
Oil and Gas | A Roadmap for unlocking future growth ... - CSIRO
Upload
khangminh22Category
view
1download
0
Volume 37 Number 1 March 2016Volume 37 Number 1 March 2016
Parasitic infections
OFFICIAL JOURNAL OF THE AUSTRALIAN SOCIETY FOR MICROBIOLOGY INC.OFFICIAL JOURNAL OF THE AUSTRALIAN SOCIETY FOR MICROBIOLOGY INC.

Flexible, reliable performance and quality results. Thermo Scientifi c Mycobacteria
Solutions offer a comprehensive solution for mycobacteria testing, with the fl exibility to
utilise a variety of sample types, including: Blood, sputum, urine, sterile body fl uids and
tissue. When it comes to treating mycobacterial infections, better patient care starts with
innovative technologies that deliver trusted results.
to mycobacteria testing
Bringing more
© 2
014 T
herm
o F
isher
Scie
ntifi c
Inc. A
ll rights
reserv
ed
.
Mycobacteria detection
instruments
VersaTREK System • Perform four FDA-cleared tests
on a single platform: mycobacteria
detection, Mycobacterium tuberculosis
(Mtb) susceptibility testing, blood
culture and sterile body fl uids
• Unique growth matrix provides better
detection on all mycobacteria species
• One system, one VersaTREK™
Myco Media bottle for all
mycobacteria testing
Mycobacteria MIC
testing systems
Sensititre:Sensititre™ Mycobacterium
tuberculosis MIC Plate includes
12 fi rst and second-line antimicrobics
on a single plate. Or, choose
Sensititre Rapid or Slow Growing
Mycobacterium MIC plates, all with
manual viewer or Sensititre
Vizion read options
Versatrek:VersaTREK System offers four FDA-
cleared, primary antituberculosis
drugs for automated Mtb susceptibility
testing, including rifampin,
isoniazid, ethambutol and PZA,
as well as streptomycin
Thermo Scientifi c
Prepared Media
Culture mycobacteria specimens with
a full range of mycobacteria media,
including Thermo Lowenstein-Jensen,
MacConkey Agar w/o Crystal Violet and
Middlebrook Agar Media.
• For more information email MicroTechSupport@thermofi sher.com

OFFICIAL JOURNAL OF THE AUSTRALIAN SOCIETY FOR MICROBIOLOGY INC.
Volume 37 Number 1 March 2016
ContentsFrom the Editorial Team 2
Jo, Ian, and Hayley Macreadie
Guest Editorial 3Parasitic infections: overlooked, under-diagnosed and under-researched 3
Harsha Sheorey and Richard S Bradbury
In Focus 4The laboratory diagnosis of Strongyloides stercoralis 4
Matthew R Watts, Gemma Robertson and Richard S Bradbury
Under the Microscope 10Current WHO protocols for mass drug administration in helminth control 10
Richard S Bradbury and Patricia M Graves
Zoonotic tissue parasites of Australian wildlife 12
David M Spratt
Assessing enteric helminths in refugees, asylum seekers and new migrants 15
Sarah Hanieh, Norbert Ryan and Beverley-Ann Biggs
Free living amoebae and human disease 20
Evan Bursle and Jennifer Robson
Tapeworm cysts in the brain: can we prevent it happening? 25
Marshall Lightowlers
Seafood-borne parasitic diseases in Australia: how much
do we know about them? 27
Shokoofeh Shamsi
Children, snails and worms: the Brachylaima cribbi story 30
Andrew R Butcher
Malaria: global challenges for malaria eradication 34
Graham Brown and Stephen Rogerson
Plasmodium knowlesi: an update 39
Balbir Singh
Diagnosis of human taeniasis 43
Abdul Jabbar, Charles Gauci and Marshall W Lightowlers
Lab Report 46Protozoa PCR: boon or bane 46
Colin Pham and Harsha Sheorey
ASM Affairs 50FT020 Water Microbiology Australian Standards Committee 50
2016 ASM Communication Ambassador Program 50
Stopping dengue: recent advances and new challenges 51
MICROBIOLOGY AUSTRALIA • MARCH 2016 1
Dr Gary Lum Dr John MerlinoProf. Wieland MeyerProf. William RawlinsonDr Paul SelleckDr David SmithMs Helen SmithDr Jack Wang
The Australian Societyfor Microbiology Inc.9/397 Smith StreetFitzroy, Vic. 3065Tel: 1300 656 423Fax: 03 9329 1777Email: [email protected] 24 065 463 274
For Microbiology Australiacorrespondence, see address below.
Editorial teamProf. Ian Macreadie, Mrs Jo Macreadieand Mrs Hayley Macreadie
Editorial BoardDr Chris Burke (Chair)Prof. Mary BartonProf. Linda BlackallProf. Sharon ChenProf. Peter ColoeDr Narelle FeganDr Geoff HoggProf. Jonathan IredellDr I
.pek Kurtböke
Subscription ratesCurrent subscription rates are availablefrom the ASM Melbourne offi ce.
Editorial correspondenceProf. Ian Macreadie/Mrs Jo MacreadieTel: 0402 564 308 (Ian)Email: [email protected]
Published four times a year in print and open access online by
Unipark, Building 1, Level 1 195 Wellington Road, Clayton, Vic. 3168http://microbiology.publish.csiro.au
Publishing enquiriesJenny FosterEmail: [email protected]
Production enquiriesHelen PavlatosEmail: [email protected]
Advertising enquiriesDoug WaltersTel: 03 9545 8505Mobile: 0419 357 779Email: [email protected]© 2016 The Australian Society for Microbiology Inc. The ASM, through CSIRO Publishing, reserve all rights to the content, artwork and photographs in Microbiology Australia. Permission to reproduce text, photos and artwork must be sought from CSIRO Publishing.
The Australian Copyright Act 1968 and subsequent amendments permit downloading and use of an article by an individual or educational institution for non-commercial personal use or study. Multiple reproduction of any Microbiology Australia article in a study block is governed by rights agreement managed by Copyright Agency Limited and fees may apply.
Authors published in Microbiology Australia have the moral right under Australian law to be acknowledged as the creator.
ISSN 1324-4272eISSN 2201-9189
While reasonable effort has been made to ensure the accuracy of the content, the Australian Society for Microbiology, CSIRO, and CSIRO Publishing accept no responsibility for any loss or damage from the direct or indirect use of or reliance on the content. The opinions expressed in articles, letters, and advertisements in Microbiology Australia are not necessarily those of the Australian Society for Microbiology, the Editorial Board, CSIRO, and CSIRO Publishing. Cover image: Protoscolex of Echinococcus granulosus, showing row of hooklets; obtained from liver
aspirate of a case of hydatid cyst. Courtesy of Associate Professor Rob Baird, Royal Darwin Hospital.

Welcome to the first issue of Microbiology Australia for 2016.
As members of The Australian Society for Microbiology know, our
electronic issues are freely available. To be notified of a new issue go
toourpublisher’swebsitehttp://microbiology.publish.csiro.au/ and
register to get alerts when each issue becomes available.
ASM members receive print copies by mail if they have opted to
receive them that way. Contact the ASMOffice if youwish to receive
print copies. Additional copies are often made for special issues.
For example, ~1000 copies were made for delegates of the ISHAM
conference in Melbourne last year. The print copies are highly
valued by many members and are great to share with others.
The online issues are highly accessed as a reference and learning
resource. Individual articles as well as entire issues can be readily
downloaded from the above website.
Themes of issues and Guest Editors are chosen by the Editorial
Board whomeet by teleconference five times each year. The Board
always values input from ASMmembers for future issues. All articles
are peer-reviewed and invited by theGuest Editorswho have a deep
understanding of the themes they present. Themes are chosen to
be topical and of interest to ASM members: authors write to reach
a broad audience.
On occasions urgent updates are needed and such updates are
presented asHot Topics. However, we are aware that there is a need
to have an occasional non-themed issue. Please contact us if you
have suggestions for articles that will have broad interest to ASM
members.
We are grateful for the support the Editorial Board and for Guest
Editors who are experts in their fields. Guest Editors present the
latest knowledge in their field through the selection of contributors
who write compact articles that are readable and informative to the
ASM’s diverse membership. We also thank the reviewers, who
provide valuable comments on each contribution. They also have
expertise in the articles they review, and they provide valuable
comments to contributors to ensure the articles are appropriately
presented. We also thank the authors for their contributions. They
provide a great amount of current information about their field and
seek to educate us all in the exciting developments ofmicrobiology.
We thank The Australian Society for Microbiology and those who
lead it. In thechallengingworldofpublishing ithas takenboldness to
enableMicrobiology Australia to be produced as a publication that
is freely available to all. We know from the download data provided
byCSIROPublishing, thatMicrobiology Australia is highly accessed
around the world, and that the articles continue as an ongoing
resource many years after their first publication. Articles published
more than a decade ago still enjoy thousands of downloads every
year. There is no doubt that they serve as resource material for
teachers, students and those seeking reliable scientific information
from experts.
We trust that you enjoy your reading of this year’s issues, which will
cover:
* Parasitic Infections* Education to enhance microbiology graduate employability* Diseases of Aquaculture* Microbiology of Travel [a special joint issue with The MicrobiologySociety in the UK].
Jo, Ian, and Hayley Macreadie
Access to Microbiology Australia
Online early articles: Articles appear on the Microbiology Australia website (http://microbiology.publish.csiro.au/) when authors have approved the pdf of their article.
Completed issues: Register at http://microbiology.publish.csiro.au/ to receive notification thatan issue is complete. You will get an email showing the titles and abstracts of the completedissue. You will have one click access to any article or the whole issue.
Print issue: ASM members are welcome to receive the print version of Microbiology Australiawithout charge. To receive the print version you need to notify the ASM National Office (http://www.theasm.org.au/).
2 10.1071/MA16001 MICROBIOLOGY AUSTRALIA * MARCH 2016
From theEditorial Team

Parasitic infections: overlooked, under-diagnosedand under-researched
Harsha Sheorey
Microbiology DepartmentSt Vincent’s Hospital MelbourneFitzroy, Vic., AustraliaEmail: [email protected]
Richard S Bradbury
School of Medical and AppliedSciencesCentral Queensland UniversityRockhampton, Qld, AustraliaEmail: [email protected]
ProfessorGeorgeNelson (1924–2009) once stated that, ‘Parasitology
is the preserve of the diagnostically destitute’. Little has changed to
thisday,withpotentially relevantparasiticcausesof illnessesoftennot
being considered early in the differential diagnoses of clinical pre-
sentations. Parasitic infections are sometimes overlooked as causes
of morbidity and (in some cases) mortality in both the medical and
veterinary fields. In Australia there remain significant problems
associated with giardiasis, cryptosporidiosis, strongyloidiasis and
other parasitic diseases, particularly in remote, underserved and
tropical regionsof the country and also in the immuno-compromised
individuals (HIV, immunosuppressive drugs etc.). The burden of
many parasitic diseases is greater in tropical and sub-tropical areas
of non-industrialised countries. With increasingly adventurous travel
and dining, increasing numbers of Australians returning from travel
overseas with added souvenirs of common or exotic parasitoses
every year and refugees and migrants arriving in Australia, these
infections are becoming increasingly important.
Recent advances in diagnostic techniques for the detection of
parasitic infections have revolutionised how we undertake such
diagnostic investigations. These methodologies often provide
greater sensitivity as well as in some cases providing additional
epidemiological data. As we increasingly move towards the use of
molecular methodologies and away from traditional morphological
diagnosis, newchallenges have emerged for clinicians, veterinarians
and laboratory staff. A focus of several articles in this edition is
consideration of the advantages and disadvantages of these new
methods, in what circumstances they are best applied and how the
results of such investigations should be best interpreted. Molecular
methods are not without potential sources of error, hence in some
situations, morphological and serological techniques still remain
relevant.
This edition also considers zoonotic parasitic infections, the treat-
ment of parasitic infections, both from the current WHO recom-
mendations formass drug administration inhighly endemic settings
to the rise of resistance to anti-parasitic agents in protozoa such as
malarial parasites and Giardia intestinalis. Whilst antimicrobial
resistance is greatly investigated in bacterial infections, its emer-
genceandprevalence inparasitic infectionsofhumanandveterinary
importance will require further investigation and attention in the
future.
We hope that this edition of Microbiology Australia will update
knowledge andserve to informall our readersof the importance and
relevance of parasitic disease. Whether one is involved in medical,
veterinary, food, environmental or other microbiological work, it is
likely that aspects of your work will at some stage involve this
important and sometimesneglectedfieldof our scientificdiscipline.
As guest editors, we are grateful and excited to be involved in the
planning and execution of this edition. We would like to thank the
editorial staff and all of the authors and reviewers who have kindly
contributed their time and expertise into the preparation of this
edition. We hope that all members of the society will find it helpful,
interesting and that it may spark the interest of many into this most
fascinating and under-researched area.
Biographies
The biography for Dr Harsha Sheorey is on page 49.
The biography for Dr Richard Bradbury is on page 9.
GuestEditorial
MICROBIOLOGY AUSTRALIA * MARCH 2016 10.1071/MA16002 3

The laboratory diagnosis of Strongyloidesstercoralis
Matthew R WattsA,*, Gemma RobertsonB,* and Richard S BradburyC,D
ACentre for Infectious Diseases and Microbiology, Pathology West – ICMPR, and Marie Bashir Institute, University of Sydney, Westmead Hospital,Westmead, Sydney, NSW, Australia, Tel: +61 2 9845 6255, Email: [email protected]
BMelbourne Pathology, Collingwood and James Cook University, Tel: +61 3 9287 7700, Email: [email protected]
CSchool of Medical and Applied Sciences, Central Queensland University, Rockhampton, Qld, Australia
DCorresponding author. Email: [email protected]
It is estimated that over 30million people worldwide are
infected by the nematode, Strongyloides stercoralis1. It is
endemic in sub-tropical and tropical parts of Australia, with
high rates of infection documented in some indigenous
communities2. Due to the potential for chronic autoinfec-
tion, that may persist for decades, migration leads to the
presence of the infection in non-endemic areas1. Transmis-
sion tohumans isgenerally throughthepenetrationof larvae
through the skin, following contact with faecally contami-
nated soil1. Disease severity ranges from asymptomatic
chronic carriage to an overwhelming illness, where large
numbers spread throughout the body, usually triggered by
immunosuppression1.
Clinicians areadvised toconsider strongyloidiasis inpatientsprior to
immunosuppression, or with indicative symptoms, if there is a
history of probable exposure in an endemic area, regardless of the
elapsed time since exposure3,4. Eosinophilia is not an accurate
marker of strongyloidiasis, with a retrospective study finding that
only a quarter of patients with Strongyloides infection had a raised
eosinophil count5. The detection of strongyloidiasis is optimised by
appropriate test ordering, clinical notes, specimen transportation,
and processing by the receiving laboratory.
The gold standard for the diagnosis of strongyloidiasis is the
morphological identification of larvae in stool, tissue biopsies, and
other clinical specimens such as bronchoalveolar lavage. However,
in chronic infections, detection canbe limited by low larval output in
stool, leading to false negative results6. Consequently, in validation
studies for serological and nucleic acid tests there is a tendency
to define heavier infections as ‘true positives’. This affects serolog-
ical cut-offs, measurements of sensitivity and specificity, and
positive and negative predictive values7. Recognition of these
limitations is important for the interpretation of negative diagnostic
test results, where clinical suspicion remains. Here, we will give an
overview of currently available conventional and molecular tests
for the diagnosis of strongyloidiasis.
Stool microscopy and culture methods
Specimen transport and storage have amajor impact on the efficacy
of culture techniques in the laboratory diagnosis of S. stercoralis
from faecal samples. Fresh, unrefrigerated samples should be
delivered to the laboratory for culture as soon after collection as
possible, as the viability of larvae decreases incrementally with
storage at 48C over a 72-h period8. Rhabditiform and filariform
larvae will be found along with free-living adults of S. stercoralis in
*These two authors contributed equally.
In Focus
4 10.1071/MA16003 MICROBIOLOGY AUSTRALIA * MARCH 2016

older cultures (Figure 1). Larval stages must be differentiated from
those of hookworms, which may also be recovered.
Microscopic methodologies such as examination of Kato-Katz pre-
parations, FLOTAC, and formalin/ethyl acetate concentrates have a
low yield compared to culture9,10. Amodified formalin/ethyl acetate
method proposed by Anamnart et al. improved rates of detection9.
Overall, however, microscopic techniques alone are insensitive and
not sufficient for the exclusion of strongyloidiasis. In one of these
studies, though 30 of 254 participants were diagnosed with stron-
gyloidiasis by either agar plate culture (APC; Figure 2) or Baermann
culture techniques, no infections were identified by microscopy
using the Kato-Katz technique11.
APC is possibly the easiest culture to perform in the context of high
volume diagnostic testing. Results are available within two days,
although extended incubation up to four days increases yield8,11,12.
Two studies comparing 48 h APC with Baermann culture found an
improved recovery of S. stercoralis larvae inAPC11,12. Recovery rates
improve markedly with multiple stool cultures6,11,13
Serological diagnosis
Several tests for the serological diagnosis of strongyloidiasis have
been described, using both crude and recombinant antigens. Two
commercial ELISA kits employing somatic antigens are available
from BORDIER (Strongyloides ratti antigen) and IVD Research
(S. stercoralis antigen), respectively14. Recently, two recombinant
antigens (32 kD recombinant antigen, called NIE and S. stercoralis
immmunoreactive antigen, SsIR) have been employed for serolog-
ical testing in both ELISA and luciferase immunoprecipitation sys-
temassay (LIPS) platforms15. The reported sensitivity and specificity
of various serological platforms ranges from 56-100% and 29-100%,
dependent upon themethod, antigens, cut-offs, study populations,
and referencemethods employed16. Strongyloides serology using a
crude larval extract antigen was shown in one study to be less
sensitive for the diagnosis of returned travellers (73%) compared
topatientswhohave lived for anextendedperiod in anendemicarea
(98%)17. No definitive study of serological methods has been con-
ducted to date, and much of the available data is subject to flaws in
methodology, particularly the use ofmicroscopy only as a reference
(a) (b)
(c)
Figure 1. Life stages of Strongyloides stercoralis in agar plate culture: (a) rhabditiform larva; (b) a free-living adult male with filariform larva adjacent;and (c) a gravid, free-living adult female with filariform larva adjacent.
In Focus
MICROBIOLOGY AUSTRALIA * MARCH 2016 5

standard for positive specimens and varying S. stercoralis exposure
rates amongst tested serum groups16. A 2010 study with a reference
standard of a combination of three culture methods and sedimen-
tation concentration found NIE LIPS had a sensitivity of 97.8% (cut-
off 37.89 LU) in a study population with high endemicity but from
regions without filarial infection15. Lower sensitivity resulted when
testing the same samples by NIE ELISA(84%), NIE-SsIR LIPS (91.2)
and a S. stercoralis crude antigen extract ELISA (97%)18. All assays
tested showed 100% specificity in this study15. A more recent study
comparedan in-house crude S. stercoralisfilariform larvae immuno-
fluorescent antibody test (IFAT)with the ELISAs from IVDResearch,
Bordier, anda recombinant antigenNIEELISA andLIPS14. This study
used reference samples identified as positive by culture as well as
microscopy, and also a composite reference standard of concordant
results in at least three of five serological tests14. The in-house IFAT
was found to be the most sensitive (93.9%) when used in a test
subject groupwithnoknownpreviousexposure to S. stercoralis and
using the composite reference standard, whilst NIE LIPS was found
to be the most specific test (100%)14. Furthermore, when tested
against subjects with potential previous exposure and using the
composite reference standard, NIE LIPS was almost 100% specific
and 84.6% sensitive (cut off value 1388 LU)14. In testing against the
same sample group, the Bordier and IVD ELISAs maintained a high
specificity (almost 100%), but a lower sensitivity (70% and 79%,
respectively) and the NIE ELISA showed the highest specificity
(99%), but a low sensitivity (45%) (cut-off 76.5 U/mL)14.
Seroreversion following treatment of many, but not all, patients
was noted in a study using a Strongyloides ratti antigen ELISA3.
However, this effect is not universal and varies between studies3,17.
Immunosuppression was demonstrated to cause a reduction in
serological sensitivity (62% vs previously determined 92%), when
testing haematological patients on antineoplastic therapies18. NIE
LIPS did not cross-react with antigens from other parasites in the
study by Bisoffi et al., whereas IFAT and the two commercial ELISAs
did yield false positives, particularly from Mansonella perstans
infection14. Suchcross-reactionmaybedecreasedbypre-incubation
of serum in an extract of Onchocerca gutterosa16.
Nucleic acid tests
Nucleic acid tests complement non-molecular methodologies for
the diagnosis of S. stercoralis, and allow the use of refrigerated,
frozen, or preserved specimens11,19,20. This simplifies specimen
transportation, particularly where collection occurs some distance
from the testing laboratory, and there is no risk of laboratory-
acquired strongyloidiasis21. DNA extraction and amplification can
be performed within 1 day, however, laboratories may batch speci-
mens according to demand.
DNA extraction
It is important that DNA extractionmethods for stool specimens are
effective at removing the numerous nucleic acid test inhibitors in
stool22. A comparisonof5methodsofDNAextractiondemonstrated
that two column-basedmethodswere themost effective for the PCR
detection ofDNA from Strongyloides ratti that had been spiked into
human stool. These were the MoBio PowerSoil kit (MoBio Labora-
tories, Carlsbad, CA, USA) and a method based on modifications of
Figure 2. Larval tracks of Strongyloides stercoralis on Koga agar plate culture.
In Focus
6 MICROBIOLOGY AUSTRALIA * MARCH 2016
the QiaAmp Tissue kit (Qiagen, Hilden, Germany) by Verweij et al.,
which has been successfully automated23. The comparison
found that bead beating prior to the use of the NucliSens EasyMag
(BioMerieux, Marcy l’Etoile, France) was less effective, which indi-
cates the method of sample pretreatment prior to automated
extractionwill impact upon test sensitivity. Other investigators have
used a variety of different extractionmethods for StrongyloidesPCR,
including in-house methods, the Qiagen stool kit (unmodified and
modified), and the Nucleospin Soil kit (Macherey-Nagel, Duren,
Germany)10,24–30.
One of the inherent limitations of the molecular diagnosis of
S. stercoralis is the sampling error that can occur when relatively
small amounts are extracted in the context of low larval output6.
For example, 2g of stool can be used for agar plate culture, whereas
250mg of specimen is recommended for the MoBio PowerSoil
kit31. Methods that concentrate larger amounts of stool prior to
DNA extraction have the potential to increase test sensitivity, if
they remove inhibitors and retain larvae29.
PCR
Current PCR methods most commonly target one of four regions:
the 18S rRNA small subunit (SSU); the internal transcribed spacer
region 1 (ITS-1); the 28S rRNA gene; or the cyclooxygenase gene
(cox1)19,23–30,32–37. Published sensitivities and specificities for
Strongyloides PCR vary according to the reference methods and
are listed in Table 1. Themajority of Strongyloides PCR publications
have used a real-time method with primers and probe published
by Verweij et al.19. This has also allowed for the development of
multiplexed PCR10,30,34,38. Some studies evaluating the diagnostic
accuracy of these PCR methods have used both morphological
diagnosis and detection of PCR products as their reference stan-
dards, and are not reviewed here. Theirmethodology precludes the
calculation of sensitivity and specificity based on gold-standard,
according to an FDA Guidance39.
In the absence of a consistent gold standard in chronic infection,
positive nucleic acid test results, where conventional tests are
negative, may be due to greater sensitivity or false positive results6.
No PCR studies have reported false positive results when analytical
specificity has been tested using DNA extracted from bacteria,
viruses, fungi, protozoa, and other helminths19,23,24,27,29,30. Studies
have also assessed the specificity of the PCR products by sequence
analysis, with all finding 100% sequence homology with the target
sequence of S. stercoralis.24,25,27,29. Sitta et al. found a number of
false positives, using published genus and species-specific primers,
based on non-target sized bands on gel electrophoresis19,25. The
genus-specific primers amplified sequences that generated non-
target bands on electrophoresis in specimens that contained
Blastocytis and other helminths on microscopy, and the species-
specific primers amplified sequences that generated non-target
bands on electrophoresis in specimens positive for hookworm on
microscopy25. Similar accounts of cross-reactivity have not yet been
reported, so further data will be useful to monitor the specificity of
PCR in different populations.
Table 1. Sensitivity and specificity of stool PCR for human strongyloidiasis.
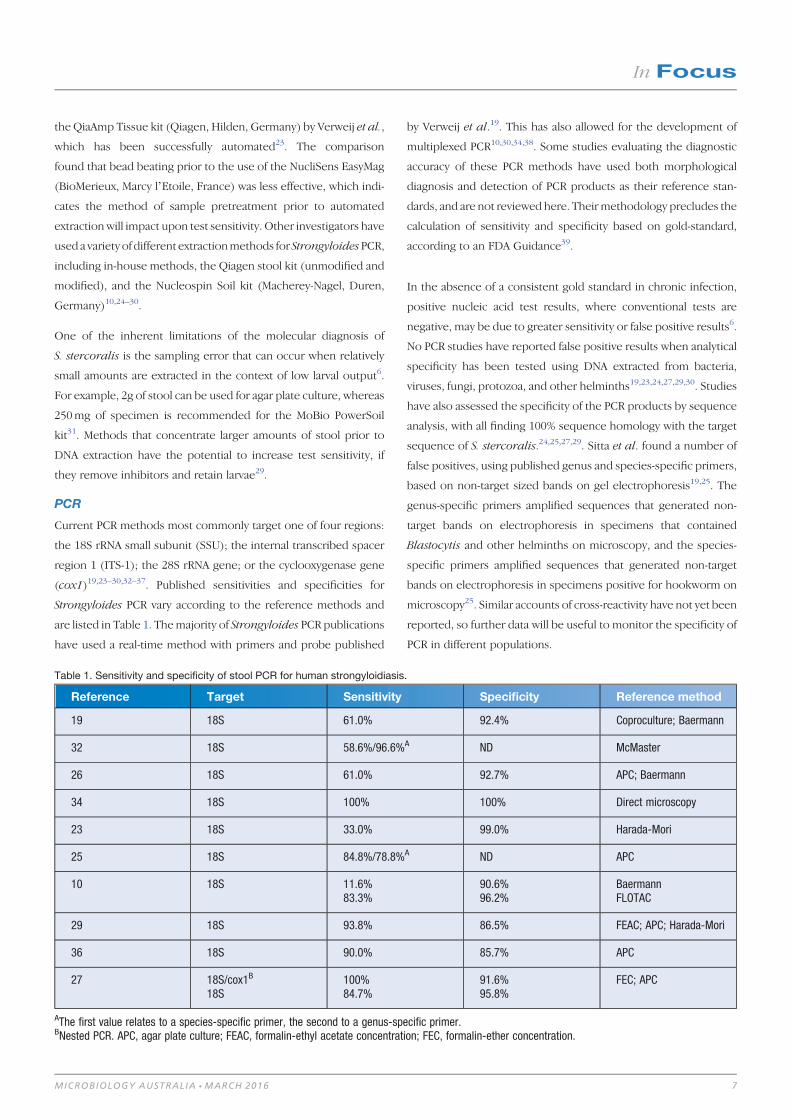
Reference Target Sensitivity Specificity Reference method
19 18S 61.0% 92.4% Coproculture; Baermann
32 18S 58.6%/96.6%A ND McMaster
26 18S 61.0% 92.7% APC; Baermann
34 18S 100% 100% Direct microscopy
23 18S 33.0% 99.0% Harada-Mori
25 18S 84.8%/78.8%A ND APC
10 18S 11.6%83.3%
90.6%96.2%
BaermannFLOTAC
29 18S 93.8% 86.5% FEAC; APC; Harada-Mori
36 18S 90.0% 85.7% APC
27 18S/cox1B
18S100%84.7%
91.6%95.8%
FEC; APC
AThe first value relates to a species-specific primer, the second to a genus-specific primer.BNested PCR. APC, agar plate culture; FEAC, formalin-ethyl acetate concentration; FEC, formalin-ether concentration.
In Focus
MICROBIOLOGY AUSTRALIA * MARCH 2016 7

