
microbial dynamics and dysbiosis in farmed European ...
248
D To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream Daniela Rosado Biodiversity, Genetics and Evolution Department of Biology 2021 Advisor Raquel Xavier, Auxiliary Researcher, CIBIO-InBIO/FCUP Co-advisors Marcos Pérez-Losada, Auxiliary Researcher, CIBIO-InBIO/George Washington University Jo Cable, Professor, Cardiff University
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of microbial dynamics and dysbiosis in farmed European ...

DTo be or not to bediseased: microbial dynamics anddysbiosis in farmedEuropean seabassand gilthead seabreamDaniela RosadoBiodiversity, Genetics and EvolutionDepartment of Biology2021
AdvisorRaquel Xavier, Auxiliary Researcher, CIBIO-InBIO/FCUP
Co-advisorsMarcos Pérez-Losada, Auxiliary Researcher, CIBIO-InBIO/George Washington UniversityJo Cable, Professor, Cardiff University

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream ii

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream iii
FOREWORD
According to the General Regulation of Doctoral Programs of the University of Porto (number
2, 4th Article) and the Decree Law 74/2006 (Article 31, 24 of March) revised under the Decree
law 230/2009 (14th of September), this thesis includes manuscripts published or in
consideration for publication in peer-reviewed scientific journals. These manuscripts are the
result of collaborations with several co-authors. The candidate declares that he actively
contributed to the ideas and the development of the research work, including the compilation,
analysis, results, discussion and writing as in its current publication form. The candidate was
supported by the National Foundation for Science and Technology (FCT), through a PhD
Grant (PD/BD/117943/2016) and through the I&D Projects (PTDC/MAR-BIO/0902/2014;
PTDC/BIA-MIC/27995/2017), financed by the European Social Fund and by the National
Ministry of Science, Technology and Higher Education (MCTES), through the Operational
Programme Human Capital (POCH), under Portugal 2020, and co financed by the European
Fund of Regional Development (FEDER), through COMPETE – Operational Program for
Competitiveness Factors (POFC). This thesis’ research was developed in the context of the
Doctoral Programme in Biodiversity, Genetics and Evolution (Faculty of Sciences, Univeristy
of Porto). The work was conducted at the Research Centre in Biodiversity and Genetic
Resources - InBIO Associate Laboratory (CIBIO-InBIO).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
iv
ACKNOWLEDGEMENTS
Although doing a PhD can sometimes feel like a lonely road to travel, I am incredibly
privileged by all the support, guidance and companionship that was given to me in what ended
up being one of the best things I’ve ever done in life. I am genuinely proud of all the work done
in the scope of this thesis, and I cannot let it end without acknowledging how grateful I am to
all the people involved. I wouldn’t have been able to do it alone.
To Raquel, the most important person in this thesis, and my personal career hero and
role model. I have never known someone with such a busy life that is, at the same time, able
to always be present. I will be forever tremendously grateful for the day that you thought of me
to enter this microbiome journey with you. Beyond that opportunity, I am grateful to you for
allowing me to always follow my own path and make my own decisions (and mistakes). I have
learned so much through you and because of you. Thank you for the endless conversations,
exchange of ideas, corrections and comments, and thank you for all the laughs. Above all,
thank you for always treating me like a person, and not just a student, and for always being
fair and kind.
To Marcos for all of his wisdom and tremendous patience in passing it on to me. Thank
you for always making time for me, my questions and doubts. Without your enormous help,
this thesis would not have gone far.
To Jo, for all the words of encouragement, kindness and guidance. I always felt very
supported by you. Thank you for being present at any time and for trusting me. You made my
time in Cardiff really pleasant by introducing me to everyone, with coffee breaks and walks on
the park. I will always think of you both as a colleague and as a friend.
To Ricardo, without whom this project wouldn’t be possible. Thank you for taking care
of all the logistics related with sample collection and shipping, for giving us ideas and for
patiently replying to all of my questions and clarifying all my doubts.
Thank you to everyone else who participated, to any extent, in the making of the
scientific articles, especifically Fernando Tavares, Ana Pereira and Pedro Tarroso. I am also
thankful to the people in the lab who initially helped me, including Diana Castro, Susana
Lopes, Sara João and Sofia Mourão.
Thank you to my sweet friends for all the love, support and patience throughout, not
only the difficult times, but also the good ones. Thank you for listening and taking care of me
and making me enjoy life a lot more. Included here, but deserving a special acknowledgement,
are the people from theater school, for one of the most incredible times of my life. Thank you

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
v
for all the conversations, either highly intelectual or completely dumb, for all the laughter and
lessons and, ultimately, for boosting my confidence.
To Dimitra, the personification of love. Thank you for all the talks and shared secrets,
all the nights out and walks together during the day. Thank you for discovering the city of Porto
with me and for being the living proof that it is possible to make good friends for life in
adulthood. Above all, thank you for being my companionship during quarantine, for the daily
yoga classes and for not making me feel alone in the past months. You are one of the greatest
people that walks on this planet and how grateful I am that our paths ever crossed.
I thank profusely to Cristela, my pillar. There was never another friend who believed
so much in me and who cared so much for me. All of your kind words made me grow in ways
that I never thought were possible. Thank you for always speaking the truth, even though you
know I might not like to hear it; that is surely a rare quality. You’ve been with me since the
beginning of my academic life and I just know I can count on you for everything, anytime.
Looking forward to being two old ladies playing cards, drinking G&T, listening to good music
and questioning the meaning of everything.
I am extremely thankful to my family for the biggest support, especially in the last
months. A special thank you to my mommy and daddy, the two strongest and most resilient
people I have ever known. For teaching me how to be confident and for always making me
believe I can do anything I want. For all the help and comfort throughout my academic and
personal life. Thank you both for the unconditional love and support, even though you don’t
really know what I do for a living. To my baby sister Maria, for all the laughter, company and
appreciation; and thank you to her sweet daughter who gave me joy everyday.
I owe a deep sense of gratitude to Inês, my soulmate. Thank you for always being
there for me, listening to me without judgement, and always knowing what I want and need.
Being in sync with you made me through a lot of bad days since you were born. You can’t
imagine how lucky I feel to have my best friend born within my family. I would need to write
another thesis dedicated to how thankful I am to you. With all my heart, thank you.
A colossal thank you to Dr Cecília for all the dedication to my person and for all the
help in my personal growth.
To Gil, the person who has seen it all. Throughout these years you have seen all my
faces and you are, undoubtedly, the most present person and one of the most important. And
I couldn’t finish this list without including Floki and her companionship in all the early morning
or late afternoon walks to clear my head.
Studying the microbiome has made me realise how no living being is truly alone, in all
senses.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream vi
ABSTRACT
Aquaculture is currently the fastest growing food industry that has the capacity to meet
the global food demand. Intensification of aquaculture practices entails several constraints to
its development, most notably by infectious diseases. The mucosal surfaces of fish harbour
microbial communities that play a key role in host health and fitness. In addition to immunity,
other microbial functions benefit the host’s physiology and metabolism. Fish microbiota,
however, is highly variable and influenced by several biotic and abiotic factors. In particular,
host taxonomy, physiology, body site, diet, age, habitat and water parameters govern the
microbial dynamics of fish. Disruptions to favourable conditions can lead to microbial
imbalance, i.e. dysbiosis, affecting fish health and, eventually, leading to disease. Particularly
important in aquaculture are bacterial infections, stress, antibiotic usage and other chemical
treatments, which can induce dysbiosis in farmed fish. Monitoring microbial dynamics and
dysbiosis in farmed fish and their surroundings is paramount to improve aquaculture practices
as well as prevent and mitigate disease. Advances in molecular technology are facilitating
microbiome research, allowing the identification of microbes and the assessment of their
diversity and function.
This doctoral thesis assessed the microbial composition, diversity and structure of the
skin and gill of the European seabass Dicentrarchus labrax and gilthead seabream Sparus
aurata. These are two of the most commercially important farmed finfishes in Europe;
however, their productivity is greatly affected by infectious diseases. In close collaboration
with a fish farm, non-invasive weekly sampling of the external mucosae of both species was
performed between August 2016 and January 2018. I used a metataxomic approach to
sequence the V4 region of the 16S rRNA bacterial gene and analysed differences in the
microbial communities observed under different conditions. I studied the effects of several
biotic and abiotic factors in fish microbiota, including: host taxonomy, body site, age group,
water microbial communities, water temperature, disease and antibiotic treatment.
The first chapter sets the context of this dissertation, presenting a literature review of
aquaculture and microbiomes, enphasizing the microbiome of fish. Additionally, it reviews
microbiome applications in aquaculture settings as well as tools available for microbiome
assessment. Chapters 2 to 5 are presented as scientific papers, whose results interconnect
to reveal comprehensive knowledge about the complex bacterial dynamics and dysbiosis
occurring in farmed European seabass and gilthead seabream. A general discussion of those
results is presented in Chapter 6.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream vii
I identified consistent predominant bacterial taxa in both fish species and tissues,
through time and across host health conditions. The two mucosal tissues presented
differences in diversity and response to external stressors, although they seemed to be
governed by the same factors. Overall, the skin microbiota was more variable and more
susceptible to dysbiosis, while the gill microbiota was more stable, but less resilient to disease
and antibiotic treatment. The microbiota of both tissues was significantly different from that of
the sourrounding waters. Nonetheless, skin and water had a more similar microbial structure
than gill and water. Continuous microbial exchange was high between tissues, also occurring
from the surrounding water.
The European seabass and gilthead seabream harboured differences in microbial
diversity, which were most pronounced in its structure (beta-diversity). Differences in microbial
structure and predicted function were also detected between fish belonging to different age
groups in both species. However, only the seabass showed significant differences in the
microbial composition across age groups. Water temperature governed skin and gill bacterial
dynamics of the European seabass through the year. Changes in the abundance of several
potentially pathogenic genera (Aliivibrio, Pseudomonas, Photobacterium and Vibrio) were also
correlated with changes in temperature. The increase in abundance of one or more of these
potentially pathogenic genera on two occasions seemed to have led to dysbiosis in both
tissues, although disease was not observed. Dysbiosis also occurred as a result of infection
and antibiotic treatment in both adult and juvenile seabass. In the case of adult seabass,
infection by Photobacterium damselae and treatment with oxytetracycline induced asymmetric
responses in the skin and gill microbiota, further demonstrating an effect of body tissue. In
seabass fingerlings, co-infection of Photobacterium damseale subsp. piscicida and Vibrio
harveyi and flumequine treatment led to an increase in diversity in the skin microbiota, a
response less commonly reported for fish microbiota. Short-term recovery assessment in both
age groups indicated that microbial homeostasis was not fully attained after 3 weeks in adults
and 1 week in fingerlings.
This research showed a high variability of the skin and gill microbiota of farmed
European seabass and gilthead seabream through time driven by host related and
environmental factors. Additionally, both disease and antibiotic treatments showed a
detrimental effect in fish microbiota. Monitoring the microbial communities of farmed species
proved to be highly insightful towards identifying biomarkers for health and disease. These
biomarkers can ultimately be the answer to fundamentally improve aquaculture practices.
Keywords: fish microbiota, aquaculture, fish pathogens, dysbiosis, skin, gill, Dicentrarchus
labrax, Sparus aurata, bacteria, 16S rRNA, metataxonomics, water microbiota, ontogenesis,
antibiotics.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream viii
RESUMO
A aquacultura é atualmente a indústria alimentar em maior crescimento com a
capacidade de atender à demanda alimentar global. A intensificação das práticas de
aquacultura acarreta várias limitações ao seu desenvolvimento, em especial devido a
doenças infeciosas. As superfícies mucosas dos peixes abrigam comunidades microbianas
que desempenham um papel fundamental na saúde e aptidão do hospedeiro. Para além da
imunidade, outras funções microbianas beneficiam a fisiologia e metabolismo do hospedeiro.
No entanto, o microbioma dos peixes é altamente variável e influenciado por vários fatores
bióticos e abióticos. Em particular, a taxonomia do hospedeiro, fisiologia, tecido, dieta, idade,
habitat e parâmetros da água, determinam a dinâmica microbiana dos peixes. Alterações às
condições favoráveis podem levar ao desiquilíbrio microbiano, isto é, disbiose, e afetar a
saúde dos peixes, podendo, eventualmente, provocar doença. Particularmente importantes
na aquacultura são as infeções bacterianas, o stress, o uso de antibióticos e outros
tratamentos químicos que podem induzir a disbiose nos peixes. A monitorização das
dinâmicas microbianas e da disbiose nos peixes de piscicultura, bem como do seu ambiente,
é fundamental para melhorar as práticas de aquacultura bem como prevenir e mitigar
doenças. Avanços na tecnologia molecular têm facilitado a investigação na área dos
microbiomas, permitindo a identificação de micróbios e estimar a sua diversidade e função.
Esta tese de doutoramento aferiu a composição, diversidade e estrutura microbiana
da pele e brânquias do robalo Dicentrarchus labrax e dourada Sparus aurata. Estas espécies
são duas das espécies de peixe de cultivo mais importantes comercialmente na Europa; no
entanto, a sua produção é bastante afetada por doenças infeciosas. Em estreita colaboração
com uma piscicultura, amostragem semanal não-invasiva das mucosas externas de ambas
as espécies foi realizada entre Agosto de 2016 e Janeiro de 2018. Usei uma abordagem
metataxonómica para sequenciar a região V4 do gene bacteriano 16S rRNA e analisei as
diferenças nas comunidades microbianas observadas em diferentes condições. Estudei os
efeitos de vários fatores bióticos e abióticos na microbiota dos peixes, incluindo: taxonomia
do hospedeiro, tecido, idade, comunidades microbianas da água, temperatura da água,
doença e tratamento com antibiótico.
O primeiro capítulo contextualiza esta dissertação, apresentado uma revisão da
literatura sobre aquacultura e microbiomas, com um ênfase especial em microbiomas de
peixes. Além disso, analisa as aplicações práticas do estudo do microbioma em ambiente de
aquacultura, bem como as ferramentas disponíveis para a caracterização do microbioma. Os
capítulos 2 a 5 são apresentados como um conjunto de artigos científicos, cujos resultados

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream ix
se interligam, revelando um conhecimento abrangente sobre a complexidade das dinâmicas
microbianas e disbiose que ocorreu no robalo e na dourada. Uma discussão geral destes
resultados é apresentada no capítulo 6.
Identifiquei taxa bacterianos consistentemente predominantes nas duas espécies e
tecidos ao longo do tempo, independentemente da condição de saúde dos indivíduos. Os
dois tecidos mucosos apresentaram diferenças na diversidade e resposta a fatores de stress
externos, embora pareçam ser governados pelos mesmos fatores. No geral, a microbiota da
pele foi mais variável e mais susceptível a disbiose, enquanto a microbiota das brânquias foi
mais estável, embora menos resiliente a doença e tratamento com antibiótico. A microbiota
de ambos os tecidos foi significativamente diferente da microbiota da água. No entanto, a
pele apresentou uma estrutura microbiana mais semelhante à da água do que as brânquias.
O intercâmbio microbiano entre tecidos foi elevado, e ocorreu também entre tecidos e a água
circundante.
O robalo e a dourada apresentaram diferenças na diversidade microbiana entre si,
sendo essas diferenças mais pronunciadas a nível da estrutura (beta-diversidade). Diferenças
na estrutura microbiana e possível função também foram detetadas entre grupos de
indivíduos de ambas as espécies que pertenciam a diferentes idades. A temperatura da água
teve uma grande influência nas dinâmicas bacterianas da pele e brânquias do robalo ao longo
de um ano. Alterações na abundância de vários géneros potencialmente patogénicos
(Aliivibrio, Pseudomonas, Photobacterium e Vibrio) também foram correlacionadas com
mudanças na temperatura. O aumento da abundância de um ou mais desses géneros em
duas ocasiões parece ter levado à disbiose nos dois tecidos, embora não tenha sido detetada
nenhuma doença. A disbiose também ocorreu como consequência de infeção e tratamento
com antibióticos, tanto em robalos adultos como em juvenis. No caso do robalo adulto, infeção
provocada por Photobacterium damselae e tratamento com oxytetracyclina induziram
respostas assimétricas na microbiota da pele e brânquias, demonstrando mais uma vez um
efeito do tecido. No robalo juvenil, co-infeção provocada por Photobacterium damseale subsp.
piscicida e Vibrio harveyi e tratamento com flumequina, provocaram um aumento da
diversidade microbiana na pele, uma resposta menos reportada na microbiota de peixes. A
avaliação da recuperação a curto prazo em ambos os grupos etários indicou que a
homeostase microbiana não tinha sido totalmente atingida ao fim de 3 semanas em adultos
e 1 semana em juvenis.
Esta pesquisa mostrou uma alta variabilidade da microbiota da pele e brânquias do
robalo e dourada de aquacultura ao longo do tempo, influenciada por fatores ligados ao
hospedeiro e ao ambiente. Além disso, tanto doenças como tratamentos com antibiótico
mostraram um efeito prejudicial para a microbiota dos peixes. A monitorização das
comunidades microbianas de espécies de peixe de piscicultura provou ser bastante

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream x
informativo para a identificação de biomarcadores de saúde. Estes biomarcadores têm o
potencial de ser a resposta para melhorar as práticas de aquacultura.
Palavras-chave: microbiota de peixe, aquacultura, patógenos de peixe, disbiose, pele,
brânquias, Dicentrarchus labrax, Sparus aurata, bactérias, 16S rRNA, metataxonómica,
microbiota da água, ontogénese, antibióticos.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream xi
Table of Contents
Foreword iii
Acknowledgements iv
Abstract vi
Resumo viii
Table of Contents xi
List of Figures xvi
List of Tables xviii
List of Abbreviations xix
Chapter 1: General introduction 1
1.1 Literature review 1
1.1.1 Aquaculture overview 1
1.1.1.1 European seabass Dicentrarchus labrax and gilthead seabream
Sparus aurata aquaculture 3
1.1.2 Microbiome 4
1.1.3 Microbiome of fish 7
1.1.3.1 Fish microbial dynamics 8
1.1.3.2 Fish microbial imbalance and disease 12
1.1.3.3 Fish microbial function 15
1.1.4 Microbiome applications in aquaculture settings 18
1.1.5 Available tools for microbiome assessment 21
1.1.5.1 Sampling 21
1.1.5.2 Laboratory procedures for culture independent microbial
assessments 22
1.1.5.3 Data analyses 24
1.2 Thesis structure and objectives 27
1.3 References 28

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream xii
Chapter 2: Characterization of the skin and gill microbiomes of the
farmed seabass (Dicentrarchus labrax) and seabream (Sparus
aurata) 66
2.1 Abstract 66
2.2 Introduction 67
2.3 Material and methods 68
2.3.1 Sample collection and preparation 68
2.3.1 Data and statistical analyses 69
2.4 Results 70
2.4.1 Taxonomic bacterial composition and core microbiome of seabass and
seabream 70
2.4.2 Microbial diversity 73
2.5 Discussion 75
2.5.1 Core microbiome composition 75
2.5.2 Potential pathogens detected in the core microbiomes 77
2.6 Conclusion 77
2.7 Acknowledgements 78
2.8 References 78
2.9 Supplementary material 87
Chapter 3: Effects of aging on the skin and gill microbiota of farmed
seabass and seabream 88
3.1 Abstract 88
3.2 Introduction 89
3.3 Material and methods 90
3.3.1 Fish species, sampling and preparation 90
3.3.2 Data processing and statistical analysis 92
3.4 Results 94
3.4.1 Microbial diversity across age groups 94
3.4.1.1 Alpha-diversity 94
3.4.1.2 Beta-diversity 97
3.4.1.3 Bacterial taxa 97
3.4.2 Microbial predicted functional diversity across age groups 100
3.4.3 Fish and water microbiota comparisons 102
3.5 Discussion 103

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream xiii
3.5.1 Microbial diversity across age groups 103
3.5.2 Microbial predicted functional diversity across age groups 105
3.5.3 Fish and water microbiota comparisons 106
3.6 Conclusion 107
3.7 Acknowledgements 108
3.8 References 108
3.9 Supplementary material 118
Chapter 4: Longitudinal sampling of external mucosae in farmed
European seabass reveals the impact of water temperature on
bacterial dynamics 125
4.1 Abstract 125
4.2 Introduction 126
4.3 Material and methods 128
4.3.1 Experimental design, sampling and processing 128
4.3.2 Data processing and statistical analysis 128
4.4 Results 130
4.4.1 Bacterial composition and temporal dynamics of the microbiota 130
4.4.2 Dynamics of potentially pathogenic (PP) genera 134
4.4.3 Effect of water temperature on fish microbiota 135
4.5 Discussion 139
4.5.1 Temporal dynamics of the microbiota 139
4.5.2 Water temperature effects in the diversity of fish microbiota 140
4.5.3 Water temperature effects in the predicted microbiota function 142
4.6 Conclusion 143
4.7 Acknowledgements 143
4.8 References 143
4.9 Supplementary material 152
Chapter 5: Effects of disease and antibiotic treatment on the
microbiota of juvenile and adult seabass 163
Chapter 5.1: Effects of disease, antibiotic treatment and
recovery trajectory on the microbiome of farmed seabass
(Dicentrarchus labrax) 163

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream xiv
5.1.1 Abstract 163
5.1.2 Introduction 164
5.1.3 Material and methods 165
5.1.3.1 Ethical statement 165
5.1.3.2 Experimental design, sample collection and preparation 165
5.1.3.3 Data and statistical analyses 167
5.1.4 Results 167
5.1.4.1 Gill bacterial composition and diversity 168
5.1.4.2 Skin bacterial composition and diversity 174
5.1.5 Discussion 174
5.1.6 Conclusion 177
5.1.7 Acknowledgements 178
5.1.8 References 178
5.1.9 Supplementary material 187
Chapter 5.2: Monitoring disease and antibiotic treatment in the
skin microbiota of farmed seabass fingerlings 188
5.2.1 Abstract 188
5.2.2 Introduction 188
5.2.3 Material and methods 190
5.2.3.1 Experimental design, sampling and preparations 190
5.2.3.2 Data and statistical analysis 191
5.2.4 Results 192
5.2.4.1 Microbial diversity and composition 192
5.2.4.2 Microbial predicted function 195
5.2.5 Discussion 196
5.2.5.1 Disease effects in skin microbiota of seabass fingerlings: healthy
vs diseased states 196
5.2.5.2 Flumequine effects in skin microbiota of seabass fingerlings:
diseased vs treatment states 198
5.2.5.3 Recovery of the skin microbiota in seabass fingerlings: healthy
vs recovery states 199
5.2.6 Conclusion 199
5.2.7 Acknowledgements 200
5.2.8 References 200
5.2.9 Supplementary material 205

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream xv
Chapter 6: General discussion 208
6.1 Variability associated with the skin and gill microbiota in farmed seabass
and seabream 208
6.2 Host related factors shaping the skin and gill microbiota in farmed seabass
and seabream 210
6.3 Water related factors shaping the skin and gill microbiota in farmed seabass
and seabream 211
6.4 Dysbiosis in the skin and gill microbiota of farmed seabass and seabream 212
6.5 Microbial monitoring implications for aquaculture 214
6.6 Future perspectives 216
6.7 Conclusions 217
6.8 References 218
Appendix A: List of additional publications 227

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream xvi
List of Figures
Chapter 1 Figure 1.1 Pictures of the European seabass Dicentrarchus labrax and the gilthead seabream
Sparus aurata.
Chapter 2 Figure 2.1 Barplots of individual taxonomic composition.
Figure 2.2 Barplots of the core microbiotas.
Figure 2.3 Boxplots of the alpha-diversity values.
Figure 2.4 PCoA plots of the beta-diversity.
Figure S2.1 Rarefaction curves.
Chapter 3 Figure 3.1 Boxplots of the Shannon diversity values in each age group.
Figure 3.2 PCoA plots and boxplots of the Bray-Curtis distance in each age group.
Figure 3.3 Barplots of the most abundant phyla and genera in each age group.
Figure 3.4 LDA score of differently abundant predicted enriched pathways in each age group.
Figure 3.5 Venn diagrams of shared ASVs between tissues and water microbiota in each age
group.
Figure S3.1 Illustrative scheme of the semi-intensive fish farm.
Figure S3.2 Boxplots of the alpha-diversity values for tissue and water microbiota in each age
group.
Figure S3.3 Boxplots of the PD, ACE and Fisher alpha-diversity values in each age group.
Figure S3.4 PCoA plots of the Bray-Curtis distance in each tissue and water microbiota.
Chapter 4 Figure 4.1 Venn diagrams of shared core ASVs between tissues and water microbiota.
Figure 4.2 Alluvial plots of the most abundant potentially pathogenic (PP) genera, and other
PP genera they interact with, per month.
Figure 4.3 Temperature models, and beta- and alpha-diversity estimates, per month.
Figure 4.4 Barplots of microbiota contribution per month.
Figure 4.5 Graph depicting the relative frequency of the differentially enriched potential
pathways during cold and warm months.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream xvii
Figure S4.1 Daily average temperature of the water surface throughout the year.
Figure S4.2 Barplots of the most abundant phyla and genera per month.
Chapter 5 Figure 5.1.1 Schematic illustration of the experimental design.
Figure 5.1.2 Boxplots of the alpha-diversity values per state.
Figure 5.1.3 PCoA plots of the beta-diversity.
Figure 5.1.4 Alluvial plots of the most abundant taxa per state.
Figure 5.1.5 Barplots of the core microbiota per state.
Figure S5.1.1 Barplots of individual taxonomic composition.
Figure 5.2.1 Schematic illustration of the experimental design.
Figure 5.2.2 Alpha- and beta-diversity estimates, coloured by state.
Figure 5.2.3 Heatmaps of the most abundant phyla and genera per state.
Figure 5.2.4 Alluvial plots of potentially pathogenic taxa per state.
Figure 5.2.5 LDA score of differently abundant predicted enriched pathways per state.
Figure S5.2.1 Dendrograms of the beta-diversity values based on UniFrac weighted and Bray-
Curtis distances, coloured by state.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream xviii
List of Tables
Chapter 2 Table 2.1 Relative proportions of the most abundant ASVs, sequences and core ASVs.
Table 2.2 Significance of diversity indices and relative proportions of dominant taxa.
Chapter 3 Table 3.1 Significance of diversity indices between age groups.
Table 3.2 Significance of beta-diversity and Mantel tests between tissue and water microbiota.
Table S3.1 Relative proportions of the most abundant phyla and genera in each age group.
Table S3.2 Significance of relative proportions of dominant taxa between age groups.
Table S3.3 Significantly enriched predicted pathways in each age group.
Chapter 4 Table 4.1 Significance of the temperature models and season on the diversity indices.
Table 4.2 Significance of the correlation between temperature variables and bacterial
diversity, including the abundance of potentially pathogenic (PP) genera.
Table S4.1 Relative mean proportions of the most abundant phyla and genera per month.
Table S4.2 Core ASVs present in the skin, gill and water microbiota in a year.
Table S4.3 Significance of pairwise alpha- and beta-diversity comparisons across consecutive
months.
Table S4.4 Dynamics of the most abundant potentially pathogenic (PP) genera.
Table S4.5 Significantly enriched potential pathways in the cold and warm months.
Chapter 5 Table 5.1.1 Significance of diversity indices and relative proportions of dominant taxa.
Table 5.1.2 Most abundant ASVs and sequences.
Table 5.2.1 Significance of alpha- and beta-diversity indices between states.
Table S5.2.1 Relative mean proportions of the most abundant phyla and genera per state.
Table S5.2.2 Core ASVs present in each state.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream xix
List of Abbreviations
°C Degrees Celsius
ACE Abundance-based coverage estimator
ANOVA Analysis of variance
ASV Amplicon sequence variant
ATP Adenosine triphosphate
AXOS Arabinoxylooligosaccharides
bp base pair(s)
ca circa
cm centimeter(s)
CO2 Carbon dioxide
DADA Divise amplicon denoising algorithm
DNA Deoxyribonucleic acid
dpf days post feed
dph days post hatch
FEAST Fast expectation-maximization microbial source tracking
FOS Fructooligosaccharides
g gram(s)
GALT Gut-associated lymphoid tissue
GI Gastrointestinal
GIALT Gill-associated lymphoid tissue
gls Generalized least squares
GOS Galactooligosaccharides
h hour(s)
IMO Isomaltooligosaccharides
IMTA Integrated multi-trophic aquaculture
ITS Internal transcribed spacer
KEEG Kyoto encyclopedia of genes and genomes
Kg Kilogram(s)

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream xx
L Liter(s)
LDA Linear discriminant analysis
LEfSe Linear discriminant analysis effect size
lm linear models
LME Linear mixed effects
m meter(s)
MALT Mucosa associated lymphoid tissue
MAMP Microbe-associated molecular patterns
min minute(s)
mm milimeter(s)
MOS Mannanoligosaccharides
NAD Nicotinamide-Adenine dinucleotide
NALT Nasopharynx-associated lymphoid tissue
nm nanometer
NSTI Nearest sequenced taxon index
OTU Operational taxonomic unit
PCoA Principal coordinates analysis
PCR Polymerase chain reaction
PD Phylogenetic diversity
PERMANOVA Permutational multivariate analysis of variance
PICRUST Phylogenetic investigation of communities by reconstruction of unobserved states
PRR Pattern recognition receptors
QIIME Quantitative insights into microbial ecology
RAS Recirculating aquaculture systems
RNA Ribonucleic acid
rRNA Ribosomal ribonucleic acid
RRPP Randomized residuals in a permutation procedure
SALT Skin-associated lymphoid tissue
scFOS Short-chain fructooligosaccharides
TCA Tricarboxylic acid cycle
TIME Temporal insights into microbial ecology
tns tonne(s)
TOS Trans-galactooligosaccharides

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream xxi
USA United States of America
wph weeks post hatch
XOS Xylooligosaccharides
μm micrometre(s)

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
1
CHAPTER 1
General introduction
1.1 Literature review
1.1.1 Aquaculture overview Fish is currently one of the main sources of animal protein consumption for humans,
reaching 20.3 kg per capita per year (FAO, 2018). Fish are also an important source of several
essential amino acids, such as lysine and methionine; vitamins, such as vitamin D, A and B;
and minerals, such as calcium, phosphorus, iodine, zinc, iron and selenium (Béné et al., 2015).
Aquaculture accounts for almost half of the global fish supply and is currently the fastest
growing food-supply industry globally, with an estimated value of US$ 232 billion and a
production peak of 171 million tonnes in 2016, from which 80 million tonnes were finfish (FAO,
2018). In order to meet the global demand, arising from the exponential growth of the human
population, production practices are intensifying and ensuring food security is becoming
increasingly challenging (Béné et al., 2015; Fisher et al., 2017).
There are multiple forms of aquaculture in freshwater, brackish and marine
environments (FAO, 2018). Cages and net pens, frequent in marine environments and located
in natural water bodies, are key for rearing Atlantic salmon, the most common marine
aquaculture species (FAO, 2018). Inland aquaculture, usually practiced in freshwater
environments, represented 64.2% of the global farmed fish production, with finfish farming
representing 92.5% of the inland aquaculture in 2016 (FAO, 2018). In inland aquaculture, fish
are commonly grown in ponds, which are usually excavated and require a water supply from
an external source as well as drainage structures (Boyd and McNevin, 2015). Flow through
raceways is also used and is, in principle, similar to ponds, but can harbour higher stock
densities and are positioned in series with water flowing from one unit to another (Boyd and
McNevin, 2015; Soderberg, 1994). A more environmentally friendly method is Recirculating
Aquaculture Systems (RAS), which are closed culture systems that reuse waste water after it
has been filtered and purified (Boyd and McNevin, 2015). A more recent approach, Integrated
Multi-trophic Aquaculture (IMTA), rears multiple species from different trophic levels in a single
system in order to more efficiently take advantage of organic waste (Bouwmeester et al.,
2020). Moreover, aquaculture systems can be classified as extensive, semi-intensive,
intensive or hyper-intensive, with increasing amount of feed, stocking density and water
exchange from the first to the latter (Lane et al., 2014). All production technologies have

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
2
different profiles of environmental issues that are essential to ensure sustainable aquaculture
development (Lane et al., 2014).
Aquaculture conditions such as stocking densities, low oxygen concentrations, diet or
decreased water quality act as chemical and biological stressors for fish (Ashley, 2007). Stress
is a key factor influencing the health of farmed fish, impacting fish homeostasis by triggering
physiological responses and impairment of the immune system, which has direct
consequences on fish susceptibility to disease (Ashley, 2007; Sneddon et al., 2016). Infectious
diseases are the biggest constraint on aquaculture development, diminishing its economic
value and quality and reducing species growth and survival (FAO, 2018). Infectious agents
enter aquaculture mainly through water, or, to a less extent, through feed, broodstock or others
(Saksida et al., 2014). Although these diseases occur naturally in the wild, aquaculture
broodstock did not coevolve with diseases as wild hosts, which makes them less resistant in
general and makes disease consequences highly unpredictable (Lafferty et al., 2015).
Amongst the multitude of diseases that predominantly infect fish, the majority are attributed to
bacteria, followed by viruses, protists and metazoans (Lafferty et al., 2015). Bacterial diseases
are especially problematic for aquaculture, since bacteria can survive in the aquatic
environment without a host (Pridgeon and Klesius, 2012). Several bacterial genera can be
pathogenic to aquatic animals, mainly the gram-negative Aeromonas, Edwardsiella,
Flavobacterium, Francisella, Photobacterium, Piscirickettsia, Pseudomonas, Tenacibaculum,
Vibrio and Yersinia, and the gram-positive Lactococcus, Renibacterium and Streptococcus
(Austin and Austin, 2016; Toranzo et al., 2005). The pathogenicity and clinical signs caused
by bacterial pathogens are dependent on the stage of the disease and on several host
characteristics, such as species and age group (Toranzo et al., 2005). For example, fish early
life-stages are the most vulnerable to stress and disease, due to a lack of fully functional
immune, digestive and osmotic systems (Rehman et al., 2017). Moreover, certain
environmental variables, such as temperature, photoperiod or precipitation are well known to
affect the physiology and behaviour of animals (Bowden et al., 2007). Especially important in
the aquatic environment, these factors govern the dynamics of many infectious diseases (e.g.,
Mohamad et al., 2019). Hence, perturbations to the host-pathogen-environment balance will
result in disease (Toranzo et al., 2005). In order to develop appropriate measures that can
prevent and control marine diseases, studies must integrate the characteristics of host species
and etiological agents of disease as well as the environmental factors that affect them
(Toranzo et al., 2005).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
3
1.1.1.1 European seabass Dicentrarchus labrax and gilthead
seabream Sparus aurata aquaculture The European seabass Dicentrarchus labrax is a fish species from the Moronidae
family with a silvery grey and rather elongated body (Figure 1.1A). The seabass is a slow
growing species with males maturing early between years 2 and 3, while females mature after
3 years (Carillo et al., 1995; Felip et al., 2001). Being an eurytherm (5-28°C) and euryhaline
(3-35‰) species, it mostly lives in coastal marine waters as well as in brackish waters
estuaries and lagoons and can occasionally be found in rivers (FAO, 2016a). Up until the late
60s, the European seabass was typically cultured in coastal lagoons and tidal reservoirs, after
which it started to be massively reared in France and Italy (FAO, 2016a). By the late 70s,
mass-production of the European seabass was implemented across the Mediterranean, being
the first non-salmonid species to be commercially cultured in Europe (FAO, 2016a). Currently,
seabass is farmed in open-water sea cages although production in seawater ponds and
lagoons across the Mediterranean is still very common with a global production of 191,003
tonnes in 2016 (FAO, 2016a).
The gilthead seabream Sparus aurata is a species from the Sparidae family with an
oval, deep and compressed body, and silvery grey colour (Figure 1.1B). The seabream is a
protandrous hermaphrodite, first maturing as male between years 1 and 2 and then as female
in the following 2 to 3 years (FAO, 2016b; Mehanna, 2007). It is also an euryhaline (2-39‰)
and eurytherm (4-26°C) species, being found in marine and brackish waters, for example
coastal lagoons and estuaries (Chervinski, 1984; FAO, 2016b; Kır, 2020). Historically, the
gilthead seabream was extensively cultured in coastal lagoons and saltwater ponds (FAO,
2016b). After the 1980s, when intensive rearing systems were developed and artificial
breeding became successful, this species achieved large-scale production in Spain, Italy and
Greece (FAO, 2016b). Currently it is mostly farmed in coastal ponds and lagoons or in sea
cages and land-based installations across the Mediterranean, with global productions of
185,980 tonnes in 2016 (FAO, 2016b).
Figure 1.1 Seabass (A) and seabream (B). After FAO 2016a, 2016b.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
4
Among the 369 finfish species farmed in the world, the European seabass and the
gilthead seabream are considered high-value species, being two of the most important farmed
finfish species traded in Europe (Clarke and Bostock, 2017). Their production, however, is
greatly affected by infectious diseases that account for losses of up to 15% in seabass and of
up to 40% in seabream (Lane et al., 2014). The main bacterial pathogens to which seabass
and seabream are susceptible include Photobacterium damselae, causing photobacteriosis,
Vibrio spp., causing vibriosis, and Tenacibaculum maritimum, causing flexibacteriosis (FAO,
2016a, 2016b; Toranzo et al., 2005). Given the unavoidable constant interaction with aquatic
potential pathogens and the role of the seabass and seabream in the global food economy,
studying their mucosal immune system will help design more efficient prevention and
treatment strategies.
1.1.2 Microbiome More than 300 years ago, Antoine van Leeuwenhoek observed the first host-
associated microbial communities that he scraped from his own teeth and called “animalcules”
(Robinson et al., 2010). Many others followed in the field of microbiology, passing by Pasteur’s
concept of culturing isolated bacteria, Koch’s determination of infectious diseases etiology, to
Staley and Konopka’s “great plate anomaly”, that was based on the observation that most
microbes seen under the microscope could not be cultured (Robinson et al., 2010; Staley and
Konopka, 1985). After several definitions, Berg et al. (2020) recently described the microbiome
as the community of organisms in an environment, including their produced molecules,
structural elements (such as nucleic acids, proteins, lipids and polysaccharides), metabolites,
and molecules produced by coexisting hosts and environmental conditions. All living
microorganisms within the microbiome, namely bacteria, archaea, fungi, algae and small
protists, form the microbiota (Berg et al., 2020). Although many technological and research
advances in the field were made, we are still far from completely grasping the magnitude and
role of the microbial communities associated with any host or environment (Gilbert et al., 2018;
Legrand et al., 2018; Robinson et al., 2010). Nevertheless, the importance of the host-
associated microbial communities to the host health is undeniable (Gilbert et al., 2018;
Legrand et al., 2018; Robinson et al., 2010).
Microbes are one of the major reservoirs of diversity on Earth and have been evolving
for roughly 3.5 billion years (McFall-Ngai et al., 2005). Microbes show an assortment of
lifestyles, being able to live in symbiosis with animals (Smith and Douglas, 1987). Mutualistic
interactions between animals and microbes are one of the most important forms of symbiosis,
having significant ecological and evolutionary effects (McFall-Ngai et al., 2005). The
microbiota of vertebrates has co-evolved with their hosts for almost half a billion years and is

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
5
now considered the “extended self” (Lloyd-Price et al., 2016). At homeostasis, this symbiotic
relationship provides mutual benefits. While the microbiota benefits from a safe growing
environment, the host is provided with immune, physiologic and metabolic aid (Gilbert et al.,
2018; Martin et al., 2007; Maynard et al., 2012).
The microbiome is one of the most important components of the vertebrate’s immune
system. All higher vertebrates have a complex immune system composed of the innate and
adaptive immune systems (Medzhitov and Janeway, 1998). While the innate immune system
is enrolled in an early response during infection, the adaptive immune system is activated in
the late phase of infection, eliminating pathogens and generating immunological memory
(Mogensen, 2009). The innate immune system is based primarily on physical and chemical
barriers to infection, including the epidermis, ciliated respiratory epithelium, vascular
endothelium, and the mucosal surfaces (Basset et al., 2003). The mucosal surfaces of animals
are part of the mucosal immune system and incorporate the mucosa associated lymphoid
tissues (MALTs), responsible for both symbiont colonization tolerance and protection against
pathogens (Gomez et al., 2013). Within the MALT there is a complex and diverse bacterial
community, the microbiota, that is involved in the mucosal immune system development and
functioning (Salinas and Magadán, 2017). Specifically, the microbiota is the first line of
defense against pathogens, by occupying potential binding sites or by secreting inhibitory
compounds (Basset et al., 2003). In this regard, studying the microbiome is crucial to
understand the microbe-host-pathogen interplays and holds promise to treat and prevent
disease. Besides providing immune benefits, the microbiome also has the ability to increase
the host’s capacity to harvest energy from its diet (e.g., Heintz-Buschart & Wilmes, 2018;
LeBlanc et al., 2013; Turnbaugh et al., 2006), affect lipid metabolism by modifying bile acid
(Martin et al., 2007), repair damaged mucosal barriers (Pull et al., 2005), alter host gene
expression encoding antimicrobial compounds (Cash et al., 2006), increase locomotor activity
(Bäckhed et al., 2007), educate skin T cells to respond to pathogens (Grice and Segre, 2011),
charge nucleotides and synthetize ATP (Huttenhower et al., 2012), prevent overgrowth of
harmful bacteria (Kamada et al., 2013), and maintain a homeostatic pH (Hickey et al., 2012),
just to name a few. Moreover, it is now known that the gut microbiome communicates with the
central nervous system participating in the “gut-brain axis” through microbial-derived
intermediates that either enter the systemic circulation and cross the blood-brain barrier or
interact directly with the immune system to spread signaling (Osadchiy et al., 2019). Insights
into the specific functional contributions of the microbiome opens the door for the use of
specific commensal microbes as therapy (Hall and Robinson, 2021; Mimee et al., 2016; Park
and Im, 2020).
There are several biotic factors that shape the microbiota, including host genotype and
physiology, body site and lifestyle, including diet, as well as abiotic factors, such as

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
6
temperature and pH (Gilbert et al., 2018; Martin et al., 2007a). The composition and diversity
of the microbiome is based on the microbes available at the time of colonization and the order
in which they colonize, together with the specific characteristics of the colonization habitat
(Martin et al., 2007a). For example, human fetuses do not have a microbiota; consequently,
mode of birth is essential in defining the newborn’s first microbiome, with neonates born
vaginally presenting a different microbiota from those born via Cesarean section (Dominguez-
Bello et al., 2010). Moreover, because the conditions that allow microbial colonization in one
body habitat are not encountered in others, the microbiota will manifest differently in different
ecological niches (Proctor et al., 2019). The human gut microbiota, for example, is significantly
different from the microbiota of the skin, which, in turn, also presents differences depending
on the body site (Costello et al., 2009). Additionally, it is thought that the human microbiome
is highly plastic over time (Caporaso et al., 2011; David et al., 2014), with a great level of
personalization (Flores et al., 2014), which vastly exceeds that of the human genome, to the
point that it might have forensic applications (Fierer et al., 2010; Robinson et al., 2021).
Despite these differences, it is possible to detect certain similarities within habitats or species.
The set of genes that is present across all individuals of a species or habitat is called the core
microbiome (Martin et al., 2007). In sum, the microbiome is composed of the core and the
variable or transient microbiome and is dependent on a wide range of factors (Martin et al.,
2007). Some specific life events that occur on longer time frames, such as aging (Heintz and
Mair, 2014), sexual maturation (Kundu et al., 2017) or reproduction (Koren et al., 2012), also
alter the microbiota and consequently animal-microbial relationships, leading to a state of
“altered symbiosis”.
Changes in conditions can alter microbial composition and diversity switching from
homeostasis to microbial imbalance, i.e., dysbiosis, which impacts the host’s health and
eventually leads to disease (Petersen and Round, 2014). Dysbiosis is the breakdown in the
symbiotic relationship, generally related to one or more major stressors, such as pollutants
(e.g., Jin et al., 2017), infection (e.g., Llewellyn et al., 2017), toxins (e.g., Tsiaoussis et al.,
2019) or climate change (e.g., Greenspan et al., 2020). Dysbiosis can occur through
pathobiont expansion, reduced microbial diversity, loss of beneficial microbes, or a
combination of any of these three events, all of which are linked to disease (Petersen and
Round, 2014). For example, common perturbations to the microbiome are related to obesity
(e.g., Fei & Zhao, 2013), diabetes (e.g., Zhou et al., 2019), inflammatory bowel diseases,
including Crohn’s disease (e.g., Erickson et al., 2012; Frank et al., 2011; Lloyd-Price et al.,
2019), disruption of the host circadian cycle (e.g., Leone et al., 2015), rheumatoid arthritis
(e.g., Zhang et al., 2015), atopic dermatitis (e.g., Kong et al., 2012), psoriasis (e.g.,
Alekseyenko et al., 2013), acne (e.g., Dessinioti & Katsambas, 2010), cardiovascular disease
(e.g., Peng et al., 2018), multiple sclerosis (e.g., Jangi et al., 2016), asthma (e.g., Sokolowska

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
7
et al., 2018), autism (e.g., Hsiao et al., 2013), Parkinson’s disease (e.g., Sampson et al.,
2016), Alzheimer’s disease (e.g., Jiang et al., 2017), cancer (e.g., Gopalakrishnan et al.,
2018), among many others. To this end, microbiome research is imperative and crucial to
provide new biomarkers of disease (Gilbert et al., 2018). In fact, it has been suggested that
changes in the microbiota can predict disease progression faster and more easily than clinical
symptoms or the presence of pathogenic agents (Woodhams et al., 2020). Although research
has mainly focused on the human microbiome, increasingly more studies have extended the
focus towards other taxa, to the point where the health of the entire Earth’s living organisms
and environments is considered to be dependent on the “smaller majority” of ubiquitous
microorganisms (Blaser et al., 2016).
1.1.3 Microbiome of fish The mucosal immune system of teleost species is a persistent vigilant of the
environment, protecting the host from infection (Salinas and Magadán, 2017). Fish incorporate
four different MALTs, namely the gut-associated lymphoid tissue (GALT), the gill-associated
lymphoid tissue (GIALT), the skin-associated lymphoid tissue (SALT), and the nasopharynx-
associated lymphoid tissue (NALT) (Salinas and Magadán, 2017). A key element of all teleost
MALTs is the mucosal layer, a perfect niche for microbial adherence and growth composed of
mucins, ions and lipids (Gomez et al., 2013). The mucus layer covers the gut, gill, skin and
nasal mucosal surfaces of fish and contains immunological molecules that interact directly
with the commensal microbial communities (Kelly and Salinas, 2017). The microbiome of fish
is thus considered as the first line of defense against pathogens and is likely shaped by the
physicochemical properties of the mucus layer (Kelly and Salinas, 2017).
Being constantly immersed in the aquatic environment, fish are endlessly exposed to
planktonic microbiota that are in contact with their mucosal surfaces (Salinas, 2015). Although
the microbiota of fish is highly diverse, some bacterial taxa dominate across tissues and
species, in particular those from the Phylum Proteobacteria (Legrand et al., 2020b; Llewellyn
et al., 2014). Additionally, the phyla Actinobacteria, Bacteroidetes, Firmicutes and
Fusobacteria are also dominant in the fish microbiota (Legrand et al., 2020b; Llewellyn et al.,
2014). As in other vertebrates, the microbiota of fish is composed of both resident and
transient microbes (Legrand et al., 2020b). While the resident bacteria occur within the
mucosa and are intimately associated with host cells, the transient communities are mostly
free-living bacteria that are part of the fish microbiota for short periods of time (Banerjee and
Kumar Ray, 2017). Resident microbiota is mostly regulated by host factors, while transient
bacteria are mainly influenced by environmental factors (Legrand et al., 2020b). While
perturbations can change microbial composition, it has been proposed that functionally

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
8
redundant members have the ability to recolonize and preserve microbial functionality (Kelly
and Salinas, 2017). On the other hand, ubiquitous pathogenic bacteria naturally colonize the
water (e.g., Martins et al., 2018) and can also integrate the microbial community of healthy
fish (e.g., Givens et al., 2015). Several stressors can alter the properties of the mucosae (Kelly
and Salinas, 2017), leading to a shift in the abundance of these pathogens, inducing
pathogenicity and leading to dysbiosis and disease (e.g., Hess et al., 2015; Legrand et al.,
2018). In addition, there is no complete distinction between symbionts and pathogens and
high abundances of certain commensals can trigger inflammatory responses (Kelly and
Salinas, 2017).
Bacteria from skin, gills and specially the gastrointestinal (GI) tract have been the focus
of research on fish microbiota, given their role in fish immunity and because these tissues are
the major pathways of pathogen entrance in fish (Merrifield and Rodiles, 2015). Fish microbial
immune response is primarily accomplished through competition between commensals and
potential pathogens for adhesion and nutrients, or through production of secondary
metabolites by the commensal community, such as antimicrobial compounds, that antagonize
potential pathogens (Kelly and Salinas, 2017; Merrifield and Rodiles, 2015; Trivedi, 2012).
The microbiota of fish eggs and larval stages (i.e., whole larvae) has also gained attention
given the importance of the initial steps of microbial colonization (e.g., Abdul Razak et al.,
2019; Bledsoe et al., 2016). Studying the microbiome of fish and their environment will
hopefully promote sustainable aquaculture by preventing and combating pathogens, boosting
host immune response and increasing water quality (Legrand et al., 2020b).
1.1.3.1 Fish microbial dynamics In order to understand microbe-host relationships, research must investigate factors
influencing these dynamics. Such factors could be the result of random dispersal of microbes,
that likely explain the high inter-individual variation presented by fish (Burns et al., 2017); or
of host and environmental selective pressures (Talwar et al., 2018).
Ontogeny is a fundamental and inevitable factor shaping microbial diversity. Fish egg
surfaces are colonized by microbes in the surrounding water (Abdul Razak et al., 2019). As
larvae hatch, these microbes further colonize mucosal tissues, including the GI tract through
water and feed ingestion (e.g., Wilkes Walburn et al., 2019). Until adulthood, fish microbiota
undergoes continuous developmental changes that are stage related. For example, the gut
microbiota of yellowtail kingfish experiences significant changes between 0- and 53-days post
hatch (dph), with significant differences in diversity even between 0- and 3-dph (Wilkes
Walburn et al., 2019). Similarly, the gut microbiota of catfish is also significantly different
between 3-, 65- and 125-dph, but stabilized between 125- and 193-dph (Bledsoe et al., 2016).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
9
On the other hand, while the gut microbiota of seabream significantly changes between 5- and
15-dph, no further differences were found between groups up until 71-dph, when the study
ended (Nikouli et al., 2019). A longer term study assessing the gut microbiota of the grass
carp, Chinese perch and Chinese largemouth catfish revealed diversity differences between
all life stages, including larvae from different phases, juveniles and adults (Yan et al., 2016).
On the contrary, the gut microbiota of zebrafish showed no differences between several stages
between 0- and 380-days post feed (dpf), except when fish underwent sexual maturation
between 35- and 75-dpf (Stephens et al., 2016). Lokesh et al. (2017) studied the Atlantic
salmon gut microbiota for 80-weeks post hatch (wph), including several early developmental,
juvenile and adult stages. Differences were found in diversity between all life cycles, with
marked shifts occurring concomitantly with major life events, including hatching and transition
between freshwater and seawater (Lokesh et al., 2019). Additionally, cross-sectional studies
also reported differences in diversity between the gut microbiota of pre-settlement larvae and
post-settlement juveniles/adults of damselfish and cardinalfish (Parris et al., 2016), and
between the skin microbiota of juveniles and adults of two damselfish species (Xavier et al.,
2020). In summary, ontogenetic studies have so far revealed that fish microbiota is highly
dynamic, diversifying with age, and that early life exposure is critical to ensure adult health
and nutrition (Legrand et al., 2020b; Llewellyn et al., 2014). It has been suggested that
microbial colonization is determined by neutral processes in the early life stages, becoming
progressively governed by deterministic processes with age (Talwar et al., 2018). The vast
majority of these studies focused on the gut microbiota and were either cross-sectional or
based on a short time window. Nevertheless, age related microbial differences in fish seems
to be many times linked to important life events (e.g., diet transitions, Wilkes Walburn et al.,
2019; environmental transitions, Lokesh et al., 2019; sexual differentiation, Stephens et al.,
2016) suggesting a compound effect.
In an early study trying to correlate fish microbiota and host genetics, Rawls et al.
(2006) performed reciprocal gut microbial transplants between zebrafish and mice. The results
revealed that, after transplant, although composition was similar to that of the original host,
the abundance of each lineage was more similar to the recipient host (Rawls et al., 2006).
Recently, a study comparing the gill and intestine microbiota of 53 fish reef species (belonging
to 15 families) showed that microbiota composition varied significantly according to host family
(Pratte et al., 2018). Moreover, the authors also saw that similarity between tissues within an
individual was significantly greater than that in the same tissue from different individuals
(Pratte et al., 2018). Other studies also reported microbial differences between species
sharing the same habitat (e.g., 2 fish species reared in the same pond, Li et al., 2014; 2 guppy
ecotypes from the same stream, Sullam et al., 2015; 4 butterflyfish species from the island of
Moorea, Reverter et al., 2017; 44 coral reef species from Mayotte island, Chiarello et al., 2018;

