
Traditional and geometric morphometrics for studying skull ...
12
Traditional and geometric morphometrics for studying skull morphology during growth in Mastomys natalensis (Rodentia: Muridae) MATTEO BRENO,* HERWIG LEIRS, AND STEFAN VAN DONGEN Evolutionary Ecology Group, Department of Biology, University of Antwerp, Groenenborgerlaan 171, B-2020 Antwerp, Belgium (MB, HL, SVD) Danish Pest Infestation Laboratory, Department of Integrated Pest Management, Faculty of Agricultural Sciences, University of Aarhus, Skovbrynet 14, DK-2800 Lyngby, Denmark (HL) * Correspondent: [email protected] Geometric morphometrics is a powerful tool for the study of morphological variation that possesses numerous advantages over the more traditional approach based on linear measurements. We analyzed skull morphology, comparing traditional with geometric morphometrics, of 3 different developmental pathways in Mastomys natalensis (Rodentia: Muridae) from a single population. During early development growth patterns were influenced by environmental factors, specifically rainfall pattern, consistent with previous reports that growth trajectories vary according to the amount and distribution of rain. Results confirmed that early growth rate is one of the main determinants of size and shape differences in the skull in the 3 developmental pathways (generation types) of M. natalensis. Other factors, such as food quality and consistency, also could play an important role. Overall, geometric morphometrics appeared more sensitive than the traditional method in detecting variation in skull morphology, but both approaches led to very comparable conclusions. Phenotypic plasticity is an alternative explanation to local adaptations for ecogeographical morphological variation. Key words: geometric morphometrics, growth, phenotypic plasticity, skull morphology, traditional morphometrics E 2011 American Society of Mammalogists DOI: 10.1644/10-MAMM-A-331.1 Morphometrics, the quantitative approach to the study of morphological variation, combines tools for the description and the statistical analysis of many relevant biological questions (Rohlf 1990). It is used in a broad range of disciplines from paleontology to quantitative genetics. Com- monly, the field of morphometrics is divided into traditional or multivariate morphometrics and geometric morphometrics (Rohlf and Marcus 1993). The former term refers to the application of multivariate statistics to linear measurements and ratios, whereas the latter concerns the development of coordinate-based methods begun in the 1970s (Bookstein 1991; Dryden and Mardia 1998; Zelditch et al. 2004). Traditional morphometrics has been shown to have certain limitations when compared to geometric morphometrics. In the former, data consist of lengths, depths, and widths containing little information about the geometry of the structure being studied (Zelditch et al. 2004). Geometric relationships among the variables are not retained so that it is not possible to depict graphical representations of shape (Adams et al. 2004). In addition, overlap of variables occurs frequently, making it difficult to describe local shape changes. Within the traditional framework a formal distinction between size and shape is difficult, and little agreement is found among the many methods of size correction proposed (Klingenberg 1996, 1998; Zelditch et al. 2004). To overcome these limitations a new method was developed starting in the 1970s. The results of the morphometrics synthesis (Bookstein 1996) led to what is now known as the morphometrics revolution (Rohlf and Marcus 1993). Geometric morphomet- rics methods capture the geometry of an object using coordinate-based data. A statistical theory of shape has been developed allowing ‘‘… the combined use of multivariate statistical methods and methods for the direct visualization in biological form’’ (Adams et al. 2004:6). Direct and indirect comparisons between traditional and geometric morphometrics have been few but generally have shown the advantages of geometric morphometrics compared to the traditional approach. For example, Maderbacher et al. (2008) reported that both traditional and geometric morpho- metrics were able to discriminate among 3 populations of the www.mammalogy.org Journal of Mammalogy, 92(6):1395–1406, 2011 1395 Downloaded from https://academic.oup.com/jmammal/article/92/6/1395/961102 by guest on 11 September 2022
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Traditional and geometric morphometrics for studying skull ...

Traditional and geometric morphometrics for studying skullmorphology during growth in Mastomys natalensis (Rodentia: Muridae)
MATTEO BRENO,* HERWIG LEIRS, AND STEFAN VAN DONGEN
Evolutionary Ecology Group, Department of Biology, University of Antwerp, Groenenborgerlaan 171, B-2020 Antwerp,
Belgium (MB, HL, SVD)
Danish Pest Infestation Laboratory, Department of Integrated Pest Management, Faculty of Agricultural Sciences,
University of Aarhus, Skovbrynet 14, DK-2800 Lyngby, Denmark (HL)
* Correspondent: [email protected]
Geometric morphometrics is a powerful tool for the study of morphological variation that possesses numerous
advantages over the more traditional approach based on linear measurements. We analyzed skull morphology,
comparing traditional with geometric morphometrics, of 3 different developmental pathways in Mastomys
natalensis (Rodentia: Muridae) from a single population. During early development growth patterns were
influenced by environmental factors, specifically rainfall pattern, consistent with previous reports that growth
trajectories vary according to the amount and distribution of rain. Results confirmed that early growth rate is
one of the main determinants of size and shape differences in the skull in the 3 developmental pathways
(generation types) of M. natalensis. Other factors, such as food quality and consistency, also could play an
important role. Overall, geometric morphometrics appeared more sensitive than the traditional method in
detecting variation in skull morphology, but both approaches led to very comparable conclusions. Phenotypic
plasticity is an alternative explanation to local adaptations for ecogeographical morphological variation.
Key words: geometric morphometrics, growth, phenotypic plasticity, skull morphology, traditional morphometrics
E 2011 American Society of Mammalogists
DOI: 10.1644/10-MAMM-A-331.1
Morphometrics, the quantitative approach to the study of
morphological variation, combines tools for the description
and the statistical analysis of many relevant biological
questions (Rohlf 1990). It is used in a broad range of
disciplines from paleontology to quantitative genetics. Com-
monly, the field of morphometrics is divided into traditional
or multivariate morphometrics and geometric morphometrics
(Rohlf and Marcus 1993). The former term refers to the
application of multivariate statistics to linear measurements
and ratios, whereas the latter concerns the development of
coordinate-based methods begun in the 1970s (Bookstein
1991; Dryden and Mardia 1998; Zelditch et al. 2004).
Traditional morphometrics has been shown to have certain
limitations when compared to geometric morphometrics. In
the former, data consist of lengths, depths, and widths
containing little information about the geometry of the
structure being studied (Zelditch et al. 2004). Geometric
relationships among the variables are not retained so that it is
not possible to depict graphical representations of shape
(Adams et al. 2004). In addition, overlap of variables occurs
frequently, making it difficult to describe local shape changes.
Within the traditional framework a formal distinction between
size and shape is difficult, and little agreement is found among
the many methods of size correction proposed (Klingenberg
1996, 1998; Zelditch et al. 2004). To overcome these
limitations a new method was developed starting in the
1970s. The results of the morphometrics synthesis (Bookstein
1996) led to what is now known as the morphometrics
revolution (Rohlf and Marcus 1993). Geometric morphomet-
rics methods capture the geometry of an object using
coordinate-based data. A statistical theory of shape has been
developed allowing ‘‘… the combined use of multivariate
statistical methods and methods for the direct visualization in
biological form’’ (Adams et al. 2004:6).
Direct and indirect comparisons between traditional and
geometric morphometrics have been few but generally have
shown the advantages of geometric morphometrics compared
to the traditional approach. For example, Maderbacher et al.
