentex (40 specimens). The
hosts were collected from the north-western Mediterranean
(Gulf of Marseille, and Scandola Natural Reserve, France).
2.2. Molecular data
DNA of several C. labracis specimens from both hosts was
extracted in guanidine lysis buffer (Maniatis, 1982), precipi-
tated with isopropanol and dissolved in distilled water. PCR
ampli®cations were performed in a total volume of 50ml with
an ampli®cation pro®le consisting of 40 cycles of 30 s at
948C, 30 s at 528C and 120 s at 728C, followed by 5 min at
728C for ®nal extension. The ITS region of the rDNA was
ampli®ed using a trematode-speci®c primer located 195 bp
from the 3 0 end of the 18S rDNA (S20T2: 5 0-GGTAAGTG-
CAAGTCATAAGC-3 0) and an universal primer at 5 0 end of
the 28S rDNA (L5: 5 0-TTCACTCGCCATTACTT-3 0).Ampli®ed PCR products were puri®ed using High Pure
PCR Puri®cation Kit (Roche Diagnostics), and sequenced
O. Jousson, P. Bartoli / International Journal for Parasitology 31 (2001) 706±714 707
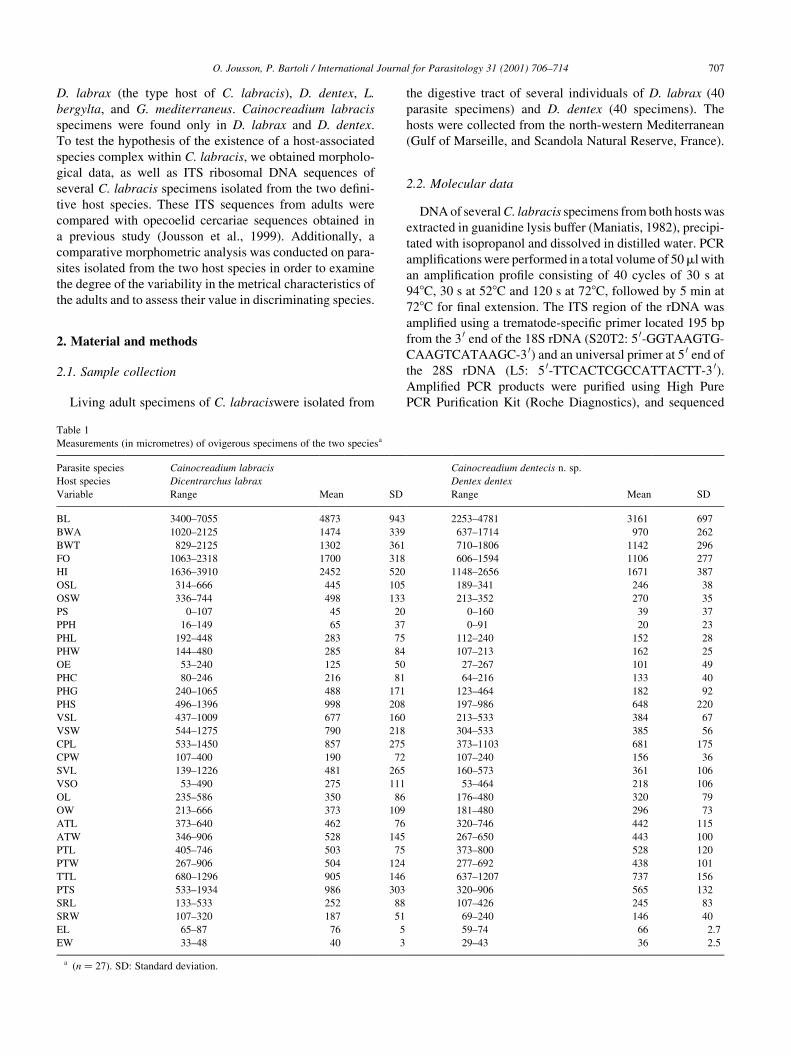
Table 1
Measurements (in micrometres) of ovigerous specimens of the two speciesa
Parasite species Cainocreadium labracis Cainocreadium dentecis n. sp.
Host species Dicentrarchus labrax Dentex dentex
Variable Range Mean SD Range Mean SD
BL 3400±7055 4873 943 2253±4781 3161 697
BWA 1020±2125 1474 339 637±1714 970 262
BWT 829±2125 1302 361 710±1806 1142 296
FO 1063±2318 1700 318 606±1594 1106 277
HI 1636±3910 2452 520 1148±2656 1671 387
OSL 314±666 445 105 189±341 246 38
OSW 336±744 498 133 213±352 270 35
PS 0±107 45 20 0±160 39 37
PPH 16±149 65 37 0±91 20 23
PHL 192±448 283 75 112±240 152 28
PHW 144±480 285 84 107±213 162 25
OE 53±240 125 50 27±267 101 49
PHC 80±246 216 81 64±216 133 40
PHG 240±1065 488 171 123±464 182 92
PHS 496±1396 998 208 197±986 648 220
VSL 437±1009 677 160 213±533 384 67
VSW 544±1275 790 218 304±533 385 56
CPL 533±1450 857 275 373±1103 681 175
CPW 107±400 190 72 107±240 156 36
SVL 139±1226 481 265 160±573 361 106
VSO 53±490 275 111 53±464 218 106
OL 235±586 350 86 176±480 320 79
OW 213±666 373 109 181±480 296 73
ATL 373±640 462 76 320±746 442 115
ATW 346±906 528 145 267±650 443 100
PTL 405±746 503 75 373±800 528 120
PTW 267±906 504 124 277±692 438 101
TTL 680±1296 905 146 637±1207 737 156
PTS 533±1934 986 303 320±906 565 132
SRL 133±533 252 88 107±426 245 83
SRW 107±320 187 51 69±240 146 40
EL 65±87 76 5 59±74 66 2.7
EW 33±48 40 3 29±43 36 2.5
a (n � 27). SD: Standard deviation.