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
10
13 species within the Mediterranean Sea, Ruiz-Rodríguez et al., 2020; 3 species from
Amazonas, Sylvain et al., 2020). On the other hand, a recent study evaluating the gut
microbiota of blue and channel catfish reported significant differences in structure but not
diversity across fish species or strains (Bledsoe et al., 2018), indicating that host factors,
although predominant, are not straightforward. These results strongly attest for a role of the
host genetics in microbiota assembly and diversification in fish, although the specific
mechanisms through which that happens are still largely unknown.
Diet, or specific diet content, is one of the most straightforward host factors impacting
the gut microbiota, and it can also impact the skin and gill microbiota (Chiarello et al., 2018;
Pratte et al., 2018). Lipids, proteins, vitamins, functional glycomic ingredients and minerals are
linked to gut microbial health that ultimately impact fish metabolism and energy utilization,
performance and immunity (reviewed in Ringø et al., 2016). Several studies have
demonstrated the effect of diet in shaping fish gut microbiota, including the gibel carp (e.g.,
Chen et al., 2014), common carp (e.g., Liu et al., 2014), surgeon fish (e.g., Miyake et al.,
2015), zebrafish (e.g., Koo et al., 2017), gilthead seabream (e.g., Rimoldi et al., 2018),
European seabass (e.g., Pérez-Pascual et al., 2020), rainbow trout (e.g., Michl et al., 2017),
brown trout (e.g., Michl et al., 2019), Atlantic salmon (e.g., Villasante et al., 2019), Nile tilapia
(e.g., Hassaan et al., 2020), among other fish species (e.g., Bolnick et al., 2014). Differences
in microbial composition and diversity in fish gut are mostly related to the nutrient content of
the diet of each fish species, coupled with gut morphology and host phylogeny (Legrand et al.,
2020b). Specifically, gut-colonizing microbes are highly dependent on species-specific
characteristics such as gut size and structure, pH, osmolality, redox potential, as well as diet
content (Merrifield and Ringø, 2014). In this regard, unique bacterial taxa are found in the gut
microbiota of fish with different feeding habits, namely herbivorous, carnivorous, omnivorous
and filter-feeding species (e.g., Li et al., 2014; Liu et al., 2016; Pratte et al., 2018). There is
thus a causal link between fish diet, gut microbiota and fish health. Importantly in aquaculture,
diets fed to farmed fish have negative impacts on gut health, having major consequences in
fish fecundity, growth rate, appetite and susceptibility to disease (Hossain et al., 2018). In this
regard, recent research has focused on gut microbial manipulation by altering or adding
supplements to fish diet (Ringø et al., 2016). An increasing body of literature has exposed the
benefits of probiotics, prebiotics, immunostimulants, medical plant extracts and microalgae in
controlling the microbiota and optimizing fish health in aquaculture (reviewed in Ringø et al.,
2014). Feed supplementation is a central strategy to improve diet quality and thus growth, but
also to mitigate the negative impacts of aquaculture practices, with important outcomes for
health and productivity of fish farms (Egerton et al., 2018; Legrand et al., 2020b; Reverter et
al., 2021).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
11
Teleost fish present microbial differences across, as well as within, body habitats.
Studies so far have confirmed differences between the microbiota of distinct tissues in several
fish species, including the rainbow trout (skin, gill, gut and olfactory organ, Lowrey et al., 2015),
Atlantic salmon (gill and intestine, Schmidt et al., 2016), yellowtail kingfish (skin, gill and gut,
Horlick et al., 2020; Legrand et al., 2020a, 2018), grass carp and southern catfish (intestine,
skin and gill, Zhang et al., 2019), rabbitfish (skin, gill, stomach and hindgut, Wu et al., 2020),
among several other fish species (gill and intestines, Pratte et al., 2018; skin, gill and
intestines, Ruiz-Rodríguez et al., 2020; skin and gut, Sylvain et al., 2020; liver and kidney,
Meron et al., 2020). In addition, microbial differences within the same tissue were seen in the
skin of the European seabass and gilthead seabream (Chiarello et al., 2015) and along the
intestine of the grass carp (Yang et al., 2019), rabbitfish (Jones et al., 2018), Asian silver carp
and gizzard shad (Ye et al., 2014). These variations reflect the diverse microbial colonization
patterns due to differences in physicochemical properties of different tissues, such as pH,
oxygen content or nutrient availability (Egerton et al., 2018; Wang et al., 2018). Moreover,
different body niches exert different pressures, for example comprising different MALTs,
attesting for microbial specialization in each tissue (Kelly and Salinas, 2017).
Many studies have reported higher bacterial diversity in water compared to fish skin
(Chiarello et al., 2019, 2018, 2015; Larsen et al., 2015a; Uren Webster et al., 2018; Wu et al.,
2020; Zhang et al., 2019), gills (Pratte et al., 2018; Wu et al., 2020), intestine (Bledsoe et al.,
2016; Parris et al., 2016; Reinhart et al., 2019; Wilkes Walburn et al., 2019; Wu et al., 2020;
Zhang et al., 2018), stomach (Wu et al., 2020) and whole larvae (Nikouli et al., 2019). The
vast majority of studies reported significant differences between the mucosal microbiota of fish
and the water microbial community (Bledsoe et al., 2016; Boutin et al., 2013; Carlson et al.,
2017; Chiarello et al., 2019, 2018, 2015; Larsen et al., 2015a; Legrand et al., 2018; Llewellyn
et al., 2016; Nikouli et al., 2019; Parris et al., 2016; Pratte et al., 2018; Schmidt et al., 2018;
Uren Webster et al., 2018; Wu et al., 2020; Yan et al., 2016; Zhang et al., 2018, 2019).
Nevertheless, compositional similarities, i.e., shared operational taxonomic units (OTUs) or
amplicon sequence variants (ASVs), are sometimes reported between fish and water
microbiota (Bledsoe et al., 2016; Boutin et al., 2013; Llewellyn et al., 2016; Nikouli et al., 2019;
Parris et al., 2016; Pratte et al., 2018; Schmidt et al., 2015; Uren Webster et al., 2018; Wilkes
Walburn et al., 2019; Zhang et al., 2018). This suggests that each body niche selects from a
nested subset of the water microbial communities (Pratte et al., 2018).
Together with host related factors, habitat or season also play an important role in fish
microbial dynamics. Fish skin, gill and gut microbial diversity and structure vary in association
with multiple ecological variables related to specific habitat characteristics, including
geographic distance (Dunn et al., 2020; Pimentel et al., 2017; Sylvain et al., 2020; Yildirimer
and Brown, 2018), habitat type (Jones et al., 2018; Smith et al., 2015; Uren Webster et al.,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
12
2018; Xavier et al., 2019) or lake morphology (Smith et al., 2015). There are exceptions
though, for example, the gut microbiota diversity and structure of the European seabass and
gilthead seabream reared in 5 distantly located aquaculture sea cages revealed no influence
of geographic location (Nikouli et al., 2018). Additionally, it has been suggested that the gut
microbiota of some fish species is mostly modulated by host factors (Steury et al., 2019;
Sylvain et al., 2020). Seasonal variation of water temperature characteristics, such as
temperature, pH, ionic composition, dissolved oxygen, conductivity or chlorophyll can also
cause differences in the microbial diversity of fish sampled across different habitats and
seasons (Hovda et al., 2012; Krotman et al., 2020; Minich et al., 2020a; Ornelas-García et al.,
2018; Sylvain et al., 2020; Vasemägi et al., 2017; Zarkasi et al., 2014). Furthermore, microbial
composition seems to reflect fish tolerance to temperature changes (Kokou et al., 2018).
Importantly, microbial differences due to environmental factors could reflect the interaction of
several covarying variables such as habitat and feeding habits (Smith et al., 2015), or even
social behaviour (Xavier et al., 2019).
Fish microbial variation between habitats can be attributed to differential exposure to
microbes or to different evolutive ecological pressures (Smith et al., 2015; Sylvain et al., 2020).
Additionally, microbial changes in fish due to habitat transition were described, including the
gut and skin of Atlantic salmon migrating from freshwater to seawater (Llewellyn et al., 2016;
Lokesh and Kiron, 2016; Rudi et al., 2017), the skin of hatchery-reared common snook
acclimating to the wild (Tarnecki et al., 2019), and the entire microbiota of the Molly
experimentally acclimated to different salinities (Schmidt et al., 2015). Such important events
in the life cycle of many fish species require them to undergo changes in physiology,
geography and diet, ultimately exposing them to different microbial communities from the
surrounding environment (Le and Wang, 2020). A deeper understanding of the role of the
environment in fish microbial dynamics could have important implications for host adaptation
to local selective pressures.
1.1.3.2 Fish microbial imbalance and disease In fish, microbial imbalance (dysbiosis) resulting from changes in the environment
caused by chemicals, antibiotic usage or aquaculture practices can lead to disease (de Bruijn
et al., 2018). In fish farms, several factors, most commonly related to water parameters, cause
stress to fish populations; in particular, hypoxia (Boutin et al., 2013), high ammonia
concentrations (Qi et al., 2017), suboptimal pH (Sylvain et al., 2016) or unfavourable salinity
(Tian et al., 2020; Zhang et al., 2016) can lead to microbial imbalance and susceptibility to
disease. Other fish farm practices also cause microbial imbalances, such as netting and
transfer (Minniti et al., 2017; Tacchi et al., 2015), rearing density (Du et al., 2019; Y. Wang et

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
13
al., 2019) or starvation (Tran et al., 2018a; Xia et al., 2014). In the majority of the cases,
exposure to such stressors leads to an increase in pathobionts or a decrease in beneficial
microbiota, confirming dysbiosis (Boutin et al., 2013; Minniti et al., 2017; Qi et al., 2017; Tran
et al., 2018a; Y. Wang et al., 2019; Zhang et al., 2016). Importantly, differences in microbial
diversity have been found between farmed and wild species, such as in the gut of fine flounder
(Ramírez and Romero, 2017a), gut, skin and gills of yellowtail kingfish (Legrand et al., 2018;
Ramírez and Romero, 2017b), and the gut of Atlantic salmon (Uren Webster et al., 2018).
Moreover, a study showed that selective breeding for resistant farmed fish species alters gut
but not gill microbiota (Brown et al., 2019). However, whether such changes are linked to
increased susceptibility to disease remains unknown. Cultivation likely exerts pressure on
host-microbe interactions, although more comparative studies are needed to fully understand
this interaction. Still, there is an urgent need to mitigate aquaculture negative impacts on fish
health and studying their microbial communities is proving to answer many questions.
The link between infection and microbial imbalance is undeniable. Many studies have
established a relationship between specific pathogens and dysbiosis in fish. For example, the
impact on microbial diversity was previously described for the skin of Atlantic salmon infected
with Lepeophtheirus salmonis (see Llewellyn et al., 2017) and alphavirus (see Reid et al.,
2017), the gut of brown trout infected with Tetracapsuloides bryosalmonae (see Vasemägi et
al., 2017), the gut of gibel carp infected with herpesvirus (see She et al., 2017), the skin of
rainbow trout infected with Ichthyophthirius multifilis (see Zhang et al., 2018), the gut, skin,
kidney and brain of Asian seabass infected with Tenacibaculum singaporense (see Miyake et
al., 2020), the gut of grass carp with enteric infection (see Tran et al., 2018b), the gut and
stomach of rainbow trout infected with Caligus lacustri (see Parshukov et al., 2019), the buccal
mucosa of the rainbow trout infected with hematopoietic necrosis virus (see Dong et al., 2019),
and the skin of orbicular batfish infected with Tenacibaculum maritimum (see Le Luyer et al.,
2021). Given the serious impact of diseases in aquaculture, studying the effects of infection in
fish microbiota proves useful for understanding pathogen abundance dynamics as well as
providing health status biomarkers (e.g., Legrand et al., 2018; Llewellyn et al., 2017; Reid et
al., 2017; She et al., 2017; Zhang et al., 2018). Additionally, dysbiosis due to disease opens
the door for secondary infections by opportunistic bacteria (e.g., Dong et al., 2019; Llewellyn
et al., 2017; Reid et al., 2017; Zhang et al., 2018). Importantly, infection causing disease in a
specific organ can lead to dysbiosis in other tissues (Legrand et al., 2018). It is, thus,
imperative that host-microbiome-pathogen studies incorporate broader approaches, including
several fish tissues as well as environmental factors.
One of the most important environmental characteristics that is related to microbial
imbalance and disease is water temperature. All bacteria, including ubiquitous pathogens,
have optimal temperature ranges that maximize their growth (Corkrey et al., 2012).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
14
Temperature shifts can confer pathogens competitive advantages and create colonization
opportunities (Mouquet et al., 2005). Indeed, positive relationships between temperature and
the abundance of potentially pathogenic genera, have been reported for Vibrio and
Flavobacterium (see Sugita et al., 1989), as well as Photobacterium (see Horlick et al., 2020;
Minich et al., 2020a). Lower temperatures are also correlated with higher incidences of severe
outbreaks of Aliivibrio (e.g., Guijarro et al., 2015; Khider et al., 2018) and Pseudomonas (e.g.,
Huang et al., 2019; Tao et al., 2016). Seasonal distributions of bacterial infections are common
in fish (Bellos et al., 2015; Habiba et al., 2015; Schade et al., 2016), including within
aquaculture settings (Baker-Austin et al., 2013; Eissa et al., 2018; Kayansamruaj et al., 2014;
Matanza and Osorio, 2018).
The most common way to treat diseases in aquaculture is through pharmacological
interventions. Vaccination is one approach, although procedures are costly, cause substantial
stress and most only confer short term immunity (Bakopoulos et al., 2003). Antibiotic treatment
remains the best option, even being sometimes used in a prophylactic manner (Cabello,
2006). Despite their reported negative impacts on animals, it is estimated that antimicrobial
use in food producing animals surpasses human use (Van Boeckel et al., 2017). Several
studies have evaluated the impact on fish microbial balance of several antibiotics, including
rifampicin (Carlson et al., 2017, 2015), olaquindox (He et al., 2017b), streptomycin (Pindling
et al., 2018), sulfamethoxazole (Zhou et al., 2018), florfenicol (E. Wang et al., 2019; Gupta et
al., 2019; Sáenz et al., 2019), oxytetracycline (Almeida et al., 2019b, 2019a; Kim et al., 2019;
López Nadal et al., 2018; Navarrete et al., 2008; Zhou et al., 2018), oxolinic acid (Gupta et al.,
2019), amoxicillin (Kim et al., 2019), and a combination treatment of oxytetracycline,
erythromycin and metronidazole (Legrand et al., 2020a). The vast majority of these studies
also report the enrichment of opportunistic pathogens linked to an increased susceptibility to
secondary infections (Almeida et al., 2019a; Carlson et al., 2017, 2015; He et al., 2017b; Kim
et al., 2019; Legrand et al., 2020a; Navarrete et al., 2008; Sáenz et al., 2019; E. Wang et al.,
2019). Importantly, persistent antibiotic treatment is driving antimicrobial resistance in fish
pathogens (Miller and Harbottle, 2018) and commensal microbes (Hong et al., 2018). For
example, several strains of Photobacterium damselae showed increased resistance to
tetracycline and streptomycin both in wild and farmed fish (Abdel-Aziz et al., 2013; Chiu et al.,
2013; Essam et al., 2016). Additionally, it has been suggested that antibiotic oral
administration increases mobile genetic element exchange of antibiotic resistant genes in fish,
enriching their route of dispersion in aquaculture systems (Sáenz et al., 2019). Thus,
aquaculture environments, including fish, water and biofilms can become reservoirs for
antibiotic resistant genes, posing elevated human, animal and ecosystem threat (Schar et al.,
2020).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
15
Several studies reported shifts in microbial composition and diversity caused by
environmental contaminants, including biocides commonly used in aquaculture (potassium
permanganate, Mohammed and Arias, 2015; triclosan, Gaulke et al., 2016; Narrowe et al.,
2015; copper sulfate, Tarnecki et al., 2021), hydrocarbons from crude oil (e.g., Bagi et al.,
2018; DeBofsky et al., 2020), lead (e.g., Zhang et al., 2021), agriculture pesticides (e.g., X.
Wang et al., 2019), microplastics (e.g., Jin et al., 2018; Lu et al., 2019), and nanomaterials
(e.g., titanium dioxide, Chen et al., 2018; molybdenum oxide, Aleshina et al., 2020). Of special
concern, nanomaterials are particles of dimensions between 1 and 100 nm and widely used
in both industrial and consumer products, which inevitably leads to their release to the aquatic
environment (Piccinno et al., 2012; Weir et al., 2012). Several types of nanomaterials are
added to products due to their antimicrobial properties, conferring them the ability to modulate
the microbiota, thus enhancing the concern regarding their effect on commensal microbial
communities (Adamovsky et al., 2018).
1.1.3.3 Fish microbial function Half a billion years of co-evolution between vertebrates and their commensal bacteria
have perfected symbiotic relationships that ultimately benefits fish immunity and physiology
through microbial metabolism (Kelly and Salinas, 2017). Early studies on herbivorous fish
showed that some anaerobic species of the gut microbiota have the ability to help in
fermentative digestion (e.g., Fishelson et al., 1985; Rimmer and Wiebe, 1987). Since then,
several studies have established a circumstantial relationship between host fish physiology
and microbial function. For example, higher plant-fiber intake increased the diversity of
cellulolytic bacteria in grass carp (Li et al., 2016, 2008), while other commensal taxa were
reported to provide assimilable carbon to a wood eating fish (McDonald et al., 2012; Watts et
al., 2013). Carnivorous fish species harbor higher diversity of commensals that produce lipase
and protease, which aids in digestion (Liu et al., 2016). A non-dominant bacterial strain
recovered from the gut of Nile tilapia increased intestinal permeability, lipid accumulation and
triglyceride absorption efficiency when added to the diet (Zhang et al., 2020), while another
study has linked gut microbial diversity with fish metabolism phenotypes (Dvergedal et al.,
2020). Fish microbiota immunity aid may occur through several mechanisms, such as niche
exclusion, resource competition and antibiosis, i.e., production of antibiotic compounds (de
Bruijn et al., 2018; Kelly and Salinas, 2017). For example, commensals are able to compete
with pathogens by producing siderophores with higher affinity for iron, essential for microbial
growth (Ahmed and Holmström, 2014). In the gill microbiota of the Atlantic salmon, several
lactic acid isolates can produce antimicrobial compounds that inhibit the growth of the
pathogen Aeromonas salmonicida (see Ringø and Holzapfel, 2000). Using in vitro assays,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
16
another study showed that bacterial isolates from the skin microbiota of the rainbow trout were
able to inhibit the growth of two aquatic fungal pathogens, revealing a skin community with
antifungal properties (Lowrey et al., 2015). In zebrafish, the microbiota enhanced with
probiotics, led to a reduction in the stress response and anxious behaviour (e.g., Davis et al.,
2016). Moreover, gut microbiota was correlated to differentially expressed genes and
physiological parameters in the yellowfin seabream under hypo-osmotic stress, suggesting a
role of the commensals in the fish immune response (Lin et al., 2020).
Studies using gnotobiotic (germ-free) zebrafish have helped further explore the role of
gut microbiota functions. For example, gut microbiota of zebrafish can regulate the expression
of genes related to stimulation of epithelial proliferation (e.g., Cheesman et al., 2011; Rawls
et al., 2006, 2004), promote nutrient metabolism (e.g., Rawls et al., 2004; Semova et al., 2012)
and innate immune response (e.g., Galindo-Villegas et al., 2012; Kanther et al., 2011). On the
other hand, the absence of gut microbiota is related to impaired physiological functions in
zebrafish (e.g., Bates et al., 2006; Rawls et al., 2004).
Although extremely scarce in fish, studies using functional annotation of the
predominant bacterial taxa can offer in-depth insight into fish microbiota functional
composition. The gut microbiota of freshwater carp presented pathways broadly related to
biosynthesis, degradation, energy metabolism and fermentation (Tyagi et al., 2019).
Additionally, aligned to their herbivorous diet, the gut microbiota of freshwater carp also
presented pathways related to the degradation of cellulose and other complex carbohydrates
(Tyagi et al., 2019). In the same study, the authors were able to relate the high prevalence of
Actinobacteria to antibiotic biosynthesis pathways, highlighting a potential as a probiotic (Tyagi
et al., 2019). Another study revealed that functions related to DNA repair and antibiotic
response were enhanced in the feces microbiota of free-living populations of the brown-
marbled grouper when compared to mariculture populations (Hennersdorf et al., 2016).
As microbial diversity is governed by both host and environmental factors, so too is
microbial function. Current inferring tools, based mainly on metabolic information from humans
and their gut microbiota, can be very insightful for predicting which fish microbial functions are
altered by certain factors, although such predictions must be interpreted with caution. For
example, potential pathways of bacterial communities associated with nutrient metabolism,
antimicrobial biosynthesis and virulence factors of pathogens have been inferred from the gut
of two species of catfish (Bledsoe et al., 2018). However, minor differences in the microbial
predicted function were seen between the two species (Bledsoe et al., 2018). An experimental
setting showed that predicted metabolic functions of the gut related to biosynthesis of other
secondary metabolites, metabolism of cofactors and vitamins, and digestive system were
significantly different in perch fed different food ratios (Zha et al., 2018). Additionally,
differences in composition along the intestinal microbiota of grass carp were translated into

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
17
differential carbohydrate metabolism capacity by the microbiota, which increased along the
intestine (Yang et al., 2019). Regarding habitat, differences in the abundance of lipid and fatty
acid metabolism were found between the gut microbiota of rainbow trout from distinct
phylogeographic distributions (Yildirimer and Brown, 2018). The skin and gut of the South-
west European nase from different populations also showed differences in the predicted
metabolic pathways mainly related to amino acid and energy metabolism (Guivier et al., 2020).
Predation is another factor influencing predicted functional categories of the gut
microbiota of perch related to transport, signaling molecules and interactions, and
environmental adaptation (Zha et al., 2018). A study evaluating microbial differences in the
gut of the yellowtail kingfish of wild and aquaculture origins revealed that fish cultivation led to
differences in microbial predicted functions related to metabolism of cofactors, vitamins, amino
acids and carbohydrates (Ramírez and Romero, 2017b). On the other hand, the gut of Atlantic
salmon from hatcheries presented enhanced microbial predicted functions related to nitrogen
fixation and denitrifying when compared to their wild counterparts (Uren Webster et al., 2018).
Additionally, transition from freshwater to seawater was marked by alterations in the potential
metabolic function of the skin microbiota of the Atlantic salmon, suggesting its ability to
structurally change in order to maintain signaling (Lokesh and Kiron, 2016). Host thermal
tolerance was also linked to differences in the predicted metabolic functions related to cellular
processes and signaling pathways of the gut microbiota of tropical tilapia, suggesting that cold
exposure leads to microbial functional shifts related to stress and cell defense (Kokou et al.,
2018).
Predicted microbial function is altered by disease as was seen in the gut of the Yunlong
grouper, with pathways related to metabolism of terpenoids and polyketides, biosynthesis of
other secondary metabolites and metabolic diseases (under the “human diseases” broader
pathway) upregulated in diseased fish (Ma et al., 2019). In another study, infection with Vibrio
anguillarum induced changes in the predicted pathways involved in infectious diseases,
genetic information processing, and ion channels in the gut microbiota of ayu (Nie et al., 2017).
Of all the negative impacts of antibiotic administration, impaired gut potential function is one,
seen, for example, in the zebrafish exposed to oxytetracycline and sulfamethoxazole (Almeida
et al., 2019a; Zhou et al., 2018). Detailed knowledge of the function capabilities of the fish
microbiota are of great relevance for aquaculture management (Ghanbari et al., 2015). Further
work is necessary to identify fish microbial biomarkers in order to optimize fish microbiota and
mitigate disease (Xiong et al., 2019).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
18
1.1.4 Microbiome applications in aquaculture settings Given the link between fish, their microbiome and disease, microbial manipulation is a
promising and ecological approach to mitigate fish disease in aquaculture settings (de Bruijn
et al., 2018; Perry et al., 2020). Knowledge of microbial composition, diversity and function will
allow us to steer the microbial communities to select for specific functions (Hoseinifar et al.,
2016). In aquaculture that could be accomplished by using feed additives such as probiotics,
prebiotics and synbiotics (Hoseinifar et al., 2018, 2016).
The term probiotic derives from the Greek words “pro” and “bios”, meaning “pro-life”.
Probiotics are originally defined as “live microorganisms that, when administered in adequate
amounts, confer a health benefit on the host” (Hill et al., 2014). However, given the difference
between terrestrial and aquatic ecosystems, a slightly modified definition for probiotics was
proposed for aquaculture as “any microbial cell provided via the diet or rearing water that
benefits the host fish, fish farmer or fish consumer, which is achieved, in part at least, by
improving the microbial balance of fish” (Merrifield et al., 2010). Probiotics in aquaculture can
include bacteria, bacteriophages, microalgae and yeast (Llewellyn et al., 2014). Probiotics
have different modes of action on disease resistance, such as modulation of immune
parameters, competition for binding sites, production of antibacterial substances or
competition for nutrients (Hoseinifar et al., 2018). Probiotics contain conserved microbe-
associated molecular patterns (MAMPs) that are recognized by pattern recognition receptors
(PRRs) initiating the production of certain molecules, inducing the host immune response
(Bron et al., 2012; Remus et al., 2012). For example, a strain of Psychrobacter sp. was able
to prompt the immune response of the orange-spotted grouper (Sun et al., 2014). Moreover,
probiotics have the ability to bind to the adhesion receptors of the gut mucosa, reducing
pathogen colonization (Chabrillon et al., 2005). Among the most studied probiotics, lactic acid
bacteria are commonly used in aquaculture and include strains of Carnobacteria,
Lactobacillus, Lactococcus, Leuconostoc, Pediococcus, Enterococcus, Vagococcus and
Bacillus genera (Hoseinifar et al., 2018). For instance, the genera Bacillus and Lactobacillus
stimulate inflammatory expression in fish gut (He et al., 2017a), improve digestion and
metabolism (Falcinelli et al., 2015), and regulate genetic components involved in growth and
appetite (Falcinelli et al., 2016; Giorgia et al., 2018). Additionally, Lactobacillus is thought to
be involved in the gut-brain axis, improving learning and memory capacity (Borrelli et al., 2016;
Zang et al., 2019). Some gram-negative bacteria with probiotic activity include strains of
Pseudomonas (e.g., Giri et al., 2012; Vinoj et al., 2015), Aeromonas (e.g., Chi et al., 2014;
Pieters et al., 2008), Shewanella (e.g., Chabrillon et al., 2006; García de la Banda et al., 2012),
Enterobacter (e.g., Burbank et al., 2011; Rodriguez-Estrada et al., 2013) and Roseobacter
(e.g., Planas et al., 2006; Sonnenschein et al., 2020). Probiotics also have the ability to

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
19
improve water quality, by improving organic matter decomposition or reducing detrimental
elements, metabolic wastes or even pathogenic bacteria (Hai, 2015). Although there are
several constraints to probiotics related with environmental consistency, specificity,
production, storage and transport (Bentzon-Tilia et al., 2016; Borges et al., 2020; Cruz et al.,
2012), several probiotics are currently commercially available (Caipang et al., 2020; Shefat,
2018).
Prebiotics are nutrients that are not digested by the host and they promote growth
and/or activity of one or more beneficial microorganisms (Roberfroid, 2007). They are
fermented by the gut microbiota, leading to the production of short-chain fatty acids that may
be used as energy source by the host (Gibson and Roberfroid, 1995). Several common
prebiotics have been studied in fish, including inulin, fructooligosaccharides (FOS), short-
chain fructooligosaccharides (scFOS), oligofructose, mannanoligosaccharides (MOS), trans-
galactooligosaccharides (TOS), galactooligosaccharides (GOS), xylooligosaccharides (XOS),
arabinoxylooligosaccharides (AXOS) and isomaltooligosaccharides (IMO) (Ringø et al.,
2014). Prebiotics have been effectively tested in a number of fish species, including salmonids
(e.g., Atlantic salmon, Dimitroglou et al., 2011; rainbow trout, Staykov et al., 2007), gadoids
(e.g., Atlantic cod, Lokesh et al., 2012), cyprinids (e.g., common carp, Ebrahimi et al., 2012;
Indian carp, Andrews et al., 2009), sturgeons (e.g., beluga, Razeghi Mansour et al., 2012;
Siberian sturgeon, Mahious et al., 2006), catfishes (e.g., channel catfish, Welker et al., 2007),
among others (e.g., European seabass, Torrecillas et al., 2013; gilthead seabream, Cerezuela
et al., 2013; Nile tilapia, Ibrahem et al., 2010; Senegalese sole, Dimitroglou et al., 2011). The
mode of action of prebiotics includes growth of beneficial bacteria (Merrifield et al., 2010),
increase of fermentation products (Gibson and Roberfroid, 1995), or interaction with pattern
recognition patterns (Torrecillas et al., 2014) that ultimately enhance host health. For instance,
Lactobacillus and Bifidobacterium can be increased in abundance in favour of other less
beneficial bacteria, due to their ability to produce enzymes necessary for the fermentation of
prebiotics (e.g., Hoseinifar et al., 2014). Another example derives from attempts to find dietary
alternatives to the use of fishmeal in farmed fish diets. In this regard, insects, rich in chitin, if
added to fish diet, can act as prebiotics by increasing beneficial commensal bacteria in the gut
(e.g., Bruni et al., 2018). However, these effects are species specific, thus having limited
applications (Ringø et al., 2012). Moreover, there are several factors that influence mode of
action of prebiotics, including source, dosage, rearing conditions, fish species and age and
diet composition (Guerreiro et al., 2018).
Synbiotics are defined as “a mixture of probiotics and prebiotics that beneficially affects
the host by improving the survival and implantation of live microbial dietary supplements in the
gastrointestinal tract, by selectively stimulating the growth and/or by activating the metabolism
of one or a limited number of health-promoting bacteria, and thus improving host welfare”

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
20
(Gibson and Roberfroid, 1995). Synbiotics are nutritional supplements that can be
administered through feed or baths (Amenyogbe et al., 2020; Cerezuela et al., 2011). For
example, rainbow trout fed with a synbiotic additive that contained the probiotic Enterococcus
faecalis and the prebiotic MOS exhibited increased growth, haematology and immune
response, and resistance to the fish pathogen Aeromonas salmonicida (see Rodriguez-
Estrada et al., 2013). Other synbiotics had similar responses in a number of fish, but the
mechanisms are still largely unknown (reviewed by Huynh et al., 2017). Nevertheless, it is
known that their metabolic processes are able to release enzyme and bioactive products that
promote fish growth. Additionally, they can also promote amino acid production through
microbial metabolism processes that can be utilized as essential nutrients or ultimately
improve gut nutrient absorptive capability (Huynh et al., 2017). Further understanding about
the specific metabolic processes behind the mode of action of synbiotics is needed to increase
implementation in aquaculture (Cerezuela et al., 2011; Huynh et al., 2017).
In aquaculture, it is possible to select for specific microbial communities, either through
directly supplying the system or through environment fertilization (Bentzon-Tilia et al., 2016).
For example, biofloc technology is based on manipulation of microbiota through nutrient
limitation of the microbes that are not able to produce their own food (heterotrophs) in zero-
exchange aquaculture systems (Emerenciano et al., 2013). This technology not only improves
water quality, but leads to the formation of flocs, i.e., bacterial macroaggregates, that can be
eaten by the fish (Emerenciano et al., 2013). Another approach is applied in RAS
(Recirculating Aquaculture Systems) technology, which keeps the water within the system and
reconditions the microbial communities of the rearing water, leading to the biofilm formation of
certain bacteria that are able to remove toxic ammonia and produce nitrate (Espinal and
Matulić, 2019).
Assessing aquaculture microbiota could give insights into changes in specific microbial
communities that are influenced by different chemical parameters, acting as an early warning
indicator of the water quality (Bentzon-Tilia et al., 2016). Additionally, a microbiome-based
monitoring system could also assess undesirable microorganisms such as pathogenic
bacteria, both in water as well as in fish (Bentzon-Tilia et al., 2016; Perry et al., 2020). One
new direction for disease management in aquaculture is to use the detection of dysbiosis in
farmed fish for predicting the onset of disease and as indicator tool for productivity, which can
ultimately be used to optimize aquaculture practices (Infante-Villamil et al., 2020; Perry et al.,
2020).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
21
1.1.5 Available tools for microbiome assessment With advances in molecular technology, microbiome assessments have become a
practical reality. Currently, many different techniques allow for the identification of the entire
microbiome, as well as its diversity and (predicted) function (Tarnecki et al., 2017).
1.1.5.1 Sampling Designing a microbiome study need to take into consideration the species, the
environment and factors that will be addressed (Knight et al., 2018). Regarding the fish
microbiome, many are the studies that take advantage of collaborations with fish farms and
aquaculture settings (e.g., Minich et al., 2020a; Pimentel et al., 2017; Tarnecki et al., 2019),
while several others are conducted in controlled laboratory environments (e.g., Lokesh and
Kiron, 2016; Pindling et al., 2018; Zhang et al., 2018). Inoculation of germ-free models with
specific bacteria can also be advantageous to create direct links between microbes and
function, although the lack of a homeostatic community context can lead to misinterpretation
of results (Laukens et al., 2015).
In fish, sampling the microbial communities of internal organs is invasive, requiring
animal sacrifice, while assessing the microbiome of external organs can be done non-
invasively. The gut microbiome can be sampled invasively by dissecting the intestine (e.g.,
Xia et al., 2014), or non-invasively, by sampling the microbiome of the feces (e.g., Zarkasi et
al., 2014). Swabbing is the preferred method for microbiome sampling, being least invasive,
and there is little difference to other methods, such as scraping or punch biopsy (Grice et al.,
2008). Increasingly studies are more integrative, assessing the microbiome from different
potential sources; for fish, these include water, feed, sediment, biofilm and nets (e.g., Canada
et al., 2020; León-Zayas et al., 2020; Minich et al., 2020b; Nikouli et al., 2019). Considering
the influence of factors affecting the microbiome such as age, diet, habitat, antibiotic usage
(see section 1.1.3), is also of great importance when planning the study (Kim et al., 2017).
Given the high variability of the microbiome over time, it is important to reflect on
whether a single-time sampling is sufficient or longitudinal sampling is more appropriate (Sinha
et al., 2015). For example, the human gut microbiome displays a circadian rhythm (e.g., Liang
et al., 2014). When cross-sectional design is the only option available, it is paramount to know
if the single time point is representative of the actual microbial diversity and disclose it when it
might not be (Kim et al., 2017). Additionally, appropriate sample size and replicates should be
selected to properly estimate diversity and variability and avoid biases and misinterpretations
(Bharti and Grimm, 2021; Goodrich et al., 2014; Prosser, 2010).
Appropriate transportation and storage are very important in microbiome studies to
preserve the integrity of the microbial DNA/RNA and community composition, and maintain

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
22
the sample potential for multiple studies, techniques and platforms (Lindahl, 1993; Song et al.,
2016; Sinha et al., 2015). A study evaluating the efficacy of different storage media in the gut
and gill microbiota of rainbow trout showed that storage in 96% ethanol outperforms other
methods in terms of cost, labor, DNA quality and quantity (Hildonen et al., 2019). For
metatranscriptomic analysis, RNAlaterâ buffer is the most efficient to preserve RNA integrity
(Reck et al., 2015). Long term storage temperature can significantly impact results (e.g.,
Carda-Diéguez et al., 2014; Larsen et al., 2015b), especially in communities with lower
diversity (Hill et al., 2016), but is not a problem for short-term DNA storage (e.g., Kim et al.,
2017; Lauber et al., 2010). For RNA studies, however, storage conditions are more critical
(Goodrich et al., 2014). It is recommended to perform DNA/RNA extractions as quickly as
possible after collection and storage (Kuczynski et al., 2012). Importantly, all samples within
a study must have consistent storage conditions (Kim et al., 2017).
1.1.5.2 Laboratory procedures for culture independent microbial
assessments Three fundamental things need to be taken into account regarding laboratory
procedures for microbial assessments: DNA/RNA extraction protocols, PCR conditions, and
sequencing platforms and techniques (Tarnecki et al., 2017). Although extraction protocols
use the same basic steps, different types of cell lysis can be applied, affecting the results (e.g.,
Larsen et al., 2015b; Smith, 2011). Additionally, variation due to different batches of extraction
kits was also previously found (Bushon et al., 2010). Importantly, bias due to extraction
techniques are often confounded by high inter-individual variation observed in some species
(Wagner Mackenzie et al., 2015). Hence, it is important to use the same kit and batch for all
the samples in a study (Kim et al., 2017).
Different approaches using culture independent methods have been used for microbial
characterization (Tarnecki et al., 2017). The most recently developed, more comprehensive
and most commonly used approach nowadays is metataxonomics, which sequences targeted
amplicons of phylogenetically informative markers (Tarnecki et al., 2017). Initially this method
was achieved through Sanger sequencing and was expensive, but the transition to Next
Generation Sequencing technologies allowed for high-throughput and cost-effective detailed
characterization of the microbial communities present in any taxa, as well as in any
environment (Tarnecki et al., 2017). The 16S marker gene is sequenced for bacteria, while
18S is sequenced for eukaryotes and ITS (internal transcribed spacer) for fungi (Kim et al.,
2017). Most commonly used in fish studies, sequencing the 16S rRNA gene allows for the
identification of bacterial isolates (Rosselló-Mora, 2005). The 16S rRNA gene is present in all
living organisms and has the advantage of containing slow evolving regions, useful to design

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
23
broad-spectrum primers, as well as fast evolving regions, providing powerful classification at
finer taxonomic levels, such as genera (Kuczynski et al., 2012). The 16S rRNA gene
comprises nine variable regions, V1 to V9 (Yarza et al., 2014), with the V4 region being the
most commonly sequenced in fish studies (Tarnecki et al., 2017). Result bias due to the
different regions selected are mainly related to primer choice, which can differentially detect
taxa, both within or between regions (Walker et al., 2015). Different sequence variability
between regions can also lead to dissimilarities in microbial diversity (Engelbrektson et al.,
2010). Prior to sequencing, amplification of the 16S rRNA variable regions is done through
PCR; however, PCR conditions can also lead to potential biases (Tarnecki et al., 2017).
However, a single “best” methodology is yet to be standardized and the best practice is to
consistently apply the same protocol to all samples in a study (Goodrich et al., 2014).
It is possible to sequence all DNA in a sample through shotgun metagenomics
sequencing (Kim et al., 2017). Metagenomics refers to the study of the genetic material
recovered from a particular environment or environmental niche and is dedicated to the
analysis of microbial communities with molecular techniques (Handelsman et al., 1998;
Weisburg et al., 1991). In metagenome sequencing, it is possible to determine which genes
are present and their overall functionalities or pathways (Di Bella et al., 2013). However, this
technique has depth limitations, being challenging to generate high-quality de novo
assemblies, especially in samples that contain large fractions of host DNA, such as mucosal
sites (Kuczynski et al., 2012). Recently emerged techniques in the field of metatranscriptomics
enable characterization of dynamic, context-specific functional profiles of microbial
communities (Shakya et al., 2019). Metatranscriptomics aims to sequence all RNAs in a
sample, determining which genes are being transcribed under particular conditions (Di Bella
et al., 2013), and ultimately can retrieve the entire metatranscriptome. Several bioinformatic
tools are currently available to annotate and map RNA reads to the corresponding metabolic
pathways (Shakya et al., 2019). Metagenomic and metatranscriptomic studies are far more
challenging than amplicon sequence studies, given the fact that coverage is considerably
lower for whole genome sequencing (Di Bella et al., 2013). In order to accurately resolve
differences between samples, high coverage is imperative (Di Bella et al., 2013). Additionally,
different abundances of bacterial taxa and transcription levels of every gene across samples
add extra difficulty to sequence assembly, thus requiring relatively high number of samples,
all with good coverage (Di Bella et al., 2013).
Different sequencing technologies are used in microbial studies, with distinct
characteristics pertaining to read length, sequencing depth, run time, reads per run, relative
cost and error rate (Kuczynski et al., 2012). Different sequencing technologies can also lead
to bias, although bias related to primer choice seems to have greater impact (Tarnecki et al.,
2017). In this regard, third-generation sequencing techniques that do not amplify the DNA and

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
24
thus, do not require amplification prior to sequencing, are promising in reducing bias and
allowing research standardization (Di Bella et al., 2013).
Especially important during sequencing, positive controls usually include a mixture of
cultured curated organisms, called “mock communities” (Kim et al., 2017; Sinha et al., 2015).
This procedure allows the accuracy of the sequencing to be determined. On the other hand,
there are many possible contamination sources during laboratory procedures, including
human, reagents, kits, dust, or crossover between samples (Kim et al., 2017). A marginal
contamination is virtually inevitable when dealing with microbes, especially with low microbial
biomass samples (Kim et al., 2017). In this regard, negative controls are equally important
during extraction, PCR amplification and sequencing (Kim et al., 2017; Sinha et al., 2015).
These refer to samples without any DNA content, or blanks, and allow identification of false
positives derived from contamination (Kim et al., 2017).
In summary, metataxonomics of the 16S are the most cost effective and popular in
microbiota studies, allowing a metataxonomic analysis (e.g., identification of OTUs/ASVs and
answering which bacteria are present in a sample) (Adamovsky et al., 2018). On the other
hand, metagenomic techniques aim to identify as many genes as possible, and are more
suitable for understanding microbiota dynamics (Adamovsky et al., 2018). Finally,
metatranscriptomic methods aim to retrieve as many RNA transcripts as possible, identifying
differential gene expression and understanding microbiota functionality (Adamovsky et al.,
2018).
1.1.5.3 Data analyses While different methods have been developed to analyze the microbiome, the common
objective is to properly characterize their composition, diversity and function (Goodrich et al.,
2014). The most widely used software options to address microbial data from target amplicon
sequence studies are QIIME (Caporaso et al., 2010), mothur (Schloss et al., 2009) and
DADA2 (Callahan et al., 2016). In metagenomic analyses, software such as IBDA-UD (Peng
et al., 2012), metaSPAdes (Nurk et al., 2017), MEGAHIT (Li et al., 2015), or MetaVelvet
(Namiki et al., 2012) are commonly used. The software SOAPdenovo (Xie et al., 2014) is
generally employed in metatranscriptomic analysis. It is recommended to always use the most
recent version of the software, as they are constantly being updated and improved (Galloway-
Peña and Hanson, 2020).
General bioinformatic pipelines for targeted amplicon sequence studies (e.g., 16S)
encompass quality checks, denoising sequences, clustering sequences into OTUs or ASVs,
inferring taxonomy or phylogeny, and performing multivariate analysis and data visualization
(Kuczynski et al., 2012). In microbiome studies, it is important to perform data filtering, which

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
25
are sequencing-platform specific (Goodrich et al., 2014). This step will discard sequences that
fail to meet minimum quality requirements, erroneous sequences (e.g., PCR chimaeras), and
those that do not match targeted organisms (Goodrich et al., 2014). It is also common to
remove sequences with single or low reads, as they can be considered contaminants or
sequencing artifacts (Goodrich et al., 2014). To deal with inter-sample differences in biomass
or sequencing depths, normalization can be done by rarefying the OTU/ASV frequency by the
total sample sequence count (Goodrich et al., 2014). However, this approach is not
consensual, as it is not able to detect differentially abundant species and can lead to high
rates of false positives (McMurdie and Holmes, 2014). As such, other approaches, such as
the negative binomial distribution, are preferred (McMurdie and Holmes, 2014).
Clustering sequences into OTUs or ASVs by sequence identity followed by taxonomy
assignment will then provide identification of the bacterial groups in the samples (Goodrich et
al., 2014). In the case of OTUs, this is done through picking algorithms, which can be de novo,
closed reference or open reference, and the chosen method will impact results (Goodrich et
al., 2014). Reference-based methods will cluster sequences, normally using a cut-off point of
97% or higher sequence similarity (Rosselló-Mora, 2005), and compare them against
reference databases that are regularly updated, and thus results can also change between
the taxonomic database versions (Goodrich et al., 2014). On the other hand, ASVs are inferred
de novo without arbitrary thresholds and distinguish sequence variants by as little as one
nucleotide (Callahan et al., 2017). ASVs methods have higher resolution, are better at
discriminating ecological patterns and allow to reliably compare ASVs across studies
(Callahan et al., 2017). Common databases of reference sequences and taxonomies for the
16S rRNA include greengenes (DeSantis et al., 2006), SILVA (Pruesse et al., 2007), and the
Ribosomal Database Project (Cole et al., 2009). Many OTUs/ASVs will be typically identified
to the genus or higher taxonomic levels; species level identification is rare and should always
be prudently considered (Goodrich et al., 2014).
Microbial diversity analysis is performed by comparing species composition and
abundance within and between samples (Lozupone and Knight, 2008). Alpha-diversity
quantifies diversity within individuals and can be compared across sample groups (Knight et
al., 2018). Alpha-diversity metrics can measure species richness (e.g., Chao1 index or Faith’s
phylogenetic diversity), evenness (e.g., Pielou’s evenness), or both (e.g., Shannon or Inverse
Simpson indices). Beta-diversity analyses compare dissimilarities between samples,
generating distance matrices between all pairs of samples (Knight et al., 2018). Beta-diversity
metrics can be qualitative, based on sequence presence or absence (e.g., Jaccard or
unweighted UniFrac) or quantitative, taking into account sequence abundance (e.g., Bray-
Curtis or weighted UniFrac). UniFrac metrics have the advantage of accounting for
phylogenetic distances between sets of taxa (Lozupone and Knight, 2005).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
26
Recently, software has been developed to predict microbial functional profile using 16S
rRNA data (Galloway-Peña and Hanson, 2020). Programs such as PICRUSt (Langille et al.,
2013) or Tax4Fun (Aßhauer et al., 2015) use taxa relative abundance to predict the function
of the gene content based on reference genomes. These techniques are highly reliant on
currently available reference genomes, which are biased towards human health
microorganisms (Choi et al., 2017). It is important to notice that these methods do not take
into account actual gene or protein expression, hence results represent a rough estimation
and should be carefully interpreted (Galloway-Peña and Hanson, 2020).
When conducting metagenomic or metatranscriptomic studies, the bioinformatic
pipeline involves assembling sequences into contigs, predicting genes, identifying sequence
function using annotation databases, identifying metabolic pathways, comparing to published
results and finally perform multivariate analysis and visualization (Kuczynski et al., 2012).
Assemblies can be done based on reference genomes, de novo, or a combination of both,
with the first being highly dependent on the availability of reference genomes and quality of
the database (Galloway-Peña and Hanson, 2020). After assembling, genes are identified and
sequence function is annotated through protein sequence homology-based searches against
databases (Galloway-Peña and Hanson, 2020). Sequence reads are mapped to reference
genomes and pathways, such as KEGG (Kanehisa et al., 2019), in order to identify active
microbes and the function of their expressed genes (Galloway-Peña and Hanson, 2020).
Software such as KEGGscape is then able to infer metabolic networks (Nishida et al., 2014).
After microbial assessment, several statistical tools can be implemented to analyze
variation in microbial diversity, taxa abundance or functional components and evaluate overall
patterns of microbial variation with covariates (Galloway-Peña and Hanson, 2020; Mallick et
al., 2017). Statistical methods for microbiome analysis have been developed to specifically
tackle the complex microbiome datasets, such as PERMANOVA analysis to compare
microbial distance matrices (Kelly et al., 2015). Other conventional statistical approaches can
be applied to compare alpha-diversity or abundance variation, such as ANOVA (Sthle and
Wold, 1989) or Kruskal-Wallis tests (Kruskal and Wallis, 1952). These analyses can be
performed using the R packages phyloseq (McMurdie and Holmes, 2013) or vegan (Oksanen
et al., 2008) in target amplicon sequence studies. With metagenomic and metatranscriptomic
data, pathway statistical analysis can be performed, for example, via the pathfindR R package
(Ulgen et al., 2019). Linear discriminant analysis of effect sizes (LEfSe) can be also used to
identify differentially abundant taxa or predicted metabolic pathways (Segata et al., 2011).
Finally, it is important that researchers disclose as much information as possible and
discuss the results taking into account a global improvement and standardization of
microbiome research (Goodrich et al., 2014). Deposition of all sequence files and metadata to
public databases using standard formats is essential (Goodrich et al., 2014).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
27
1.2 Thesis structure and objectives The main objectives of this doctoral thesis were to assess the composition, diversity
and structure of the skin and gill microbiota of two commercially important fish species, the
European seabass Dicentrarchus labrax and the gilthead seabream Sparus aurata in an
aquaculture setting. This thesis was developed in collaboration with Piscicultura do Vale da
Lama, a fish farm based in southern Portugal (Ria de Alvor). This is a semi-intensive fish farm
focused on the production of the seabass and seabream. In this farm, grow out phases occur
in exterior ponds, water is captured from the estuary and naturally renewed with the tide (i.e.,
twice a day). My specific goals were to perform baseline characterization of the microbiota of
the skin and gill of juveniles and adults of both species and understand the impact of the
following factors on their microbial dynamics: ontogeny, water temperature, disease and
antibiotic treatment. Towards this goal I used high-throughput amplicon sequencing of the V4
region of the 16S rRNA of bacteria and metataxonomic alpha- and beta-diversity analyses.
Analysis of the predicted function was also applied to some datasets.
The thesis is divided into six chapters. Following this literature review Chapter,
Chapters 2 to 5 are presented as scientific papers. Specifically, Chapters 2, 3 and 4 and
Subchapter 5.1 were published in the international scientific journals Aquaculture, Animal
Microbiome, ISME Communications and Scientific Reports, respectively, whereas Subchapter
5.2 is under review.
Chapter 2 is a baseline characterization of the diversity of the skin and gill microbiota
of the seabass and seabream, assessed by monitoring microbial composition and structure of
these species during 3 winter months (December to February). This study identified the most
abundant phyla and genera and revealed significant differences in the microbial diversity and
structure between tissues and species. It was also possible to identify the presence of several
potentially pathogenic genera within the microbiota of healthy fish.
In Chapter 3 I assessed the effects of aging on the skin and gill microbiota of farmed
seabass and seabream. Three different age groups were studied, including early and late
juveniles and mature adults. Age was seen to shape the microbiota of both species, but varied
for each species. Additionally, the microbiota of the surrounding water was significantly distinct
from the fish microbiota, regardless of the age group.
Chapter 4 comprises a longitudinal study that aimed to characterize the skin and gill
microbiota of seabass and the surrounding water over a period of 12 months, while also
assessing the impact of water temperature on microbial dynamics. Results showed similar
trends in the microbial diversity of both tissues. The microbiota were highly dynamic with
periods where several potentially pathogenic genera were highly abundant in both tissues.
The potential pathogens also exhibited complex interactions with other bacterial genera. The

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
28
surrounding water temperature influenced fish microbiota, with episodes of dysbiosis
coinciding with warmer months and during transitions between cold/warm months.
Chapter 5 encloses 2 scientific articles, both related to the effects of disease and
antibiotic treatment on the microbiota of seabass. Subchapter 5.1 evaluated the effects of a
natural disease outbreak of Photobacterium damselae and subsequent antibiotic treatment
with oxytetracycline on the gill and skin microbiota of adult seabass. Both infection and
antibiotic treatment caused significant changes in fish microbiota, although such changes
were asymmetrical between tissues. Effects of dysbiosis were more pronounced in the skin
but the gill microbiota presented less resilience and did not return to the diversity levels
observed previous to disease and treatment. Subchapter 5.2 also assessed the effects of a
natural disease outbreak, caused by a coinfection of Photobacterium damselae spp. piscicida
and Vibrio harveyi, this time on the skin of seabass fingerlings. It also analyzed the response
to subsequent antibiotic treatment with flumequine. As in the previous study, both disease and
antibiotic treatment led to dysbiosis, although in this case a significant increase in microbial
diversity was observed. Nevertheless, changes in the abundance of potential pathogenic
genera and other opportunistic taxa confirmed that dysbiosis occurred.
To finalize, Chapter 6 summarizes and discusses all results of this thesis and the future
perspectives based on the outcome of the study of the microbial dynamics and dysbiosis in
farmed European seabass and gilthead seabream.
1.3 References Abdel-Aziz, M., Eissa, A.E., Hanna, M., Okada, M.A., 2013. Identifying some pathogenic
Vibrio/Photobacterium species during mass mortalities of cultured Gilthead seabream
(Sparus aurata) and European seabass (Dicentrarchus labrax) from some Egyptian
coastal provinces. Int. J. Vet. Sci. Med. 1, 87–95.
https://doi.org/10.1016/j.ijvsm.2013.10.004
Abdul Razak, S., Griffin, M.J., Mischke, C.C., Bosworth, B.G., Waldbieser, G.C., Wise, D.J.,
Marsh, T.L., Scribner, K.T., 2019. Biotic and abiotic factors influencing channel catfish
egg and gut microbiome dynamics during early life stages. Aquaculture 498, 556–567.
https://doi.org/10.1016/j.aquaculture.2018.08.073
Adamovsky, O., Buerger, A.N., Wormington, A.M., Ector, N., Griffitt, R.J., Bisesi, J.H.,
Martyniuk, C.J., 2018. The gut microbiome and aquatic toxicology: An emerging concept
for environmental health. Environ. Toxicol. Chem. 37, 2758–2775.
https://doi.org/10.1002/etc.4249
Ahmed, E., Holmström, S.J.M., 2014. Siderophores in environmental research: roles and
applications. Microb. Biotechnol. 7, 196–208. https://doi.org/10.1111/1751-7915.12117