(2008) reported that both traditional and geometric morpho-
metrics were able to discriminate among 3 populations of the
w w w . m a m m a l o g y . o r g
Journal of Mammalogy, 92(6):1395–1406, 2011
1395
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022

cichlid fish Tropheus moorii, but a landmarks approach
permitted a far better quantification and visualization of
among-population differences. Abdel-Rahman et al. (2009)
found a statistically significant sexually dimorphism in
the skull of the African Nile rat (Arvicanthis niloticus) that
was not detected in previous studies based on traditional
morphometrics, and Bernal (2007) showed that a considerable
amount of information was added by using landmarks-based
methods in a study of variation in molar shape in humans.
Finally, Blanco and Godfrey (2006) used geometric morpho-
metrics to show that ontogenetic scaling is insufficient to
describe sexual dimorphism in the mantled howling monkey
(Alouatta palliata).
The aim of this work is to explore within-population
morphological pattern in the skull of Mastomys natalensis
using both traditional and geometric morphometrics. Tradi-
tional and geometric approaches are applied to data collected
in the field with a 2-fold purpose: a nontechnical comparison
between multivariate and geometric morphometrics and the
study of morphological variation in relation to environmental
conditions. In a recent paper Fadda and Leirs (2009) showed
that environmental conditions affect growth trajectories of the
skull within a population of M. natalensis in Tanzania. The
peculiarity of this population is that it presents different
growth patterns related to the amount of rainfall. Three
different generation types (developmental pathways) have
been identified according to the presence and the length of a
period of reduced growth (Leirs et al. 1993). Generation a is
born in the middle of the year (main breeding season) and
shows the longest period of reduced growth, reaching the
maximum size in the next breeding season. Alternatively,
generation b is born in the main breeding season as well, but
due to abundant off-season rain (October–December) it has a
shorter period of reduced growth and is able to mature at the
very beginning of the next year. Its offspring, generation c,
appear in the off-season period (January–March) and reach
maturity in the main breeding season of that same year. A
growth stop phase has been identified as a major determinant
of skull shape, and its duration relates to the achievement of
target skull morphology (Fadda and Leirs 2009).
Among the many studies about within-species variation
in the morphology of the skull, most are concerned with
ecogeographical variation (Lawing and Polly 2009), whereas
studies investigating the direct effect of environment (mainly,
quality and consistency of food) on shape often are performed
in the laboratory (Kiliaridis 2006). For example, effects of
habitat on skull morphology in M. natalensis have been
reported, where 2 populations existing in different environ-
ments differed significantly in morphology (Lalis et al.
2009a). In studies like the latter it often is assumed that the
observed ecogeographical variation in morphology represents
a local adaptation and not differential reaction to different
environments (i.e., plasticity). However, laboratory studies have
shown an important role of plasticity (Myers et al. 1996).
Because we studied a single genetically homogeneous population
(Van Hooft et al. 2008) under varying developmental conditions,
we have the opportunity to study phenotypic plasticity in the
field.
MATERIALS AND METHODS
The data we analyzed come from the same trapping
campaign as in Fadda and Leirs (2009). However, we
considered only a subset of individuals for which a precise
age estimation was available. For this subset landmark data of
the skull were collected in addition to linear measurements.
Specimens were collected in Morogoro, Tanzania, between
September 1987 and October 1988. Animals were aged using
eye lens weight (Leirs 1995) and then assigned to generation
types and age classes according to their estimated date of birth
and age. Three 2-month age classes (0–2, 2–4, and 4–6 months)
were established, but a individuals belonging to the 3rd age
class were not available. In total, 856 skulls were used, of which
111, 563, and 182 were assigned to generation types a, b, and c,
respectively. Division into age classes resulted in the assign-
ment of 233 skulls to the 1st age class, 290 to the 2nd age class,
and 333 to the 3rd age class. Dermestid beetles were used to
skeletonize skulls, after which 22 cranial linear parameters were
measured to the nearest 0.05 mm with digital calipers (Fig. 1).
A subset of the original data set was investigated by using
geometric morphometrics; 292 (62 of generation a, 125
of generation b, and 105 of generation c) specimens were
photographed in dorsal view, and 23 landmarks (Fig. 1) were
registered twice using ImageJ (Rasband 2010). Numbers of
specimens were 95, 126, and 71 for the 1st, 2nd, and 3rd age
classes, respectively. Other studies have applied geometric
morphometrics to the study of skull morphology in M.
natalensis (Lalis et al. 2009a, 2009b); however, it was not
possible to locate exactly the same landmarks on the dorsal
side of the skull. All analyses were performed using MorphoJ
(Klingenberg 2011) and R (R Development Core Team 2009).
Analysis of linear measurements.—Univariate analysis of
variance and multivariate analysis of variance (MANOVA)
were carried out to identify sources of morphological variation
between or among generation types, age classes, and sex. Data
were graphically inspected for outliers and normality by QQ
plots. Proportional contributions of the sum of squares
(%SSQ) of each source of variation to the total SSQ were
reported (Bronner et al. 2007; Leamy 1983) for each variable.
In comparison to morphological differences observed between
age classes (Pillai’s trace 5 0.69, F44,1,646 5 19.80, P ,
0.001) and generation types (Pillai’s trace 5 0.77, F44,1,646 5
23.48, P , 0.001), sexual dimorphism was relatively small but
significant (Pillai’s trace 5 0.124, F22,822 5 5.30, P , 0.001).
The %SSQ identified age and generation type as the 2
major sources of variation for almost all variables (variation
explained by age class and generation type ranged from 1.6%
to 75% and 0.8% to 6%, respectively). For sexual dimorphism,
however, this range was between 0.1% and 1.2%. Because no
interactions with sex were observed, males and females were
pooled for all further analyses (sex*age class: Pillai’s trace 5
0.07, F44,1,646 5 1.45, P 5 0.03; sex*generation type: Pillai’s
1396 JOURNAL OF MAMMALOGY Vol. 92, No. 6
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022

trace 5 0.06, F44,1,646 5 1.06, P 5 0.36). A significant
interaction was observed between age class and generation type
(Pillai’s trace 5 0.15, F66,2,472 5 1.94, P , 0.001), indicating
that morphological differences between generation types
changed with age.
Next, we performed a principal component analysis (PCA)
on untransformed data based on the correlation matrix for
each generation type separately. Age-related changes in size
were visualized by plotting the 1st principal component
(PC1) against age. This approach is common in traditional
FIG. 1.—Linear measurements made on skulls of Mastomys natalensis. 1, Greatest length of the skull (GL); 2, condylonasal length (CL);
3, henselion–basion distance (HB); 4, henselion–palation distance (HP); 5, length of palatal foramen (PL); 6, length of diastema (DL); 7,
distance alveolus M1–upper incisor (MoID); 8, smallest interorbital breadth (IB); 9, zygomatic breadth (BB); 10, smallest palatal breadth (PB);
11, length of upper cheek-teeth alveoli (UAL); 12, breadth of upper dental arch (UDB); 13, greatest breadth of M1 (Mo1); 14, breadth of
zygomatic plate (ZPB); 15, greatest breadth of nasals (NB); 16, greatest length of nasals (NL); 17, distance M1–M3 (MoJ); 18, length of bulla
(TBL); 19, greatest breadth of braincase (BrB); 20, rostrum height (RH); 21, greatest rostrum breadth (RB); 22, greatest jaw height (JH). Open
black squares represent landmarks.