directly using an ABI-377 DNA automated sequencer
(Perkin±Elmer), all according to the instructions of the
manufacturers. For DNA sequencing, the ampli®cation
primers were used, together with an internal reverse primer
located at 5 0 end of the 5.8S rDNA (5.8S1: 5 0-GCTGCGCTCTTCATCGACA-3 0). The sequences were
aligned manually using the GDE 2.2 (Larsen et al., 1993)
and their divergence was analysed using the Phylo-win
program (Galtier and Gouy, 1996).
2.3. Morphological data
Hosts (D. labrax and D. dentex) were necropsied imme-
diately after death for parasite studies. The digeneans
collected were studied while still living and later in perma-
nent preparations. Only ovigerous specimens have been
used in the descriptions. Cainocreadium labracis specimens
from each host were ®xed in Bouin's ¯uid between a slide
and a coverglass without pressure, coloured in acetic
carmine and mounted in Canada balsam. Studies were
conducted using a differential interference contrast micro-
scope and illustrations were made with a camera lucida.
Measurements are given in micrometres.
2.4. Variables
The following metrical features (variables) were
subjected to analyses (see Table 1): (1) body length (BL);
(2) body width at acetabulum level (BWA); (3) body width
at anterior testis level (BWT); (4) forebody (FO); (5) hind-
body (HI); (6) oral sucker length (OSL); (7) oral sucker
width (OSW); (8) preoral space (PS); (9) prepharynx
(PPH); (10) pharynx length (PHL); (11) pharynx width
(PHW); (12) oesophagus (OE); (13) pharynx to caecal bifur-
cation (PHC); (14) pharynx to genital aperture (PHG); (15)
pharynx to ventral sucker (PHS); (16) ventral sucker length
(VSL); (17) ventral sucker width (VSW); (18) cirrus pouch
length (CPL); (19) cirrus pouch width (CPW); (20) seminal
vesicle length (SVL); (21) ventral sucker to ovary (VSO);
(22) ovary length (OL); (23) ovary width (OW); (24) ante-
rior testis length (ATL); (25) anterior testis width (ATW);
(26) posterior testis length (PTL); (27) posterior testis width
(PTW); (28) two testes length (TTL); (29) post-testicular
space (PTS); (30) seminal receptacle length (SRL); (31)
seminal receptacle width (SRW); (32) egg length (EL);
(33) egg width (EW).
2.5. Statistical analyses
Initially, the descriptive univariate statistics (means and
SD) were calculated from original data. Statistical analyses
were performed on 33 metric characters (Table 1) using the
ADE-4 package (Thioulousse et al., 1997). An initial prin-
O. Jousson, P. Bartoli / International Journal for Parasitology 31 (2001) 706±714708