LAMP
Loop-mediated isothermal amplification (LAMP) is an additional
nucleic acid detection method. LAMP uses a DNA polymerase with
strand-displacement activity, so it doesn’t require the temperature
cycling of PCR, and can be performed with a simple source of
constant temperature such as a heating block40. LAMP has been
successfully applied in resource limited-settings for the detection
of pathogens40.
The Strongyloides LAMP assay uses primers that are genus specific
and bind to the 28S rRNA gene20. The reaction runs at 608C for
1 hour. Pre-heating of the reagents andDNA template to 958C, prior
to the addition of enzyme, increases the limit of detection and
eliminates the need to pre-heat the template and keep it at 48C20.
A novel use of Syto-82 dye (Life Technologies, Carlsbad, CA, USA)
enables the detection of positive results in real-time or visually on
completionof the reaction20. Analytical sensitivity and specificity are
comparable to PCR, according to themethodof Verweij et al.19,20,23.
When 28 human stool specimens that were microscopy and PCR
positive for S. stercoralis were tested with the LAMP method, 27
were positive20. The negative specimen had a high cycle threshold
(38.44) on PCR20. Further validation of the LAMP assay with clinical
specimens is currently in progress.
Conclusion
The diagnosis of strongyloidiasis can be made through the mor-
phological identification of larvae, usually in the stool, serological
testing, and nucleic acid tests. While each methodology has advan-
tages, there are limitations that need to be taken into account when
assessing the significance of negative test results. Often the most
important aspect of patient management is to consider the possi-
bility of S. stercoralis infection.
References1. Olsen, A. et al. (2009) Strongyloidiasis – the most neglected of the neglected
tropical diseases? Trans. R. Soc. Trop. Med. Hyg. 103, 967–972. doi:10.1016/
j.trstmh.2009.02.013
2. Adams, M. et al. (2003) Strongyloidiasis: an issue in Aboriginal communities.
Rural Remote Health 3, 152.
3. Page, W.A. et al. (2006) Utility of serological follow-up of chronic strongyloidiasis
after antihelminthic chemotherapy.Trans.R. Soc. Trop.Med.Hyg.100, 1056–1062.
doi:10.1016/j.trstmh.2005.12.006
4. Bailey, M.S. et al. (2006) Helminth infections in British troops following an
operation in Sierra Leone. Trans. R. Soc. Trop. Med. Hyg. 100, 842–846.
doi:10.1016/j.trstmh.2005.10.001
5. Naidu, P et al. (2013) Eosinophilia: a poor predictor of Strongyloides infection in
refugees. Can. J. Infect. Dis Med. Microbiol. 24, 93–96.
6. Dreyer, G. et al. (1996) Patterns of detection of Strongyloides stercoralis in stool
specimens: implications for diagnosis and clinical trials. J. Clin. Microbiol. 34,
2569–2571.
7. Siddiqui, A.A. and Berk, S.L. (2001) Diagnosis of Strongyloidies stercoralis
infection. Clin. Infect. Dis. 33, 1040–1047. doi:10.1086/322707
8. Inês, EdeJ. et al. (2011) Efficacy of parasitological methods for the diagnosis
of Strongyloides stercoralis and hookworm in faecal specimens. Acta Trop. 120,
206–210. doi:10.1016/j.actatropica.2011.08.010
9. Anamnart, W. et al. (2010) Factors affecting the recovery of Strongyloides
stercoralis larvae: an approach to a newly modified formalin-ether concentra-
tiontechnique for diagnosis of strongyloidiasis. J. Clin. Microbiol. 48, 97–100.
doi:10.1128/JCM.01613-09
10. Knopp, S. et al. (2014) Diagnostic accuracy of Kato-Katz, FLOTAC, Baermann, and
PCR methods for the detection of light-intensity hookworm and Strongyloides
stercoralis infections in Tanzania. Am. J. Trop. Med. Hyg. 90, 535–545.
doi:10.4269/ajtmh.13-0268
11. Steinmann, P. et al. (2007) Occurrence of Strongyloides stercoralis in Yunnan
Province, China, and comparison of diagnostic methods. PLoS Negl. Trop. Dis. 1,
e75. doi:10.1371/journal.pntd.0000075
12. Khieu, V. et al. (2013) Diagnosis, treatment and risk factors of Strongyloides
stercoralis in schoolchildren in Cambodia. PLoS Negl. Trop. Dis. 7, e2035.
doi:10.1371/journal.pntd.0002035
13. Hirata, T. et al. (2007) Short report: increased detection rate of Strongyloides
stercoralis by repeated stool examinations using the agar plate culture method.
Am. J. Trop. Med. Hyg. 77, 683–684.
14. Bisoffi, Z. et al. (2014) Diagnostic accuracy of five serologic tests for Strongyloides
stercoralis infection. PLoS Negl. Trop. Dis. 8, e2640. doi:10.1371/journal.pntd.
0002640
15. Krolewiecki, A.J. et al. (2010) Improved diagnosis of Strongyloides stercoralis
using recombinant antigen-based serologies in a community-wide study in
northern Argentina. Clin. Vaccine Immunol. 17, 1624–1630. doi:10.1128/
CVI.00259-10
16. Requena-Méndez, A, Chiodini, P, Bisoffi, Z, Buonfrate, D, Gotuzzo, E andMunoz, J
(2013) The laboratory diagnosis and follow up of strongyloidiasis: a systematic
review. PLoS Negl Trop Dis. 7, e2002.
17. Sudarshi, S. et al. (2003) Clinical presentation and diagnostic sensitivity of
laboratory tests for Strongyloides stercoralis in travellers compared with immi-
grants in a non-endemic country. Trop. Med. Int. Health 8, 728–732. doi:10.1046/
j.1365-3156.2003.01069.x
18. Schaffel, R. et al. (2001) The value of an immunoenzymatic test (enzyme-
linked immunosorbent assay) for the diagnosis of strongyloidiasis in patients
immunosuppressed by haematologic malignancies. Am. J. Trop. Med. Hyg. 65,
346–350.
19. Verweij, J.J. et al. (2009) Molecular diagnosis of Strongyloides stercoralis in faecal
samples using real-time PCR. Trans. R. Soc. Trop. Med. Hyg. 103, 342–346.
doi:10.1016/j.trstmh.2008.12.001
20. Watts, M.R. et al. (2014) A loop-mediated isothermal amplification (LAMP) assay
for Strongyloides stercoralis in stool that uses a visual detection method with
SYTO-82 fluorescent dye. Am. J. Trop. Med. Hyg. 90, 306–311. doi:10.4269/ajtmh.
13-0583
21. Herwaldt, B.L. (2001) Laboratory-acquired parasitic infections from accidental
exposures. Clin. Microbiol. Rev. 14, 659–688. doi:10.1128/CMR.14.3.659-688.
2001
22. Schrader, C. et al. (2012) PCR inhibitors – occurrence, properties and removal.
J. Appl. Microbiol. 113, 1014–1026. doi:10.1111/j.1365-2672.2012.05384.x
23. Sultana, Y. et al. (2013) Real-time polymerase chain reaction for detection of
Strongyloides stercoralis in stool. Am. J. Trop. Med. Hyg. 88, 1048–1051.
doi:10.4269/ajtmh.12-0437
24. Repetto, S.A. et al. (2013) An improved DNA isolation technique for PCR
detection of Strongyloides stercoralis in stool samples. Acta Trop. 126,
110–114. doi:10.1016/j.actatropica.2013.02.003
25. Sitta, R.B. et al. (2014) Conventional PCR for molecular diagnosis of human
strongyloidiasis. Parasitology 141, 716–721. doi:10.1017/S0031182013002035
26. Schär, F. et al. (2013) Evaluation of real-time PCR for Strongyloides stercoralis
and hookworm as diagnostic tool in asymptomatic schoolchildren in Cambodia.
Acta Trop. 126, 89–92. doi:10.1016/j.actatropica.2012.12.012
27. Sharifdini, M. et al. (2015) Comparison of nested polymerase chain reaction and
real-time polymerase chain reaction with parasitological methods for detection of
Strongyloides stercoralis in human fecal samples. Am. J. Trop. Med. Hyg. 93,
1285–1291.
In Focus
8 MICROBIOLOGY AUSTRALIA * MARCH 2016

28. Angal, L. et al. (2015) Determining intestinal parasitic infections (IPIs) in inmates
from Kajang Prison, Selangor, Malaysia for improved prison management. BMC
Infect. Dis. 15, 467. doi:10.1186/s12879-015-1178-3
29. Saugar, J.M. et al. (2015) Application of real-time PCR for the detection of
Strongyloides spp. in clinical samples in a reference center in Spain. Acta Trop.
142, 20–25. doi:10.1016/j.actatropica.2014.10.020
30. Janwan, P. et al. (2011) Rapid detection of Opisthorchis viverrini and Strongy-
loides stercoralis in human fecal samples using a duplex real-time PCR and
melting curve analysis. Parasitol. Res. 109, 1593–1601. doi:10.1007/
s00436-011-2419-z
31. Koga, K. et al. (1991) Amodified agar plate method for detection of Strongyloides
stercoralis. Am. J. Trop. Med. Hyg. 45, 518–521.
32. Marra, N.M. et al. (2010) Faecal examination and PCR to detect Strongyloides
venezuelensis in experimentally infected Lewis rats. Mem. Inst. Oswaldo Cruz
105, 57–61. doi:10.1590/S0074-02762010000100008
33. Basuni, M. et al. (2011) A pentaplex real-time polymerase chain reaction assay for
detection of four species of soil-transmitted helminths. Am. J. Trop. Med. Hyg. 84,
338–343. doi:10.4269/ajtmh.2011.10-0499
34. Mejia, R. et al. (2013) A novel, multi-parallel, real-time polymerase chain reaction
approach for eight gastrointestinal parasites provides improved diagnostic
capabilities to resource-limited at-risk populations. Am. J. Trop. Med. Hyg. 88,
1041–1047. doi:10.4269/ajtmh.12-0726
35. Zueter, A.M. et al. (2014) Detection of Strongyloides stercoralis infection among
cancer patients in a major hospital in Kelantan, Malaysia. Singapore Med. J. 55,
367–371. doi:10.11622/smedj.2014088
36. de Paula, F.M. et al. (2015)Molecular diagnosis of strongyloidiasis in tropical areas:
a comparison of conventional and real-time polymerase chain reaction with
parasitological methods. Mem. Inst. Oswaldo Cruz 110, 272–274. doi:10.1590/
0074-02760140371
37. Moghaddassani, H. et al. (2011) Molecular diagnosis of Strongyloides stercoralis
infection by PCR detection of specific DNA in human stool samples. Iran.
J. Parasitol. 6, 23–30.
38. Basuni, M. et al. (2012) Detection of selected intestinal helminths and protozoa at
Hospital Universiti Sains Malaysia using multiplex real-time PCR. Trop. Biomed.
29, 434–442.
39. U.S. FoodandDrugAdministration (2007) Statistical guidanceon reporting results
from studies evaluating diagnostic tests. http://www.fda.gov/RegulatoryInforma-
tion/Guidances/ucm071148.htm.
40. Mori, Y. et al. (2013) Loop-mediated isothermal amplification (LAMP): recent
progress in research and development. J. Infect. Chemother. 19, 404–411.
doi:10.1007/s10156-013-0590-0
Biographies
Matthew Watts is an Infectious Diseases Physician and Clinical
Microbiologist based at the Centre for Infectious Diseases and
Microbiology, PathologyWest-ICPMRand theMarieBashir Institute,
University of Sydney, Westmead Hospital. His interests include
parasitic and zoonotic infections.
Gemma Robertson is a final year microbiology trainee at Mel-
bourne Pathology. She has an interest in tropical medicine and
parasitology, andwill beundertaking aPhD tocontinueher research
into soil-transmitted helminthiases in Aboriginal communities.
Dr Richard Bradbury is an Australian Parasitologist with an
interest in all fields of parasitology. He was recently appointed as
the Team Lead in the Parasite Diagnostics and Biology Laboratory
of the Centers for Disease Control and Prevention in Atlanta, USA.
He is writing this work in both his personal capacity and in his
capacity as an adjunct academic at Central Queensland University.
In Focus
MICROBIOLOGY AUSTRALIA * MARCH 2016 9

Current WHO protocols for mass drugadministration in helminth control
Richard S Bradbury
School of Medical and AppliedSciencesCentral Queensland UniversityRockhampton, Qld, AustraliaEmail: [email protected]
Patricia M Graves
College of Public HealthMedical and Veterinary SciencesDivision of Tropical Health andMedicineJames Cook UniversityCairns Qld, AustraliaEmail: [email protected]
Soil transmitted helminths (STH), comprising Ascaris,
Trichuris, Strongyloides and the hookworms remain a sig-
nificant cause ofmorbidity amongst people inmany parts of
the world, including Australia. Other important helminth
infections include lymphatic filariasis (LF), schistosomiasis
and onchocerciasis. Preventive chemotherapy (mass drug
administration [MDA]) campaigns are frequently conducted
for thesehelminth infections inendemicareas,but the target
population groups, duration of campaigns, cointerventions
(e.g. vector control) criteria for inclusion, drugs used and
doses of drugs differ.
The benefits of deworming individuals, especially children, who are
infected with soil-transmitted helminths and schistosomiasis,
include reduction in anaemia and improved growth. Rarer, but
more severe presentations, such as intestinal obstruction with
A. lumbricoides and rectal prolapse due to T. trichiura infection,
will also be reduced1. Treatment for filariasis and onchcocerciasis
in childhood will prevent the development of later severe conse-
quences of these diseases including lymphoedema, hydrocoele,
elephantiasis and blindness.
In situations ofmoderate to high endemicity, it has been considered
more efficient and cost-effective to treat the entire eligible popu-
lation in particular age groups or communities for these diseases,
rather thanfirst testing individuals todeterminewho is infected.The
goals of such MDA are morbidity control in some cases and inter-
ruption of transmission through vectors in others. For these rea-
sons, MDA for STH is usually undertaken for school age children,
while for schistosomiasis, MDA is performed either in children or
in eligible people of all ages, depending on the endemicity level.
For the insect borne helminths, onchocerciasis and LF, the goal is
transmission interruption and the entire community is eligible for
MDA.
The frequency of MDA for STH in school-age children is dependent
on the prevalence of infections in a given population (Table 1).
Current WHO protocols for STH control recommend MDA with a
single oral dose of albendazole (400mg), mebendazole (500mg) or
levamisole (80mg)1. Mebendazole is more effective than albenda-
zole for T. trichiura, whilst albendazole is slightly more effective
against hookworm than mebendazole. The two drugs have equally
Table 1. Current recommended regularity of mass drug administration(MDA) for helminth (STH) infections in school-ageA children (adaptedfrom WHO 2011)1.
PrevalenceB Regularity of MDA
For control of soil-transmitted helminth (STH) infections
�50% Twice per year, or every 4 months if at thehigh end of prevalence
�20 and <50% Once yearly
<20% Treat on case-by-case basis
For control of schistosomiasis
�50% Once per year, or every 4 months if at thehigh end of prevalence
�10 and <50%B Once every 2 years
<10% Treat on case-by-case basis
AUsually defined as children between 5 and 14 years of age.BAs determined by parasitological methods; cut-off is�30% if based only onquestionnaires for visible haematuria.
Under theMicroscope
10 10.1071/MA16004 MICROBIOLOGY AUSTRALIA * MARCH 2016

high efficacy when used against Ascaris lumbricoides1. Despite
these differences, in practice when administered biennially over a
number of years, either drug used on its own is effective for overall
STH control1.
Onchocerciasis control and/or elimination requires annual or bian-
nual treatment with ivermectin for many years (up to 20 in some
cases), while lymphatic filariasis uses annual administration with
albendazole and either ivermectin or diethylcarbamazine (DEC)
with at least 65% population coverage for at least five years. Thus
ivermectin and/or albendazole administration may occur in some
communities annually or biannually as part of onchocerciasis or LF
elimination programs. Where this occurs, STH control programs
should be harmonised to ensure that there is a 6 month delay
between albendazole administrations1. In communities with en-
demic schistosomiasis, the addition of praziquantel (40mg/kg) is
recommended (Table 1). Reductions in the frequency of MDA may
be considered after 5–6 years of consistent >75% population cov-
erage, after testing of the residual prevalence of helminths in that
population. Such a decision is based on several factors, specific
details of which may be found in the World Health Organization
guide for managers of control programmes1.
Reliance on albendazole and mebendazole in WHO recommenda-
tions for MDA will result in a lower impact on the clearance of
Strongyloides stercoralis, for which ivermectin and thiabendazole
are more effective drugs2. Thiabendazole was discontinued in
Australia in 2003 anddue to the lower rate of side effects, ivermectin
has been recommended by some as the treatment of choice2. Due
to the auto-infective cycle of this helminth, some authors have
recommended re-treatment at one and two months to ensure
elimination3. Only one randomised trial has been performed thus
far, in which treatment twice at 2 weeks apart was found to have no
greater benefit than a single dose4. Both the authors of this paper
and others2 recommend further studies into the optimal dose
schedules for ivermectin in the control of Strongyloidiasis within
a larger cohort of participants.
Recently, there has been discussion about the outcomes and
optimal age range for MDA programs to control STH and more
evidence on the impacts is required5–7; this controversy is outside
the scope of this review. A novel concept of elimination of STH by
MDA ‘one village at a time’ rather than by wide-scale MDA has been
proposed for remote areas with low populations, such as many
remote Australian Aboriginal communities. This approach is cur-
rently being trialled in a remote area of the Solomon Islands8. It
advocates allowing individual communities to act as autonomous
units and to employ control options specifically tailored to the
geographic, cultural, economic, aetiological and environmental
factors influencing STH transmission in their own community8
(Figure 1).
Deworming of school-age children has been amainstay of helminth
control for many years. Discussion continues on the optimal meth-
od ofMDA for this purpose. In some cases, such as where high rates
of strongyloidiasis or onchocerciasis are present, the addition of
other drugs may be warranted. Further consideration of new ther-
apies, combination therapies, the reconsideration of use of the
use of ‘old’ anti-helminthic therapies have all been postulated as
mechanisms by which to improve absolute cure rates in MDA
programsand to reduce thepossibledevelopmentof antihelminthic
resistance9.Ways to improveMDAparticipation, aswell as paediatric
formulations of praziquantel for schistosomiasis prevention in pre-
school children may also be added to this list. The current WHO
protocols for MDA provide an important baseline guide to those
undertaking MDA for helminth control in endemic areas.
References1. World Health Organization (2011) Helminth control in school-age children:
a guide for managers of control programmes. 2nd edn. Geneva: World Health
Organization.
2. Bisoffi, Z. et al. (2011) Randomized clinical trial on ivermectin versus thiabendazole
for the treatment of strongyloidiasis. PLoS Negl. Trop. Dis. 5, e1254. doi:10.1371/
journal.pntd.0001254
3. Adams, M. et al. (2003) Strongyloidiasis: an issue in Aboriginal communities. Rural
Remote Health 3, 152.
4. Suputtamongkol, Y. et al. (2011) Efficacy and safety of single and double doses of
ivermectin versus 7-day high dose albendazole for chronic strongyloidiasis. PLoS
Negl. Trop. Dis. 5, e1044. doi:10.1371/journal.pntd.0001044
5. Anderson, R.M. et al. (2015) Should the goal for the treatment of soil transmitted
Helminth (STH) infections be changed from morbidity control in children to
community-wide transmission elimination? PLoS Negl. Trop. Dis. 9, e0003897.
doi:10.1371/journal.pntd.0003897
6. Taylor-Robinson, D.C. et al. (2015)Deworming drugs for soil-transmitted intestinal
worms in children: effects on nutritional indicators, haemoglobin and school
Figure 1. A Solomon Islander researcher undertaking a community wideSTH prevalence survey as part of an integrated STH control program(photograph by Richard Bradbury).
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 11

performance. Cochrane Database Syst. Rev. 7, CD000371. doi:10.1002/14651858.
CD000371.pub6
7. Hicks, J.H. et al. (2015) The case for mass treatment of intestinal helminths in
endemic areas PLoS Negl. Trop. Dis. 9, e0004214. doi:10.1371/journal.pntd.000
4214
8. Harrington, H. et al. (2015) Prevalence of soil transmitted helminths in remote
villages in East Kwaio, Solomon Islands. Western Pac. Surveill. Response J. 6, 1–8.
doi:10.5365/wpsar.2015.6.1.016
9. Savioli, L. (2014) Preventive anthelmintic chemotherapy – expanding the arma-
mentarium. N. Engl. J. Med. 370, 665–666. doi:10.1056/NEJMe1312403
Biographies
Dr Richard Bradbury is an Australian Parasitologist with an
interest in all fields of parasitology. He was recently appointed as
the Team Lead in the Parasite Diagnostics and Biology Laboratory
of the Centers for Disease Control and Prevention in Atlanta, USA.
He is writing this work in both his personal capacity and in his
capacity as an adjunct academic at Central Queensland University.
Dr Patricia Graves is a vector-borne disease epidemiologist with
research, field, laboratory and consulting experience in the control
and elimination of malaria and filariasis. She is Director of the
JCU/WHO Collaborating Centre for Control of LF, STH and other
NTDs in the Division of Tropical Health and Medicine, James
Cook University.
Zoonotic tissue parasites of Australian wildlife
David M Spratt
Australian National WildlifeCollectionNational Research CollectionsAustralia, CSIROGPO Box 1700Canberra, ACT 2601, AustraliaTel: +61 2 6242 1648Email: [email protected]
Increasing use of bushlands for recreational, commercial
and scientific activities fosters movement across the urban-
bushland interface. This may facilitate the transmission of
parasitic diseases from wildlife to humans (zoonoses). The
fashionable trend to consumption of game meats such as
feral pig and crocodile, and raw fish such as sushi, sashimi
and pickled herring has exacerbated the zoonotic potential
of parasites of wildlife.
Transmission from wildlife to humans
Angiostrongyliasis
Angiostrongylus cantonensis is a nematode parasite of the pulmo-
nary arteries and right ventricle ofRattus rattus andR. norvegicus in
Australia1. It is the causative agent of eosinophilic meningoenceph-
alitis, a zoonotic infection of humans. The life cycle includes an
obligatory period of larval development in terrestrial or aquatic
snails and slugs, and also may involve a range of paratenic or
transport hosts (freshwater prawns, land crabs, planarians, frogs,
lizards), which feed on gastropods. Rats become infected by ingest-
ing intermediate or paratenic hosts. In the rat, the nematode
undergoes an obligatory migration through the spinal column and
brain en route to the final site in the pulmonary arteries of the lungs.
Humans become infected by accidentally or deliberately eating
infected gastropods or paratenic hosts, or unwashed salad greens
containing these.Theparasitehasbeen reported fromdomestic and
zoo animals, mammalian and avian wildlife and humans in Brisbane
and Sydney2–4. The clinical signs of headache, vomiting, paralysis
and sometimes death are induced as a consequence of the oblig-
atory period of development of the parasite in the central nervous
system. This occurs in young children who deliberately or acciden-
tally ingest snails or slugs containing infective larvae5–7, or foolish
young adults who do so for a bet8,9.
Muspiceoidosis
Haycocknema perplexum is aminutemuspiceoid nematode living
as adults inside individual skeletal muscle cells of humans in
Australia10. Eight cases have been documented, 4 in Tasmania and
4 in north Queensland11–13 Gasser (personal communication).
Eight to twelve eggshatch inside theuterusof the female, develop to
third-stage infective larvae andburst fromthehead regionkilling the
adult, an efficient mechanism for auto-re-infection. Escaped larvae
invade uninfected muscle cells. The occurrence ofH. perplexum in
intramyofibres results in eosinophilic polymyositis but no reaction
within the invaded cell itself11. Progressive myopathy occurs and
infection becomes life threatening. Early human diagnosis by mus-
cle biopsy is imperative in cases of progressivemyopathy associated
with blood eosinophilia and elevated creatine kinase levels. Steroid
Under theMicroscope
12 10.1071/MA16005 MICROBIOLOGY AUSTRALIA * MARCH 2016

treatment of patients exacerbates their infections to a life-threat-
ening illness and may delay diagnosis by masking key diagnostic
features. Treatment with albendazole, 400mg twice daily for 8–
9 weeks, is recommended12. Haycocknema perplexum is consid-
ered a zoonosis although the source of infection of humans –water,
soil, plants or animals – remains unknown.
Halicephalobus gingivalis
Halicephalobus gingivalis formerly known asMicronemadeletrix,
is a free-livingnematodeof soil,manure anddecayinghumusknown
to cause opportunistic infections, primarily in horses but also in
humans14.Themajorityof cases inhorseshavebeen fatal andusually
not diagnosed before necropsy. All human cases have involved fatal
meningoencephalitis including the first human case in Australia, a
74-year-old woman from Eyre Penninsula, South Australia15. In
tissues, only ova, larvae and adult females are seen, reproduction
in the parasitic phase presumed to be by parthenogenesis16. It is not
known how H. gingivalis infects humans or horses although expo-
sure through an oromaxillary route may explain common neuro-
logical involvement15.
Transmission from domestic animals to humans
and wildlife
Toxoplasmosis
Felids, domestic cats in particular, are the only definitive host of the
obligate intracellular protozoan parasite, Toxoplasma gondii17.
Most species of mammals and birds are susceptible to infection
and may act as intermediate hosts. Infection is usually systemic
resulting in a short period of rapid multiplication in various tissues
followed by the establishment of tissue cysts in the muscles and
brain. These are transmitted only if ingested by predation or
scavenging or if passed vertically across the placenta from mother
to foetus.
The localisation of T. gondii cysts in the forebrain of rats and mice
together with the immune reaction to the cysts is related to altered
behaviour, in particular the attenuation of predator odour aversion
and anxiety. This facilitates ingestion of the intermediate host by the
cat definitive host and perpetuation of the life cycle17.
Humans become infected with T. gondii mainly by ingesting un-
cooked meat containing viable tissue cysts or by ingesting food or
water contaminated with oocysts from the faeces of infected cats.
Transplacental transmission may occur in women during their first
trimester of pregnancy and this infection is then passed to the
developingembryovia theplacenta. This frequently results in severe
brain and eye lesions in the newborn or death of the developing
embryo. Research results from the past 15 years have shown that
T. gondii infection is associated with several neuropsychiatric
diseases and behavioural changes in humans as well animals18.
Although the mechanisms are unknown a growing body of data
indicates that they are complex, comprising humoral, immune,
neurotransmitter, epigenetic, genetic, and structural effects19.
Echinococcosis (hydatid disease)
Australia has, on average, >80 new cases of human hydatidosis per
annum caused by the larval stage of the cestode, Echinococcus
granulosus20. The parasite was introduced into Australia with
domestic livestock and dogs. However, a cycle in wildlife is main-
tained through a predator/prey interaction between dingoes, wild
dogs and less importantly foxes and kangaroos and wallabies, less
importantly feral pigs (Sus scrofa)21,22. The establishment of a
dingo/wild dog-macropod cycle, which effectively maintains para-
site transmission, acts as a spill-back reservoir of infection for sheep
and cattle23. This is a major problem for control strategies focussed
onhumaneducation andhusbandrypractices to break thedomestic
‘dog-sheep’ cycle. In contrast to the situation in livestock, hydatid
cysts occur primarily in the lungs rather than the liver of marsupials.
Infectionoccurspredominantly in theeasternStatesbut theparasite
has established recently in wildlife in water catchment and forestry
areas outside Perth21. Western grey kangaroos and feral pigs act as
intermediate hosts. This focus of transmission may have been
initiated through E. granulosus-infected domestic pig hunting dogs
Table 1. Potential zoonotic tissue parasite infections in Australia.
Disease name Organism Type Source Reference
Leishmaniasis Leishmania sp. Trypanosomatid Forcipomyia spp. 26,27
Protozoan
Sparganosis Spirometra erinaceieuropaei Cestode (larval, plerocercoid) Eating raw/undercooked meat 28
Pentastomiasis Armillifer spp. Modified brachiuran Eating undercooked snake/mammal flesh, drinkingcontaminated water
29,30,31
Linguatula spp. Crustacean
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 13