December 2011 BRENO ET AL.—TRADITIONAL AND GEOMETRIC MORPHOMETRICS 1397
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022

morphometrics, and PC1 usually is interpreted as a size com-
ponent if all of the coefficients of the 1st eigenvector have the
same sign and similar magnitude (Bever 2008; Krystufek
2002; Smith 1998; but see Sundberg 1989 for a discussion of
the topic). PC2 and PC3 were inspected by plotting them
against PC1 to explore differences between age classes and
generation types not associated with the main vector of
variation (i.e., PC1). To further explore differences in shape
between the 3 generation types a discriminant analysis was
performed for each age class. To visualize the dimensions that
optimally divide generation types within each age class,
canonical variate analysis (CVA) was conducted, and the 1st
and 2nd canonical variates (CVs) were plotted. Mahalanobis
distances were calculated among generation types for each age
class and were compared using permutation tests (10,000
permutations). To compare each trait among different
generation types allometric trajectories of both the c and bgeneration types were described using bivariate and multivar-
iate analysis (described below). Generation a was excluded
from the following analyses because it did not contain any
individual older than 4 months. In the bivariate case overall
size was estimated by means of total skull length. The relation
between each variable and skull length was studied using the
allometry equation: log y 5 log b0 + b1 + log e (derived from
power equation y~b0xb1 e—Abdala et al. 2001; Bever 2008;
Giannini et al. 2004). Deviations from isometry were tested by
constructing confidence intervals of the slopes (b1). Slopes
significantly . 1 indicate fast development relative to total
skull length, and slopes , 1 are indicative of a relative slow
growth. Analysis of covariance was used to test for differences
in allometric patterns between generation types for all traits. A
multivariate generalization of the allometry equation was
proposed by Jolicoeur (1963) as the PC1 from the variance–
covariance matrix of the log-transformed data. In this case
isometry is expressed by the isometric vector 1/n0.5, where
n is the number of variables. Values of the 1st eigenvector
represent the allometric coefficients of each variable. A
bootstrap procedure (with 5,000 resamples) was used to obtain
confidence intervals for each allometric coefficient where
differences in allometric coefficients between generation types
were tested by subtracting the bootstrap values of 1 generation
type from those of the other. If the confidence intervals of
these differences did not include zero, the 2 coefficients were
considered different between generation types.
Analysis of landmark data using geometric morphomet-
rics.—Several geometric methods exist, and among them the
Procrustes method (Goodall 1991) is the most widespread.
Procrustes superimposition involves 3 steps to eliminate all
nonshape variation. First, configurations of landmarks are
translated so that their centroid is placed at the origin of the
coordinate system. Next, they are scaled to a common unit
size. Finally, configurations are rotated around their centroid
to minimize the squared distance between corresponding
landmarks. When .2 configurations exist the process
becomes iterative, and all of the configurations are rotated
repeatedly to fit a temporal consensus. The procedure ends
when 2 subsequent average configurations match. Within- and
between-species variation is one among the many phenom-
ena that can be addressed with a morphometric approach.
Ultimately, Procrustes allows a more straightforward distinc-
tion between shape and size variations. Statistical analyses can
be applied to both sources of variation, and variation in shape
can be visualized easily (Zelditch et al. 2004).
The presence of outliers and the distribution of the data
were inspected graphically by plotting the cumulative
distribution of the squared Mahalanobis distance against a
multivariate normal distribution fitted to the data as imple-
mented by MorphoJ (Klingenberg 2011). Overall shape
variation and variation within age classes were visualized by
PCA of Procrustes superimposed landmark configurations of
the entire data set and for each age class. Relative warps (PC
scores of Procrustes coordinates) extracted from Procrustes
superimposition performed separately for each age class were
used to test for morphological differences among generation
types within age classes. The advantage of using relative
warps is that, in contrast to Procrustes coordinates, they
possess the correct number of dimensions and can be
submitted to further analyses without correcting for degrees
of freedom. However, a modified version of MANOVA for
Procrustes data implemented in MorphoJ (Klingenberg 2011)
and MANOVA of relative warps gave the same results.
Preliminary analyses were conducted on the entire data set
by means of MANOVA of relative warps, with age class,
generation type, sex, and their interactions as effects. Sig-
nificant differences were found for generation type (Pillai’s
trace 5 0.68, F42,520 5 6.49, P , 0.001) and age class (Pillai’s
trace 5 0.75, F42,520 5 7.44, P , 0.001) but not for sex
(Pillai’s trace 5 0.11, F21,259 5 1.56, P 5 0.06). Moreover, an
interaction between age class and generation type was found
(Pillai’s trace 5 0.44, F63,783 5 2.16, P , 0.001), and because
no significant interactions involving sex were present (sex*age
class: Pillai’s trace 5 0.12, F42,520 5 0.82, P 5 0.78;
sex*generation type: Pillai’s trace 5 0.17, F42,520 5 1.14, P 5
0.26), males and females were pooled. CVA then was
performed on Procrustes coordinates for each age class using
generation type as a grouping factor. Mahalanobis distances
and Procrustes distances among generation types were
computed in each age class and tested by a permutation test
(10,000 permutations).
RESULTS
Analysis of linear measurements.—Differences among
generation types were highly significant in each age class
(age class 1: Pillai’s trace 5 0.78, F2,123 5 3.02, P , 0.001;
age class 2: Pillai’s trace 5 0.71, F2,323 5 7.65, P , 0.001;
age class 3: Pillai’s trace 5 0.33, F1,224 5 4.68, P , 0.001).
Univariate analyses of variance revealed a number of
differences in all age classes, especially in the 2nd age class
(Table 1). PC1 explained 73% of the total variance. PC1 could
be interpreted as a size vector because the factor loadings
of this 1st eigenvector all had the same sign and similar
1398 JOURNAL OF MAMMALOGY Vol. 92, No. 6
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022

TA
BL
E1
.—A
nal
ysi
so
fv
aria
nce
tab
leo
fth
eco
mp
aris
on
of
lin
ear
mea
sure
men
ts(X
6S
D)
amo
ng
gen
erat
ion
typ
es(a
lph
a,b
eta,
and
gam
ma)
wit
hin
each
age
clas
s.S
eeF
ig.
1fo
r
def
init
ion
so
fm
easu
rem
ents
.*
P,
0.0
5;
**
P,
0.0
01
.