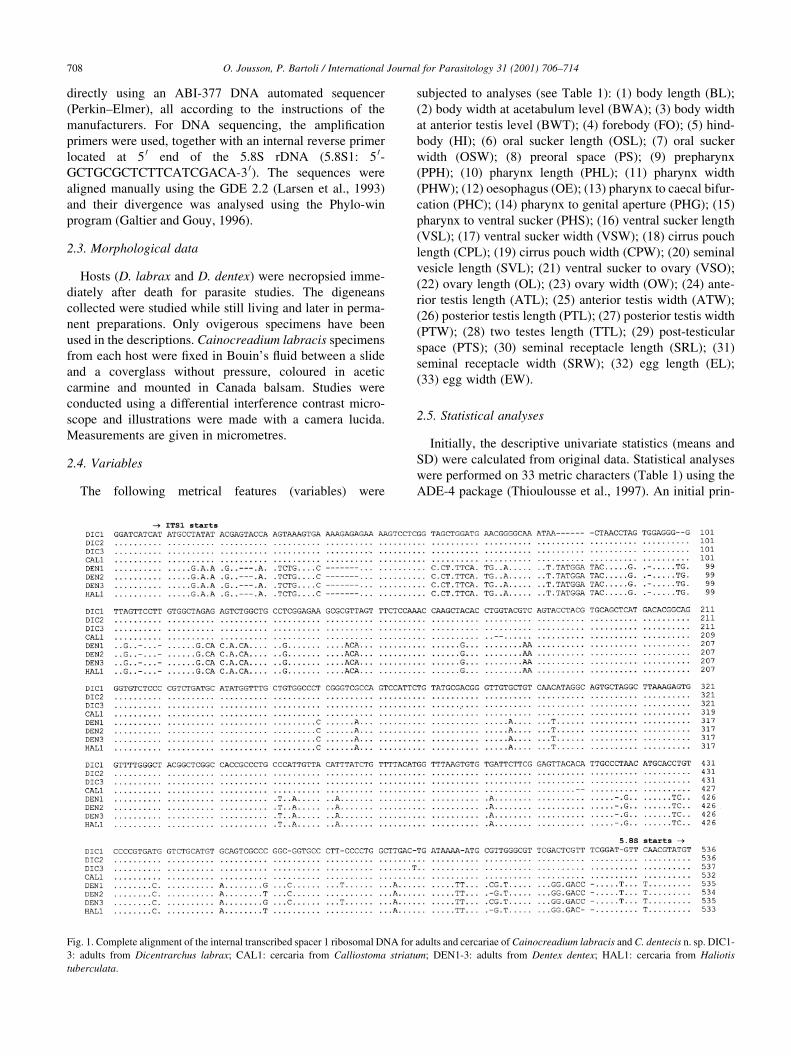
Fig. 1. Complete alignment of the internal transcribed spacer 1 ribosomal DNA for adults and cercariae of Cainocreadium labracis and C. dentecis n. sp. DIC1-
3: adults from Dicentrarchus labrax; CAL1: cercaria from Calliostoma striatum; DEN1-3: adults from Dentex dentex; HAL1: cercaria from Haliotis
tuberculata.
cipal component analysis (PCA) was performed using
normalised data. The size effect was controlled by measur-
ing only ovigerous specimens. A discriminant analysis was
performed to determine which part of the variance can be
explained by the differences between the two groups (i.e. the
specimens isolated from both hosts) and to verify results
obtained by PCA. A variance analysis (ANOVA) was
performed to determine which characters signi®cantly
discriminate the two groups.
3. Results
3.1. Molecular data
The analysed data set of the ITS region (ITS1 1 5.8S 1ITS2) and part of the ¯anking genes coding for 18S rRNA
and 28S rRNA of Cainocreadium specimens comprises
1164 sites. When all gaps-containing sites are removed,
there remain 1125 sites, of which 84 are variable (7.5%).
Two types of ITS sequences were found; one type includes
all adult specimens isolated from D. labrax and the cercaria
found in Calliostoma striatum. A second type includes all
adult specimens isolated from D. dentex and the cercaria
found in Haliotis tuberculata (Fig. 1). The ITS sequence
divergence between the two types reaches 7.5% (84 variable
sites), whereas the divergence within each type does not
exceed 0.8% (®ve variables sites). Both sequence types do
not differ signi®cantly regarding GC content (51.4±51.8%)
and complete ITS length (938±945 nucleotides). The
presence of a particular ITS type in parasites of each de®-
nitive host species suggests the occurrence of two species of
Cainocreadium: C. labracis from D. labrax and C. dentecis
n. sp. from D. dentex.
3.2. Morphological data
A complete description of C. labracis from D. dentex was
given by Bartoli et al. (1989b). Common features of adults
C. labracisfrom D. labrax and C. dentecis n. sp. from D.
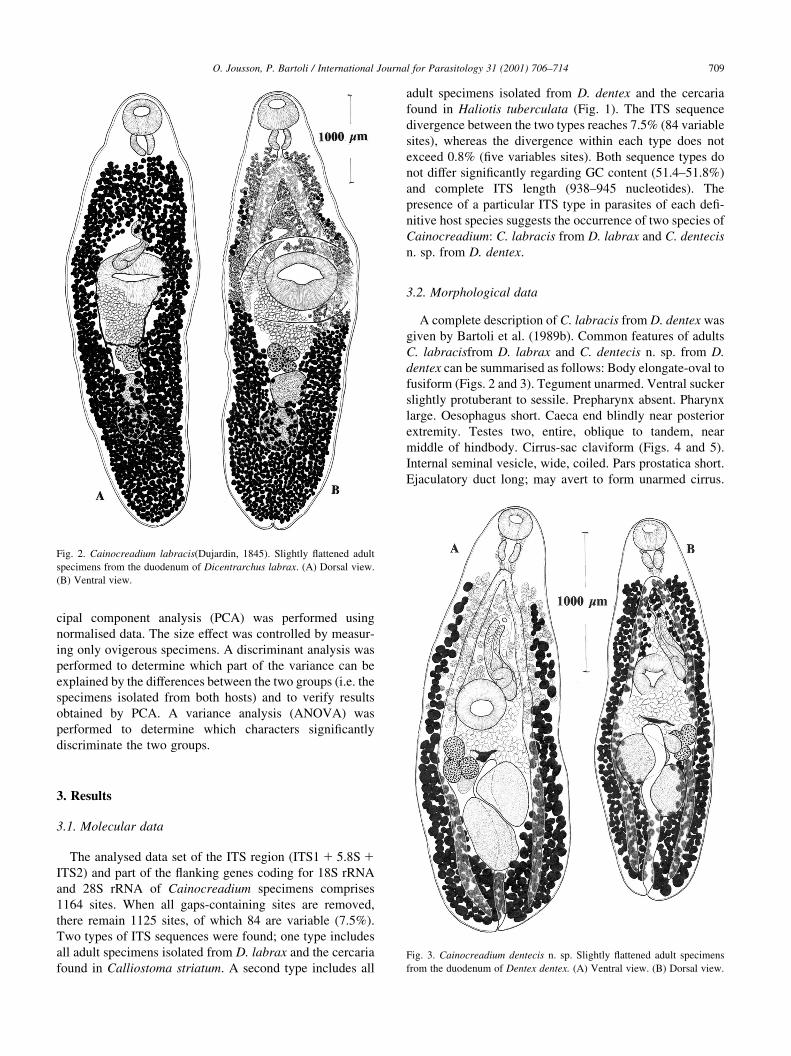
dentex can be summarised as follows: Body elongate-oval to
fusiform (Figs. 2 and 3). Tegument unarmed. Ventral sucker
slightly protuberant to sessile. Prepharynx absent. Pharynx
large. Oesophagus short. Caeca end blindly near posterior
extremity. Testes two, entire, oblique to tandem, near
middle of hindbody. Cirrus-sac claviform (Figs. 4 and 5).