from the eastern states. Transmission appears to be perpetuated by
dogs of local pig hunters infected through being fed the offal of
locally shot kangaroos21. Hydatid disease is also an important
conservation issue, especially for small endangered species and
populations of Macropodidae by severely reducing effective lung
volume24,25. Such reductions impact the fitness of the animals
enhancing susceptibility to predation, thus ensuring perpetuation
of the cycle.
Several potential zoonotic tissue parasite infections in Australia are
listed in Table 1.
References1. Mackerras, M.J. and Sandars, D.F. (1955) The life history of the rat lung-worm,
Angiostrongylus cantonensis (Chen) (Nematoda: Metastrongylidae). Aust.
J. Zool. 3, 1–21. doi:10.1071/ZO9550001
2. Spratt, D.M. (2005a) Australian ecosystems, capricious food chains and parasitic
consequences for people. Int. J. Parasitol.35, 717–724. doi:10.1016/j.ijpara.2005.
01.014
3. Spratt, D.M. (2005b) Neuroangiostrongyliasis: disease in wildlife and humans.
Microbiol. Aust. 26, 63–64.
4. Ma, G. (2013) Tawny frogmouths and brushtail possums as sentinels for Angios-
trongylus cantonensis, the rat lungworm. Vet. Parasitol. 192, 158–165.
doi:10.1016/j.vetpar.2012.11.009
5. Prociv, P. and Tiernan, J.R. (1987) Eosinophilic meningoencephalitis with per-
manent sequelae. Med. J. Aust. 147, 294–295.
6. Prociv, P. et al. (2000) Neuro-angiostrongyliasis: unresolved issues. Int.
J. Parasitol. 30, 1295–1303. doi:10.1016/S0020-7519(00)00133-8
7. Morton, N.J. et al. (2013) Severe haemorrhagic meningoencephalitis due to
Angiostrongylus cantonensis among young children in Sydney, Australia. Clin.
Infect. Dis. 57, 1158–1161. doi:10.1093/cid/cit444
8. Senanayake, S.N. (2003) First case of human angiostrongyliasis acquired in
Sydney. Med. J. Aust. 179, 430–431.
9. Blair, N.F. et al. (2013) Angiostrongylus meningoencephalitis: survival from
minimally conscious state to rehabilitation. Med. J. Aust. 198, 440–442.
doi:10.5694/mja12.11085
10. Spratt, D.M. et al. (1999) Haycocknema perplexum n.g., n. sp. (Nematoda:
Robertdollfusidae): an intramyofibre parasite in man. Syst. Parasitol. 43,
123–131. doi:10.1023/A:1006158218854
11. Dennett, X. et al. (1998) Polymyositis caused by a new nematode. Med. J. Aust.
168, 226–227.
12. Basuroy, R. et al. (2008) Parasitic myositis in tropical Australia. Med. J. Aust. 188,
254–256.
13. McKelvie, P. et al. (2013) A further patient with parasitic myositis due to Hay-
cocknemaperplexum, a rare entity. J. Clin.Neurosci.20, 1019–1022.doi:10.1016/
j.jocn.2012.08.009
14. Anderson, R.C. et al. (1998) Halicephalobus gingivalis (Stefanski, 1954) from a
fatal infection of a horse in Ontario, Canada with comments on the validity of
H. deletrix and a review of the genus. Parasite 5, 255–261. doi:10.1051/parasite/
1998053255
15. Lim, C.K. et al. (2015) First human case of fatal Halicephalobus gingivalis
meningoencephalitis in Australia. J. Clin. Microbiol. 53, 1768–1774. doi:10.1128/
JCM.00032-15
16. Blunden, A.S. et al. (1987) Halicephalobus deletrix infection in a horse. Equine
Vet. J. 19, 255–260. doi:10.1111/j.2042-3306.1987.tb01399.x
17. Dubey, J.P. et al. (1998) Structures of Toxoplasma gondii tachyzoites, brady-
zoites, and sporozoites and biology and development of tissue cysts. Clin.
Microbiol. Rev. 11, 267–299.
18. Evans, A.K. (2014) Patterns of Toxoplasma gondii cyst distribution in the
forebrain associated with individual variation in predator odor avoidance and
anxiety-related behavior in male Long-Evans rats. Brain Behav. Immun. 37,
122–133. doi:10.1016/j.bbi.2013.11.012
19. Hinze-Selch, D. (2015) Toxoplasma gondii infection and neuropsychiatric dis-
ease: current insight. Rep. Parasitol. 4, 43–51. doi:10.2147/RIP.S52980
20. Thompson, R.C.A. and McManus, D.P. (2002) Towards a taxonomic revision of
the genus Echinococcus. Trends Parasitol.18, 452–457. doi:10.1016/S1471-4922
(02)02358-9
21. Thompson, R.C.A. et al. (1988) Hydatid disease in urban areas of Western
Australia: an unusual cycle involving western grey kangaroos (Macropus
fuliginosus), feral pigs and domestic dogs. Aust. Vet. J. 65, 188–190.
doi:10.1111/j.1751-0813.1988.tb14298.x
22. Jenkins, D.J. and Morris, B. (2003) Echinococcus granulosus in and around
the Kosciuszko National Park, south-eastern Australia. Aust. Vet. J. 81, 81–85.
doi:10.1111/j.1751-0813.2003.tb11440.x
23. Durie, P.H. and Riek, R.F. (1952) The role of the dingo and wallaby in the
infestation of cattle with hydatids (Echinococcus granulosus (Batsch, 1786)
Rudolphi, 1805) in Queensland. Aust. Vet. J. 28, 249–254. doi:10.1111/
j.1751-0813.1952.tb13436.x
24. Johnson, P.M. et al. (1998) Mortality in wild and captive rock wallabies and nailtail
wallabies due to hydatid disease caused by Echinococcus granulosus. Aust.
Mammal. 20, 419–423.
25. Barnes, T.S. et al. (2008) Cystic echinococcosis in a wild population of the brush-
tailed rock-wallaby (Petrogale penicillata), a threatened macropodid. Parasitol-
ogy 135, 715–723. doi:10.1017/S0031182008004423
26. Rose, K. (2004) Cutaneous leishmaniasis in red kangaroos: isolation and charac-
terisation of the causative organisms. Int. J. Parasitol. 34, 655–664. doi:10.1016/
j.ijpara.2004.03.001
27. Dougall, A.M. et al. (2011) Evidence incriminating midges (Diptera: Ceratopogo-
nidae) as potential vectors of Leishmania in Australia. Int. J. Parasitol. 41,
571–579. doi:10.1016/j.ijpara.2010.12.008
28. Liu,Q. (2015)Human sparganosis, aneglected foodborne zoonosis. Lancet Infect.
Dis. 15, 1226–1235. doi:10.1016/S1473-3099(15)00133-4
29. Paré, J.A. (2008) An overview of Pentastomiasis in reptiles and other vertebrates.
J. Exot. Pet Med. 17, 285–294. doi:10.1053/j.jepm.2008.07.005
30. Kelehear, C. et al. (2014) Pentastomids of wild snakes in the Australian tropics.
Intl. J. Parasitol.: Para. Wldlfe. 3, 20–31. [with online supplementary table]
31. Drabick, J.J. (1987) Pentastomiasis. Rev. Infect. Dis. 9, 1087–1094. doi:10.1093/
clinids/9.6.1087
Biography
Dave Spratt is an Honorary Fellow at the Australian National
Wildlife Collection, National Research Collections Australia, CSIRO,
in Canberra. Themajor themes of Dr Spratt’s research are the study
and understanding of the diseases of wildlife including disease
ecology, parasite taxonomy, helminth biodiversity and zoonoses.
Research topics of interest includemetastrongyloid, filarioid, trichi-
nelloid and muspiceoid nematodes, pentastomes, and small mam-
mal succession and recolonisation of their helminth communities
following wildfire.
Under theMicroscope
14 MICROBIOLOGY AUSTRALIA * MARCH 2016

Assessing enteric helminths in refugees, asylumseekers and new migrants
Sarah HaniehA,D,*, Norbert RyanB,* and Beverley-Ann BiggsB,C
AVictorian Infectious Diseases Reference Laboratory, Peter Doherty Institute for Immunity and Infection, Parkville, Vic. 3050, Australia
BDepartment of Medicine, The University of Melbourne, Peter Doherty Institute for Immunity and Infection, Parkville, Vic. 3050, Australia
CThe Victorian Infectious Diseases Service, Royal Melbourne Hospital, Parkville, Vic. 3050, Australia
DCorresponding author. Department ofMedicine, The University of Melbourne, Doherty Institute, Parkville, Vic. 3050, Australia, Tel: +61 3 8344 3257,Fax: +61 3 9347 1863, Email: [email protected]
Currently there are 59.5million people forcibly displaced
worldwide as a result of conflict, human rights violations,
generalised violence or persecution. Of these, 19.5million
are refugees and 1.8million are asylum seekers. Each year
Australia accepts 13750 refugees through the offshore
Humanitarian program, and in 2016 that number will
almost double with the addition of 12000 refugees from
Syria and Iraq. Many refugees have complex medical needs
and have reached Australia after a difficult journey, often
involving time in refugee camps and exposure to traumatic
events including physical hardship and illness. Refugees
often come from parts of the world where parasitic and
tropical infectious diseases are prevalent and untreated.
This article provides a review of enteric helminth infections
in refugees, including asylum seekers and those from a
refugee-like background.
Parasitic infections in refugees and new migrants reflect the under-
lying epidemiology of parasites in areas where refugees may have
been exposed, including the country of origin, migration journey to
Australia, andplaceof detention. Factors such aspoverty, disruption
of basic services, poor sanitation/hygiene (e.g. quality of drinking
water, access to running water, access to footwear) and insufficient
access to adequate health care and treatment may significantly
increase the risk of exposure to intestinal parasitic disease in the
refugee population1. Other practices such as the use of night-soil as
fertiliser, dietary habits, and past occupational exposures are likely
to play an important role in increasing the burden of disease. Soil
transmitted helminth (STH) infections are very common in those
living in resource constrained settings1. Treatment of refugees and
asylum seekers is often empirical, in refugee camps, before depar-
ture as part of the pre-departure health check, or after arrival in
Australia. More serious infections, such as strongyloidiasis, schisto-
somiasis, opisthorchus and Taenia solium, require diagnosis
and specific treatment. Table 1 summarises the findings of recent
prevalence studies of strongyloidisis and schistosomiasis in refugee
groups from Australia and overseas.
Strongyloidiasis
The highest prevalence of Strongyloides stercoralis infection
occurs in refugees from Africa and South-East Asia2. Of those
arriving in the last decade, the Burmese groups (e.g. Karen, Chin)
have the highest prevalence (26.0%). Earlier data show an even
*Joint first authors.
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 10.1071/MA16006 15
higher prevalence in Lao and Cambodian refugees3–5. Infections
may persist for decades after leaving an endemic area due to a
continual cycle of auto-infection. Although many patients are
asymptomatic or have minimal clinical symptoms, they remain at
risk for subsequent hyperinfection if immunosuppressed6. Stron-
gyloides antibodies decline after effective treatment7.
Schistosomiasis
The highest prevalence of schistosomiasis is found in Africa, ac-
counting for an estimated 95% of global cases. Species involved are
S. mansoni, S. haematobium and S. intercalatum. S. japonicum is
present along the Yangtze River in China and the Philippines.
S. mekongi is found in the Mekong river valley. Schistosoma
infection may also be encountered in other areas such as parts of
Indonesia, the Caribbean, the Arabian Peninsula, Madagascar, the
Middle east and Turkey8. Prevalence of schistosomiasis infection in
refugees in Australia, as determined by serology, has been shown
to range from 5.4% in Burmese9 to 37% in Africans10.
Schistosoma antibodies are thought to persist in those from en-
demic areas despite prior treatment. A long-term study of schisto-
somiasis serology post-treatment showed an immediate increase in
titre and thena fourfolddecline inmost travellers after 6–12months.
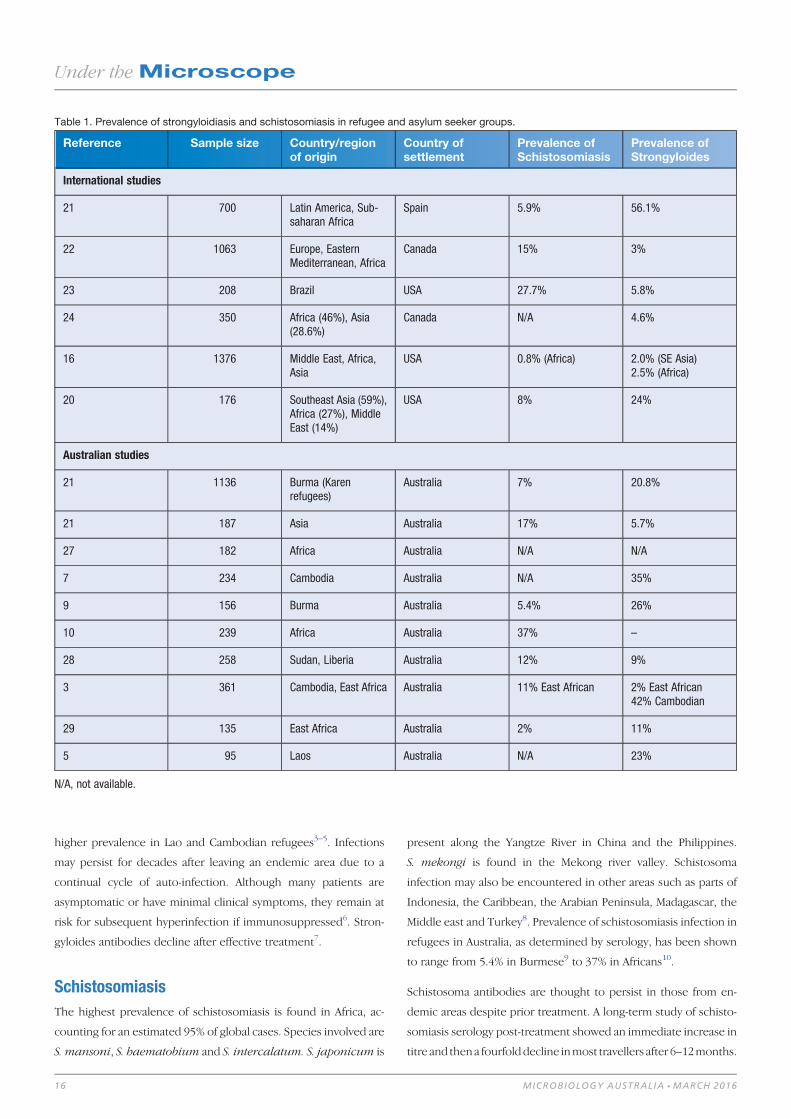
Table 1. Prevalence of strongyloidiasis and schistosomiasis in refugee and asylum seeker groups.
Reference Sample size Country/regionof origin
Country ofsettlement
Prevalence ofSchistosomiasis
Prevalence ofStrongyloides
International studies
21 700 Latin America, Sub-saharan Africa
Spain 5.9% 56.1%
22 1063 Europe, EasternMediterranean, Africa
Canada 15% 3%
23 208 Brazil USA 27.7% 5.8%
24 350 Africa (46%), Asia(28.6%)
Canada N/A 4.6%
16 1376 Middle East, Africa,Asia
USA 0.8% (Africa) 2.0% (SE Asia)2.5% (Africa)
20 176 Southeast Asia (59%),Africa (27%), MiddleEast (14%)
USA 8% 24%
Australian studies
21 1136 Burma (Karenrefugees)
Australia 7% 20.8%
21 187 Asia Australia 17% 5.7%
27 182 Africa Australia N/A N/A
7 234 Cambodia Australia N/A 35%
9 156 Burma Australia 5.4% 26%
10 239 Africa Australia 37% –
28 258 Sudan, Liberia Australia 12% 9%
3 361 Cambodia, East Africa Australia 11% East African 2% East African42% Cambodian
29 135 East Africa Australia 2% 11%
5 95 Laos Australia N/A 23%
N/A, not available.
Under theMicroscope
16 MICROBIOLOGY AUSTRALIA * MARCH 2016

However, for immigrants, serology remained elevated even
three years after effective treatment in a proportion of patients11.
Soil-transmitted helminths (STH)
Many refugees will have received empirical albendazole as part of
the predeparture medical assessment conducted by the Interna-
tional Organisation of Migration on behalf of the Australian
Government. This has significantly altered the prevalence and
patternsof intestinal helminths in refugees12.However, albendazole
has limited effectiveness against Trichuris trichiura, and is not an
effective treatment for some less common helminths found in
refugees and asylum seekers (see below).
Less common helminth infections
Themajority of Asian refugees currently entering Australia are from
Myanmar, from camps on the Thai border. Within Thailand, espe-
cially the north east, Opisthorchis viverrini is highly prevalent.
Infection results from consumption of raw, uncooked or fermented
fish containing metacerciae. Long-term infection may cause cho-
langitis, obstructive jaundice, cholecystitis, periductal fibrosis and
bile duct cancer, contributing to a liver cancer rate in excess of
70 per 100 000 in NE Thailand. There is a paucity of data on faecal
microscopy findings for refugees from Myanmar. However, the
presence of vector cyprinoid fish and substantial wetlands suggests
that infection with Opisthorchis viverrini is also likely.
Other serious infections in refugees from Asia include Taenia
solium (pork tapeworm) with the potential risk of cysticercosis for
both patient and household members. In Thailand faecal tests
for helminth eggs have revealed a prevalence of Taenia spp of
2.3–3.7% in communities with high migrant populations along the
Thai border. In the remote western border area of Kanchanaburi
three species of tapeworm T. saginata, T. solium and T. asiatica
co-exist in the human population13.
Considerations in the Syrian refugee population
In September 2015, it was announced that 12 000 Syrian and Iraqi
refugees would be accepted to Australia as part of theHumanitarian
Program during 2016–17. The recent prevalence of enteric parasite
infections is not well documented in Syria. However, schistosomi-
asis is considered to have low endemnicity in Iraq (0.1%) and
Syria (<10%prevalence in 2010)14. There is no information available
on the prevalence of S. stercoralis in Syria; however, a hospital-
based survey in Iraq reported a prevalence of 24.2%15. Chang et al.
reported a low prevalence of other parasitic infections in refugees
from Iran and Iraq16.
Diagnosis
In Australia, the number of faecal microscopy tests performed has
fallen in some States (e.g. NSW), with more emphasis being placed
on empirical treatment of STH and the use of serology for the
diagnosis of S. stercoralis or Schistosoma spp. Current recom-
mended diagnostic tests for enteric parasites are shown in Table 2.
Challenges and limitations of testing for
schistosoma and strongyloides infections
in a refugee population
Serology may overestimate the prevalence of disease due to cross-
reactivity with other nematode infections and there is difficulty
distinguishing recent from past (and cured) infections. Serological
titres (OD values) in the equivocal and low positive ranges are
difficult to interpret. Follow-up serology should preferably be done
in the same laboratory and in parallel with previous specimens
where available. The interpretation of Schistosoma serology in
Table 2. Recommended diagnostic tests for enteric parasites.
Enteric parasite Diagnosis
Strongyloides stercoralis Positive serologyStool microscopy should be performed to rule out other enteric infections
Schistosoma spp. Positive serologyStool/urine microscopy for ova should be performed in those with positive serology
Hookworm (Necator americanus and Ancylostoma duodenale) Microscopic finding of ova in faecal specimensConcentration methods are necessary to detect light infections
Ascaris lumbricoides Microscopic identification of ova in faecal specimens
Trichuris trichiura Microscopic identification of ova in faecal specimens
Taenia spp. Microscopic identification of ova and proglottids in faecal specimens
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 17

Australia presents several challenges as test methodology varies
between laboratories. VIDRL in Melbourne, use the Fumouze
indirect haemagglutination test (IHA). This is based on an antigen
derived from adult worms of S. mansoni. Estimates in one com-
parison study show that the sensitivity of this test is 76.2% for
Schistosomamansoni, and slightly lower for S. haematobium, with
a specificity of 99%17. However, in NSW at ICPMR, an ‘in house’
ELISA assay is the preferred assay used, based on S. mansoni egg
antigen. Two commercial assays based on the use of a similar
antigen showed 71.4–85.7% sensitivity but reduced specificity of
76.9–88.4%. Both ELISA assays showed cross-reactivity with ces-
tode, nematode and trematode infections17. The sensitivity of these
assays for other species of Schistosoma, such as S. haematobium,
S. intercalatum, S. mekongi and S. japonicum, is not specified;
however, it is likely to be reduced.
The sensitivity and specificity of Strongyloides stercoralis serology
is reported to be up to 94.6% and 99.6% respectively, depending
on the assay used18. However, as there is no gold standard test for
comparison, these are estimations only. Serological titres decline
with effective treatment over a 12 month period7,18.
Persistence of parasitic infections in refugee
populations
Several studies have demonstrated that serious intestinal parasitic
infectionsmay persist formany years after arrival in Australia. A 2002
study of Laotian refugees who had arrived in Australia during
1974–91 showed that strongyloides serology was positive in 24%
and 3 carried Opisthorchis, compared to respective prevalences
of 19.2% and 41%, on initial screening by faecal microscopy4. As
liver flukes survive for approximately 7–10 years, the three cases of
Opisthorchis identified may well represent reinfection on subse-
quent visits to Laos.
In a second study of East African refugees, who arrived in Australia
in the late 90s, screening for strongyloides and schistosoma
serology was positive in 11% and 15%, respectively, of patients
some 16 years later in 20063. In a further aspect of the same study,
42% of 234 Cambodians who had arrived in the late 80s still tested
positive for strongyloides serology3. This compared with 7.8% of
Cambodians who were found to be positive for strongyloides by
microscopy at initial health screening4.
Post arrival health assessment for refugees
and asylum seekers
Community Health Centres and GP services in suburbs with high
rates of migrant and refugee settlement are now responsible for
much of the post-arrival refugee health screening, with support
from specialist Refugee Health Services. Guidelines for post-arrival
assessment for people of refugee-like background were published
by the Australian Society for Infectious Diseases in 2009 and are
currently being revised19. If possible, a full health assessment of new
arrivals is ideally conducted within onemonth of arrival in Australia,
including serology for Strongyloides for all, and Schistosoma in
those who have lived or travelled through an endemic area. For
many of the clinics in suburbs with high migrant populations,
the special pathology requirements are met by private pathology
providers.
Summary
Intestinal helminth infections in refugees are common and should
remain a high priority for health workers. These populations often
have specific needs that should be considered in diagnosis and
managementof these infections.Burdenofdisease is likely to reflect
the country of origin, journey of migration to Australia, pre-depar-
ture treatment and place of detention. Other socio-economic and
cultural factors are also likely to play a significant role in exposure
risk. Helminth infections may be chronic and persist in humans for
more than four decades resulting in seriousmorbidity andmortality
and highlighting the need for early diagnosis.
Acknowledgements
We thank Ms Christalla Hajisava for technical assistance.
References1. Freeman, M.C. et al. (2015) Associations between school- and household-level
water, sanitation and hygiene conditions and soil-transmitted helminth infection
among Kenyan school children. Parasit. Vectors 8, 412. doi:10.1186/
s13071-015-1024-x
2. Puthiyakunnon, S. et al. (2014) Strongyloidiasis–an insight into its global prev-
alence and management. PLoS Negl. Trop. Dis. 8, e3018. doi:10.1371/journal.
pntd.0003018
3. Caruana, S.R. et al. (2006) Undiagnosed and potentially lethal parasite infections
among immigrants and refugees in Australia. J. Travel Med. 13, 233–239.
doi:10.1111/j.1708-8305.2006.00045.x
4. Ryan, N. et al. (1988) Parasitic infections of refugees.Med. J. Aust. 148, 491–494.
5. de Silva, S. et al. (2002) Chronic Strongyloides stercoralis infection in Laotian
immigrants and refugees 7-20 years after resettlement in Australia. Epidemiol.
Infect. 128, 439–444. doi:10.1017/S0950268801006677
6. Olsen, A. et al. (2009) Strongyloidiasis–the most neglected of the neglected
tropical diseases? Trans. R. Soc. Trop. Med. Hyg. 103, 967–972. doi:10.1016/
j.trstmh.2009.02.013
7. Biggs, B.A. et al. (2009)Management of chronic strongyloidiasis in immigrants and
refugees: is serologic testing useful? Am. J. Trop. Med. Hyg. 80, 788–791.
8. Steinmann, P. et al. (2006) Schistosomiasis and water resources development:
systematic review, meta-analysis, and estimates of people at risk. Lancet Infect.
Dis. 6, 411–425. doi:10.1016/S1473-3099(06)70521-7
9. Chaves, N.J. et al. (2009) Screening practices for infectious diseases among
Burmese refugees in Australia. Emerg. Infect. Dis. 15, 1769–1772. doi:10.3201/
eid1511.090777
10. Sheikh, M. et al. (2009) The epidemiology of health conditions of newly arrived
refugee children: a review of patients attending a specialist health clinic in Sydney.
J. Paediatr. Child Health 45, 509–513. doi:10.1111/j.1440-1754.2009.01550.x
Under theMicroscope
18 MICROBIOLOGY AUSTRALIA * MARCH 2016

11. Yong, M.K. et al. (2010) Long-term follow-up of schistosomiasis serology post-
treatment in Australian travelers and immigrants. J. Travel Med. 17, 89–93.
doi:10.1111/j.1708-8305.2009.00379.x
12. Swanson, S.J. et al. (2012) Albendazole therapy and enteric parasites in United
States-bound refugees. N. Engl. J. Med. 366, 1498–1507. doi:10.1056/
NEJMoa1103360
13. Anantaphruti, M.T. et al. (2010) Molecular and serological survey on taeniasis and
cysticercosis in Kanchanaburi Province, Thailand. Parasitol. Int. 59, 326–330.
doi:10.1016/j.parint.2010.03.007
14. Hotez, P.J. et al. (2012) Neglected tropical diseases of the Middle East and North
Africa: reviewof their prevalence, distribution, and opportunities for control.PLoS
Negl. Trop. Dis. 6, e1475. doi:10.1371/journal.pntd.0001475
15. Schär, F. et al. (2013) Strongyloides stercoralis: global distribution and risk factors.
PLoS Negl. Trop. Dis. 7, e2288. doi:10.1371/journal.pntd.0002288
16. Chang, A.H. et al. (2013) Decreasing intestinal parasites in recent Northern
California refugees. Am. J. Trop. Med. Hyg. 88, 191–197. doi:10.4269/ajtmh.2012.
12-0349
17. Kinkel, H.F. et al. (2012) Evaluation of eight serological tests for diagnosis of
imported schistosomiasis. Clin. Vaccine Immunol. 19, 948–953. doi:10.1128/
CVI.05680-11
18. Bisoffi, Z. et al. (2014) Diagnostic accuracy of five serologic tests for Strongyloides
stercoralis infection. PLoS Negl. Trop. Dis. 8, e2640. doi:10.1371/journal.pntd.
0002640
19. Australian Society of Infectious Diseases (2009) Diagnosis, management and
prevention of infections in recently arrived refugees Sydney, NSW: ASID.
Biographies
Dr Sarah Hanieh is a paediatric infectious diseases physician and
NHMRC Early Career Research Fellow in the Immigrant and Inter-
national Health Group, at the Peter Doherty Institute for Infection
and Immunity, The University of Melbourne. She has interests in
child nutrition, infectious diseases and refugee health. Her current
research aims to understand the nutritional and infectious causes of
impaired child growth and development in resource poor settings.
Norbert Ryan is a Senior Scientist in the Bacteriology Laboratory at
Victorian Infectious Diseases Reference Laboratory, Doherty Insti-
tute, Melbourne. Areas of interest include parasitology, diagnosis of
Legionnaires’diseaseandsexually transmitted infections.Theuseof
staining methods for diagnosis of protozoal infections including
microsporidia has been a specialty field. He was involved in proces-
sing of specimens during the systematic health screening of refugee
and family reunion migrant groups conducted by the Victorian
Department of health during the late 80s-early 90s.
Professor Beverley-Ann Biggs heads the International and Im-
migrant Health Group in the Department of Medicine, The Univer-
sity of Melbourne and is an Infectious Diseases Physician in the
Victorian Infectious Diseases Service at the Royal Melbourne
Hospital. She has a special interest in parasitic and other infectious
diseases in refugees and immigrants living in Australia, and has
published extensively in this area.
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 19