Ag
ecl
ass
1A
ge
clas
s2
Ag
ecl
ass
3
Alp
ha
Bet
aG
amm
aF
2,1
23
Alp
ha
Bet
aG
amm
aF
2,3
23
Bet
aG
amm
aF
1,2
24
GL
2,5
35
.86
75
.22
,532
.36
11
2.8
2,5
81
.66
95
.53
.57
*2
,608
.66
92
.22
,71
0.2
61
23
.62
,805
.96
12
6.2
38
.27
**
2,8
88
.56
11
5.0
2,9
47
.56
75
.71
0.1
9*
*
CL
2,3
26
.46
91
.02
,341
.76
10
9.0
2,3
59
.26
10
5.9
1.3
42
,419
.46
94
.22
,51
7.4
61
26
.22
,610
.86
12
7.4
34
.90
**
2,6
96
.36
12
1.0
2,7
49
.96
84
.67
.55*
*
HB
1,9
78
.86
84
.71
,983
.06
92
.82
,00
6.6
69
5.2
1.1
62
,064
.26
87
.42
,14
5.5
61
16
.32
,236
.46
11
8.4
34
.88
**
2,3
17
.76
11
2.5
2,3
69
.06
75
.68
.06*
*
HP
1,1
15
.66
44
.81
,127
.06
58
.11
,12
5.2
65
4.8
0.6
41
,161
.86
48
.31
,20
2.6
66
0.4
1,2
43
.96
64
.22
7.4
1*
*1
,286
.66
55
.21
,30
5.3
64
2.8
4.3
6*
PL
60
7.4
63
1.0
59
9.3
62
7.8
61
1.8
63
7.7
0.7
66
36
.56
34
.16
50
.66
33
.86
65
.56
41
.39
.55*
*6
93
.86
34
.96
98
.76
32
.90
.70
DL
63
5.1
63
0.4
64
2.0
63
5.8
64
7.4
63
3.3
1.8
26
69
.76
30
.97
00
.46
41
.37
26
.56
45
.22
6.6
2*
*7
62
.26
40
.27
77
.46
34
.75
.26*
Mo
ID6
65
.86
32
.96
87
.36
37
.86
87
.76
36
.75
.97
**
70
4.0
63
6.0
75
9.6
64
5.7
78
2.7
64
9.9
36
.15
**
82
8.5
64
4.0
84
1.8
63
3.8
3.4
7
IB3
92
.66
16
.44
01
.06
12
.63
94
.16
13
.31
.92
40
0.5
61
5.4
41
3.1
61
5.5
41
3.6
61
4.9
10
.48
**
42
1.3
61
3.7
41
7.2
61
3.2
3.2
0
BB
1,2
21
.26
48
.71
,231
.96
42
.41
,25
0.5
63
9.2
4.7
8*
1,2
47
.06
47
.31
,30
4.4
65
2.7
1,3
37
.16
54
.53
7.5
4*
*1
,381
.26
60
.31
,39
9.6
65
1.3
3.4
1
PB
22
7.2
61
4.5
23
8.0
61
0.5
23
6.7
61
3.1
7.7
4*
*2
36
.66
13
.12
52
.76
14
.12
51
.16
14
.81
8.2
3*
*2
65
.26
16
.22
61
.66
13
.31
.85
UA
L4
80
.66
16
.54
87
.36
13
.94
89
.76
14
.04
.41
*4
82
.46
18
.34
93
.96
18
.05
03
.96
17
.92
0.1
5*
*5
04
.96
19
.75
11
.96
15
.84
.65*
UD
B5
34
.76
15
.85
47
.06
15
.35
49
.86
11
.81
4.0
3*
*5
44
.26
17
.55
65
.06
18
.55
71
.66
18
.32
7.5
9*
*5
89
.06
18
.85
85
.56
14
.11
.32
Mo
11
46
.16
5.7
14
6.3
64
.81
50
.26
5.9
6.1
8*
*1
46
.86
4.1
14
7.8
66
.91
51
.06
5.8
9.8
2*
*1
48
.86
6.3
15
1.6
66
.37
.08*
*
ZP
B2
63
.36
14
.32
66
.36
15
.92
66
.76
17
.50
.65
27
1.1
61
8.2
28
4.6
61
9.3
29
4.6
61
9.8
20
**
30
1.5
61
8.5
31
7.4
61
5.6
27
.30
**
NB
28
6.5
61
5.6
27
5.7
61
2.9
28
3.6
61
2.0
3.5
7*
29
2.9
61
4.0
29
1.4
61
8.7
30
0.2
61
5.6
8.4
7*
*3
10
.86
19
.33
09
.56
16
.30
.15
NL
94
3.6
64
9.9
96
0.0
64
8.4
98
1.2
65
2.0
6.6
7*
*9
88
.06
52
.31
,06
3.2
66
9.8
1,1
01
.76
69
.23
5.3
5*
*1
,155
.26
60
.61
,17
4.9
65
1.6
3.8
7
Mo
J4
28
.66
12
.54
33
.36
8.4
43
7.9
61
1.4
7.3
5*
*4
30
.26
13
.84
35
.96
14
.64
45
.26
17
.01
7.1
28
**
44
3.7
61
7.5
44
8.5
61
5.2
2.6
9
TB
L4
61
.66
19
.34
64
.06
18
.14
73
.46
17
.34
.70
*4
61
.56
18
.94
66
.86
18
.94
81
.06
20
.82
1.1
9*
*4
76
.86
17
.24
84
.76
16
.37
.31*
*
BrB
1,1
20
.96
35
.61
,116
.76
30
.61
,13
7.1
62
8.0
3.2
6*
1,1
36
.66
28
.51
,15
1.8
62
7.9
1,1
72
.56
32
.32
4.7
7*
*1
,183
.16
30
.01
,18
0.3
63
5.8
0.2
8
RH
56
2.5
62
5.5
57
3.7
62
1.6
57
7.3
62
8.2
4.2
5*
58
7.1
62
2.3
61
6.2
63
1.3
64
1.8
63
2.7
43
.25
**
66
6.7
63
3.9
67
0.9
62
7.8
0.5
9
RB
43
1.9
62
3.5
43
9.3
62
1.2
44
7.3
61
6.2
6.0
1*
44
5.3
62
2.5
46
4.2
62
6.0
48
5.1
62
8.9
34
.25
**
49
9.0
62
7.7
51
2.3
62
2.8
8.6
1*
*
JH6
96
.06
44
.77
00
.06
23
.27
00
.86
34
.20
.19
72
3.8
62
8.4
76
4.9
65
2.6
78
6.6
64
9.8
20
.24
**
82
6.4
64
9.5
84
2.9
63
2.6
4.3
3*
December 2011 BRENO ET AL.—TRADITIONAL AND GEOMETRIC MORPHOMETRICS 1399
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022

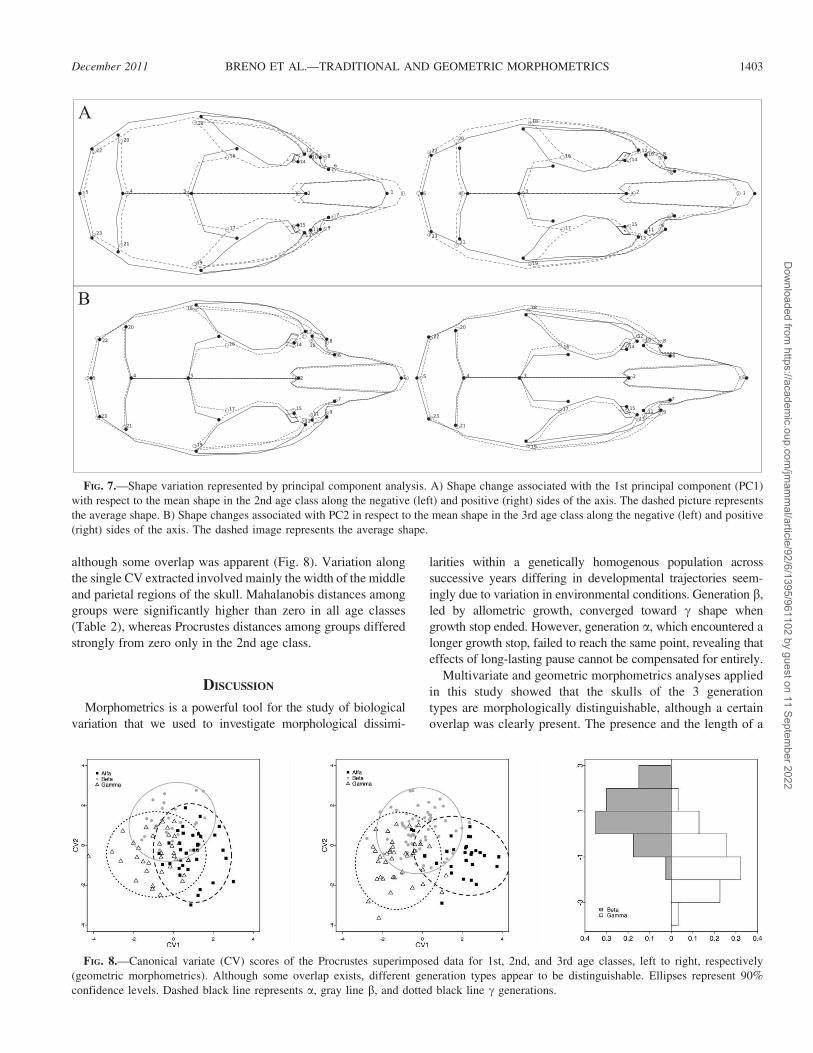
magnitude. Variation represented by PC2 was mainly within
a, and removing the a generation before performing PCA led
to a PC1 containing 93% of the total variance; thus only PC1
was considered for further analysis. This interpretation also
was confirmed by plotting the individual scores for PC1
against age (Fig. 2), where highest scores were related to
oldest individuals. Finally, plotting PC1 against PC2 (explain-
ing 7% of the total variance) led to a subdivision between the
youngest and oldest individuals (figure not shown).