Internal seminal vesicle, wide, coiled. Pars prostatica short.
Ejaculatory duct long; may avert to form unarmed cirrus.
O. Jousson, P. Bartoli / International Journal for Parasitology 31 (2001) 706±714 709
Fig. 2. Cainocreadium labracis(Dujardin, 1845). Slightly ¯attened adult
specimens from the duodenum of Dicentrarchus labrax. (A) Dorsal view.
(B) Ventral view.
Fig. 3. Cainocreadium dentecis n. sp. Slightly ¯attened adult specimens
from the duodenum of Dentex dentex. (A) Ventral view. (B) Dorsal view.
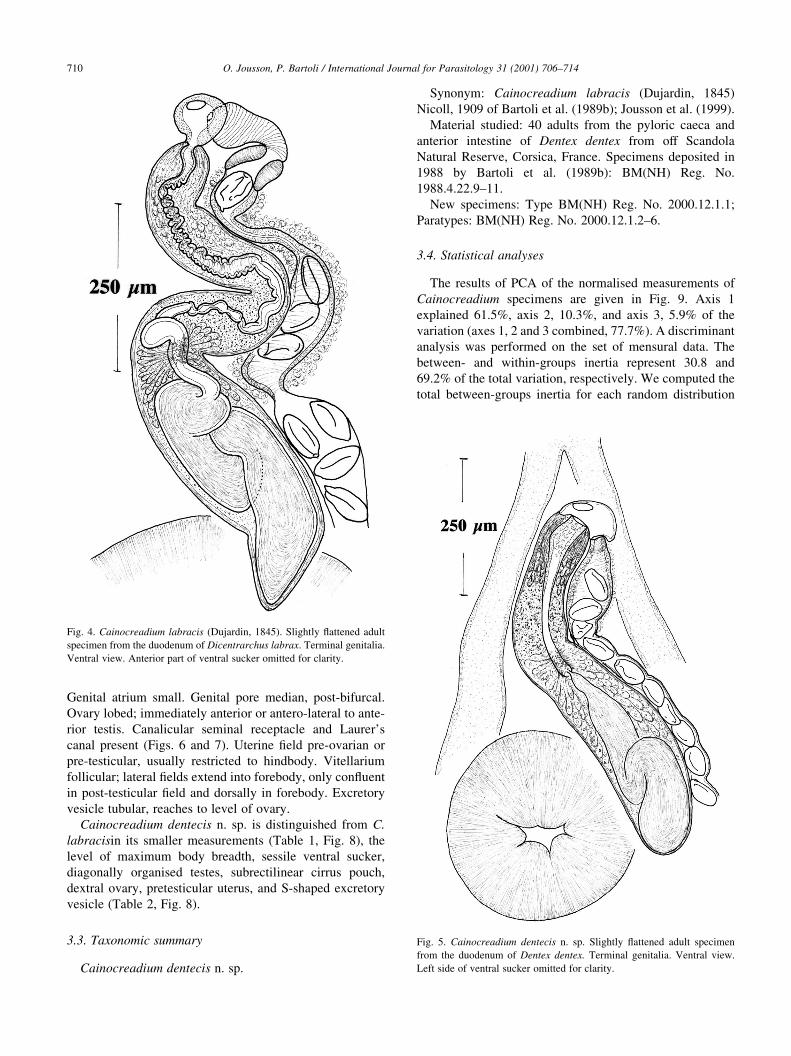
Genital atrium small. Genital pore median, post-bifurcal.