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
29
Alekseyenko, A. V, Perez-Perez, G.I., De Souza, A., Strober, B., Gao, Z., Bihan, M., Li, K.,
Methé, B.A., Blaser, M.J., 2013. Community differentiation of the cutaneous microbiota
in psoriasis. Microbiome 1, 31. https://doi.org/10.1186/2049-2618-1-31
Aleshina, E., Miroshnikova, E., Sizova, E., 2020. Transformation of microbiota of fish intestines
and gills against the background of molybdenum oxide nanoparticles in environment. Int.
J. Environ. Sci. Technol. 17, 721–732. https://doi.org/10.1007/s13762-019-02509-x
Almeida, A.R., Alves, M., Domingues, I., Henriques, I., 2019a. The impact of antibiotic
exposure in water and zebrafish gut microbiomes: A 16S rRNA gene-based
metagenomic analysis. Ecotoxicol. Environ. Saf. 186, 109771.
https://doi.org/10.1016/j.ecoenv.2019.109771
Almeida, A.R., Tacão, M., Machado, A.L., Golovko, O., Zlabek, V., Domingues, I., Henriques,
I., 2019b. Long-term effects of oxytetracycline exposure in zebrafish: A multi-level
perspective. Chemosphere 222, 333–344.
https://doi.org/10.1016/j.chemosphere.2019.01.147
Amenyogbe, E., Chen, G., Wang, Z., Huang, J., Huang, B., Li, H., 2020. The exploitation of
probiotics, prebiotics and synbiotics in aquaculture: present study, limitations and future
directions. : a review. Aquac. Int. 28, 1017–1041. https://doi.org/10.1007/s10499-020-
00509-0
Andrews, S.R., Sahu, N.P., Pal, A.K., Kumar, S., 2009. Haematological modulation and growth
of Labeo rohita fingerlings: effect of dietary mannan oligosaccharide, yeast extract,
protein hydrolysate and chlorella. Aquac. Res. 41, 61–69. https://doi.org/10.1111/j.1365-
2109.2009.02304.x
Ashley, P.J., 2007. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 104,
199–235. https://doi.org/10.1016/j.applanim.2006.09.001
Aßhauer, K.P., Wemheuer, B., Daniel, R., Meinicke, P., 2015. Tax4Fun: predicting functional
profiles from metagenomic 16S rRNA data: Fig. 1. Bioinformatics 31, 2882–2884.
https://doi.org/10.1093/bioinformatics/btv287
Austin, B., Austin, D.A., 2016. Bacterial fish pathogens, 6th ed. Springer, Stirling, UK.
https://doi.org/10.1007/978-3-319-32674-0
Bäckhed, F., Manchester, J.K., Semenkovich, C.F., Gordon, J.I., 2007. Mechanisms
underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci.
U. S. A. 104, 979–984. https://doi.org/10.1073/pnas.0605374104
Bagi, A., Riiser, E.S., Molland, H.S., Star, B., Haverkamp, T.H.A., Sydnes, M.O., Pampanin,
D.M., 2018. Gastrointestinal microbial community changes in Atlantic cod (Gadus
morhua) exposed to crude oil. BMC Microbiol. 18, 1–14. https://doi.org/10.1186/s12866-
018-1171-2
Baker-Austin, C., Trinanes, J.A., Taylor, N.G.H., Hartnell, R., Siitonen, A., Martinez-Urtaza, J.,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
30
2013. Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim.
Chang. 3, 73–77. https://doi.org/10.1038/nclimate1628
Bakopoulos, V., Volpatti, D., Gusmani, L., Galeotti, M., Adams, A., Dimitriadis, G.J., 2003.
Vaccination trials of sea bass, Dicentrarchus labrax (L.), against Photobacterium
damselae subsp. piscicida, using novel vaccine mixtures. J. Fish Dis. 26, 77–90.
https://doi.org/10.1046/j.1365-2761.2003.00438.x
Banerjee, G., Kumar Ray, A., 2017. Bacterial symbiosis in the fish gut and its role in health
and metabolism. Symbiosis 72, 1–11. https://doi.org/10.1007/s13199-016-0441-8
Basset, C., Holton, J., O’Mahony, R., Roitt, I., 2003. Innate immunity and pathogen-host
interaction. Vaccine. https://doi.org/10.1016/S0264-410X(03)00195-6
Bates, J.M., Mittge, E., Kuhlman, J., Baden, K.N., Cheesman, S.E., Guillemin, K., 2006.
Distinct signals from the microbiota promote different aspects of zebrafish gut
differentiation. Dev. Biol. 297, 374–386. https://doi.org/10.1016/j.ydbio.2006.05.006
Bellos, G., Angelidis, P., Miliou, H., 2015. Effect of temperature and seasonality principal
epizootiological risk factor on vibriosis and photobacteriosis outbreaks for European sea
bass in Greece (1998-2013). ournal Aquac. Res. Dev. 6.
Béné, C., Barange, M., Subasinghe, R., Pinstrup-Andersen, P., Merino, G., Hemre, G.I.,
Williams, M., 2015. Feeding 9 billion by 2050 – Putting fish back on the menu. Food
Secur. 7, 261–274. https://doi.org/10.1007/s12571-015-0427-z
Bentzon-Tilia, M., Sonnenschein, E.C., Gram, L., 2016. Monitoring and managing microbes in
aquaculture - Towards a sustainable industry. Microb. Biotechnol. 9, 576–584.
https://doi.org/10.1111/1751-7915.12392
Berg, G., Rybakova, D., Fischer, D., Cernava, T., Vergès, M.C.C., Charles, T., Chen, X.,
Cocolin, L., Eversole, K., Corral, G.H., Kazou, M., Kinkel, L., Lange, L., Lima, N., Loy, A.,
Macklin, J.A., Maguin, E., Mauchline, T., McClure, R., Mitter, B., Ryan, M., Sarand, I.,
Smidt, H., Schelkle, B., Roume, H., Kiran, G.S., Selvin, J., Souza, R.S.C. de, Van
Overbeek, L., Singh, B.K., Wagner, M., Walsh, A., Sessitsch, A., Schloter, M., 2020.
Microbiome definition re-visited: old concepts and new challenges. Microbiome.
https://doi.org/10.1186/s40168-020-00875-0
Bharti, R., Grimm, D.G., 2021. Current challenges and best-practice protocols for microbiome
analysis. Brief. Bioinform. 22, 178–193. https://doi.org/10.1093/bib/bbz155
Blaser, M.J., Cardon, Z.G., Cho, M.K., Dangl, J.L., Donohue, T.J., Green, J.L., Knight, R.,
Maxon, M.E., Northen, T.R., Pollard, K.S., Brodie, E.L., 2016. Toward a Predictive
Understanding of Earth’s Microbiomes to Address 21st Century Challenges.
https://doi.org/10.1128/mBio.00714-16
Bledsoe, J.W., Peterson, B.C., Swanson, K.S., Small, B.C., 2016. Ontogenetic
Characterization of the Intestinal Microbiota of Channel Catfish through 16S rRNA Gene

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
31
Sequencing Reveals Insights on Temporal Shifts and the Influence of Environmental
Microbes. PLoS One 11, e0166379. https://doi.org/10.1371/journal.pone.0166379
Bledsoe, J.W., Waldbieser, G.C., Swanson, K.S., Peterson, B.C., Small, B.C., 2018.
Comparison of Channel Catfish and Blue Catfish Gut Microbiota Assemblages Shows
Minimal Effects of Host Genetics on Microbial Structure and Inferred Function. Front.
Microbiol. 9, 1073. https://doi.org/10.3389/fmicb.2018.01073
Bolnick, D.I., Snowberg, L.K., Hirsch, P.E., Lauber, C.L., Knight, R., Caporaso, J.G.,
Svanbäck, R., 2014. Individuals’ diet diversity influences gut microbial diversity in two
freshwater fish (threespine stickleback and Eurasian perch). Ecol. Lett. 17, 979–987.
https://doi.org/10.1111/ele.12301
Borges, N., Keller-Costa, T., Sanches-Fernandes, G.M.M., Louvado, A., Gomes, N.C.M.,
Costa, R., 2020. Bacteriome Structure, Function, and Probiotics in Fish Larviculture: The
Good, the Bad, and the Gaps. Annu. Rev. Anim. Biosci. 9, 423–452.
https://doi.org/10.1146/annurev-animal-062920
Borrelli, L., Aceto, S., Agnisola, C., De Paolo, S., Dipineto, L., Stilling, R.M., Dinan, T.G.,
Cryan, J.F., Menna, L.F., Fioretti, A., 2016. Probiotic modulation of the microbiota-gut-
brain axis and behaviour in zebrafish. Sci. Rep. 6, 1–9. https://doi.org/10.1038/srep30046
Boutin, S., Bernatchez, L., Audet, C., Derôme, N., 2013. Network Analysis Highlights Complex
Interactions between Pathogen, Host and Commensal Microbiota. PLoS One 8, e84772.
https://doi.org/10.1371/journal.pone.0084772
Bouwmeester, M.M., Goedknegt, M.A., Poulin, R., Thieltges, D.W., 2020. Collateral diseases:
Aquaculture impacts on wildlife infections. J. Appl. Ecol. https://doi.org/10.1111/1365-
2664.13775
Bowden, T.J., Thompson, K.D., Morgan, A.L., Gratacap, R.M.L., Nikoskelainen, S., 2007.
Seasonal variation and the immune response: A fish perspective. Fish Shellfish Immunol.
22, 695–706. https://doi.org/10.1016/j.fsi.2006.08.016
Boyd, C., McNevin, A., 2015. Aquaculture, Resource Use, and the Environment. John Wiley
& Sons.
Bron, P.A., Tomita, S., van Swam, I.I., Remus, D.M., Meijerink, M., Wels, M., Okada, S., Wells,
J.M., Kleerebezem, M., 2012. Lactobacillus plantarum possesses the capability for wall
teichoic acid backbone alditol switching. Microb. Cell Fact. 11, 123.
https://doi.org/10.1186/1475-2859-11-123
Brown, R.M., Wiens, G.D., Salinas, I., 2019. Analysis of the gut and gill microbiome of resistant
and susceptible lines of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol.
86, 497–506. https://doi.org/10.1016/j.fsi.2018.11.079
Bruni, L., Pastorelli, R., Viti, C., Gasco, L., Parisi, G., 2018. Characterisation of the intestinal
microbial communities of rainbow trout (Oncorhynchus mykiss) fed with Hermetia illucens

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
32
(black soldier fly) partially defatted larva meal as partial dietary protein source.
Aquaculture 487, 56–63. https://doi.org/10.1016/j.aquaculture.2018.01.006
Burbank, D.R., Shah, D.H., LaPatra, S.E., Fornshell, G., Cain, K.D., 2011. Enhanced
resistance to coldwater disease following feeding of probiotic bacterial strains to rainbow
trout (Oncorhynchus mykiss). Aquaculture 321, 185–190.
https://doi.org/10.1016/j.aquaculture.2011.09.004
Burns, A.R., Miller, E., Agarwal, M., Rolig, A.S., Milligan-Myhre, K., Seredick, S., Guillemin,
K., Bohannan, B.J.M., 2017. Interhost dispersal alters microbiome assembly and can
overwhelm host innate immunity in an experimental zebrafish model. Proc. Natl. Acad.
Sci. U. S. A. 114, 11181–11186. https://doi.org/10.1073/pnas.1702511114
Bushon, R.N., Kephart, C.M., Koltun, G.F., Francy, D.S., Schaefer, F.W., Alan Lindquist, H.D.,
2010. Statistical assessment of DNA extraction reagent lot variability in real-time
quantitative PCR. Lett. Appl. Microbiol. 50, 276–282. https://doi.org/10.1111/j.1472-
765X.2009.02788.x
Cabello, F.C., 2006. Heavy use of prophylactic antibiotics in aquaculture: a growing problem
for human and animal health and for the environment. Environ. Microbiol. 8, 1137–1144.
https://doi.org/10.1111/j.1462-2920.2006.01054.x
Caipang, C.M.A., Suharman, I., Avillanosa, A.L., Bargoyo, V.T., 2020. Host-derived Probiotics
for Finfish Aquaculture Host-derived Probiotics for Finfish Aquaculture, in: IOP
Conference Series: Earth and Environmental Science. IOP Publishing.
https://doi.org/10.1088/1755-1315/430/1/012026
Callahan, B.J., McMurdie, P.J., Holmes, S.P., 2017. Exact sequence variants should replace
operational taxonomic units in marker-gene data analysis. ISME J. 11, 2639–2643.
https://doi.org/10.1038/ismej.2017.119
Callahan, B.J., McMurdie, P.J., Rosen, M.J., Han, A.W., Johnson, A.J.A., Holmes, S.P., 2016.
DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13,
581–583. https://doi.org/10.1038/nmeth.3869
Canada, P., Pereira, A., Nogueira, N., Png-Gonzalez, L., Andrade, C., Xavier, R., 2020.
Analysis of bacterial microbiome associated with nylon and copper nets in an aquaculture
context. Aquaculture 516, 734540. https://doi.org/10.1016/j.aquaculture.2019.734540
Caporaso, J.G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F.D., Costello, E.K.,
Fierer, N., Pẽa, A.G., Goodrich, J.K., Gordon, J.I., Huttley, G.A., Kelley, S.T., Knights, D.,
Koenig, J.E., Ley, R.E., Lozupone, C.A., McDonald, D., Muegge, B.D., Pirrung, M.,
Reeder, J., Sevinsky, J.R., Turnbaugh, P.J., Walters, W.A., Widmann, J., Yatsunenko,
T., Zaneveld, J., Knight, R., 2010. QIIME allows analysis of high-throughput community
sequencing data. Nat. Methods. https://doi.org/10.1038/nmeth.f.303
Caporaso, J.G., Lauber, C.L., Costello, E.K., Berg-Lyons, D., Gonzalez, A., Stombaugh, J.,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
33
Knights, D., Gajer, P., Ravel, J., Fierer, N., Gordon, J.I., Knight, R., 2011. Moving pictures
of the human microbiome. Genome Biol. 12, R50. https://doi.org/10.1186/gb-2011-12-5-
r50
Carda-Diéguez, M., Mira, A., Fouz, B., 2014. Pyrosequencing survey of intestinal microbiota
diversity in cultured sea bass (Dicentrarchus labrax) fed functional diets. FEMS Microbiol.
Ecol. 87, 451–459. https://doi.org/10.1111/1574-6941.12236
Carillo, M., Zanuy, S., Prat, F., Cerdá, J., Ramos, J., Mañanós, E., Bromage, N, 1995.
Seabass (Dicentrarchus labrax), in: Bromage, NR, Roberts, R. (Eds.), Broodstock
Management and Egg and Larval Quality. Blackwell Science, Oxford, pp. 138–68.
Carlson, J.M., Hyde, E.R., Petrosino, J.F., Manage, A.B.W., Primm, T.P., 2015. The host
effects of Gambusia affinis with an antibiotic-disrupted microbiome. Comp. Biochem.
Physiol. Part - C Toxicol. Pharmacol. 178, 163–168.
https://doi.org/10.1016/j.cbpc.2015.10.004
Carlson, J.M., Leonard, A.B., Hyde, E.R., Petrosino, J.F., Primm, T.P., 2017. Microbiome
disruption and recovery in the fish Gambusia affinis following exposure to broad-
spectrum antibiotic. Infect. Drug Resist. 10, 143–154.
https://doi.org/10.2147/IDR.S129055
Cash, H.L., Whitham, C. V., Behrendt, C.L., Hooper, L. V., 2006. Symbiotic Bacteria Direct
Expression of an Intestinal Bactericidal Lectin. Science (80). 313, 1126–1130.
https://doi.org/10.1126/science.1127119
Cerezuela, R., Fumanal, M., Tapia-Paniagua, S.T., Meseguer, J., Moriñigo, M. ángel,
Esteban, M. ángeles, 2013. Changes in intestinal morphology and microbiota caused by
dietary administration of inulin and Bacillus subtilis in gilthead sea bream (Sparus aurata
L.) specimens. Fish Shellfish Immunol. 34, 1063–1070.
https://doi.org/10.1016/j.fsi.2013.01.015
Cerezuela, R., Meseguer, J., Esteban, M.A., 2011. Current Knowledge in Synbiotic Use for
Fish Aquaculture: A Review. J. Aquac. Res. Dev. S 1, 1–7. https://doi.org/10.4172/2155-
9546.S1-008
Chabrillon, M., Arijo, S., Diaz-Rosales, P., Balebona, M.C., Morinigo, M.A., 2006. Interference
of Listonella anguillarum with potential probiotic microorganisms isolated from farmed
gilthead seabream (Sparus aurata, L.). Aquac. Res. 37, 78–86.
https://doi.org/10.1111/j.1365-2109.2005.01400.x
Chabrillon, M., Rico, R.M., Arijo, S., Diaz-Rosales, P., Balebona, M.C., Morinigo, M.A., 2005.
Interactions of microorganisms isolated from gilthead sea bream, Sparus aurata L., on
Vibrio harveyi, a pathogen of farmed Senegalese sole, Solea senegalensis (Kaup). J.
Fish Dis. 28, 531–537. https://doi.org/10.1111/j.1365-2761.2005.00657.x
Cheesman, S.E., Neal, J.T., Mittge, E., Seredick, B.M., Guillemin, K., 2011. Epithelial cell

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
34
proliferation in the developing zebrafish intestine is regulated by the Wnt pathway and
microbial signaling via Myd88. Proc. Natl. Acad. Sci. U. S. A. 108, 4570–4577.
https://doi.org/10.1073/pnas.1000072107
Chen, L., Guo, Y., Hu, C., Lam, P.K.S., Lam, J.C.W., Zhou, B., 2018. Dysbiosis of gut
microbiota by chronic coexposure to titanium dioxide nanoparticles and bisphenol A:
Implications for host health in zebrafish. Environ. Pollut. 234, 307–317.
https://doi.org/10.1016/j.envpol.2017.11.074
Chen, Y., Zhu, X., Yang, Y., Han, D., Jin, J., Xie, S., 2014. Effect of dietary chitosan on growth
performance, haematology, immune response, intestine morphology, intestine
microbiota and disease resistance in gibel carp (Carassius auratus gibelio). Aquac. Nutr.
20, 532–546. https://doi.org/10.1111/anu.12106
Chervinski, J., 1984. Salinity tolerance of young gilthead sea bream. Bamidgeh 36, 121–124.
Chi, C., Jiang, B., Yu, X.B., Liu, T.Q., Xia, L., Wang, G.X., 2014. Effects of three strains of
intestinal autochthonous bacteria and their extracellular products on the immune
response and disease resistance of common carp, Cyprinus carpio. Fish Shellfish
Immunol. 36, 9–18. https://doi.org/10.1016/j.fsi.2013.10.003
Chiarello, M., Auguet, J.C., Bettarel, Y., Bouvier, C., Claverie, T., Graham, N.A.J.,
Rieuvilleneuve, F., Sucré, E., Bouvier, T., Villéger, S., 2018. Skin microbiome of coral
reef fish is highly variable and driven by host phylogeny and diet. Microbiome 6, 147.
https://doi.org/10.1186/s40168-018-0530-4
Chiarello, M., Paz-Vinas, I., Veyssière, C., Santoul, F., Loot, G., Ferriol, J., Boulêtreau, S.,
2019. Environmental conditions and neutral processes shape the skin microbiome of
European catfish (Silurus glanis) populations of Southwestern France. Environ. Microbiol.
Rep. 11, 605–614. https://doi.org/10.1111/1758-2229.12774
Chiarello, M., Villéger, S., Bouvier, C., Bettarel, Y., Bouvier, T., 2015. High diversity of skin-
associated bacterial communities of marine fishes is promoted by their high variability
among body parts, individuals and species. FEMS Microbiol. Ecol. 91.
https://doi.org/10.1093/femsec/fiv061
Chiu, T.-H., Kao, L.-Y., Chen, M.-L., 2013. Antibiotic resistance and molecular typing of
Photobacterium damselae subsp. damselae, isolated from seafood. J. Appl. Microbiol.
114, 1184–1192. https://doi.org/10.1111/jam.12104
Choi, J., Yang, F., Stepanauskas, R., Cardenas, E., Garoutte, A., Williams, R., Flater, J.,
Tiedje, J.M., Hofmockel, K.S., Gelder, B., Howe, A., 2017. Strategies to improve
reference databases for soil microbiomes. ISME J. 11, 829–834.
https://doi.org/10.1038/ismej.2016.168
Clarke, R., Bostock, J., 2017. Regional Review on Status and Trends in Aquaculture
Development in Europe - 2015.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
35
Cole, J.R., Wang, Q., Cardenas, E., Fish, J., Chai, B., Farris, R.J., Kulam-Syed-Mohideen,
A.S., McGarrell, D.M., Marsh, T., Garrity, G.M., Tiedje, J.M., 2009. The Ribosomal
Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids
Res. 37, 141–145. https://doi.org/10.1093/nar/gkn879
Corkrey, R., Olley, J., Ratkowsky, D., McMeekin, T., Ross, T., 2012. Universality of
thermodynamic constants governing biological growth rates. PLoS One 7, e32003.
https://doi.org/10.1371/journal.pone.0032003
Costello, E.K., Lauber, C.L., Hamady, M., Fierer, N., Gordon, J.I., Knight, R., 2009. Bacterial
Community Variation in Human Body Habitats Across Space and Time. Science (80).
326, 1694–1697. https://doi.org/10.1126/science.1177486
Cruz, P.M., Ibáñez, A.L., Monroy Hermosillo, O.A., Saad, H.C.R., 2012. Use of Probiotics in
Aquaculture. Int. Sch. Res. Netw. ISRN Microbiol. 2012, 13.
https://doi.org/10.5402/2012/916845
David, L.A., Materna, A.C., Friedman, J., Campos-Baptista, M.I., Blackburn, M.C., Perrotta,
A., Erdman, S.E., Alm, E.J., 2014. Host lifestyle affects human microbiota on daily
timescales. Genome Biol. 15, R89. https://doi.org/10.1186/gb-2014-15-7-r89
Davis, D.J., Bryda, E.C., Gillespie, C.H., Ericsson, A.C., 2016. Microbial modulation of
behavior and stress responses in zebrafish larvae. Behav. Brain Res. 311, 219–227.
https://doi.org/10.1016/j.bbr.2016.05.040
de Bruijn, I., Liu, Y., Wiegertjes, G.F., Raaijmakers, J.M., 2018. Exploring fish microbial
communities to mitigate emerging diseases in aquaculture. FEMS Microbiol. Ecol. 94,
161. https://doi.org/10.1093/femsec/fix161
DeBofsky, A., Xie, Y., Jardine, T.D., Hill, J.E., Jones, P.D., Giesy, J.P., 2020. Effects of the
husky oil spill on gut microbiota of native fishes in the North Saskatchewan River,
Canada. Aquat. Toxicol. 229, 105658. https://doi.org/10.1016/j.aquatox.2020.105658
DeSantis, T.Z., Hugenholtz, P., Larsen, N., Rojas, M., Brodie, E.L., Keller, K., Huber, T.,
Dalevi, D., Hu, P., Andersen, G.L., 2006. Greengenes, a chimera-checked 16S rRNA
gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 72, 5069–
5072. https://doi.org/10.1128/AEM.03006-05
Dessinioti, C., Katsambas, A.D., 2010. The role of Propionibacterium acnes in acne
pathogenesis: facts and controversies. Clin. Dermatol. 28, 2–7.
https://doi.org/10.1016/j.clindermatol.2009.03.012
Di Bella, J.M., Bao, Y., Gloor, G.B., Burton, J.P., Reid, G., 2013. High throughput sequencing
methods and analysis for microbiome research. J. Microbiol. Methods 95, 401–414.
https://doi.org/10.1016/j.mimet.2013.08.011
Dimitroglou, A., Moate, R., Janssens, T., Spring, P., Sweetman, J.W., Davies, S.J., 2011. Field
Observations on the Effect of a Mannan Oligosaccharide on Mortality and Intestinal

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
36
Integrity of Sole (Solea senegalensis, Kaup) Infected by Photobacterium damselae
subsp. piscicida. J. Aquac. Res. Dev. https://doi.org/10.4172/2155-9546.S1-013
Dominguez-Bello, M.G., Costello, E.K., Contreras, M., Magris, M., Hidalgo, G., Fierer, N.,
Knight, R., 2010. Delivery mode shapes the acquisition and structure of the initial
microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. U. S. A. 107,
11971–11975. https://doi.org/10.1073/pnas.1002601107
Dong, S., Ding, L., Cao, J., Liu, X., Xu, H., Meng, K., Yu, Y., Wang, Q., Xu, Z., 2019. Viral-
Infected Change of the Digestive Tract Microbiota Associated With Mucosal Immunity in
Teleost Fish. Front. Immunol. 10, 2878. https://doi.org/10.3389/fimmu.2019.02878
Du, F., Li, Y., Tang, Y., Su, S., Yu, J., Yu, F., Li, J., Li, H., Wang, M., Xu, P., 2019. Response
of the gut microbiome of Megalobrama amblycephala to crowding stress. Aquaculture
500, 586–596. https://doi.org/10.1016/j.aquaculture.2018.10.067
Dunn, C.D., Campbell, L.J., Wallace, E.M., Danylchuk, A.J., Cooke, S.J., Shultz, A.D., Black,
B.D., Brownscombe, J.W., Griffin, L.P., Philipp, D.P., Adams, A.J., Goldberg, T.L., 2020.
Bacterial communities on the gills of bonefish (Albula vulpes) in the Florida Keys and The
Bahamas show spatial structure and differential abundance of disease-associated
bacteria 167, 85. https://doi.org/10.1007/s00227-020-03698-7
Dvergedal, H., Sandve, S.R., Angell, I.L., Klemetsdal, G., Rudi, K., 2020. Association of gut
microbiota with metabolism in juvenile Atlantic salmon. Microbiome 8, 160.
https://doi.org/10.1186/s40168-020-00938-2
Ebrahimi, G., Ouraji, H., Khalesi, M.K., Sudagar, M., Barari, A., Zarei Dangesaraki, M., Jani
Khalili, K.H., 2012. Effects of a prebiotic, Immunogen® , on feed utilization, body
composition, immunity and resistance to Aeromonas hydrophila infection in the common
carp Cyprinus carpio (Linnaeus) fingerlings. J. Anim. Physiol. Anim. Nutr. (Berl). 96, 591–
599. https://doi.org/10.1111/j.1439-0396.2011.01182.x
Egerton, S., Culloty, S., Whooley, J., Stanton, C., Ross, R.P., 2018. The gut microbiota of
marine fish. Front. Microbiol. https://doi.org/10.3389/fmicb.2018.00873
Eissa, I.A.M., Derwa, H.I., Ismail, M., El-lamie, M., Dessouki, A.A., Elsheshtawy, H., Bayoumy,
E.M., 2018. Molecular and phenotypic characterization of Photobacterium damselae
among some marine fishes in Lake Temsah. Microb. Pathog. 114, 315–322.
https://doi.org/10.1016/j.micpath.2017.12.006
Emerenciano, M., Gaxiola, G., Cuzon, G., 2013. Biofloc technology (BFT): a review for
aquaculture application and animal food industry., in: Matovic, M.D. (Ed.), Biomass Now:
Cultivation and Utilization. BoD–Books on Demand, pp. 301–328.
Engelbrektson, A., Kunin, V., Wrighton, K.C., Zvenigorodsky, N., Chen, F., Ochman, H.,
Hugenholtz, P., 2010. Experimental factors affecting PCR-based estimates of microbial
species richness and evenness. ISME J. 4, 642–647.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
37
https://doi.org/10.1038/ismej.2009.153
Erickson, A.R., Cantarel, B.L., Lamendella, R., Darzi, Y., Mongodin, E.F., Pan, C., Shah, M.,
Halfvarson, J., Tysk, C., Henrissat, B., Raes, J., Verberkmoes, N.C., Fraser, C.M.,
Hettich, R.L., Jansson, J.K., 2012. Integrated Metagenomics/Metaproteomics Reveals
Human Host-Microbiota Signatures of Crohn’s Disease. PLoS One 7, e49138.
https://doi.org/10.1371/journal.pone.0049138
Espinal, C.A., Matulić, D., 2019. Recirculating Aquaculture Technologies, in: Goddek, S.,
Joyce, A., Kotzen, B., Burnell, G.M. (Eds.), Aquaponics Food Production Systems:
Combined Aquaculture and Hydroponic Production Technologies for the Future. Springer
Open, pp. 35–55.
Essam, H.M., Abdellrazeq, G.S., Tayel, S.I., Torky, H.A., Fadel, A.H., 2016. Pathogenesis of
Photobacterium damselae subspecies infections in sea bass and sea bream. Microb.
Pathog. 99, 41–50. https://doi.org/10.1016/j.micpath.2016.08.003
Falcinelli, S., Picchietti, S., Rodiles, A., Cossignani, L., Merrifield, D.L., Taddei, A.R.,
Maradonna, F., Olivotto, I., Gioacchini, G., Carnevali, O., 2015. Lactobacillus rhamnosus
lowers zebrafish lipid content by changing gut microbiota and host transcription of genes
involved in lipid metabolism. Sci. Rep. 5, 1–11. https://doi.org/10.1038/srep09336
Falcinelli, S., Rodiles, A., Unniappan, S., Picchietti, S., Gioacchini, G., Merrifield, D.L.,
Carnevali, O., 2016. Probiotic treatment reduces appetite and glucose level in the
zebrafish model. Sci. Rep. 6, 1–13. https://doi.org/10.1038/srep18061
FAO, 2018. The state of world fisheries and aquaculture 2018 - Meeting the sustainable
development goals. Rome.
FAO, 2016a. FAO Fisheries & Aquaculture - Cultured Aquatic Species Information
Programme - Dicentrarchus labrax (Linnaeus, 1758) [WWW Document]. URL
http://www.fao.org/fishery/culturedspecies/Dicentrarchus_labrax/en (accessed 2.12.21).
FAO, 2016b. FAO Fisheries & Aquaculture - Cultured Aquatic Species Information
Programme - Sparus aurata (Linnaeus, 1758) [WWW Document]. URL
http://www.fao.org/fishery/culturedspecies/Sparus_aurata/en (accessed 2.12.21).
Fei, N., Zhao, L., 2013. An opportunistic pathogen isolated from the gut of an obese human
causes obesity in germfree mice. ISME J. 7, 880–884.
https://doi.org/10.1038/ismej.2012.153
Felip, A., Piferrer, F., Zanuy, S., Carrillo, M., 2001. Comparative growth performance of diploid
and triploid European sea bass over the first four spawning seasons. J. Fish Biol. 58, 76–
88. https://doi.org/10.1111/j.1095-8649.2001.tb00500.x
Fierer, N., Lauber, C.L., Zhou, N., McDonald, D., Costello, E.K., Knight, R., 2010. Forensic
identification using skin bacterial communities. Proc. Natl. Acad. Sci. U. S. A. 107, 6477–
6481. https://doi.org/10.1073/pnas.1000162107

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
38
Fishelson, L., Montgomery, W.L., Myrberg, A.A., 1985. A unique symbiosis in the gut of
tropical herbivorous surgeonfish (Acanthuridae: teleostei) from the red sea. Science (80).
229, 49–51. https://doi.org/10.1126/science.229.4708.49
Fisher, B., Naidoo, R., Guernier, J., Johnson, K., Mullins, D., Robinson, D., Allison, E.H., 2017.
Integrating fisheries and agricultural programs for food security. Agric. Food Secur. 6, 1.
https://doi.org/10.1186/s40066-016-0078-0
Flores, G.E., Caporaso, J.G., Henley, J.B., Rideout, J.R. a., Domogala, D., Chase, J., Leff,
J.W., Vázquez-Baeza, Y., Gonzalez, A., Knight, R., Dunn, R.R., Fierer, N., 2014.
Temporal variability is a personalized feature of the human microbiome. Genome Biol.
15, 531. https://doi.org/10.1186/s13059-014-0531-y
Frank, D.N., Robertson, C.E., Hamm, C.M., Kpadeh, Z., Zhang, T., Chen, H., Zhu, W., Sartor,
R.B., Boedeker, E.C., Harpaz, N., Pace, N.R., Li, E., 2011. Disease phenotype and
genotype are associated with shifts in intestinal-associated microbiota in inflammatory
bowel diseases. Inflamm. Bowel Dis. 17, 179–184. https://doi.org/10.1002/ibd.21339
Galindo-Villegas, J., Garciá-Moreno, D., De Oliveira, S., Meseguer, J., Mulero, V., 2012.
Regulation of immunity and disease resistance by commensal microbes and chromatin
modifications during zebrafish development. Proc. Natl. Acad. Sci. U. S. A. 109, E2605–
E2614. https://doi.org/10.1073/pnas.1209920109
Galloway-Peña, J., Hanson, B., 2020. Tools for Analysis of the Microbiome. Dig. Dis. Sci.
https://doi.org/10.1007/s10620-020-06091-y
Gao, Z., Tseng, C.H., Pei, Z., Blaser, M.J., 2007. Molecular analysis of human forearm
superficial skin bacterial biota. Proc. Natl. Acad. Sci. U. S. A. 104, 2927–2932.
https://doi.org/10.1073/pnas.0607077104
García de la Banda, I., Lobo, C., Chabrillón, M., León-Rubio, J.M., Arijo, S., Pazos, G., María
Lucas, L., Moriñigo, M.Á., 2012. Influence of dietary administration of a probiotic strain
Shewanella putrefaciens on senegalese sole (Solea senegalensis, Kaup 1858) growth,
body composition and resistance to Photobacterium damselae subsp piscicida. Aquac.
Res. 43, 662–669. https://doi.org/10.1111/j.1365-2109.2011.02871.x
Gaulke, C.A., Barton, C.L., Proffitt, S., Tanguay, R.L., Sharpton, T.J., 2016. Triclosan
Exposure Is Associated with Rapid Restructuring of the Microbiome in Adult Zebrafish.
PLoS One 11, e0154632. https://doi.org/10.1371/journal.pone.0154632
Ghanbari, M., Kneifel, W., Domig, K.J., 2015. A new view of the fish gut microbiome: Advances
from next-generation sequencing. Aquaculture.
https://doi.org/10.1016/j.aquaculture.2015.06.033
Gibson, G.R., Roberfroid, M.B., 1995. Dietary modulation of the human colonic microbiota:
Introducing the concept of prebiotics. J. Nutr. https://doi.org/10.1093/jn/125.6.1401
Gilbert, J.A., Blaser, M.J., Caporaso, J.G., Jansson, J.K., Lynch, S. V., Knight, R., 2018.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
39
Current understanding of the human microbiome. Nat. Med. 24, 392–400.
https://doi.org/10.1038/nm.4517
Giorgia, G., Elia, C., Andrea, P., Cinzia, C., Stefania, S., Ana, R., Daniel, M.L., Ike, O., Oliana,
C., 2018. Effects of Lactogen 13, a New Probiotic Preparation, on Gut Microbiota and
Endocrine Signals Controlling Growth and Appetite of Oreochromis niloticus Juveniles.
Microb. Ecol. 76, 1063–1074. https://doi.org/10.1007/s00248-018-1177-1
Giri, S.S., Sen, S.S., Sukumaran, V., 2012. Effects of dietary supplementation of potential
probiotic Pseudomonas aeruginosa VSG-2 on the innate immunity and disease
resistance of tropical freshwater fish, Labeo rohita. Fish Shellfish Immunol. 32, 1135–
1140. https://doi.org/10.1016/j.fsi.2012.03.019
Givens, C., Ransom, B., Bano, N., Hollibaugh, J., 2015. Comparison of the gut microbiomes
of 12 bony fish and 3 shark species. Mar. Ecol. Prog. Ser. 518, 209–223.
https://doi.org/10.3354/meps11034
Gomez, D., Sunyer, J.O., Salinas, I., 2013. The mucosal immune system of fish: The evolution
of tolerating commensals while fighting pathogens. Fish Shellfish Immunol.
https://doi.org/10.1016/j.fsi.2013.09.032
Goodrich, J.K., Di Rienzi, S.C., Poole, A.C., Koren, O., Walters, W.A., Caporaso, J.G., Knight,
R., Ley, R.E., 2014. Conducting a microbiome study. Cell.
https://doi.org/10.1016/j.cell.2014.06.037
Gopalakrishnan, V., Spencer, C.N., Nezi, L., Reuben, A., Andrews, M.C., Karpinets, T. V.,
Prieto, P.A., Vicente, D., Hoffman, K., Wei, S.C., Cogdill, A.P., Zhao, L., Hudgens, C.W.,
Hutchinson, D.S., Manzo, T., Petaccia De Macedo, M., Cotechini, T., Kumar, T., Chen,
W.S., Reddy, S.M., Szczepaniak Sloane, R., Galloway-Pena, J., Jiang, H., Chen, P.L.,
Shpall, E.J., Rezvani, K., Alousi, A.M., Chemaly, R.F., Shelburne, S., Vence, L.M.,
Okhuysen, P.C., Jensen, V.B., Swennes, A.G., McAllister, F., Marcelo Riquelme
Sanchez, E., Zhang, Y., Le Chatelier, E., Zitvogel, L., Pons, N., Austin-Breneman, J.L.,
Haydu, L.E., Burton, E.M., Gardner, J.M., Sirmans, E., Hu, J., Lazar, A.J., Tsujikawa, T.,
Diab, A., Tawbi, H., Glitza, I.C., Hwu, W.J., Patel, S.P., Woodman, S.E., Amaria, R.N.,
Davies, M.A., Gershenwald, J.E., Hwu, P., Lee, J.E., Zhang, J., Coussens, L.M., Cooper,
Z.A., Futreal, P.A., Daniel, C.R., Ajami, N.J., Petrosino, J.F., Tetzlaff, M.T., Sharma, P.,
Allison, J.P., Jenq, R.R., Wargo, J.A., 2018. Gut microbiome modulates response to anti-
PD-1 immunotherapy in melanoma patients. Science (80). 359, 97–103.
https://doi.org/10.1126/science.aan4236
Greenspan, S.E., Migliorini, G.H., Lyra, M.L., Pontes, M.R., Carvalho, T., Ribeiro, L.P., Moura-
Campos, D., Haddad, C.F.B., Toledo, L.F., Romero, G.Q., Becker, C.G., 2020. Warming
drives ecological community changes linked to host-associated microbiome dysbiosis.
Nat. Clim. Chang. 10, 1057–1061. https://doi.org/10.1038/s41558-020-0899-5

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
40
Grice, E.A., Kong, H.H., Renaud, G., Young, A.C., Bouffard, G.G., Blakesley, R.W., Wolfsberg,
T.G., Turner, M.L., Segre, J.A., 2008. A diversity profile of the human skin microbiota.
Genome Res. 18, 1043–1050. https://doi.org/10.1101/gr.075549.107
Grice, E.A., Segre, J.A., 2011. The skin microbiome. Nat. Rev. Microbiol.
https://doi.org/10.1038/nrmicro2537
Guerreiro, I., Oliva-Teles, A., Enes, P., 2018. Prebiotics as functional ingredients: focus on
Mediterranean fish aquaculture. Rev. Aquac. 10, 800–832.
https://doi.org/10.1111/raq.12201
Guijarro, J.A., Cascales, D., García-Torrico, A.I., García-Domínguez, M., Méndez, J., 2015.
Temperature-dependent expression of virulence genes in fish-pathogenic bacteria.
Front. Microbiol. https://doi.org/10.3389/fmicb.2015.00700
Guivier, E., Pech, N., Chappaz, R., Gilles, A., 2020. Microbiota associated with the skin, gills,
and gut of the fish Parachondrostoma toxostoma from the Rhône basin. Freshw. Biol. 65,
446–459. https://doi.org/10.1111/fwb.13437
Gupta, S., Fernandes, J., Kiron, V., 2019. Antibiotic-Induced Perturbations Are Manifested in
the Dominant Intestinal Bacterial Phyla of Atlantic Salmon. Microorganisms 7, 233.
https://doi.org/10.3390/microorganisms7080233
Habiba, M.E., Tayel, S.I., Torky, H.A., 2015. Occurrence and Characterization of
Photobacteria damselae in Cultured Marine Fish. Alexandria J. Vet. Sci. 44, 103–110.
https://doi.org/10.5455/ajvs.129
Hai, N.V., 2015. The use of probiotics in aquaculture. J. Appl. Microbiol. 119, 917–935.
https://doi.org/10.1111/jam.12886
Hall, L.J., Robinson, S.D., 2021. Bacterial strains augment cancer therapeutics. Nat. Microbiol.
6, 275–276. https://doi.org/10.1038/s41564-021-00871-6
Handelsman, J., Rondon, M.R., Brady, S.F., Clardy, J., Goodman, R.M., 1998. Molecular
biological access to the chemistry of unknown soil microbes: A new frontier for natural
products. Chem. Biol. 5, R245–R249. https://doi.org/10.1016/S1074-5521(98)90108-9
Hassaan, M.S., Mohammady, E.Y., Soaudy, M.R., Palma, J., Shawer, E.E., El-Haroun, E.,
2020. The effect of dietary sericite on growth performance, digestive enzymes activity,
gut microbiota and haematological parameters of Nile tilapia, Oreochromis niloticus (L.)
fingerlings. Anim. Feed Sci. Technol. 262, 114400.
https://doi.org/10.1016/j.anifeedsci.2020.114400
He, S., Ran, C., Qin, C., Li, S., Zhang, H., De Vos, W.M., Ringø, E., Zhou, Z., 2017a. Anti-
Infective Effect of Adhesive Probiotic Lactobacillus in Fish is Correlated with Their Spatial
Distribution in the Intestinal Tissue. Sci. Rep. 7, 1–12. https://doi.org/10.1038/s41598-
017-13466-1
He, S., Wang, Q., Li, S., Ran, C., Guo, X., Zhang, Z., Zhou, Z., 2017b. Antibiotic growth

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
41
promoter olaquindox increases pathogen susceptibility in fish by inducing gut microbiota
dysbiosis. Sci. China Life Sci. 60, 1260–1270. https://doi.org/10.1007/s11427-016-9072-
6
Heintz-Buschart, A., Wilmes, P., 2018. Human Gut Microbiome: Function Matters. Trends
Microbiol. https://doi.org/10.1016/j.tim.2017.11.002
Heintz, C., Mair, W., 2014. You are what you host: Microbiome modulation of the aging
process. Cell. https://doi.org/10.1016/j.cell.2014.01.025
Hennersdorf, P., Mrotzek, G., Abdul-Aziz, M.A., Saluz, H.P., 2016. Metagenomic analysis
between free-living and cultured Epinephelus fuscoguttatus under different
environmental conditions in Indonesian waters. Mar. Pollut. Bull. 110, 726–734.
https://doi.org/10.1016/j.marpolbul.2016.05.009
Hess, S., Wenger, A.S., Ainsworth, T.D., Rummer, J.L., 2015. Exposure of clownfish larvae to
suspended sediment levels found on the Great Barrier Reef: Impacts on gill structure and
microbiome. Sci. Rep. 5, 1–8. https://doi.org/10.1038/srep10561
Hickey, R.J., Zhou, X., Pierson, J.D., Ravel, J., Forney, L.J., 2012. Understanding vaginal
microbiome complexity from an ecological perspective. Transl. Res.
https://doi.org/10.1016/j.trsl.2012.02.008
Hildonen, M., Kodama, M., Puetz, L.C., Gilbert, M.T.P., Limborg, M.T., 2019. A comparison of
storage methods for gut microbiome studies in teleosts: Insights from rainbow trout
(Oncorhynchus mykiss). J. Microbiol. Methods 160, 42–48.
https://doi.org/10.1016/j.mimet.2019.03.010
Hill, C., Guarner, F., Reid, G., Gibson, G.R., Merenstein, D.J., Pot, B., Morelli, L., Canani,
R.B., Flint, H.J., Salminen, S., Calder, P.C., Sanders, M.E., 2014. Expert consensus
document: The international scientific association for probiotics and prebiotics consensus
statement on the scope and appropriate use of the term probiotic. Nat. Rev.
Gastroenterol. Hepatol. 11, 506–514. https://doi.org/10.1038/nrgastro.2014.66
Hill, C.J., Brown, J.R.M., Lynch, D.B., Jeffery, I.B., Ryan, C.A., Ross, R.P., Stanton, C.,
O’Toole, P.W., 2016. Effect of room temperature transport vials on DNA quality and
phylogenetic composition of faecal microbiota of elderly adults and infants. Microbiome
4, 1–10. https://doi.org/10.1186/s40168-016-0164-3
Hong, B., Ba, Y., Niu, L., Lou, F., Zhang, Z., Liu, H., Pan, Y., Zhao, Y., 2018. A Comprehensive
Research on Antibiotic Resistance Genes in Microbiota of Aquatic Animals. Front.
Microbiol. 9, 1617. https://doi.org/10.3389/fmicb.2018.01617
Horlick, J., Booth, M.A., Tetu, S.G., 2020. Alternative dietary protein and water temperature
influence the skin and gut microbial communities of yellowtail kingfish (Seriola lalandi).
PeerJ 2020, e8705. https://doi.org/10.7717/peerj.8705
Hoseinifar, S.H., Ringø, E., Shenavar Masouleh, A., Esteban, M.Á., 2016. Probiotic, prebiotic

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
42
and synbiotic supplements in sturgeon aquaculture: a review. Rev. Aquac. 8, 89–102.
https://doi.org/10.1111/raq.12082
Hoseinifar, S.H., Sharifian, M., Vesaghi, M.J., Khalili, M., Esteban, M.Á., 2014. The effects of
dietary xylooligosaccharide on mucosal parameters, intestinal microbiota and
morphology and growth performance of Caspian white fish (Rutilus frisii kutum) fry. Fish
Shellfish Immunol. 39, 231–236. https://doi.org/10.1016/j.fsi.2014.05.009
Hoseinifar, S.H., Sun, Y.Z., Wang, A., Zhou, Z., 2018. Probiotics as means of diseases control
in aquaculture, a review of current knowledge and future perspectives. Front. Microbiol.
https://doi.org/10.3389/fmicb.2018.02429
Hossain, S., Shefat, T., Karim, A., 2018. Nutritional Diseases of Fish in Aquaculture and Their
Management: A Review. Acta Sci. Pharm. Sci. 50–58.
Hovda, M.B., Fontanillas, R., Mcgurk, C., Obach, A., Rosnes, J.T., 2012. Seasonal variations
in the intestinal microbiota of farmed Atlantic salmon (Salmo salar L.) 43, 154–159.
https://doi.org/10.1111/j.1365-2109.2011.02805.x
Hsiao, E.Y., McBride, S.W., Hsien, S., Sharon, G., Hyde, E.R., McCue, T., Codelli, J.A., Chow,
J., Reisman, S.E., Petrosino, J.F., Patterson, P.H., Mazmanian, S.K., 2013. Microbiota
modulate behavioral and physiological abnormalities associated with
neurodevelopmental disorders. Cell 155, 1451–1463.
https://doi.org/10.1016/j.cell.2013.11.024
Huang, L., Zuo, Y., Jiang, Q., Su, Y., Qin, Y., Xu, X., Zhao, L., Yan, Q., 2019. A metabolomic
investigation into the temperature-dependent virulence of Pseudomonas plecoglossicida
from large yellow croaker (Pseudosciaena crocea). J. Fish Dis. 42, 431–446.
https://doi.org/10.1111/jfd.12957
Huttenhower, C., Gevers, D., Knight, R., Abubucker, S., Badger, J.H., Chinwalla, A.T., Creasy,
H.H., Earl, A.M., Fitzgerald, M.G., Fulton, R.S., Giglio, M.G., Hallsworth-Pepin, K., Lobos,
E.A., Madupu, R., Magrini, V., Martin, J.C., Mitreva, M., Muzny, D.M., Sodergren, E.J.,
Versalovic, J., Wollam, A.M., Worley, K.C., Wortman, J.R., Young, S.K., Zeng, Q.,
Aagaard, K.M., Abolude, O.O., Allen-Vercoe, E., Alm, E.J., Alvarado, L., Andersen, G.L.,
Anderson, S., Appelbaum, E., Arachchi, H.M., Armitage, G., Arze, C.A., Ayvaz, T., Baker,
C.C., Begg, L., Belachew, T., Bhonagiri, V., Bihan, M., Blaser, M.J., Bloom, T., Bonazzi,
V., Paul Brooks, J., Buck, G.A., Buhay, C.J., Busam, D.A., Campbell, J.L., Canon, S.R.,
Cantarel, B.L., Chain, P.S.G., Chen, I.M.A., Chen, L., Chhibba, S., Chu, K., Ciulla, D.M.,
Clemente, J.C., Clifton, S.W., Conlan, S., Crabtree, J., Cutting, M.A., Davidovics, N.J.,
Davis, C.C., Desantis, T.Z., Deal, C., Delehaunty, K.D., Dewhirst, F.E., Deych, E., Ding,
Y., Dooling, D.J., Dugan, S.P., Michael Dunne, W., Scott Durkin, A., Edgar, R.C., Erlich,
R.L., Farmer, C.N., Farrell, R.M., Faust, K., Feldgarden, M., Felix, V.M., Fisher, S., Fodor,
A.A., Forney, L.J., Foster, L., Di Francesco, V., Friedman, J., Friedrich, D.C., Fronick,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
43
C.C., Fulton, L.L., Gao, H., Garcia, N., Giannoukos, G., Giblin, C., Giovanni, M.Y.,
Goldberg, J.M., Goll, J., Gonzalez, A., Griggs, A., Gujja, S., Kinder Haake, S., Haas, B.J.,
Hamilton, H.A., Harris, E.L., Hepburn, T.A., Herter, B., Hoffmann, D.E., Holder, M.E.,
Howarth, C., Huang, K.H., Huse, S.M., Izard, J., Jansson, J.K., Jiang, H., Jordan, C.,
Joshi, V., Katancik, J.A., Keitel, W.A., Kelley, S.T., Kells, C., King, N.B., Knights, D.,
Kong, H.H., Koren, O., Koren, S., Kota, K.C., Kovar, C.L., Kyrpides, N.C., La Rosa, P.S.,
Lee, S.L., Lemon, K.P., Lennon, N., Lewis, C.M., Lewis, L., Ley, R.E., Li, K., Liolios, K.,
Liu, B., Liu, Y., Lo, C.C., Lozupone, C.A., Dwayne Lunsford, R., Madden, T., Mahurkar,
A.A., Mannon, P.J., Mardis, E.R., Markowitz, V.M., Mavromatis, K., McCorrison, J.M.,
McDonald, D., McEwen, J., McGuire, A.L., McInnes, P., Mehta, T., Mihindukulasuriya,
K.A., Miller, J.R., Minx, P.J., Newsham, I., Nusbaum, C., Oglaughlin, M., Orvis, J.,
Pagani, I., Palaniappan, K., Patel, S.M., Pearson, M., Peterson, J., Podar, M., Pohl, C.,
Pollard, K.S., Pop, M., Priest, M.E., Proctor, L.M., Qin, X., Raes, J., Ravel, J., Reid, J.G.,
Rho, M., Rhodes, R., Riehle, K.P., Rivera, M.C., Rodriguez-Mueller, B., Rogers, Y.H.,
Ross, M.C., Russ, C., Sanka, R.K., Sankar, P., Fah Sathirapongsasuti, J., Schloss, J.A.,
Schloss, P.D., Schmidt, T.M., Scholz, M., Schriml, L., Schubert, A.M., Segata, N., Segre,
J.A., Shannon, W.D., Sharp, R.R., Sharpton, T.J., Shenoy, N., Sheth, N.U., Simone,
G.A., Singh, I., Smillie, C.S., Sobel, J.D., Sommer, D.D., Spicer, P., Sutton, G.G., Sykes,
S.M., Tabbaa, D.G., Thiagarajan, M., Tomlinson, C.M., Torralba, M., Treangen, T.J.,
Truty, R.M., Vishnivetskaya, T.A., Walker, J., Wang, L., Wang, Z., Ward, D. V., Warren,
W., Watson, M.A., Wellington, C., Wetterstrand, K.A., White, J.R., Wilczek-Boney, K.,
Wu, Y., Wylie, K.M., Wylie, T., Yandava, C., Ye, L., Ye, Y., Yooseph, S., Youmans, B.P.,
Zhang, L., Zhou, Y., Zhu, Y., Zoloth, L., Zucker, J.D., Birren, B.W., Gibbs, R.A.,
Highlander, S.K., Methé, B.A., Nelson, K.E., Petrosino, J.F., Weinstock, G.M., Wilson,
R.K., White, O., 2012. Structure, function and diversity of the healthy human microbiome.
Nature 486, 207–214. https://doi.org/10.1038/nature11234
Huynh, T.G., Shiu, Y.L., Nguyen, T.P., Truong, Q.P., Chen, J.C., Liu, C.H., 2017. Current
applications, selection, and possible mechanisms of actions of synbiotics in improving
the growth and health status in aquaculture: A review. Fish Shellfish Immunol. 64, 367–
382. https://doi.org/10.1016/j.fsi.2017.03.035
Ibrahem, M.D., Fathi, M., Mesalhy, S., Abd El-Aty, A.M., 2010. Effect of dietary
supplementation of inulin and vitamin C on the growth, hematology, innate immunity, and
resistance of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 29, 241–246.
https://doi.org/10.1016/j.fsi.2010.03.004
Infante-Villamil, S., Huerlimann, R., Jerry, D.R., 2020. Microbiome diversity and dysbiosis in
aquaculture. Rev. Aquac. 1–20. https://doi.org/10.1111/raq.12513
Jangi, S., Gandhi, R., Cox, L.M., Li, N., Von Glehn, F., Yan, R., Patel, B., Mazzola, M.A., Liu,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
44
S., Glanz, B.L., Cook, S., Tankou, S., Stuart, F., Melo, K., Nejad, P., Smith, K., Topçuolu,
B.D., Holden, J., Kivisäkk, P., Chitnis, T., De Jager, P.L., Quintana, F.J., Gerber, G.K.,
Bry, L., Weiner, H.L., 2016. Alterations of the human gut microbiome in multiple sclerosis.
Nat. Commun. 7, 1–11. https://doi.org/10.1038/ncomms12015
Jiang, C., Li, G., Huang, P., Liu, Z., Zhao, B., 2017. The Gut Microbiota and Alzheimer’s
Disease. J. Alzheimer’s Dis. https://doi.org/10.3233/JAD-161141
Jin, Y., Wu, S., Zeng, Z., Fu, Z., 2017. Effects of environmental pollutants on gut microbiota.
Environ. Pollut. https://doi.org/10.1016/j.envpol.2016.11.045
Jin, Y., Xia, J., Pan, Z., Yang, J., Wang, W., Fu, Z., 2018. Polystyrene microplastics induce
microbiota dysbiosis and inflammation in the gut of adult zebrafish. Environ. Pollut. 235,
322–329. https://doi.org/10.1016/j.envpol.2017.12.088
Jones, J., DiBattista, J.D., Stat, M., Bunce, M., Boyce, M.C., Fairclough, D. V., Travers, M.J.,
Huggett, M.J., 2018. The Microbiome of the Gastrointestinal Tract of a Range-Shifting
Marine Herbivorous Fish. Front. Microbiol. 9, 2000.
https://doi.org/10.3389/fmicb.2018.02000
Kamada, N., Chen, G.Y., Inohara, N., Núñez, G., 2013. Control of pathogens and pathobionts
by the gut microbiota. Nat. Immunol. https://doi.org/10.1038/ni.2608
Kanehisa, M., Sato, Y., Furumichi, M., Morishima, K., Tanabe, M., 2019. New approach for
understanding genome variations in KEGG. Nucleic Acids Res. 47, D590–D595.
https://doi.org/10.1093/nar/gky962
Kanther, M., Sun, X., Mhlbauer, M., MacKey, L.C., Flynn, E.J., Bagnat, M., Jobin, C., Rawls,
J.F., 2011. Microbial colonization induces dynamic temporal and spatial patterns of NF-
κB activation in the zebrafish digestive tract. Gastroenterology 141, 197–207.
https://doi.org/10.1053/j.gastro.2011.03.042
Kayansamruaj, P., Pirarat, N., Hirono, I., Rodkhum, C., 2014. Increasing of temperature
induces pathogenicity of Streptococcus agalactiae and the up-regulation of inflammatory
related genes in infected Nile tilapia (Oreochromis niloticus). Vet. Microbiol. 172, 265–
271. https://doi.org/10.1016/j.vetmic.2014.04.013
Kelly, B.J., Gross, R., Bittinger, K., Sherrill-Mix, S., Lewis, J.D., Collman, R.G., Bushman, F.D.,
Li, H., 2015. Power and sample-size estimation for microbiome studies using pairwise
distances and PERMANOVA. Bioinformatics 31, 2461–2468.
https://doi.org/10.1093/bioinformatics/btv183
Kelly, C., Salinas, I., 2017. Under pressure: Interactions between commensal microbiota and
the teleost immune system. Front. Immunol. https://doi.org/10.3389/fimmu.2017.00559
Khider, M., Willassen, N.P., Hansen, H., 2018. The alternative sigma factor RpoQ regulates
colony morphology, biofilm formation and motility in the fish pathogen Aliivibrio
salmonicida. BMC Microbiol. 18, 1–13. https://doi.org/10.1186/s12866-018-1258-9