Analysis of the PC1 scores showed significant differences
among generation types in all 3 age classes (1st age class:
F2,123 5 5.18, P , 0.01; 2nd age class: F2,323 5 38.19, P ,
0.001; 3rd age class: F1,224 5 4.49, P , 0.05). Pairwise
comparisons (after Bonferroni’s correction) showed a signif-
icant difference only between a and c in the 1st age class (P ,
0.01), with b intermediate. The strongest differences were
found in the 2nd age class, where all generation types differed
significantly from each other (always P , 0.001; Fig. 2). In
the 1st age class (Fig. 3) the 2 CVs explained 64% and 36% of
the between-group variance, respectively. The measurements
showing highest loadings were smallest palatal breadth,
distance between M1 and the upper incisor, and greatest
breadth of the nasals. In the 2nd age class (Fig. 3) the 1st CV
described 52% of the between-group variance, and the 2nd CV
explained 48%. The highest loadings were observed for
smallest palatal breadth, smallest interorbital breadth, and
greatest breadth of M1 (Fig. 3). In the 1st age class the 2 CVs
contained a different amount of variance, but in the 2nd age
class they accounted for nearly the same amount.
Plotting the 1st CV against the 2nd one for the first 2 age
classes highlighted the presence of 3 distinct, although
overlapping, groups corresponding to the generation types.
In the 1st age class the 1st CV separated a from b and c. The
2nd CV differentiated between b and c. In age class 2 the 1st
CV separated b and c, and the 2nd separated b and c from a
(Fig. 3). The axis of group separation that divides a from band c was always perpendicular to the one that divides b from
c, although the relative importance of the 2 axes changes from
the 1st to 2nd age class. Observing CV loadings and the
FIG. 2.—Average scores of the 1st principal component (PC1 scores)
based on linear measurements for the 3 generation types in the different
age classes. Group scores on PC1 were significantly different (see
‘‘Results’’). Lines connecting the points represent growth trajectories
for the 3 generation types. Vertical bars depict SEs.
FIG. 3.—First and 2nd canonical variates (CVs) for the 1st and 2nd age classes (linear measurements). Although some overlap can be
observed, 3 groups are distinguishable. Lines represent the direction and importance of each measurement in discriminating among groups.
Ellipses represent 90% confidence levels. Dashed black line represents a, gray line b, and dotted black line c generations.
1400 JOURNAL OF MAMMALOGY Vol. 92, No. 6
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022

amount of variation accounted for by the 1st CV and 2nd CV
in both age classes, it is appears that differences among
generation types seem to be led by the same measurements
in both 1st and 2nd age class, and in the 2nd age class
morphological differences between c and b become more
prominent in comparison to the 1st age class.
In the 3rd age class (Fig. 4)—containing only individuals
from the b and c generation such that only 1 linear discriminant
function could be determined—the measurements that contrib-
uted most were greatest breadth of M1, breadth of zygomatic
plate, and breadth of upper dental arch measured on M1.
Permutation tests indicated that all Mahalanobis distances were
significant (Table 2). Bivariate allometry coefficients were
mostly consistent with their multivariate counterparts (Table 3).
All measurements had the same allometric pattern in the 2
generation types considered, except for breadth of zygomatic
plate (isometric in b and positively allometric in c), breadth of
the nasals (isometric in b and negatively allometric in c), and
greatest rostrum breadth (positively allometric in b and
isometric in c). Among the measurements that showed the
same allometric pattern zygomatic breadth and breadth of the
upper dental arch were more negative in c individuals.
Multivariate allometry coefficients (Table 3; Fig. 5) showed 7
variables—condylobasal length, henselion–basion distance,
henselion–palation distance, diastema length, M1–upper incisor
distance, nasals length, and jaw greatest height—to be
positively allometric in both generation types. Negative
allometry was shown by 7 measurements in both generation
types: interorbital breadth, length of upper cheek-teeth alveoli,
breadth of the upper dental arch, greatest breadth of the 1st
upper molar, M1–M3 distance (jaw), length of the bulla, and
greatest breadth of braincase. Differences in allometric patterns
between b and c individuals were found for 5 measurements:
length of palatal foramen, zygomatic breadth, breadth of
zygomatic plate, greatest breadth of nasals, and greatest rostrum
breadth. Length of palatal foramen was isometric in b and
slightly positively allometric in c; zygomatic breadth showed
isometry in b and negative allometry in c. Both generation types
showed positive allometry for breadth of zygomatic plate but
near isometry in b individuals; the greatest breadth of nasals
was slightly positively allometric in b and negatively allometric
in c. Finally, the greatest rostrum breadth was isometric in c and
positively allometric in b. The confidence intervals for the
difference between the 2 bootstrap tables showed that the
coefficients differed not only for the traits with different
FIG. 4.—Canonical variate (CV) scores for the 3rd age class (linear
measurements). Because only the b and c generations are represented
in this age class, only 1 CV is estimated.
TABLE 2.—Mahalanobis distances among generation types for each
age class for traditional (linear) and geometric (landmarks)
morphometrics data. See ‘‘Results’’ for details on statistical significance.
Linear Landmarks
Alpha Beta Alpha Beta
Age class 1
Beta 2.5609 1.8587
Gamma 1.9831 2.4004 1.8062 1.8198
Age class 2
Beta 2.4820 2.4503
Gamma 2.640 1.6916 3.1636 1.8372
Age class 3
Gamma 1.8061 2.1546
TABLE 3.—Allometry and bivariate and multivariate allometric
coefficients for each linear measurement. Symbols used indicate
different trends in the allometric pattern: +, positive allometry;
5, isometry; 2, negative allometry. * P , 0.05. See Fig. 1 for
definitions of measurements.
Multivariate Bivariate Coefficients
Beta Gamma Beta Gamma Beta Gamma
GL 5 5 0.219 0.219
CL + + + + 0.244 0.255
HB + + + + 0.268 0.273
HP + + + + 0.238 0.253
PL 5 + 5 5 0.221 0.242
DL + + + + 0.289 0.305
MoID + + + + 0.304 0.321
IB 2 2 2 2 0.087 0.089
BB 5 2* 2 2* 0.213 0.173
PB 2 2 2 2 0.177 0.177
UAL 2 2 2 2 0.092 0.081
UDB 2 2* 2 2* 0.135 0.110
Mo1 2 2 2 2 0.027 0.023
ZPB + +* 5 +* 0.234 0.287
NB + 2* 5 2* 0.228 0.152
NL + + + + 0.279 0.289
MoJ 2 2 2 2 0.071 0.053
TBL 2 2 2 2 0.091 0.077
BrB 2 2 2 2 0.089 0.077
RH + + + + 0.254 0.251
RB + 5* + 5* 0.269 0.227
JH + + + + 0.289 0.294
December 2011 BRENO ET AL.—TRADITIONAL AND GEOMETRIC MORPHOMETRICS 1401
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022

allometric patterns but also for a few that had similar allometry:
condylobasal length, length of diastema, M1–upper incisor
distance, and breadth of the upper dental arch.