Ovary lobed; immediately anterior or antero-lateral to ante-
rior testis. Canalicular seminal receptacle and Laurer's
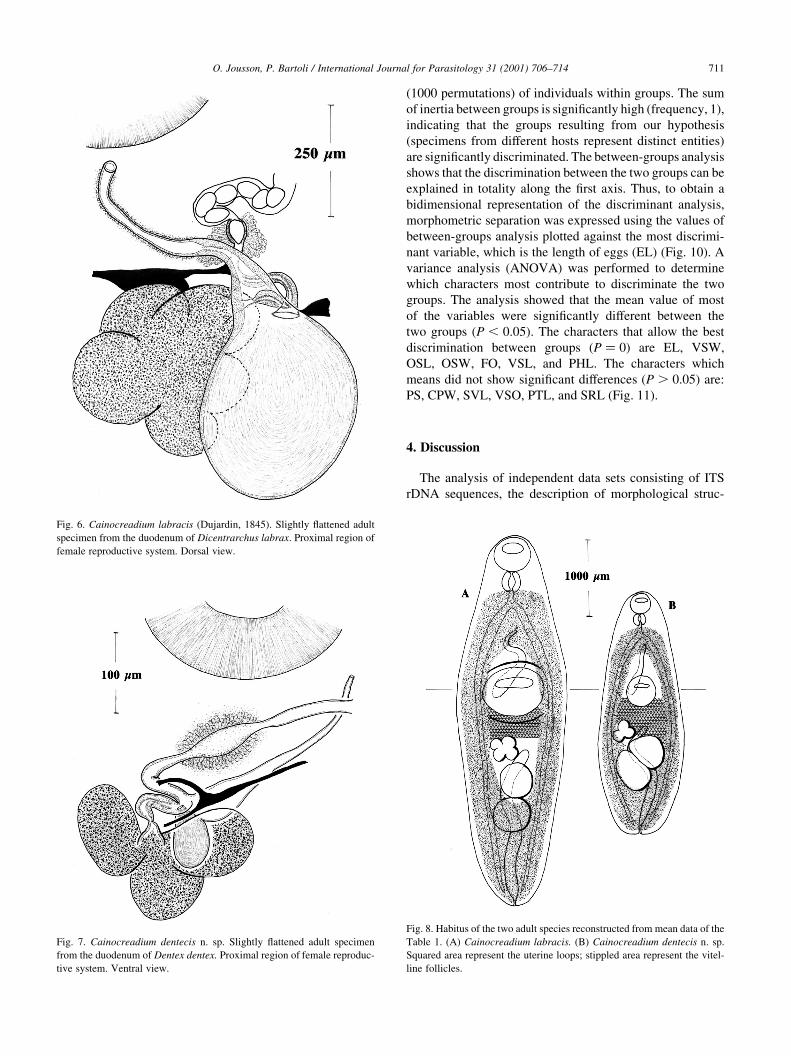
canal present (Figs. 6 and 7). Uterine ®eld pre-ovarian or
pre-testicular, usually restricted to hindbody. Vitellarium
follicular; lateral ®elds extend into forebody, only con¯uent
in post-testicular ®eld and dorsally in forebody. Excretory
vesicle tubular, reaches to level of ovary.
Cainocreadium dentecis n. sp. is distinguished from C.
labracisin its smaller measurements (Table 1, Fig. 8), the
level of maximum body breadth, sessile ventral sucker,
diagonally organised testes, subrectilinear cirrus pouch,
dextral ovary, pretesticular uterus, and S-shaped excretory
vesicle (Table 2, Fig. 8).
3.3. Taxonomic summary
Cainocreadium dentecis n. sp.
Synonym: Cainocreadium labracis (Dujardin, 1845)
Nicoll, 1909 of Bartoli et al. (1989b); Jousson et al. (1999).
Material studied: 40 adults from the pyloric caeca and
anterior intestine of Dentex dentex from off Scandola
Natural Reserve, Corsica, France. Specimens deposited in
1988 by Bartoli et al. (1989b): BM(NH) Reg. No.
1988.4.22.9±11.
New specimens: Type BM(NH) Reg. No. 2000.12.1.1;
Paratypes: BM(NH) Reg. No. 2000.12.1.2±6.
3.4. Statistical analyses
The results of PCA of the normalised measurements of
Cainocreadium specimens are given in Fig. 9. Axis 1
explained 61.5%, axis 2, 10.3%, and axis 3, 5.9% of the
variation (axes 1, 2 and 3 combined, 77.7%). A discriminant
analysis was performed on the set of mensural data. The
between- and within-groups inertia represent 30.8 and
69.2% of the total variation, respectively. We computed the
total between-groups inertia for each random distribution
O. Jousson, P. Bartoli / International Journal for Parasitology 31 (2001) 706±714710
Fig. 4. Cainocreadium labracis (Dujardin, 1845). Slightly ¯attened adult
specimen from the duodenum of Dicentrarchus labrax. Terminal genitalia.
Ventral view. Anterior part of ventral sucker omitted for clarity.
Fig. 5. Cainocreadium dentecis n. sp. Slightly ¯attened adult specimen
from the duodenum of Dentex dentex. Terminal genitalia. Ventral view.
Left side of ventral sucker omitted for clarity.