Free living amoebae and human disease
Evan Bursle
Sullivan Nicolaides PathologyWhitmore StreetTaringa, Qld 4068, AustraliaEmail: [email protected]
Jennifer Robson
Sullivan Nicolaides PathologyWhitmore StreetTaringa, Qld 4068, AustraliaEmail: [email protected]
Pathogenic FLA are ubiquitous protozoans and despite fre-
quent human contact remain a rare cause of often devastat-
ing infectionwith poor prognosis. Given changes in climate,
human encroachment into the environment, increasing im-
munosuppression, and improving diagnostic capacity, it is
likely we will see increased cases in the future. Early diag-
nosis is challenging but crucial to achieving a favourable
outcome. It is best facilitated by improved awareness of
FLA disease, appropriate clinical suspicion and early diag-
nostic testing.
Free living amoebae (FLA) are a cosmopolitan group of protozoan
organisms that do not require a host to survive. Despite their
ubiquitous nature, these organisms are uncommon human patho-
gens.However, four genera contain species known to cause invasive
disease in humans: Acanthamoeba, Naegleria, Balamuthia and
Sappinia. Acanthamoeba infectionsmaypresent as granulomatous
amoebic encephalitis (GAE), disseminated disease (e.g. cutaneous,
sinus or pulmonary infection) or keratitis, with Balamuthia man-
drillaris causing similar cutaneous infections and GAE. Naegleria
fowleri is responsible for the rapidly progressive primary amoebic
meningoencephalitis (PAM). In addition, a single case of human
central nervous system (CNS) infection with Sappinia pedata has
been reported1, along with isolated cases of corneal infection with
Vahlkampfia spp.,Hartmannella spp. and Paravahlkamfia spp.2.
This review aims to provide an overview of human disease caused
by the three most common genera involved, Acanthamoeba,
Naegleria and Balamuthia, including their laboratory diagnosis.
Epidemiology
Pathogenic FLAare foundworldwideand serological studies suggest
humanexposure is common3–5. Three cases ofAcanthamoebaGAE
have been described in Australia6–8. Acanthamoeba sp tolerate a
wide rangeof temperature, pH andosmolarity9 andmay be found in
air, soil and water samples. They are one of the most commonly
isolated FLA in the environment10, and themost common in human
infection2. The strongest risk factor for GAE or disseminated infec-
tion is immunodeficiency, while keratitis most commonly affects
immunocompetent contact lenswearerswhooftenhave ahistory of
poor lens hygiene. Exposure is thought to occur by inhalation,
mucosal contact or direct inoculation10. Acanthamoeba species
were previously classified based on morphology, but are now
grouped into 17 genotypes based on 18S rRNA sequencing, with
the majority of pathogenic species belonging to the T4 genotype9,11.
Naegleria fowleri may be found in rivers, lakes and soil but does
not survive in sea water. As thermophiles, their presence in fresh
water is related to temperature and they may even be recovered
from thermally pollutedwaters at high latitudes12. Human exposure
occurs through contact with intact or disrupted nasal mucosa,
commonly through recreational or nasal ablution practices13 and
contaminated drinking water has been implicated as a source of
infection in some cases14. Australia has featured prominently in
the history of Naegleria fowleri infection, with cases described in
Queensland, New South Wales and Western Australia following the
first description of the disease in 1965 by two South Australian
pathologists, Fowler and Carter15,16. This discovery was related to
an outbreak of 20 cases, attributed to a contaminated overland
water pipeline which reached optimal temperatures for Naegleria
proliferation during the summer months. Cases continued from
1947 to 1972 when public health measures including adequate
chlorination were applied17. Several cases inWestern Australia have
also been associated with overland water pipelines, and Australian
drinking water guidelines suggest a Naegleria monitoring and
response protocol for water supplies that seasonally exceed 308C,
or 258C continually18. Today, despite significant advancement in
Under theMicroscope
20 10.1071/MA16009 MICROBIOLOGY AUSTRALIA * MARCH 2016

knowledge regarding risk reduction measures, the incidence of
infection with Naegleria fowleri appears to be increasing world-
wide, with factors such as global warming, substandard water
management and sanitation, and changing recreational practices
likely involved13.
Balmuthia mandrillaris is most commonly found in soil, though
it may also be recovered from water samples4. Infection tends to
occur in immunocompetent individuals, most commonly children,
probably through inhalation or nasal or cutaneous inoculation.
Transmission via organ transplantation has also been described19.
Six human cases have been described in Australia20–24 (Western
Australia, Tasmania, Victoria and Queensland), with another being
described in a Victorian dog25.
Clinical manifestations
Acanthamoeba sp.
The predominant clinicalmanifestations of Acanthamoeba infection
are disseminated disease (e.g. cutaneous, nasopharyngeal or pulmo-
nary infection), GAE and keratitis. With the exception of rare case
reports, GAE and disseminated disease occur in the immunocom-
promised or debilitated. The incubation period is unknown, but
thought to be weeks to months in duration. Cutaneous infection
usually begins as fibronodular lesions, which progress to non-healing
ulcerated lesions over time10. While GAE may be fatal within days of
symptom onset, it generally assumes a more chronic course, with
slow progression over weeks to months. Clinical features of GAE
are myriad and include fever, symptoms of meningism, personality
and mental status change and, later, focal neurological deficits,
coma and death. Imaging of the brain may show single or multiple
space occupying lesions which can be ring enhancing.
Acanthamoeba keratitis is usually a disease of immunocompetent
patients, with the strongest risk factor being contact lens use and
poor lens hygiene. The disease is usually unilateral, with symptoms
including lacrimation, pain, photophobia and foreign body sensa-
tion. Signs include a typical corneal ring infiltrate, stromal infiltrates,
epitheliopathy and hypopyon26. In the absence of effective therapy,
it may progress to corneal perforation and loss of vision. The clinical
diagnosis of AK can be difficult, as lesions may resemble bacterial
or fungal disease, or the dendritic ulcer of HSV infection26. Further-
more the clinical course may be characterised by periods of tem-
porary remission, leading to false impressions of response to
antibacterial or viral agents2.
Naegleria fowleri
Naegleria fowleri causes primary amoebic meningoencephalitis.
Symptoms generally occur 2–5 days after exposure and may begin
with changes in taste or smell, followed by fever, nausea, vomiting,
photophobia and headache. The disease is fulminant, with rapid
progression to coma and death.
Balamuthia mandrillaris
Balamuthia mandrillaris causes GAE in immunocompromised
and immunocompetent individuals. The onset of meningoenceph-
alitis is often subacute or chronic, with symptoms developing over a
period of 2 weeks to 2 years27. It also has the propensity to cause
cutaneous lesions that may precede CNS involvement and are
similar in appearance to those of Acanthamoeba sp. These lesions
appear as poorly defined plaques and may be single or with
bordering satellite lesions4. They often involve the central face and
appear to be more common in South America4. Cutaneous disease
generally progresses to CNS involvement; however, it may resolve
with therapy4.
Laboratory diagnosis
Diagnosis of FLA infection, particularly systemic disease, is chal-
lenging: it may masquerade as bacterial or viral infection, exposure
events may not be apparent and specialised diagnostic testing
availability is limited. Unfortunately as a result, many CNS infections
are diagnosed post-mortem. Successful early diagnosis depends on
appropriate clinical suspicion and collection of suitable diagnostic
material, usually tissue or CSF.
Microscopy
CSF samples in cases of PAM appear purulent, with no bacteria
evident on gram stain, a polymorphonuclear pleocytosis, elevated
protein and decreased glucose. Wet preparations may show motile
Naegleria fowleri trophozoites (Figure 1a), as there are usually
large numbers of organisms in theCSF. Thesefindings contrast with
those of GAE caused by Acanthamoeba or Balamuthia. While CSF
from cases of GAE also demonstrates elevated protein and lowered
glucose, these changes aremoremodest, andamononuclear, rather
than polymorphonuclear inflammatory response is seen. Further-
more,AcanthamoebaorBalamuthia trophozoites are not typically
seen in CSF preparations.
Trophozoites from all pathogenic FLA species can be difficult to
differentiate from host inflammatory cells, especially in stained
tissue sections. Thenuclear characteristics of amoeba canbehelpful
in differentiating these parasites from host cells, with Naegleria
fowleri possessing a nucleus with a large, round, central nucleolus
and Acanthamoeba and Balamuthia (Figure 1b) a rounded
nucleus with large, central nucleolus forming a halo. Polyclonal
andmonoclonal antibodies, with a secondary detecting fluorescent
anti-IgG antibody (such as FITC), may be used to identify and
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 21
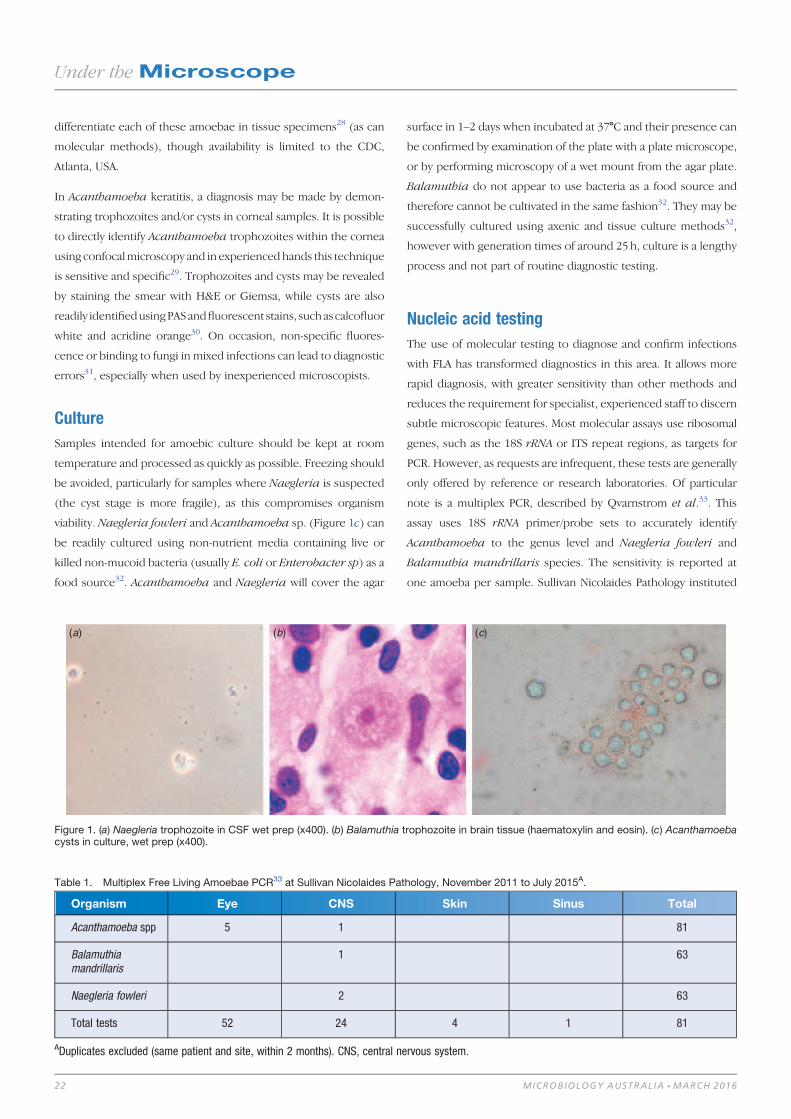
differentiate each of these amoebae in tissue specimens28 (as can
molecular methods), though availability is limited to the CDC,
Atlanta, USA.
In Acanthamoeba keratitis, a diagnosis may be made by demon-
strating trophozoites and/or cysts in corneal samples. It is possible
to directly identify Acanthamoeba trophozoites within the cornea
using confocalmicroscopy and in experienced hands this technique
is sensitive and specific29. Trophozoites and cysts may be revealed
by staining the smear with H&E or Giemsa, while cysts are also
readily identifiedusingPASandfluorescent stains, suchascalcofluor
white and acridine orange30. On occasion, non-specific fluores-
cence or binding to fungi in mixed infections can lead to diagnostic
errors31, especially when used by inexperienced microscopists.
Culture
Samples intended for amoebic culture should be kept at room
temperature and processed as quickly as possible. Freezing should
be avoided, particularly for samples where Naegleria is suspected
(the cyst stage is more fragile), as this compromises organism
viability. Naegleria fowleri and Acanthamoeba sp. (Figure 1c) can
be readily cultured using non-nutrient media containing live or
killed non-mucoid bacteria (usually E. coli or Enterobacter sp) as a
food source32. Acanthamoeba and Naegleria will cover the agar
surface in 1–2 days when incubated at 378C and their presence can
be confirmed by examination of the plate with a plate microscope,
or by performing microscopy of a wet mount from the agar plate.
Balamuthia do not appear to use bacteria as a food source and
therefore cannot be cultivated in the same fashion32. They may be
successfully cultured using axenic and tissue culture methods32,
however with generation times of around 25 h, culture is a lengthy
process and not part of routine diagnostic testing.
Nucleic acid testing
The use of molecular testing to diagnose and confirm infections
with FLA has transformed diagnostics in this area. It allows more
rapid diagnosis, with greater sensitivity than other methods and
reduces the requirement for specialist, experienced staff to discern
subtle microscopic features. Most molecular assays use ribosomal
genes, such as the 18S rRNA or ITS repeat regions, as targets for
PCR. However, as requests are infrequent, these tests are generally
only offered by reference or research laboratories. Of particular
note is a multiplex PCR, described by Qvarnstrom et al.33. This
assay uses 18S rRNA primer/probe sets to accurately identify
Acanthamoeba to the genus level and Naegleria fowleri and
Balamuthia mandrillaris species. The sensitivity is reported at
one amoeba per sample. Sullivan Nicolaides Pathology instituted
(a) (b) (c)
Figure 1. (a) Naegleria trophozoite in CSF wet prep (x400). (b) Balamuthia trophozoite in brain tissue (haematoxylin and eosin). (c) Acanthamoebacysts in culture, wet prep (x400).
Table 1. Multiplex Free Living Amoebae PCR33 at Sullivan Nicolaides Pathology, November 2011 to July 2015A.
Organism Eye CNS Skin Sinus Total
Acanthamoeba spp 5 1 81
Balamuthiamandrillaris
1 63
Naegleria fowleri 2 63
Total tests 52 24 4 1 81
ADuplicates excluded (same patient and site, within 2 months). CNS, central nervous system.
Under theMicroscope
22 MICROBIOLOGY AUSTRALIA * MARCH 2016

this test in 2010 and our experience is summarised in
Table 1. As a significant number of ocular specimens have been
tested against all three targets, a large number of negatives for
Naegleria fowleri and Balamuthiamandrillaris are expected. It is
alsonotable thatwehave recently reported aprobable false negative
Acanthamoeba result. The referred CSF specimen tested negative
at our laboratory, but positive at the CDC (using the same
multiplex PCR). It is possible the small volume of CSF received at
our laboratory contributed to this result (only 100mL was received,
where 200mL is the standard volume for extraction).
Treatment and prognosis
Primary amoebic meningitis
There have been few survivors of primary amoebic meningitis
and the factors that have resulted in successful therapy are poorly
defined, though early diagnosis and institution of therapy appears
critical. The treatment of choice is the antifungal amphotericin B,
which is often used both intravenously and intrathecally. Other
agents used in survivors include miltefosine, sulfisoxazole, flucon-
azole, miconazole and rifampicin.
Granulomatous amoebic encephalitis
Acanthamoeba
Suboptimal efficacy of antimicrobial agents, the high morbidity of
patients affected and tendency to late diagnosis contribute to a
poor prognosis in Acanthamoeba GAE, with a mortality rate of
>90%34. In the few reported survivors of GAE or cutaneous
infection, most have received combination therapy. The agents
used have included trimethoprim-sulfamethoxazole, flucytosine,
sulfadiazine, penicillin G, chloramphenicol, pentamidine, flucona-
zole and itraconazole. More recently, it appears that the inclusion
of miltefosine in combination regimens may result in improved
survival35.
Balamuthia
Early recognition of cutaneous disease is critical to allow early
therapy and prevent progression to CNS infection. Unfortunately,
the overall prognosis in Balamuthia CNS infection is extremely
poor. However, there are several case reports of survival in the
literature36, including an Australian case24. These cases have re-
ceived varied combination therapy regimens, with agents including
pentamidine, flucytosine, fluconazole, macrolides, sulfadiazine,
miltefosine, thioridazine, amphotericin B, albendazole and trimeth-
oprim-sulfamethoxazole. Miltefosine, in particular, has demonstrat-
ed amoebicidal activity in vitro37 and its inclusion in combination
regimens may offer a survival advantage35.
Acanthamoeba keratitis
Like disseminated disease, success in treatment is dependent on
early diagnosis and institution of therapy. Fortunately, success
rates in treating Acanthamoebic keratitis are more promising, with
cure rates in the literature generally greater than 75–85%38,39.
Topical chlorhexidine and polyhexamethylenebiguanide (PHMB)
are effective against trophozoites and cysts and form the mainstay
of therapy. These are usually used in combination with diamidine,
propamidine or hexamidine, though other agents including keto-
conazole, itraconazole, voriconazole and topical imidazoles have
been used. Surgical intervention, including enucleation, is some-
times required in severe cases40.
References1. Qvarnstrom, Y. et al. (2009) Molecular confirmation of Sappinia pedata as a
causative agent of amoebic encephalitis. J. Infect. Dis. 199, 1139–1142.
doi:10.1086/597473
2. Mandell, G.L. et al. (2015) Mandell, Douglas, and Bennett’s principles and
practice of infectious diseases. Philadelphia, PA: Churchill Livingstone/Elsevier.
3. Chappell, C.L. et al. (2001) Standardized method of measuring Acanthamoeba
antibodies in sera from healthy human subjects. Clin. Diagn. Lab. Immunol. 8,
724–730.
4. Bravo, F.G. and Seas, C. (2012) Balamuthia mandrillaris amoebic encephalitis:
an emerging parasitic infection. Curr. Infect. Dis. Rep. 14, 391–396. doi:10.1007/
s11908-012-0266-4
5. Marciano-Cabral, F. et al. (1987) Specificity of antibodies from human sera for
Naegleria species. J. Clin. Microbiol. 25, 692–697.
6. Carter, R.F. et al. (1981) A fatal case of meningoencephalitis due to a free-living
amoeba of uncertain identity–probably Acanthamoeba Sp. Pathology 13, 51–68.
doi:10.3109/00313028109086829
7. Harwood, C.R. et al. (1988) Isolation of Acanthamoeba from a cerebral abscess.
Med. J. Aust. 148, 47.
8. Azzam, R. et al. (2015) Acanthamoeba encephalitis: isolation of genotype T1
in mycobacterial liquid culture medium. J. Clin. Microbiol. 53, 735–739.
doi:10.1128/JCM.02887-14
9. Trabelsi, H. et al. (2012) Pathogenic free-living amoebae: epidemiology
and clinical review. Pathol. Biol. (Paris) 60, 399–405. doi:10.1016/j.patbio.
2012.03.002
10. Marciano-Cabral, F. and Cabral, G. (2003) Acanthamoeba spp. as agents of
disease in humans. Clin. Microbiol. Rev. 16, 273–307. doi:10.1128/CMR.16.
2.273-307.2003
11. Siddiqui, R. and Khan, N.A. (2012) Biology and pathogenesis of Acanthamoeba.
Parasit. Vectors 5, 6. doi:10.1186/1756-3305-5-6
12. Kemble, S.K. et al. (2012) Fatal Naegleria fowleri infection acquired in
Minnesota: possible expanded range of a deadly thermophilic organism. Clin.
Infect. Dis. 54, 805–809. doi:10.1093/cid/cir961
13. Siddiqui, R. and Khan, N.A. (2014) Primary amoebic meningoencephalitis
caused by Naegleria fowleri: an old enemy presenting new challenges. PLoS
Negl. Trop. Dis. 8, e3017. doi:10.1371/journal.pntd.0003017
14. Cooter, R. (2002) The history of the discovery of primary amoebic meningoen-
cephalitis. Aust. Fam. Physician 31, 399–400.
15. Fowler, M. and Carter, R.F. (1965) Acute pyogenic meningitis probably due to
Acanthamoeba sp.: a preliminary report. BMJ 2, 734–742. doi:10.1136/bmj.2.
5464.734-a
16. Carter, R.F. (1968) Primary amoebic meningo-encephalitis: clinical, pathological
and epidemiological features of six fatal cases. J. Pathol. Bacteriol. 96, 1–25.
doi:10.1002/path.1700960102
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 23

17. Dorsch, M.M. et al. (1983) The epidemiology and control of primary amoebic
meningoencephalitis with particular reference to South Australia. Trans. R. Soc.
Trop. Med. Hyg. 77, 372–377. doi:10.1016/0035-9203(83)90167-0
18. National Health and Medical Research Council Australia (2011) Natural
Resource Management Ministerial C. Australian drinking water guidelines 2011:
national water quality management strategy. Canberra: National Health and
Medical Research Council.
19. Schlessinger, S. et al. (2010) Balamuthia mandrillaris transmitted through
organ transplantation – Mississippi, 2009. Morbidity and Mortality Weekly
Report 59, 1165–1170.
20. Carter, R.F. et al. (1981) A fatal case of meningoencephalitis due to a free-living
amoeba of uncertain identity–probably Acanthamoeba sp. Pathology 13, 51–68.
doi:10.3109/00313028109086829
21. Reed, R.P. et al. (1997) Fatal granulomatous amoebic encephalitis caused by
Balamuthia mandrillaris. Med. J. Aust. 167, 82–84.
22. Hill, C.P. et al. (2011) Balamuthia amebic meningoencephalitis and mycotic
aneurysms in an infant. Pediatr. Neurol. 45, 45–48. doi:10.1016/j.pediatrneurol.
2011.05.003
23. Doyle, J.S. et al. (2011) Balamuthia mandrillaris brain abscess successfully
treated with complete surgical excision and prolonged combination antimicrobial
therapy. J. Neurosurg. 114, 458–462. doi:10.3171/2010.10.JNS10677
24. Moriarty, P. et al. (2014) Balamuthia mandrillaris encephalitis: survival of a
child with severe meningoencephalitis and review of the literature. J. Pediatric
Infect. Dis. Soc. 3, e4–e9. doi:10.1093/jpids/pit033
25. Finnin, P.J. et al. (2007) Multifocal Balamuthia mandrillaris infection in a dog
in Australia. Parasitol. Res. 100, 423–426. doi:10.1007/s00436-006-0302-0
26. Patel, D.V. and McGhee, C.N. (2009) Acanthamoeba keratitis: a comprehensive
photographic reference of common and uncommon signs. Clin. Experiment.
Ophthalmol. 37, 232–238. doi:10.1111/j.1442-9071.2008.01913.x
27. Visvesvara, G.S. et al. (2007) Pathogenic and opportunistic free-living
amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri,
and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 50, 1–26. doi:10.1111/
j.1574-695X.2007.00232.x
28. da Rocha-Azevedo, B, Tanowitz, HB and Marciano-Cabral, F (2009) Diagnosis
of infections caused by pathogenic free-living amoebae. Interdisciplinary
perspectives on infectious diseases 2009, 251406. doi:10.1155/2009/251406
29. Tu, E.Y. et al. (2008) The relative value of confocal microscopy and superficial
corneal scrapings in the diagnosis of Acanthamoeba keratitis. Cornea 27,
764–772. doi:10.1097/ICO.0b013e31816f27bf
30. Hahn, T.-W. et al. (1998) Acridine orange staining for rapid diagnosis of Acantha-
moeba keratitis. Jpn. J. Ophthalmol. 42, 108–114. doi:10.1016/S0021-5155(97)
00127-5
31. Grossniklaus, H.E. et al. (2003) Evaluation of hematoxylin and eosin and special
stains for the detection of Acanthamoeba keratitis in penetrating keratoplasties.
Am. J. Ophthalmol. 136, 520–526. doi:10.1016/S0002-9394(03)00322-2
32. Schuster, F.L. (2002) Cultivation of pathogenic and opportunistic free-living
amebas. Clin. Microbiol. Rev. 15, 342–354. doi:10.1128/CMR.15.3.342-354.2002
33. Qvarnstrom, Y. et al. (2006) Multiplex real-time PCR assay for simultaneous
detection of Acanthamoeba spp., Balamuthia mandrillaris, and Naegleria
fowleri. J. Clin. Microbiol. 44, 3589–3595. doi:10.1128/JCM.00875-06
34. Barratt, J.L.N. et al. (2010) Importance of nonenteric protozoan infections in
immunocompromised people. Clin. Microbiol. Rev. 23, 795–836. doi:10.1128/
CMR.00001-10
35. Cope, J. et al. (2013) Increased patient survival: miltefosine for treatment of
free-living ameba infections caused by Acanthamoeba and Balamuthia
[Poster]. Council of State and Territorial Epidemiologists’ Annual Conference.
Pasadena, California.
36. Deetz, T.R. et al. (2003) Successful treatment of Balamuthia amoebic
encephalitis: presentation of 2 cases. Clin. Infect. Dis. 37, 1304–1312.
doi:10.1086/379020
37. Schuster, F.L. et al. (2006) In-vitro activity of miltefosine and voriconazole on
clinical isolates of free-living amebas: Balamuthia mandrillaris, Acanthamoeba
spp., and Naegleria fowleri. J. Eukaryot. Microbiol. 53, 121–126. doi:10.1111/
j.1550-7408.2005.00082.x
38. Dart, J. K. et al. (2009) Acanthamoeba keratitis: diagnosis and treatment update
2009. Am. J. Ophthalmol. 148, 487–99e2. doi:10.1016/j.ajo.2009.06.009
39. Butler, T.K.H. et al. (2005) Six-year review of Acanthamoeba keratitis in New
South Wales, Australia: 1997–2002. Clin. Experiment. Ophthalmol. 33, 41–46.
doi:10.1111/j.1442-9071.2004.00911.x
40. Vemuganti, G.K. et al. (2005) Granulomatous inflammation in Acanthamoeba
keratitis: an immunohistochemical study of five cases and review of literature.
Indian J. Med. Microbiol. 23, 231–238.
Biographies
Dr Evan Bursle, BSc, MBBS, is a microbiology and infectious
diseases registrar based at Sullivan Nicolaides Pathology. Although
his microbiological tastes are broad and still being refined, he has a
keen interest in parasitology.
Dr Jenny Robson, MBBS (Hons I); FRACP, FRCPA, FACTM, is an
infectious disease physician andmicrobiologist who has worked for
the past 26 years at Sullivan Nicolaides Pathology. She has a broad
range of interests, which includes travel and tropical medicine.
Future issues of Microbiology Australia
May 2016: Education to enhance microbiology graduate employability
Guest Editor: Danilla Grando
September 2016: Diseases of Aquaculture
Guest Editor: Nicky Buller
November 2016: Microbiology of Travel
Guest Editor: Ipek Kurtboke; joint issue with the Society for General Microbiology
March 2017: Bat-associated diseases
Guest Editor: Glenn Marsh
Under theMicroscope
24 MICROBIOLOGY AUSTRALIA * MARCH 2016

Tapeworm cysts in the brain: can we preventit happening?
Marshall Lightowlers
Facultyof VeterinaryandAgriculturalSciencesThe University of Melbourne250 Princes HighwayWerribee, Vic. 3030, AustraliaEmail: [email protected]
Imagine theconsternation; youareamemberofanorthodox
Jewish family and you and another family member are
diagnosed with larvae of a pork tapeworm in your brain.
You have recurrent seizures as a result. Ridiculous? Not
for members of a Jewish community in New York where a
Mexican domesticworker harbouring aTaenia solium tape-
worm had apparently contaminated the family’s food with
eggs from her tapeworm1.
T. solium is a cestode parasite that is transmitted between pigs and
people. Pigs may harbour the parasite’s larval stage in their muscles
(Figure 1). Ifwe accidentally eat oneof these larvae in poorly cooked
pig meat, the adult tapeworm will develop in our small intestine.
Humans are the only definitive host (the host in which the parasite
undergoes sexual reproduction) for this species of tapeworm. A
person with the tapeworm releases the parasite’s eggs in their
faeces, and the lifecycle is completed if foods contaminated with
these eggs, or indeed the faeces themselves, are consumed by a pig.
The disease is fully transmitted only where pigs roam freely and
humans defecate in areaswhere thepigs are free roaming. This does
nothappen inNewYork.However, it doeshappenacross large areas
of central and South America, Africa and east and south-east Asia.
In former times T. solium and the human brain disease that the
parasite causes, neurocysticercosis, were endemic throughout
Europe and other parts of the current-day First World; however,
improved public sanitation and hygienic standards for raising pigs
have seen thedisease in pigs eliminatedwithout any efforts directed
specifically to the disease. Likely the same will happen, in time,
through economic development in those areas of the world where
thedisease remains endemic today. Unfortunately, this is unlikely to
occur in our lifetimes. Meanwhile, those living in the T. solium
endemic areas of the world continue to suffer epilepsy and death
due to the presence of T. solium cysts in their brain. Keep in mind
that people from poor countries can travel anywhere with their
intestinal residents and deliver their tapeworm eggs to you or me,
just as happened in the New York community referred to above.
No point turning vego. Consider this: I have a worm and I am
not particularly hygienic when I go to the toilet. I make your salad;
bingo – see you in the neurology clinic.
T. solium is not an obscure parasite. The Food and Agriculture
Organization of the United Nations considers it to be the most
important foodborne parasitic infection from a global perspective2.
T. solium is the most frequent preventable cause of seizure dis-
orders, being associated with 29% of people with epilepsy3. The
World Health Organization list T. solium as one of 16 Neglected
Tropical Diseases, and is actively promoting efforts to reduce the
parasite’s transmission4.
For those living in poor countries in which T. solium is endemic,
help is at hand from, of all places, Australia. I say ‘of all places’
because the parasite is not, and has never been known to be,
endemic here. Nevertheless, a research program at the University
of Melbourne with an original genesis way back in the 1970s led
eventually to the development of both the first effective non-living
vaccine against a eukaryotic parasite and eventually also to a vaccine
that can stop pigs being infected with T. solium. The vaccine uses a
recombinant antigen known as TSOL18. Several independent ex-
perimental trials of TSOL18 have confirmed that it is extraordinarily
effective5. A field trial of the vaccine was undertaken in which pairs
of young piglets were distributed to families living in a T. solium
endemic region of north-east Cameroon6. One animal from each
pair was vaccinated and one acted as a control. When the animals
were of normal eating-age (~12 months), they were recovered
from the farmers and assessed for T. solium infection. About 20%
of the controls were infected but not a single parasite was found in
any of the 110 vaccinated animals.
So far, so good. However the field trial involved a single cohort of
animals that was vaccinated. In a real life situation, new disease
susceptible piglets are born into the community more-or-less
every day. Difficulties arise when we try to work out a feasible
and sustainable program to deliver vaccination regularly enough
to prevent pigs becoming infected on an on-going basis. The
‘standard’ vaccination protocol calls for two immunizations
about a month apart. It is difficult to give any veterinary care to
pigs in the communities where T. solium is transmitted; however
having to deliver two vaccines a month apart would be close to
impossible.
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 10.1071/MA16010 25