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
45
Kim, A., Kim, N., Roh, H.J., Chun, W.K., Ho, D.T., Lee, Y., Kim, D.H., 2019. Administration of
antibiotics can cause dysbiosis in fish gut. Aquaculture 512, 734330.
https://doi.org/10.1016/j.aquaculture.2019.734330
Kim, D., Hofstaedter, C.E., Zhao, C., Mattei, L., Tanes, C., Clarke, E., Lauder, A., Sherrill-Mix,
S., Chehoud, C., Kelsen, J., Conrad, M., Collman, R.G., Baldassano, R., Bushman, F.D.,
Bittinger, K., 2017. Optimizing methods and dodging pitfalls in microbiome research.
Microbiome. https://doi.org/10.1186/s40168-017-0267-5
Kır, M., 2020. Thermal tolerance and standard metabolic rate of juvenile gilthead seabream
(Sparus aurata) acclimated to four temperatures. J. Therm. Biol. 93, 102739.
https://doi.org/10.1016/j.jtherbio.2020.102739
Knight, R., Vrbanac, A., Taylor, B.C., Aksenov, A., Callewaert, C., Debelius, J., Gonzalez, A.,
Kosciolek, T., McCall, L.I., McDonald, D., Melnik, A. V., Morton, J.T., Navas, J., Quinn,
R.A., Sanders, J.G., Swafford, A.D., Thompson, L.R., Tripathi, A., Xu, Z.Z., Zaneveld,
J.R., Zhu, Q., Caporaso, J.G., Dorrestein, P.C., 2018. Best practices for analysing
microbiomes. Nat. Rev. Microbiol. https://doi.org/10.1038/s41579-018-0029-9
Kokou, F., Sasson, G., Nitzan, T., Doron-Faigenboim, A., Harpaz, S., Cnaani, A., Mizrahi, I.,
2018. Host genetic selection for cold tolerance shapes microbiome composition and
modulates its response to temperature. Elife 7. https://doi.org/10.7554/eLife.36398
Kong, H.H., Oh, J., Deming, C., Conlan, S., Grice, E.A., Beatson, M.A., Nomicos, E., Polley,
E.C., Komarow, H.D., Mullikin, J., Thomas, J., Blakesley, R., Young, A., Chu, G.,
Ramsahoye, C., Lovett, S., Han, J., Legaspi, R., Sison, C., Montemayor, C., Gregory, M.,
Hargrove, A., Johnson, T., Riebow, N., Schmidt, B., Novotny, B., Gupta, J., Benjamin, B.,
Brooks, S., Coleman, H., Ho, S.L., Schandler, K., Stantripop, M., Maduro, Q., Bouffard,
G., Dekhtyar, M., Guan, X., Masiello, C., Maskeri, B., McDowell, J., Park, M., Vemulapalli,
M., Murray, P.R., Turner, M.L., Segre, J.A., 2012. Temporal shifts in the skin microbiome
associated with disease flares and treatment in children with atopic dermatitis. Genome
Res. 22, 850–859. https://doi.org/10.1101/gr.131029.111
Koo, H., Hakim, J.A., Powell, M.L., Kumar, R., Eipers, P.G., Morrow, C.D., Crowley, M.,
Lefkowitz, E.J., Watts, S.A., Bej, A.K., 2017. Metagenomics approach to the study of the
gut microbiome structure and function in zebrafish Danio rerio fed with gluten formulated
diet. J. Microbiol. Methods 135, 69–76. https://doi.org/10.1016/j.mimet.2017.01.016
Koren, O., Goodrich, J.K., Cullender, T.C., Spor, A., Laitinen, K., Kling Bäckhed, H., Gonzalez,
A., Werner, J.J., Angenent, L.T., Knight, R., Bäckhed, F., Isolauri, E., Salminen, S., Ley,
R.E., 2012. Host remodeling of the gut microbiome and metabolic changes during
pregnancy. Cell 150, 470–480. https://doi.org/10.1016/j.cell.2012.07.008
Krotman, Y., Yergaliyev, T.M., Alexander Shani, R., Avrahami, Y., Szitenberg, A., 2020.
Dissecting the factors shaping fish skin microbiomes in a heterogeneous inland water

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
46
system. Microbiome 8, 9. https://doi.org/10.1186/s40168-020-0784-5
Kruskal, W.H., Wallis, W.A., 1952. Use of Ranks in One-Criterion Variance Analysis. J. Am.
Stat. Assoc. 47, 583–621. https://doi.org/10.1080/01621459.1952.10483441
Kuczynski, J., Lauber, C.L., Walters, W.A., Parfrey, L.W., Clemente, J.C., Gevers, D., Knight,
R., 2012. Experimental and analytical tools for studying the human microbiome. Nat. Rev.
Genet. https://doi.org/10.1038/nrg3129
Kundu, P., Blacher, E., Elinav, E., Pettersson, S., 2017. Our Gut Microbiome: The Evolving
Inner Self. Cell. https://doi.org/10.1016/j.cell.2017.11.024
Lafferty, K.D., Harvell, C.D., Conrad, J.M., Friedman, C.S., Kent, M.L., Kuris, A.M., Powell,
E.N., Rondeau, D., Saksida, S.M., 2015. Infectious Diseases Affect Marine Fisheries and
Aquaculture Economics. Ann. Rev. Mar. Sci. 7, 471–496.
https://doi.org/10.1146/annurev-marine-010814-015646
Lane, A., Hough, C., Bostock, J., 2014. The Long Term Economic and Ecological Impact of
Larger Sustainable Aquaculture.
Langille, M.G.I., Zaneveld, J., Caporaso, J.G., McDonald, D., Knights, D., Reyes, J.A.,
Clemente, J.C., Burkepile, D.E., Vega Thurber, R.L., Knight, R., Beiko, R.G.,
Huttenhower, C., 2013. Predictive functional profiling of microbial communities using 16S
rRNA marker gene sequences. Nat. Biotechnol. 31, 814–821.
https://doi.org/10.1038/nbt.2676
Larsen, A.M., Bullard, S.A., Womble, M., Arias, C.R., 2015a. Community Structure of Skin
Microbiome of Gulf Killifish, Fundulus grandis, Is Driven by Seasonality and Not Exposure
to Oiled Sediments in a Louisiana Salt Marsh. Microb. Ecol. 70, 534–544.
https://doi.org/10.1007/s00248-015-0578-7
Larsen, A.M., Mohammed, H.H., Arias, C.R., 2015b. Comparison of DNA extraction protocols
for the analysis of gut microbiota in fishes. FEMS Microbiol. Lett. 362, 31.
https://doi.org/10.1093/femsle/fnu031
Lauber, C.L., Zhou, N., Gordon, J.I., Knight, R., Fierer, N., 2010. Effect of storage conditions
on the assessment of bacterial community structure in soil and human-associated
samples. FEMS Microbiol. Lett. 307, 80–86. https://doi.org/10.1111/j.1574-
6968.2010.01965.x
Laukens, D., Brinkman, B.M., Raes, J., De Vos, M., Vandenabeele, P., 2015. Heterogeneity
of the gut microbiome in mice: Guidelines for optimizing experimental design. FEMS
Microbiol. Rev. https://doi.org/10.1093/femsre/fuv036
Le Luyer, J., Schull, Q., Auffret, P., Lopez, P., Crusot, M., Belliard, C., Basset, C., Carradec,
Q., Poulain, J., Planes, S., Saulnier, D., 2021. Dual RNAseq Highlights the Kinetics of
Skin Microbiome and Fish Host Responsiveness to Bacterial Infection.
https://doi.org/10.21203/rs.3.rs-108248/v1

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
47
Le, M.H., Wang, D., 2020. Structure and membership of gut microbial communities in multiple
fish cryptic species under potential migratory effects. Sci. Rep. 10, 1–12.
https://doi.org/10.1038/s41598-020-64570-8
LeBlanc, J.G., Milani, C., de Giori, G.S., Sesma, F., van Sinderen, D., Ventura, M., 2013.
Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin.
Biotechnol. https://doi.org/10.1016/j.copbio.2012.08.005
Legrand, T.P.R.A., Catalano, S.R., Wos-Oxley, M.L., Stephens, F., Landos, M., Bansemer,
M.S., Stone, D.A.J., Qin, J.G., Oxley, A.P.A., 2018. The Inner Workings of the Outer
Surface: Skin and Gill Microbiota as Indicators of Changing Gut Health in Yellowtail
Kingfish. Front. Microbiol. 8, 2664. https://doi.org/10.3389/fmicb.2017.02664
Legrand, T.P.R.A., Catalano, S.R., Wos-Oxley, M.L., Wynne, J.W., Weyrich, L.S., Oxley,
A.P.A., 2020a. Antibiotic-induced alterations and repopulation dynamics of yellowtail
kingfish microbiota. Anim. Microbiome 2, 26. https://doi.org/10.1186/s42523-020-00046-
4
Legrand, T.P.R.A., Wynne, J.W., Weyrich, L.S., Oxley, A.P.A., 2020b. A microbial sea of
possibilities: current knowledge and prospects for an improved understanding of the fish
microbiome. Rev. Aquac. 12, 1101–1134. https://doi.org/10.1111/raq.12375
León-Zayas, R., McCargar, M., Drew, J.A., Biddle, J.F., 2020. Microbiomes of fish, sediment
and seagrass suggest connectivity of coral reef microbial populations. PeerJ 8, e10026.
https://doi.org/10.7717/peerj.10026
Leone, V., Gibbons, S.M., Martinez, K., Hutchison, A.L., Huang, E.Y., Cham, C.M., Pierre,
J.F., Heneghan, A.F., Nadimpalli, A., Hubert, N., Zale, E., Wang, Y., Huang, Y., Theriault,
B., Dinner, A.R., Musch, M.W., Kudsk, K.A., Prendergast, B.J., Gilbert, J.A., Chang, E.B.,
2015. Effects of diurnal variation of gut microbes and high-fat feeding on host circadian
clock function and metabolism. Cell Host Microbe 17, 681–689.
https://doi.org/10.1016/j.chom.2015.03.006
Li, D., Liu, C.-M., Luo, R., Sadakane, K., Lam, T.-W., 2015. MEGAHIT: an ultra-fast single-
node solution for large and complex metagenomics assembly via succinct de Bruijn
graph. Bioinformatics 31, 1674–1676. https://doi.org/10.1093/bioinformatics/btv033
Li, H., Wu, S., Wirth, S., Hao, Y., Wang, W., Zou, H., Li, W., Wang, G., 2016. Diversity and
activity of cellulolytic bacteria, isolated from the gut contents of grass carp
(Ctenopharyngodon idellus) (Valenciennes) fed on Sudan grass (Sorghum sudanense)
or artificial feedstuffs. Aquac. Res. 47, 153–164. https://doi.org/10.1111/are.12478
Li, H., Zheng, Z., Cong-xin, X., Bo, H., Chao-yuan, W., Gang, H., 2008. Isolation of cellulose—
producing microbes from the intestine of grass carp (Ctenopharyngodon idellus).
Springer, Dordrecht, pp. 131–135. https://doi.org/10.1007/978-90-481-3458-8_19
Li, X.M., Zhu, Y.J., Yan, Q.Y., Ringø, E., Yang, D.G., 2014. Do the intestinal microbiotas differ

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
48
between paddlefish (Polyodon spathala) and bighead carp (Aristichthys nobilis) reared in
the same pond? J. Appl. Microbiol. 117, 1245–1252. https://doi.org/10.1111/jam.12626
Liang, X., Bushman, F.D., Fitzgerald, G.A., 2014. Time in motion: The molecular clock meets
the microbiome. Cell. https://doi.org/10.1016/j.cell.2014.10.020
Lin, G., Zheng, M., Li, S., Xie, J., Fang, W., Gao, D., Huang, J., Lu, J., 2020. Response of gut
microbiota and immune function to hypoosmotic stress in the yellowfin seabream
(Acanthopagrus latus). Sci. Total Environ. 745, 140976.
https://doi.org/10.1016/j.scitotenv.2020.140976
Lindahl, T., 1993. Instability and decay of the primary structure of DNA. Nature.
https://doi.org/10.1038/362709a0
Liu, H., Guo, X., Gooneratne, R., Lai, R., Zeng, C., Zhan, F., Wang, W., 2016. The gut
microbiome and degradation enzyme activity of wild freshwater fishes influenced by their
trophic levels. Sci. Rep. 6, 1–12. https://doi.org/10.1038/srep24340
Liu, W., Yang, Y., Zhang, J., Gatlin, D.M., Ringo, E., Zhou, Z., 2014. Effects of dietary
microencapsulated sodium butyrate on growth, intestinal mucosal morphology, immune
response and adhesive bacteria in juvenile common carp (Cyprinus carpio) pre-fed with
or without oxidised oil. Br. J. Nutr. 112, 15–29.
https://doi.org/10.1017/S0007114514000610
Llewellyn, M.S., Boutin, S., Hoseinifar, S.H., Derome, N., 2014. Teleost microbiomes: The
state of the art in their characterization, manipulation and importance in aquaculture and
fisheries. Front. Microbiol. https://doi.org/10.3389/fmicb.2014.00207
Llewellyn, M.S., Leadbeater, S., Garcia, C., Sylvain, F.E., Custodio, M., Ang, K.P., Powell, F.,
Carvalho, G.R., Creer, S., Elliot, J., Derome, N., 2017. Parasitism perturbs the mucosal
microbiome of Atlantic Salmon. Sci. Rep. 7, 1–10. https://doi.org/10.1038/srep43465
Llewellyn, M.S., McGinnity, P., Dionne, M., Letourneau, J., Thonier, F., Carvalho, G.R., Creer,
S., Derome, N., 2016. The biogeography of the atlantic salmon (Salmo salar) gut
microbiome. ISME J. 10, 1280–1284. https://doi.org/10.1038/ismej.2015.189
Lloyd-Price, J., Abu-Ali, G., Huttenhower, C., 2016. The healthy human microbiome. Genome
Med. https://doi.org/10.1186/s13073-016-0307-y
Lloyd-Price, J., Arze, C., Ananthakrishnan, A.N., Schirmer, M., Avila-Pacheco, J., Poon, T.W.,
Andrews, E., Ajami, N.J., Bonham, K.S., Brislawn, C.J., Casero, D., Courtney, H.,
Gonzalez, A., Graeber, T.G., Hall, A.B., Lake, K., Landers, C.J., Mallick, H., Plichta, D.R.,
Prasad, M., Rahnavard, G., Sauk, J., Shungin, D., Vázquez-Baeza, Y., White, R.A.,
Bishai, J., Bullock, K., Deik, A., Dennis, C., Kaplan, J.L., Khalili, H., McIver, L.J., Moran,
C.J., Nguyen, L., Pierce, K.A., Schwager, R., Sirota-Madi, A., Stevens, B.W., Tan, W.,
ten Hoeve, J.J., Weingart, G., Wilson, R.G., Yajnik, V., Braun, J., Denson, L.A., Jansson,
J.K., Knight, R., Kugathasan, S., McGovern, D.P.B., Petrosino, J.F., Stappenbeck, T.S.,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
49
Winter, H.S., Clish, C.B., Franzosa, E.A., Vlamakis, H., Xavier, R.J., Huttenhower, C.,
2019. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature
569, 655–662. https://doi.org/10.1038/s41586-019-1237-9
Lokesh, J., Fernandes, J.M.O., Korsnes, K., Bergh, Ø., Brinchmann, M.F., Kiron, V., 2012.
Transcriptional regulation of cytokines in the intestine of Atlantic cod fed yeast derived
mannan oligosaccharide or β-Glucan and challenged with Vibrio anguillarum. Fish
Shellfish Immunol. 33, 626–631. https://doi.org/10.1016/j.fsi.2012.06.017
Lokesh, J., Kiron, V., 2016. Transition from freshwater to seawater reshapes the skin-
associated microbiota of Atlantic salmon. Sci. Rep. 6, 1–10.
https://doi.org/10.1038/srep19707
Lokesh, J., Kiron, V., Sipkema, D., Fernandes, J.M.O., Moum, T., 2019. Succession of
embryonic and the intestinal bacterial communities of Atlantic salmon (Salmo salar)
reveals stage-specific microbial signatures. Microbiologyopen 8, e00672.
https://doi.org/10.1002/mbo3.672
López Nadal, A., Peggs, D., Wiegertjes, G.F., Brugman, S., 2018. Exposure to Antibiotics
Affects Saponin Immersion-Induced Immune Stimulation and Shift in Microbial
Composition in Zebrafish Larvae. Front. Microbiol. 9, 2588.
https://doi.org/10.3389/fmicb.2018.02588
Lowrey, L., Woodhams, D.C., Tacchi, L., Salinas, I., 2015. Topographical mapping of the
rainbow trout (Oncorhynchus mykiss) microbiome reveals a diverse bacterial community
with antifungal properties in the skin. Appl. Environ. Microbiol. 81, 6915–6925.
https://doi.org/10.1128/AEM.01826-15
Lozupone, C., Knight, R., 2005. UniFrac: A new phylogenetic method for comparing microbial
communities. Appl. Environ. Microbiol. 71, 8228–8235.
https://doi.org/10.1128/AEM.71.12.8228-8235.2005
Lozupone, C.A., Knight, R., 2008. Species divergence and the measurement of microbial
diversity. FEMS Microbiol. Rev. 32, 557–578. https://doi.org/10.1111/j.1574-
6976.2008.00111.x
Lu, L., Luo, T., Zhao, Y., Cai, C., Fu, Z., Jin, Y., 2019. Interaction between microplastics and
microorganism as well as gut microbiota: A consideration on environmental animal and
human health. Sci. Total Environ. https://doi.org/10.1016/j.scitotenv.2019.02.380
Ma, C., Chen, C., Jia, L., He, X., Zhang, B., 2019. Comparison of the intestinal microbiota
composition and function in healthy and diseased Yunlong Grouper. AMB Express 9,
187. https://doi.org/10.1186/s13568-019-0913-3
Mahious, A.S., Van Loo, J., Ollevier, F., 2006. Impact of the prebiotics, inulin and oligofructose
on microbial fermentation in the spiral valve of Siberian sturgeon (Acipenser baerii),
World Aquaculture Society. Florence, Italy.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
50
Mallick, H., Ma, S., Franzosa, E.A., Vatanen, T., Morgan, X.C., Huttenhower, C., 2017.
Experimental design and quantitative analysis of microbial community multiomics.
Genome Biol. https://doi.org/10.1186/s13059-017-1359-z
Martin, F.J., Dumas, M., Wang, Y., Legido-Quigley, C., Yap, I.K.S., Tang, H., Zirah, S.,
Murphy, G.M., Cloarec, O., Lindon, J.C., Sprenger, N., Fay, L.B., Kochhar, S., Bladeren,
P. van, Holmes, E., Nicholson, eremy K., 2007a. The Human Microbiome Project.
Nature. https://doi.org/10.1038/nature06244
Martin, F.J., Dumas, M., Wang, Y., Legido-Quigley, C., Yap, I.K.S., Tang, H., Zirah, S.,
Murphy, G.M., Cloarec, O., Lindon, J.C., Sprenger, N., Fay, L.B., Kochhar, S., van
Bladeren, P., Holmes, E., Nicholson, J.K., 2007b. A top-down systems biology view of
microbiome-mammalian metabolic interactions in a mouse model. Mol. Syst. Biol. 3, 112.
https://doi.org/10.1038/msb4100153
Martins, P., Coelho, F.J.R.C., Cleary, D.F.R., Pires, A.C.C., Marques, B., Rodrigues, A.M.,
Quintino, V., Gomes, N.C.M., 2018. Seasonal patterns of bacterioplankton composition
in a semi-intensive European seabass (Dicentrarchus labrax) aquaculture system.
Aquaculture 490, 240–250. https://doi.org/10.1016/j.aquaculture.2018.02.038
Matanza, X.M., Osorio, C.R., 2018. Transcriptome changes in response to temperature in the
fish pathogen Photobacterium damselae subsp. damselae: Clues to understand the
emergence of disease outbreaks at increased seawater temperatures. PLoS One 13,
e0210118. https://doi.org/10.1371/journal.pone.0210118
Maynard, C.L., Elson, C.O., Hatton, R.D., Weaver, C.T., 2012. Reciprocal interactions of the
intestinal microbiota and immune system. Nature. https://doi.org/10.1038/nature11551
McDonald, R., Schreier, H.J., Watts, J.E.M., 2012. Phylogenetic Analysis of Microbial
Communities in Different Regions of the Gastrointestinal Tract in Panaque nigrolineatus,
a Wood-Eating Fish. PLoS One 7, e48018. https://doi.org/10.1371/journal.pone.0048018
McFall-Ngai, M.J., Henderson, B., Ruby, E.G. (Eds.), 2005. The Influence of Cooperative
Bacteria on Animal Host Biology - Google Books. Cambridge University Press.
McMurdie, P.J., Holmes, S., 2014. Waste Not, Want Not: Why Rarefying Microbiome Data Is
Inadmissible. PLoS Comput. Biol. 10, e1003531.
https://doi.org/10.1371/journal.pcbi.1003531
Medzhitov, R., Janeway, C.A., 1998. Innate immune recognition and control of adaptive
immune responses. Semin. Immunol. 10, 351–353.
https://doi.org/10.1006/smim.1998.0136
Mehanna, S.F., 2007. A Preliminary Assessment and Management of Gilthead Bream Sparus
aurata in the Port Said Fishery, the Southeastern Mediterranean, Egypt, Turkish Journal
of Fisheries and Aquatic Sciences.
Meron, D., Davidovich, N., Ofek-Lalzar, M., Berzak, R., Scheinin, A., Regev, Y., Diga, R.,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
51
Tchernov, D., Morick, D., 2020. Specific pathogens and microbial abundance within liver
and kidney tissues of wild marine fish from the Eastern Mediterranean Sea. Microb.
Biotechnol. 13, 770–780. https://doi.org/10.1111/1751-7915.13537
Merrifield, D.L., Dimitroglou, A., Foey, A., Davies, S.J., Baker, R.T.M., Bøgwald, J., Castex,
M., Ringø, E., 2010. The current status and future focus of probiotic and prebiotic
applications for salmonids. Aquaculture.
https://doi.org/10.1016/j.aquaculture.2010.02.007
Merrifield, D.L., Ringø, E., 2014. Aquaculture Nutrition: Gut Health, Probiotics and Prebiotics.
John Wiley & Sons.
Merrifield, D.L., Rodiles, A., 2015. The fish microbiome and its interactions with mucosal
tissues, in: Mucosal Health in Aquaculture. Elsevier Inc., pp. 273–295.
https://doi.org/10.1016/B978-0-12-417186-2.00010-8
Michl, S.C., Beyer, M., Ratten, J.M., Hasler, M., LaRoche, J., Schulz, C., 2019. A diet-change
modulates the previously established bacterial gut community in juvenile brown trout
(Salmo trutta). Sci. Rep. 9, 1–12. https://doi.org/10.1038/s41598-019-38800-7
Michl, S.C., Ratten, J.-M., Beyer, M., Hasler, M., LaRoche, J., Schulz, C., 2017. The malleable
gut microbiome of juvenile rainbow trout (Oncorhynchus mykiss): Diet-dependent shifts
of bacterial community structures. PLoS One 12, e0177735.
https://doi.org/10.1371/journal.pone.0177735
Miller, R.A., Harbottle, H., 2018. Antimicrobial Drug Resistance in Fish Pathogens, in:
Antimicrobial Resistance in Bacteria from Livestock and Companion Animals. American
Society of Microbiology, pp. 501–520. https://doi.org/10.1128/microbiolspec.arba-0017-
2017
Mimee, M., Citorik, R.J., Lu, T.K., 2016. Microbiome therapeutics — Advances and
challenges. Adv. Drug Deliv. Rev. https://doi.org/10.1016/j.addr.2016.04.032
Minich, J., Petrus, S., Michael, J.D., Michael, T.P., Knight, R., Allen, E.E., 2020a. Temporal,
Environmental, and Biological Drivers of the Mucosal Microbiome in a Wild Marine Fish,
Scomber japonicus. Msphere. 5. https://doi.org/10.1128/mSphere.00401-20
Minich, J., Poore, G.D., Jantawongsri, K., Johnston, C., Bowie, K., Bowman, J., Knight, R.,
Nowak, B., Allen, E.E., 2020b. Microbial ecology of atlantic salmon (Salmo salar)
hatcheries: Impacts of the built environment on fish mucosal microbiota. Appl. Environ.
Microbiol. 86. https://doi.org/10.1128/AEM.00411-20
Minniti, G., Hagen, L.H., Porcellato, D., Jørgensen, S.M., Pope, P.B., Vaaje-Kolstad, G., 2017.
The Skin-Mucus Microbial Community of Farmed Atlantic Salmon (Salmo salar). Front.
Microbiol. 8, 2043. https://doi.org/10.3389/fmicb.2017.02043
Miyake, S., Ngugi, D.K., Stingl, U., 2015. Diet strongly influences the gut microbiota of
surgeonfishes. Mol. Ecol. 24, 656–672. https://doi.org/10.1111/mec.13050

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
52
Miyake, S., Soh, M., Nursyafiq Azman, M., Yan Ngoh, S., Orbán, L., Seedorf, H., 2020.
Insights into the microbiome of farmed Asian sea bass (Lates calcarifer) with symptoms
of tenacibaculosis and description of Tenacibaculum singaporense sp. nov. Antonie Van
Leeuwenhoek 113, 737–752. https://doi.org/10.1007/s10482-020-01391-9
Mogensen, T.H., 2009. Pathogen recognition and inflammatory signaling in innate immune
defenses. Clin. Microbiol. Rev. https://doi.org/10.1128/CMR.00046-08
Mohamad, N., Amal, M.N.A., Yasin, I.S.M., Zamri Saad, M., Nasruddin, N.S., Al-saari, N.,
Mino, S., Sawabe, T., 2019. Vibriosis in cultured marine fishes: a review. Aquaculture
512, 734289. https://doi.org/10.1016/j.aquaculture.2019.734289
Mohammed, H.H., Arias, C.R., 2015. Potassium permanganate elicits a shift of the external
fish microbiome and increases host susceptibility to columnaris disease. Vet. Res. 46,
82. https://doi.org/10.1186/s13567-015-0215-y
Mouquet, N., Hoopes, M.F., Amarasekare, P., 2005. The world is patchy and heterogeneous.
Trade-off and source-sink dynamics in competitive metacommunities, in: Holyoak, M.,
Leibold, M.A., Holt, R.D. (Eds.), Metacommunities: Spatial Dynamics and Ecological
Communities. The University of Chicago Press, Chicago, pp. 237–262.
Namiki, T., Hachiya, T., Tanaka, H., Sakakibara, Y., 2012. MetaVelvet: An extension of Velvet
assembler to de novo metagenome assembly from short sequence reads. Nucleic Acids
Res. 40, e155–e155. https://doi.org/10.1093/nar/gks678
Narrowe, A.B., Albuthi-Lantz, M., Smith, E.P., Bower, K.J., Roane, T.M., Vajda, A.M., Miller,
C.S., 2015. Perturbation and restoration of the fathead minnow gut microbiome after low-
level triclosan exposure. Microbiome 3, 6. https://doi.org/10.1186/s40168-015-0069-6
Navarrete, P., Mardones, P., Opazo, R., Espejo, R., Romero, J., 2008. Oxytetracycline
treatment reduces bacterial diversity of intestinal microbiota of Atlantic salmon. J. Aquat.
Anim. Health 20, 177–183. https://doi.org/10.1577/H07-043.1
Nie, L., Zhou, Q.J., Qiao, Y., Chen, J., 2017. Interplay between the gut microbiota and immune
responses of ayu (Plecoglossus altivelis) during Vibrio anguillarum infection. Fish
Shellfish Immunol. 68, 479–487. https://doi.org/10.1016/j.fsi.2017.07.054
Nikouli, E., Meziti, A., Antonopoulou, E., Mente, E., Kormas, K., 2018. Gut Bacterial
Communities in Geographically Distant Populations of Farmed Sea Bream (Sparus
aurata) and Sea Bass (Dicentrarchus labrax). Microorganisms 6, 92.
https://doi.org/10.3390/microorganisms6030092
Nikouli, Meziti, Antonopoulou, Mente, Kormas, 2019. Host-Associated Bacterial Succession
during the Early Embryonic Stages and First Feeding in Farmed Gilthead Sea Bream
(Sparus aurata). Genes (Basel). 10, 483. https://doi.org/10.3390/genes10070483
Nishida, K., Ono, K., Kanaya, S., Takahashi, K., 2014. KEGGscape: A Cytoscape app for
pathway data integration. F1000Research 3.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
53
https://doi.org/10.12688/f1000research.4524.1
Nurk, S., Meleshko, D., Korobeynikov, A., Pevzner, P.A., 2017. MetaSPAdes: A new versatile
metagenomic assembler. Genome Res. 27, 824–834.
https://doi.org/10.1101/gr.213959.116
Oksanen, J., Kindt, R., Legendre, P., O’Hara, B., Simpson, G.L., Stevens, M.H.H., Wagner,
H., 2008. The vegan package: community ecology package, version 1.13-1 [WWW
Document].
Ornelas-García, P., Pajares, S., Sosa-Jiménez, V.M., Rétaux, S., Miranda-Gamboa, R.A.,
2018. Microbiome differences between river-dwelling and cave-adapted populations of
the fish Astyanax mexicanus (De Filippi, 1853). PeerJ 2018, e5906.
https://doi.org/10.7717/peerj.5906
Osadchiy, V., Martin, C.R., Mayer, E.A., 2019. The Gut–Brain Axis and the Microbiome:
Mechanisms and Clinical Implications. Clin. Gastroenterol. Hepatol.
https://doi.org/10.1016/j.cgh.2018.10.002
Park, J.C., Im, S.H., 2020. Of men in mice: the development and application of a humanized
gnotobiotic mouse model for microbiome therapeutics. Exp. Mol. Med.
https://doi.org/10.1038/s12276-020-0473-2
Parris, D.J., Brooker, R.M., Morgan, M.A., Dixson, D.L., Stewart, F.J., 2016. Whole gut
microbiome composition of damselfish and cardinalfish before and after reef settlement.
PeerJ 2016, e2412. https://doi.org/10.7717/PEERJ.2412
Parshukov, A.N., Kashinskaya, E.N., Simonov, E.P., Hlunov, O.V., Izvekova, G.I., Andree,
K.B., Solovyev, M.M., 2019. Variations of the intestinal gut microbiota of farmed rainbow
trout, Oncorhynchus mykiss (Walbaum), depending on the infection status of the fish. J.
Appl. Microbiol. 127, 379–395. https://doi.org/10.1111/jam.14302
Paulino, L.C., Tseng, C.H., Strober, B.E., Blaser, M.J., 2006. Molecular analysis of fungal
microbiota in samples from healthy human skin and psoriatic lesions. J. Clin. Microbiol.
44, 2933–2941. https://doi.org/10.1128/JCM.00785-06
Peirson, S.N., Butler, J.N., 2007. RNA Extraction From Mammalian Tissues. Humana Press,
pp. 315–327. https://doi.org/10.1007/978-1-59745-257-1_22
Peng, J., Xiao, X., Hu, M., Zhang, X., 2018. Interaction between gut microbiome and
cardiovascular disease. Life Sci. https://doi.org/10.1016/j.lfs.2018.10.063
Peng, Y., Leung, H.C.M., Yiu, S.M., Chin, F.Y.L., 2012. IDBA-UD: a de novo assembler for
single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics
28, 1420–1428. https://doi.org/10.1093/bioinformatics/bts174
Pérez-Pascual, D., Estellé, J., Dutto, G., Rodde, C., Bernardet, J.F., Marchand, Y., Duchaud,
E., Przybyla, C., Ghigo, J.M., 2020. Growth performance and adaptability of european
sea bass (Dicentrarchus labrax) gut microbiota to alternative diets free of fish products.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
54
Microorganisms 8, 1–18. https://doi.org/10.3390/microorganisms8091346
Perry, W.B., Lindsay, E., Payne, C.J., Brodie, C., Kazlauskaite, R., 2020. The role of the gut
microbiome in sustainable teleost aquaculture. Proc. R. Soc. B Biol. Sci.
https://doi.org/10.1098/rspb.2020.0184
Petersen, C., Round, J.L., 2014. Defining dysbiosis and its influence on host immunity and
disease. Cell. Microbiol. 16, 1024–1033. https://doi.org/10.1111/cmi.12308
Piccinno, F., Gottschalk, F., Seeger, S., Nowack, B., 2012. Industrial production quantities and
uses of ten engineered nanomaterials in Europe and the world. J. Nanoparticle Res.
https://doi.org/10.1007/s11051-012-1109-9
Pieters, N., Brunt, J., Austin, B., Lyndon, A.R., 2008. Efficacy of in-feed probiotics against
Aeromonas bestiarum and Ichthyophthirius multifiliis skin infections in rainbow trout
(Oncorhynchus mykiss, Walbaum). J. Appl. Microbiol. 105, 723–732.
https://doi.org/10.1111/j.1365-2672.2008.03817.x
Pimentel, T., Marcelino, J., Ricardo, F., Soares, A.M.V.M., Calado, R., 2017. Bacterial
communities 16S rDNA fingerprinting as a potential tracing tool for cultured seabass
Dicentrarchus labrax. Sci. Rep. 7, 1–10. https://doi.org/10.1038/s41598-017-11552-y
Pindling, S., Azulai, D., Zheng, B., Dahan, D., Perron, G.G., 2018. Dysbiosis and early
mortality in zebrafish larvae exposed to subclinical concentrations of streptomycin. FEMS
Microbiol. Lett. 365, 188. https://doi.org/10.1093/femsle/fny188
Planas, M., Pérez-Lorenzo, M., Hjelm, M., Gram, L., Uglenes Fiksdal, I., Bergh, Ø., Pintado,
J., 2006. Probiotic effect in vivo of Roseobacter strain 27-4 against Vibrio (Listonella)
anguillarum infections in turbot (Scophthalmus maximus L.) larvae. Aquaculture 255,
323–333. https://doi.org/10.1016/j.aquaculture.2005.11.039
Pratte, Z.A., Besson, M., Hollman, R.D., Stewart, F.J., 2018. The Gills of Reef Fish Support a
Distinct Microbiome Influenced by Host-Specific Factors. https://doi.org/10.1128/AEM
Pridgeon, J.W., Klesius, P.H., 2012. Major bacterial diseases in aquaculture and their vaccine
development. Anim. Sci. Rev. 7, 1–16.
Proctor, L.M., Creasy, H.H., Fettweis, J.M., Lloyd-Price, J., Mahurkar, A., Zhou, W., Buck,
G.A., Snyder, M.P., Strauss, J.F., Weinstock, G.M., White, O., Huttenhower, C., 2019.
The Integrative Human Microbiome Project. Nature 569, 641–648.
https://doi.org/10.1038/s41586-019-1238-8
Prosser, J.I., 2010. Replicate or lie. Environ. Microbiol. 12, 1806–1810.
https://doi.org/10.1111/j.1462-2920.2010.02201.x
Pruesse, E., Quast, C., Knittel, K., Fuchs, B.M., Ludwig, W., Peplies, J., Glöckner, F.O., 2007.
SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA
sequence data compatible with ARB. Nucleic Acids Res. 35, 7188–7196.
https://doi.org/10.1093/nar/gkm864

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
55
Pull, S.L., Doherty, J.M., Mills, J.C., Gordon, J.I., Stappenbeck, T.S., 2005. Activated
macrophages are an adaptive element of the colonic epithelial progenitor niche
necessary for regenerative responses to injury. Proc. Natl. Acad. Sci. U. S. A. 102, 99–
104. https://doi.org/10.1073/pnas.0405979102
Qi, X.Z., Xue, M.Y., Yang, S.B., Zha, J.W., Wang, G.X., Ling, F., 2017. Ammonia exposure
alters the expression of immune-related and antioxidant enzymes-related genes and the
gut microbial community of crucian carp (Carassius auratus). Fish Shellfish Immunol. 70,
485–492. https://doi.org/10.1016/j.fsi.2017.09.043
Ramírez, C., Romero, J., 2017a. Fine Flounder (Paralichthys adspersus) Microbiome Showed
Important Differences between Wild and Reared Specimens. Front. Microbiol. 08, 271.
https://doi.org/10.3389/fmicb.2017.00271
Ramírez, C., Romero, J., 2017b. The Microbiome of Seriola lalandi of Wild and Aquaculture
Origin Reveals Differences in Composition and Potential Function. Front. Microbiol. 8,
1844. https://doi.org/10.3389/fmicb.2017.01844
Rawls, J.F., Mahowald, M.A., Ley, R.E., Gordon, J.I., 2006. Reciprocal Gut Microbiota
Transplants from Zebrafish and Mice to Germ-free Recipients Reveal Host Habitat
Selection. Cell 127, 423–433. https://doi.org/10.1016/j.cell.2006.08.043
Rawls, J.F., Samuel, B.S., Gordon, J.I., 2004. Gnotobiotic zebrafish reveal evolutionarily
conserved responses to the gut microbiota. Proc. Natl. Acad. Sci. U. S. A. 101, 4596–
4601. https://doi.org/10.1073/pnas.0400706101
Razeghi Mansour, M., Akrami, R., Ghobadi, S.H., Amani Denji, K., Ezatrahimi, N., Gharaei,
A., 2012. Effect of dietary mannan oligosaccharide (MOS) on growth performance,
survival, body composition, and some hematological parameters in giant sturgeon
juvenile (Huso huso Linnaeus, 1754). Fish Physiol. Biochem. 38, 829–835.
https://doi.org/10.1007/s10695-011-9570-4
Reck, M., Tomasch, J., Deng, Z., Jarek, M., Husemann, P., Wagner-Döbler, I., 2015. Stool
metatranscriptomics: A technical guideline for mRNA stabilisation and isolation. BMC
Genomics 16, 1–18. https://doi.org/10.1186/s12864-015-1694-y
Rehman, S., Gora, A.H., Ahmad, I., Rasool, I., 2017. Stress in aquaculture hatcheries: source,
impact and mitigation. Int. J. Curr. Microbiol. Appl. Sci. 6, 3030–3045.
https://doi.org/10.20546/ijcmas.2017.610.357
Reid, K.M., Patel, S., Robinson, A.J., Bu, L., Jarungsriapisit, J., Moore, L.J., Salinas, I., 2017.
Salmonid alphavirus infection causes skin dysbiosis in Atlantic salmon (Salmo salar L.)
post-smolts. PLoS One 12, e0172856. https://doi.org/10.1371/journal.pone.0172856
Reinhart, E.M., Korry, B.J., Rowan-Nash, A.D., Belenky, P., 2019. Defining the Distinct Skin
and Gut Microbiomes of the Northern Pike (Esox lucius). Front. Microbiol. 10, 2118.
https://doi.org/10.3389/fmicb.2019.02118

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
56
Remus, D.M., van Kranenburg, R., van Swam, I.I., Taverne, N., Bongers, R.S., Wels, M.,
Wells, J.M., Bron, P.A., Kleerebezem, M., 2012. Impact of 4 Lactobacillus plantarum
capsular polysaccharide clusters on surface glycan composition and host cell signaling.
Microb. Cell Fact. 11, 149. https://doi.org/10.1186/1475-2859-11-149
Reverter, M., Sasal, P., Tapissier-Bontemps, N., Lecchini, D., Suzuki, M., 2017.
Characterisation of the gill mucosal bacterial communities of four butterflyfish species: a
reservoir of bacterial diversity in coral reef ecosystems. FEMS Microbiol. Ecol. 93, 51.
https://doi.org/10.1093/femsec/fix051
Reverter, M., Tapissier-Bontemps, N., Sarter, S., Sasal, P., Caruso, D., 2021. Moving towards
more sustainable aquaculture practices: a meta-analysis on the potential of plant-
enriched diets to improve fish growth, immunity and disease resistance. Rev. Aquac. 13,
537–555. https://doi.org/10.1111/raq.12485
Rimmer, D.W., Wiebe, W.J., 1987. Fermentative microbial digestion in herbivorous fishes. J.
Fish Biol. 31, 229–236. https://doi.org/10.1111/j.1095-8649.1987.tb05228.x
Rimoldi, S., Gliozheni, E., Ascione, C., Gini, E., Terova, G., 2018. Effect of a specific
composition of short- and medium-chain fatty acid 1-Monoglycerides on growth
performances and gut microbiota of gilthead sea bream (Sparus aurata). PeerJ 2018,
e5355. https://doi.org/10.7717/peerj.5355
Ringø, E., Dimitroglou, A., Hoseinifar, S.H., Davies, S.J., 2014. Prebiotics in Finfish: An
Update, in: Aquaculture Nutrition. John Wiley & Sons, Ltd, Chichester, UK, pp. 360–400.
https://doi.org/10.1002/9781118897263.ch14
Ringø, E., Holzapfel, W., 2000. Identification and characterization of carnobacteria associated
with the gills of Atlantic salmon (Salmo salar L.). Syst. Appl. Microbiol. 23, 523–527.
https://doi.org/10.1016/S0723-2020(00)80026-0
Ringø, E., Zhou, Z., Olsen, R.E., Song, S.K., 2012. Use of chitin and krill in aquaculture - the
effect on gut microbiota and the immune system: a review. Aquac. Nutr. 18, 117–131.
https://doi.org/10.1111/j.1365-2095.2011.00919.x
Ringø, E., Zhou, Z., Vecino, J.L.G., Wadsworth, S., Romero, J., Krogdahl, Olsen, R.E.,
Dimitroglou, A., Foey, A., Davies, S., Owen, M., Lauzon, H.L., Martinsen, L.L., De
Schryver, P., Bossier, P., Sperstad, S., Merrifield, D.L., 2016. Effect of dietary
components on the gut microbiota of aquatic animals. A never-ending story? Aquac. Nutr.
22, 219–282. https://doi.org/10.1111/anu.12346
Roberfroid, M., 2007. Prebiotics: The concept revisited, in: Journal of Nutrition. American
Institute of Nutrition, pp. 830S-837S. https://doi.org/10.1093/jn/137.3.830s
Robinson, C.J., Bohannan, B.J.M., Young, V.B., 2010. From Structure to Function: the
Ecology of Host-Associated Microbial Communities. Microbiol. Mol. Biol. Rev. 74, 453–
476. https://doi.org/10.1128/mmbr.00014-10

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
57
Robinson, J.M., Pasternak, Z., Mason, C.E., Elhaik, E., 2021. Forensic Applications of
Microbiomics: A Review. Front. Microbiol. https://doi.org/10.3389/fmicb.2020.608101
Rodriguez-Estrada, U., Satoh, S., Haga, Y., Fushimi, H., Sweetman, J., 2013. Effects of
inactivated Enterococcus faecalis and mannan oligosaccharide and their combination on
growth, immunity, and disease protection in rainbow trout. N. Am. J. Aquac. 75, 416–428.
https://doi.org/10.1080/15222055.2013.799620
Rosselló-Mora, R., 2005. Updating prokaryotic taxonomy. J. Bacteriol.
https://doi.org/10.1128/JB.187.18.6255-6257.2005
Rudi, K., Angell, I.L., Pope, P.B., Olav Vik, J., Sandve, S.R., Snipen, L.-G., 2017. A stable
core gut microbiota across fresh-to saltwater transition for farmed Atlantic salmon. Am.
Soc. Microbiol. https://doi.org/10.1128/AEM.01974-17
Ruiz-Rodríguez, M., Scheifler, & M., Sanchez-Brosseau, & S., Magnanou, & E., West, & N.,
Suzuki, & M., Duperron, & S., Desdevises, Y., 2020. Host Species and Body Site Explain
the Variation in the Microbiota Associated to Wild Sympatric Mediterranean Teleost
Fishes. Microb. Ecol. 80, 212–222. https://doi.org/10.1007/s00248-020-01484-y
Sáenz, J.S., Marques, T.V., Barone, R.S.C., Cyrino, J.E.P., Kublik, S., Nesme, J., Schloter,
M., Rath, S., Vestergaard, G., 2019. Oral administration of antibiotics increased the
potential mobility of bacterial resistance genes in the gut of the fish Piaractus
mesopotamicus. Microbiome 7, 24. https://doi.org/10.1186/s40168-019-0632-7
Saksida, S.M., Gardner, I., Lent, M.L., 2014. Transmission of infected agents between wild
and farmed fish, in: Woo, P.T.K., Bruno, D.W. (Eds.), Saksida SM, Gardner I, Kent ML.
2014. Transmission of Infectious Agents between Wild and Farmed Fish. In Diseases
and Disorders of Finfish in Cage Culture. CABI, Wallingford, UK, p. 313.
Salinas, I., 2015. The mucosal immune system of teleost fish. Biology (Basel).
https://doi.org/10.3390/biology4030525
Salinas, I., Magadán, S., 2017. Omics in fish mucosal immunity. Dev. Comp. Immunol. 75,
99–108. https://doi.org/10.1016/j.dci.2017.02.010
Sampson, T.R., Debelius, J.W., Thron, T., Janssen, S., Shastri, G.G., Ilhan, Z.E., Challis, C.,
Schretter, C.E., Rocha, S., Gradinaru, V., Chesselet, M.F., Keshavarzian, A., Shannon,
K.M., Krajmalnik-Brown, R., Wittung-Stafshede, P., Knight, R., Mazmanian, S.K., 2016.
Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s
Disease. Cell 167, 1469-1480.e12. https://doi.org/10.1016/j.cell.2016.11.018
Schade, F.M., Raupach, M.J., Mathias Wegner, K., 2016. Seasonal variation in parasite
infection patterns of marine fish species from the Northern Wadden Sea in relation to
interannual temperature fluctuations. J. Sea Res. 113, 73–84.
https://doi.org/10.1016/j.seares.2015.09.002
Schar, D., Klein, E.Y., Laxminarayan, R., Gilbert, M., Van Boeckel, T.P., 2020. Global trends

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
58
in antimicrobial use in aquaculture. Sci. Rep. 10, 21878. https://doi.org/10.1038/s41598-
020-78849-3
Schloss, P.D., Westcott, S.L., Ryabin, T., Hall, J.R., Hartmann, M., Hollister, E.B., Lesniewski,
R.A., Oakley, B.B., Parks, D.H., Robinson, C.J., Sahl, J.W., Stres, B., Thallinger, G.G.,
Van Horn, D.J., Weber, C.F., 2009. Introducing mothur: Open-source, platform-
independent, community-supported software for describing and comparing microbial
communities. Appl. Environ. Microbiol. 75, 7537–7541.
https://doi.org/10.1128/AEM.01541-09
Schmidt, J.G., Thompson, Kim D, Padrós, Francesc, Thompson, K D, Padrós, F, 2018.
Emerging skin diseases in aquaculture. Bull. Eur. Assoc. Fish Pathol. 38, 122–129.
Schmidt, V., Amaral-Zettler, L., Davidson, J., Summerfelt, S., Good, C., 2016. Influence of
Fishmeal-Free Diets on Microbial Communities in Atlantic Salmon (Salmo salar)
Recirculation Aquaculture Systems. https://doi.org/10.1128/AEM.00902-16
Schmidt, V.T., Smith, K.F., Melvin, D.W., Amaral-Zettler, L.A., 2015. Community assembly of
a euryhaline fish microbiome during salinity acclimation. Mol. Ecol. 24, 2537–2550.
https://doi.org/10.1111/mec.13177
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W.S., Huttenhower, C.,
2011. Metagenomic biomarker discovery and explanation. Genome Biol. 12, R60.
https://doi.org/10.1186/gb-2011-12-6-r60
Semova, I., Carten, J.D., Stombaugh, J., MacKey, L.C., Knight, R., Farber, S.A., Rawls, J.F.,
2012. Microbiota regulate intestinal absorption and metabolism of fatty acids in the
zebrafish. Cell Host Microbe 12, 277–288. https://doi.org/10.1016/j.chom.2012.08.003
Shakya, M., Lo, C.C., Chain, P.S.G., 2019. Advances and challenges in metatranscriptomic
analysis. Front. Genet. https://doi.org/10.3389/fgene.2019.00904
She, R., Li, T.T., Luo, D., Li, J.B., Yin, L.Y., Li, H., Liu, Y.M., Li, X.Z., Yan, Q. gui, 2017.
Changes in the Intestinal Microbiota of Gibel Carp (Carassius gibelio) Associated with
Cyprinid herpesvirus 2 (CyHV-2) Infection. Curr. Microbiol. 74, 1130–1136.
https://doi.org/10.1007/s00284-017-1294-y
Shefat, S.H.T., 2018. Probiotic Strains Used in Aquaculture. Int. Res. J. Microbiol. 7, 43–55.
https://doi.org/10.14303/irjm.2018.023
Sinha, R., Abnet, C.C., White, O., Knight, R., Huttenhower, C., 2015. The microbiome quality
control project: Baseline study design and future directions. Genome Biol. 16, 1–6.
https://doi.org/10.1186/s13059-015-0841-8
Smith, B., 2011. Optimising Bacterial DNA Extraction from Faecal Samples: Comparison of
Three Methods. Open Microbiol. J. 5, 14–17.
https://doi.org/10.2174/1874285801105010014
Smith, C.C.R., Snowberg, L.K., Gregory Caporaso, J., Knight, R., Bolnick, D.I., 2015. Dietary

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
59
input of microbes and host genetic variation shape among-population differences in
stickleback gut microbiota. ISME J. 9, 2515–2526. https://doi.org/10.1038/ismej.2015.64
Smith, D.C., Douglas, A.E., 1987. The biology of symbiosis. Biol. symbiosis.
Sneddon, L.U., Wolfenden, D.C.C., Thomson, J.S., 2016. Stress Management and Welfare,
in: Fish Physiology. Academic press, pp. 463–539. https://doi.org/10.1016/B978-0-12-
802728-8.00012-6
Soderberg, R.W., 1994. Flowing Water Fish Culture. CRC Press.
Sokolowska, M., Frei, R., Lunjani, N., Akdis, C.A., O’Mahony, L., 2018. Microbiome and
asthma. Asthma Res. Pract. 4, 1. https://doi.org/10.1186/s40733-017-0037-y
Song, S.J., Amir, A., Metcalf, J.L., Amato, K.R., Xu, Z.Z., Humphrey, G., Knight, R., 2016.
Preservation Methods Differ in Fecal Microbiome Stability, Affecting Suitability for Field
Studies. mSystems 1, 21–37. https://doi.org/10.1128/msystems.00021-16
Sonnenschein, E.C., Jimenez, G., Castex, M., Gram, L., 2020. The Roseobacter-Group
Bacterium Phaeobacter as a Safe Probiotic Solution for Aquaculture . Appl. Environ.
Microbiol. 87. https://doi.org/10.1128/aem.02581-20
Staley, J.T., Konopka, A., 1985. Measurement of in Situ Activities of Nonphotosynthetic
Microorganisms in Aquatic and Terrestrial Habitats. Annu. Rev. Microbiol. 39, 321–346.
https://doi.org/10.1146/annurev.mi.39.100185.001541
Staykov, Y., Spring, P., Denev, S., Sweetman, J., 2007. Effect of a mannan oligosaccharide
on the growth performance and immune status of rainbow trout (Oncorhynchus mykiss).
Aquac. Int. 15, 153–161. https://doi.org/10.1007/s10499-007-9096-z
Stephens, W.Z., Burns, A.R., Stagaman, K., Wong, S., Rawls, J.F., Guillemin, K., Bohannan,
B.J.M., 2016. The composition of the zebrafish intestinal microbial community varies
across development. ISME J. 10, 644–654. https://doi.org/10.1038/ismej.2015.140
Steury, Currey, Cresko, Bohannan, 2019. Population Genetic Divergence and Environment
Influence the Gut Microbiome in Oregon Threespine Stickleback. Genes (Basel). 10, 484.
https://doi.org/10.3390/genes10070484
Sthle, L., Wold, S., 1989. Analysis of variance (ANOVA). Chemom. Intell. Lab. Syst.
https://doi.org/10.1016/0169-7439(89)80095-4
Sugita, H., Iwata, J., Miyajima, C., Kubo, T., Noguchi, T., Hashimoto, K., Deguchi, Y., 1989.
Changes in microflora of a puffer fish Fugu niphobles, with different water temperatures.
Mar. Biol. 101, 299–304. https://doi.org/10.1007/BF00428125
Sullam, K.E., Rubin, B.E.R., Dalton, C.M., Kilham, S.S., Flecker, A.S., Russell, J.A., 2015.
Divergence across diet, time and populations rules out parallel evolution in the gut
microbiomes of Trinidadian guppies. ISME J. 9, 1508–1522.
https://doi.org/10.1038/ismej.2014.231
Sun, Y.Z., Xia, H.Q., Yang, H.L., Wang, Y.L., Zou, W.C., 2014. TLR2 signaling may play a key

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
60
role in the probiotic modulation of intestinal microbiota in grouper Epinephelus coioides.
Aquaculture 430, 50–56. https://doi.org/10.1016/j.aquaculture.2014.03.042
Sylvain, F.-É., Holland, A., Bouslama, S., Audet-Gilbert, É., Lavoie, C., Val, A.L., Derome, N.,
2020. Fish Skin and Gut Microbiomes Show Contrasting Signatures of Host Species and
Habitat. Appl. Environ. Microbiol. 86. https://doi.org/10.1128/AEM.00789-20
Sylvain, F.É., Cheaib, B., Llewellyn, M., Gabriel Correia, T., Barros Fagundes, D., Luis Val,
A., Derome, N., 2016. PH drop impacts differentially skin and gut microbiota of the
Amazonian fish tambaqui (Colossoma macropomum). Sci. Rep. 6, 1–10.
https://doi.org/10.1038/srep32032
Tacchi, L., Lowrey, L., Musharrafieh, R., Crossey, K., Larragoite, E.T., Salinas, I., 2015.
Effects of transportation stress and addition of salt to transport water on the skin mucosal
homeostasis of rainbow trout (Oncorhynchus mykiss). Aquaculture 435, 120–127.
https://doi.org/10.1016/j.aquaculture.2014.09.027
Talwar, C., Nagar, S., Lal, R., Negi, R.K., 2018. Fish Gut Microbiome: Current Approaches
and Future Perspectives. Indian J. Microbiol. https://doi.org/10.1007/s12088-018-0760-y
Tao, Z., Zhou, T., Zhou, S., Wang, G., 2016. Temperature-regulated expression of type VI
secretion systems in fish pathogen Pseudomonas plecoglossicida revealed by
comparative secretome analysis. FEMS Microbiol. Lett. 363, fnw261.
https://doi.org/10.1093/femsle/fnw261
Tarnecki, A.M., Brennan, N.P., Schloesser, R.W., Rhody, N.R., 2019. Shifts in the Skin-
Associated Microbiota of Hatchery-Reared Common Snook Centropomus undecimalis
During Acclimation to the Wild. Microb. Ecol. 77, 770–781.
https://doi.org/10.1007/s00248-018-1252-7
Tarnecki, A.M., Burgos, F.A., Ray, C.L., Arias, C.R., 2017. Fish intestinal microbiome: diversity
and symbiosis unravelled by metagenomics. J. Appl. Microbiol. 123, 2–17.
https://doi.org/10.1111/jam.13415
Tarnecki, A.M., Levi, N.J., Resley, M., Main, K., 2021. Effect of copper sulfate on the external
microbiota of adult common snook (Centropomus undecimalis). Anim. Microbiome 3, 21.
https://doi.org/10.1186/s42523-021-00085-5
Tian, L., Tan, P., Yang, L., Zhu, W., Xu, D., 2020. Effects of salinity on the growth, plasma ion
concentrations, osmoregulation, non-specific immunity, and intestinal microbiota of the
yellow drum (Nibea albiflora). Aquaculture 528, 735470.
https://doi.org/10.1016/j.aquaculture.2020.735470
Toranzo, A.E., Magariños, B., Romalde, J.L., 2005. A review of the main bacterial fish
diseases in mariculture systems. Aquaculture 246, 37–61.
https://doi.org/10.1016/j.aquaculture.2005.01.002
Torrecillas, S., Makol, A., Betancor, M.B., Montero, D., Caballero, M.J., Sweetman, J.,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
61
Izquierdo, M., 2013. Enhanced intestinal epithelial barrier health status on European sea
bass (Dicentrarchus labrax) fed mannan oligosaccharides. Fish Shellfish Immunol. 34,
1485–1495. https://doi.org/10.1016/j.fsi.2013.03.351
Torrecillas, S., Montero, D., Izquierdo, M., 2014. Improved health and growth of fish fed
mannan oligosaccharides: Potential mode of action. Fish Shellfish Immunol.
https://doi.org/10.1016/j.fsi.2013.12.029
Tran, N.T., Xiong, F., Hao, Y.T., Zhang, J., Wu, S.G., Wang, G.T., 2018a. Starvation
influences the microbiota assembly and expression of immunity-related genes in the
intestine of grass carp (Ctenopharyngodon idellus). Aquaculture 489, 121–129.
https://doi.org/10.1016/j.aquaculture.2018.02.016
Tran, N.T., Zhang, J., Xiong, F., Wang, G.T., Li, W.X., Wu, S.G., 2018b. Altered gut microbiota
associated with intestinal disease in grass carp (Ctenopharyngodon idellus). World J.
Microbiol. Biotechnol. 34, 71. https://doi.org/10.1007/s11274-018-2447-2
Trivedi, B., 2012. Microbiome: the surface brigade. Nature 492, S60–S61.
Tsiaoussis, J., Antoniou, M.N., Koliarakis, I., Mesnage, R., Vardavas, C.I., Izotov, B.N.,
Psaroulaki, A., Tsatsakis, A., 2019. Effects of single and combined toxic exposures on
the gut microbiome: Current knowledge and future directions. Toxicol. Lett. 312, 72–97.
https://doi.org/10.1016/j.toxlet.2019.04.014
Turnbaugh, P.J., Ley, R.E., Mahowald, M.A., Magrini, V., Mardis, E.R., Gordon, J.I., 2006. An
obesity-associated gut microbiome with increased capacity for energy harvest. Nature
444, 1027–1031. https://doi.org/10.1038/nature05414
Tyagi, A., Singh, B., Billekallu Thammegowda, N.K., Singh, N.K., 2019. Shotgun
metagenomics offers novel insights into taxonomic compositions, metabolic pathways
and antibiotic resistance genes in fish gut microbiome. Arch. Microbiol. 201, 295–303.
https://doi.org/10.1007/s00203-018-1615-y
Ulgen, E., Ozisik, O., Sezerman, O.U., 2019. PathfindR: An R package for comprehensive
identification of enriched pathways in omics data through active subnetworks. Front.
Genet. 10, 858. https://doi.org/10.3389/fgene.2019.00858
Uren Webster, T.M., Consuegra, S., Hitchings, M., Garcia de Leaniz, C., 2018. Interpopulation
Variation in the Atlantic Salmon Microbiome Reflects Environmental and Genetic
Diversity. Appl. Environ. Microbiol. 84. https://doi.org/10.1128/aem.00691-18
Van Boeckel, T.P., Glennon, E.E., Chen, D., Gilbert, M., Robinson, T.P., Grenfell, B.T., Levin,
S.A., Bonhoeffer, S., Laxminarayan, R., 2017. Reducing antimicrobial use in food
animals. Science (80-. ). 357, 1350–1352. https://doi.org/10.1126/science.aao1495
Vasemägi, A., Visse, M., Kisand, V., 2017. Effect of Environmental Factors and an Emerging
Parasitic Disease on Gut Microbiome of Wild Salmonid Fish. mSphere 2, 418–435.
https://doi.org/10.1128/msphere.00418-17