(1000 permutations) of individuals within groups. The sum
of inertia between groups is signi®cantly high (frequency, 1),
indicating that the groups resulting from our hypothesis
(specimens from different hosts represent distinct entities)
are signi®cantly discriminated. The between-groups analysis
shows that the discrimination between the two groups can be
explained in totality along the ®rst axis. Thus, to obtain a
bidimensional representation of the discriminant analysis,
morphometric separation was expressed using the values of
between-groups analysis plotted against the most discrimi-
nant variable, which is the length of eggs (EL) (Fig. 10). A
variance analysis (ANOVA) was performed to determine
which characters most contribute to discriminate the two
groups. The analysis showed that the mean value of most
of the variables were signi®cantly different between the
two groups (P , 0:05). The characters that allow the best
discrimination between groups (P � 0) are EL, VSW,
OSL, OSW, FO, VSL, and PHL. The characters which
means did not show signi®cant differences (P . 0:05) are:
PS, CPW, SVL, VSO, PTL, and SRL (Fig. 11).
4. Discussion
The analysis of independent data sets consisting of ITS
rDNA sequences, the description of morphological struc-
O. Jousson, P. Bartoli / International Journal for Parasitology 31 (2001) 706±714 711
Fig. 6. Cainocreadium labracis (Dujardin, 1845). Slightly ¯attened adult
specimen from the duodenum of Dicentrarchus labrax. Proximal region of
female reproductive system. Dorsal view.
Fig. 7. Cainocreadium dentecis n. sp. Slightly ¯attened adult specimen
from the duodenum of Dentex dentex. Proximal region of female reproduc-
tive system. Ventral view.
Fig. 8. Habitus of the two adult species reconstructed from mean data of the
Table 1. (A) Cainocreadium labracis. (B) Cainocreadium dentecis n. sp.
Squared area represent the uterine loops; stippled area represent the vitel-
line follicles.

tures and multivariate analyses of morphometrics clearly
showed a high degree of clustering between adults of C.
labracisisolated from D. labrax and D. dentex, demonstrat-
ing the occurrence of distinct species. Cainocreadium
labracis occurs in D. labrax, whereas C. dentecis n. sp.
occurs in D. dentex.
The amount of genetic divergence between the two
species is quite high (7.5%), and greatly exceeds the intras-
peci®c variation (0.8%). This degree of variability is much
higher than values of interspeci®c ITS sequence divergence
observed for other congeneric opecoelids (Jousson et al.,
1999) and among other digenean families (Adlard et al.,
1993). The occurrence of a host-associated species complex
within C. labracis is reinforced by the fact that correspond-
ing cercariae were isolated from two species of gastropods
belonging to different families of the order Archaeogastro-
poda: Haliotis tuberculata (Haliotidae) and Calliostoma
striatum (Trochidae) (Jousson et al., 1999). These cercariae
were found to correspond to C. labracis and C. dentecis,
respectively (Fig. 1). The more obvious non-metrical char-
acters that allow to distinguish the two adult species are the
disposition of the testes, the position of the uterus, and the
shape of the cirrus pouch (Fig. 8). Discriminant statistical
analyses showed that the differences between the two
groups (i.e. specimens isolated from C. labracis and from
C. dentecis) are highly signi®cant. The variance analysis
(ANOVA) demonstrated that most of the metrical charac-
ters show signi®cant differences in mean between C. labra-
cis and C. dentecis and thus allow a good discrimination
between the two species. The characters with the best scores
O. Jousson, P. Bartoli / International Journal for Parasitology 31 (2001) 706±714712
Table 2
Main non-metrical features differentiating the two species
Parasite species Cainocreadium labracis Cainocreadium dentecisn.sp.
Host species Dicentrarchus labrax Dentex dentex
Level of maximum body width Ventral sucker Anterior testis
Ventral sucker Prominent; often with peripheral folds Sessile; peripheral folds absent
Testes Rounded, tandem Somewhat triangular, diagonal
Anterior testis Median, smaller than acetabulum Sinistral, larger than acetabulum
Cirrus pouch S-shape Rectilinear
Cirrus Heavily sinuous Rectilinear
Ovary Submedian, in front of anterior testis Dextral, at level of anterior testis
Number of ovarian lobes 3±5 3
Uterus Preovarian Pretesticular
Vitelline follicles Very abundant Less abundant
Excretory vesicle Sinuous S-shaped
Fig. 9. Principal component analysis of C. labracis (1) and C. dentecis n. sp.
(2). Star-representation of individuals scores in the plane F1 £ F2 using
normalised variables.
Fig. 10. Morphometric separation of C. labracis (1) and C. dentecis n. sp.
(2). First axis of the between-groups discriminant analysis plotted against
eggs length (EL).