Geometric morphometrics.—Multivariate analysis of vari-
ance of relative warps showed differences among generation
types for each age class (age class 1: Pillai’s trace 5 0.72,
F2,92 5 1.96, P 5 0.001; age class 2: Pillai’s trace 5 0.91,
F2,123 5 4.11, P , 0.001; age class 3: Pillai’s trace 5 0.54,
F1,69 5 2.74, P 5 0.001). The first 5 PCs explained 70% of
the total variance of the entire data set. Age classes divided
along the 1st axis, whereas other PCs did not show any pattern
related to generation types or any other grouping factors
(details not shown). PCA performed separately for each age
class showed a separation between a and c generations along
PC1, with b individuals distributed all along the axis in the
2nd age class (Fig. 6), whereas c specimens tended to cluster
together along PC2 in the 3rd age class (Fig. 6) with no clear
pattern detected in the 1st age class (details not shown). Shape
transformations relative to PC1 in the 2nd age class showed a
longer rostrum and a narrower and shorter neurocranium in
the positive side of the axis (Figs. 6 and 7A). Shape changes
associated with PC2 (Fig. 7B) in the 3rd age class were related
to the length and breadth of rostrum and breadth of the frontal
bone, depicting a narrower skull in the middle and anterior
region. First and 2nd CV axes for the 1st age class (Fig. 8)
contained 54% and 46% of among-group variation, respec-
tively. Generations a and c tended to be separated along the
1st axis, and the b generation differed from a and c along the
2nd axis (Fig. 8). Variation retained by the 1st CV was
manifested mainly in the length of the parietal region, whereas
variation in the 2nd CV involved primarily length and breadth
of the middle and anterior regions. This analysis for the 2nd
age class showed a similar pattern as in 1st age class (Fig. 8),
but the 1st CV explained more variation than the 2nd (72%
compared to 28%). Shape variation associated with the 1st
CV referred to the length of the rostrum and width of
neurocranium. Variation in the 2nd shape transformation
involved primarily a posterolateral–anteromedial shift of
maxillary bone in the region of the infraorbital fissure. CVA
in the 3rd age class showed a separation between b and c,
FIG. 5.—Multivariate allometric coefficients (linear measure-
ments) for each trait. Vertical bars represent 95% bootstrapped
confidence intervals. The horizontal line represents the pure
isometric vector. See Fig. 1 for definitions of measurements.
FIG. 6.—Plot of the 1st and 2nd principal component scores (PC1 versus PC2) in the 2nd (left graph) and 3rd (right graph) age classes
(geometric morphometrics). The 3 generation types separate along PC1 in the 2nd age class. PC2 appears to separate the b and c generations in
the 3rd age class.
1402 JOURNAL OF MAMMALOGY Vol. 92, No. 6
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022
although some overlap was apparent (Fig. 8). Variation along
the single CV extracted involved mainly the width of the middle
and parietal regions of the skull. Mahalanobis distances among
groups were significantly higher than zero in all age classes
(Table 2), whereas Procrustes distances among groups differed
strongly from zero only in the 2nd age class.
DISCUSSION
Morphometrics is a powerful tool for the study of biological
variation that we used to investigate morphological dissimi-
larities within a genetically homogenous population across
successive years differing in developmental trajectories seem-
ingly due to variation in environmental conditions. Generation b,
led by allometric growth, converged toward c shape when
growth stop ended. However, generation a, which encountered a
longer growth stop, failed to reach the same point, revealing that
effects of long-lasting pause cannot be compensated for entirely.
Multivariate and geometric morphometrics analyses applied
in this study showed that the skulls of the 3 generation
types are morphologically distinguishable, although a certain
overlap was clearly present. The presence and the length of a
FIG. 8.—Canonical variate (CV) scores of the Procrustes superimposed data for 1st, 2nd, and 3rd age classes, left to right, respectively
(geometric morphometrics). Although some overlap exists, different generation types appear to be distinguishable. Ellipses represent 90%
confidence levels. Dashed black line represents a, gray line b, and dotted black line c generations.
FIG. 7.—Shape variation represented by principal component analysis. A) Shape change associated with the 1st principal component (PC1)
with respect to the mean shape in the 2nd age class along the negative (left) and positive (right) sides of the axis. The dashed picture represents
the average shape. B) Shape changes associated with PC2 in respect to the mean shape in the 3rd age class along the negative (left) and positive
(right) sides of the axis. The dashed image represents the average shape.
December 2011 BRENO ET AL.—TRADITIONAL AND GEOMETRIC MORPHOMETRICS 1403
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022

period of reduced growth seemed to influence the patterns of
morphological variations directly. As expected, when evalu-
ating growth, generation b and c diverged strongly early in
development and then tended to reach a common point later
(in the 3rd age class). Moreover, b developed somewhat
between the slowest (a) and the fastest (c) growing
generations. PC1 scores of the 2nd age class, obtained with
geometric data, illustrated a clear separation between a and cgenerations, with b individuals distributed all along this axis
of variation. The 1st PC derived from Procrustes coordi-
nates was interpreted as the allometric component of shape
variation, a common feature of PCA when performed on
variance–covariance matrices of Procrustes data (Mitteroecker
et al. 2004). The distribution of specimens along this axis
showed that growth rate had a direct effect on morphological
variation among generation types. In contrast, neither PC1
nor PC2 extracted from the variance–covariance matrix of
superimposed coordinates in the 3rd age class had any
relationship with allometric change (details not shown).
Although caution should be exercised in interpreting PCs
other than the 1st one, when PC1 is the allometric component
(Mitteroecker and Gunz 2009), the clustering of c individuals
along PC2 in the 3rd age class can be seen as a source of shape
variation (15% of the total variation) independent from size.
The c generation was born from b, thus b individuals that were
born late in the main breeding season experienced environ-
mental conditions similar to those encountered by individuals
from the c generation, and that would explain why some the
individuals from the b generation cluster with those from c.
Both traditional and geometric approaches led to discrimination
among generation types across age classes despite a smaller
sample size for geometric analyses (292, compared to 856 for
the traditional approach). Moreover, data based on Procrustes
superimposition capture the geometry of the structures being
studied and thus allow for a better description of morphological
variation. This capability allowed us to recognize at least 2 main
sources of divergence among generation types in subsequent
age classes. Moreover, description of shape change associated
with the biological factors being considered is one of the most
remarkable features of geometric morphometrics. This charac-
teristic led to a clear depiction of the morphological differ-
ences among generation types. Depictions of shape variation
associated with CV loadings in the 2nd and 3rd age classes and
with PC2 coefficients in the 3rd age class, derived from
geometric data, showed that generation b had a shorter and
wider skull than generation c. A similar picture can be acquired
from multivariate allometric coefficients of linear measurements,
which showed that generation c presented a relatively longer and
narrower rostrum and a relatively shorter zygomatic breadth.
The presence of a period of reduced growth led to a
difference in the growth trajectories; c grew with a curve often
observed in rodents (Gaillard et al. 1997; Jackson and van
Aarde 2003), whereas b grew at a slower but constant rate for a
longer period. Allometric growth, shape change associated with
size and with age, thus is expected to drive the divergence at the
beginning and converge toward a common point when the
catch-up process occurs in the generation b. In the meantime
local differences in available resources are likely to act directly
on the skull, adding another source of shape variation.
Availability and quality of food sources across different years
are likely to affect the morphology of the skull both as a
consequence of a general influence on growth rates and locally at
the level of skull structure (i.e., changes in strain and loadings).