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
62
Villasante, A., Ramírez, C., Catalán, N., Opazo, R., Dantagnan, P., Romero, J., 2019. Effect
of Dietary Carbohydrate-to-Protein Ratio on Gut Microbiota in Atlantic Salmon (Salmo
salar). Animals 9, 89. https://doi.org/10.3390/ani9030089
Vinoj, G., Jayakumar, R., Chen, J.C., Withyachumnarnkul, B., Shanthi, S., Vaseeharan, B.,
2015. N-hexanoyl-L-homoserine lactone-degrading Pseudomonas aeruginosa
PsDAHP1 protects zebrafish against Vibrio parahaemolyticus infection. Fish Shellfish
Immunol. 42, 204–212. https://doi.org/10.1016/j.fsi.2014.10.033
Wagner Mackenzie, B., Waite, D.W., Taylor, M.W., 2015. Evaluating variation in human gut
microbiota profiles due to DNA extraction method and inter-subject differences. Front.
Microbiol. 6, 130. https://doi.org/10.3389/fmicb.2015.00130
Walker, A.W., Martin, J.C., Scott, P., Parkhill, J., Flint, H.J., Scott, K.P., 2015. 16S rRNA gene-
based profiling of the human infant gut microbiota is strongly influenced by sample
processing and PCR primer choice. Microbiome 3, 1–11. https://doi.org/10.1186/s40168-
015-0087-4
Wang, A.R., Ran, C., Ringø, E., Zhou, Z.G., 2018. Progress in fish gastrointestinal microbiota
research. Rev. Aquac. 10, 626–640. https://doi.org/10.1111/raq.12191
Wang, E., Yuan, Z., Wang, K., Gao, D., Liu, Z., Liles, M.R., 2019. Consumption of florfenicol-
medicated feed alters the composition of the channel catfish intestinal microbiota
including enriching the relative abundance of opportunistic pathogens. Aquaculture 501,
111–118. https://doi.org/10.1016/j.aquaculture.2018.11.019
Wang, X., Shen, M., Zhou, J., Jin, Y., 2019. Chlorpyrifos disturbs hepatic metabolism
associated with oxidative stress and gut microbiota dysbiosis in adult zebrafish. Comp.
Biochem. Physiol. Part - C Toxicol. Pharmacol. 216, 19–28.
https://doi.org/10.1016/j.cbpc.2018.11.010
Wang, Y., Zhu, J., Ge, X., Sun, S., Su, Y., Li, B., Hou, Y., Ren, M., 2019. Effects of stocking
density on the growth performance, digestive enzyme activities, antioxidant resistance,
and intestinal microflora of blunt snout bream (Megalobrama amblycephala) juveniles.
Aquac. Res. 50, 236–246. https://doi.org/10.1111/are.13889
Watts, J., McDonald, R., Daniel, R., Schreier, H., 2013. Examination of a Culturable Microbial
Population from the Gastrointestinal Tract of the Wood-Eating Loricariid Catfish Panaque
nigrolineatus. Diversity 5, 641–656. https://doi.org/10.3390/d5030641
Weir, A., Westerhoff, P., Fabricius, L., Hristovski, K., Von Goetz, N., 2012. Titanium dioxide
nanoparticles in food and personal care products. Environ. Sci. Technol. 46, 2242–2250.
https://doi.org/10.1021/es204168d
Weisburg, W.G., Barns, S.M., Pelletier, D.A., Lane, D.J., 1991. 16S ribosomal DNA
amplification for phylogenetic study. J. Bacteriol. 173, 697–703.
https://doi.org/10.1128/jb.173.2.697-703.1991

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
63
Welker, T.L., Lim, C., Yildirim-Aksoy, M., Shelby, R., Klesius, P.H., 2007. Immune Response
and Resistance to Stress and Edwardsiella ictaluri Challenge in Channel Catfish,
Ictalurus punctatus, Fed Diets Containing Commercial Whole-Cell Yeast or Yeast
Subcomponents. J. World Aquac. Soc. 38, 24–35. https://doi.org/10.1111/j.1749-
7345.2006.00070.x
Wilkes Walburn, J., Wemheuer, B., Thomas, T., Copeland, E., O’Connor, W., Booth, M.,
Fielder, S., Egan, S., 2019. Diet and diet-associated bacteria shape early microbiome
development in Yellowtail Kingfish (Seriola lalandi). Microb. Biotechnol. 12, 275–288.
https://doi.org/10.1111/1751-7915.13323
Woodhams, D.C., Bletz, M.C., Becker, C.G., Bender, H.A., Buitrago-Rosas, D., Diebboll, H.,
Huynh, R., Kearns, P.J., Kueneman, J., Kurosawa, E., Labumbard, B.C., Lyons, C.,
McNally, K., Schliep, K., Shankar, N., Tokash-Peters, A.G., Vences, M., Whetstone, R.,
2020. Host-associated microbiomes are predicted by immune system complexity and
climate. Genome Biol. 21, 1–20. https://doi.org/10.1186/s13059-019-1908-8
Wu, Y., Xiao, F., Wang, C., Shu, L., Zheng, X., Xu, K., Yu, X., Zhang, K., Luo, H., Yang, Y.,
He, Z., Yan, Q., 2020. The Beta-Diversity of Siganus fuscescens-Associated Microbial
Communities From Different Habitats Increases With Body Weight. Front. Microbiol. 11,
1562. https://doi.org/10.3389/fmicb.2020.01562
Xavier, R., Mazzei, R., Pérez-Losada, M., Rosado, D., Santos, J.L., Veríssimo, A., Soares,
M.C., 2019. A Risky Business? Habitat and Social Behavior Impact Skin and Gut
Microbiomes in Caribbean Cleaning Gobies. Front. Microbiol. 10, 716.
https://doi.org/10.3389/fmicb.2019.00716
Xavier, R., Pereira, A., Pagan, A., Hendrick, G.C., Nicholson, M.D., Rosado, D., Soares, M.C.,
Pérez-Losada, M., Sikkel, P.C., 2020. The effects of environment and ontogeny on the
skin microbiome of two Stegastes damselfishes (Pomacentridae) from the eastern
Caribbean Sea. Mar. Biol. 167, 102. https://doi.org/10.1007/s00227-020-03717-7
Xia, J.H., Lin, G., Fu, G.H., Wan, Z.Y., Lee, M., Wang, L., Liu, X.J., Yue, G.H., 2014. The
intestinal microbiome of fish under starvation. BMC Genomics 15, 1–11.
https://doi.org/10.1186/1471-2164-15-266
Xie, Y., Wu, G., Tang, J., Luo, R., Patterson, J., Liu, S., Huang, W., He, G., Gu, S., Li, S.,
Zhou, X., Lam, T.-W., Li, Y., Xu, X., Wong, G.K.-S., Wang, J., 2014. SOAPdenovo-Trans:
de novo transcriptome assembly with short RNA-Seq reads. Bioinformatics 30, 1660–
1666. https://doi.org/10.1093/bioinformatics/btu077
Xiong, J.B., Nie, L., Chen, J., 2019. Current understanding on the roles of gut microbiota in
fish disease and immunity. Zool. Res. https://doi.org/10.24272/j.issn.2095-
8137.2018.069
Yan, Q., Li, J., Yu, Y., Wang, J., He, Z., Van Nostrand, J.D., Kempher, M.L., Wu, L., Wang,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
64
Y., Liao, L., Li, X., Wu, S., Ni, J., Wang, C., Zhou, J., 2016. Environmental filtering
decreases with fish development for the assembly of gut microbiota. Environ. Microbiol.
18, 4739–4754. https://doi.org/10.1111/1462-2920.13365
Yang, G., Jian, S.Q., Cao, H., Wen, C., Hu, B., Peng, M., Peng, L., Yuan, J., Liang, L., 2019.
Changes in microbiota along the intestine of grass carp (Ctenopharyngodon idella):
Community, interspecific interactions, and functions. Aquaculture 498, 151–161.
https://doi.org/10.1016/j.aquaculture.2018.08.062
Yarza, P., Yilmaz, P., Pruesse, E., Glöckner, F.O., Ludwig, W., Schleifer, K.H., Whitman, W.B.,
Euzéby, J., Amann, R., Rosselló-Móra, R., 2014. Uniting the classification of cultured and
uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol.
12, 635–645. https://doi.org/10.1038/nrmicro3330
Ye, L., Amberg, J., Chapman, D., Gaikowski, M., Liu, W.T., 2014. Fish gut microbiota analysis
differentiates physiology and behavior of invasive Asian carp and indigenous American
fish. ISME J. 8, 541–551. https://doi.org/10.1038/ismej.2013.181
Yildirimer, C.C., Brown, K.H., 2018. Intestinal microbiota lipid metabolism varies across
rainbow trout (Oncorhynchus mykiss) phylogeographic divide. J. Appl. Microbiol. 125,
1614–1625. https://doi.org/10.1111/jam.14059
Zang, L., Ma, Y., Huang, W., Ling, Y., Sun, L., Wang, X., Zeng, A., Dahlgren, R.A., Wang, C.,
Wang, H., 2019. Dietary Lactobacillus plantarum ST-III alleviates the toxic effects of
triclosan on zebrafish (Danio rerio) via gut microbiota modulation. Fish Shellfish Immunol.
84, 1157–1169. https://doi.org/10.1016/j.fsi.2018.11.007
Zarkasi, K.Z., Abell, G.C.J., Taylor, R.S., Neuman, C., Hatje, E., Tamplin, M.L., Katouli, M.,
Bowman, J.P., 2014. Pyrosequencing-based characterization of gastrointestinal bacteria
of Atlantic salmon (Salmo salar L.) within a commercial mariculture system. J. Appl.
Microbiol. 117, 18–27. https://doi.org/10.1111/jam.12514
Zha, Y., Eiler, A., Johansson, F., Svanbäck, R., 2018. Effects of predation stress and food
ration on perch gut microbiota. Microbiome 6, 1–12. https://doi.org/10.1186/s40168-018-
0400-0
Zhang, M.-L., Li, M., Sheng, Y., Tan, F., Chen, L., Cann, I., Du, Z.-Y., 2020. Citrobacter
Species Increase Energy Harvest by Modulating Intestinal Microbiota in Fish:
Nondominant Species Play Important Functions . mSystems 5.
https://doi.org/10.1128/msystems.00303-20
Zhang, M., Sun, Y., Liu, Y., Qiao, F., Chen, L., Liu, W.T., Du, Z., Li, E., 2016. Response of gut
microbiota to salinity change in two euryhaline aquatic animals with reverse salinity
preference. Aquaculture 454, 72–80. https://doi.org/10.1016/j.aquaculture.2015.12.014
Zhang, X., Zhang, D., Jia, H., Feng, Q., Wang, D., Liang, D., Wu, X., Li, Junhua, Tang, L., Li,
Yin, Lan, Z., Chen, B., Li, Yanli, Zhong, H., Xie, H., Jie, Z., Chen, Weineng, Tang, S., Xu,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
65
Xiaoqiang, Wang, X., Cai, X., Liu, S., Xia, Y., Li, Jiyang, Qiao, X., Al-Aama, J.Y., Chen,
H., Wang, L., Wu, Q.J., Zhang, F., Zheng, W., Li, Yongzhe, Zhang, M., Luo, G., Xue, W.,
Xiao, L., Li, Jun, Chen, Wanting, Xu, Xun, Yin, Y., Yang, H., Wang, Jian, Kristiansen, K.,
Liu, L., Li, T., Huang, Q., Li, Yingrui, Wang, Jun, 2015. The oral and gut microbiomes are
perturbed in rheumatoid arthritis and partly normalized after treatment. Nat. Med. 21,
895–905. https://doi.org/10.1038/nm.3914
Zhang, Xiaoting, Ding, L., Yu, Y., Kong, W., Yin, Y., Huang, Z., Zhang, Xuezhen, Xu, Z., 2018.
The Change of Teleost Skin Commensal Microbiota Is Associated With Skin Mucosal
Transcriptomic Responses During Parasitic Infection by Ichthyophthirius multifillis. Front.
Immunol. 9, 2972. https://doi.org/10.3389/fimmu.2018.02972
Zhang, Y., Zhang, P., Shang, X., Lu, Y., Li, Y., 2021. Exposure of lead on intestinal structural
integrity and the diversity of gut microbiota of common carp. Comp. Biochem. Physiol.
Part - C Toxicol. Pharmacol. 239, 108877. https://doi.org/10.1016/j.cbpc.2020.108877
Zhang, Z., Li, D., Xu, W., Tang, R., Li, L., 2019. Microbiome of Co-cultured Fish Exhibits Host
Selection and Niche Differentiation at the Organ Scale. Front. Microbiol. 10, 2576.
https://doi.org/10.3389/fmicb.2019.02576
Zhou, L., Limbu, S.M., Shen, M., Zhai, W., Qiao, F., He, A., Du, Z.Y., Zhang, M., 2018.
Environmental concentrations of antibiotics impair zebrafish gut health. Environ. Pollut.
235, 245–254. https://doi.org/10.1016/j.envpol.2017.12.073
Zhou, W., Sailani, M.R., Contrepois, K., Zhou, Y., Ahadi, S., Leopold, S.R., Zhang, M.J., Rao,
V., Avina, M., Mishra, T., Johnson, J., Lee-McMullen, B., Chen, S., Metwally, A.A., Tran,
T.D.B., Nguyen, H., Zhou, X., Albright, B., Hong, B.Y., Petersen, L., Bautista, E., Hanson,
B., Chen, L., Spakowicz, D., Bahmani, A., Salins, D., Leopold, B., Ashland, M., Dagan-
Rosenfeld, O., Rego, S., Limcaoco, P., Colbert, E., Allister, C., Perelman, D., Craig, C.,
Wei, E., Chaib, H., Hornburg, D., Dunn, J., Liang, L., Rose, S.M.S.F., Kukurba, K.,
Piening, B., Rost, H., Tse, D., McLaughlin, T., Sodergren, E., Weinstock, G.M., Snyder,
M., 2019. Longitudinal multi-omics of host–microbe dynamics in prediabetes. Nature 569,
663–671. https://doi.org/10.1038/s41586-019-1236-x

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
66
CHAPTER 2
Characterization of the skin and gill microbiomes of
the farmed seabass (Dicentrarchus labrax) and
seabream (Sparus aurata)
Daniela Rosado, Marcos Pérez-Losada, Ricardo Severino, Jo Cable,
Raquel Xavier
2019. Aquaculture 500, 57-64. https://doi.org/10.1016/j.aquaculture.2018.09.063.
2.1 Abstract There is substantial evidence showing that the microbiome of teleosts plays a key role
in host health and wellbeing. Aquaculture practices increase the risk of dysbiosis (i.e. microbial
imbalance), which is known to facilitate pathogen infections. The skin and gills are the primary
defense organs against pathogens, thus, characterizing their microbiome composition in
farmed fish is pivotal for detecting potential alterations that may lead to disease susceptibility.
Here, we assessed the skin and gill microbiomes of two of the most important adult fish
species farmed in southern Europe, the seabass and the seabream, during winter months.
We coupled next-generation sequencing (MiSeq) of the 16S rRNA V4 region with the DADA2
bioinformatic pipeline to assess microbial composition and structure. Variation in microbial
alpha-diversity (intra-sample) and taxa proportions were assessed using analysis of variance.
Differences in beta-diversity (between-sample) were tested using permutational multivariate
analysis of variance. Microbiomes of both tissues (n=30 per species) identified 19 bacteria
phyla, dominated by the phyla Proteobacteria (44 - 68%) and Bacteroidetes (15 - 37%); the
families Flavobacteriaceae (11 - 28%), Rhodobacteraeae (4 - 8%) and Vibrionaceae (2 - 17%);
and the genera Rubritalea (4 - 13%), Pseudomonas (4 - 8%) and the NS3a marine group (4 -
12%). Mean relative proportion of these taxa, some alpha-diversity indices and all beta-
diversity distances varied significantly between tissues within and between species. ASVs
belonging to the genera Polaribacter and Vibrio, which include several species that are
pathogenic, were detected in the core microbiomes of seabass or seabream.
Keywords: aquaculture, 16S rRNA, microbiome, pathogens, fish farm

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
67
2.2 Introduction Seminal studies conducted in mammals established a link between the microbiome
and the host’s innate immune response, with implications for host health and wellbeing (see
for example reviews Belkaid and Hand, 2014; Lynch and Pedersen, 2016; Nelson et al., 2014).
Furthermore, differences in microbial composition can account for differential disease
susceptibility in humans (e.g., Börnigen et al., 2013; Dunn et al., 2016; Pérez-Losada et al.,
2018, 2015) and teleosts (reviewed by Kelly and Salinas, 2017). Particularly problematic for
fish, pathogenic bacteria that naturally reside in the aquatic environment can also form part of
their microbiomes (e.g., Borchardt et al., 2003; Califano et al., 2017; Rivas et al., 2011; Rud
et al., 2017) and cause disease if there is a shift in abundance (i.e., dysbiosis) (e.g., Hess et
al., 2015). While the skin microbiome of unstressed fish is dominated by taxa known for their
probiotic and antimicrobial activity, the microbiome of stressed fish is dominated by potential
pathogens (see Boutin et al., 2013). Although mucosal surfaces, such as skin, gills and the
gut, do act as primary barriers to disease (reviewed by Gómez and Balcázar, 2008), they can
be affected by several pathogens (e.g. Aeromonas septicemia, see Balebona et al., 1998;
Doukas et al., 1998), which may cause significant losses. Aquaculture practices also impact
microbial communities in the epidermal mucosa of fish. Overcrowding and low oxygen
concentrations, typical in fish farms, result in host stress and induce dysbiosis in the skin
microbiome, facilitating the proliferation of opportunistic pathogens (e.g., Boutin et al., 2013).
At the same time, infectious diseases that frequently affect farmed fish can also induce
dysbiosis, generally favouring increased abundance of opportunistic bacteria creating
complex feedback mechanisms (e.g., Llewellyn et al., 2017; Reid et al., 2017).
Seabass (Dicentrarchus labrax) and seabream (Sparus aurata) are the two most
important fish species farmed in southern Europe; their productivity, however, is greatly
affected by infectious diseases, which can account for losses of 15% to 40%, respectively
(Lane et al., 2014). Given the role skin and gill microbiomes play in fish innate immunity
(Gourzioti et al., 2016; Pellizzari et al., 2013) and the economic impact of diseases in fish
aquaculture, characterizing the microbiomes of these two fishes is paramount. Additionally,
anthropogenic stressors (e.g., rise of sea temperature and pollution) and farming conditions
(e.g., high densities) aggravate bacterial diseases causing external lesions in skin and gills
(e.g., photobacteriosis and vibriosis) of farmed seabass and seabream (e.g., Avendaño-
Herrera et al., 2006; Bakopoulos et al., 2018; Frans et al., 2011; Gourzioti et al., 2016;
Pellizzari et al., 2013; Weber et al., 2010). To this end, identifying potential fish pathogens
could help to design more efficient prevention and treatment strategies. The first assessment
of the skin microbiome of adult seabass and seabream showed that inter-individual variability
was comparable to interspecific variability (Chiarello et al., 2015). Recently, Tapia-Paniagua

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
68
et al. (2018) found a reduction in beneficial bacteria from the skin microbiomes of ulcered
compared to healthy seabream. Differences in microbiome diversity of the skin of seabass
have also been assessed in three different fish farms located in Ria de Aveiro, northern
Portugal (Pimentel et al., 2017). Despite high inter-individual variation, microbial composition
was found to act as a unique signature of each individual’s geographic origin (Pimentel et al.,
2017). Although the authors controlled for ontogenetic effects, which affect skin microbiome
(e.g., Sylvain and Derome, 2017), they acknowledged that other factors, such as different
farming practices and probiotic use, may have explained some of the observed differences
(Pimentel et al., 2017). Moreover, previous disease history (e.g., Llewellyn et al., 2017; Reid
et al., 2017; Tapia-Paniagua et al., 2018) and host physiology (e.g., Apprill et al., 2014) may
have influenced the composition of fish skin microbiomes. Other key factor impacting microbial
composition differences between groups is sample size and longitudinal (time) variation
(Knight et al., 2018). Not surprisingly, skin and gill microbiome composition of seabass and
seabream varied greatly between previous studies, as they have been cross-sectional (one
time point) and included few individuals (Chiarello et al., 2015; Pimentel et al., 2017; Tapia-
Paniagua et al., 2018).
In the present study, we monitored the microbiome composition and structure of the
skin and gills of 30 seabass and 30 seabream healthy adults over winter (December to
February) using 16S rRNA next-generation sequencing (MiSeq). Our main aims here were to
characterize the baseline diversity of the skin and gill microbiomes of these two farmed
species and identify potential pathogens or opportunistic bacteria.
2.3 Material and methods
2.3.1 Sample collection and preparation Thirty individuals of both seabass and seabream were collected in 19 of December
2016, 16 of January 2017 and 13 of February 2017 (10 specimens of each species per month)
from a commercial fish farm located in an estuarine environment, the Ria Formosa (Portimão),
southern Portugal. Seabass and seabream sampled were about 2 years old and individuals
weighted on average, 384 g and 318 g. The fish were reared in two separate ponds, at a
density of ca. 4.4 kg/m3 (ca. 130 individual seabass) and 5.2 kg/m3 (ca. 150 individual
seabream), with the same open water circulation systems, thus subjected to the same
environmental conditions. The mean water temperature 30 days before each sampling point
was 16.6 ºC, 15.3 ºC and 14.4 ºC, and the photoperiod for each sampling point was 9 h 35
min, 9 h 54 min and 10 h 45 min, respectively. All fish were fed with the same commercial
feed and they shared the same clinical history. All fish were considered healthy, with no
external lesions and no pathologies detected during the sampling period. Individuals were

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
69
randomly caught from each tank using a fishing pole, and skin and gill swabs were collected
using tubed sterile dry swabs (Medical Wire & Equipment, UK). Skin samples were taken by
swabbing several times along the right upper lateral part of the fish from head to tail; gill swabs
were taken from the right filaments between the first and second arch. Swabs were
immediately stored at -20ºC until transported on dry ice to the CIBIO laboratory by airmail
where they were kept at -80ºC until processing. DNA from a total of 120 samples (60 skin and
60 gills) was extracted using the PowerSoil DNA Isolation Kit (QIAGEN, Netherlands),
following the manufacturer’s protocol. DNA concentration was measured with the NanoDropTM
2000 Spectrophotometer (Thermo Fisher Scientific, USA) and extractions were sent on dry
ice by airmail to the University of Michigan Medical School (USA) for amplification and
sequencing according to the protocol of Kozich et al. (2013). Each sample was amplified for
the V4 hypervariable region of the 16S rRNA gene (~250 bp), which has been widely used to
characterize microbiomes from vertebrates (Earth Microbiome Project, Gilbert et al., 2014),
including fish (e.g., Carlson et al., 2017; Llewellyn et al., 2014; Thompson et al., 2017; Wang
et al., 2017). Amplicon libraries were sequenced in a single run of the Illumina MiSeq
sequencing platform.
2.3.2 Data and statistical analyses Raw FASTQ files were analyzed using the Quantitative Insights Into Microbial Ecology
2 (QIIME2; release 2018.4) platform. Clean sequences were aligned against the SILVA (132
release) reference database (Quast et al., 2013) with DADA2 pipeline (Callahan et al., 2016).
Samples were rarefied to the minimum read count (9,087) and a feature table containing
amplicon sequence variants (ASVs) was constructed. ASVs with less than 0.01% of reads
across samples were eliminated (Nelson et al., 2014). The core microbiome was assessed for
the skin and gill of seabass and seabream, separately. An ASV was considered as part of the
core microbiome if present in 100% of samples in each group. Rarefaction curves were
performed to examine sampling depth (Figure S2.1).
Microbial taxonomic alpha-diversity (intra-sample) was calculated using Shannon,
ACE, Fisher and Faith’s phylogenetic diversity (PD) indices as implemented in the R package
phyloseq (McMurdie and Holmes, 2013). Species beta-diversity (inter-sample) was estimated
using phylogenetic Unifrac (unweighted and weighted) and Bray-Curtis distances. Dissimilarity
between samples was assessed by principal coordinates analysis (PCoA). Variation in
microbial alpha-diversity and taxa composition were assessed using one-way analysis of
variance (ANOVA). Differences in community composition (beta-diversity) were tested using
permutational multivariate analysis of variance for Unweighted and Weighted Unifrac and
Bray-Curtis indices with 1,000 permutations, as implemented in the adonis function of the R

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
70
vegan package. In our microbiome statistical analyses we compared i) tissues within each fish
species (skin x gills) and ii) fish species within each tissue (seabass x seabream) – see Table
2.2. We used the three sampling months (December to February) as temporal replicates,
rendering a total of 30 microbiome samples per comparison per tissue. All analyses were
performed in R studio v1.0.143 (RStudio, 2012).
2.4 Results
2.4.1 Taxonomic bacterial composition and core microbiome of
seabass and seabream Approximately 3.2 million raw reads were retrieved (1.7 million for seabass and 1.5
million for seabream) and the number of sequences per sample ranged between 9,087 and
3,537,652. These sequences corresponded to 8,136 unique ASVs, from which ASVs with less
than 0.01% of sequences across all samples and ASVs belonging to Archaea were removed,
resulting in 556 unique ASVs and 3,246,429 sequences. Of the 457 ASVs found in the skin of
the seabass, only 24 were common to all individuals sampled, thus forming the core
microbiome (Table 2.1). Of the 466 ASVs found on the gills of the seabass, only 7 were shared
among all individuals. The same pattern was observed in the seabream, where 15 out of 532
skin ASVs and 2 out of 539 gill ASVs were present in all individuals (Table 2.1). These results
highlight the high inter-individual variability found in both tissues, especially the gills (Table
2.1, Figures 2.1 and 2.2).
Of the total 19 bacteria phyla identified across all samples, Proteobacteria and
Bacteroidetes were the most abundant in both tissues (Figure 2.1, Table 2.1). ASVs from four
(Proteobacteria, Bacteroidetes, Chlamydiae and Verrucomicrobia) of these 19 phyla formed
part of the core microbiome (Figure 2.2). Moreover, the phyla Dependentiae (0.2% of ASVs
and sequences) and Patescibacteria (0.2% of ASV, 0.1% of sequences) were unique to the
microbiome of seabream, while the phyla Spirochaetes (0.2% of ASVs and 0.1% of
sequences) was unique to the gill microbiome of seabass.
The phyla Proteobacteria and Bacteroidetes accounted for 69% to 72% of all ASVs
and 62% to 87% of all sequences in both species and for 50% to 93% of all phyla in the core
microbiomes (Figure 2.2, Table 2.1). It was possible to identify 106 families, from which ASVs
belonging to 16 families formed the core microbiome of both species. Altogether,
Flavobacteriaceae (Bacteroidetes), Rhodobacteraeae (Proteobacteria) and Vibrionaceae
(Proteobacteria) accounted for 19% to 21% of ASVs, 17% to 51% of sequences, and 29% to
50% of all families in the core microbiome of both tissues (Figure 2.2, Table 2.1). From the
117 genera identified, ASVs belonging to 16 of these genera formed the core microbiome of
both species. The NS3a marine group (4% - 12%), Rubritalea (4% - 13%) and Pseudomonas
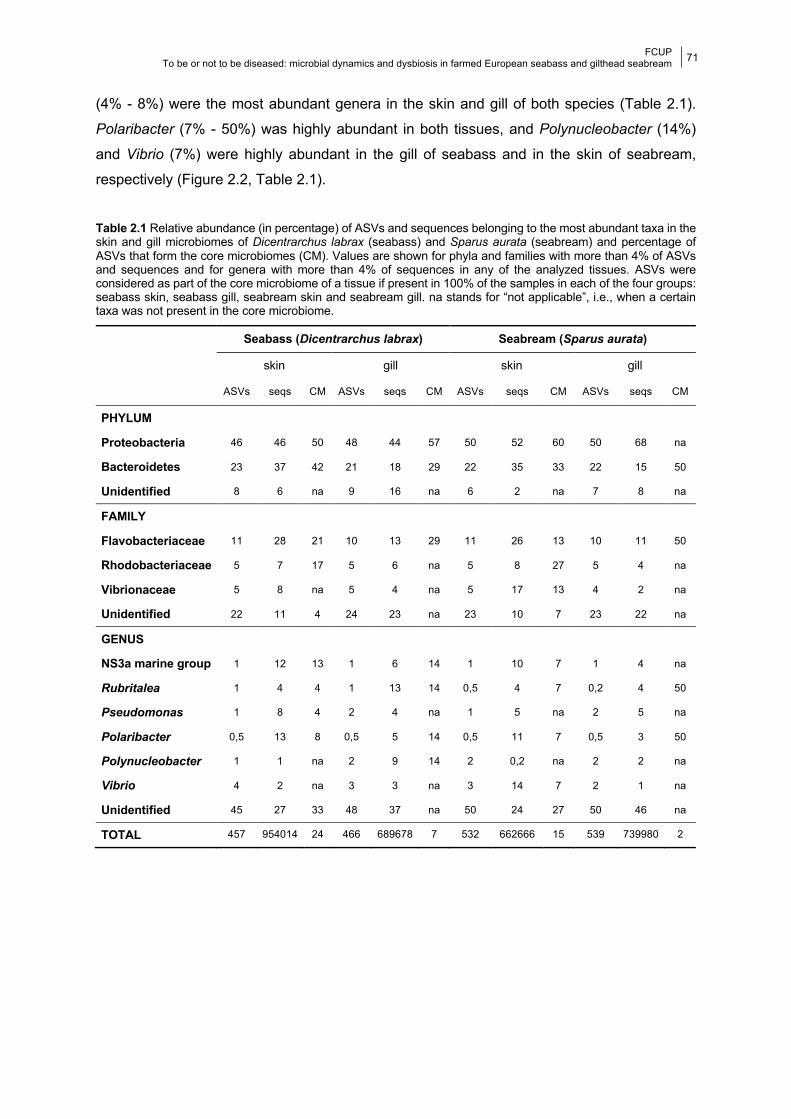
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
71
(4% - 8%) were the most abundant genera in the skin and gill of both species (Table 2.1).
Polaribacter (7% - 50%) was highly abundant in both tissues, and Polynucleobacter (14%)
and Vibrio (7%) were highly abundant in the gill of seabass and in the skin of seabream,
respectively (Figure 2.2, Table 2.1).
Table 2.1 Relative abundance (in percentage) of ASVs and sequences belonging to the most abundant taxa in the skin and gill microbiomes of Dicentrarchus labrax (seabass) and Sparus aurata (seabream) and percentage of ASVs that form the core microbiomes (CM). Values are shown for phyla and families with more than 4% of ASVs and sequences and for genera with more than 4% of sequences in any of the analyzed tissues. ASVs were considered as part of the core microbiome of a tissue if present in 100% of the samples in each of the four groups: seabass skin, seabass gill, seabream skin and seabream gill. na stands for “not applicable”, i.e., when a certain taxa was not present in the core microbiome.
Seabass (Dicentrarchus labrax) Seabream (Sparus aurata)
skin gill skin gill
ASVs seqs CM ASVs seqs CM ASVs seqs CM ASVs seqs CM
PHYLUM
Proteobacteria 46 46 50 48 44 57 50 52 60 50 68 na
Bacteroidetes 23 37 42 21 18 29 22 35 33 22 15 50
Unidentified 8 6 na 9 16 na 6 2 na 7 8 na
FAMILY
Flavobacteriaceae 11 28 21 10 13 29 11 26 13 10 11 50
Rhodobacteriaceae 5 7 17 5 6 na 5 8 27 5 4 na
Vibrionaceae 5 8 na 5 4 na 5 17 13 4 2 na
Unidentified 22 11 4 24 23 na 23 10 7 23 22 na
GENUS
NS3a marine group 1 12 13 1 6 14 1 10 7 1 4 na
Rubritalea 1 4 4 1 13 14 0,5 4 7 0,2 4 50
Pseudomonas 1 8 4 2 4 na 1 5 na 2 5 na
Polaribacter 0,5 13 8 0,5 5 14 0,5 11 7 0,5 3 50
Polynucleobacter 1 1 na 2 9 14 2 0,2 na 2 2 na
Vibrio 4 2 na 3 3 na 3 14 7 2 1 na
Unidentified 45 27 33 48 37 na 50 24 27 50 46 na
TOTAL 457 954014 24 466 689678 7 532 662666 15 539 739980 2
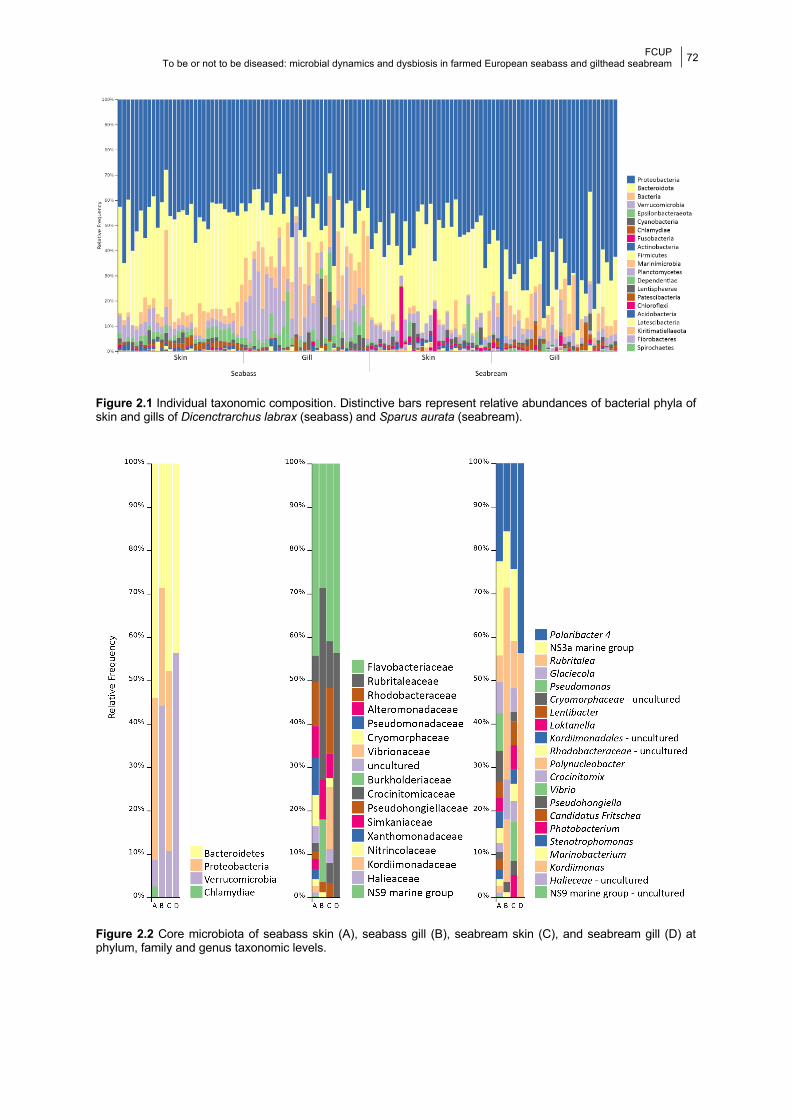
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
72
Figure 2.1 Individual taxonomic composition. Distinctive bars represent relative abundances of bacterial phyla of skin and gills of Dicenctrarchus labrax (seabass) and Sparus aurata (seabream).
Figure 2.2 Core microbiota of seabass skin (A), seabass gill (B), seabream skin (C), and seabream gill (D) at phylum, family and genus taxonomic levels.
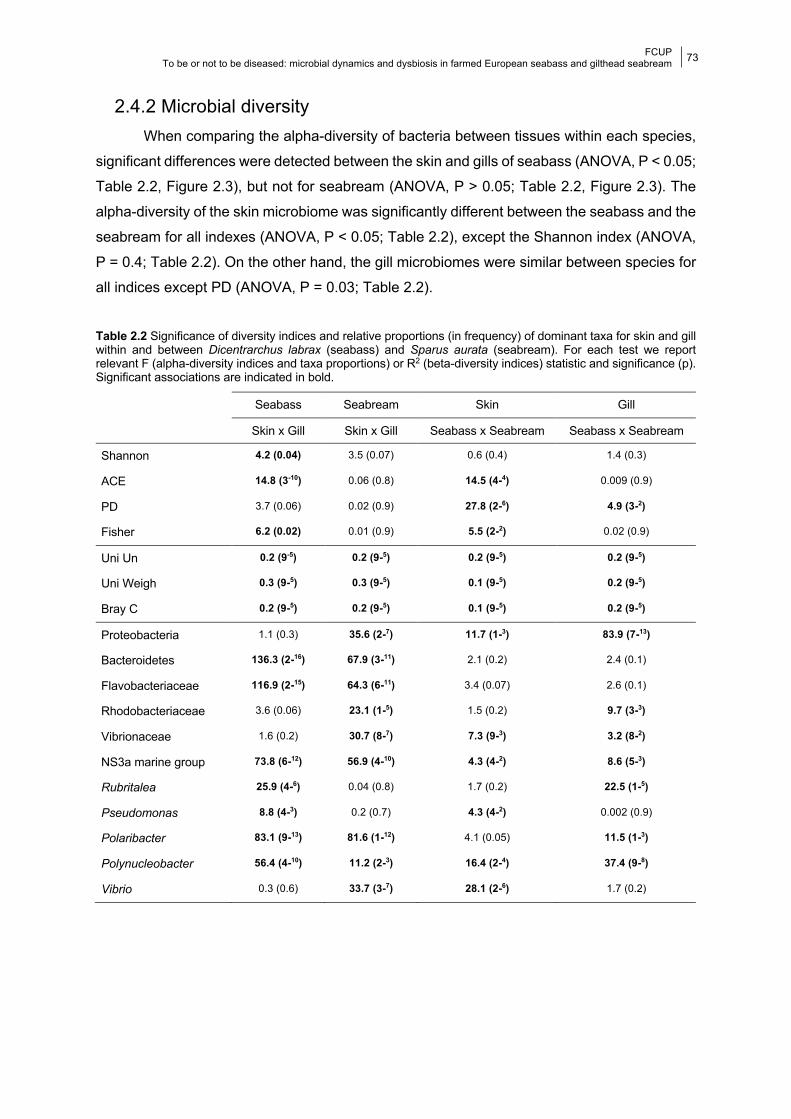
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
73
2.4.2 Microbial diversity When comparing the alpha-diversity of bacteria between tissues within each species,
significant differences were detected between the skin and gills of seabass (ANOVA, P < 0.05;
Table 2.2, Figure 2.3), but not for seabream (ANOVA, P > 0.05; Table 2.2, Figure 2.3). The
alpha-diversity of the skin microbiome was significantly different between the seabass and the
seabream for all indexes (ANOVA, P < 0.05; Table 2.2), except the Shannon index (ANOVA,
P = 0.4; Table 2.2). On the other hand, the gill microbiomes were similar between species for
all indices except PD (ANOVA, P = 0.03; Table 2.2).
Table 2.2 Significance of diversity indices and relative proportions (in frequency) of dominant taxa for skin and gill within and between Dicentrarchus labrax (seabass) and Sparus aurata (seabream). For each test we report relevant F (alpha-diversity indices and taxa proportions) or R2 (beta-diversity indices) statistic and significance (p). Significant associations are indicated in bold.
Seabass Seabream Skin Gill
Skin x Gill Skin x Gill Seabass x Seabream Seabass x Seabream
Shannon 4.2 (0.04) 3.5 (0.07) 0.6 (0.4) 1.4 (0.3)
ACE 14.8 (3-10) 0.06 (0.8) 14.5 (4-4) 0.009 (0.9)
PD 3.7 (0.06) 0.02 (0.9) 27.8 (2-6) 4.9 (3-2)
Fisher 6.2 (0.02) 0.01 (0.9) 5.5 (2-2) 0.02 (0.9)
Uni Un 0.2 (9-5) 0.2 (9-5) 0.2 (9-5) 0.2 (9-5)
Uni Weigh 0.3 (9-5) 0.3 (9-5) 0.1 (9-5) 0.2 (9-5)
Bray C 0.2 (9-5) 0.2 (9-5) 0.1 (9-5) 0.2 (9-5)
Proteobacteria 1.1 (0.3) 35.6 (2-7) 11.7 (1-3) 83.9 (7-13)
Bacteroidetes 136.3 (2-16) 67.9 (3-11) 2.1 (0.2) 2.4 (0.1)
Flavobacteriaceae 116.9 (2-15) 64.3 (6-11) 3.4 (0.07) 2.6 (0.1)
Rhodobacteriaceae 3.6 (0.06) 23.1 (1-5) 1.5 (0.2) 9.7 (3-3)
Vibrionaceae 1.6 (0.2) 30.7 (8-7) 7.3 (9-3) 3.2 (8-2)
NS3a marine group 73.8 (6-12) 56.9 (4-10) 4.3 (4-2) 8.6 (5-3)
Rubritalea 25.9 (4-6) 0.04 (0.8) 1.7 (0.2) 22.5 (1-5)
Pseudomonas 8.8 (4-3) 0.2 (0.7) 4.3 (4-2) 0.002 (0.9)
Polaribacter 83.1 (9-13) 81.6 (1-12) 4.1 (0.05) 11.5 (1-3)
Polynucleobacter 56.4 (4-10) 11.2 (2-3) 16.4 (2-4) 37.4 (9-8)
Vibrio 0.3 (0.6) 33.7 (3-7) 28.1 (2-6) 1.7 (0.2)
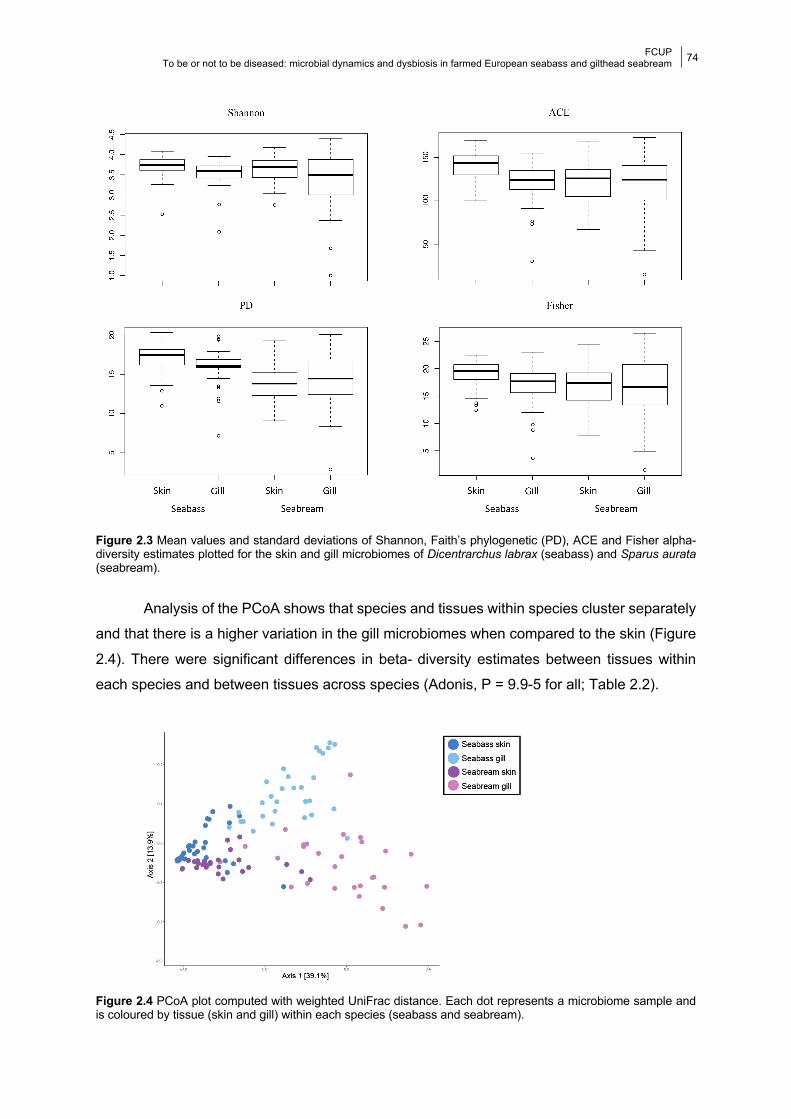
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
74
Figure 2.3 Mean values and standard deviations of Shannon, Faith’s phylogenetic (PD), ACE and Fisher alpha-diversity estimates plotted for the skin and gill microbiomes of Dicentrarchus labrax (seabass) and Sparus aurata (seabream).
Analysis of the PCoA shows that species and tissues within species cluster separately
and that there is a higher variation in the gill microbiomes when compared to the skin (Figure
2.4). There were significant differences in beta- diversity estimates between tissues within
each species and between tissues across species (Adonis, P = 9.9-5 for all; Table 2.2).
Figure 2.4 PCoA plot computed with weighted UniFrac distance. Each dot represents a microbiome sample and is coloured by tissue (skin and gill) within each species (seabass and seabream).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
75
Mean proportions of bacterial taxa varied between the two fish species and tissues
(Table 2.2). In the seabass, the abundance of Bacteroidetes, Flavobacteriaceae, NS3a marine
group, Rubritalea, Pseudomonas, Polaribacter and Polynucleobacter were significantly
different between the skin and gill (ANOVA, P < 0.05, Table 2.2). In the seabream, the mean
proportion of Proteobacteria, Bacteroidetes, Flavobacteriaceae, Rhodobacteriaceae,
Vibrionaceae, NS3a marine group, Polaribacter, Polynucleobacter and Vibrio varied
significantly (ANOVA, P < 0.05) between the skin and gill microbiomes (Table 2.2). Finally,
Proteobacteria, Vibrionaceae, NS3a marine group, Pseudomonas, Polaribacter,
Polynucleobacter, and Vibrio varied significantly between the skin microbiomes of seabass
and seabream (ANOVA, P < 0.05, Table 2.2), while Proteobacteria, Rhodobacteriaceae,
Vibrionaceae, NS3a marine group, Rubritalea, Polaribacter and Polynucleobacter varied
significantly between their gill microbiomes (ANOVA, P < 0.05, Table 2.2).
2.5 Discussion Characterizing the microbiome composition and structure of the mucosal surfaces of
economically important fish species, such as the seabass and the seabream, is of paramount
importance in order to detect imbalances and prevent potential disease outbreaks in fish
farms. Here, we showed significant differences in both the composition and structure of the
microbial communities residing in the skin and gills of seabass and seabream, which is in line
with previous findings of both fish species (Chiarello et al., 2015). The skin microbiomes were
found to be species-specific as in other fish species (e.g., the stripped mullet, red snapper,
spotted seatrout, sand seatrout, pinfish and Atlantic croaker; Larsen et al., 2013). Despite the
high inter-individual variation, overall, the seabream microbiomes were less diverse than those
of the seabass (Figure 2.3).
2.5.1 Core microbiome composition Proteobacteria (50 - 60%) and Bacteroidetes (29 - 50%, Table 2.1) formed the main
components of the skin and gill microbiomes of seabass and seabream. Proteobacteria is the
most common phylum reported in the skin and gill microbiomes of teleosts (see for example
the review by Llewellyn et al., 2014), including the skin microbiome of seabass and seabream
(Chiarello et al., 2015; Pimentel et al., 2017; Tapia-Paniagua et al., 2018). A predominance of
the phylum Bacteroidetes has also been previously reported in seabass and seabream
(Chiarello et al., 2015; Tapia-Paniagua et al., 2018), as well as in the skin of many other fishes,
such as in the brook char (Boutin et al., 2014), rainbow trout (Lowrey et al., 2015), channel
catfish (Larsen et al., 2014), tambaqui (Sylvain et al., 2016), among others (see Doane et al.,
2017; Larsen et al., 2013, 2015; Legrand et al., 2018; Leonard et al., 2014). The gill

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
76
microbiome of the bluefin tuna (Valdenegro-Vega et al., 2013), rainbow trout (Lowrey et al.,
2015) and yellowtail kingfish (Legrand et al., 2018) were also found to be dominated by
Bacteroidetes.
In the present study, from the 16 genera identified in the core microbiome of the skin
and gill of adult seabass and seabream, the highest percentage of amplicon sequence variants
(ASVs) belonged to the NS3a marine group, Rubritalea and Pseudomonas genera. Besides
these three, the microbiome of seabass also exhibited an elevated abundance of the genera
Polaribacter, Polynucleobacter and Arcobacter; while the microbiome of seabream included
high abundance of Polaribacter and Vibrio. The genus Pseudomonas has been previously
reported to be highly represented in the skin microbiome of seabass (Pimentel et al., 2017),
cod (Wilson et al., 2008), mosquitofish (Leonard et al., 2014), gulf killifish (Larsen et al., 2015)
and others (see Colwell and Liston, 1932; Horsley, 1977, 1973; Larsen et al., 2013). However,
we found some differences in microbial composition at the genus level in comparison with
previous studies of seabass and seabream; Tapia-Paniagua et al. (2018), for example, found
Staphylococcus and Lactobacillus to be the most abundant in the skin microbiome of
seabream. This in not unexpected since, the skin microbiome of seabass comprises genera
that are unique signatures of specific earth growth ponds, even though these ponds were
geographically close (Pimentel et al., 2017). Besides spatial variation in fish location,
environmental conditions (such as water temperatures and water supply [e.g., Lokesh and
Kiron, 2016; Tapia-Paniagua et al., 2018]), host physiology and even clinical history (Apprill et
al., 2014; Llewellyn et al., 2017) could contribute to explain the observed differences.
Ontogenetic shifts in microbiome composition have been described in several fish
species (e.g., Atlantic salmon, Llewellyn et al., 2016; Zarkasi et al., 2014; Zebrafish, Stephens
et al., 2016; discus, Sylvain and Derome, 2017). The larval microbiome tends to reflect more
the microbial community of the surrounding water (Stephens et al., 2016; Sylvain and Derome,
2017), while adult fish harbour a more adapted and stable microbial community (e.g., Llewellyn
et al., 2016). Califano et al. (2017) even reported an increase in the microbiome composition
of seabream larvae between day 2 and day 34. This pattern, however, is far from being
universal as decreased diversity with age has been reported in other fish species (Stephens
et al., 2016; Yan et al., 2016).
As with most microbiome research, it is important to note any methodological
differences that might explain variation. One of such methodological differences relates to our
skin sampling method; Chiarello et al. (2015) used tissue from different fins, while in the
present study and in Tapia-Paniagua et al. (2018) and Pimentel et al. (2017) we targeted skin
mucous. Lowrey et al. (2015) uncovered high diversity of bacteria in the different dermal layers
of skin, suggesting that mucosal diversity is an underestimation of the actual skin microbial
diversity. Moreover, specifically for this study, the sequenced 16S variable region and the