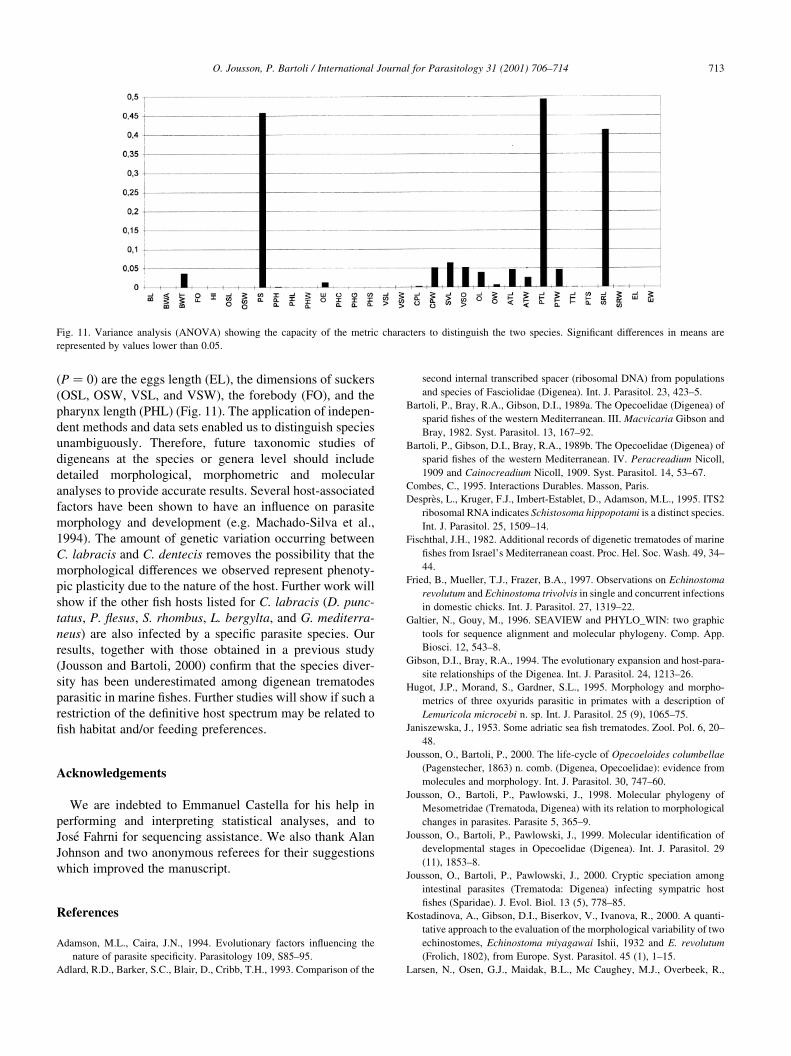
(P � 0) are the eggs length (EL), the dimensions of suckers
(OSL, OSW, VSL, and VSW), the forebody (FO), and the
pharynx length (PHL) (Fig. 11). The application of indepen-
dent methods and data sets enabled us to distinguish species
unambiguously. Therefore, future taxonomic studies of
digeneans at the species or genera level should include
detailed morphological, morphometric and molecular
analyses to provide accurate results. Several host-associated
factors have been shown to have an in¯uence on parasite
morphology and development (e.g. Machado-Silva et al.,
1994). The amount of genetic variation occurring between
C. labracis and C. dentecis removes the possibility that the
morphological differences we observed represent phenoty-
pic plasticity due to the nature of the host. Further work will
show if the other ®sh hosts listed for C. labracis (D. punc-
tatus, P. ¯esus, S. rhombus, L. bergylta, and G. mediterra-
neus) are also infected by a speci®c parasite species. Our
results, together with those obtained in a previous study
(Jousson and Bartoli, 2000) con®rm that the species diver-
sity has been underestimated among digenean trematodes
parasitic in marine ®shes. Further studies will show if such a
restriction of the de®nitive host spectrum may be related to
®sh habitat and/or feeding preferences.
Acknowledgements
We are indebted to Emmanuel Castella for his help in
performing and interpreting statistical analyses, and to
Jose Fahrni for sequencing assistance. We also thank Alan
Johnson and two anonymous referees for their suggestions
which improved the manuscript.
References
Adamson, M.L., Caira, J.N., 1994. Evolutionary factors in¯uencing the
nature of parasite speci®city. Parasitology 109, S85±95.
Adlard, R.D., Barker, S.C., Blair, D., Cribb, T.H., 1993. Comparison of the
second internal transcribed spacer (ribosomal DNA) from populations
and species of Fasciolidae (Digenea). Int. J. Parasitol. 23, 423±5.
Bartoli, P., Bray, R.A., Gibson, D.I., 1989a. The Opecoelidae (Digenea) of
sparid ®shes of the western Mediterranean. III. Macvicaria Gibson and
Bray, 1982. Syst. Parasitol. 13, 167±92.
Bartoli, P., Gibson, D.I., Bray, R.A., 1989b. The Opecoelidae (Digenea) of
sparid ®shes of the western Mediterranean. IV. Peracreadium Nicoll,
1909 and Cainocreadium Nicoll, 1909. Syst. Parasitol. 14, 53±67.
Combes, C., 1995. Interactions Durables. Masson, Paris.
DespreÁs, L., Kruger, F.J., Imbert-Establet, D., Adamson, M.L., 1995. ITS2
ribosomal RNA indicates Schistosoma hippopotami is a distinct species.
Int. J. Parasitol. 25, 1509±14.
Fischthal, J.H., 1982. Additional records of digenetic trematodes of marine
®shes from Israel's Mediterranean coast. Proc. Hel. Soc. Wash. 49, 34±
44.