Using knowledge about the parasite’s development in pigs and
information from the successful Cameroonian field trial, various
scenarios involving vaccination in pigs were compared for their
predicted effectiveness to control T. solium transmission7. One
scenario that appeared relatively practical was to vaccinate at
4-monthly intervals. While the scenario looked good theoretically,
it could not be recommended because we had no information
about whether the vaccine would raise a protective response if
the interval between primary and secondary injections was longer
than 4 weeks.
Recently we have completed an experiment in which pigs received
their secondary immunizationwithTSOL18at4,8,12,16or20weeks
after the first injection8. The results were very promising. Antibody
responses to the vaccine generally increased beyond the ‘standard’
4-week interval. Responses seen in the animals vaccinated at a
12 week interval were the best, and field evaluation of T. solium
interventions are about to begin in several endemic regions of Africa
that will involve vaccinations at 3 or 4 monthly intervals.
Despite the solid progress that has beenmade so far, much remains
to be achieved before we would be likely to see pig vaccination
contribute to reducing the incidence of neurocysticercosis in peo-
ple. One major difficulty with implementation of pig vaccination to
prevent transmission of T. solium is that the owners of the animals
often have little incentive to undertake control measures because
the infection does not often directly cause illness or death in pigs.
In the future, combination vaccines that include TSOL18 and also
antigens that can provide protection against pig deaths caused by
Classical Swine Fever (CSF) in the Americas, or African Swine Fever
(ASF) in African countries, would improve the acceptability of pig
vaccination by providing an economic incentive for the animal
owners to vaccinate. While the potential for a combination with
CSF is something that can be explored immediately because com-
mercial vaccines alreadyexist, there is yet tobeacommercial vaccine
for ASF.
As for the Jewish community in New York who use domestic staff
sourced from T. solium endemic countries, and of course all the
people who live permanently in those endemic countries, the risk
of exposure to T. solium remains. Hopefully we will be able to
overcome the practical difficulties around working with pigs in
T. solium endemic areas so that implementation of pig vaccination
can reduce T. solium transmission and decrease the incidence of
human neurocysticercosis as a result.
References1. Schantz, P.M. et al. (1992) Neurocysticercosis in an Orthodox Jewish community in
New York City.N. Engl. J. Med. 327, 692–695. doi:10.1056/NEJM199209033271004
2. Robertson, L.J. et al. (2013) Have foodborne parasites finally become a global
concern? Trends Parasitol. 29, 101–103. doi:10.1016/j.pt.2012.12.004
3. Ndimubanzi, P.C. et al. (2010) A systematic review of the frequency of neurocy-
ticercosis with a focus on people with epilepsy. PLoS Negl. Trop. Dis. 4, e870.
doi:10.1371/journal.pntd.0000870
4. Organization, W.H. (2015) Investing to overcome the global impact of neglected
tropical diseases. Third WHO report on neglected tropical diseases. WHO/HTM/
NTD/2015.1.
5. Lightowlers, M.W. (2010) Eradication of Taenia solium cysticercosis: a role for
vaccination of pigs. Int. J. Parasitol. 40, 1183–1192. doi:10.1016/j.ijpara.2010.05.
001
6. Assana, E. et al. (2010) Elimination of Taenia solium transmission to pigs in a field
trial of theTSOL18 vaccine inCameroon. Int. J. Parasitol.40, 515–519. doi:10.1016/
j.ijpara.2010.01.006
7. Lightowlers, M.W. (2013) Control of Taenia solium taeniasis/cysticercosis: past
practices and new possibilities. Parasitology 140, 1566–1577. doi:10.1017/S003
1182013001005
8. Lightowlers, M.W. et al. (2016) Anamnestic responses in pigs to the Taenia solium
TSOL18 vaccine and implications for control strategies. Parasitology, in press.
BiographyMarshall Lightowlers has been a full-time research scientist
supported by medical research funding for more-or-less all of his
working life.He currently holds appointments as Laureate Professor
at The University of Melbourne’s Faculty of Veterinary and Agricul-
tural Sciences, and Principal Research Fellow with the NHMRC.
(a)
(b)
Figure 1. (a) Taenia solium cysts (cysticerci) in the tongue of a naturallyinfected pig (Photo: M. Donadeu). (b) Cysticerci in the muscles of aheavily infected pig.
Under theMicroscope
26 MICROBIOLOGY AUSTRALIA * MARCH 2016

Seafood-borne parasitic diseases in Australia:how much do we know about them?
Shokoofeh Shamsi
School of Animal and VeterinarySciencesCharles Sturt UniversityWagga Wagga, NSW 2650, AustraliaTel: +61 2 6933 4887Email: [email protected]
Fish are host to many parasites, some of which can cause
disease inhumans.With the increase in cultural andculinary
diversity and the increased popularity of eating raw or
slightly cooked seafood dishes in Australia it is speculated
that seafood-borne parasitic infections in Australian consu-
mers may rise. Seafood-borne zoonotic parasites are recog-
nised as a significant public health concern worldwide.
In Australia there are few reports of infection in humans in
themedical literature. Australian Government enforcement
agencies rate the risk of seafood-borne zoonosis as low;
however, the prevalence of seafood-borne zoonoses may be
under-reported in Australia due to misdiagnosis. Although
food safety regulations and import controls for seafood
in Australia are strict, the focus is more on the control of
food-borne bacterial, viral and chemical contaminant rela-
ted illnesses rather than parasitic diseases.
Increasing demand for raw and exotic seafood has significantly
expanded the geographical and demographic limits of fish-borne
parasitic infections. Medical literature on the regular consumption
of seafood is plentiful. It is heavily promoted for prolonging life,
aiding in childhood development, increasing brain stimulation
and even to help our pets. However, there is an increasing risk of
contracting parasitic zoonoses from consuming raw and under-
cooked seafood, which should be of concern in Australia. Zoonotic
parasites are common in Australian seafood, and can pose a major
health risk to people who consume seafood or work in a fishing
industry1. Among zoonotic parasites in seafood, Anisakid nema-
todes are of the greatest significance due to their high prevalence in
wild caught fish2 and also due to the severity of the disease they
cause, which is known as anisakidosis. Other nematodes such as
Gnathostoma andAngiostrongylus aswell as platyhelminths suchas
Diphylobothrium, Clonorchis and Paragonimus, can occasionally
cause seafood-borne zoonotic infections3.
Infection in humans (zoonosis)Anisakidosis results from accidental infection with one or more
larvae of species of certain genera of anisakids, including Anisakis
spp, Contracaecum spp and Pseudoterranova spp. Infection with
larvae of Anisakis and Pseudoterranova is of common occurrence,
whereas infection with other genera, such as Contracaecum has
been reported less frequently. Humans usually become infected
with these parasites after eating raw, undercooked or improperly
processed fish or seafood (Figure 1). Clinically, several different
types of human anisakidosis have been described based on the
location of the parasite. This includes gastro-intestinal, visceral,
oropharyngeal and transient luminal anisakidosis4. The first two
types are associatedwith severe symptoms such as vomiting, severe
pain in lower abdomen and fever.
In recent years, it has been recognised that an allergic response can
occur in humans due to live anisakids or food in which worms have
been killed by cooking or pasteurisation5. Other allergic disorders,
such as chronic urticaria may also result from parasitism6. There is
anecdotal evidence in other countries (e.g. Spain) that some allergic
reactions to seafood are indeed allergic reaction to anisakids in
seafood.
The Australian scenarioIn Australia our knowledge about these important parasites is poor.
Although anisakid nematodes are commonly found in Australian
marine fish (Figure 2), such as mackerel, flathead, snapper and
whiting2,7,8, all forms of seafood are available and popular in Aus-
tralia and 40% of fish consumed raw in Australia have been consid-
ered as infected with Anisakis at the time of processing for trade1,
there are only a handful of documented confirmed cases acquired in
Australia.
Case 1: There has been only one documented confirmed case of
infection with anisakid nematodes acquired in Australia9. This case
involved a 41-year-old South Australian woman of Tongan descent
who became ill after eating raw mackerel that had been caught
locally. She was hospitalised with severe gastrointestinal pain,
diarrhoea and vomiting that progressively worsened for 3 weeks.
The diagnosis only occurred after a worm was passed in her faeces.
The initial presumptive identification of the larva was an intestinal
nematode, possibly a species of the Trichostrongylus or Ascaris
genera. However, on further detailed microscopic examination,
the larva was identified as a species of Contracaecum. Had the
worm not been found, or properly examined by a taxonomist,
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 10.1071/MA16015 27
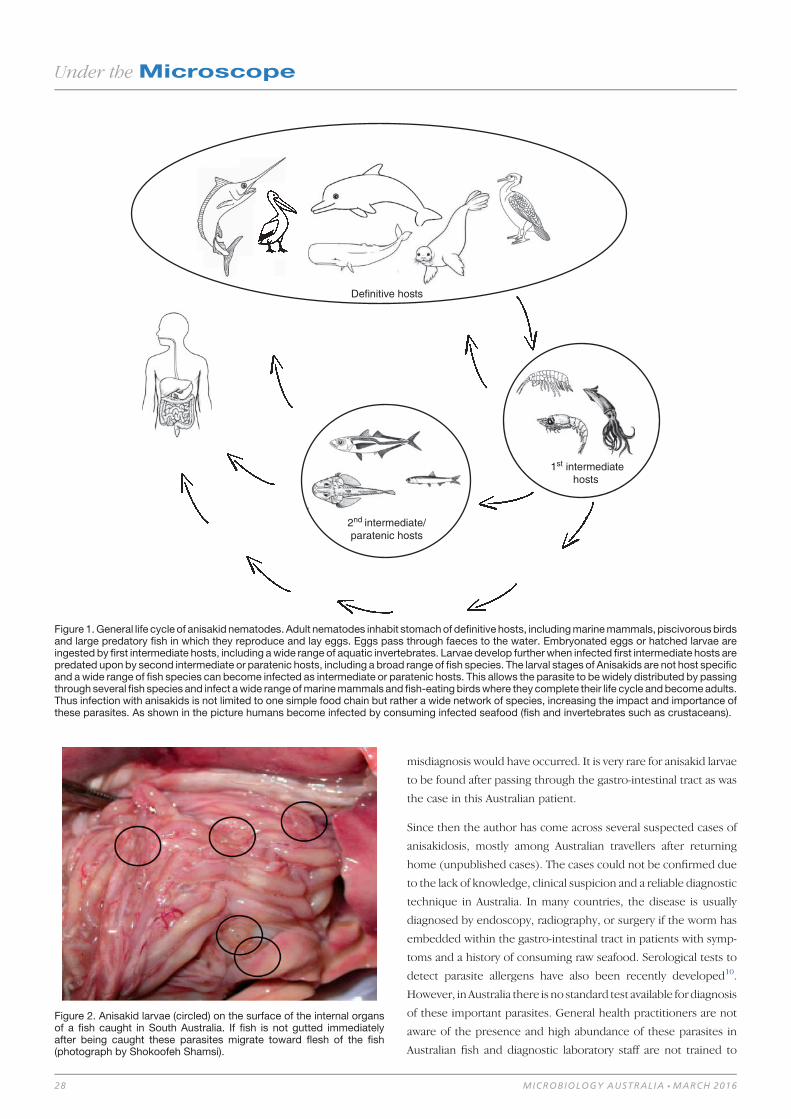
misdiagnosis would have occurred. It is very rare for anisakid larvae
to be found after passing through the gastro-intestinal tract as was
the case in this Australian patient.
Since then the author has come across several suspected cases of
anisakidosis, mostly among Australian travellers after returning
home (unpublished cases). The cases could not be confirmed due
to the lack of knowledge, clinical suspicion and a reliable diagnostic
technique in Australia. In many countries, the disease is usually
diagnosed by endoscopy, radiography, or surgery if the worm has
embedded within the gastro-intestinal tract in patients with symp-
toms and a history of consuming raw seafood. Serological tests to
detect parasite allergens have also been recently developed10.
However, inAustralia there is no standard test available for diagnosis
of these important parasites. General health practitioners are not
aware of the presence and high abundance of these parasites in
Australian fish and diagnostic laboratory staff are not trained to
Definitive hosts
1st intermediatehosts
2nd intermediate/paratenic hosts
Figure 1.General life cycle of anisakid nematodes. Adult nematodes inhabit stomachof definitive hosts, includingmarinemammals, piscivorousbirdsand large predatory fish in which they reproduce and lay eggs. Eggs pass through faeces to the water. Embryonated eggs or hatched larvae areingested by first intermediate hosts, including awide range of aquatic invertebrates. Larvae develop furtherwhen infected first intermediate hosts arepredated uponby second intermediate or paratenic hosts, including a broad range of fish species. The larval stages of Anisakids are not host specificand awide range of fish species can become infected as intermediate or paratenic hosts. This allows the parasite to bewidely distributed by passingthrough several fish species and infect awide rangeofmarinemammals and fish-eating birdswhere they complete their life cycle andbecomeadults.Thus infection with anisakids is not limited to one simple food chain but rather a wide network of species, increasing the impact and importance ofthese parasites. As shown in the picture humans become infected by consuming infected seafood (fish and invertebrates such as crustaceans).
Figure 2. Anisakid larvae (circled) on the surface of the internal organsof a fish caught in South Australia. If fish is not gutted immediatelyafter being caught these parasites migrate toward flesh of the fish(photograph by Shokoofeh Shamsi).
Under theMicroscope
28 MICROBIOLOGY AUSTRALIA * MARCH 2016

identify these worms. As a result, the full extent of hidden cases of
the disease in Australia remains unknown.
Case 2: Another confirmed case report of fish-borne parasitic
disease in humans in Australia occurred in 201111. In this case, a
couple became infected withGnathostoma species after consump-
tion of ‘Black Bream’ (possibly Acanthopagrus berda or Hephaes-
tus jenkinsi), which they caught in the Calder River in Western
Australia. They presented with recurring skin swellings. The fish
had been cooked on a camp fire, but the cooking time was unclear.
The diagnosis was based on eosinophilia and a positive serology.
Humans become infected accidentally by consuming third-stage
larvae. Gnathostoma larvae are unable to mature inside the human
body, and instead migrate through visceral and cutaneous tissues,
causing a variety of generalised, non-specific symptoms. Gnathos-
tomiasis is generally endemic in parts of the world where seafood is
consumed raw, such as in South-East Asia and Japan, but also more
recently in LatinAmerica, India andAfrica, and in travellers returning
from these areas12.
Another aspect of Australia’s seafood consumption that must be
considered is the large volume of imported seafood that Australians
consume. The most important sources of seafood products are
Thailand, New Zealand, Vietnam and China (frdc.com.au/knowl-
edge/Factsheets/Factsheet_Imported_Seafood_in_Australia.pdf).
The literature describes several zoonotic parasites affecting fish
endemic to these countries13. CSIRO conducted a comprehensive
review of AQIS’s imported seafood testing protocols14 and con-
cluded that imported seafood does not pose any greater health risk
to the consumer than locally produced seafood. Nevertheless
proper precautions when preparing seafood in order to reduce
the risk of illness or disease has been recommended. However,
recommendations to eliminate or reduce parasites have not been
dealt with in details.
PreventionGastro-intestinal anisakidosis can beprevented byproperly cooking
fish to an internal temperature of approximately 638C or freezing at
or below�208C for 7 days15. Thiswillmost likely preventmost other
parasites as well.
Consuming raw seafood in reputable restaurants only, where chefs
are trained to recognise infected seafood.
ConclusionIn the absence of standard diagnostic techniques it is difficult to
have a realistic estimation of occurrence of seafood borne parasitic
diseases in the country. As Chai et al.12 stated it appears that fish
borne parasitic disease in Australia remains a ‘public health orphan’
with very little research having been carried out on what the real
risks are. The economic value of the fishing industry and health
of consumers must be protected. Therefore further research is
required to fill in the current knowledge gaps of the biology and
ecology of these parasites and the risk they pose to consumers and
workers.
References1. Anantanawat, S. et al. (2012) A semi-quantitative risk assessment of harmful
parasites inAustralianfinfish. SouthAustralian Research&Development Institute,
2012.
2. Shamsi, S. et al. (2011)Occurrence and abundance of anisakid nematode larvae in
five species of fish from southern Australian waters. Parasitol. Res. 108, 927–934.
doi:10.1007/s00436-010-2134-1
3. Butt, A.A. et al. (2004) Infections related to the ingestion of seafood. Part II:
parasitic infections and food safety. Lancet Infect. Dis. 4, 294–300. doi:10.1016/
S1473-3099(04)01005-9
4. Smith, J.W. (1999) Ascaridoid nematodes and pathology of the alimentary tract
and its associated organs in vertebrates, including man: a literature review.
Helminthological Abstracts 68, 49–96.
5. Audicana, M.T. and Kennedy, M.W. (2008) Anisakis simplex: from obscure
infectious worm to inducer of immune hypersensitivity. Clin. Microbiol. Rev.
21, 360–379. doi:10.1128/CMR.00012-07
6. Daschner, A. and Pascual, C.Y. (2005) Anisakis simplex: sensitization and clinical
allergy. Curr. Opin. Allergy Clin. Immunol. 5, 281–285. doi:10.1097/01.all.0000
168795.12701.fd
7. Doupe, R.G. et al. (2003) Larval anisakid infections of some tropical fish species
from north-west Australia. J. Helminthol. 77, 363–365.
8. Shamsi, S. et al. (2011) Mutation scanning-coupled sequencing of nuclear ribo-
somal DNA spacers (as a taxonomic tool) for the specific identification of different
Contracaecum (Nematoda: Anisakidae) larval types.Mol. Cell. Probes 25, 13–18.
9. Shamsi, S. andButcher, A.R. (2011) First report of human anisakidosis in Australia.
Med. J. Aust. 194, 199–200.
10. García, M. et al. (1997) The use of IgE immunoblotting as a diagnostic tool in
Anisakis simplex allergy. J. Allergy Clin. Immunol. 99, 497–501. doi:10.1016/
S0091-6749(97)70076-9
11. Jeremiah,C.J. et al. (2011)Gnathostomiasis in remotenorthernWesternAustralia:
the first confirmed cases acquired in Australia. Med. J. Aust. 195, 42–44.
12. Herman, J.S. and Chiodini, P.L. (2009) Gnathostomiasis, another emerging
imported disease. Clin. Microbiol. Rev. 22, 484–492. doi:10.1128/CMR.00003-09
13. Chai, J.-Y. et al. (2005) Fish-borne parasitic zoonoses: status and issues. Int.
J. Parasitol. 35, 1233–1254. doi:10.1016/j.ijpara.2005.07.013
14. Moir, C. (2009) Review of the current testing protocols for imported seafood
products. CSIRO Report reference number R-661-03-11. 90 pp.
15. Hochberg, N.S. and Hamer, D.H. (2010) Anisakidosis: perils of the deep. Clin.
Infect. Dis. 51, 806–812.
BiographyDr Shokoofeh Shamsi is a senior lecturer in veterinary Parasito-
logy. After completing a Masters Degree in Medical Parasitology
(Tehran University of Medical Sciences) she gained her PhD in
Veterinary Parasitology from The University of Melbourne. She is a
taxonomist who couples conventional morphological techniques
with modern molecular technologies to diagnose infections and
identify parasite species, especially aquatic parasites. This has
resulted in the discovery of several new pathogenic species and
strains, and their recognition as disease causing agents in humans
and animals in Australia and worldwide.
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 29

Children, snails and worms: the Brachylaimacribbi story
Andrew R Butcher
SA PathologyMicrobiology and InfectiousDiseasesSA, AustraliaEmail:[email protected]
Brachylaimids are parasitic trematode fluke worms that
have a terrestrial life cycle involving land snails and slugs
as thefirst and/or second intermediate hosts for the cercarial
and metacercarial larval stages. A wide range of mammals,
birds, reptiles andamphibians are thedefinitivehosts for the
adult worm. Brachylaima spp. have been reported from
most continents including Europe, Africa, Asia, North and
South America and Australia. There are over 70 described
species in the genus with seven species indigenous to Aus-
tralia. Although Brachylaima spp. are a cosmopolitan ter-
restrial trematode they have not been recorded to infect
humans other than the three Brachylaima cribbi infections
reported in two children and an adult from South Australia.
The publication of a new species of brachylaimid, Brachylaima
cribbibyButcher andGrove in2001was the culminationof a 12-year
scientific journey that would not have been possible without a
broad scientific network of colleagues and some ‘scientific luck’,
which enabled the establishment of a laboratory life cycle using
parasite eggs recovered from the stool of an infected human1.
My long-term association, as an active member of the Australian
Society for Microbiology and the special interest groups played a
significant role in having a network of colleagues to help find
techniques and data to assist in the study of this parasite.
The publication of the first human Brachylaima sp. infections
followed the detection of a fluke worm egg, in the stool of two
South Australian children, who had never travelled overseas. The
eggs were identified as belonging to a genus of trematode worm
endemic in the local area2. After conversations with many local and
interstate colleagues, a veterinary parasitology colleague Michael
O’Callaghan remembered the post-doctoral work of Thomas Cribb
in South Australia. He had studied and published work on a
brachylaimid in South Australia that infected mice3,4. This provided
the data on a local trematode that had an eggmorphologymatching
the eggs detected in the children’s stools.
However, was this a true human infection that resulted in the
establishment ofmature gravidworms producing the eggs detected
in their stool or was it a spurious infection? The answer to this
questionwas resolved 18months laterwhen anelderly lady from the
mid-north of South Australia presented with chronic diarrhoea.
Microscopic examination of her stool detected Brachylaima eggs,
which matched the morphology of the eggs detected in the two
children. In addition, on the first day following treatment with
praziquantel a gravid degenerate adult Brachylaima worm was
recovered from her stool5. This provided the evidence that a
Brachylaima sp.was infecting humans and completing its life cycle.
(a) (b)
Figure 1. (a) European helicid and hygromiid land snails aestivate over summer to escape the ground heat by attaching to fence posts,which presentsan easy meal for natural definitive hosts like birds. (b) Fertile Brachylaima cribbi egg being smooth shelled with an inconspicuous operculum,an abopercular knob or thickening and measuring 26–32mm (29.1mm) long and 16–17.5mm (16.6mm) wide.
Under theMicroscope
30 10.1071/MA16012 MICROBIOLOGY AUSTRALIA * MARCH 2016

However, due to the advanced state of decomposition of the worm
there was insufficient morphological detail to establish the species
identification.
Previous to the finding of the third case described above I had
commenced the cultivation of introduced European helicid and
hygromiid land snails (Figure 1a) in homemade terrariums using
techniques published for snail farming6. My office in the diagnostic
Microbiology laboratory had now become part snail culture labo-
ratory. These snailswereused to study the larval stages in an attempt
to understand how humans could have become infected. At the
same time I had obtained a copy of an article by Mas-Coma and
Montoliu detailing the life cycle of Brachylaima ruminae from the
duodenumofa rodent fromtheMediterranean islandofFormentera
(Spain)7. This article provided methods for the establishment of a
laboratory life cycle. With these methods in hand and my snail
cultures thriving I set myself the task to attempt to establish a
laboratory life cycle using eggs recovered from the stool of the third
patient. Faecal sediment of the patient’s stool containing washed
concentrated eggs was smeared on wet filter paper and was fed to
laboratory snails in Petri dishes overnight. Eight weeks later, when
the snails were exposed to moisture in a Petri dish, cercariae by
the thousands were observed emerging from the snails (Figure 2).
Cercariae were collected in water and used to infect the common
brown garden snails Cornu aspersum (Helix aspersa) via the
respiratory orifice. After 8–10 weeks mature metacercariae were
harvested from kidneys of the snails. Mice were inoculated orally
withmetacercariae. Sevenweeks later eggswere detected inmouse
(a)
(b) (c)
Figure 3.Brachylaimacribbi adult worm. (a) Oral (os) and ventral suckers (vs) and genital pore (gp). Bar = 500mm. (b) Ventral viewof the oral and ventralsuckers showing rows of tegumental spines. Bar = 100mm. (c) Genital pore showing extended cirrus (c) releasing sperm (s). Bar = 100mm.
Figure 2. Scanning electron microscopy ventral view of a Brachylaimacribbi cercaria showing oral (os), ventral suckers (vs) and elongatedsensory papilla (sp). Bar = 50mm.
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 31

faeces. Gravid adult Brachylaima worms were recovered from the
small intestine for characterisation andmorphological identification
(Figure 3). Eggs frommouse faeces and dissectedwormswere used
to continue the life cycle (Figure 4) in the laboratory to study the life
cycle kinetics.
At the same time as I was using my spare time to tinker with this life
cycle experiment the former Institute of Medical and Veterinary
Science (IMVS) was undergoing amalgamation with the Queen
Elizabeth Hospital (TQEH) Laboratories. Professor David Grove
was the Clinical Microbiologist Head of TQEH Microbiology
Department and a world recognised expert clinical parasitologist.
At that time Professor Grove offeredme the opportunity to join the
TQEH Microbiology team under the amalgamation program to
continue the Brachylaima study and commence a PhD under his
supervision. That is scientific luck, right place at the right time and
the rest is now history, with the following years being the most
simulating and productive science of my career.
(e)
(d)
(a)
(b)
(c)
Figure 4. Life cycle of Brachylaima cribbi. The definitive host sheds eggs (a) in their faeces. The egg is eaten by introduced European land snails(b), which hatch in the snail’s gut to release themiracidium. Themiracidiumdevelops into a sporocyst in the snail’s digestive gland.Mature cercariae(c) released from the sporocyst emerge from snail passing into the environment. Cercariae infect other land snails and develop into metacercariae(d) in the snail’s kidney. The definitive host (mammals, birds and reptiles) (e) ingest infected snails, releasing metacercariae, which migrate to thesmall intestine, attach and develop into mature adult worms. Humans are incidental definitive hosts being infected after eating raw snails containingmetacercariae.
Under theMicroscope
32 MICROBIOLOGY AUSTRALIA * MARCH 2016

Human cases
Since the publication of the three human cases to my knowledge
there have been a further 12 laboratory confirmedhuman infections
(A.R. Butcher and D.I. Grove, unpublished observations). Part of
my PhD studies was the follow-up of all human infections with a
standard questionnaire to obtain clinical history. The main present-
ing symptoms were mucoid, watery diarrhoea, abdominal pain,
anorexia and weight loss or poor weight gain in children. All
identified cases were in patients who lived in rural or semi-rural
districts where helicid and hygromiid land snails infected with
B. cribbimetacercariae were abundant. The likely infection source
was the ingestionof rawsnails. Themajorityof infections (80%)were
in children under the age of two years. At this age the mouthing of
objects is a common behavioural characteristic with parents of the
infected children detailing their child’s consumption of snails. For
the three adults infected they had all eaten home-grown garden
vegetables contaminated with snails. The duration of symptoms
ranged from 1 month to 2 years with the majority of infections
diagnosed within 1–2 months of the first signs of symptoms. All
patientswere treated successfully with praziquantel with no adverse
side-effects. The most commonly used dose in both adults and
children was 20 mg/kg once per day for 3 days.
Laboratory diagnosis
Diagnosis of the infection relies on the detection of typical eggs in
the faeces which are asymmetrical, being ovoid, with one side
slightly flattened. They have a smooth shell, inconspicuous oper-
culum, an abopercular knob or thickening and measure 26–32mm
by 16–17.5mm. The eggs generally contain a well-developed mira-
cidium (Fig. 1b). However, in chronic infections many of the eggs
may be infertile, being of similar morphology but smaller size and
lacking a developed miracidium. Infected land snails have been
reported from coastal and inland Western Australia, South Australia
and western Victoria8. Therefore, the potential exists for further
human infections, especially in children living in these regional
districts.
References1. Butcher, A.R. and Grove, D.I. (2001) Description of the life-cycle stages of Brachy-
laima cribbi n. sp. (Digenea: Brachylaimidae) derived from eggs recovered
from human faeces in Australia. Syst. Parasitol. 49, 211–221. doi:10.1023/A:1010
616920412
2. Butcher, A.R. et al. (1996) Locally acquired Brachylaima sp. (Digenea: Brachylai-
midae) intestinal fluke infection in two South Australian infants.Med. J. Aust. 164,
475–478.
3. Cribb, T.H. (1990) Introduction of a Brachylaima species (Digenea: Brachylaimi-
dae) to Australia. Int. J. Parasitol.20, 789–796. doi:10.1016/0020-7519(90)90013-D
4. Cribb, T.H. and O’Callaghan, M. (1992) An unusual trematode infecting domestic
chickens. Aust. Vet. J. 69, 69–70. doi:10.1111/j.1751-0813.1992.tb07456.x
5. Butcher, A.R. et al. (1998) First report of the isolation of an adult wormof the genus
Brachylaima (Digenea: Brachylaimidae), from the gastrointestinal tract of a
human. Int. J. Parasitol. 28, 607–610. doi:10.1016/S0020-7519(97)84372-X
6. Baker, G.H. (1991) Production of eggs and young snails by adult Theba pisana
(Müller) and Cernuella virgata (da Costa) (Mollusca: Helicidae) in laboratory
culture and field populations. Aust. J. Zool. 39, 673–679. doi:10.1071/ZO9910673
7. Mas-Coma, S. andMontoliu, I. (1986) The life cycle of Brachylaima ruminae n. sp.
(Trematoda: Brachylaimidae), a parasite of rodents. Z. Parasitenkd. 72, 739–753.
doi:10.1007/BF00925095
8. Butcher, A.R. and Grove, D.I. (2003) Field prevalence and laboratory susceptibility
of southern Australian land snails to Brachylaima cribbi sporocyst infection.
Parasite 10, 119–125. doi:10.1051/parasite/2003102119
Biography
Dr Andrew Butcher is a retired medical scientist with 38 years
experience as a diagnostic medical microbiologist with a special
interest in parasitology. He has been an active member of the ASM
Parasitology and Tropical Medicine Special Interest Group for
many years being national convenor for 7 years. He has been
involved in teaching diagnostic medical parasitology and one
of the founding committee members of the ASM Parasitology
Master Class.
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 33