Quantitative data about abundance and composition of food were
not available, but we can speculate. Rainfall pattern seems to
have a direct effect on availability and composition of food in
Morogoro. Food is seen as the means by which environmental
conditions can affect the growth pattern of M. natalensis in
Morogoro (Leirs et al. 1994). According to several authors,
seasonal variation in diet occurs in M. natalensis, with arthropods
and grasses preferred over other kinds of resources during wet
seasons when their availability increases (Monadjem 1998;
Odhiambo et al. 2008). Thus, a shift in the composition of the
food resource is likely to happen among generation types. In rats
transitional perturbations can lead to a catch-up process occurring
after the perturbation to recuperate normal growth; rats fed a
high-protein diet had a higher growth rate than rats fed a low-
protein diet, although the latter grew significantly longer so that
few differences were apparent in the adult traits studied in the 2
groups (Reichling and German 2000). The low-protein diet,
however, affected craniofacial growth, leading to shorter and
relative wider viscerocranium regions. These results clearly
showed a change in adult shape but not in final size (Miller and
German 1999). In contrast, squirrel monkeys (Saimiri sciureus
boliviensis) fed low- and high-protein diets, and studied with both
traditional and geometric morphometrics, had similar shape but
different sizes of the skull (Ramirez Rozzi et al. 2005). Not only
the nutritional quality but also the structure of food has been shown
to affect skull morphology in rodents. In the prairie deer mouse
(Peromyscus maniculatus bairdii) individuals fed a soft diet
showed smaller masseteric tubercles, a narrowed zygomatic plate,
and anteriorly shifted incisors (Myers 1996). Shifting of incisors
also was observed in rats (Kiliaridis 1986), whereas a narrowed
zygomatic plate was not reported in other studies that showed
differences in width of palate and maxillae and in the braincase
(Beecher and Corruccini 1981; Moore 1965). Although many
studies have assessed the relationship between craniofacial growth
and masticatory strains in mammals, it is difficult to recognize
comparable patterns due to the great variability in experimental
methods and results (Kiliaridis 2006). Nevertheless, several
findings support the hypothesis that nonallometric divergence in
b and c could be caused by differences in food consistency. A
relative increase of skull width in animals fed a hard diet has been
reported (Abed et al. 2007; Ciochon et al. 1997; Corrucini and
Beecher 1984; Lieberman et al. 2004). Moreover, M. natalensis
living in a swampy area had a longer and narrower skull when
compared to a population from an agricultural area (Lalis et al.
2009a). These results are consistent with our observations.
Generation c individuals likely fed mainly on arthropods and
green vegetation, whereas b individuals tended to eat hard seeds,
the same diet composition reported, respectively, for the
‘‘swampy’’ and the ‘‘rural’’ populations of Lalis et al. (2009a).
1404 JOURNAL OF MAMMALOGY Vol. 92, No. 6
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022

Although with the current data we could not assess any
specific relationship between diet and morphological variation
among generation types, quality and composition of food
likely are the major determinants of the observed differences
in skull growth and skull shape among successive generations
of M. natalensis in Morogoro. This argues for plasticity being
an important source of shape variation, and one that should be
considered in studies of intraspecific ecogeographic variation.
ACKNOWLEDGMENTS
Fieldwork was carried out by different Tanzanian and visiting
staff at the SUA Pest Management Center, Sokoine University of
Agriculture, Morogoro, Tanzania. Financial support was provided by
the Belgian Directorate-General for Development Cooperation and
the Flemish Interuniversity Council–University Development Coop-
eration. C. Bogaerts and W. Wendelen cleaned and measured all the
skull material for this work. MB holds a Ph.D. Fellowship from the
Research Foundation–Flanders.
LITERATURE CITED
ABDALA, F., D. A. FLORES, AND N. P. GIANNINI. 2001. Post weaning
ontogeny of the skull of Didelphis albiventris. Journal of
Mammalogy 82:190–200.
ABDEL-RAHMAN, E. H., P. J. TAYLOR, G. CONTRAFATTO, J. M. LAMB,
P. BLOOMER, AND C. T. CHIMIMBA. 2009. Geometric craniometric
analysis of sexual dimorphism and ontogenetic variation: a case
study based on two geographically disparate species, Aethomys
ineptus from southern Africa and Arvicanthis niloticus from Sudan
(Rodentia: Muridae). Mammalian Biology 74:361–373.
ABED, G. S., P. H. BUSCHANG, R. TAYLOR, AND R. J. HINTON. 2007.
Maturational and functional related differences in rat craniofacial
growth. Archive of Oral Biology 52:1018–1025.
ADAMS, D. C., F. J. ROHLF, AND D. E. SLICE. 2004. Geometric
morphometrics: ten years of progress following the ‘revolution.’
Italian Journal of Zoology 71:5–16.
BEECHER, R. M., AND R. S. CORRUCCINI. 1981. Effects of dietary
consistency on craniofacial and occlusal development in the rat.
Angle Orthodontist 51:61–69.
BERNAL, V. 2007. Size and shape analysis of human molars:
comparing traditional and geometric morphometric techniques.
Homo—Journal of Comparative Human Biology 58:279–296.
BEVER, G. S. 2008. Comparative growth in the postnatal skull of the
extant North American turtle Pseudemys texana (Testudinoidea:
Emydidae). Acta Zoologica (Stockholm) 89:107–131.
BLANCO, M. B., AND L. R. GODFREY. 2006. Craniofacial sexual
dimorphism in Alouatta palliata, the mantled howling monkey.
Journal of Zoology (London) 270:268–276.
BOOKSTEIN, F. L. 1991. Morphometric tools for landmark data: geometry
and biology. Cambridge University Press, Cambridge, United Kingdom.
BOOKSTEIN, F. L. 1996. Biometrics, biomathematics and the morpho-
metrics synthesis. Bulletin of Mathematical Biology 58:313–365.
BRONNER, G. N., M. VAN DER MERWE, AND K. NJOBE. 2007.
Nongeographic cranial variation in two medically important
rodents from South Africa, Mastomys natalensis and Mastomys
coucha. Journal of Mammalogy 88:1179–1194.
CIOCHON, R. L., R. A. NISBETT, AND R. S. CORRUCCINI. 1997. Dietary
consistency and craniofacial development related to masticatory
function in minipigs. Journal of Craniofacial Genetics and
Developmental Biology 17:96–102.
CORRUCINI, R. S., AND R. BEECHER. 1984. Occlusofacial morphological
integration lowered in baboons raised on soft diet. Journal of
Craniofacial Genetics and Developmental Biology 4:135–142.
DRYDEN, I. L., AND K. V. MARDIA. 1998. Statistical analysis of shape.
John Wiley & Sons, Inc., New York.
FADDA, C., AND H. LEIRS. 2009. The role of growth stop as morphogenetic
factor in Mastomys natalensis (Rodentia: Muridae). Biological Journal
of the Linnean Society 97:791–800.
GAILLARD, J. M., D. PONTIER, D. ALLAINE, A. LOISON, J. C. HERVE, AND
A. HEIZMAN. 1997. Variation in growth form and precocity at birth
in eutherian mammals. Proceedings of the Royal Society of
London, B. Biological Sciences 264:859–868.
GIANNINI, N. P., F. ABDALA, AND D. A. FLORES. 2004. Comparative
postnatal ontogeny of the skull in Dromiciops gliroides (Marsupi-
alia: Microbiotheriidae). American Museum Novitates 3460:1–17.
GOODALL, C. 1991. Procrustes methods in the statistical analysis of shape.
Journal of the Royal Statistical Society, B. Methodological 53:285–339.