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
77
sequencing platform might have impacted taxonomic assignment. We sequenced the V4
region by synthesis (MiSeq), while Chiarello et al. (2015), Pimentel et al. (2017) and Tapia-
Paniagua et al. (2018) used different combinations of other 16S regions and sequencing
platforms (pyrosequencing and sequencing by synthesis). While differences in outputs
provided by different sequencing methods are widely acknowledged (e.g., Frey et al., 2014;
Li et al., 2014), debate regarding the most appropriate region for microbiome studies is still
ongoing (e.g., Guo et al., 2013; Mizrahi-Man et al., 2013). Finally, these results are likely to
have been affected by the different analytical pipelines used to analyze the sequence data -
amplicon sequence variants (ASVs) in this study versus Operational Taxonomic Units (OTUs)
in previous studies.
2.5.2 Potential pathogens detected in the core microbiomes Several ASVs belonging to genera comprising opportunistic and potential pathogenic
bacteria were recovered from the skin and gill core microbiomes of apparently healthy
individuals of seabass and seabream. Polaribacter is one such taxa, recovered from the skin
and gills of both species (Figure 2.2, Table 2.1). This genus is often found in diseased fishes,
being considered opportunistic and colonizing already weakened hosts (Bornø and Linaker,
2015). Species from this genus seem to be common in fish farms and have been reported in
the water and biofilm of recirculating and semiclosed aquaculture systems rearing Atlantic
salmon, turbot and the Senegalese sole (Martins et al., 2013; Rud et al., 2017). The genus
Vibrio, present in the gills of seabass and in both the gills and skin core microbiomes of
seabream, harbours species associated with several diseases in these fish and many are
considered opportunistic pathogens (e.g., Pujalte et al., 2003; Weber et al., 2010). Vibrio
anguillarum and V. alginolyticus, for example, cause skin lesions, and V. splendidus has been
involved in several disease outbreaks (e.g., Frans et al., 2011; Pujalte et al., 2003). V. harveyi
is another important pathogen causing tail rot disease in farmed seabream (Austin and Zhang,
2006; Haldar et al., 2010), comprising many strains that are fatal to seabass (Pujalte et al.,
2003b). However, due to taxonomic assignment limitations, it was not possible to ascertain to
which species these ASVs belonged to and if they are, indeed, pathogenic. If so, the high
prevalence of these ASVs means that, in case of dysbiosis, these bacteria may overgrow and
impact fish health.
2.6 Conclusion The skin and gills of fish are exposed to many pathogens present in the marine and
freshwater environment and represent an important barrier preventing pathogen invasion
(e.g., Trivedi, 2012). The links between microbiome composition and disease resistance are

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
78
now well established in mammals and teleosts (e.g., Britton and Young, 2014; Gomez et al.,
2013; Gómez and Balcázar, 2008; Kelly and Salinas, 2017), and alterations in the microbiome
often precede the onset of disease (reviewed by Munang’andu et al., 2018). Microbial
imbalance, however, is not easily detected unless baseline information regarding microbiome
composition and structure are established. Here, we describe the skin and gills microbiomes
of farmed healthy seabass and seabream adults through three sampling months, thus yielding
a more robust assessment of the microbiome of these two species. Our results show that
seabass and seabream host different microbiomes despite sharing the same environment.
Furthermore, high levels of intra- and inter-individual variability were found across tissues.
Additionally, several potential pathogens were detected in the core microbiome of both
species, which could lead to potential disease outbreaks during dysbiosis.
2.7 Acknowledgements This work was funded by the European Regional Development Fund (ERDF) through the
COMPETE program and by National Funds through FCT - Foundation for Science and
Technology (project PTDC/MAR-BIO/0902/2014 -POCI01-0145-FEDER-016550; project
POCI-01-0145-FEDER-027995; and by a “Projecto de Investigação Exploratória”:
IF/00764/2013); the Welsh Government and Higher Education Funding Council for Wales
(HEFCW) AquaWales Project through the Sêr Cymru National Research Network for Low
Carbon Energy and Environment (NRNLCEE). DR, MPL and RX are supported by FCT under
the Programa Operacional Potencial Humano – Quadro de Referência Estratégico Nacional
funds from the European Social Fund and Portuguese Ministério da Educação e Ciência (DR
doctoral grant SFRH/BD/117943/2016; MPL: IF/00764/2013; RX: IF/00359/2015).
2.8 References Apprill, A., Robbins, J., Eren, A.M., Pack, A.A., Reveillaud, J., Mattila, D., Moore, M.,
Niemeyer, M., Moore, K.M.T., Mincer, T.J., 2014. Humpback Whale Populations Share a
Core Skin Bacterial Community: Towards a Health Index for Marine Mammals? PLoS
One 9, e90785. https://doi.org/10.1371/journal.pone.0090785
Austin, B., Zhang, X.-H., 2006. Vibrio harveyi: a significant pathogen of marine vertebrates
and invertebrates. Lett. Appl. Microbiol. 43, 119–124. https://doi.org/10.1111/j.1472-
765X.2006.01989.x
Avendaño-Herrera, R., Toranzo, A., Magariños, B., 2006. Tenacibaculosis infection in marine
fish caused by Tenacibaculum maritimum: a review. Dis. Aquat. Organ. 71, 255–266.
https://doi.org/10.3354/dao071255
Bakopoulos, V., Kosma, I., Laspa, E., 2018. Quantitative and qualitative analysis of sea

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
79
bream, Sparus aurata (L.), humoral immune response, vaccinated with commercial and
experimental vaccines against vibriosis and photobacteriosis. J. Mar. Biol. Assoc. United
Kingdom 98, 105–115. https://doi.org/10.1017/S0025315417001631
Balebona, M.C., Zorrilla, I., Moriñigo, M.A., Borrego, J.J., 1998. Survey of bacterial pathologies
affecting farmed gilt-head sea bream (Sparus aurata L.) in southwestern Spain from 1990
to 1996. Aquaculture 166, 19–35. https://doi.org/10.1016/S0044-8486(98)00282-8
Belkaid, Y., Hand, T.W., 2014. Role of the microbiota in immunity and inflammation. Cell.
https://doi.org/10.1016/j.cell.2014.03.011
Borchardt, M.A., Bertz, P.D., Spencer, S.K., Battigelli, D.A., 2003. Incidence of enteric viruses
in groundwater from household wells in Wisconsin. Appl. Environ. Microbiol. 69, 1172–
1180. https://doi.org/10.1128/AEM.69.2.1172-1180.2003
Börnigen, D., Morgan, X.C., Franzosa, E.A., Ren, B., Xavier, R.J., Garrett, W.S., Huttenhower,
C., 2013. Functional profiling of the gut microbiome in disease-associated inflammation.
Genome Med. https://doi.org/10.1186/gm469
Bornø, G., Linaker, M.L., 2015. The health situation in Norwegian aquaculture, 2014.
Boutin, S., Bernatchez, L., Audet, C., Derôme, N., 2013. Network Analysis Highlights Complex
Interactions between Pathogen, Host and Commensal Microbiota. PLoS One 8, e84772.
https://doi.org/10.1371/journal.pone.0084772
Boutin, S., Sauvage, C., Bernatchez, L., Audet, C., Derome, N., 2014. Inter Individual
Variations of the Fish Skin Microbiota: Host Genetics Basis of Mutualism? PLoS One 9,
e102649. https://doi.org/10.1371/journal.pone.0102649
Britton, R.A., Young, V.B., 2014. Role of the intestinal microbiota in resistance to colonization
by Clostridium difficile. Gastroenterology 146, 1547–1553.
https://doi.org/10.1053/j.gastro.2014.01.059
Califano, G., Castanho, S., Soares, F., Ribeiro, L., Cox, C.J., Mata, L., Costa, R., 2017.
Molecular Taxonomic Profiling of Bacterial Communities in a Gilthead Seabream (Sparus
aurata) Hatchery. Front. Microbiol. 8, 204. https://doi.org/10.3389/fmicb.2017.00204
Callahan, B.J., McMurdie, P.J., Rosen, M.J., Han, A.W., Johnson, A.J.A., Holmes, S.P., 2016.
DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13,
581–583. https://doi.org/10.1038/nmeth.3869
Carlson, J.M., Leonard, A.B., Hyde, E.R., Petrosino, J.F., Primm, T.P., 2017. Microbiome
disruption and recovery in the fish Gambusia affinis following exposure to broad-
spectrum antibiotic. Infect. Drug Resist. 10, 143–154.
https://doi.org/10.2147/IDR.S129055
Chiarello, M., Villéger, S., Bouvier, C., Bettarel, Y., Bouvier, T., 2015. High diversity of skin-
associated bacterial communities of marine fishes is promoted by their high variability
among body parts, individuals and species. FEMS Microbiol. Ecol. 91.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
80
https://doi.org/10.1093/femsec/fiv061
Colwell, R.R., Liston, J., 1932. Bacterial flora of seven species of fish collected at Rongelap
and Eniwetok atolls, Fellers. Kiser and Beckwith.
Doane, M.P., Haggerty, J.M., Kacev, D., Papudeshi, B., Dinsdale, E.A., 2017. The skin
microbiome of the common thresher shark (Alopias vulpinus) has low taxonomic and
gene function β-diversity. Environ. Microbiol. Rep. 9, 357–373.
https://doi.org/10.1111/1758-2229.12537
Doukas, V., Athanassopoulou, F., Karagouni, E., Dotsika, E. (Department of P.I.H.P.I.A.
(Greece)), 1998. Aeromonas hydrophila infection in cultured sea bass, Dicentrarchus
labrax L., and Puntazzo puntazzo Cuvier from the Aegean sea. J. Fish Dis. (United
Kingdom).
Dunn, K.A., Moore-Connors, J., MacIntyre, B., Stadnyk, A., Thomas, N.A., Noble, A., Mahdi,
G., Rashid, M., Otley, A.R., Bielawski, J.P., Van Limbergen, J., 2016. The Gut
Microbiome of Pediatric Crohnʼs Disease Patients Differs from Healthy Controls in Genes
That Can Influence the Balance Between a Healthy and Dysregulated Immune
Response. Inflamm. Bowel Dis. 22, 2607–2618.
https://doi.org/10.1097/MIB.0000000000000949
Frans, I., Michiels, C.W., Bossier, P., Willems, K.A., Lievens, B., Rediers, H., 2011. Vibrio
anguillarum as a fish pathogen: virulence factors, diagnosis and prevention. J. Fish Dis.
34, 643–661. https://doi.org/10.1111/j.1365-2761.2011.01279.x
Frey, K.G., Herrera-Galeano, J.E., Redden, C.L., Luu, T. V., Servetas, S.L., Mateczun, A.J.,
Mokashi, V.P., Bishop-Lilly, K.A., 2014. Comparison of three next-generation sequencing
platforms for metagenomic sequencing and identification of pathogens in blood. BMC
Genomics 15, 1–14. https://doi.org/10.1186/1471-2164-15-96
Gilbert, J.A., Jansson, J.K., Knight, R., 2014. The Earth Microbiome project: Successes and
aspirations. BMC Biol. https://doi.org/10.1186/s12915-014-0069-1
Gomez, D., Sunyer, J.O., Salinas, I., 2013. The mucosal immune system of fish: The evolution
of tolerating commensals while fighting pathogens. Fish Shellfish Immunol.
https://doi.org/10.1016/j.fsi.2013.09.032
Gómez, G.D., Balcázar, J.L., 2008. A review on the interactions between gut microbiota and
innate immunity of fish: Table 1. FEMS Immunol. Med. Microbiol. 52, 145–154.
https://doi.org/10.1111/j.1574-695X.2007.00343.x
Gourzioti, E., Kolygas, M.N., Athanassopoulou, F., Babili, V., 2016. Tenacibaculosis in
aquaculture farmed marine fish. J. Hell. Vet. Med. Soc.
https://doi.org/10.12681/jhvms.15620
Guo, F., Ju, F., Cai, L., Zhang, T., 2013. Taxonomic Precision of Different Hypervariable
Regions of 16S rRNA Gene and Annotation Methods for Functional Bacterial Groups in

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
81
Biological Wastewater Treatment. PLoS One 8, e76185.
https://doi.org/10.1371/journal.pone.0076185
Haldar, S., Maharajan, A., Chatterjee, S., Hunter, S.A., Chowdhury, N., Hinenoya, A., Asakura,
M., Yamasaki, S., 2010. Identification of Vibrio harveyi as a causative bacterium for a tail
rot disease of sea bream Sparus aurata from research hatchery in Malta. Microbiol. Res.
165, 639–648. https://doi.org/10.1016/j.micres.2009.12.001
Hess, S., Wenger, A.S., Ainsworth, T.D., Rummer, J.L., 2015. Exposure of clownfish larvae to
suspended sediment levels found on the Great Barrier Reef: Impacts on gill structure and
microbiome. Sci. Rep. 5, 1–8. https://doi.org/10.1038/srep10561
Horsley, R.W., 1977. A review of the bacterial flora of teleosts and elasmobranchs, including
methods for its analysis. J. Fish Biol. 10, 529–553. https://doi.org/10.1111/j.1095-
8649.1977.tb04086.x
Horsley, R.W., 1973. The Bacterial Flora of the Atlantic Salmon ( Salmo salar L.) in Relation
to its Environment. J. Appl. Bacteriol. 36, 377–386. https://doi.org/10.1111/j.1365-
2672.1973.tb04119.x
Kelly, C., Salinas, I., 2017. Under pressure: Interactions between commensal microbiota and
the teleost immune system. Front. Immunol. https://doi.org/10.3389/fimmu.2017.00559
Knight, R., Vrbanac, A., Taylor, B.C., Aksenov, A., Callewaert, C., Debelius, J., Gonzalez, A.,
Kosciolek, T., McCall, L.I., McDonald, D., Melnik, A. V., Morton, J.T., Navas, J., Quinn,
R.A., Sanders, J.G., Swafford, A.D., Thompson, L.R., Tripathi, A., Xu, Z.Z., Zaneveld,
J.R., Zhu, Q., Caporaso, J.G., Dorrestein, P.C., 2018. Best practices for analysing
microbiomes. Nat. Rev. Microbiol. https://doi.org/10.1038/s41579-018-0029-9
Kozich, J.J., Westcott, S.L., Baxter, N.T., Highlander, S.K., Schloss, P.D., 2013. Development
of a dual-index sequencing strategy and curation pipeline for analyzing amplicon
sequence data on the miseq illumina sequencing platform. Appl. Environ. Microbiol. 79,
5112–5120. https://doi.org/10.1128/AEM.01043-13
Lane, A., Hough, C., Bostock, J., 2014. The Long Term Economic and Ecological Impact of
Larger Sustainable Aquaculture.
Larsen, A., Tao, Z., Bullard, S.A., Arias, C.R., 2013. Diversity of the skin microbiota of fishes:
evidence for host species specificity. FEMS Microbiol. Ecol. 85, 483–494.
https://doi.org/10.1111/1574-6941.12136
Larsen, A.M., Bullard, S.A., Womble, M., Arias, C.R., 2015. Community Structure of Skin
Microbiome of Gulf Killifish, Fundulus grandis, Is Driven by Seasonality and Not Exposure
to Oiled Sediments in a Louisiana Salt Marsh. Microb. Ecol. 70, 534–544.
https://doi.org/10.1007/s00248-015-0578-7
Larsen, A.M., Mohammed, H.H., Arias, C.R., 2014. Characterization of the gut microbiota of
three commercially valuable warmwater fish species. J. Appl. Microbiol. 116, 1396–1404.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
82
https://doi.org/10.1111/jam.12475
Legrand, T.P.R.A., Catalano, S.R., Wos-Oxley, M.L., Stephens, F., Landos, M., Bansemer,
M.S., Stone, D.A.J., Qin, J.G., Oxley, A.P.A., 2018. The Inner Workings of the Outer
Surface: Skin and Gill Microbiota as Indicators of Changing Gut Health in Yellowtail
Kingfish. Front. Microbiol. 8, 2664. https://doi.org/10.3389/fmicb.2017.02664
Leonard, A.B., Carlson, J.M., Bishoff, D.E., Sendelbach, S.I., Yung, S.B., Ramzanali, S.,
Manage, A.B.W., Hyde, E.R., Petrosino, J.F., Primm, T.P., 2014. The Skin Microbiome
of Gambusia affinis Is Defined and Selective. Adv. Microbiol. 04, 335–343.
https://doi.org/10.4236/aim.2014.47040
Li, J.Z., Chapman, B., Charlebois, P., Hofmann, O., Weiner, B., Porter, A.J., Samuel, R.,
Vardhanabhuti, S., Zheng, L., Eron, J., Taiwo, B., Zody, M.C., Henn, M.R., Kuritzkes,
D.R., Hide, W., 2014. Comparison of Illumina and 454 Deep Sequencing in Participants
Failing Raltegravir-Based Antiretroviral Therapy. PLoS One 9, e90485.
https://doi.org/10.1371/journal.pone.0090485
Llewellyn, M.S., Boutin, S., Hoseinifar, S.H., Derome, N., 2014. Teleost microbiomes: The
state of the art in their characterization, manipulation and importance in aquaculture and
fisheries. Front. Microbiol. https://doi.org/10.3389/fmicb.2014.00207
Llewellyn, M.S., Leadbeater, S., Garcia, C., Sylvain, F.E., Custodio, M., Ang, K.P., Powell, F.,
Carvalho, G.R., Creer, S., Elliot, J., Derome, N., 2017. Parasitism perturbs the mucosal
microbiome of Atlantic Salmon. Sci. Rep. 7, 1–10. https://doi.org/10.1038/srep43465
Llewellyn, M.S., McGinnity, P., Dionne, M., Letourneau, J., Thonier, F., Carvalho, G.R., Creer,
S., Derome, N., 2016. The biogeography of the atlantic salmon (Salmo salar) gut
microbiome. ISME J. 10, 1280–1284. https://doi.org/10.1038/ismej.2015.189
Lokesh, J., Kiron, V., 2016. Transition from freshwater to seawater reshapes the skin-
associated microbiota of Atlantic salmon. Sci. Rep. 6, 1–10.
https://doi.org/10.1038/srep19707
Lowrey, L., Woodhams, D.C., Tacchi, L., Salinas, I., 2015. Topographical mapping of the
rainbow trout (Oncorhynchus mykiss) microbiome reveals a diverse bacterial community
with antifungal properties in the skin. Appl. Environ. Microbiol. 81, 6915–6925.
https://doi.org/10.1128/AEM.01826-15
Lynch, S. V., Pedersen, O., 2016. The Human Intestinal Microbiome in Health and Disease.
N. Engl. J. Med. 375, 2369–2379. https://doi.org/10.1056/NEJMra1600266
Martins, P., Cleary, D.F.R., Pires, A.C.C., Rodrigues, A.M., Quintino, V., Calado, R., Gomes,
N.C.M., 2013. Molecular Analysis of Bacterial Communities and Detection of Potential
Pathogens in a Recirculating Aquaculture System for Scophthalmus maximus and Solea
senegalensis. PLoS One 8, e80847. https://doi.org/10.1371/journal.pone.0080847
McMurdie, P.J., Holmes, S., 2013. phyloseq: An R Package for Reproducible Interactive

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
83
Analysis and Graphics of Microbiome Census Data. PLoS One 8, e61217.
https://doi.org/10.1371/journal.pone.0061217
Mizrahi-Man, O., Davenport, E.R., Gilad, Y., 2013. Taxonomic Classification of Bacterial 16S
rRNA Genes Using Short Sequencing Reads: Evaluation of Effective Study Designs.
PLoS One 8, e53608. https://doi.org/10.1371/journal.pone.0053608
Munang’andu, H., Galindo-Villegas, J., David, L., 2018. Teleosts Genomics: Progress and
Prospects in Disease Prevention and Control. Int. J. Mol. Sci. 19, 1083.
https://doi.org/10.3390/ijms19041083
Nelson, M.C., Morrison, H.G., Benjamino, J., Grim, S.L., Graf, J., 2014. Analysis, Optimization
and Verification of Illumina-Generated 16S rRNA Gene Amplicon Surveys. PLoS One 9,
e94249. https://doi.org/10.1371/journal.pone.0094249
Pellizzari, C., Krasnov, A., Afanasyev, S., Vitulo, N., Franch, R., Pegolo, S., Patarnello, T.,
Bargelloni, L., 2013. High mortality of juvenile gilthead sea bream (Sparus aurata) from
photobacteriosis is associated with alternative macrophage activation and anti-
inflammatory response: Results of gene expression profiling of early responses in the
head kidney. Fish Shellfish Immunol. 34, 1269–1278.
https://doi.org/10.1016/j.fsi.2013.02.007
Pérez-Losada, M., Castro-Nallar, E., Bendall, M.L., Freishtat, R.J., Crandall, K.A., 2015. Dual
Transcriptomic Profiling of Host and Microbiota during Health and Disease in Pediatric
Asthma. PLoS One 10, e0131819. https://doi.org/10.1371/journal.pone.0131819
Pérez-Losada, M., Graham, R.J., Coquillette, M., Jafarey, A., Castro-Nallar, E., Aira, M.,
Hoptay, C., Freishtat, R.J., Mansbach, J.M., 2018. Tracheal Microbiota in Patients with a
Tracheostomy Before, during and after an Acute Respiratory Infection. Pediatr. Infect.
Dis. J. 37, E269–E271. https://doi.org/10.1097/INF.0000000000001952
Pimentel, T., Marcelino, J., Ricardo, F., Soares, A.M.V.M., Calado, R., 2017. Bacterial
communities 16S rDNA fingerprinting as a potential tracing tool for cultured seabass
Dicentrarchus labrax. Sci. Rep. 7, 1–10. https://doi.org/10.1038/s41598-017-11552-y
Pujalte, M.J., Sitjà-Bobadilla, A., Álvarez-Pellitero, P., Garay, E., 2003a. Carriage of potentially
fish-pathogenic bacteria in Sparus aurata cultured in Mediterranean fish farms. Dis.
Aquat. Organ. 54, 119–126. https://doi.org/10.3354/dao054119
Pujalte, M.J., Sitjà-Bobadilla, A., Macián, M.C., Belloch, C., Álvarez-Pellitero, P., Pérez-
Sánchez, J., Uruburu, F., Garay, E., 2003b. Virulence and molecular typing of Vibrio
harveyi strains isolated from cultured dentex, gilthead sea bream and European sea
bass. Syst. Appl. Microbiol. 26, 284–292. https://doi.org/10.1078/072320203322346146
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., Peplies, J., Glöckner,
F.O., 2013. The SILVA ribosomal RNA gene database project: Improved data processing
and web-based tools. Nucleic Acids Res. 41, D590–D596.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
84
https://doi.org/10.1093/nar/gks1219
Reid, K.M., Patel, S., Robinson, A.J., Bu, L., Jarungsriapisit, J., Moore, L.J., Salinas, I., 2017.
Salmonid alphavirus infection causes skin dysbiosis in Atlantic salmon (Salmo salar L.)
post-smolts. PLoS One 12, e0172856. https://doi.org/10.1371/journal.pone.0172856
Rivas, A.J., Balado, M., Lemos, M.L., Osorio, C.R., 2011. The Photobacterium damselae
subsp. damselae hemolysins damselysin and HlyA are encoded within a new virulence
plasmid. Infect. Immun. 79, 4617–4627. https://doi.org/10.1128/IAI.05436-11
Rud, I., Kolarevic, J., Holan, A.B., Berget, I., Calabrese, S., Terjesen, B.F., 2017. Deep-
sequencing of the bacterial microbiota in commercial-scale recirculating and semi-closed
aquaculture systems for Atlantic salmon post-smolt production. Aquac. Eng. 78, 50–62.
https://doi.org/10.1016/j.aquaeng.2016.10.003
Stephens, W.Z., Burns, A.R., Stagaman, K., Wong, S., Rawls, J.F., Guillemin, K., Bohannan,
B.J.M., 2016. The composition of the zebrafish intestinal microbial community varies
across development. ISME J. 10, 644–654. https://doi.org/10.1038/ismej.2015.140
Studio, R., 2012. RStudio: integrated development environment for R.
Sylvain, F.É., Cheaib, B., Llewellyn, M., Gabriel Correia, T., Barros Fagundes, D., Luis Val,
A., Derome, N., 2016. PH drop impacts differentially skin and gut microbiota of the
Amazonian fish tambaqui (Colossoma macropomum). Sci. Rep. 6, 1–10.
https://doi.org/10.1038/srep32032
Sylvain, F.É., Derome, N., 2017. Vertically and horizontally transmitted microbial symbionts
shape the gut microbiota ontogenesis of a skin-mucus feeding discus fish progeny. Sci.
Rep. 7, 1–14. https://doi.org/10.1038/s41598-017-05662-w
Tapia-Paniagua, S.T., Ceballos-Francisco, D., Balebona, M.C., Esteban, M.Á., Moriñigo,
M.Á., 2018. Mucus glycosylation, immunity and bacterial microbiota associated to the
skin of experimentally ulcered gilthead seabream (Sparus aurata). Fish Shellfish
Immunol. 75, 381–390. https://doi.org/10.1016/j.fsi.2018.02.006
Thompson, L.R., Sanders, J.G., McDonald, D., Amir, A., Ladau, J., Locey, K.J., Prill, R.J.,
Tripathi, A., Gibbons, S.M., Ackermann, G., Navas-Molina, J.A., Janssen, S., Kopylova,
E., Vázquez-Baeza, Y., González, A., Morton, J.T., Mirarab, S., Xu, Z.Z., Jiang, L.,
Haroon, M.F., Kanbar, J., Zhu, Q., Song, S.J., Kosciolek, T., Bokulich, N.A., Lefler, J.,
Brislawn, C.J., Humphrey, G., Owens, S.M., Hampton-Marcell, J., Berg-Lyons, D.,
McKenzie, V., Fierer, N., Fuhrman, J.A., Clauset, A., Stevens, R.L., Shade, A., Pollard,
K.S., Goodwin, K.D., Jansson, J.K., Gilbert, J.A., Knight, R., Agosto Rivera, J.L., Al-
Moosawi, L., Alverdy, J., Amato, K.R., Andras, J., Angenent, L.T., Antonopoulos, D.A.,
Apprill, A., Armitage, D., Ballantine, K., Bárta, J., Baum, J.K., Berry, A., Bhatnagar, A.,
Bhatnagar, M., Biddle, J.F., Bittner, L., Boldgiv, B., Bottos, E., Boyer, D.M., Braun, J.,
Brazelton, W., Brearley, F.Q., Campbell, A.H., Caporaso, J.G., Cardona, C., Carroll, J.L.,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
85
Cary, S.C., Casper, B.B., Charles, T.C., Chu, H., Claar, D.C., Clark, R.G., Clayton, J.B.,
Clemente, J.C., Cochran, A., Coleman, M.L., Collins, G., Colwell, R.R., Contreras, M.,
Crary, B.B., Creer, S., Cristol, D.A., Crump, B.C., Cui, D., Daly, S.E., Davalos, L.,
Dawson, R.D., Defazio, J., Delsuc, F., Dionisi, H.M., Dominguez-Bello, M.G., Dowell, R.,
Dubinsky, E.A., Dunn, P.O., Ercolini, D., Espinoza, R.E., Ezenwa, V., Fenner, N., Findlay,
H.S., Fleming, I.D., Fogliano, V., Forsman, A., Freeman, C., Friedman, E.S., Galindo, G.,
Garcia, L., Garcia-Amado, M.A., Garshelis, D., Gasser, R.B., Gerdts, G., Gibson, M.K.,
Gifford, I., Gill, R.T., Giray, T., Gittel, A., Golyshin, P., Gong, D., Grossart, H.P., Guyton,
K., Haig, S.J., Hale, V., Hall, R.S., Hallam, S.J., Handley, K.M., Hasan, N.A., Haydon,
S.R., Hickman, J.E., Hidalgo, G., Hofmockel, K.S., Hooker, J., Hulth, S., Hultman, J.,
Hyde, E., Ibáñez-Álamo, J.D., Jastrow, J.D., Jex, A.R., Johnson, L.S., Johnston, E.R.,
Joseph, S., Jurburg, S.D., Jurelevicius, D., Karlsson, A., Karlsson, R., Kauppinen, S.,
Kellogg, C.T.E., Kennedy, S.J., Kerkhof, L.J., King, G.M., Kling, G.W., Koehler, A. V.,
Krezalek, M., Kueneman, J., Lamendella, R., Landon, E.M., Lanede Graaf, K., LaRoche,
J., Larsen, P., Laverock, B., Lax, S., Lentino, M., Levin, I.I., Liancourt, P., Liang, W., Linz,
A.M., Lipson, D.A., Liu, Y., Lladser, M.E., Lozada, M., Spirito, C.M., MacCormack, W.P.,
MacRae-Crerar, A., Magris, M., Martín-Platero, A.M., Martín-Vivaldi, M., Martínez, L.M.,
Martínez-Bueno, M., Marzinelli, E.M., Mason, O.U., Mayer, G.D., McDevitt-Irwin, J.M.,
McDonald, J.E., McGuire, K.L., McMahon, K.D., McMinds, R., Medina, M., Mendelson,
J.R., Metcalf, J.L., Meyer, F., Michelangeli, F., Miller, K., Mills, D.A., Minich, J., Mocali,
S., Moitinho-Silva, L., Moore, A., Morgan-Kiss, R.M., Munroe, P., Myrold, D., Neufeld,
J.D., Ni, Y., Nicol, G.W., Nielsen, S., Nissimov, J.I., Niu, K., Nolan, M.J., Noyce, K.,
O’Brien, S.L., Okamoto, N., Orlando, L., Castellano, Y.O., Osuolale, O., Oswald, W.,
Parnell, J., Peralta-Sánchez, J.M., Petraitis, P., Pfister, C., Pilon-Smits, E., Piombino, P.,
Pointing, S.B., Pollock, F.J., Potter, C., Prithiviraj, B., Quince, C., Rani, A., Ranjan, R.,
Rao, S., Rees, A.P., Richardson, M., Riebesell, U., Robinson, C., Rockne, K.J.,
Rodriguezl, S.M., Rohwer, F., Roundstone, W., Safran, R.J., Sangwan, N., Sanz, V.,
Schrenk, M., Schrenzel, M.D., Scott, N.M., Seger, R.L., Seguinorlando, A., Seldin, L.,
Seyler, L.M., Shakhsheer, B., Sheets, G.M., Shen, C., Shi, Y., Shin, H., Shogan, B.D.,
Shutler, D., Siegel, J., Simmons, S., Sjöling, S., Smith, D.P., Soler, J.J., Sperling, M.,
Steinberg, P.D., Stephens, B., Stevens, M.A., Taghavi, S., Tai, V., Tait, K., Tan, C.L.,
Taş, N., Taylor, D.L., Thomas, T., Timling, I., Turner, B.L., Urich, T., Ursell, L.K., Van Der
Lelie, D., Van Treuren, W., Van Zwieten, L., Vargas-Robles, D., Thurber, R.V., Vitaglione,
P., Walker, D.A., Walters, W.A., Wang, S., Wang, T., Weaver, T., Webster, N.S., Wehrle,
B., Weisenhorn, P., Weiss, S., Werner, J.J., West, K., Whitehead, A., Whitehead, S.R.,
Whittingham, L.A., Willerslev, E., Williams, A.E., Wood, S.A., Woodhams, D.C., Yang,
Y., Zaneveld, J., Zarraonaindia, I., Zhang, Q., Zhao, H., 2017. A communal catalogue

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
86
reveals Earth’s multiscale microbial diversity. Nature 551, 457–463.
https://doi.org/10.1038/nature24621
Trivedi, B., 2012. Microbiome: the surface brigade. Nature 492, S60–S61.
Valdenegro-Vega, V., Naeem, S., Carson, J., Bowman, J.P., Tejedor del Real, J.L., Nowak,
B., 2013. Culturable microbiota of ranched southern bluefin tuna (Thunnus maccoyii
Castelnau). J. Appl. Microbiol. 115, 923–932. https://doi.org/10.1111/jam.12286
Wang, J., Tao, Q., Wang, Z., Mai, K., Xu, W., Zhang, Y., Ai, Q., 2017. Effects of fish meal
replacement by soybean meal with supplementation of functional compound additives on
intestinal morphology and microbiome of Japanese seabass ( Lateolabrax japonicus ).
Aquac. Res. 48, 2186–2197. https://doi.org/10.1111/are.13055
Weber, B., Chen, C., Milton, D.L., 2010. Colonization of fish skin is vital for Vibrio anguillarum
to cause disease. Environ. Microbiol. Rep. 2, 133–139. https://doi.org/10.1111/j.1758-
2229.2009.00120.x
Wilson, B., Danilowicz, B.S., Meijer, W.G., 2008. The Diversity of Bacterial Communities
Associated with Atlantic Cod Gadus morhua. Microb. Ecol. 55, 425–434.
https://doi.org/10.1007/s00248-007-9288-0
Yan, Q., Li, J., Yu, Y., Wang, J., He, Z., Van Nostrand, J.D., Kempher, M.L., Wu, L., Wang,
Y., Liao, L., Li, X., Wu, S., Ni, J., Wang, C., Zhou, J., 2016. Environmental filtering
decreases with fish development for the assembly of gut microbiota. Environ. Microbiol.
18, 4739–4754. https://doi.org/10.1111/1462-2920.13365
Zarkasi, K.Z., Abell, G.C.J., Taylor, R.S., Neuman, C., Hatje, E., Tamplin, M.L., Katouli, M.,
Bowman, J.P., 2014. Pyrosequencing-based characterization of gastrointestinal bacteria
of Atlantic salmon (Salmo salar L.) within a commercial mariculture system. J. Appl.
Microbiol. 117, 18–27. https://doi.org/10.1111/jam.12514
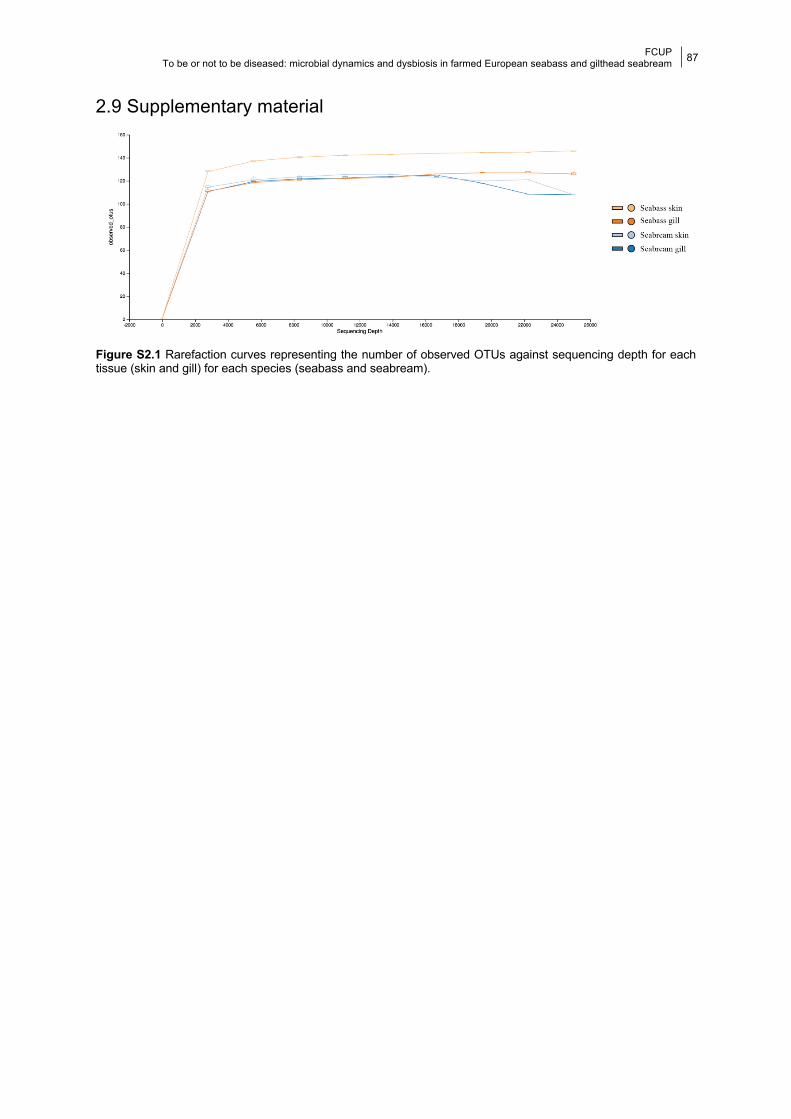
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
87
2.9 Supplementary material
Figure S2.1 Rarefaction curves representing the number of observed OTUs against sequencing depth for each tissue (skin and gill) for each species (seabass and seabream).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
88
CHAPTER 3
Effects of aging on the skin and gill microbiota of
farmed seabass and seabream
Daniela Rosado, Marcos Pérez-Losada, Ana Pereira, Ricardo Severino,
Raquel Xavier
2021. Animal Microbiome 3, 10. https://doi.org/10.1186/s42523-020-00072-2.
3.1 Abstract Important changes in microbial composition related to sexual maturation have been
already reported in the gut of several vertebrates including mammals, amphibians and fish.
Such changes in fish are linked to reproduction and growth during developmental stages, diet
transitions and critical life events. We used amplicon (16S rRNA) high-throughput sequencing
to characterize the skin and gill bacterial microbiota of farmed seabass and seabream
belonging to three different developmental age groups: early and late juveniles and mature
adults. We also assessed the impact of the surrounding estuarine water microbiota in shaping
the fish skin and gill microbiota. Microbial diversity, composition and predicted metabolic
functions varied across fish maturity stages. Alpha-diversity in the seabass microbiota varied
significantly between age groups and was higher in older fish. Conversely, in the seabream,
no significant differences were found in alpha-diversity between age groups. Microbial
structure varied significantly across age groups; moreover, high structural variation was also
observed within groups. Different bacterial metabolic pathways were predicted to be enriched
in the microbiota of both species. Finally, we found that the water microbiota was significantly
distinct from the fish microbiota across all the studied age groups, although a high percentage
of ASVs was shared with the skin and gill microbiotas. We report important microbial
differences in composition and potential functionality across different ages of farmed seabass
and seabream. These differences may be related to somatic growth and the onset of sexual
maturation. Importantly, some of the inferred metabolic pathways could enhance the fish
coping mechanisms during stressful conditions. Our results provide new evidence suggesting
that growth and sexual maturation have an important role in shaping the microbiota of the fish
external mucosae and highlight the importance of considering different life stages in microbiota
studies.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
89
Keywords: Dicentrarchus labrax, Sparus aurata, bacteria, ontogenesis, microbiome
3.2 Introduction Research on animal microbial communities (microbiota) is growing exponentially as
the link between microbiota and host health is strongly validated by emerging evidence (Jin
Song et al., 2019; Kelly and Salinas, 2017; Legrand et al., 2018; Murphy, 2018; Reid et al.,
2017; Ross et al., 2019; Tarnecki et al., 2018; Zhang et al., 2018). Age-related fluctuations in
the microbiota are well studied in humans and are considered as “natural, inevitable and
benign” (Nagpal et al., 2017). Critical microbial changes occur during infancy and old age,
coinciding with stages when the immune system is also more fragile (Nagpal et al., 2017).
Results linking changes in the gut microbiota to reproduction and growth (e.g., monkeys,
Amato et al., 2014) or disease resistance in early life stages (e.g., amphibians, Warne et al.,
2019) have been also found in other vertebrates.
In piscine hosts, most of the microbiota studies related to the effects of age are focused
on the gut (Bledsoe et al., 2016; Llewellyn et al., 2016, 2014; Lokesh et al., 2019; Nikouli et
al., 2019; Parris et al., 2016; Pratte et al., 2018; Stephens et al., 2016; Wilkes Walburn et al.,
2019; Wong et al., 2015; Yan et al., 2016). Some of these studies showed that microbial
communities in the surrounding waters influence the gut microbiota during early life stages,
which becomes increasingly unique with age (Stephens et al., 2016; Wilkes Walburn et al.,
2019). Indeed, initial microbiota colonization in animals is highly dependent on the
environment (e.g., Collado et al., 2015; Frese et al., 2015; Mueller et al., 2015; Quercia et al.,
2019; Villamil et al., 2018). Ecological factors, such as diet transitions (e.g., Wilkes Walburn
et al., 2019) or critical life events (e.g., habitat transition, Lokesh et al., 2019), which in turn
are intrinsically linked to sexual maturation, also play a major role in shaping the fish gut
microbiota. Importantly, most studies testing the role of age on fish microbiota were cross-
sectional and based on a single time point or a short time window (e.g., Llewellyn et al., 2016;
Nikouli et al., 2019; Parris et al., 2016; Wilkes Walburn et al., 2019; Wong et al., 2015). Thus,
given the high susceptibility of the fish microbiota to environmental changes and the high
interindividual microbiota variability (e.g., Larsen et al., 2015; Uren Webster et al., 2020, 2018),
the compound effect of all these factors could be hard to interpret (Thompson et al., 2017).
Fish skin and gills and their associated mucous and microbes form a natural physical
and chemical barrier to pathogens (Dash et al., 2018; Kelly and Salinas, 2017; Trivedi, 2012).
Despite this protective role, little is known about potential host developmental effects on skin
and gill microbiota. Filling out this knowledge gap is particularly important in fish farming,
where diseases are a main concern causing high mortality rates (e.g., Schmidt et al., 2018).
Two previous studies in wild reef fish comparing the gill (Pratte et al., 2018) and skin (Xavier

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
90
et al., 2020) microbiota of juvenile and mature adult fish from several species, showed a
general pattern of differentiation between life-stages with differences attributed to intraspecific
niche partitioning (Pratte et al., 2018; Xavier et al., 2020). Additionally, increases in body
weight were seen to be associated with an increase in the microbial structure (i.e., beta-
diversity) of the skin and gill microbiota of wild rabbitfish (Wu et al., 2020).
The European seabass (Dicentrarchus labrax) and the gilthead seabream (Sparus
aurata) are two of the most important farmed fish in Europe, with a global production of
191,003 tns and 185,980 tns, respectively, in 2016 (FAO, 2018). The gilthead seabream is a
protandric hermaphrodite, maturing first as males between years 1 and 2, with sex reversal
occurring in the following 2–3 years (Chaoui et al., 2006; Mehanna, 2007; Zohar and Gordin,
1979). The European seabass reaches sexual maturity between years 2 and 3 in males, and
after year 3 in females (Barnabe, 1991; Carillo et al., 1995; Felip et al., 2001). Typically, in
semi-extensive production systems, both fish are reared until they reach their first commercial
size (18–24 months). However, demand for larger fish sizes has been increasing, meaning
that both species can reach sexual maturity before harvest.
Here we used amplicon (16S rRNA) high-throughput sequencing to characterize the
skin and gill bacterial microbiota of farmed seabass and seabream from different ages
(juvenile stages and mature adults). Our main goal was to describe differences in composition,
structure and potential metabolic functions in their microbiota. Additionally, we investigated
the impact of the microbial communities present in the water column on the skin and gill
microbiotas.
3.3 Material and methods
3.3.1 Fish species, sampling and preparation Fish were sampled at a semi-intensive open-water farm in the Alvor Estuary (Ria
Formosa, Portimão, Portugal). In this fish farm, seabass and seabream production can take
up to 36 months to reach minimum commercial size, so having a healthy mucosa during this
time is of utter importance. The gilthead seabream is a protandric hermaphrodite, maturing
first as males between years 1 and 2 in the wild, with sex reversal occurring in the following
2–3 years (Chaoui et al., 2006; Mehanna, 2007; Zohar and Gordin, 1979). The European
seabass reaches sexual maturity between years 2 and 3 in males, and after year 3 in females
(Barnabe, 1991; Carillo et al., 1995; Felip et al., 2001). In this particular fish farm, seabass
typically reaches sexual maturity at approximately 275 g, whereas for seabream maturity is
usually attained at 300 g. We monitored the skin and gill microbiota of seabass and seabream
of different age cohorts, including juveniles and adults. Due to commercial restrictions within
the fish farm, sampling was strictly non-invasive and fish could not be dissected to confirm

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
91
sexual maturation. The categorization of the age group cohorts was based on previous studies
(e.g., Felip et al., 2001; Zohar and Gordin, 1979) and the weight at maturity records available
at the farm.
We collected samples every other week (12 sampling time points) between August
2017 and January 2018 (6 months). We simultaneously sampled three seabass age groups
with approximately 1 year difference. Seabass specimens were categorized as early juveniles
(9 months and an average weight of 22 g at the beginning of the study and 15 months and an
average weight of 76 g at the last sampling point), late juveniles (18 months and an average
weight of 151 g at the beginning of the study and 24 months and an average weight of 277 g
at the last sampling point), and mature adults (32 months and an average weight of 467 g at
the beginning of the study and 38 months and an average weight of 669 g at the last sampling
point). We also simultaneously sampled two seabream cohorts categorized as juveniles (15
months and an average weight of 103 g initially and 21 months and an average weight of 250
g at the last sampling point), and mature adults (31 months and an average weight of 411 g
at the beginning of the study and 37 months and an average weight of 476 g at the last
sampling point). Seabream from an intermediate age-cohort were not available.
Each age group and species was reared in separate but not distant open-water ponds
(maximum 344 m and 380 m apart for seabass and seabream, respectively; Figure S3.1). In
this fish farm, all ponds share the same water inflow, which is taken from a single point in the
estuary. Water in each pond is naturally recycled at each high tide (twice a day) and never
shared between ponds. Hence, fish share roughly the same water quality and environment.
Additionally, fry were bought from commercial hatcheries, which genetic variation is
considered low (Araki and Schmid, 2010).
Fish were caught from each tank using a fishing line, and skin and gill samples were
non-invasively taken using sterile swabs (Medical Wire & Equipment, UK). We swabbed the
right filaments between the first and second arches of the gill and the right upper lateral part
of the fish skin from head to tail. Afterwards fish were released unharmed. We collected water
samples (1 L) from the five different culture ponds at the same time as fish swabbing was
performed, except during the month of December, when no water samples could be collected.
Water samples were filtered through 0.2 μm cellulose nitrate filters on collection day. Swabs
and filters were immediately frozen at − 20 °C and then transported in dry ice to the CIBIO-
InBIO laboratory where they were kept at − 80 °C until processing.
We sampled five fish per week per age group, totaling 60 specimens per species and
age group. We processed 360 seabass samples (60 skin and 60 gills × 3 age groups) plus 29
water samples from their corresponding fishponds. We also processed 240 seabream
samples (60 skin and 60 gills × 2 age groups) plus 16 water samples from their corresponding
fishponds. Seabass and their corresponding water samples were processed using the

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
92
PowerSoil DNA Isolation Kit (QIAGEN, Netherlands), while seabream and corresponding
water samples were processed using the PureLink Microbiome DNA Purification Kit
(ThermoFisher Scientific, UK). We used two different DNA extraction kits due to supply
shortage at the time of extraction. This technical difference did not impact the goals of our
study since we studied each fish species separately (i.e., mucosa microbiotas are not
compared between fish species). We measured DNA concentration and quality in a
NanoDropTM 2000 Spectrophotometer (ThermoFisher Scientific, USA). DNA extractions were
shipped in dry ice to the University of Michigan Medical School (USA) for amplification and
sequencing according to the protocol of Kozich et al. (2013). Each sample was amplified for
the V4 hyper-variable region of the 16S rRNA gene (~ 250 bp). All amplicon libraries were
pooled and sequenced in a single run of the Illumina MiSeq sequencing platform.
Approximately 8,313,608 and 6,943,265 16S rRNA sequences were retrieved for
seabass and seabream, respectively. The number of sequences per sample ranged from 726
to 46,001 in seabass and from 5145 to 151,713 in seabream. After normalization and removal
of non-bacterial reads, 8724 and 5754 ASVs were assigned to the skin and gill, respectively,
of seabass; while 5308 and 3423 ASVs were assigned to the skin and gill, respectively, of
seabream. A total of 2543 ASVs were retrieved from the water samples collected in seabass
fishponds, while 1440 ASVs were retrieved from the waters of seabream fishponds. Microbial
taxa showing a mean relative proportion ≥ 5% in any group were considered the most
abundant taxa in that group.
3.3.2 Data processing and statistical analysis Raw FASTQ files were denoised using the DADA2 pipeline in R with the parameters
for filtering and trimming being trimLeft = 20, truncLen = c (220,200), maxN = 0, maxEE = c
(2,2), truncQ = 2 (Callahan et al., 2016). We estimated a midpoint rooted tree of ASVs using
the Quantitative Insights Into Microbial Ecology 2 package (QIIME2; release 2019.7). We
constructed a table containing amplicon sequence variants (ASVs) and made taxonomic
inferences against the SILVA (138 release) reference database (Quast et al., 2013). We
normalized ASV abundances using the negative binomial distribution (McMurdie and Holmes,
2014), which accounts for library size differences and biological variability.
Microbial taxonomic alpha-diversity (intra-sample) was calculated using Shannon,
Faith’s phylogenetic diversity (PD), ACE and Simpson 1-D indices as implemented in the R
package phyloseq (McMurdie and Holmes, 2013). We assessed variation in microbial
composition (alpha-diversity) and the mean proportions of the most abundant taxa (≥5% of all
sequences) using Linear Mixed Effects models (LME) with the lmer R package (Gałecki and
Burzykowski, 2013). Since we were interested in assessing whether microbial diversity varied