Malaria: global challenges for malaria eradication
Graham Brown
The University of MelbourneVic. 3010, AustraliaEmail: [email protected]
Stephen Rogerson
The University of MelbourneVic. 3010, AustraliaEmail: [email protected]
Theenormousdecline in theannualmorbidity andmortality
frommalaria is the spectacular global public health success
of the past decade. This achievement results largely from
increased finance for investment in measures known to
prevent malaria: bednets treated with long-lasting insecti-
cides, chemoprophylaxis, and rapid access to effective treat-
ment.Suchhasbeenthesuccessof thesemeasures thatplans
are being put in place to achieve the vision of a malaria-free
world within the next three decades. Large financial and
political commitments and ongoing research will be re-
quired to maintain the gains, overcome known and un-
known challenges such as drug and insecticide resistance,
and to achieve those goals. Effective vaccines ormethods for
reducingmosquito vectorial capacity would add enormous-
ly to the chance of achieving this goal. The aim of this article
is to summarise the current status of malaria control, the
recent research successes, the challenges being addressed,
and the plan for progress to elimination of malaria in the
longer term.
Spectacular progress in reducing the burden
of malaria
Funding for major efforts in implementation of known effective
preventive measures in the past 15 years has caused an almost 40%
reduction in incidenceofmalaria disease episodes. Thesemeasures,
together with increased access to effective treatment, have led to
a reduction in malaria death rates of 60% globally, and 71% in
children under five1. It is estimated that between 2001 and 2015,
~6.2million lives were saved by increasing the provision of these
services. Approximately 110 countries are free from malaria, ~40
have committed to an elimination timetable, while 70 focus
predominantly on malaria control. Many factors contributed to this
success from the many constituencies that make up the Roll Back
Malaria Partnership2, but most of the progress in Africa is usually
attributed to widespread distribution of bednets treated with long-
lasting insecticides, combined with increased access to highly-
effective drugs3.
The current situation
According to the latest information provided by the WHO, ~1.2
billion people are at risk ofmalaria with themajor burden of disease
carried by young children and pregnant women living in endemic
areas.1
The burden of malaria is not evenly distributed across the world.
About 90% of malaria deaths occur in sub-Saharan Africa and recent
estimates suggest that 80% of malaria deaths occur in only 15
countries. The largemorbidity resulting fromP. vivax is seenmainly
in the Asia Pacific region. Further general information on malaria is
available in themost recentWorldMalariaReport fromWHO1.There
has been great recent interest in the finding that infections with the
simian malaria P. knowlesi have become increasingly important in
Malaysia and surrounding countries.
The strategy for malaria control leading
to elimination
The overall guiding strategy for addressingmalaria is summarised in
theGlobal Technical Strategy forMalaria Elimination 2016–2030,
which was approved and adopted by the World Health Assembly in
May 20154. The strategy calls for accelerated action towards malaria
elimination in countries and regions, but does not set a time frame
for global eradication. Notably this document recognises the con-
tinuum of activities from malaria control to elimination, the
Under theMicroscope
34 10.1071/MA16013 MICROBIOLOGY AUSTRALIA * MARCH 2016

importance of amulti-disciplinary approach, and the need for tailor-
made strategies for different situations, even within one country, in
other words, ‘One size does not fit all’. One area, for example a
mountainous region, may have only the problem of introduced
malaria occurring in people of all ages, whereas another highly
endemic area may need to focus on children and pregnant women
who remain at risk and suffer the consequences all year round.
The Global Malaria Programme of WHO provides support to coun-
tries, compiles the Annual Global Malaria Report, and through the
Malaria Policy Advisory Committee provides a normative role in
reviewing new data or new problems to provide advice for policy
implementation at country level.
This WHO strategy is complemented by the Roll Back Malaria
advocacy plan, Action and Investment to Defeat Malaria
2016–2030, that was approved in 20155. The publication outlines
both the health and economic benefits that wouldflow frommalaria
elimination, and like the Global Technical Strategy was the result of
extensive consultative processes involving the participation ofmore
than 400 malaria experts from 70 countries. Ambitious but achiev-
able global targets were established, including:
* reducing malaria case incidence by at least 90% by 2030* reducing malaria mortality rates by at least 90% by 2030* eliminating malaria in at least 35 countries by 2030* preventing a resurgenceofmalaria in all countries that aremalaria-free.
The timeline of 2016–2030 is aligned with the 2030 Agenda for
Sustainable Development, the new global development framework
adopted by all UN Member States in September.
Malaria elimination
The major progress already referred to is the result of implemen-
tation of known effective preventive measures, including vector
control with long-lasting insecticide-treated nets and indoor resid-
ual spraying; chemoprophylaxis including intermittent preventive
treatment of malaria in pregnancy (IPTp); intermittent preventive
treatment of malaria in infants (IPTi) or malaria chemoprevention
during the season of malaria transmission (SMC); and provision of
ready access to case management with rapidly-effective drugs,
primarily artemisinin combination treatments (ACTs).
As countries, or areas within countries, move towards elimination,
the emphasis changes from population level data and disease
control to intense surveillance for identification of individual infec-
tions, followed by planning and, most importantly, implementation
of a response. For example, teams might test everyone in a com-
pound or a village surrounding a person presenting with illness,
apply control measures locally, and search for the source of intro-
duced cases.
MalERA (Malaria Eradication Research Agenda)
Following the call for malaria eradication, the Bill & Melinda Gates
Foundation recognised the need for new and better tools for
elimination, andnewandbetter systems toguidedeliveryof services
to achieve those goals. Following extensive discussions by working
groups, a research agenda was developed (MalERA)6 that included:
* Better and safer drugs for ease of administration to populations.The ‘dream’ product profile is ‘SERCAP’, (Single EncounterRadical Cure And Prophylaxis), i.e. a single dose treatment to killnot only asexual blood-stage parasites but also liver and sexualstages. An intermediate goal would be an alternative and moreeffective drug to kill hypnozoites, the dormant liver stage respon-sible for relapse in P. vivax, but without the troublesome side-effects of primaquine, the only drug currently available for thatpurpose.
* Basic researchenablingcultureof all life cycle stagesof all parasitesinfecting humans would be required for development of vaccinesand for production of better drugs for eradication of hypnozoites.This would entail investment in culture of hepatic stages ofP. vivax, and achieving the dream of laboratory culture of spor-ozoites from gametocytes.
* The emphasis on elimination draws attention to P. vivax, com-mon outside Africa, and with separate challenges such as long-term relapses from hypnozoites, and transmission by vectorsbiting outdoors and during the daytime, which are therefore notamenable to the standard interventions for vector control.
Many considered that elimination would not be achieved without a
major breakthrough such as effective vaccines to reduce morbidity
and transmission, or a technology for vast reduction in vectorial
capacity of Anopheles gambiae, the most efficient and important
vector of P. falciparum in Africa.
It is a source of optimism that since the MalERA agenda was
proposed, fundingwasdirected to address important gaps, aMalaria
Eradication Scientific Alliance (MESA) was established, and now
through its MESA Track database tool can capture relevant research
anddevelopment projects. By the endofOctober 2015, 685projects
from 647 institutions and 84 countries had been documented7.
MESA is currently coordinating several expert groups to perform a
5-year ‘refresh’ of the MalERA agenda.
Progress is being made onmany other fronts, including conference
reports of the in vitro culture of sporozoites from gametocytes;
successful culture of hepatic stages of plasmodia; and development
of new antimalarial drugs, through ‘Medicines for Malaria Venture’
(MMV), a highly successful not-for-profit public-private partnership.
The timeframe for initial testing of newdrugs (or vaccines) has been
dramatically shortened by the development of a blood stage chal-
lenge model by Queensland-based researchers.
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 35

The challenges for malaria control
and elimination
See Table 1 for a more comprehensive list. Some of the biological,
societal and economic requirements for elimination are listed in
Table 2.
Artemisinin resistance
The time andmodeof action of artemisinin are notwell understood,
and in vitro tests are not sufficiently developed and standardised to
replace the requirement for in vivo testing for the presence of
resistance.Awarning sign, beforeemergenceof clinical resistance, is
the detection of delayed parasite clearance from the blood over
the first few days of treatment. Because of the fear of emergence
of resistance, combination therapy is essential, for example with
piperaquine or lumefantrine, so the efficacy of partner drugs is also
of great importance to ensure complete parasite clearance. Recent-
ly, drug resistance has been associated with some but not all
mutations of the kelch gene in P. falciparum that will probably
assist in mapping resistance8,9. Melbourne-based researchers have
developed synthetic artemisinin-like compounds, which are prob-
ably not susceptible to kelch-based resistance.
Insecticide resistance
Chemical resistance to insecticides is a well known threat. Strong
movements against DDT that is cheap and effective for indoor
residual spraying (because of past evidence of harmful environmen-
tal effects when very widely distributed in agriculture) mean that
there are few suppliers globally, and alternatives are very expensive.
Behavioural resistance is another well known complication of the
introduction of effective vector control directed at mosquitoes that
bite indoors in the evening. Repeated studies have documented the
gradual increase in proportion of mosquitoes biting earlier in the
evening or later in themorning and resting outdoors where they are
not susceptible.
Another huge challenge, particularly for control of P. vivax is that
the majority of vectors for this parasite bite outdoors and during
the daytime. No satisfactory method for long-term control of these
vectors is available.
Elimination as an approach to drug resistance
in the Mekong region
Where artemisinin has been widely used in the Mekong region,
delayed parasite clearance has been noted inmany countries, and it
is felt that in the absence of alternative drugs, the only appropriate
approach is to eliminate malaria in the area where resistance has
already been detected. Pilot studies have been initiated to test the
feasibility of this approach10.
Fortunately, delayed parasite clearance due to artemisinin resis-
tance has not yet been seen in parasites from India and Africa, but
in the recent past, development of resistance to chloroquine
or sulphadoxine/pyrimethamine accompanied by ongoing use of
those drugs led to many unnecessary deaths.
Table 1. Some of the challenges for malaria control and elimination.
Parasite and vector factors
Artemisinin resistance (and resistance to partner drugs)
P. vivax hypnozoites
Detecting infections at low parasitaemia
Vector resistance (chemical and behavioural)
Substandard and counterfeit drugs
Societal and economic
Inadequate health services to deliver interventions
Lack of financial commitment
Lack of political will
Cross border coordination
Underserved minorities at high risk
Civil unrest
Table 2. Preparing for elimination.
Biological approaches
SERCAP a drug for ‘Single Encounter Radical Cure And Prophylaxis’
Highly sensitive non-invasive point-of-care tests
Highly effective malaria vaccines
Novel vector control strategies
Societal and economic
Political will for a long-term program at high cost per case
Financial commitment
Experienced work force
Technical and operational capacity
Ongoing research
Under theMicroscope
36 MICROBIOLOGY AUSTRALIA * MARCH 2016

Funding shortfall
The major advances of the past two decades are at great risk from
lack of funding. Not only could progress stall, but major epidemics
of malaria could return to cleared areas, as has been seen so often
in the past11, with serious effects on people of all ages whose
immunity has declined through lack of boosting by recurrent
sub-clinical infections.
The case is now strong for the economic benefits that will follow
from the elimination of malaria5. This message needs to reach the
highest levels of government to ensure financial and political
commitment and the engagement not just of the health system,
but all sectors of society.
Strengthened Health Services
As themalariamapshrinks, the lastpocketsof transmission areoften
found in marginalised poor or rural populations with least access to
services. As episodes become infrequent, success will depend on a
general health service with the capacity to recognise and diagnose
infections, then respond with appropriate increased surveillance
to detect the origin of the infection and interventions to prevent
further spread.
Regional initiatives
APMEN Recognising the need for a partnership across regions and
across borders, the Asia Pacific Malaria Elimination Network was
establishedwith the supportof theAustraliangovernment in2008 to
facilitate collaboration amongst nations of the region that hadmade
commitments to achieve elimination within a defined time period.
This network enables regional collaboration and facilitates capacity
building and knowledge transfer among nations of the region that
share similar challenges such as artemisinin resistance, inadequate
finance for malaria, or a vast market penetration of counterfeit
drugs.
AICEM, the Australian Initiative for Control and Elimination of
Malaria, is funded by the Australian government to strengthen
malaria control in the Solomon Islands and Vanuatu, and at the
same time establish pilot projects for elimination of malaria in
certain island populations.
APLMA (www.aplma.org), the Asia Pacific Leaders Malaria Alli-
ance, co-chaired by the Heads of Government of Australia and
Vietnam, modelled loosely on the African Leaders Malaria Alliance,
aims to bring together Heads of State, politicians, and opinion
leaders of civil society to advocate for political and financial com-
mitment to the goals of elimination of malaria. Special task forces
are designed to address critical issues that could be barriers to
achievement of elimination such as financing, or the safety and
quality of medicines (to tackle the problem of counterfeit drugs
and ensure that only certified effective products are available in the
public andprivate sectors). Recently, theAllianceendorsedaMalaria
Elimination Roadmap towards a malaria-free Asia Pacific by 2030.
Elimination to eradication
The malaria eradication campaign of 50 years ago succeeded in
eliminatingmalaria frommanycountriesbutdidnot achieve its goals
in areas of most intense transmission such as heartland Africa.
However, many important lessons were learned about the need
for community engagement, political will, intensive surveillance as a
response, and the need for continued vigilance to prevent return of
malaria to a now susceptible entire population that has benefited
from a few years of interrupted transmission. As transmission
declines and malaria-specific services are phased out, it is critical
for awareness of the disease in all itsmanifestations to be retained at
a high level in general services for detection, management, surveil-
lance and response to any new episodes.
Lessons of the eradication campaign are being refreshed, for ex-
ample on mass drug administration to entire populations (MDA),
and comparing this strategy with treatment only of those found
with highly sensitive tests to be infected, thus avoiding the unnec-
essary use of drugs that could have side-effects, so called Mass
Screening and Treatment, or MSAT. Research on application of
moremodern tools such as rapid diagnostic tests, including nucleic
acid amplification or data transmission by mobile phones are
providing further data to guide strategy. For example, it is important
to decide whether the cost of a more sensitive but more expensive
and technically difficult nucleic acid detection test adds significant
benefit in elimination campaigns, or how PCR-based parasite
‘barcoding’ (genetic epidemiology) can be used to provide rapid
analysis of the source of emergent infections, the spread of drug
resistance, or the risk of the parasite’s appearance in susceptible
areas.
Othermajor lessons of the eradication campaignwere that research
must continue throughout the programs to deal with new chal-
lenges, and that every countryneeds todevelop thehuman resource
capacity to deal with the multi-disciplinary approach to this grand
challenge.
Recent vaccine breakthroughs; progress
with P. falciparum but little progress with
other species
Successful Phase 3 field trials of RTS,S vaccine in African
children. Development of the RTS,S vaccine, which provides
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 37

35% efficacy against clinical episodes of P. falciparummalaria, was
an important milestone for the malaria vaccine field. The vaccine
includes a fusion protein containing the immunogenic repeat
regions of the circumsporozoite protein, T-cell epitopes of
the same molecule and the hepatitis B surface antigen that is
co-expressed with free hepatitis B virus surface antigen leading to
formation of a viral-like particle known as RTS,S. In recent very large
trials, participants had good access to malaria prevention and rapid
access to treatment, somortalitywas lowandnoeffect of vaccination
on mortality could be detected. There was a concerning, but
unexplained, slight increase in episodes of meningitis in the RTS,S
arm, requiring further evaluation. A recent review of RTS,S byWHO
recommended that the next step should be to initiate 3–5 pilot
implementation studies12. Further analysis of trial data showed that
the vaccine had higher efficacy against strains homologous to the
vaccine, suggesting both a strain-specific as well as a strain-inde-
pendentmechanism13. This vaccine is a very importantfirst step but
on its own, is unlikely to be the solution that is required.
Attenuated sporozoite vaccines
The most impressive of all malaria vaccine trial results are those
achieved with intravenous inoculation of radiation-attenuated
whole sporozoites in which up to 100% protection has been
achieved in Phase 1 studies using multiple doses of vaccine14. Field
studies in Mali and Tanzania have also demonstrated efficacy and
further trials are planned to optimise dose and delivery schedule14.
Conclusion
With the spectacular reduction of the burden of malaria in the past
15 years, the time is ripe to re-double efforts both to prevent
resurgence and to increase resources with the goal of achieving
the health and economic benefits that would result from disease
elimination. Major progress is being reported in developing the
tools to add to current successful interventions from basic research
into biology of hypnozoites and transition of gametocytes to spor-
ozoites, to field trials of vaccines and innovations in surveillance and
epidemiology. A multi-disciplinary response coupled with strong
community engagement, and ongoing political and financial sup-
port will be required to maintain the current rate of decline in
malaria, and hopefully achieve the ambitious goal of elimination.
References1. World Health Organization (2015) Malaria: fact sheet no. 94. http://www.who.int/
mediacentre/factsheets/fs094/en/
2. World Health Organization (2016) Roll back malaria partnership. http://www.
rollbackmalaria.org
3. Bhatt, S. et al. (2015) The effect of malaria control on Plasmodium falciparum in
Africa between 2000 and 2015. Nature 526, 207–211. doi:10.1038/nature15535
4. World Health Organization (2015) Global technical strategy for malaria 2016–
2030. http://www.who.int/malaria/publications/atoz/9789241564991/en/.
5. World Health Organization Secretariat, on behalf of the Roll Back Malaria Pro-
gramme. (2015)Action and investment todefeatmalaria 2016–2030. For amalaria-
free world. http://www.rollbackmalaria.org/about/about-rbm/aim-2016-2030
6. (2011) malERA – a research agenda for malaria eradication (PLoS medicine
monographic volume). http://collections.plos.org/malera?issue=info:doi/10.1371/
issue.pcol.v07.i13
7. Malaria Eradication Scientific Alliance (2015) MESA track. http://www.malariaer-
adication.org/mesa-track
8. Ariey, F. et al. (2014) A molecular marker of artemisinin-resistant Plasmodium
falciparum malaria. Nature 505, 50–55. doi:10.1038/nature12876
9. Ashley, E.A. et al. (2014) Spread of artemisinin resistance in Plasmodium
falciparummalaria. N. Engl. J. Med. 371, 411–423. doi:10.1056/NEJMoa1314981
10. World Health Organization (2015) Draft strategy for malaria elimination in the
GreaterMekong subregion (2015–2030). http://www.who.int/malaria/areas/great-
er_mekong/consultation-elimination-strategy/en/
11. Cohen, J.M. et al. (2012) Malaria resurgence: a systematic review and assessment
of its causes. Malar. J. 11, 122. doi:10.1186/1475-2875-11-122
12. World Health Organization (2015) Pilot implementation of first malaria vaccine
recommended by WHO advisory groups. http://www.who.int/mediacentre/news/
releases/2015/sage/en/
13. Neafsey, D.E. et al. (2015) Genetic diversity and protective efficacy of the RTS,S/
AS01 malaria vaccine. N. Engl. J. Med. 373, 2025–2037. doi:10.1056/NEJMoa
1505819
14. Richie, T.L. et al. (2015) Progress with Plasmodium falciparum sporozoite
(PfSPZ)-based malaria vaccines. Vaccine 33, 7452–7461. doi:10.1016/j.vaccine.
2015.09.096
Biographies
Graham Brown, AM Professor Emeritus at The University of
Melbourne, currently serves as the Deputy Chair of the Board,
and Chair of the Executive Committee of Roll Back Malaria. As a
clinician-researcher, his interests include malaria vaccine develop-
ment, antigenic variation and clinical infectious diseases.
Stephen Rogerson is a Professor at The University of Melbourne.
His research interests include the pathogenesis of malaria in preg-
nant women and young children, and tools for prevention of
malaria in these high-risk groups.
Member of Australian Society for Microbiology (MASM)
Want to upgrade from an associate to a full member?No points or cash required and only one supporting referee needed.Go to the ASM website, download and complete the form, and forward it with a CV to the NationalOffice.We will do the rest.
Under theMicroscope
38 MICROBIOLOGY AUSTRALIA * MARCH 2016

Plasmodium knowlesi: an update
Balbir Singh
Malaria Research CentreUniversity Malaysia Sarawak94300 Kota SamarahanSarawak, MalaysiaTel: +6 082 581000Fax: +6 082 665152Email: [email protected]
There were only four species of Plasmodium that were
thought to cause malaria in humans until a large number
of human infections by Plasmodium knowlesi, a malaria
parasite typically found in long-tailed and pig-tailed maca-
ques,werereported in2004 inMalaysianBorneo.Since then,
cases of knowlesi malaria have been reported throughout
South-east Asia and also in travellers returning from the
region. This article describes the molecular, entomological
and epidemiological data which indicate that P. knowlesi is
an ancient parasite that is primarily zoonotic, and there are
three highly divergent sub-populations. It also describes the
detectionmethods forP. knowlesi, which ismorphologicaly
similar to P. malariae, and the clinical features and treat-
ment of this malaria parasite that is potentially fatal.
Malaria parasites and discovery of large focus
of human knowlesi malaria cases
Malaria is caused by parasites that belong to the genus Plasmodium
and there are more than 150 species of Plasmodium that infect
reptiles, birds andmammals1. These parasites, in general, tend to be
host-specific. Long-tailed and pig-tailed macaques (Macaca fasci-
cularis and M. nemestrina respectively) are hosts to five species
(P. knowlesi, P. inui, P. cynomolgi, P. fieldi and P. coatneyi). Only
four species of Plasmodium, namely P. falciparum, P. vivax,
P. malariae and P. ovale, were thought to causemalaria in humans
until a large number of human cases due to P. knowlesi were
reported in Sarawak, Malaysian Borneo over 11 years ago2. The
study in Kapit was prompted by observations that cases diagnosed
by microscopy as P. malariae had high parasitaemias, required
hospitalization and that 95% of patients were adults. This was in
contrast to P.malariae infections which typically are asymptomatic
with low parasitaemia and occur in all age groups. When blood
samples from 208 malaria patients at Kapit Hospital were analysed
by PCR assays, none were identified as P. malariae, although 141
had been diagnosed as P. malariae by microscopy. Fifty-eight
percent (120) were either single P. knowlesi infections or mixed
infections of P. knowlesi with P. falciparum and P. vivax. Misdi-
agnosis had occurred because the blood stages of P. knowlesi and
P. malariae are morphologically indistinguishable3.
Epidemiology and risk factors of acquiring
knowlesi malaria
Human infections with P. knowlesi have been reported throughout
Malaysia and in Thailand, Singapore, the Philippines, Vietnam,
Cambodia, Indonesia, Brunei, Myanmar and in the Nicobar and
Andaman Islands of India4,5. InMalaysia, P. falciparum and P. vivax
cases have declined over the pastfive years and P. knowlesihas now
become the most common cause of human malaria6,7. The true
incidence of knowlesi malaria is not known in other parts of South-
east Asia since not many large-scale studies have been undertaken
with molecular detection assays.
The geographical distribution of human P. knowlesi infections is
similar to that of the natural hosts ofP. knowlesi, the long-tailed and
pig-tailed macaques8. Reports from 1931 to 1970 identified maca-
ques as hosts of P. knowlesi in Peninsular Malaysia, Singapore and
the Philippines9, and a banded leaf monkey (Presbytis melalophos)
in Peninsular Malaysia9. Since 2007, P. knowlesi infections detected
by molecular methods have been described in macaques in Penin-
sular Malaysia, Malaysian Borneo, Singapore and Thailand4.
The transmission of P. knowlesi in nature has been shown to be
restricted to mosquitoes belonging to the Anopheles leucosphyrus
group10. The members of this forest-dwelling group of mosquitoes
that have been identified as vectors include An. latens (in Sarawak,
Malaysian Borneo)11, An. balabacensis balabacensis (in Sabah,
Malaysian Borneo), An. dirus (in Vietnam)12 and An. hackeri and
An. cracens (in Peninsular Malaysia)1,13.
People that are at risk of acquiring knowlesi malaria are those that
enter thehabitat of themacaque reservoir hosts and theAnopheline
vectors at dusk or later as this coincides with the peak biting time of
the vectors14,15. These include subsistence farmers, timber camp
workers, hunters, army personnel and also travelers to forests or
forest-fringe areas. Visitors to South-east Asia from Australia, USA,
Finland, Sweden,Germany, France, NewZealand, Taiwan and Japan
have acquired knowlesi malaria following holidays or working visits
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 10.1071/MA16014 39

to Malaysian Borneo, Peninsular Malaysia, Brunei, Thailand,
Indonesia and the Philippines16.
Molecular and whole genome studies
In order to understand the molecular epidemiology and demo-
graphic history of knowlesimalaria, themitochondrial (mt) genome
sequences of P. knowlesi were initially studied17. Certain mt hap-
lotypes were shared between humans and macaques and there
were no haplotypes that were associated exclusively with either
host; further evidence supportingP.knowlesi as a zoonoticparasite.
Additional analyses indicated that P. knowlesi was as old as, if not
older than, P. falciparum and P. vivax, and that it underwent a
population expansion between 30,000 to 40,000 years ago. Maca-
ques colonized Asia over 5million years ago18 and are probably the
original hosts forP. knowlesi. A recent study, where 599P. knowlesi
samples from Peninsular Malaysia and Malaysian Borneo were
analysed by a panel of ten microsatellite markers, showed there
are twohighly divergent sub-populations ofP. knowlesi, andeachof
these subpopulations correspond with parasites from either long-
tailed or pig-tailed macaques19. More recently, genome-wide se-
quence analysis of clinical P. knowlesi isolates from Malaysian
Borneo shows sub-population structure that matches the analysis
using microsatellite markers and also demonstrate there is a third
sub-population of parasites, corresponding to laboratory strains
isolated over 50 years ago from Peninsular Malaysia and the
Philippines20. No signals of positive selection were observed in
P. knowlesi around five orthologues of known P. falciparum
drug resistance genes, indicating that the parasites in the reservoir
macaque hosts have not been under antimalarial drug selection,
thereby providing further evidence that knowlesi malaria is a
zoonosis.
Diagnosis
In laboratories in malaria-endemic countries, malaria is diagnosed
by examination of blood films by microscopy. Under the micro-
scope, the early blood forms of P. knowlesi are identical to those
of P. falciparum, while the other developmental stages, including
the ‘band forms’, are similar to those of P. malariae3. There are
minor morphological differences between these two species. The
mature schizonts of P. knowlesi can contain up to 16 merozoites,
whereas those of P. malariae have between 6–123. However,
mature schizonts are not found in all blood films examined and in
diagnostic laboratories, where technologists are only trained to
recognise P. falciparum, P. vivax, P. ovale and P. malariae, most
P. knowlesi infections have been identified by microscopy as
P. malariae2,4,21. Although morphologically similar, P. knowlesi
parasites multiply every 24 h in the blood while this erythrocytic
cycle is 72 h for P. malariae9.
Molecular detection methods are the most sensitive and accurate
techniques for identification of P. knowlesi. These include single
and nested PCR assays, real-time PCR assays and loop-mediated
isothermal assays4. However, these assays are relatively expensive,
not rapid and are not readily available in resource-poor laboratories
where the majority of P. knowlesi infections are detected. Rapid
diagnostic tests (RDTs) for malaria are available, but the overall
sensitivity of detection of a small number of RDTs that have been
evaluated against knowlesi malaria cases varied between 26–74%
andwas even lower (0–45%) for parasitaemias below1000parasites/
mL22–24. Due to the rapid multiplication rate of P. knowlesi in
the blood of 24 h, sensitive RDTs capable of detecting knowlesi
malaria at the early phase of infection are urgently required for rural
laboratories.
Clinical and laboratory features
of knowlesi malaria
P. knowlesi causes a wide spectrum of disease, from asymptomatic
infections9,25 to fatal ones26–29. Themost common presenting signs
and symptoms reported are fever with chills, followed by headache,
myalgia, poor appetite, arthralgia, cough, abdominal pain and
diarrhoea27. These are not significantly different to those observed
in patients with vivax and falciparum malaria. The majority of cases
(93.5%27 and 84.5%30) at district hospitals in Sarawak had uncom-
plicated malaria with a fatality rate of 2%, whereas in a retrospective
study in a referral hospital in Sabah, 61% of 56 cases were uncom-
plicated and the fatality rate was 27%31. However, subsequently at
the same referral hospital, the use of intravenous artesunate for
severe malaria cases and artemisinin combination therapy for non-
severe cases, resulted in no deaths among 130 knowlesi malaria
patients29. Typical complications of severe knowlesi malaria in
adults include jaundice, acute kidney injury, hypotension, acute
respiratory distress syndrome andmetabolic acidosis26,27,29,30,32. In
adults, severe anaemia has not been observed and neither has
cerebral malaria, while severe disease has not been noted in the
relatively small number of children with knowlesi malaria4,33.
Thrombocytopaenia is very common, occurring in 97.3 to 100% of
knowlesi malaria patients, and together with parasitaemia, corre-
lates with severity of disease27,30,31. Following a case control study,
it was recommended that any patient with a platelet count of
<45 000/mL or parasitaemia of >35 000 parasites/mL should be
regarded at risk of developing complications and should be treated
for severe malaria30.
Under theMicroscope
40 MICROBIOLOGY AUSTRALIA * MARCH 2016