Fried, B., Mueller, T.J., Frazer, B.A., 1997. Observations on Echinostoma
revolutum and Echinostoma trivolvis in single and concurrent infections
in domestic chicks. Int. J. Parasitol. 27, 1319±22.
Galtier, N., Gouy, M., 1996. SEAVIEW and PHYLO_WIN: two graphic
tools for sequence alignment and molecular phylogeny. Comp. App.
Biosci. 12, 543±8.
Gibson, D.I., Bray, R.A., 1994. The evolutionary expansion and host-para-
site relationships of the Digenea. Int. J. Parasitol. 24, 1213±26.
Hugot, J.P., Morand, S., Gardner, S.L., 1995. Morphology and morpho-
metrics of three oxyurids parasitic in primates with a description of
Lemuricola microcebi n. sp. Int. J. Parasitol. 25 (9), 1065±75.
Janiszewska, J., 1953. Some adriatic sea ®sh trematodes. Zool. Pol. 6, 20±
48.
Jousson, O., Bartoli, P., 2000. The life-cycle of Opecoeloides columbellae
(Pagenstecher, 1863) n. comb. (Digenea, Opecoelidae): evidence from
molecules and morphology. Int. J. Parasitol. 30, 747±60.
Jousson, O., Bartoli, P., Pawlowski, J., 1998. Molecular phylogeny of
Mesometridae (Trematoda, Digenea) with its relation to morphological
changes in parasites. Parasite 5, 365±9.
Jousson, O., Bartoli, P., Pawlowski, J., 1999. Molecular identi®cation of
developmental stages in Opecoelidae (Digenea). Int. J. Parasitol. 29
(11), 1853±8.
Jousson, O., Bartoli, P., Pawlowski, J., 2000. Cryptic speciation among
intestinal parasites (Trematoda: Digenea) infecting sympatric host
®shes (Sparidae). J. Evol. Biol. 13 (5), 778±85.
Kostadinova, A., Gibson, D.I., Biserkov, V., Ivanova, R., 2000. A quanti-
tative approach to the evaluation of the morphological variability of two
echinostomes, Echinostoma miyagawai Ishii, 1932 and E. revolutum
(Frolich, 1802), from Europe. Syst. Parasitol. 45 (1), 1±15.
Larsen, N., Osen, G.J., Maidak, B.L., Mc Caughey, M.J., Overbeek, R.,
O. Jousson, P. Bartoli / International Journal for Parasitology 31 (2001) 706±714 713
Fig. 11. Variance analysis (ANOVA) showing the capacity of the metric characters to distinguish the two species. Signi®cant differences in means are
represented by values lower than 0.05.

Macke, T.J., Marsh, T.L., Woese, C.R., 1993. The ribosomal database
project. Nucleic Acids Res. 21, 3021±3.
Linstow, O.F.B., 1878. Compendium der Helminthologie. Hahn'sche
Buchhandlung, Hannover, p. 382.
Luton, K., Walker, D., Blair, D., 1992. Comparisons of ribosomal internal
transcribed spacer from two congeneric species of ¯ukes (Platyhel-
minthes: Trematoda: Digenea). Mol. Biochem. Parasitol. 56, 323±8.
Machado-Silva, J.R., Galvao, C., Presgrave, O.A., Rey, L., Gomes, D.C.,
1994. Host-induced morphological changes of Schistosoma mansoni
Sambon, 1907 male worms. Mem. Inst. Oswaldo Cruz. 89 (3), 411±6.
Maillard, C., 1971. Cycle eÂvolutif de Cainocreadium labracis (Dujardin,
1845) (Trematoda, Allocreadiidae). C. R. Acad. Sci. 272, 3303±6.
Maniatis, T., 1982. Molecular Cloning, Cold Spring Harbor Laboratory,
New York.
Nunez, P.E., De Jong-Brink, M., 1997. The suppressive excretory-secretory
product of Trichobilharzia ocellata: a possible factor for determining
compatibility in parasite-host interactions. Parasitology 115, 193±203.
Osmanov, S.U., 1940. Studies on the parasite fauna of ®sh from the Black
Sea. Uchenye Zapiski Leningradskogo Gosudarstwennogo Pedagogi-
cheskogo Instituta im Gertsena 30, 187±265 (In Russian).
Ozaki, 1925; queried.
Stock, S.P., Kaya, H.K., 1996. A multivariate analysis of morphometric
characters of Heterorhabditis species (Nemata: Heterorhabditidae) and
the role of morphometrics in the taxonomy of species of the genus. J.
Parasitol. 82 (5), 806±13.
Stossich, M., 1905. Note distomologiche III-IV-V. Bolletino della SocietaÁ
Adriatica di Scienze Naturali in Trieste 22, 211±27.
Thioulousse, J., Chessel, D., DoleÂdec, S., Olivier, J.M., 1997. ADE-4: a
multivariate analysis and graphical display software. Stat. Comput. 7
(1), 75±83.
O. Jousson, P. Bartoli / International Journal for Parasitology 31 (2001) 706±714714
Copyright © 2022 FDOKUMEN




