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
93
across fish age groups (predictor), we used age groups as a fixed factor and sampling date
(with 12 sampling time points) as a random factor. The final general LME formula was
expressed as: microbial diversity~fish age group + (1|sampling time point). Microbial structure
(beta-diversity) was estimated using phylogenetic Unifrac (unweighted and weighted) and
Bray-Curtis distances. Dissimilarity in microbial structure between samples was visualized
using principal coordinates analysis (PCoA). Additionally, differences in community structure
driven by fish age group were further tested using permutational multivariate analysis of
variance (PERMANOVA) as implemented in the adonis function of the vegan R package
(Oksanen et al., 2008). We used the strata argument to permutate sampling dates and ran
1000 permutations.
Previous fish studies of skin and gill microbiota (e.g., Legrand et al., 2018; Reinhart et
al., 2019; Wu et al., 2020; Zhang et al., 2018), including seabass and seabream (Rosado et
al., 2019a), have shown remarkable differences in microbial composition and structure across
host species and tissues. Although the two species studied here are farmed in the same
location, provided feeds were different throughout the sampling period. For this reason, and
because fish samples were processed using two different DNA extraction kits, we did not
compare mucosa microbiotas between fishes (e.g., Kennedy et al., 2014; Wilkes Walburn et
al., 2019). Additionally, a previous study by our group (Rosado et al., 2019b) showed that
disease and antibiotic treatment in seabass leads to asymmetrical shifts in skin and gill
microbial communities. Therefore, we carried out all our statistical analyses separately for
each fish species and tissue.
Microbial potential metabolic functions were predicted using the metagenomic
Phylogenetic Investigation of Communities by Reconstruction of Unobserved States software
(PICRUSt2) embedded in QIIME2 (Douglas et al., 2019), applying a weighted nearest
sequenced taxon index (NSTI) cutoff of 0.03. Predicted metagenomes were collapsed using
the Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway metadata (Kanehisa et al.,
2019). We identified differentially abundant metabolic pathways in the skin and gill microbiota
of seabass and seabream across age groups using linear discriminant analysis (LDA) in
LEfSe, using age groups as classes (Segata et al., 2011). As suggested by the authors, we
used a P-value cut-off of 0.05 and a LDA effect size cut-off of 2 (Segata et al., 2011).
Finally, to assess to what extent water microbial communities shaped skin and gill
microbiota across fish age groups, we estimated the number of shared ASVs between fish
and water microbiota and constructed Venn diagrams in R. We used PERMANOVA and
mantel testes (Mantel, 1967) to assess differences in community structure and correlations
between tissues and water microbiota, respectively, in both species.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
94
3.4 Results Skin, gill and water microbial samples from different fish age groups were collected
simultaneously (same day and at approximately the same time) from separate ponds (Figure
S3.1). Three age cohorts were sampled for the seabass, which included fish in their 1st, 2nd
and 3rd year of age; while two age cohorts were sampled for the seabream, which included
fish in their 2nd and 3rd year of age. Since our sampling was non-invasive, to classify fishes
into age groups we coupled available information from the literature (Barnabe, 1991; Chaoui
et al., 2006; Felip et al., 2001; Mehanna, 2007; Zohar and Gordin, 1979) with weight and age
of maturation estimates provided by the fish farm. The three seabass age cohorts were thus
classified as early juveniles, late juveniles and mature adults, respectively; while the two
seabream age cohorts were classified as juveniles and mature adults, respectively – see
Material and methods section for more details. Differences in the average weight estimated
for each age group at the beginning and end of our sampling showed a 245% growth for the
seabass early juvenile group, an 83% growth for the late juvenile group and a 43% growth for
the mature adult group. Similarly, a 143% growth was estimated for the seabream juveniles
and a 16% growth for the mature adults. Descriptive analyses were performed for each age
group separately and comparative statistical analyses were performed between age groups.
3.4.1 Microbial diversity across age groups
3.4.1.1 Alpha-diversity Microbial alpha-diversity was calculated using Shannon, Faith’s phylogenetic diversity
(PD), ACE and Simpson 1-D indices. In general, the skin microbiota showed higher mean
values for the alpha-diversity indices than the gill microbiota across all age groups in both fish
species, except for the Simpson 1-D index in the late juveniles and adults of the seabass
(Figure S3.2). In seabass, the skin and gill microbiota of late juveniles and mature adult fish
presented higher mean values of alpha-diversity than the microbiota of the early juveniles
(Figures 3.1A, S3.3). In seabream, skin and gill microbiotas showed similar alpha-diversity in
both cohorts (Figures 3.1B, S3.3). Linear Mixed Effects (LME) model analysis (diversity~age
group + (1|sampling date)) showed most alpha-diversity estimates varied significantly between
seabass age groups in both tissues. Pairwise comparisons between age groups in seabass
showed significant (p < 0.05) differences in alpha-diversity for almost all the tests comparing
early vs late juveniles and early juveniles vs mature adults (Table 3.1). Seabass late juvenile
vs mature adult alpha-diversity comparisons were never significant (p > 0.05, Table 3.1) for
both fish mucosae. In the seabream, only the Shannon and PD indices of the gill microbiota
varied significantly between juveniles and mature adults (p < 0.04, Table 3.1).
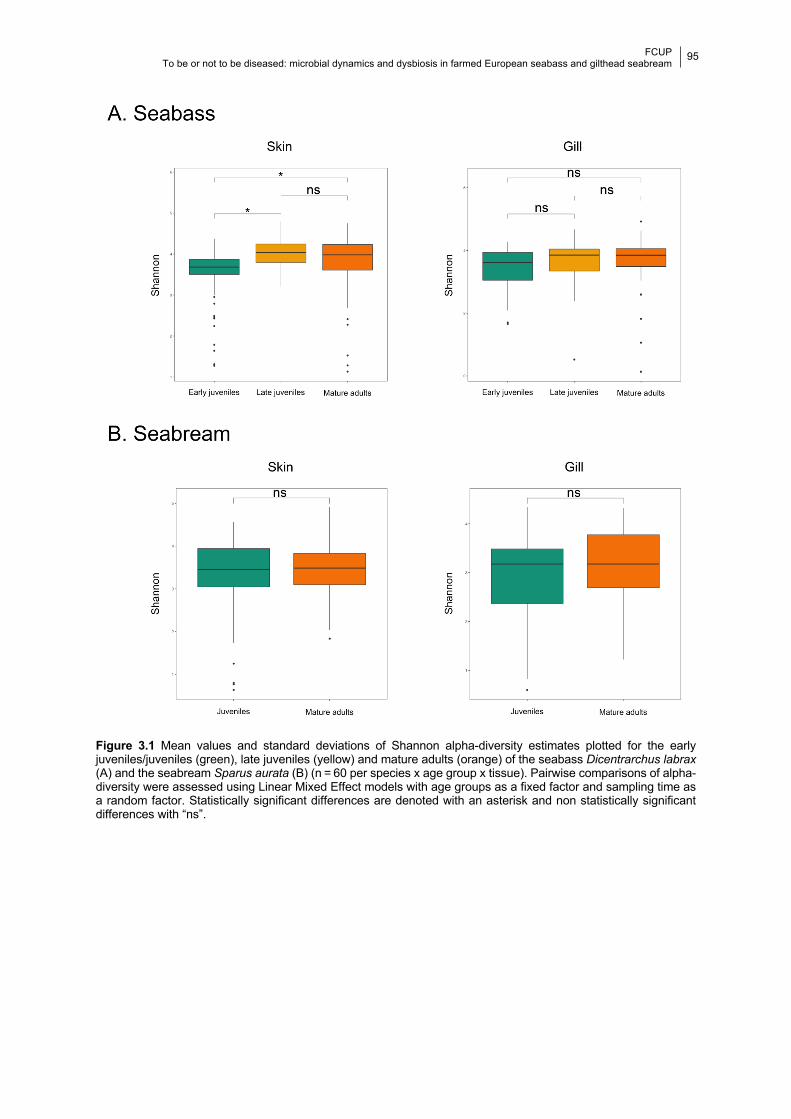
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
95
Figure 3.1 Mean values and standard deviations of Shannon alpha-diversity estimates plotted for the early juveniles/juveniles (green), late juveniles (yellow) and mature adults (orange) of the seabass Dicentrarchus labrax (A) and the seabream Sparus aurata (B) (n = 60 per species x age group x tissue). Pairwise comparisons of alpha-diversity were assessed using Linear Mixed Effect models with age groups as a fixed factor and sampling time as a random factor. Statistically significant differences are denoted with an asterisk and non statistically significant differences with “ns”.
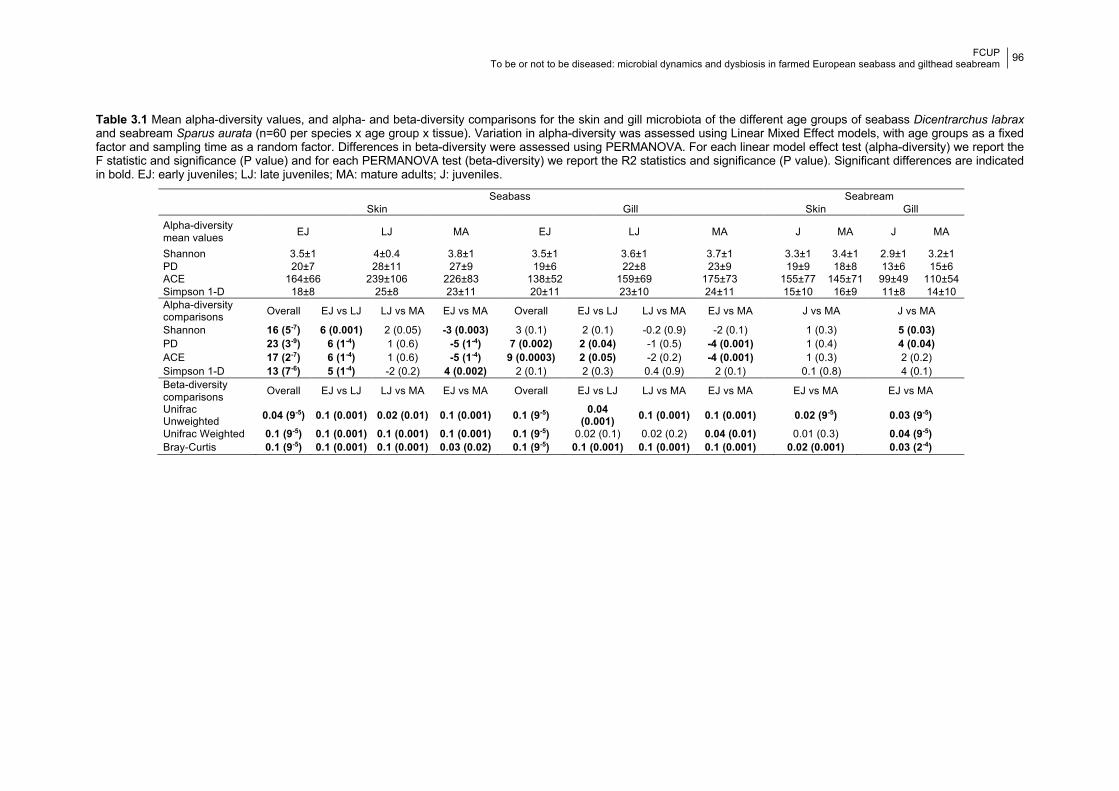
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
96
Table 3.1 Mean alpha-diversity values, and alpha- and beta-diversity comparisons for the skin and gill microbiota of the different age groups of seabass Dicentrarchus labrax and seabream Sparus aurata (n=60 per species x age group x tissue). Variation in alpha-diversity was assessed using Linear Mixed Effect models, with age groups as a fixed factor and sampling time as a random factor. Differences in beta-diversity were assessed using PERMANOVA. For each linear model effect test (alpha-diversity) we report the F statistic and significance (P value) and for each PERMANOVA test (beta-diversity) we report the R2 statistics and significance (P value). Significant differences are indicated in bold. EJ: early juveniles; LJ: late juveniles; MA: mature adults; J: juveniles.
Seabass Seabream Skin Gill Skin Gill Alpha-diversity mean values EJ LJ MA EJ LJ MA J MA J MA
Shannon 3.5±1 4±0.4 3.8±1 3.5±1 3.6±1 3.7±1 3.3±1 3.4±1 2.9±1 3.2±1 PD 20±7 28±11 27±9 19±6 22±8 23±9 19±9 18±8 13±6 15±6 ACE 164±66 239±106 226±83 138±52 159±69 175±73 155±77 145±71 99±49 110±54 Simpson 1-D 18±8 25±8 23±11 20±11 23±10 24±11 15±10 16±9 11±8 14±10 Alpha-diversity comparisons Overall EJ vs LJ LJ vs MA EJ vs MA Overall EJ vs LJ LJ vs MA EJ vs MA J vs MA J vs MA
Shannon 16 (5-7) 6 (0.001) 2 (0.05) -3 (0.003) 3 (0.1) 2 (0.1) -0.2 (0.9) -2 (0.1) 1 (0.3) 5 (0.03) PD 23 (3-9) 6 (1-4) 1 (0.6) -5 (1-4) 7 (0.002) 2 (0.04) -1 (0.5) -4 (0.001) 1 (0.4) 4 (0.04) ACE 17 (2-7) 6 (1-4) 1 (0.6) -5 (1-4) 9 (0.0003) 2 (0.05) -2 (0.2) -4 (0.001) 1 (0.3) 2 (0.2) Simpson 1-D 13 (7-6) 5 (1-4) -2 (0.2) 4 (0.002) 2 (0.1) 2 (0.3) 0.4 (0.9) 2 (0.1) 0.1 (0.8) 4 (0.1) Beta-diversity comparisons Overall EJ vs LJ LJ vs MA EJ vs MA Overall EJ vs LJ LJ vs MA EJ vs MA EJ vs MA EJ vs MA
Unifrac Unweighted 0.04 (9-5) 0.1 (0.001) 0.02 (0.01) 0.1 (0.001) 0.1 (9-5) 0.04
(0.001) 0.1 (0.001) 0.1 (0.001) 0.02 (9-5) 0.03 (9-5) Unifrac Weighted 0.1 (9-5) 0.1 (0.001) 0.1 (0.001) 0.1 (0.001) 0.1 (9-5) 0.02 (0.1) 0.02 (0.2) 0.04 (0.01) 0.01 (0.3) 0.04 (9-5) Bray-Curtis 0.1 (9-5) 0.1 (0.001) 0.1 (0.001) 0.03 (0.02) 0.1 (9-5) 0.1 (0.001) 0.1 (0.001) 0.1 (0.001) 0.02 (0.001) 0.03 (2-4)

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
97
3.4.1.2 Beta-diversity Microbial structure was estimated using phylogenetic UniFrac (unweighted and
weighted) and Bray-Curtis distances. The PERMANOVA analyses of dissimilarity
(diversity~age group, strata = sampling date) showed significant differences between age
groups in both species (p < 0.02, Table 3.1), except for the UniFrac Weighted distance
between the gills of early and late seabass juveniles (p = 0.1, Table 3.1), seabass late juveniles
and mature adults (p = 0.2, Table 3.1), and the skin of juveniles and seabream adults (p = 0.3,
Table 3.1). Moreover, high inter-individual variability within age groups was also observed.
However, differences in beta-diversity dispersion within age groups for the skin and gill
microbiotas of both species were small (Bray-Curtis distance, Figure 3.2).
3.4.1.3 Bacterial taxa Proteobacteria and Bacteroidetes were the most abundant (≥5%) phyla in the skin
(averaging 41 ± 4% and 39 ± 2% of the sequences in seabass and 55 ± 4% and 31 ± 4% in
seabream) and gill (averaging 52 ± 7% and 25 ± 5% in seabass and 69 ± 4% and 12 ± 1% in
seabream) microbiotas of all studied age groups (Table S3.1). The NS3a marine group and a
genus belonging to the Flavobacteriaceae family were the most abundant (≥5%) genera in the
skin (10 ± 1 and 11 ± 2, respectively) and gill (6 ± 1 for both) of all the age groups in seabass;
while Burkholderia-Caballeronia-Paraburkholderia was the most abundant genus in the skin
(17 ± 1) and gill (25 ± 0) of both age groups in seabream (Table S3.1). The most abundant
microbial phyla and genera found in both fish species varied between age groups and tissues
(Figure 3.3, Table S3.1). LME models showed that the relative abundance of all those phyla
was significantly different between age groups, except in the gill microbiota of the seabream,
where the relative abundance of Cyanobacteria did not vary (Table S3.2). LME analyses also
revealed that 100 and 63% of the genera varied between age groups in the skin and gill of the
seabass, respectively, while 40 and 50% varied in the skin and gill of the seabream,
respectively (Table S3.2). Pairwise comparisons of taxa across age groups in seabass yielded
a higher percentage of significant differences between early juveniles and mature adults in
both tissues (100% in the skin and 38% in the gill) than between early and late juveniles (67%
in the skin and 13% in the gill), or between late juveniles and mature adults (0% in the skin
and 25% in the gill) (Table S3.2).
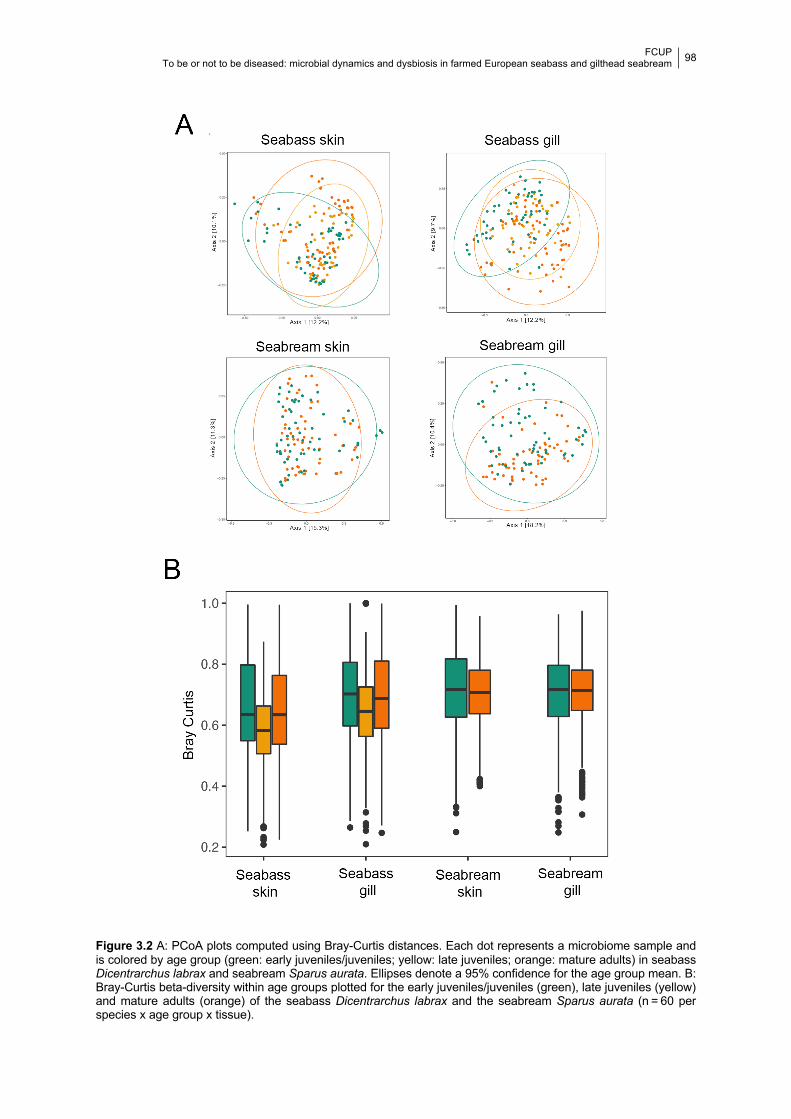
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
98
Figure 3.2 A: PCoA plots computed using Bray-Curtis distances. Each dot represents a microbiome sample and
is colored by age group (green: early juveniles/juveniles; yellow: late juveniles; orange: mature adults) in seabass
Dicentrarchus labrax and seabream Sparus aurata. Ellipses denote a 95% confidence for the age group mean. B:
Bray-Curtis beta-diversity within age groups plotted for the early juveniles/juveniles (green), late juveniles (yellow)
and mature adults (orange) of the seabass Dicentrarchus labrax and the seabream Sparus aurata (n = 60 per
species x age group x tissue).
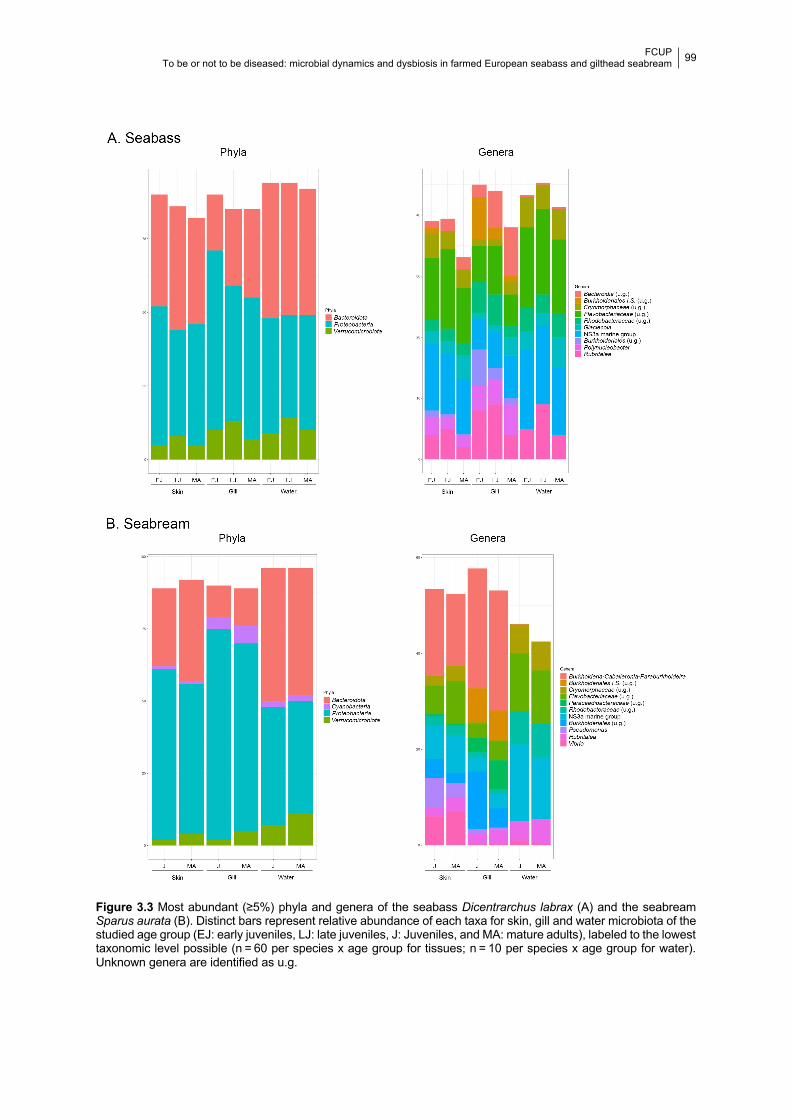
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
99
Figure 3.3 Most abundant (≥5%) phyla and genera of the seabass Dicentrarchus labrax (A) and the seabream
Sparus aurata (B). Distinct bars represent relative abundance of each taxa for skin, gill and water microbiota of the
studied age group (EJ: early juveniles, LJ: late juveniles, J: Juveniles, and MA: mature adults), labeled to the lowest
taxonomic level possible (n = 60 per species x age group for tissues; n = 10 per species x age group for water).
Unknown genera are identified as u.g.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
100
3.4.2 Microbial predicted functional diversity across age groups About 462 ± 18 KEGG pathways were inferred in the skin and gill microbiota of the
seabass, while 455 ± 4 pathways were inferred in the skin and gill microbiota of the seabream.
Linear discriminant analysis of the metagenomic predictions performed in LEfSe showed that
different pathways were significantly enriched for each age group in both species (Figure 3.4,
Table S3.3). Overall, there were more enriched pathways in the older age groups (seabass
late juveniles and mature adults of both species) than in the younger age groups of both
species (Figure 3.4, Table S3.3).
While there were no significantly enriched pathways in the skin microbiota of early
juvenile seabass, enriched pathways in the gill microbiota of this age group were related to
metabolic regulator biosynthesis, purine nucleotide degradation, sugar degradation and
fermentation of pyruvate. In the skin microbiota of late juvenile seabass, enriched pathways
were related to thiamine biosynthesis, aldehyde degradation and L-arabinose degradation;
while in the gill microbiota they were related to denitrification, galactose degradation and
nitrogen compound metabolism. In mature seabass, pyrimidine and purine
deoxyribonucleotide de novo biosynthesis were enriched in both tissues. Additionally, the gill
microbiota was also enriched by pathways related to the biosynthesis of chlorophyll, folate,
hemiterpene, L-alanine, L-tyrosine, NAD, secondary metabolite and ubiquinol, chloroaromatic
compound degradation, fermentation to lactate and glycolysis (Figure 3.4, Table S3.3).
In the skin microbiota of seabream juveniles, enriched pathways were related to amine
and polyamine biosynthesis and degradation, choline biosynthesis, and sugar acid and
toluene degradation; whereas in the gill microbiota only pyrimidine and purine
deoxyribonucleotide de novo biosynthesis were identified. The enriched pathways of the
seabream mature adults were related to fatty acid, L-methionine, NAD, palmitate,
palmitoleate, siderophore, stearate and unsaturated fatty acid biosynthesis, pyrimidine and
purine nucleotide salvage, aspartate superpathway and TCA cycle in the skin microbiota;
whereas pyrimidine and purine deoxyribonucleotide de novo biosynthesis, autotrophic CO2
fixation and fermentation of pyruvate were enriched in the gill microbiota (Figure 3.4, Table
S3.3).
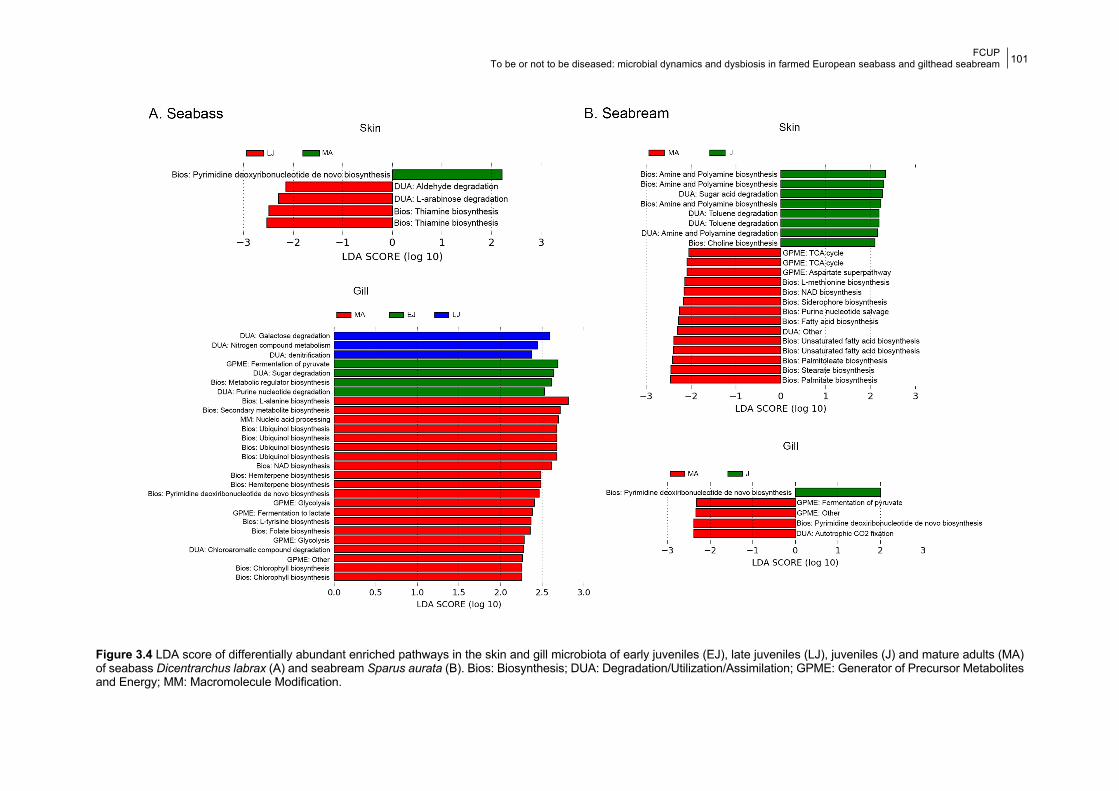
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
101
Figure 3.4 LDA score of differentially abundant enriched pathways in the skin and gill microbiota of early juveniles (EJ), late juveniles (LJ), juveniles (J) and mature adults (MA) of seabass Dicentrarchus labrax (A) and seabream Sparus aurata (B). Bios: Biosynthesis; DUA: Degradation/Utilization/Assimilation; GPME: Generator of Precursor Metabolites and Energy; MM: Macromolecule Modification.
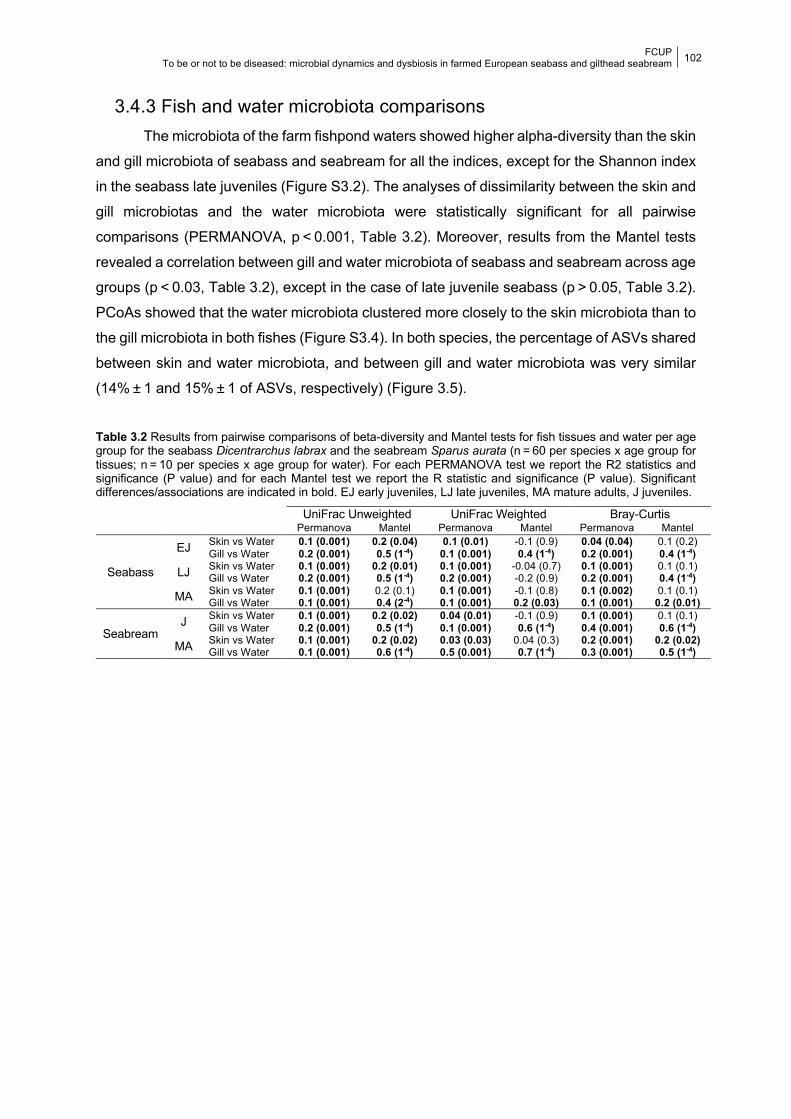
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
102
3.4.3 Fish and water microbiota comparisons The microbiota of the farm fishpond waters showed higher alpha-diversity than the skin
and gill microbiota of seabass and seabream for all the indices, except for the Shannon index
in the seabass late juveniles (Figure S3.2). The analyses of dissimilarity between the skin and
gill microbiotas and the water microbiota were statistically significant for all pairwise
comparisons (PERMANOVA, p < 0.001, Table 3.2). Moreover, results from the Mantel tests
revealed a correlation between gill and water microbiota of seabass and seabream across age
groups (p < 0.03, Table 3.2), except in the case of late juvenile seabass (p > 0.05, Table 3.2).
PCoAs showed that the water microbiota clustered more closely to the skin microbiota than to
the gill microbiota in both fishes (Figure S3.4). In both species, the percentage of ASVs shared
between skin and water microbiota, and between gill and water microbiota was very similar
(14% ± 1 and 15% ± 1 of ASVs, respectively) (Figure 3.5).
Table 3.2 Results from pairwise comparisons of beta-diversity and Mantel tests for fish tissues and water per age group for the seabass Dicentrarchus labrax and the seabream Sparus aurata (n = 60 per species x age group for tissues; n = 10 per species x age group for water). For each PERMANOVA test we report the R2 statistics and significance (P value) and for each Mantel test we report the R statistic and significance (P value). Significant differences/associations are indicated in bold. EJ early juveniles, LJ late juveniles, MA mature adults, J juveniles.
UniFrac Unweighted UniFrac Weighted Bray-Curtis Permanova Mantel Permanova Mantel Permanova Mantel
Seabass
EJ Skin vs Water 0.1 (0.001) 0.2 (0.04) 0.1 (0.01) -0.1 (0.9) 0.04 (0.04) 0.1 (0.2) Gill vs Water 0.2 (0.001) 0.5 (1-4) 0.1 (0.001) 0.4 (1-4) 0.2 (0.001) 0.4 (1-4)
LJ Skin vs Water 0.1 (0.001) 0.2 (0.01) 0.1 (0.001) -0.04 (0.7) 0.1 (0.001) 0.1 (0.1) Gill vs Water 0.2 (0.001) 0.5 (1-4) 0.2 (0.001) -0.2 (0.9) 0.2 (0.001) 0.4 (1-4)
MA Skin vs Water 0.1 (0.001) 0.2 (0.1) 0.1 (0.001) -0.1 (0.8) 0.1 (0.002) 0.1 (0.1) Gill vs Water 0.1 (0.001) 0.4 (2-4) 0.1 (0.001) 0.2 (0.03) 0.1 (0.001) 0.2 (0.01)
Seabream J Skin vs Water 0.1 (0.001) 0.2 (0.02) 0.04 (0.01) -0.1 (0.9) 0.1 (0.001) 0.1 (0.1)
Gill vs Water 0.2 (0.001) 0.5 (1-4) 0.1 (0.001) 0.6 (1-4) 0.4 (0.001) 0.6 (1-4) MA Skin vs Water 0.1 (0.001) 0.2 (0.02) 0.03 (0.03) 0.04 (0.3) 0.2 (0.001) 0.2 (0.02)
Gill vs Water 0.1 (0.001) 0.6 (1-4) 0.5 (0.001) 0.7 (1-4) 0.3 (0.001) 0.5 (1-4)

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
103
Figure 3.5 Venn diagrams showing the number and percentage of shared ASVs between skin (yellow), gill (pink) and water (blue) microbiota of the different age groups for the seabass Dicentrarchus labrax (A) and the seabream Sparus aurata (B) (n = 60 per species x age group for tissues; n = 10 per species x age group for water).
3.5 Discussion We characterized the skin and gill microbiota of different age groups of farmed
European seabass and gilthead seabream using 16S rRNA amplicon high-throughput
sequencing. By taking into account potential environmental and seasonal effects, our study
shows that fish age, in particular sexual maturation and growth, impact skin and gill microbial
diversity (Table 3.1; Figure 3.1), composition (Table S3.1; Figures 3.3, 3.5) and predicted
microbial functions (Table S3.3; Figure 3.4).
3.5.1 Microbial diversity across age groups Fish growth and sexual maturation are usually accompanied by extreme
morphological and physiological changes (e.g., Varsamos et al., 2005; Wilkins and Jancsar,
1979). Importantly, some of these changes reported for the skin and gills have been suggested
to also affect their microbiota. For example, changes in epidermal structure derived from
sexual maturation (e.g., increases in the number, size and activity of the mucous cells) have

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
104
been detected in several fish species (e.g., Whitear, 2009; Wilkins and Jancsar, 1979), and
suggested to increase infection rate with Saprolegnia fungus in sea trout and brown trout
(Richards and Pickering, 1978). Likewise, changes in the hormones expressed in the skin alter
the biochemistry of the skin mucous and also potentially affect its microbiota (Roberts and
Bullock, 1980). Fish growth and sexual maturation also impact gill morphology and function in
some fish species. For example, the ability to osmoregulate at different salinities was seen to
increase throughout the developmental stages (larva to juvenile) of seabass (Varsamos et al.,
2005). Additionally, body size was also identified as the main factor affecting morphological
variation in gill rakes and gill pore size in the Silver Carp and Gizzard Shad, suggesting that
the overall filtering ability of these species is related to size and maturation (Walleser et al.,
2014). Importantly, a recent study showed that body weight increase is accompanied by higher
microbial community structuring in the skin and gill of rabbit fish (Wu et al., 2020). We thus
hypothesize that such physiological and morphological changes occurring during fish growth
have led to the changes in microbial diversity, composition and predicted functionality
observed in the present study.
The seabass skin and gill microbiotas of older age groups showed significantly higher
alpha-diversity than those of early juveniles. Additionally, a higher percentage of significant
differences in the relative abundance of the most abundant phyla and genera occurred
between early juveniles and older age groups (67 ± 27% and 55 ± 38% in the skin and gill,
respectively; Table S3.2). This suggests that the skin and gill microbiotas of the seabass were
highly dynamic, diversifying with age. Conversely, the skin and gill microbiotas of the
seabream juveniles and adults showed similar alpha-diversity means, although a high
percentage of the most abundant phyla and genera varied between age groups (70 ± 42%
and 63 ± 18% in the skin and gill, respectively; Table S3.2). Variation in microbiota alpha-
diversity between different age groups has been previously reported for many fish species.
For example, studies on the zebrafish and salmon gut microbiota, have reported differences
between mature and immature life stages; however, those differences also coincide with other
major ecological changes in the fish, such as diet (Stephens et al., 2016) or environment
transitions (Lokesh et al., 2019). Moreover, the relative abundance of predominant bacterial
groups also changed with aging in other fish species (e.g., Lokesh et al., 2019; Pratte et al.,
2018; Stephens et al., 2016).
We detected significant differences in microbial structure across all age groups in both
species. Similar results have been also reported in other fish (e.g., several reef fish species,
Pratte et al., 2018; Xavier et al., 2020, and Salmo salar; Llewellyn et al., 2016), being
particularly evident in longitudinal studies encompassing several months (Lokesh et al., 2019;
Stephens et al., 2016; Yan et al., 2016). High inter-individual variability within age groups was
also previously reported for other fish species (e.g., Pratte et al., 2018; Thompson et al., 2017;

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
105
Uren Webster et al., 2018). However, our results showed that fish age only explained a low
percentage of the variation in the bacterial community structure (R2 < 0.1). This suggests that
microbial differences between age groups are small at the community level, but clearly
noticeable at the species level, with a high proportion of the predominant bacterial taxa (61 ±
39% and 46 ± 32% in the skin and gill of seabass, respectively; 70 ± 42% and 63 ± 18% in the
skin and gill of the seabream, respectively) changing their abundance with sexual maturation.
Although our statistical models accounted for sampling dates as a random factor, other biotic
and abiotic factors (e.g., variation in the environment and individual weights) could be
responsible for most of the variation observed in community structure (e.g., Llewellyn et al.,
2014; Lokesh et al., 2019; Uren Webster et al., 2018).
3.5.2 Microbial predicted functional diversity across age groups The predicted functional analysis suggests that distinct significantly enriched metabolic
pathways are expressed in skin and gill microbiotas of both fish species across age groups.
Although metabolic information is particularly limited for fish microbiotas, studies on other
vertebrates, mainly in humans and their gut microbiotas, are starting to shed light on the
beneficial outcomes specific microbial metabolic functions have on the host health and
physiology (e.g., Berry and Loy, 2018).
Notwithstanding that present results should be interpreted with caution since
PICRUSt2 analysis is limited by the currently available genomes and biased towards human
health microorganisms (Choi et al., 2017), one could suggest that some of the enriched
metabolic pathways found in our analysis could also improve the seabass and seabream
health and physiology. The protective role of the microbiota is often related to the production
of secondary metabolites that provide chemical defense and mediate bacterial diversity (Kelly
and Salinas, 2017). Secondary metabolites with antimicrobial activity have been previously
isolated from microbial species inhabiting the gut microbiome of fish (Jami et al., 2015). Here,
the biosynthesis of secondary metabolites that have been associated with antimicrobial
activity, including hemiterpene (e.g., Reyes-Jurado et al., 2015), were enriched in mature
adults of both species. Additionally, the biosynthesis of chlorophyll and several amino acids,
herein enriched in older age groups of both fish species, have also been found to be expressed
in the skin and gut of healthy humans (e.g., Barnard et al., 2016; Shao et al., 2017).
Importantly, amino acid biosynthesis was reported in the gut microbiota of grass carp when
fed a protein-deficient diet, suggesting a metabolic role of the gut microbiota towards fish
nutrition (Ni et al., 2014). The biosynthesis of vitamins, here enriched in older age groups of
seabass, has been found beneficial for human skin (e.g., Saxena et al., 2018) and gut mucosa,
including folate and thiamine (Hill, 1997). In addition, polyamines are bacterial metabolites

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
106
known to have several benefits towards gut mucosa recovery (Tofalo et al., 2019). These
pathways were also enriched in the juveniles of seabream.
It is also worth noticing that some of the enriched metabolic pathways detected in the
present study could be driven by the high environmental variability of the Alvor estuary where
these fish were reared. In estuaries, salinity variations occur on a daily basis due to tides and
pollutants can be prevalent (e.g., Antunes et al., 2007). Biosynthesis of fatty acids and
unsaturated fatty acids were two of the predicted metabolic pathways enriched in the
microbiota of mature seabream. These same pathways have also been enriched in previous
analyses of the skin and gut microbiota in the Atlantic salmon (Dehler et al., 2017; Lokesh and
Kiron, 2016) and in the skin microbiota of the common snook (Tarnecki et al., 2019) when
transitioning between freshwater and seawater. Additionally, two of the predicted metabolic
pathways identified in both fish species were related to degradation of toxic compounds.
Specifically, biodegradation of the highly prevalent toxic pollutants toluene and chloroaromatic
compounds by bacteria is essential to remove them from the environment and to prevent
absorption through the skin and gills in aquatic animals (Fuchs et al., 2011; Patrolecco et al.,
2010; Van Der Meer, 1997).
Following alpha-diversity patterns, fish from older age groups, particularly in the
seabass, had greater enrichment of predicted functions related mainly to the biosynthesis and
degradation of compounds; as well as, to a lesser extent, metabolism and energy cycles. We
then hypothesize that the increase in microbial diversity observed as fish ages leads to wider
functional diversity. This could prove beneficial to those fishes, given the key physiological
modifications older fish groups are experiencing during sexual maturation and growth.
3.5.3 Fish and water microbiota comparisons The water microbiotas of fishponds were significantly distinct and more diverse than
the skin and gill microbiota of both fish species, regardless of their age. It is known that free-
living microbial communities retain higher richness than host-associated communities
(Thompson et al., 2017), with many studies showing a higher bacterial diversity in water
relative to fish skin (Chiarello et al., 2019, 2018, 2015; Larsen et al., 2015; Uren Webster et
al., 2018; Wu et al., 2020), gills (Pratte et al., 2018; Wu et al., 2020), gut (Bledsoe et al., 2016;
Parris et al., 2016; Reinhart et al., 2019; Wilkes Walburn et al., 2019; Zhang et al., 2018),
stomach (Wu et al., 2020), hindgut (Wu et al., 2020) and whole larvae (Nikouli et al., 2019).
Although some studies in fish have shown that the microbial communities found in the water
tend to be recovered in the larval gut microbiota (Stephens et al., 2016; Wilkes Walburn et al.,
2019), others have also shown that water microbiota does not influence directly the microbiota
of the fish mucosa (Bledsoe et al., 2016; Boutin et al., 2013; Carlson et al., 2017; Chiarello et

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
107
al., 2019, 2018, 2015; Larsen et al., 2015; Legrand et al., 2018; Llewellyn et al., 2016; Nikouli
et al., 2019; Parris et al., 2016; Pratte et al., 2018; Schmidt et al., 2018; Uren Webster et al.,
2018; Wu et al., 2020; Yan et al., 2016; Zhang et al., 2018). Importantly, a previous study of
the skin microbiotas of seabass and seabream also showed significant differences with
planktonic communities (Chiarello et al., 2015). However, in that study only a low number of
Operational Taxonomic Units (3%) was shared between skin and water microbiota; whereas
in the present study higher percentages of ASVs were shared between the skin (14% ± 1) and
the gill (15% ± 1) of both fish species and the surrounding water.
Microbial dissimilarities depicted by PCoAs showed that, although significantly
different, the skin microbiota of both species clustered more closely to the water microbiota
than the gill microbiota. However, only a small percentage of the variation (PC 1 – average
18% ± 2; PC 2 – average 10% ± 1) was explained by this analysis. On the other hand, the
results from the Mantel tests showed a significant (p < 0.03) correlation between the water
and gill microbiotas, but not between the water and skin microbiotas. This suggests that
although both skin and gill are permanently in contact with water, the gill environment may be
more susceptible to variations in the water microbiota.
3.6 Conclusion Skin and gill are important mucosal barriers that protect the fish from the external
environment. They are in permanent contact with the water column and thus prone to
pathogenic bacterioplankton colonization. However, most studies so far investigating microbial
changes related to fish age have either strictly focused on early life stages (i.e., larvae
development) or on the gut microbiota. In the present study, we demonstrate that, to some
extent, changes occurring later in life can also be correlated with aging factors such as growth
and sexual maturation. We uncovered important differences in the diversity, composition and
predicted functionality of the skin and gill microbiotas across age groups of farmed seabass
and seabream. Fish included in this study were exposed to a heterogeneous estuarine
environment, that varies seasonally as well as daily (e.g., salinity and temperature
fluctuations). Hence, although we observed significant differences in microbial community
structure due to age, other biotic and abiotic factors not considered here may have more
deeply structured the fish microbiota. Growth rate decreased drastically with age, being much
higher in juveniles (243 and 83% for early and late seabass juveniles, and 143% in seabream)
relative to adults (43 and 16% in adult seabass and seabream, respectively). We, thus,
conclude that growth and sexual maturation are likely the main drivers of the microbial
differences attributed to age observed here. Overall, our results agree well with other studies
of the skin (Wu et al., 2020; Xavier et al., 2020) and gill (Pratte et al., 2018; Wu et al., 2020)

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
108
microbiotas of several wild reef fish, suggesting this could be a general pattern across fish.
Our results also highlight the importance of aging in farm fish studies focused on microbial
dysbiosis and disease dynamics.
3.7 Acknowledgements This work was funded by the European Regional Development Fund (ERDF) through the
COMPETE program and by National Funds through FCT - Foundation for Science and
Technology (project PTDC/ MAR-BIO/0902/2014 -POCI-01-0145-FEDER-016550; project
PTDC/BIA-MIC/27995/2017 POCI-01-0145- FEDER-027995); DR, AP, MP-L and RX were
supported by FCT under the Programa Operacional Potencial Humano – Quadro de
Referência Estratégico Nacional funds from the European Social Fund and Portuguese
Ministério da Educação e Ciência (DR doctoral grant SFRH/ BD/117943/2016; AP doctoral
Grant SFRH/BD/144928/2019; MPL: IF/00764/2013; RX: IF/00359/2015).
3.8 References Amato, K.R., Leigh, S.R., Kent, A., Mackie, R.I., Yeoman, C.J., Stumpf, R.M., Wilson, B.A.,
Nelson, K.E., White, B.A., Garber, P.A., 2014. The role of gut microbes in satisfying the
nutritional demands of adult and juvenile wild, black howler monkeys (Alouatta pigra).
Am. J. Phys. Anthropol. 155, 652–664. https://doi.org/10.1002/ajpa.22621
Antunes, P., Gil, O., Ferreira, M., Vale, C., Reis-Henriques, M.A., 2007. Depuration of PCBs
and DDTs in mullet under captivity clean conditions. Chemosphere 67, S58–S64.
https://doi.org/10.1016/j.chemosphere.2006.05.091
Araki, H., Schmid, C., 2010. Is hatchery stocking a help or harm?. Evidence, limitations and
future directions in ecological and genetic surveys. Aquaculture.
https://doi.org/10.1016/j.aquaculture.2010.05.036
Barnabe, G., 1991. La cría de lubina y de dorada. Acuicultura 573–612.
Barnard, E., Shi, B., Kang, D., Craft, N., Li, H., 2016. The balance of metagenomic elements
shapes the skin microbiome in acne and health. Sci. Rep. 6, 1–12.
https://doi.org/10.1038/srep39491
Berry, D., Loy, A., 2018. Stable-Isotope Probing of Human and Animal Microbiome Function.
Trends Microbiol. https://doi.org/10.1016/j.tim.2018.06.004
Bledsoe, J.W., Peterson, B.C., Swanson, K.S., Small, B.C., 2016. Ontogenetic
Characterization of the Intestinal Microbiota of Channel Catfish through 16S rRNA Gene
Sequencing Reveals Insights on Temporal Shifts and the Influence of Environmental
Microbes. PLoS One 11, e0166379. https://doi.org/10.1371/journal.pone.0166379
Boutin, S., Bernatchez, L., Audet, C., Derôme, N., 2013. Network Analysis Highlights Complex

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
109
Interactions between Pathogen, Host and Commensal Microbiota. PLoS One 8, e84772.
https://doi.org/10.1371/journal.pone.0084772
Callahan, B.J., McMurdie, P.J., Rosen, M.J., Han, A.W., Johnson, A.J.A., Holmes, S.P., 2016.
DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13,
581–583. https://doi.org/10.1038/nmeth.3869
Carillo, M., Zanuy, S., Prat, F., Cerdá, J., Ramos, J., Mañanós, E., Bromage, N, 1995.
Seabass (Dicentrarchus labrax), in: Bromage, NR, Roberts, R. (Eds.), Broodstock
Management and Egg and Larval Quality. Blackwell Science, Oxford, pp. 138–68.
Carlson, J.M., Leonard, A.B., Hyde, E.R., Petrosino, J.F., Primm, T.P., 2017. Microbiome
disruption and recovery in the fish Gambusia affinis following exposure to broad-
spectrum antibiotic. Infect. Drug Resist. 10, 143–154.
https://doi.org/10.2147/IDR.S129055
Chaoui, L., Hichem Kara, M., Faure, E., Quignard, J.P., 2006. Growth and reproduction of the
gilthead seabream Sparus aurata in Mellah lagoon (north-eastern Algeria). Sci. Mar. 70,
545–552. https://doi.org/10.3989/scimar.2006.70n3545
Chiarello, M., Auguet, J.C., Bettarel, Y., Bouvier, C., Claverie, T., Graham, N.A.J.,
Rieuvilleneuve, F., Sucré, E., Bouvier, T., Villéger, S., 2018. Skin microbiome of coral
reef fish is highly variable and driven by host phylogeny and diet. Microbiome 6, 147.
https://doi.org/10.1186/s40168-018-0530-4
Chiarello, M., Paz-Vinas, I., Veyssière, C., Santoul, F., Loot, G., Ferriol, J., Boulêtreau, S.,
2019. Environmental conditions and neutral processes shape the skin microbiome of
European catfish (Silurus glanis) populations of Southwestern France. Environ. Microbiol.
Rep. 11, 605–614. https://doi.org/10.1111/1758-2229.12774
Chiarello, M., Villéger, S., Bouvier, C., Bettarel, Y., Bouvier, T., 2015. High diversity of skin-
associated bacterial communities of marine fishes is promoted by their high variability
among body parts, individuals and species. FEMS Microbiol. Ecol. 91.
https://doi.org/10.1093/femsec/fiv061
Choi, J., Yang, F., Stepanauskas, R., Cardenas, E., Garoutte, A., Williams, R., Flater, J.,
Tiedje, J.M., Hofmockel, K.S., Gelder, B., Howe, A., 2017. Strategies to improve
reference databases for soil microbiomes. ISME J. 11, 829–834.
https://doi.org/10.1038/ismej.2016.168
Collado, M.C., Cernada, M., Neu, J., Pérez-Martínez, G., Gormaz, M., Vento, M., 2015.
Factors influencing gastrointestinal tract and microbiota immune interaction in preterm
infants. Pediatr. Res. https://doi.org/10.1038/pr.2015.54
Dash, S., Das, S.K., Samal, J., Thatoi, H.N., 2018. Epidermal mucus, a major determinant in
fish health: A review. Iran. J. Vet. Res. https://doi.org/10.22099/ijvr.2018.4849
Dehler, C.E., Secombes, C.J., Martin, S.A.M., 2017. Seawater transfer alters the intestinal

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
110
microbiota profiles of Atlantic salmon (Salmo salar L.). Sci. Rep. 7, 1–11.
https://doi.org/10.1038/s41598-017-13249-8
Douglas, G.M., Maffei, V.J., Zaneveld, J., Yurgel, S.N., Brown, J.R., Taylor, C.M.,
Huttenhower, C., Langille, M.G.I., 2019. PICRUSt2: An improved and extensible
approach for metagenome inference. bioRxiv. https://doi.org/10.1101/672295
FAO, 2018. The state of world fisheries and aquaculture 2018 - Meeting the sustainable
development goals. Rome.
Felip, A., Piferrer, F., Zanuy, S., Carrillo, M., 2001. Comparative growth performance of diploid
and triploid European sea bass over the first four spawning seasons. J. Fish Biol. 58, 76–
88. https://doi.org/10.1111/j.1095-8649.2001.tb00500.x
Frese, S.A., Parker, K., Calvert, C.C., Mills, D.A., 2015. Diet shapes the gut microbiome of
pigs during nursing and weaning. Microbiome 3, 28. https://doi.org/10.1186/s40168-015-
0091-8
Fuchs, G., Boll, M., Heider, J., 2011. Microbial degradation of aromatic compounds- From one
strategy to four. Nat. Rev. Microbiol. https://doi.org/10.1038/nrmicro2652
Gałecki, A., Burzykowski, T., 2013. Linear Mixed-Effects Models Using R: a step-by-step
approach, Springer Texts in Statistics. Springer New York, New York, NY.
https://doi.org/10.1007/978-1-4614-3900-4
Hill, M.J., 1997. Intestinal flora and endogenous vitamin synthesis. Eur. J. Cancer Prev. 6,
S43–S45. https://doi.org/10.1097/00008469-199703001-00009
Jami, M., Ghanbari, M., Kneifel, W., Domig, K.J., 2015. Phylogenetic diversity and biological
activity of culturable Actinobacteria isolated from freshwater fish gut microbiota.
Microbiol. Res. 175, 6–15. https://doi.org/10.1016/j.micres.2015.01.009
Jin Song, S., Woodhams, D.C., Martino, C., Allaband, C., Mu, A., Javorschi-Miller-
Montgomery, S., Suchodolski, J.S., Knight, R., 2019. Engineering the microbiome for
animal health and conservation. Exp. Biol. Med. 244, 494–504.
https://doi.org/10.1177/1535370219830075
Kanehisa, M., Sato, Y., Furumichi, M., Morishima, K., Tanabe, M., 2019. New approach for
understanding genome variations in KEGG. Nucleic Acids Res. 47, D590–D595.
https://doi.org/10.1093/nar/gky962
Kelly, C., Salinas, I., 2017. Under pressure: Interactions between commensal microbiota and
the teleost immune system. Front. Immunol. https://doi.org/10.3389/fimmu.2017.00559
Kennedy, N.A., Walker, A.W., Berry, S.H., Duncan, S.H., Farquarson, F.M., Louis, P.,
Thomson, J.M., Satsangi, J., Flint, H.J., Parkhill, J., Lees, C.W., Hold, G.L., 2014. The
Impact of Different DNA Extraction Kits and Laboratories upon the Assessment of Human
Gut Microbiota Composition by 16S rRNA Gene Sequencing. PLoS One 9, e88982.
https://doi.org/10.1371/journal.pone.0088982

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
111
Kozich, J.J., Westcott, S.L., Baxter, N.T., Highlander, S.K., Schloss, P.D., 2013. Development
of a dual-index sequencing strategy and curation pipeline for analyzing amplicon
sequence data on the miseq illumina sequencing platform. Appl. Environ. Microbiol. 79,
5112–5120. https://doi.org/10.1128/AEM.01043-13
Larsen, A.M., Bullard, S.A., Womble, M., Arias, C.R., 2015. Community Structure of Skin
Microbiome of Gulf Killifish, Fundulus grandis, Is Driven by Seasonality and Not Exposure
to Oiled Sediments in a Louisiana Salt Marsh. Microb. Ecol. 70, 534–544.
https://doi.org/10.1007/s00248-015-0578-7
Legrand, T.P.R.A., Catalano, S.R., Wos-Oxley, M.L., Stephens, F., Landos, M., Bansemer,
M.S., Stone, D.A.J., Qin, J.G., Oxley, A.P.A., 2018. The Inner Workings of the Outer
Surface: Skin and Gill Microbiota as Indicators of Changing Gut Health in Yellowtail
Kingfish. Front. Microbiol. 8, 2664. https://doi.org/10.3389/fmicb.2017.02664
Llewellyn, M.S., Boutin, S., Hoseinifar, S.H., Derome, N., 2014. Teleost microbiomes: The
state of the art in their characterization, manipulation and importance in aquaculture and
fisheries. Front. Microbiol. https://doi.org/10.3389/fmicb.2014.00207
Llewellyn, M.S., McGinnity, P., Dionne, M., Letourneau, J., Thonier, F., Carvalho, G.R., Creer,
S., Derome, N., 2016. The biogeography of the atlantic salmon (Salmo salar) gut
microbiome. ISME J. 10, 1280–1284. https://doi.org/10.1038/ismej.2015.189
Lokesh, J., Kiron, V., 2016. Transition from freshwater to seawater reshapes the skin-
associated microbiota of Atlantic salmon. Sci. Rep. 6, 1–10.
https://doi.org/10.1038/srep19707
Lokesh, J., Kiron, V., Sipkema, D., Fernandes, J.M.O., Moum, T., 2019. Succession of
embryonic and the intestinal bacterial communities of Atlantic salmon (Salmo salar)
reveals stage-specific microbial signatures. Microbiologyopen 8, e00672.
https://doi.org/10.1002/mbo3.672
Mantel, N., 1967. The Detection of Disease Clustering and a Generalized Regression
Approach. Cancer Res. 27.
McMurdie, P.J., Holmes, S., 2014. Waste Not, Want Not: Why Rarefying Microbiome Data Is
Inadmissible. PLoS Comput. Biol. 10, e1003531.
https://doi.org/10.1371/journal.pcbi.1003531
McMurdie, P.J., Holmes, S., 2013. phyloseq: An R Package for Reproducible Interactive
Analysis and Graphics of Microbiome Census Data. PLoS One 8, e61217.
https://doi.org/10.1371/journal.pone.0061217
Mehanna, S.F., 2007. A Preliminary Assessment and Management of Gilthead Bream Sparus
aurata in the Port Said Fishery, the Southeastern Mediterranean, Egypt, Turkish Journal
of Fisheries and Aquatic Sciences.
Mueller, N.T., Bakacs, E., Combellick, J., Grigoryan, Z., Dominguez-Bello, M.G., 2015. The

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
112
infant microbiome development: Mom matters. Trends Mol. Med.
https://doi.org/10.1016/j.molmed.2014.12.002
Murphy, J.F., 2018. The human microbiome: an emerging paradigm for better health.
MedCrave online J. Immunol. 6, 54–55. https://doi.org/10.15406/moji.2018.06.00192
Nagpal, R., Kurakawa, T., Tsuji, H., Takahashi, T., Kawashima, K., Nagata, S., Nomoto, K.,
Yamashiro, Y., 2017. Evolution of gut Bifidobacterium population in healthy Japanese
infants over the first three years of life: A quantitative assessment. Sci. Rep. 7, 1–11.
https://doi.org/10.1038/s41598-017-10711-5
Ni, J., Yan, Q., Yu, Y., Zhang, T., 2014. Factors influencing the grass carp gut microbiome
and its effect on metabolism. FEMS Microbiol. Ecol. 87, 704–714.
https://doi.org/10.1111/1574-6941.12256
Nikouli, Meziti, Antonopoulou, Mente, Kormas, 2019. Host-Associated Bacterial Succession
during the Early Embryonic Stages and First Feeding in Farmed Gilthead Sea Bream
(Sparus aurata). Genes (Basel). 10, 483. https://doi.org/10.3390/genes10070483
Oksanen, J., Kindt, R., Legendre, P., O’Hara, B., Simpson, G.L., Stevens, M.H.H., Wagner,
H., 2008. The vegan package: community ecology package, version 1.13-1 [WWW
Document].
Parris, D.J., Brooker, R.M., Morgan, M.A., Dixson, D.L., Stewart, F.J., 2016. Whole gut
microbiome composition of damselfish and cardinalfish before and after reef settlement.
PeerJ 2016, e2412. https://doi.org/10.7717/PEERJ.2412
Patrolecco, L., Ademollo, N., Capri, S., Pagnotta, R., Polesello, S., 2010. Occurrence of
priority hazardous PAHs in water, suspended particulate matter, sediment and common
eels (Anguilla anguilla) in the urban stretch of the River Tiber (Italy). Chemosphere 81,
1386–1392. https://doi.org/10.1016/j.chemosphere.2010.09.027
Pratte, Z.A., Besson, M., Hollman, R.D., Stewart, F.J., 2018. The Gills of Reef Fish Support a
Distinct Microbiome Influenced by Host-Specific Factors. https://doi.org/10.1128/AEM
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., Peplies, J., Glöckner,
F.O., 2013. The SILVA ribosomal RNA gene database project: Improved data processing
and web-based tools. Nucleic Acids Res. 41, D590–D596.
https://doi.org/10.1093/nar/gks1219
Quercia, S., Freccero, F., Castagnetti, C., Soverini, M., Turroni, S., Biagi, E., Rampelli, S.,
Lanci, A., Mariella, J., Chinellato, E., Brigidi, P., Candela, M., 2019. Early colonisation
and temporal dynamics of the gut microbial ecosystem in Standardbred foals. Equine
Vet. J. 51, 231–237. https://doi.org/10.1111/evj.12983
Reid, K.M., Patel, S., Robinson, A.J., Bu, L., Jarungsriapisit, J., Moore, L.J., Salinas, I., 2017.
Salmonid alphavirus infection causes skin dysbiosis in Atlantic salmon (Salmo salar L.)
post-smolts. PLoS One 12, e0172856. https://doi.org/10.1371/journal.pone.0172856