Treatment of knowlesi malaria
Since knowlesi malaria is primarily a zoonosis, the parasites have
beenunder no antimalarial drug pressure and should be susceptible
to all antimalarials. This has been observed in hospital-based studies
as well as case reports where several antimalarials have been used
successfully to treat knowlesi malaria patients4. P. knowlesi para-
sites are highly sensitive to chloroquine34 but following an informal
consultation on the public health importance of knowlesi malaria
organised by the WHO in 2011, it was recommended that in areas
where knowlesi malaria has been detected, all infections diagnosed
as P. malariae by microscopy should be treated and managed as
for falciparum malaria35. Therefore, for uncomplicated knowlesi
malaria cases in South-east Asia, artemisinin combination therapy is
recommended. For severe knowlesi malaria, intravenous antimalar-
ials should be administered and the use of artesunate in a tertiary
referral hospital in Sabah was associated with zero mortality29.
Future directions
The available molecular, entomological and epidemiological data
strongly indicate that knowlesi malaria is primarily a zoonosis.
However, human-to-human transmission has been demonstrated
under experimental conditions9 and it is not known whether it is
currently occurring. The reasons for the increase in the number of
knowlesi malaria cases, particularly in Malaysian Borneo, are also
unknown. Whether the increase is due to increased awareness,
changes in the feeding habits of the vectors, the destruction of the
natural habitats of themacaque reservoir, humanmigration to areas
close to macaque habitats, a recent adaptation of knowlesi malaria
parasites to humans, or to some other factors needs to be investi-
gated. In addition, currently available methods of control of human
malaria involving the use of insecticide treated bednets and residual
spraying of houses are ineffective against knowlesi malaria, where
transmission primarily occurs outdoors. Therefore, effective meth-
odsofpreventionandcontrolneed tobe foundand implemented, in
order to prevent P. knowlesi from establishing itself in the human
population.
References1. Garnham, P.C.C. (1966) Malaria parasites and other haemosporidia. Blackwell
Scientific Publications.
2. Singh, B. et al. (2004) A large focus of naturally acquired Plasmodium knowlesi
infections in human beings. Lancet 363, 1017–1024. doi:10.1016/S0140-6736(04)
15836-4
3. Lee, K.S. et al. (2009) Morphological features and differential counts of Plasmo-
dium knowlesi parasites in naturally acquired human infections. Malar. J. 8, 73.
doi:10.1186/1475-2875-8-73
4. Singh, B. and Daneshvar, C. (2013) Human infections and detection of Plasmo-
dium knowlesi. Clin. Microbiol. Rev. 26, 165–184. doi:10.1128/CMR.00079-12
5. Tyagi, R.K. et al. (2013) Discordance in drug resistance-associated mutation
patterns inmarker genes of Plasmodium falciparum and Plasmodium knowlesi
during coinfections. J. Antimicrob. Chemother. 68, 1081–1088. doi:10.1093/jac/
dks508
6. William, T. et al. (2013) Increasing incidence of Plasmodium knowlesi malaria
following control of P. falciparum and P. vivax Malaria in Sabah, Malaysia. PLoS
Negl. Trop. Dis. 7, e2026. doi:10.1371/journal.pntd.0002026
7. Yusof, R. et al. (2014) High proportion of knowlesi malaria in recent malaria cases
in Malaysia. Malar. J. 13, 168. doi:10.1186/1475-2875-13-168
8. Cox-Singh, J. and Singh, B. (2008) Knowlesimalaria: newly emergent and of public
health importance? Trends Parasitol. 24, 406–410. doi:10.1016/j.pt.2008.06.001
9. Coatney, G.R. et al. (1971) The primate malarias. US Department of Health,
Education and Welfare.
10. Collins, W.E. (2012) Plasmodium knowlesi: a malaria parasite of monkeys and
humans. Annu. Rev. Entomol. 57, 107–121. doi:10.1146/annurev-ento-121510-
133540
11. Vythilingam, I. et al. (2006) Natural transmission of Plasmodium knowlesi to
humans by Anopheles latens in Sarawak, Malaysia. Trans. R. Soc. Trop. Med. Hyg.
100, 1087–1088. doi:10.1016/j.trstmh.2006.02.006
12. Marchand, R.P. et al. (2011) Co-infections of Plasmodium knowlesi,
P. falciparum, and P. vivax among humans and Anopheles dirus mosquitoes,
Southern Vietnam. Emerg. Infect. Dis. 17, 1232–1239. doi:10.3201/eid1707.
101551
13. Vythilingam, I. et al. (2008) Plasmodium knowlesi in humans, macaques and
mosquitoes in peninsular Malaysia. Parasit. Vectors 1, 26. doi:10.1186/1756-3305-
1-26
14. Tan, C.H. et al. (2008) Bionomics of Anopheles latens in Kapit, Sarawak, Malaysian
Borneo in relation to the transmission of zoonotic simian malaria parasite
Plasmodium knowlesi. Malar. J. 7, 52. doi:10.1186/1475-2875-7-52
15. Vythilingam, I. (2012) Plasmodium knowlesi and Wuchereria bancrofti: their
vectors and challenges for the future. Front. Physiol. 3, 115. doi:10.3389/
fphys.2012.00115
16. Cramer, J.P. (2015) Plasmodium knowlesimalaria: overview focussing on travel-
associated infections.Curr. Infect. Dis. Rep.17, 8. doi:10.1007/s11908-015-0469-6
17. Lee, K.S. et al. (2011) Plasmodium knowlesi: reservoir hosts and tracking the
emergence in humans and macaques. PLoS Pathog. 7, e1002015. doi:10.1371/
journal.ppat.1002015
18. Ziegler, T. et al. (2007)Molecular phylogeny and evolutionary history of Southeast
Asianmacaques forming theM. silenus group.Mol. Phylogenet. Evol.42, 807–816.
doi:10.1016/j.ympev.2006.11.015
19. Divis, P.C. et al. (2015) Admixture in humans of two divergent Plasmodium
knowlesi populations associated with different macaque host species. PLoS
Pathog. 11, e1004888. doi:10.1371/journal.ppat.1004888
20. Assefa, S. et al. (2015) Population genomic structure and adaptation in the
zoonotic malaria parasite Plasmodium knowlesi. Proc. Natl. Acad. Sci. USA
112, 13027–13032. doi:10.1073/pnas.1509534112
21. William, T. et al. (2014) Changing epidemiology of malaria in Sabah, Malaysia:
increasing incidence of Plasmodium knowlesi. Malar. J. 13, 390. doi:10.1186/
1475-2875-13-390
22. Foster, D. et al. (2014) Evaluation of three rapid diagnostic tests for the detection
of human infections with Plasmodium knowlesi. Malar. J. 13, 60. doi:10.1186/
1475-2875-13-60
23. Barber, B.E. et al. (2013) Evaluation of the sensitivity of a pLDH-based and an
aldolase-based rapid diagnostic test for diagnosis of uncomplicated and severe
malaria caused by PCR-confirmed Plasmodium knowlesi, Plasmodium falci-
parum, and Plasmodium vivax. J. Clin. Microbiol. 51, 1118–1123. doi:10.1128/
JCM.03285-12
24. Grigg, M.J. et al. (2014) Combining parasite lactate dehydrogenase-based and
histidine-rich protein 2-based rapid tests to improve specificity for diagnosis of
malaria due to Plasmodium knowlesi and other Plasmodium species in Sabah,
Malaysia. J. Clin. Microbiol. 52, 2053–2060. doi:10.1128/JCM.00181-14
25. Fornace, K.M. et al. (2015) Asymptomatic and submicroscopic carriage of
Plasmodium knowlesi malaria in household and community members of
clinical cases in Sabah, Malaysia. J. Infect. Dis. doi:10.1093/infdis/jiv475
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 41

26. Cox-Singh, J. et al. (2008) Plasmodium knowlesi malaria in humans is widely
distributed and potentially life threatening. Clin. Infect. Dis. 46, 165–171.
doi:10.1086/524888
27. Daneshvar, C. et al. (2009) Clinical and laboratory features of human
Plasmodium knowlesi infection. Clin. Infect. Dis. 49, 852–860. doi:10.1086/
605439
28. Rajahram,G.S.et al. (2012)Deathsdue toPlasmodiumknowlesimalaria in Sabah,
Malaysia: association with reporting as Plasmodium malariae and delayed
parenteral artesunate. Malar. J. 11, 284. doi:10.1186/1475-2875-11-284
29. Barber, B.E. et al. (2013) A prospective comparative study of knowlesi,
falciparum, and vivax malaria in Sabah, Malaysia: high proportion with severe
disease from Plasmodium knowlesi and Plasmodium vivax but no mortality
with early referral and artesunate therapy. Clin. Infect. Dis. 56, 383–397.
doi:10.1093/cid/cis902
30. Willmann, M. et al. (2012) Laboratory markers of disease severity in Plasmodium
knowlesi infection: a case control study. Malar. J. 11, 363. doi:10.1186/1475-
2875-11-363
31. William, T. et al. (2011) Severe Plasmodium knowlesi malaria in a tertiary care
hospital, Sabah, Malaysia. Emerg. Infect. Dis. 17, 1248–1255. doi:10.3201/eid
1707.101017
32. Cox-Singh, J. et al. (2010) Severe malaria – a case of fatal Plasmodium knowlesi
infection with post-mortem findings: a case report. Malar. J. 9, 10. doi:10.1186/
1475-2875-9-10
33. Barber, B.E. et al. (2011)Plasmodiumknowlesimalaria in children.Emerg. Infect.
Dis. 17, 814–820. doi:10.3201/eid1705.101489
34. Daneshvar, C. et al. (2010) Clinical and parasitological response to oral chloro-
quine and primaquine in uncomplicated human Plasmodium knowlesi infec-
tions. Malar. J. 9, 238. doi:10.1186/1475-2875-9-238
35. WHO (2011) Informal consultation on the public health importance of Plasmo-
diumknowlesi.WorldHealthOrganizationRegionalOffice for theWesternPacific
Press.
Biography
Professor Balbir Singh is the Director of the Malaria Research
Centre at University Malaysia Sarawak. His research interests
include the epidemiology, pathogenesis and evolution of malaria
parasites.
Under theMicroscope
42 MICROBIOLOGY AUSTRALIA * MARCH 2016

Diagnosis of human taeniasis
Abdul Jabbar, Charles Gauci and Marshall W Lightowlers
Faculty of Veterinary and Agricultural Sciences, The University of Melbourne, Werribee, Vic. 3030, Australia, Email: [email protected]
Taenia solium, T. saginata and T. asiatica are taeniid tape-
worms that cause taeniasis in humans and cysticercosis in
intermediate host animals. T. solium can also cause cysti-
cercosis in humans. A number of diagnostic methods have
been developed to diagnose Taenia species that infect
humans. This article is aimed at providing an overview of
currently available diagnosticmethods for human taeniasis.
Human taeniasis is an important zoonotic health problem and is
causedby adult stages of taeniid cestodes, includingTaenia solium,
T. saginata, and T. asiatica. These parasites have indirect life cycles
wherehumansact as definitivehostswhereaspigs (forT. solium and
T. asiatica) and cattle (T. saginata) serve as intermediate hosts.
Humans become infected by ingesting parasite cysts present in raw
or undercooked infected meat/liver; when the cyst reaches the
human intestine, it develops into an adult tapeworm, releasing
segments and/or eggs in the stools or motile segments (e.g.
T. saginata) are expelled actively. Intermediate hosts such as cattle
and pigs become infected when they ingest taeniid eggs via con-
taminated feed or water and the larval stage (cysticercus) forms in
muscular and sometimes other tissues1. The adult tapeworm stage
of Taenia spp. is relatively innocuous and does not cause patho-
genic effects in humans; however, the intermediate stage of
T. solium can also develop in human brains causing neurocysticer-
cosis, a major cause of neurological disease in many developing
countries as well as other organs causing intramuscular, ocular,
subcutaneous and spinal cysticercoses2,3. T. saginata is endemic in
Australia, whereas T. solium and T. asiatica are exotic. However,
people coming and/or returning to Australia from endemic coun-
tries can be infected with T. solium, leading to the possibility that
infected individualsmaypass segmentsof theparasite in their stools,
which can serve as a source of infection for human cysticercosis.
Therefore, differentiation ofTaenia species becomes significant for
surveillance and control of human taeniasis.
A number of diagnosticmethods have beenused todifferentiate the
common human cestodes, T. saginata and T. solium; however,
each method has its advantages and disadvantages, and careful
attention should be paid to determining which particular test is
best to use for differentiation of the two species4. The following
sections provide a quick rundown on various diagnostic methods
available to differentiate Taenia spp. that infect humans.
Microscopic diagnosisTraditionally, diagnosis of taeniasis has been based on the detection
of eggs by microscopic examination but this method lacks
sensitivity and specificity as T. solium and T. saginata eggs are
morphologically identical, making species identification impossi-
ble5.However,morphological examinationof gravidproglottids can
allow the differentiation of T. solium and T. saginata provided the
internal structures (i.e. uterine branches) are intact5. Sometimes,
even the morphological examination of proglottids does not allow
the differentiation of T. solium and T. saginata, thus requiring
alternate methods for the differentiation of human Taenia spp.
ImmunodiagnosisThefirstmethodof coproantigen (parasite antigens inhuman stool)
detection using enzyme-linked immunosorbent assay (ELISA) was
developed by Allan et al.6. Although the test displayed a higher
sensitivity and specificity than microscopic diagnosis for the detec-
tion of Taenia spp.7, it did not allow differentiation of T. saginata
andT. solium. Recently,Guezala et al.8 developed another coproan-
tigen ELISA and successfully differentiated T. solium from
T. saginata. These tests were developed in individual labs using
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 10.1071/MA16011 43
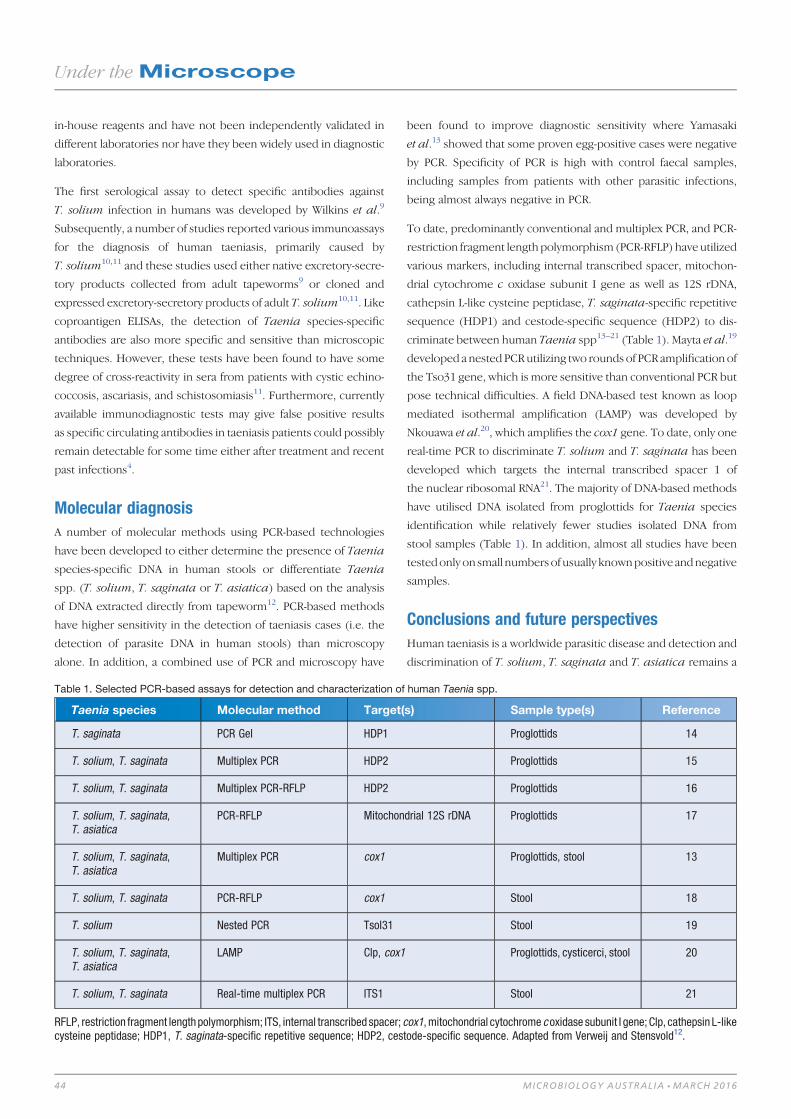
in-house reagents and have not been independently validated in
different laboratories nor have they been widely used in diagnostic
laboratories.
The first serological assay to detect specific antibodies against
T. solium infection in humans was developed by Wilkins et al.9
Subsequently, a number of studies reported various immunoassays
for the diagnosis of human taeniasis, primarily caused by
T. solium10,11 and these studies used either native excretory-secre-
tory products collected from adult tapeworms9 or cloned and
expressed excretory-secretory products of adult T. solium10,11. Like
coproantigen ELISAs, the detection of Taenia species-specific
antibodies are also more specific and sensitive than microscopic
techniques. However, these tests have been found to have some
degree of cross-reactivity in sera from patients with cystic echino-
coccosis, ascariasis, and schistosomiasis11. Furthermore, currently
available immunodiagnostic tests may give false positive results
as specific circulating antibodies in taeniasis patients could possibly
remain detectable for some time either after treatment and recent
past infections4.
Molecular diagnosisA number of molecular methods using PCR-based technologies
have been developed to either determine the presence of Taenia
species-specific DNA in human stools or differentiate Taenia
spp. (T. solium, T. saginata or T. asiatica) based on the analysis
of DNA extracted directly from tapeworm12. PCR-based methods
have higher sensitivity in the detection of taeniasis cases (i.e. the
detection of parasite DNA in human stools) than microscopy
alone. In addition, a combined use of PCR and microscopy have
been found to improve diagnostic sensitivity where Yamasaki
et al.13 showed that some proven egg-positive cases were negative
by PCR. Specificity of PCR is high with control faecal samples,
including samples from patients with other parasitic infections,
being almost always negative in PCR.
To date, predominantly conventional and multiplex PCR, and PCR-
restriction fragment lengthpolymorphism (PCR-RFLP) haveutilized
various markers, including internal transcribed spacer, mitochon-
drial cytochrome c oxidase subunit I gene as well as 12S rDNA,
cathepsin L-like cysteine peptidase, T. saginata-specific repetitive
sequence (HDP1) and cestode-specific sequence (HDP2) to dis-
criminate between human Taenia spp13–21 (Table 1). Mayta et al.19
developed a nested PCRutilizing two rounds of PCR amplification of
the Tso31 gene, which is more sensitive than conventional PCR but
pose technical difficulties. A field DNA-based test known as loop
mediated isothermal amplification (LAMP) was developed by
Nkouawa et al.20, which amplifies the cox1 gene. To date, only one
real-time PCR to discriminate T. solium and T. saginata has been
developed which targets the internal transcribed spacer 1 of
the nuclear ribosomal RNA21. The majority of DNA-based methods
have utilised DNA isolated from proglottids for Taenia species
identification while relatively fewer studies isolated DNA from
stool samples (Table 1). In addition, almost all studies have been
testedonlyon small numbersof usually knownpositive andnegative
samples.
Conclusions and future perspectivesHuman taeniasis is a worldwide parasitic disease and detection and
discrimination of T. solium, T. saginata and T. asiatica remains a
Table 1. Selected PCR-based assays for detection and characterization of human Taenia spp.
Taenia species Molecular method Target(s) Sample type(s) Reference
T. saginata PCR Gel HDP1 Proglottids 14
T. solium, T. saginata Multiplex PCR HDP2 Proglottids 15
T. solium, T. saginata Multiplex PCR-RFLP HDP2 Proglottids 16
T. solium, T. saginata,T. asiatica
PCR-RFLP Mitochondrial 12S rDNA Proglottids 17
T. solium, T. saginata,T. asiatica
Multiplex PCR cox1 Proglottids, stool 13
T. solium, T. saginata PCR-RFLP cox1 Stool 18
T. solium Nested PCR Tsol31 Stool 19
T. solium, T. saginata,T. asiatica
LAMP Clp, cox1 Proglottids, cysticerci, stool 20
T. solium, T. saginata Real-time multiplex PCR ITS1 Stool 21
RFLP, restriction fragment length polymorphism; ITS, internal transcribed spacer; cox1,mitochondrial cytochrome coxidase subunit I gene; Clp, cathepsin L-likecysteine peptidase; HDP1, T. saginata-specific repetitive sequence; HDP2, cestode-specific sequence. Adapted from Verweij and Stensvold12.
Under theMicroscope
44 MICROBIOLOGY AUSTRALIA * MARCH 2016

public health concern. Microscopic, immunological and molecular
methods have been used to detect and differentiate Taenia spp.,
and a combination of two or more methods appears to provide
higher sensitivity13,21. Almost all of the immunological and molec-
ular methods developed so far have not been independently tested
and validated on controlled negative and positive samples as well as
field samples. Amongexistingmolecularmethods, thenestedPCR19
provides thehighest sensitivity and specificityof thosemethods that
have been developed and validated using unselected faecal samples
fromparasitological proven taeniasis carriers. Although themethod
is technically challenging and could be expected to be relatively
expensive to use, the reagents required are available worldwide
and laboratories capable of undertaking PCR competently can run
this PCR4.
To the best of our knowledge, no standardised PCRs are available in
Australia to discriminate human Taenia spp. Future studies should
focus on independent validation of the nested PCR19 and the LAMP
technique20 as these two DNA-based tests offer higher sensitivity
and a user-friendly option without sophisticated equipment, re-
spectively. In addition, future studies should be aimed at extracting
DNA from sodium acetate-acetic acid-formalin (SAF) fixed proglot-
tidsusingdifferentmethods as thecurrentDNAextractionprotocols
do not provide reliable and consistent DNA yield for PCRs (Jabbar,
unpublished data). To date only one study has reported the extrac-
tion of DNA from long-term stored Taenia specimens in formalin22
and this has not been independently verified.
References1. Roberts, L.S. et al. (2013) Foundations of Parasitology, 9th edn. McGraw-Hill
Education, USA.
2. Mahanty, S. et al. (2010) Cysticercosis and neurocysticercosis as pathogens
affecting the nervous system. Prog. Neurobiol. 91, 172–184. doi:10.1016/
j.pneurobio.2009.12.008
3. García, H.H. et al. (2003) Taenia solium cysticercosis. Lancet 362, 547–556.
doi:10.1016/S0140-6736(03)14117-7
4. Lightowlers, M.W. et al. (2015) Monitoring the outcomes of interventions against
Taenia solium: option and suggestion. Parasite Immunol. In press. doi:10.1111/
pim.12291
5. Chapman, A. et al. (1995) Isolation and characterization of species-specific
DNA probes from Taenia solium and Taenia saginata and their use in an egg
detection assay. J. Clin. Microbiol. 99, 1283–1288.
6. Allan, J.C. et al. (1990) Immunodiagnosis of taeniasis by coproantigen detection.
Parasitology 101, 473–477. doi:10.1017/S0031182000060686
7. Allan, J.C. et al. (1996) Field trial of the coproantigen-based diagnosis of Taenia
solium taeniasis by enzymelinked immunosorbent assay. Am. J. Trop. Med. Hyg.
54, 352–356.
8. Guezala, M.C. et al. (2009) Development of a species-specific coproantigen ELISA
for human Taenia solium taeniasis. Am. J. Trop. Med. Hyg. 81, 433–437.
9. Wilkins, P.P. et al. (1999) Development of a serologic assay to detect Taenia
solium taeniasis. Am. J. Trop. Med. Hyg. 60, 199–204.
10. Levine, M.Z. et al. (2004) Characterization, cloning, and expression of two
diagnostic antigens for Taenia solium tapeworm infection. J. Parasitol. 90,
631–638. doi:10.1645/GE-189R
11. Levine, M.Z. et al. (2007) Development of an enzyme-linked immunoelectro-
transfer blot (EITB) assay using two baculovirus expressed recombinant antigens
for diagnosis of Taenia solium taeniasis. J. Parasitol. 93, 409–417. doi:10.1645/
GE-938R.1
12. Verweij, J.J. and Stensvold, C.R. (2014) Molecular testing for clinical diagnosis and
epidemiological investigations of intestinal parasitic infections. Clin. Microbiol.
Rev. 27, 371–418. doi:10.1128/CMR.00122-13
13. Yamasaki, H. et al. (2003) Cysticercosis/taeniasis: recent advances in serological
andmolecular diagnoses. Southeast Asian J. Trop.Med. PublicHealth34, 98–102.
14. Gonzalez, L.M. (2000) Differential diagnosis of Taenia saginata and Taenia
solium infection by PCR. J. Clin. Microbiol. 38, 737–744.
15. González, L.M. et al. (2002) Differential diagnosis of Taenia saginata and Taenia
solium infections: from DNA probes to polymerase chain reaction. Trans. R. Soc.
Trop. Med. Hyg. 96, S243–S250. doi:10.1016/S0035-9203(02)90083-0
16. González, L.M. et al. (2002) PCR tools for the differential diagnosis of Taenia
saginata and Taenia solium taeniasis/cysticercosis from different geographical
locations.Diagn.Microbiol. Infect. Dis.42, 243–249. doi:10.1016/S0732-8893(01)
00356-X
17. Rodriguez-Hidalgo, R. et al. (2002) Comparison of conventional techniques to
differentiate between Taenia solium and Taenia saginata and an improved
polymerase chain reaction-restriction fragment length polymorphism assay using
a mitochondrial 12S rDNA fragment. J. Parasitol. 88, 1007–1011. doi:10.1645/
0022-3395(2002)088[1007:COCTTD]2.0.CO;2
18. Nunes, C.M. et al. (2005) Taenia saginata: differential diagnosis of human
taeniasis by polymerase chain reaction-restriction fragment length polymorphism
assay. Exp. Parasitol. 110, 412–415. doi:10.1016/j.exppara.2005.03.008
19. Mayta, H. et al. (2008) Nested PCR for specific diagnosis of Taenia solium
taeniasis. J. Clin. Microbiol. 46, 286–289. doi:10.1128/JCM.01172-07
20. Nkouawa, A. et al. (2009) Loop-mediated isothermal amplification method for
differentiation and rapid detection of Taenia species. J. Clin. Microbiol. 47,
168–174. doi:10.1128/JCM.01573-08
21. Praet, N. et al. (2013) Bayesian modelling to estimate the test characteristics of
coprology, coproantigen ELISA and a novel real-time PCR for the diagnosis of
taeniasis. Trop. Med. Int. Health 18, 608–614. doi:10.1111/tmi.12089
22. Jeon, H.-K. et al. (2011) Molecular identification of Taenia specimens after long-
term preservation in formalin. Parasitol. Int. 60, 203–205. doi:10.1016/j.parint.
2010.12.001
BiographiesDr Abdul Jabbar is a Senior Lecturer in Veterinary Parasitology at
The University of Melbourne. His main research interests cover
epidemiology and diagnosis of parasites of socioeconomic impor-
tance using next-generation molecular tools.
Dr Charles Gauci is a Senior Research Fellow at The University of
Melbourne. He has worked with Prof Marshall Lightowlers through-
out his career and his research interests focus on recombinant
vaccines for prevention of transmission of the parasite causing
neurocysticercosis and the related parasite that causes hydatid
disease.
Professor Marshall W Lightowlers has been a full-time research
scientist supported bymedical research funding formore-or-less all
of his working life. He currently holds appointments as Laureate
Professor at the University of Melbourne’s Faculty of Veterinary
and Agricultural Sciences, and Principal Research Fellow with the
NHMRC.
Under theMicroscope
MICROBIOLOGY AUSTRALIA * MARCH 2016 45