JACKSON, T. P., AND R. I. VAN AARDE. 2003. Sex- and species-specific
growth patterns in cryptic African rodents: Mastomys natalensis
and M. coucha. Journal of Mammalogy 84:851–860.
JOLICOEUR, P. 1963. The multivariate generalization of the allometry
equation. Biometrics 19:497–499.
KILIARIDIS, S. 1986. The relationship between masticatory function and
cranial morphology: the eruption pattern of the incisors in the growing
rat fed a soft diet. European Journal of Orthodontics 8:71–79.
KILIARIDIS, S. 2006. The importance of masticatory muscle function in
dentofacial growth. Seminars in Orthodontics 12:110–119.
KLINGENBERG, C. P. 1996. Multivariate allometry. Pp. 23–49 in
Advances in morphometrics (L. F. Marcus, M. Corti, A. Loy,
G. J. P. Naylor, and D. E. Slice, eds.). Plenum Press, New York.
KLINGENBERG, C. P. 1998. Heterochrony and allometry: the analysis of
evolutionary change in ontogeny. Biological Reviews 73:79–123.
KLINGENBERG, C. P. 2011. MorphoJ: an integrated software package for
geometric morphometrics. Molecular Ecology Resources 11:353–357.
http://www.flywings.org.uk/MorphoJ_page.htm. Accessed 13 July 2011.
KRYSTUFEK, B. 2002. Cranial variability in the eastern hedgehog
Erinaceus concolor (Mammalia: Insectivora). Journal of Zoology
(London) 258:365–373.
LALIS, A., M. BAYLAC, J. F. COSSON, R. H. MAKUNDI, R. S. MACHANG’U,
AND C. DENYS. 2009a. Cranial morphometric and fine scale genetic
variability of two adjacent Mastomys natalensis (Rodentia:
Muridae) populations. Acta Theriologica 542:171–181.
LALIS, A., A. EVIN, AND C. DENYS. 2009b. Morphological identification
of sibling species: the case of West African Mastomys (Rodentia:
Muridae) in sympatry. Comptes Rendus Biologies 332:480–488.
LAWING, A. M., AND P. D. POLLY. 2009. Geometric morphometrics:
recent applications to the study of evolution and development.
Journal of Zoology (London) 280:1–7.
LEAMY, L. 1983. Variance partitioning and effects of sex and age on
morphometric traits in random bred house mice. Journal of
Mammalogy 64:55–61.
LEIRS, H. 1995. Population ecology of Mastomys natalensis (Smith,
1834). Implications for rodent control in Africa. Administration for
Development Cooperation, Agricultural Editions, Scientific Pub-
lications, Brussels, Belgium.
LEIRS, H., R. VERHAGEN, AND W. VERHEYEN. 1993. Productivity of
different generations in a population of Mastomys natalensis in
Tanzania. Oikos 68:53–60.
LEIRS, H., R. VERHAGEN, AND W. VERHEYEN. 1994. The basis of
reproductive seasonality in Mastomys rats (Rodentia: Muridae) in
Tanzania. Journal of Tropical Ecology 10:55–66.
December 2011 BRENO ET AL.—TRADITIONAL AND GEOMETRIC MORPHOMETRICS 1405
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022

LIEBERMAN, D. E., G. E. KROVITZ, F. W. YATES, M. DEVLIN, AND M. ST.
CLAIRE. 2004. Effects of food processing on masticatory strain and
craniofacial growth in a retrognathic strain and face. Journal of
Human Evolution 46:655–677.
MADERBACHER, M., C. BAUER, J. HERLER, L. POSTL, L. MAKASA, AND
C. STURMBAUER. 2008. Assessment of traditional versus geometric
morphometrics for discriminating populations of the Tropheus
moorii species complex (Teleostei: Cichlidae), a Lake Tanganyika
model for allopatric speciation. Journal of Zoological Systematics
& Evolutionary Research 46:153–161.
MILLER, J. P., AND R. Z. GERMAN. 1999. Protein malnutrition affects
the growth trajectories of the craniofacial skeleton in rats. Journal
of Nutrition 129:2061–2069.
MITTEROECKER, P., AND P. GUNZ. 2009. Advances in geometric
morphometrics. Evolutionary Biology 36:235–247.
MITTEROECKER, P., P. GUNZ, M. BERNHARD, K. SCHAEFER, AND F. L.
BOOKSTEIN. 2004. Comparison of cranial ontogenetic trajectories
among great apes and humans. Journal of Human Evolution
46:679–697.
MONADJEM, A. 1998. Reproductive biology, age structure, and diet of
Mastomys natalensis (Rodentia: Muridae) in a Swaziland grass-
land. Zeitschrift fur Saugetierkunde 63:347–356.
MOORE, W. J. 1965. Masticatory function and skull growth. Journal of
Zoology (London) 146:123–131.
MYERS, P., B. L. LUNDRIGAN, B. W. GILLESPIE, AND M. L. ZELDITCH.
1996. Phenotypic plasticity in skull and dental morphology in the
prairie deer mouse (Peromyscus maniculatus bairdii). Journal of
Morphology 229:229–237.
ODHIAMBO, R. O., R. H. MAKUNDI, H. LEIRS, AND R. VERHAGEN. 2008.
Dietary selection in Mastomys natalensis (Rodentia: Muridae) in
the maize agro-ecosystem of central and southwestern Tanzania.
Mammalia 72:169–177.
RAMIREZ ROZZI, F. V., R. GONZALES-JOSE, AND H. P. PUCCIARELLI. 2005.
Cranial growth in normal and low-protein-fed Saimiri: an environ-
mental heterochrony. Journal of Human Evolution 49:515–535.
RASBAND, W. S. 2010. ImageJ. 1997–2010. United States National
Institutes of Health, Bethesda, Maryland. http://rsb.info.nih.gov/ij/.
Accessed 13 July 2011.
R DEVELOPMENT CORE TEAM. 2009. R: a language and environment for
statistical computing. R Foundation for Statistical Computing,
Vienna, Austria. http://www.R-project.org. Accessed 13 July 2011.
REICHLING, T. D., AND R. Z. GERMAN. 2000. Bones, muscles and
visceral organs of protein-malnourished rats (Rattus norvegicus)
grow more slowly but for longer durations to reach normal final
size. Journal of Nutrition 130:2326–2332.
ROHLF, F. J. 1990. Morphometrics. Annual Review of Ecology and
Systematics 21:299–316.
ROHLF, F. J., AND L. F. MARCUS. 1993. A revolution in morphometrics.
Trends in Ecology & Evolution 8:129–132.
SMITH, J. I. 1998. Allometric influence on phenotypic variation in
the song sparrow (Melospiza melodia). Zoological Journal of the
Linnean Society 122:427–454.
SUNDBERG, P. 1989. Shape and size-constrained principal component
analysis. Systematic Zoology 38:166–168.
VAN HOOFT, P., J. F. COSSON, S. VIBE-PETERSEN, AND H. LEIRS. 2008.
Dispersal in Mastomys natalensis mice: use of fine-scale genetic
analyses for pest management. Hereditas 145:262–273.
ZELDITCH, M. L., D. L. SWIDERSKI, H. D. SHEETS, AND W. L. FINK. 2004.
Geometric morphometrics for biologists: a primer. Elsevier,
Amsterdam, Netherlands.
Submitted 28 September 2010. Accepted 15 May 2011.
Associate Editor was Elizabeth R. Dumont.
1406 JOURNAL OF MAMMALOGY Vol. 92, No. 6
Dow
nloaded from https://academ
ic.oup.com/jm
amm
al/article/92/6/1395/961102 by guest on 11 September 2022




