Protozoa PCR: boon or bane
Colin Pham
St Vincent’s HospitalMelbourne, Vic., AustraliaEmail: [email protected]
Harsha Sheorey
St Vincent’s HospitalMelbourne, Vic., AustraliaEmail: [email protected]
Parasitedetection in faeceshas traditionallybeenperformed
by microscopy, a procedure that is labour-intensive and
highly specialised. In addition, identification bymicroscopy
based on morphological features alone is subjective and
prone to wide variability. Although enzyme immunoassays
(EIA) of high sensitivity have been developed1 they can
detect only a limited range of pathogens. Given these
factors the introduction of Polymerase Chain Reaction
(PCR) into the routine diagnostic laboratory has improved
parasite detection rates2. The ability to multiplex has en-
abled the detection of multiple targets from a single sample
and provides an objective alternative to identification by
morphology.
Results
In February 2014, St Vincent’s Pathology, Melbourne, introduced a
new testing algorithm making use of a commercial multiplex PCR
(LightMix�Gastroenteritis Parasite Kit, Roche Molecular Systems,
Pleasanton, California, USA) for parasite detection. This assay is
designed to detect Giardia intestinalis, Cryptosporidium species,
Dientamoeba fragilis and Entamoeba histolytica, four human gas-
trointestinal protozoanpathogens3.Blastocystiswasnot includedasa
target in thismultiplex assay, as theprimersdonotdifferentiate the17
differentsubtypesnowknown4,manybeingofanimaloriginandmost
of doubtful clinical significance.
Since the commencement of testing by PCR, the rates of detection
have improved when compared with results obtained based on
the previous protocol that included microscopy and Giardia/
Cryptosporidium antigen testing (Table 1). Comparison over a
three month period showed that the range of common parasites
detected in the population remains similar with D. fragilis
accounting for the majority of the additional parasites detected
(Figure 1). The reduction of Cryptosporidium cases from 18 (0.8%)
to 4 (0.4%) was not statistically significant and is thought to relate
to seasonal variation in prevalence. The additional E. histolytica
cases were detected from microscopy-positive specimens referred
from other laboratories. In addition, the assay was positive for
Table 1. Three months comparison between pre and post implementation of protozoa PCR testing.
Quarter 2: 2013Pre PCR
Quarter 2: 2014Post PCR
Statistical significance
Samples 2198 1096 P < 0.001
G. intestinalis 24 (1.1%) 33 (3.0%) P < 0.001
C. parvum 18 (0.8%) 4 (0.4%) P= 0.132
E. histolytica – 4 (0.4%) P= 0.005
D. fragilis 47 (2.1%) 163 (14.9%) P < 0.001
Total positives 89 (4%) 204 (19%) P < 0.001
LabReport
46 10.1071/MA16007 MICROBIOLOGY AUSTRALIA * MARCH 2016
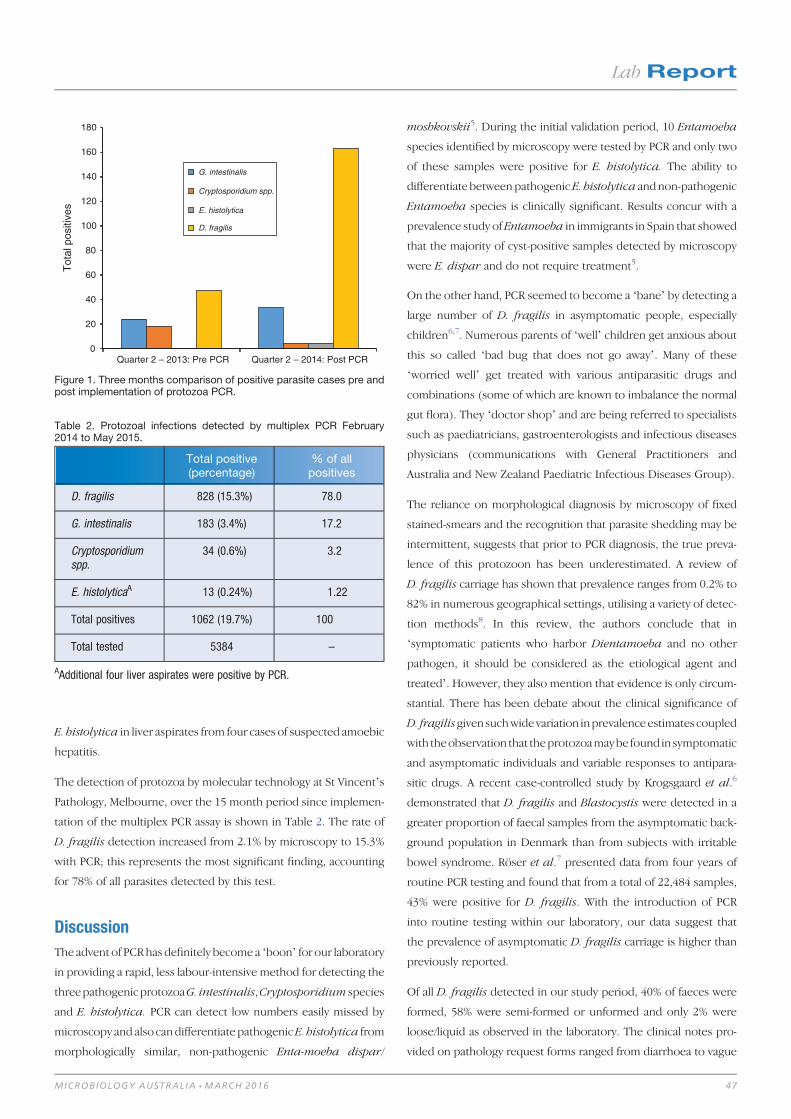
E. histolytica in liver aspirates from four cases of suspected amoebic
hepatitis.
The detection of protozoa by molecular technology at St Vincent’s
Pathology, Melbourne, over the 15 month period since implemen-
tation of the multiplex PCR assay is shown in Table 2. The rate of
D. fragilis detection increased from 2.1% by microscopy to 15.3%
with PCR; this represents the most significant finding, accounting
for 78% of all parasites detected by this test.
Discussion
The advent of PCRhas definitely become a ‘boon’ for our laboratory
in providing a rapid, less labour-intensive method for detecting the
threepathogenicprotozoaG. intestinalis,Cryptosporidium species
and E. histolytica. PCR can detect low numbers easily missed by
microscopy and also candifferentiate pathogenicE. histolytica from
morphologically similar, non-pathogenic Enta-moeba dispar/
moshkovskii5. During the initial validation period, 10 Entamoeba
species identified by microscopy were tested by PCR and only two
of these samples were positive for E. histolytica. The ability to
differentiate betweenpathogenic E. histolytica andnon-pathogenic
Entamoeba species is clinically significant. Results concur with a
prevalence study of Entamoeba in immigrants in Spain that showed
that the majority of cyst-positive samples detected by microscopy
were E. dispar and do not require treatment5.
On the other hand, PCR seemed to become a ‘bane’ by detecting a
large number of D. fragilis in asymptomatic people, especially
children6,7. Numerous parents of ‘well’ children get anxious about
this so called ‘bad bug that does not go away’. Many of these
‘worried well’ get treated with various antiparasitic drugs and
combinations (some of which are known to imbalance the normal
gut flora). They ‘doctor shop’ and are being referred to specialists
such as paediatricians, gastroenterologists and infectious diseases
physicians (communications with General Practitioners and
Australia and New Zealand Paediatric Infectious Diseases Group).
The reliance on morphological diagnosis by microscopy of fixed
stained-smears and the recognition that parasite shedding may be
intermittent, suggests that prior to PCR diagnosis, the true preva-
lence of this protozoon has been underestimated. A review of
D. fragilis carriage has shown that prevalence ranges from 0.2% to
82% in numerous geographical settings, utilising a variety of detec-
tion methods8. In this review, the authors conclude that in
‘symptomatic patients who harbor Dientamoeba and no other
pathogen, it should be considered as the etiological agent and
treated’. However, they also mention that evidence is only circum-
stantial. There has been debate about the clinical significance of
D. fragilisgivensuchwidevariation inprevalenceestimates coupled
with theobservation that theprotozoamaybe found in symptomatic
and asymptomatic individuals and variable responses to antipara-
sitic drugs. A recent case-controlled study by Krogsgaard et al.6
demonstrated that D. fragilis and Blastocystis were detected in a
greater proportion of faecal samples from the asymptomatic back-
ground population in Denmark than from subjects with irritable
bowel syndrome. Röser et al.7 presented data from four years of
routine PCR testing and found that from a total of 22,484 samples,
43% were positive for D. fragilis. With the introduction of PCR
into routine testing within our laboratory, our data suggest that
the prevalence of asymptomatic D. fragilis carriage is higher than
previously reported.
Of all D. fragilis detected in our study period, 40% of faeces were
formed, 58% were semi-formed or unformed and only 2% were
loose/liquid as observed in the laboratory. The clinical notes pro-
vided on pathology request forms ranged from diarrhoea to vague
0
20
40
60
80
100
120
140
160
180
Quarter 2 – 2013: Pre PCR Quarter 2 – 2014: Post PCR
Tot
al p
ositi
ves
G. intestinalis
Cryptosporidium spp.
E. histolytica
D. fragilis
Figure 1. Three months comparison of positive parasite cases pre andpost implementation of protozoa PCR.
Table 2. Protozoal infections detected by multiplex PCR February2014 to May 2015.
Total positive(percentage)
% of allpositives
D. fragilis 828 (15.3%) 78.0
G. intestinalis 183 (3.4%) 17.2
Cryptosporidiumspp.
34 (0.6%) 3.2
E. histolyticaA 13 (0.24%) 1.22
Total positives 1062 (19.7%) 100
Total tested 5384 –
AAdditional four liver aspirates were positive by PCR.
LabReport
MICROBIOLOGY AUSTRALIA * MARCH 2016 47
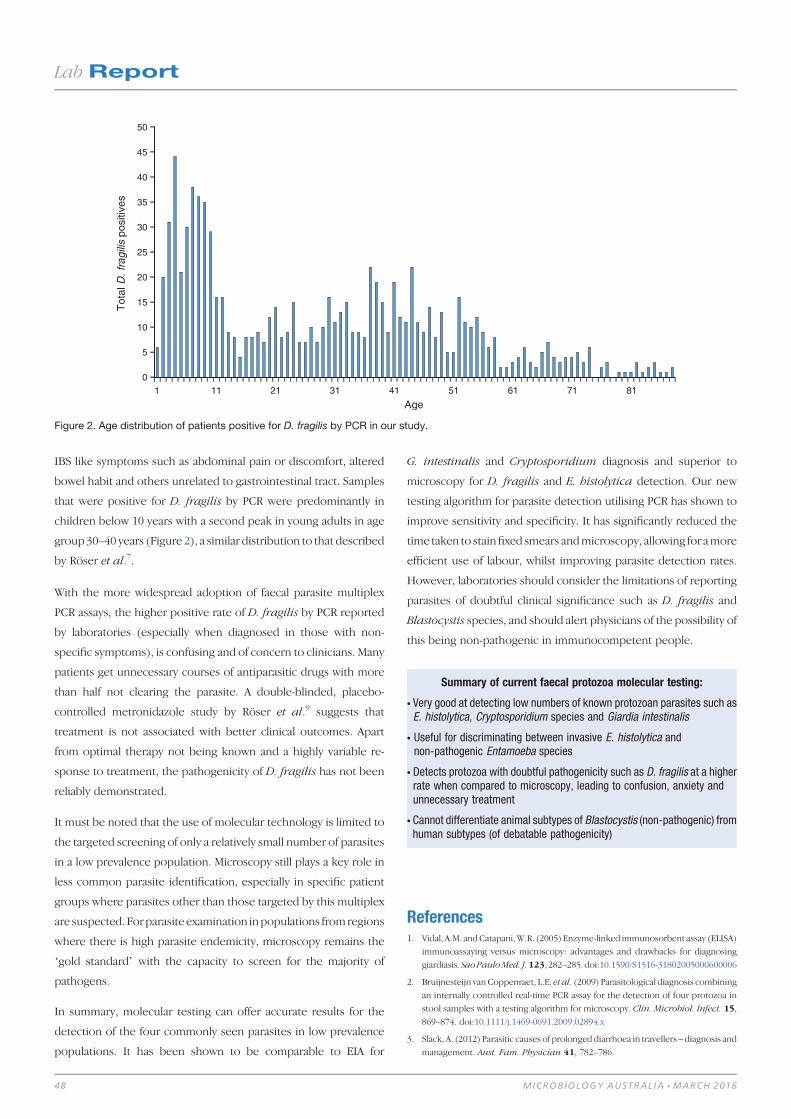
IBS like symptoms such as abdominal pain or discomfort, altered
bowel habit and others unrelated to gastrointestinal tract. Samples
that were positive for D. fragilis by PCR were predominantly in
children below 10 years with a second peak in young adults in age
group 30–40 years (Figure 2), a similar distribution to that described
by Röser et al.7.
With the more widespread adoption of faecal parasite multiplex
PCR assays, the higher positive rate of D. fragilis by PCR reported
by laboratories (especially when diagnosed in those with non-
specific symptoms), is confusing and of concern to clinicians. Many
patients get unnecessary courses of antiparasitic drugs with more
than half not clearing the parasite. A double-blinded, placebo-
controlled metronidazole study by Röser et al.9 suggests that
treatment is not associated with better clinical outcomes. Apart
from optimal therapy not being known and a highly variable re-
sponse to treatment, the pathogenicity of D. fragilis has not been
reliably demonstrated.
It must be noted that the use of molecular technology is limited to
the targeted screening of only a relatively small number of parasites
in a low prevalence population. Microscopy still plays a key role in
less common parasite identification, especially in specific patient
groups where parasites other than those targeted by this multiplex
are suspected. Forparasite examination inpopulations fromregions
where there is high parasite endemicity, microscopy remains the
‘gold standard’ with the capacity to screen for the majority of
pathogens.
In summary, molecular testing can offer accurate results for the
detection of the four commonly seen parasites in low prevalence
populations. It has been shown to be comparable to EIA for
G. intestinalis and Cryptosporidium diagnosis and superior to
microscopy for D. fragilis and E. histolytica detection. Our new
testing algorithm for parasite detection utilising PCR has shown to
improve sensitivity and specificity. It has significantly reduced the
time taken to stainfixed smears andmicroscopy, allowing for amore
efficient use of labour, whilst improving parasite detection rates.
However, laboratories should consider the limitations of reporting
parasites of doubtful clinical significance such as D. fragilis and
Blastocystis species, and should alert physicians of the possibility of
this being non-pathogenic in immunocompetent people.
References1. Vidal, A.M. andCatapani,W.R. (2005) Enzyme-linked immunosorbent assay (ELISA)
immunoassaying versus microscopy: advantages and drawbacks for diagnosing
giardiasis. SaoPauloMed. J.123, 282–285.doi:10.1590/S1516-31802005000600006
2. Bruijnesteijn van Coppenraet, L.E. et al. (2009) Parasitological diagnosis combining
an internally controlled real-time PCR assay for the detection of four protozoa in
stool samples with a testing algorithm for microscopy. Clin. Microbiol. Infect. 15,
869–874. doi:10.1111/j.1469-0691.2009.02894.x
3. Slack, A. (2012) Parasitic causes of prolonged diarrhoea in travellers – diagnosis and
management. Aust. Fam. Physician 41, 782–786.
0
5
10
15
20
25
30
35
40
45
50
1 11 21 31 41 51 61 71 81
Tot
al D
. fra
gilis
pos
itive
s
Age
Figure 2. Age distribution of patients positive for D. fragilis by PCR in our study.
Summary of current faecal protozoa molecular testing:
* Very good at detecting low numbers of known protozoan parasites such asE. histolytica, Cryptosporidium species and Giardia intestinalis
* Useful for discriminating between invasive E. histolytica andnon-pathogenic Entamoeba species
* Detects protozoa with doubtful pathogenicity such as D. fragilis at a higherrate when compared to microscopy, leading to confusion, anxiety andunnecessary treatment
* Cannot differentiate animal subtypes of Blastocystis (non-pathogenic) fromhuman subtypes (of debatable pathogenicity)
LabReport
48 MICROBIOLOGY AUSTRALIA * MARCH 2016

4. Stensvold, C.R. (2013) Blastocystis: genetic diversity and molecular methods for
diagnosis and epidemiology. Trop. Parasitol. 3, 26–34. doi:10.4103/2229-5070.
113896
5. Gutiérrez-Cisneros, M.J. et al. (2010) Application of real-time PCR for the differ-
entiation of Entamoeba histolytica and E. dispar in cyst-positive faecal samples
from 130 immigrants living in Spain. Ann. Trop. Med. Parasitol. 104, 145–149.
doi:10.1179/136485910X12607012373759
6. Krogsgaard, L.R. et al. (2015) The prevalence of intestinal parasites is not greater
among individuals with irritable bowel syndrome: a population-based case-control
study. Clin. Gastroenterol. Hepatol. 13, 507–513.
7. Röser, D. et al. (2013) Dientamoeba fragilis in Denmark: epidemiological expe-
rience derived from four years of routine real-time PCR. Eur. J. Clin. Microbiol.
Infect. Dis. 32, 1303–1310. doi:10.1007/s10096-013-1880-2
8. Barratt, J.L. et al. (2011) A review of Dientamoeba fragilis carriage in humans:
several reasons why this organism should be considered in the diagnosis of
gastrointestinal illness. Gut Microbes 2, 3–12. doi:10.4161/gmic.2.1.14755
9. Röser, D. et al. (2014) Metronidazole therapy for treating dientamoebiasis in
children is not associated with better clinical outcomes: a randomized, double-
blinded and placebo-controlled clinical trial. Clin. Infect. Dis. 58, 1692–1699.
doi:10.1093/cid/ciu188
Biographies
Colin Pham, is a Medical Scientist with over 10 years’ experience
in bacteriology, serology and molecular testing. He underwent his
training at St Vincent’s Melbourne in 2003, and started his career
in Microbiology at the Monash Medical Centre. Since re-joining
St Vincent’s, he has specialised in molecular testing and emerging
molecular technology. He completed a Masters of Applied Science
(Medical Science) at RMIT in 2014, with his thesis titled ‘Impact
of molecular testing on the laboratory diagnosis of Giardia
intestinalis, Cryptosporidium parvum, Dientamoeba fragilis and
Entamoeba histolytica from clinical stool samples’.
Dr Harsha Sheorey, is a Clinical Microbiologist at St Vincent’s
Hospital in Melbourne. His special interest is in clinical parasitology
and tropical medicine and is the author of Clinical Parasitology –
a handbook for medical practitioners and microbiologists: a
second edition has recently been published. He is an invited writer
for the Nematodes chapter in ASMManual of Clinical Microbiology.
He coordinates the Victorian branch of Parasitology & Tropical
Medicine SIG and is actively involved in organisation of the Austra-
lian Parasitology and Tropical Medicine Master Class.
LabReport
MICROBIOLOGY AUSTRALIA * MARCH 2016 49

FT020 Water Microbiology Australian StandardsCommitteeRobin Woodward
Email: [email protected]
I am the Australian Society of Microbiology representative on the
FT020 Water Microbiology Australian Standards Committee. This
committee meets when there are Australian Standards (AS) that are
up for review (approximately every 5 years) or when a new Standard
has been proposed and requires work. Before I became a member
on this committee I thought AS were put together by an elite group
of people and that was all they did. The Committee is made up of
elite members, (of course that goes without saying �) but we are
peoplewhowork in the industry andmost stillwork in the laboratory
or have at some time in our career. There is the Standards Australia
secretarywho ensures themeeting and Standards follow the correct
protocols, the Chair who runs the meeting, a NATA representative
and members comprising people like myself who are in a supervi-
sory role but still perform bench work in the real world. I still do
bench work and that is why I enjoy the challenge of being on
the Committee. It entails reading all the Standards, assessing their
relevance and at meetings discussing any issues that may arise from
the testing side. These issues can be industry driven changes,
changes to the type of testing required, the introduction and
availability of new methods or consumables, feedback from users
of the Standard or personal experience when using the method
in the laboratory. The Standards are reviewed and it is then the
decisionmust bemadeonwhat todowith theStandard. They canbe
reconfirmed (no changes), withdrawn or revised (the Committee
evaluates and revises the Standard).
When a Standard is released for public comment it is crucial that
the users of the Standard read it and make any relevant comments.
All of these comments are documented and must be discussed by
the committee at a meeting. The discussion outcome is also docu-
mented and any changes required are made and eventually the
Standard is printed. It is not an easy process. Writing an unambig-
uousStandard takes timeanda lot of discussion. Thinkback to those
tasks you had at TAFE or university when you had to write instruc-
tions on how to cook a piece of toast. It is often the little things that
can be overlooked, the things you may take for granted. There are
certain guidelines that have to be followed and are set out by
Standards Australia. The method steps are there to follow so that
everyone using the Standard is doing the same thing therefore
creating a uniform procedure that should give the same results no
matter where you are. Standard procedures are important and it is
the basis of the quality systems in laboratories.
So please comment if you feel there is something you need to bring
up when a Standard goes out for Public Comment. Even if it just a
typographical error or something you think is wrong-COMMENT.
Your company can subscribe to StandardsWatch so you are alerted
when a Standard relevant to your area is released or open for public
comment.
Thank you to the ASM and my employer (ALS) for supporting me
to be a committee member on FT020 Water Microbiology. It is
appreciated and it is only through organisations like the ASM
and generous employers that Standards Australia committees can
operate. All committee members are volunteers and must have
the relevant qualifications and knowledge to ensure that these
Standards can continue to be developed and maintained.
2016 ASM Communication Ambassador ProgramThe ASM is looking for teams of student and ECR members to
volunteer as part of a roster of Communication Ambassadors within
the society, who will:
(1) Be in charge of ASM’s online communication channels togetherwith a team of ASMmembers, bolstering the society’s networksby sharing your story with scientists, industry, media membersand policy makers;
(2) Effectively represent the society by using these channels toshowcase yourwork, reveal personal insights intoyour scientificcareer, and serve as a public advocate for microbiology.
We are looking for enthusiastic members who are keen to contrib-
ute. To apply for the 2016 program, please submit an Expression of
Interest (maximum 200 words) to [email protected] by 31
March 2016 and highlight the following:
* Your name, institution, qualifications, area of interest withinmicrobiology, and ASM membership status;
* Your strategy for broadening ASM’s online presence and collab-orating with other communication ambassadors;
* What conferences, scientific meetings, or events you can providesocial media coverage for in 2016.
Successful applicants for the 2016 Communication Ambassador
program will receive a $100 discount for registration to the 2016
ASM Annual Scientific Meeting (3–6 July 2016, Perth Convention &
Exhibition Centre).
ASMAffairs
50 10.1071/MA16016 MICROBIOLOGY AUSTRALIA * MARCH 2016

Stopping dengue: recent advances and newchallengesGayathri Manokaran, Kirsty McPherson and Cameron P Simmons
Symposium 16 November 2015
Peter Doherty Institute for Infection and Immunity,
Melbourne
Dengue remains a major problem throughout the world with an
estimated 30% of the world’s population at risk of infection. In the
past few years, major advances have been made across virology,
clinical insights, vaccines and mosquito control strategies. The first
of its kind to be ever held in Australia, the International Dengue
Symposium 2015 showcased some of the best science and clinical/
public health research being undertaken in the field.
Proudly organised by The Peter Doherty Institute for Infection and
Immunity, under the leadership of Professor Cameron Simmons,
this symposium brought together 130 national and international
dengue researchers. The organising committee acknowledges the
support of ASM as an important partner to this meeting, together
with generous support from The University of Melbourne, The
Royal Melbourne Hospital, the Oxford University Clinical Research
Unit, Vietnam and the DUKE-NUS Graduate Medical School,
Singapore. Industry sponsors included Sanofi Pasteur, BioTools,
VWR International, In Vitro Technologies, Pacific Lab Products,
Interpath Services and Alere Global.
The formal symposium was opened by Professor Cameron
Simmons. This was followed by a series of 21 brief talks spanning
across four main themes of dengue research: Virology, Public
Health, Clinical Research and Immunology/Pathogenesis. Short
breaks for networking were scheduled in between each of these
four sessions.
All of the invited speakers had considerable international profiles in
dengue and hence the quality of the presentations was very high.
Associate Professor Sheemei Lok discussed the structural basis
behind how a dengue-2-specific human monoclonal antibody
effectively protects mice from dengue infection due to its ability
to ‘lock’ the dengue envelope proteins while blocking the binding
of enhancing antibodies. Following that, Professor Ooi Eng Eong
described the novel finding of a viral RNA-host protein interaction
that allows dengue virus to evade the host immune response- this
work was recently published in the prestigious journal, Science.
Ahighlight of this symposiumwas the ‘PublicHealth’ session,which
was the first of its kind in Australia, with speakers from various
companies includingSanofiPasteur andTakeda, coming together to
discuss their respective dengue vaccines, thereby providing atten-
dees with a useful summary of all the dengue vaccines currently in
clinical development.
Novel and highly innovative medical technologies were next dis-
cussed in the ‘Clinical Research’ session, with Dr Jenny Low
explaining how in vivo imaging technologies such as positron
emission tomography are being optimised presently to track den-
gue virus progression in a mouse model. Dr Sophie Yacoub gave a
summary on changes in microvasculature of dengue patients upon
infection and how these disturbances might precede the develop-
ment of severe disease.
In the next session, Dr Laura Rivino discussed their work on
elucidating the human cellular responses during dengue infection
and how both CD4+ and CD8+ T-cell subsets remain enriched in
the skin of dengue patients up to 5months post infection. Professor
Cameron Simmons then described an interesting dengue neutra-
lisation assay developed in Vietnam using patients’ viraemic blood
and infected mosquitoes as a readout of the efficacy of monoclonal
antibodies to efficiently neutralise dengue virus. Next, Dr David
Muller introduced the concept of nanopatches that allow vaccine
delivery in a needle-free manner.
In closing, the sponsors were wholeheartedly acknowledged for
their contributions by Professor Cameron Simmons and their
support was instrumental in allowing this symposium to be a
success. Not only was the stellar line-up of talks highly educational
but importantly, this symposium provided a networking opportu-
nity for everyone interested in meeting the challenge of reducing
dengue burden throughout the world.
ASMAffairs
MICROBIOLOGY AUSTRALIA * MARCH 2016 10.1071/MA16017 51


Confirmed Plenary speakers
Annual Scientific Meeting and Trade Exhibition
www.theasm.org.au www.westernaustralia.theasm.org.au
Professor Peter Hawkey University of Birmingham Nosocomial infection control and antibiotic resistance
Professor Dan Andersson Upsalla University Environmental pollution by antibiotics and its role in the evolution of resistance
Assoc Prof Susan Lynch University of California San Francisco Colitis, Crohn's Disease and Microbiome Research
Professor Anna Durbin Johns Hopkins Dengue and vaccines
Dr Brian Conlon Northeastern University, Boston Drug discovery in soil bacteria
Watch this space for more details on the scientific and social program, speakers, ASM Public Lecture, workshops, ASM awards, student events, travel awards, abstract deadlines and much more..
Perth, WA A vibrant and beautiful city located on the banks of the
majestic Swan river. Come stay with us in WA and experience our world class wineries and restaurants, stunning national parks, beaches and much more..
NOW CONFIRMED! 2016 Rubbo Oration Professor Anne Kelso CEO NHMRC
As with previous years, ASM 2016 will be co-run with EduCon 2016: Microbiology Educators’ Conference

© 2015 Hologic, Inc. All rights reserved. Hologic, Science of Sure, Aptima, Panther and associated logos are trademarks and/or registered trademarks of Hologic, Inc. and/or its subsidiaries in the United States and/or other countries. This information is intended for medical professionals and other markets and is not intended as a product solicitation or promotion where such activities are prohibited. Because Hologic materials are distributed through websites, eBroadcasts and tradeshows, it is not always possible to control where such materials appear. For specific information on what products are available for sale in a particular country, please contact your local Hologic representative or write to [email protected]
Gen-Probe Australia Pty Ltd. ABN 91 150 687 773 . Suite 402, Level 4, 2-4 Lyon Park Road, MacquariePark, NSW, Australia, 2113 Phone: +61 2 9888 8000 Fax: (02) 9870 7555
hologic.com | [email protected] | 1800 264 073 (Australia) | [email protected] | 0800 804 904 (New Zealand)
Aptima Combo 2® assay | Aptima CT assay | Aptima GC assay | Aptima Trichomonas vaginalis assay
Aptima HPV assay | Aptima HPV 16 18/45 Genotyping assay
Aptima HIV-1 Quant Dx Assay
The highly evolvedsenses of a hunter
With continuous, random access
and no-batch processing the
Panther® system, along with our
highly sensitive and specific
Aptima® assays, provide
on-demand performance for a
wide range of infectious diseases.
aasa say | Aptima CT assay | Aptima GC assay | Aptima Trichomonas vaginalis assay
AAppttiima HPV assay | Aptima HPV assay | Aptima HPV 16 18/45 Gma HPV 16 18/45 Genotyping assayenotyping assay
Aptima HIV-1 Quant Dx Assay
Copyright © 2022 FDOKUMEN




