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
113
Reinhart, E.M., Korry, B.J., Rowan-Nash, A.D., Belenky, P., 2019. Defining the Distinct Skin
and Gut Microbiomes of the Northern Pike (Esox lucius). Front. Microbiol. 10, 2118.
https://doi.org/10.3389/fmicb.2019.02118
Reyes-Jurado, F., Franco-Vega, A., Ramírez-Corona, N., Palou, E., López-Malo, A., 2015.
Essential Oils: Antimicrobial Activities, Extraction Methods, and Their Modeling. Food
Eng. Rev. https://doi.org/10.1007/s12393-014-9099-2
Richards, R.H., Pickering, A.D., 1978. Frequency and distribution patterns of Saprolegnia
infection in wild and hatchery-reared brown trout Salmo trutta L. and char Salvelinus
alpinus (L.). J. Fish Dis. 1, 69–82. https://doi.org/10.1111/j.1365-2761.1978.tb00006.x
Roberts, R.J., Bullock, A.M., 1980. The skin surface ecosystem of teleost fishes. Proc. R. Soc.
Edinburgh. Sect. B. Biol. Sci. 79, 87–91. https://doi.org/10.1017/s0269727000010332
Rosado, D., Pérez-Losada, M., Severino, R., Cable, J., Xavier, R., 2019a. Characterization of
the skin and gill microbiomes of the farmed seabass (Dicentrarchus labrax) and
seabream (Sparus aurata). Aquaculture 500, 57–64.
https://doi.org/10.1016/j.aquaculture.2018.09.063
Rosado, D., Xavier, R., Severino, R., Tavares, F., Cable, J., Pérez-Losada, M., 2019b. Effects
of disease, antibiotic treatment and recovery trajectory on the microbiome of farmed
seabass (Dicentrarchus labrax). Sci. Rep. 9, 1–11. https://doi.org/10.1038/s41598-019-
55314-4
Ross, A.A., Rodrigues Hoffmann, A., Neufeld, J.D., 2019. The skin microbiome of vertebrates.
Microbiome. https://doi.org/10.1186/s40168-019-0694-6
Saxena, R., Mittal, P., Clavaud, C., Dhakan, D.B., Hegde, P., Veeranagaiah, M.M., Saha, S.,
Souverain, L., Roy, N., Breton, L., Misra, N., Sharma, V.K., 2018. Comparison of Healthy
and Dandruff Scalp Microbiome Reveals the Role of Commensals in Scalp Health. Front.
Cell. Infect. Microbiol. 8, 346. https://doi.org/10.3389/fcimb.2018.00346
Schmidt, J.G., Thompson, Kim D, Padrós, Francesc, Thompson, K D, Padrós, F, 2018.
Emerging skin diseases in aquaculture. Bull. Eur. Assoc. Fish Pathol. 38, 122–129.
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W.S., Huttenhower, C.,
2011. Metagenomic biomarker discovery and explanation. Genome Biol. 12, R60.
https://doi.org/10.1186/gb-2011-12-6-r60
Shao, Y., Huo, D., Peng, Q., Pan, Y., Jiang, S., Liu, B., Zhang, J., 2017. Lactobacillus
plantarum HNU082-derived improvements in the intestinal microbiome prevent the
development of hyperlipidaemia. Food Funct. 8, 4508–4516.
https://doi.org/10.1039/c7fo00902j
Stephens, W.Z., Burns, A.R., Stagaman, K., Wong, S., Rawls, J.F., Guillemin, K., Bohannan,
B.J.M., 2016. The composition of the zebrafish intestinal microbial community varies
across development. ISME J. 10, 644–654. https://doi.org/10.1038/ismej.2015.140

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
114
Tarnecki, A.M., Brennan, N.P., Schloesser, R.W., Rhody, N.R., 2019. Shifts in the Skin-
Associated Microbiota of Hatchery-Reared Common Snook Centropomus undecimalis
During Acclimation to the Wild. Microb. Ecol. 77, 770–781.
https://doi.org/10.1007/s00248-018-1252-7
Tarnecki, A.M., Rhody, N.R., Walsh, C.J., 2018. Health Characteristics and Blood Bacterial
Assemblages of Healthy Captive Red Drum: Implications for Aquaculture and Fish Health
Management. J. Aquat. Anim. Health 30, 339–353. https://doi.org/10.1002/aah.10047
Thompson, L.R., Sanders, J.G., McDonald, D., Amir, A., Ladau, J., Locey, K.J., Prill, R.J.,
Tripathi, A., Gibbons, S.M., Ackermann, G., Navas-Molina, J.A., Janssen, S., Kopylova,
E., Vázquez-Baeza, Y., González, A., Morton, J.T., Mirarab, S., Xu, Z.Z., Jiang, L.,
Haroon, M.F., Kanbar, J., Zhu, Q., Song, S.J., Kosciolek, T., Bokulich, N.A., Lefler, J.,
Brislawn, C.J., Humphrey, G., Owens, S.M., Hampton-Marcell, J., Berg-Lyons, D.,
McKenzie, V., Fierer, N., Fuhrman, J.A., Clauset, A., Stevens, R.L., Shade, A., Pollard,
K.S., Goodwin, K.D., Jansson, J.K., Gilbert, J.A., Knight, R., Agosto Rivera, J.L., Al-
Moosawi, L., Alverdy, J., Amato, K.R., Andras, J., Angenent, L.T., Antonopoulos, D.A.,
Apprill, A., Armitage, D., Ballantine, K., Bárta, J., Baum, J.K., Berry, A., Bhatnagar, A.,
Bhatnagar, M., Biddle, J.F., Bittner, L., Boldgiv, B., Bottos, E., Boyer, D.M., Braun, J.,
Brazelton, W., Brearley, F.Q., Campbell, A.H., Caporaso, J.G., Cardona, C., Carroll, J.L.,
Cary, S.C., Casper, B.B., Charles, T.C., Chu, H., Claar, D.C., Clark, R.G., Clayton, J.B.,
Clemente, J.C., Cochran, A., Coleman, M.L., Collins, G., Colwell, R.R., Contreras, M.,
Crary, B.B., Creer, S., Cristol, D.A., Crump, B.C., Cui, D., Daly, S.E., Davalos, L.,
Dawson, R.D., Defazio, J., Delsuc, F., Dionisi, H.M., Dominguez-Bello, M.G., Dowell, R.,
Dubinsky, E.A., Dunn, P.O., Ercolini, D., Espinoza, R.E., Ezenwa, V., Fenner, N., Findlay,
H.S., Fleming, I.D., Fogliano, V., Forsman, A., Freeman, C., Friedman, E.S., Galindo, G.,
Garcia, L., Garcia-Amado, M.A., Garshelis, D., Gasser, R.B., Gerdts, G., Gibson, M.K.,
Gifford, I., Gill, R.T., Giray, T., Gittel, A., Golyshin, P., Gong, D., Grossart, H.P., Guyton,
K., Haig, S.J., Hale, V., Hall, R.S., Hallam, S.J., Handley, K.M., Hasan, N.A., Haydon,
S.R., Hickman, J.E., Hidalgo, G., Hofmockel, K.S., Hooker, J., Hulth, S., Hultman, J.,
Hyde, E., Ibáñez-Álamo, J.D., Jastrow, J.D., Jex, A.R., Johnson, L.S., Johnston, E.R.,
Joseph, S., Jurburg, S.D., Jurelevicius, D., Karlsson, A., Karlsson, R., Kauppinen, S.,
Kellogg, C.T.E., Kennedy, S.J., Kerkhof, L.J., King, G.M., Kling, G.W., Koehler, A. V.,
Krezalek, M., Kueneman, J., Lamendella, R., Landon, E.M., Lanede Graaf, K., LaRoche,
J., Larsen, P., Laverock, B., Lax, S., Lentino, M., Levin, I.I., Liancourt, P., Liang, W., Linz,
A.M., Lipson, D.A., Liu, Y., Lladser, M.E., Lozada, M., Spirito, C.M., MacCormack, W.P.,
MacRae-Crerar, A., Magris, M., Martín-Platero, A.M., Martín-Vivaldi, M., Martínez, L.M.,
Martínez-Bueno, M., Marzinelli, E.M., Mason, O.U., Mayer, G.D., McDevitt-Irwin, J.M.,
McDonald, J.E., McGuire, K.L., McMahon, K.D., McMinds, R., Medina, M., Mendelson,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
115
J.R., Metcalf, J.L., Meyer, F., Michelangeli, F., Miller, K., Mills, D.A., Minich, J., Mocali,
S., Moitinho-Silva, L., Moore, A., Morgan-Kiss, R.M., Munroe, P., Myrold, D., Neufeld,
J.D., Ni, Y., Nicol, G.W., Nielsen, S., Nissimov, J.I., Niu, K., Nolan, M.J., Noyce, K.,
O’Brien, S.L., Okamoto, N., Orlando, L., Castellano, Y.O., Osuolale, O., Oswald, W.,
Parnell, J., Peralta-Sánchez, J.M., Petraitis, P., Pfister, C., Pilon-Smits, E., Piombino, P.,
Pointing, S.B., Pollock, F.J., Potter, C., Prithiviraj, B., Quince, C., Rani, A., Ranjan, R.,
Rao, S., Rees, A.P., Richardson, M., Riebesell, U., Robinson, C., Rockne, K.J.,
Rodriguezl, S.M., Rohwer, F., Roundstone, W., Safran, R.J., Sangwan, N., Sanz, V.,
Schrenk, M., Schrenzel, M.D., Scott, N.M., Seger, R.L., Seguinorlando, A., Seldin, L.,
Seyler, L.M., Shakhsheer, B., Sheets, G.M., Shen, C., Shi, Y., Shin, H., Shogan, B.D.,
Shutler, D., Siegel, J., Simmons, S., Sjöling, S., Smith, D.P., Soler, J.J., Sperling, M.,
Steinberg, P.D., Stephens, B., Stevens, M.A., Taghavi, S., Tai, V., Tait, K., Tan, C.L.,
Taş, N., Taylor, D.L., Thomas, T., Timling, I., Turner, B.L., Urich, T., Ursell, L.K., Van Der
Lelie, D., Van Treuren, W., Van Zwieten, L., Vargas-Robles, D., Thurber, R.V., Vitaglione,
P., Walker, D.A., Walters, W.A., Wang, S., Wang, T., Weaver, T., Webster, N.S., Wehrle,
B., Weisenhorn, P., Weiss, S., Werner, J.J., West, K., Whitehead, A., Whitehead, S.R.,
Whittingham, L.A., Willerslev, E., Williams, A.E., Wood, S.A., Woodhams, D.C., Yang,
Y., Zaneveld, J., Zarraonaindia, I., Zhang, Q., Zhao, H., 2017. A communal catalogue
reveals Earth’s multiscale microbial diversity. Nature 551, 457–463.
https://doi.org/10.1038/nature24621
Tofalo, R., Cocchi, S., Suzzi, G., 2019. Polyamines and Gut Microbiota. Front. Nutr. 6, 16.
https://doi.org/10.3389/fnut.2019.00016
Trivedi, B., 2012. Microbiome: the surface brigade. Nature 492, S60–S61.
Uren Webster, T.M., Consuegra, S., Hitchings, M., Garcia de Leaniz, C., 2018. Interpopulation
Variation in the Atlantic Salmon Microbiome Reflects Environmental and Genetic
Diversity. Appl. Environ. Microbiol. 84. https://doi.org/10.1128/aem.00691-18
Uren Webster, T.M., Rodriguez-Barreto, D., Castaldo, G., Gough, P., Consuegra, S., Garcia
de Leaniz, C., 2020. Environmental plasticity and colonisation history in the Atlantic
salmon microbiome: A translocation experiment. Mol. Ecol. 29, 886–898.
https://doi.org/10.1111/mec.15369
Van Der Meer, J.R., 1997. Evolution of novel metabolic pathways for the degradation of
chloroaromatic compounds, in: Antonie van Leeuwenhoek, International Journal of
General and Molecular Microbiology. Kluwer Academic Publishers, pp. 159–178.
https://doi.org/10.1023/A:1000166400935
Varsamos, S., Nebel, C., Charmantier, G., 2005. Ontogeny of osmoregulation in
postembryonic fish: A review, in: Comparative Biochemistry and Physiology - A Molecular
and Integrative Physiology. Elsevier Inc., pp. 401–429.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
116
https://doi.org/10.1016/j.cbpb.2005.01.013
Villamil, S.I., Huerlimann, R., Morianos, C., Sarnyai, Z., Maes, G.E., 2018. Adverse effect of
early-life high-fat/high-carbohydrate (“Western”) diet on bacterial community in the distal
bowel of mice. Nutr. Res. 50, 25–36. https://doi.org/10.1016/j.nutres.2017.11.008
Walleser, L.R., Sandheinrich, M.B., Howard, D.R., Gaikowski, M.P., Amberg, J.J., 2014.
Spatial and Temporal Variation of the Gill Rakers of Gizzard Shad and Silver Carp in
Three Midwestern Rivers. North Am. J. Fish. Manag. 34, 875–884.
https://doi.org/10.1080/02755947.2014.920740
Warne, R.W., Kirschman, L., Zeglin, L., 2019. Manipulation of gut microbiota during critical
developmental windows affects host physiological performance and disease
susceptibility across ontogeny. J. Anim. Ecol. 88, 1365-2656.12973.
https://doi.org/10.1111/1365-2656.12973
Whitear, M., 2009. The skin surface of bony fishes. J. Zool. 160, 437–454.
https://doi.org/10.1111/j.1469-7998.1970.tb03091.x
Wilkes Walburn, J., Wemheuer, B., Thomas, T., Copeland, E., O’Connor, W., Booth, M.,
Fielder, S., Egan, S., 2019. Diet and diet-associated bacteria shape early microbiome
development in Yellowtail Kingfish (Seriola lalandi). Microb. Biotechnol. 12, 275–288.
https://doi.org/10.1111/1751-7915.13323
Wilkins, N.P., Jancsar, S., 1979. Temporal variations in the skin of Atlantic salmon Salmo salar
L. J. Fish Biol. 15, 299–307. https://doi.org/10.1111/j.1095-8649.1979.tb03609.x
Wong, S., Zac Stephens, W., Burns, A.R., Stagaman, K., David, L.A., Bohannan, B.J.M.,
Guillemin, K., Rawls, J.F., 2015. Ontogenetic differences in dietary fat influence
microbiota assembly in the zebrafish gut. MBio 6. https://doi.org/10.1128/mBio.00687-15
Wu, Y., Xiao, F., Wang, C., Shu, L., Zheng, X., Xu, K., Yu, X., Zhang, K., Luo, H., Yang, Y.,
He, Z., Yan, Q., 2020. The Beta-Diversity of Siganus fuscescens-Associated Microbial
Communities From Different Habitats Increases With Body Weight. Front. Microbiol. 11,
1562. https://doi.org/10.3389/fmicb.2020.01562
Xavier, R., Pereira, A., Pagan, A., Hendrick, G.C., Nicholson, M.D., Rosado, D., Soares, M.C.,
Pérez-Losada, M., Sikkel, P.C., 2020. The effects of environment and ontogeny on the
skin microbiome of two Stegastes damselfishes (Pomacentridae) from the eastern
Caribbean Sea. Mar. Biol. 167, 102. https://doi.org/10.1007/s00227-020-03717-7
Yan, Q., Li, J., Yu, Y., Wang, J., He, Z., Van Nostrand, J.D., Kempher, M.L., Wu, L., Wang,
Y., Liao, L., Li, X., Wu, S., Ni, J., Wang, C., Zhou, J., 2016. Environmental filtering
decreases with fish development for the assembly of gut microbiota. Environ. Microbiol.
18, 4739–4754. https://doi.org/10.1111/1462-2920.13365
Zhang, Xiaoting, Ding, L., Yu, Y., Kong, W., Yin, Y., Huang, Z., Zhang, Xuezhen, Xu, Z., 2018.
The Change of Teleost Skin Commensal Microbiota Is Associated With Skin Mucosal

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
117
Transcriptomic Responses During Parasitic Infection by Ichthyophthirius multifillis. Front.
Immunol. 9, 2972. https://doi.org/10.3389/fimmu.2018.02972
Zohar, Y., Gordin, H., 1979. Spawning kinetics in the gilthead sea-bream, Sparus aurata L.
after low doses of human chronic gonadotropin. J. Fish Biol. 15, 665–670.
https://doi.org/10.1111/j.1095-8649.1979.tb03675.x
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
118
3.9 Supplementary material
Figure S3.1 Illustrative scheme of the semi-intensive fish farm where samples were collected. All ponds shared the same inflow of estuarine water (A) and water was never shared between ponds. Each age group and species was reared in separated but not distant open water ponds: 1 - mature adults seabass; 2 - mature adults seabream; 3 - late juveniles seabass; 4 - early juveniles seabass; 5 - early juveniles seabream.
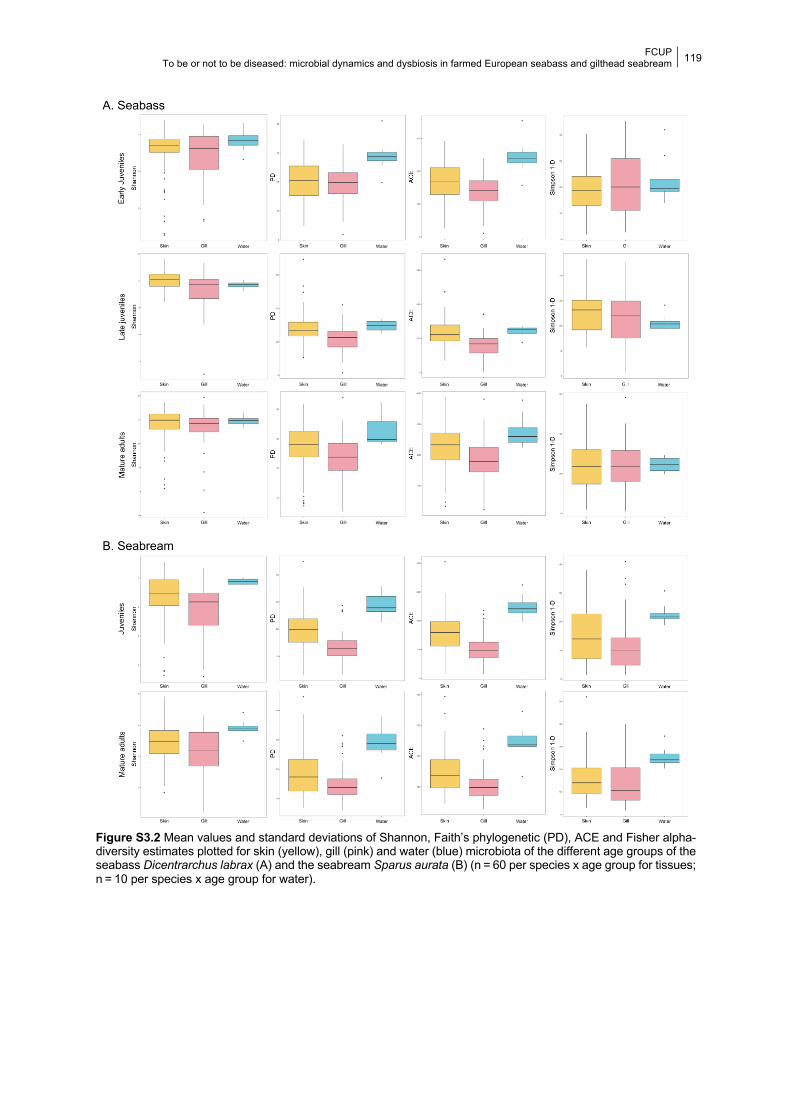
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
119
Figure S3.2 Mean values and standard deviations of Shannon, Faith’s phylogenetic (PD), ACE and Fisher alpha-diversity estimates plotted for skin (yellow), gill (pink) and water (blue) microbiota of the different age groups of the seabass Dicentrarchus labrax (A) and the seabream Sparus aurata (B) (n = 60 per species x age group for tissues; n = 10 per species x age group for water).
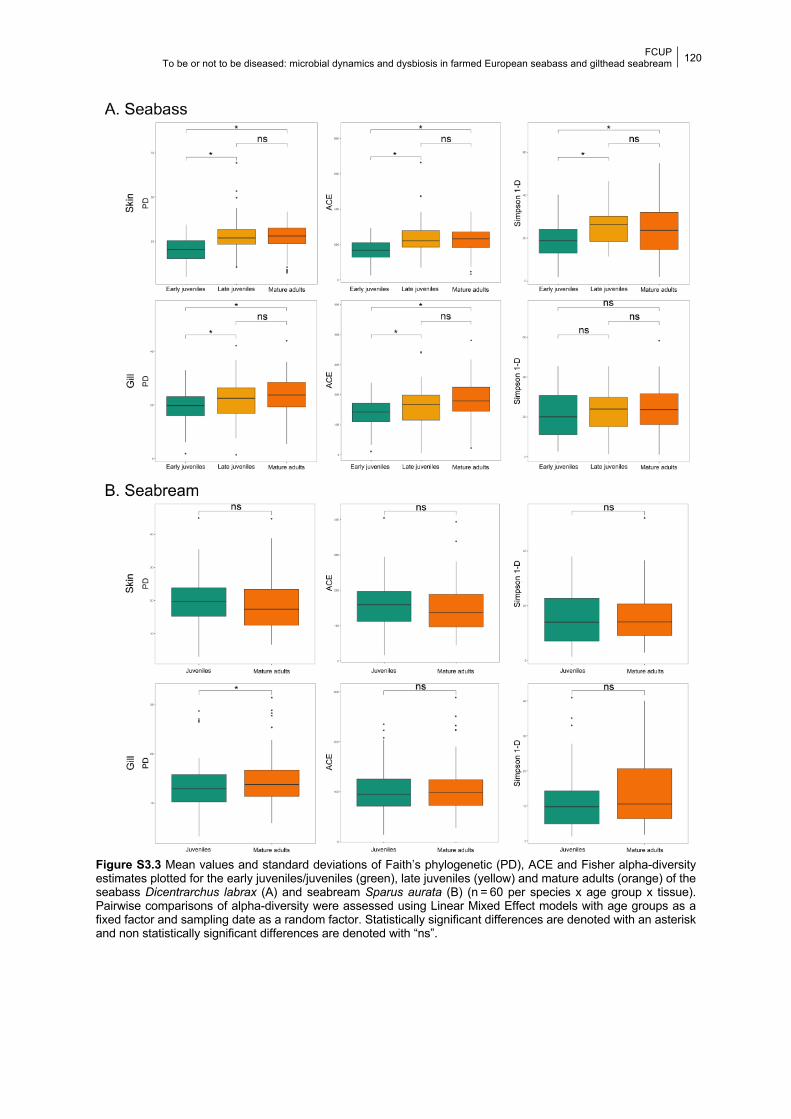
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
120
Figure S3.3 Mean values and standard deviations of Faith’s phylogenetic (PD), ACE and Fisher alpha-diversity estimates plotted for the early juveniles/juveniles (green), late juveniles (yellow) and mature adults (orange) of the seabass Dicentrarchus labrax (A) and seabream Sparus aurata (B) (n = 60 per species x age group x tissue). Pairwise comparisons of alpha-diversity were assessed using Linear Mixed Effect models with age groups as a fixed factor and sampling date as a random factor. Statistically significant differences are denoted with an asterisk and non statistically significant differences are denoted with “ns”.
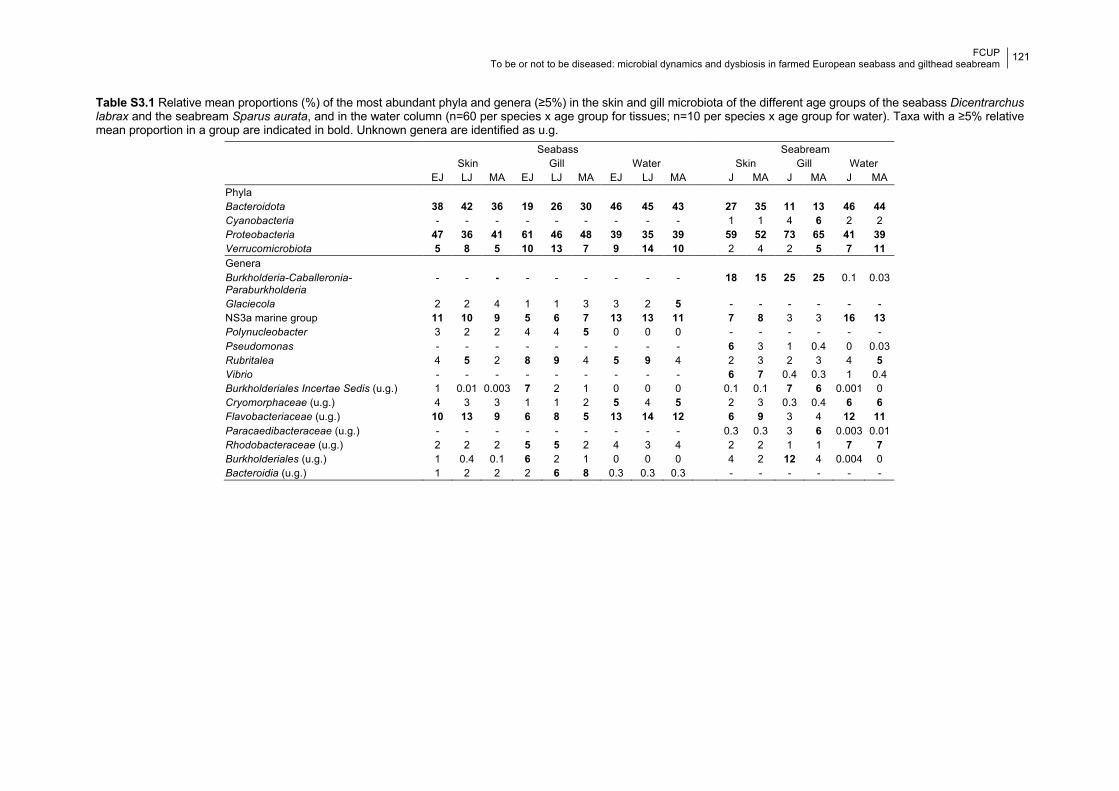
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
121
Table S3.1 Relative mean proportions (%) of the most abundant phyla and genera (≥5%) in the skin and gill microbiota of the different age groups of the seabass Dicentrarchus labrax and the seabream Sparus aurata, and in the water column (n=60 per species x age group for tissues; n=10 per species x age group for water). Taxa with a ≥5% relative mean proportion in a group are indicated in bold. Unknown genera are identified as u.g. Seabass Seabream
Skin Gill Water Skin Gill Water EJ LJ MA EJ LJ MA EJ LJ MA J MA J MA J MA Phyla Bacteroidota 38 42 36 19 26 30 46 45 43 27 35 11 13 46 44 Cyanobacteria - - - - - - - - - 1 1 4 6 2 2 Proteobacteria 47 36 41 61 46 48 39 35 39 59 52 73 65 41 39 Verrucomicrobiota 5 8 5 10 13 7 9 14 10 2 4 2 5 7 11 Genera Burkholderia-Caballeronia-Paraburkholderia
- - - - - - - - - 18 15 25 25 0.1 0.03
Glaciecola 2 2 4 1 1 3 3 2 5 - - - - - - NS3a marine group 11 10 9 5 6 7 13 13 11 7 8 3 3 16 13 Polynucleobacter 3 2 2 4 4 5 0 0 0 - - - - - - Pseudomonas - - - - - - - - - 6 3 1 0.4 0 0.03 Rubritalea 4 5 2 8 9 4 5 9 4 2 3 2 3 4 5 Vibrio - - - - - - - - - 6 7 0.4 0.3 1 0.4 Burkholderiales Incertae Sedis (u.g.) 1 0.01 0.003 7 2 1 0 0 0 0.1 0.1 7 6 0.001 0 Cryomorphaceae (u.g.) 4 3 3 1 1 2 5 4 5 2 3 0.3 0.4 6 6 Flavobacteriaceae (u.g.) 10 13 9 6 8 5 13 14 12 6 9 3 4 12 11 Paracaedibacteraceae (u.g.) - - - - - - - - - 0.3 0.3 3 6 0.003 0.01 Rhodobacteraceae (u.g.) 2 2 2 5 5 2 4 3 4 2 2 1 1 7 7 Burkholderiales (u.g.) 1 0.4 0.1 6 2 1 0 0 0 4 2 12 4 0.004 0 Bacteroidia (u.g.) 1 2 2 2 6 8 0.3 0.3 0.3 - - - - - -
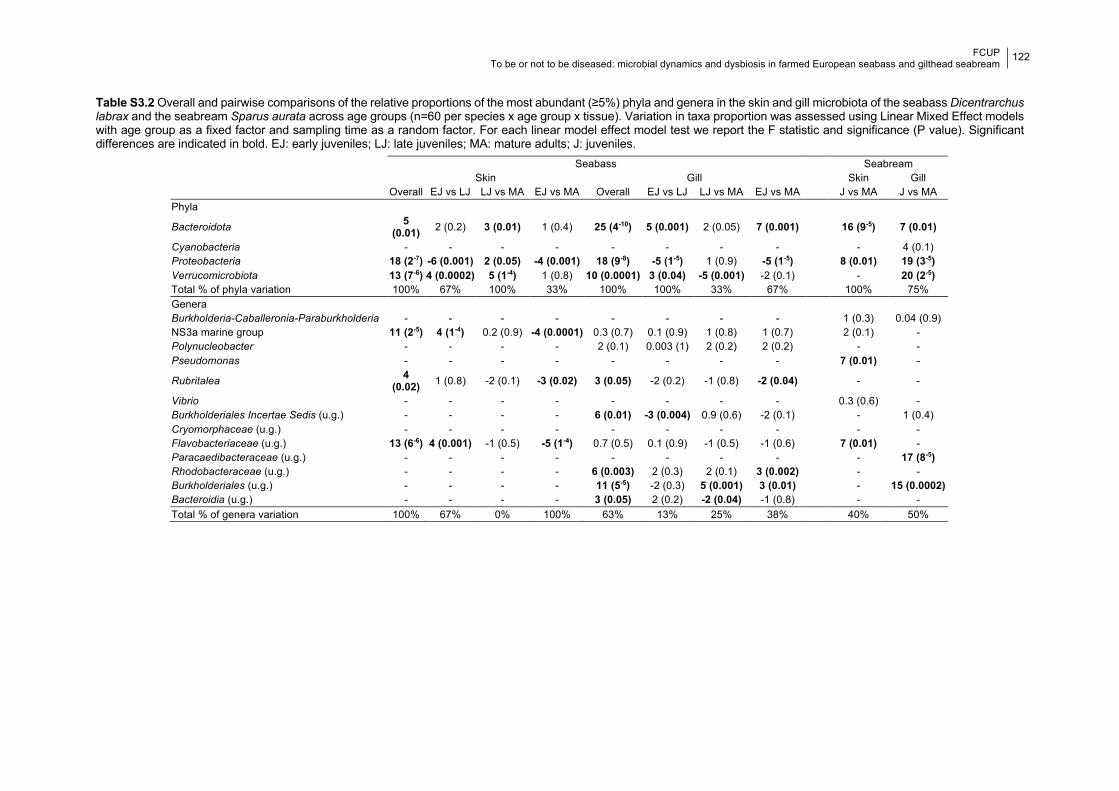
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
122
Table S3.2 Overall and pairwise comparisons of the relative proportions of the most abundant (≥5%) phyla and genera in the skin and gill microbiota of the seabass Dicentrarchus labrax and the seabream Sparus aurata across age groups (n=60 per species x age group x tissue). Variation in taxa proportion was assessed using Linear Mixed Effect models with age group as a fixed factor and sampling time as a random factor. For each linear model effect model test we report the F statistic and significance (P value). Significant differences are indicated in bold. EJ: early juveniles; LJ: late juveniles; MA: mature adults; J: juveniles.
Seabass Seabream Skin Gill Skin Gill Overall EJ vs LJ LJ vs MA EJ vs MA Overall EJ vs LJ LJ vs MA EJ vs MA J vs MA J vs MA Phyla
Bacteroidota 5 (0.01) 2 (0.2) 3 (0.01) 1 (0.4) 25 (4-10) 5 (0.001) 2 (0.05) 7 (0.001) 16 (9-5) 7 (0.01)
Cyanobacteria - - - - - - - - - 4 (0.1) Proteobacteria 18 (2-7) -6 (0.001) 2 (0.05) -4 (0.001) 18 (9-8) -5 (1-5) 1 (0.9) -5 (1-5) 8 (0.01) 19 (3-5) Verrucomicrobiota 13 (7-6) 4 (0.0002) 5 (1-4) 1 (0.8) 10 (0.0001) 3 (0.04) -5 (0.001) -2 (0.1) - 20 (2-5) Total % of phyla variation 100% 67% 100% 33% 100% 100% 33% 67% 100% 75% Genera Burkholderia-Caballeronia-Paraburkholderia - - - - - - - - 1 (0.3) 0.04 (0.9) NS3a marine group 11 (2-5) 4 (1-4) 0.2 (0.9) -4 (0.0001) 0.3 (0.7) 0.1 (0.9) 1 (0.8) 1 (0.7) 2 (0.1) - Polynucleobacter - - - - 2 (0.1) 0.003 (1) 2 (0.2) 2 (0.2) - - Pseudomonas - - - - - - - - 7 (0.01) -
Rubritalea 4 (0.02) 1 (0.8) -2 (0.1) -3 (0.02) 3 (0.05) -2 (0.2) -1 (0.8) -2 (0.04) - -
Vibrio - - - - - - - - 0.3 (0.6) - Burkholderiales Incertae Sedis (u.g.) - - - - 6 (0.01) -3 (0.004) 0.9 (0.6) -2 (0.1) - 1 (0.4) Cryomorphaceae (u.g.) - - - - - - - - - - Flavobacteriaceae (u.g.) 13 (6-6) 4 (0.001) -1 (0.5) -5 (1-4) 0.7 (0.5) 0.1 (0.9) -1 (0.5) -1 (0.6) 7 (0.01) - Paracaedibacteraceae (u.g.) - - - - - - - - - 17 (8-5) Rhodobacteraceae (u.g.) - - - - 6 (0.003) 2 (0.3) 2 (0.1) 3 (0.002) - - Burkholderiales (u.g.) - - - - 11 (5-5) -2 (0.3) 5 (0.001) 3 (0.01) - 15 (0.0002) Bacteroidia (u.g.) - - - - 3 (0.05) 2 (0.2) -2 (0.04) -1 (0.8) - - Total % of genera variation 100% 67% 0% 100% 63% 13% 25% 38% 40% 50%
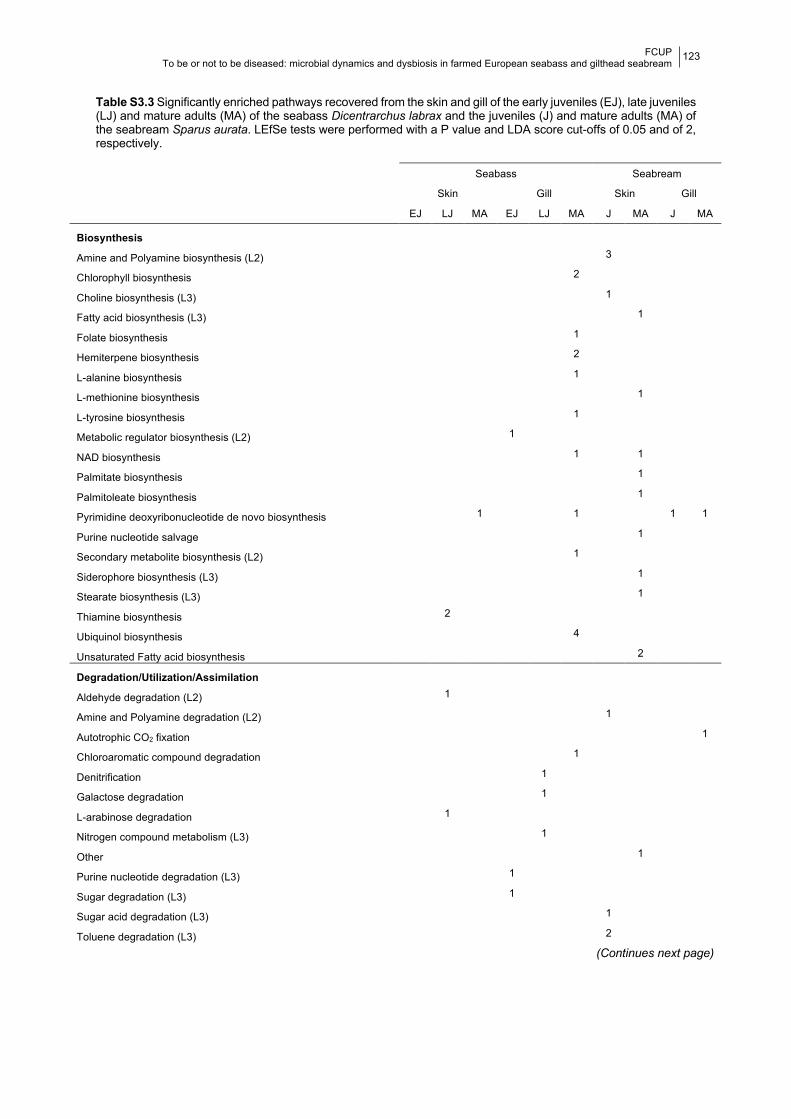
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
123
Table S3.3 Significantly enriched pathways recovered from the skin and gill of the early juveniles (EJ), late juveniles (LJ) and mature adults (MA) of the seabass Dicentrarchus labrax and the juveniles (J) and mature adults (MA) of the seabream Sparus aurata. LEfSe tests were performed with a P value and LDA score cut-offs of 0.05 and of 2, respectively.
Seabass Seabream
Skin Gill Skin Gill
EJ LJ MA EJ LJ MA J MA J MA
Biosynthesis
Amine and Polyamine biosynthesis (L2) 3
Chlorophyll biosynthesis 2
Choline biosynthesis (L3) 1
Fatty acid biosynthesis (L3) 1
Folate biosynthesis 1
Hemiterpene biosynthesis 2
L-alanine biosynthesis 1
L-methionine biosynthesis 1
L-tyrosine biosynthesis 1
Metabolic regulator biosynthesis (L2) 1
NAD biosynthesis 1 1
Palmitate biosynthesis 1
Palmitoleate biosynthesis 1
Pyrimidine deoxyribonucleotide de novo biosynthesis 1 1 1 1
Purine nucleotide salvage 1
Secondary metabolite biosynthesis (L2) 1
Siderophore biosynthesis (L3) 1
Stearate biosynthesis (L3) 1
Thiamine biosynthesis 2
Ubiquinol biosynthesis 4
Unsaturated Fatty acid biosynthesis 2
Degradation/Utilization/Assimilation
Aldehyde degradation (L2) 1
Amine and Polyamine degradation (L2) 1
Autotrophic CO2 fixation 1
Chloroaromatic compound degradation 1
Denitrification 1
Galactose degradation 1
L-arabinose degradation 1
Nitrogen compound metabolism (L3) 1
Other 1
Purine nucleotide degradation (L3) 1
Sugar degradation (L3) 1
Sugar acid degradation (L3) 1
Toluene degradation (L3) 2
(Continues next page)
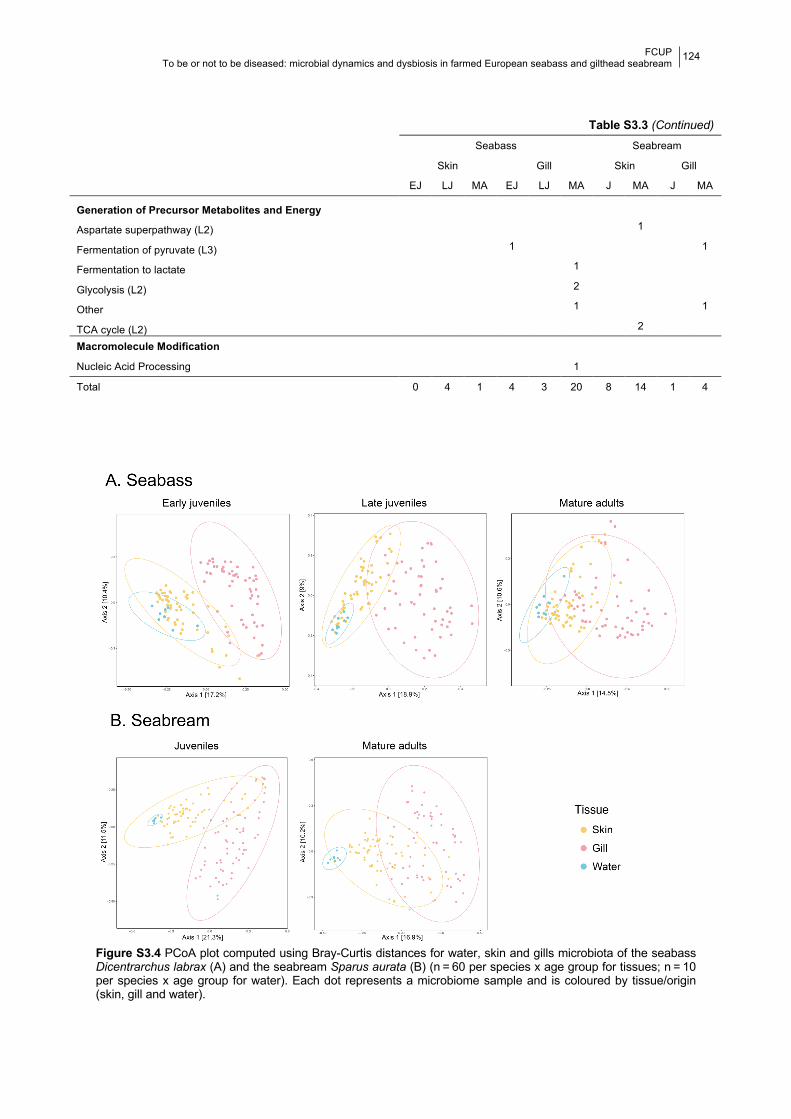
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
124
Figure S3.4 PCoA plot computed using Bray-Curtis distances for water, skin and gills microbiota of the seabass Dicentrarchus labrax (A) and the seabream Sparus aurata (B) (n = 60 per species x age group for tissues; n = 10 per species x age group for water). Each dot represents a microbiome sample and is coloured by tissue/origin (skin, gill and water).
Table S3.3 (Continued)
Seabass Seabream
Skin Gill Skin Gill
EJ LJ MA EJ LJ MA J MA J MA
Generation of Precursor Metabolites and Energy
Aspartate superpathway (L2) 1
Fermentation of pyruvate (L3) 1 1
Fermentation to lactate 1
Glycolysis (L2) 2
Other 1 1
TCA cycle (L2) 2
Macromolecule Modification
Nucleic Acid Processing 1
Total 0 4 1 4 3 20 8 14 1 4

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
125
CHAPTER 4
Longitudinal sampling of external mucosae in
farmed European seabass reveals the impact of
water temperature on bacterial dynamics
Daniela Rosado, Raquel Xavier, Jo Cable, Ricardo Severino, Pedro
Tarroso, Marcos Pérez-Losada
2021. ISME Communications. https://doi.org/10.1038/s43705-021-00019-x.
4.1 Abstract Fish microbiota are intrinsically linked to health and fitness, but they are highly variable
and influenced by both biotic and abiotic factors. Water temperature particularly limits bacterial
adhesion and growth, impacting microbial diversity and bacterial infections on the skin and
gills. Aquaculture is heavily affected by infectious diseases, especially in warmer months, and
industry practices often promote stress and microbial dysbiosis, leading to an increased
abundance of potentially pathogenic bacteria. In this regard, fish mucosa health is extremely
important because it provides a primary barrier against pathogens. We used 16 rRNA V4
metataxonomics to characterize the skin and gill microbiota of the European seabass,
Dicentrarchus labrax, and the surrounding water over 12 months, assessing the impact of
water temperature on microbial diversity and function. We show that the microbiota of external
mucosae are highly dynamic with consistent longitudinal trends in taxon diversity. Several
potentially pathogenic genera (Aliivibrio, Photobacterium, Pseudomonas and Vibrio) were
highly abundant, showing complex interactions with other bacterial genera, some of which with
recognized probiotic activity, and were also significantly impacted by changes in temperature.
The surrounding water temperature influenced fish microbial composition, structure and
function over time (days and months). Additionally, dysbiosis was more frequent in warmer
months and during transitions between cold/warm months. We also detected a strong
seasonal effect in the fish microbiota, which is likely to result from the compound action of
several unmeasured environmental factors (e.g., pH, nutrient availability) beyond temperature.
Our results highlight the importance of performing longitudinal studies to assess the impact of
environmental factors on fish microbiotas.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
126
Keywords: aquaculture, microbiome, Dicentrarchus labrax, dysbiosis, fish pathogen
4.2 Introduction Cumulative evidence shows that the diversity of commensal microbiota mirror fish
health and that microbial diversity can improve host fitness (Kelly and Salinas, 2017; Legrand
et al., 2018; Zhang et al., 2018). Fish microbial composition and structure are driven by host
related factors, including host taxonomy and ontogeny (Arias et al., 2019; Rosado et al., 2021),
but also physio-chemical properties of the water (e.g., temperature, pH, oxygen and nutrient
concentrations, Duarte et al., 2019; Kokou et al., 2018; Martins et al., 2018; Minich et al.,
2020), which can be highly dynamic (Yukgehnaish et al., 2020). Consequently, the microbiota
of fish external mucosa, i.e., skin and gills, can be highly variable (Chiarello et al., 2015). Water
temperature, in particular, can be pivotal as it can prompt modifications in the composition of
key components of the fish skin, such as mucins and mucosal immunoglobulins, that influence
the microbiota by limiting bacterial adhesion and growth (Chen et al., 2008; Gomez et al.,
2013; Legrand et al., 2020; Solem and Stenvik, 2006). Additionally, bacteria have optimal
temperature ranges that maximize their growth (Corkrey et al., 2012), thereby conferring some
taxa with a competitive advantage and creating colonization opportunities for other taxa during
temperature shifts (Mouquet et al., 2005). Several microbiome studies have reported direct or
indirect temperature-related changes in fish bacterial diversity of the skin, gill or gut in the
puffer fish (Sugita et al., 1989), brown trout (Vasemägi et al., 2017), tilapia (Kokou et al., 2018),
yellowtail kingfish (Horlick et al., 2020), chub mackerel (Minich et al., 2020) and other teleost
species (Krotman et al., 2020). Importantly, differences in host’s tolerance to temperature
seem to be related to microbial composition and their resilience to temperature changes
(Kokou et al., 2018). Such studies were either based on experimental setups to test the effects
of specific temperatures (22° and 26°C, Horlick et al., 2020; 12° and 24°C, Kokou et al., 2018;
10°, 20° and 29°C, Sugita et al., 1989) or on sampling single time points at different localities
(Krotman et al., 2020; Vasemägi et al., 2017); the only exception being Minich et al. (2020)
who conducted a year long study of the chub mackerel microbiota. Hence, the effects on fish
microbiota of natural fluctuations in sea water temperature are still largely understudied,
although there are increasing reports of a positive correlation between temperature and the
abundance of potentially pathogenic genera (e.g., Vibrio and Flavobacterium, Sugita et al.,
1989; and Photobacterium, Horlick et al., 2020; Minich et al., 2020). Indeed, several bacterial
infections in fish have seasonal distributions with higher incidence in warmer months (e.g.,
Bellos et al., 2015; Habiba et al., 2015; Schade et al., 2016). This pattern is particularly notable
in farmed fish (e.g., Baker-Austin et al., 2013; Eissa et al., 2018; Kayansamruaj et al., 2014;

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
127
Matanza and Osorio, 2018), where the abundance of pathogenic taxa is higher compared to
their wild host counterparts (Tarnecki et al., 2019).
Aquaculture is the fastest growing food-production industry; however, its sustainability
is heavily affected by infectious diseases (Pridgeon and Klesius, 2012). Fish farming practices
are known to promote microbial dysbiosis, which often involves an increase in the abundance
of potentially pathogenic bacteria and a parallel decrease in microbial diversity (e.g., Boutin et
al., 2013). This is critical in fish farms given the predicted increase of diseases due to the
ongoing rise of sea surface temperatures (Baker-Austin et al., 2018, 2017). In this regard, fish
mucosa health is of extreme importance, particularly in the case of the skin and gills, since
both act as primary physical and chemical barriers against pathogens (Beck and Peatman,
2015). Specifically, mucosal microbiota can have an important role in controlling pathogen
abundances either through direct competition or antibiotic production (Dash et al., 2018;
Trivedi, 2012). Indeed, putative bacterial pathogens will integrate within the microbiota of
healthy/asymptomatic fish, where their abundance is controlled by harmless commensal
bacteria (e.g., Califano et al., 2017; Rosado et al., 2019).
The European seabass (Dicentrarchus labrax) is one of the most profitable species
farmed in the Mediterranean region, including southern Portugal (FAO, 2016). Being a
eurythermic (from 5-28°C) and euryhaline (from 3-35%) species, it is traditionally farmed in
outdoor ponds located in protected areas, such as estuaries and coastal lagoons, or in open-
water sea cages (FAO, 2016). As such, farmed fish are subjected to natural fluctuations of
water physio-chemical properties. Among the many diseases reported in European seabass
fish farms, the most frequent have bacterial etiological agents, with vibriosis (caused by Vibrio
spp.) being the most prevalent, followed by photobacteriousis (Photobacterium damselae) and
Tenacibaculosis (Tenacibaculum maritimum) (see Bellos et al., 2015; Muniesa et al., 2020).
Temperature and seasonality are considered the main epizootiological risk factors for severe
disease caused by these pathogens (Bellos et al., 2015).
Here we used 16S rRNA high-throughput sequencing to characterize the composition
and structure of the skin and gill microbiomes of the farmed European seabass and its
surrounding waters over 12 months. We then assessed the impact of water temperature
oscillations measured during different time frames (days, weeks and months prior to sampling)
on microbial diversity and function, as well as the dynamics and effects of potentially
pathogenic bacterial genera in the microbial community.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
128
4.3 Material and methods
4.3.1 Experimental design, sampling and processing European seabass specimens were sampled between February 2017 and January
2018 from a single exterior pond of a fish farm operating in a semi-intensive regime and
located in the Alvor Estuary (Portimão, Portugal). Due to the impossibility of sampling the
same individuals throughout time, a random subset of 5 fish were sampled every fortnight,
totaling 120 fish and 24 sampling time points. Fish were caught using a fishing rod and skin
and gill mucous samples taken immediately using tubed sterile dry swabs (Medical Wire &
Equipment, UK). Water samples (1 L) were concomitantly collected and filtered through 0.2
µm filters, except through December and once in February, due to technical constraints. Fish
diet composition remained stable throughout sampling (SKRETTING/SORGAL), although
pellet size increased from 4 to 6 mm. All sampled fish were considered healthy during the
sampling period based on the lack of visible disease symptoms and mortality fraction recorded
at the sampling pond (i.e., 105 fish, 29% of total density, died in May 2017 from unknown
causes). Fish were not vaccinated against bacterial diseases and no antibiotics were
administered during the sampling period. Water temperature (°C) was measured every 10
seconds in situ throughout the duration of the experiment using a probe placed in the pond at
20 cm deep. The mean daily temperatures ranged between 13.3°C in January (the coldest
month) and 25.1°C in June (the warmest month), with an annual mean temperature of 18.6 ±
2.8°C (Figure S4.1).
Total DNA from 261 samples (120 skin, 120 gill and 21 water filters) was extracted
using the PowerSoil DNA Isolation Kit (QIAGEN, Netherlands). DNA extractions were shipped
on dry ice to the University of Michigan Medical School (USA) for amplification of the V4 hyper-
variable region of the 16S rRNA gene (~250 bp) according to the protocol of Kozich et al.
(2013). All samples were sequenced in a single run on the Illumina MiSeq sequencing
platform.
4.3.2 Data processing and statistical analysis Raw FASTQ files were denoised using the DADA2 pipeline in R vs. 4.0.2 (Callahan et
al., 2016). Microbial composition (alpha-diversity) was calculated using Shannon and Faith’s
phylogenetic diversity (PD) as implemented in the R package phyloseq (McMurdie and
Holmes, 2013). Microbial structure (beta-diversity) was estimated using phylogenetic Unifrac
(unweighted and weighted) distances. Variations in microbial composition and microbial
structure between sampling dates and months were assessed using the Kruskal-Wallis
(Kruskal and Wallis, 1952) and PERMANOVA (Anderson, 2017) tests, as implemented in the
adonis function of the vegan R package (Oksanen et al., 2008). Pairwise comparisons were

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
129
done between all sampling dates and months using the Wilcox tests with Benjamini-Hochberg
correction (stats package; R Team, 2012) and pairwise adonis (Martinez Arbizu, 2017). No
significant differences were found in bacterial diversity between dates within months (results
not shown). Additionally, dissimilarity in microbial structure between samples was visualized
using Principal Coordinates Analysis (PCoA).
We used the software FEAST (Shenhav et al., 2019) to estimate microbial monthly
exchange between skin, gills and water. This software implements an expectation-
maximization algorithm (Moon, 1996) which infers the fraction of the microbiota (i.e., the sink)
originating from different available sources (see Supplementary File S4.1 for more details).
Genera containing important bacterial pathogens affecting aquaculture were identified
according to the list compiled by Haenen (2017). Several of these potentially pathogenic (PP)
genera showed peaks of abundance throughout the year: Aliivibrio, Photobacterium,
Pseudomonas and Vibrio. Temporal Insights into Microbial Ecology (TIME; Baksi et al., 2018)
was used to visualize the temporal dynamics and predicted microbial competition involving
the most abundant PP genera (≥5%, hereon termed PP genera), as well as to assess their
influence in the community structure throughout months (see Supplementary File S4.1 for
more details).
The impact of temperature and season (coded as a categorical variable) on microbial
alpha-diversity was assessed using linear models (lm), while their impact on microbial beta-
diversity was assessed using permutational multivariate analysis of variance (PERMANOVA)
(see Supplementary File S4.1 for more details). Several temperature variables were derived
from the mean (mn) and standard deviation (SD) of the temperatures measured at different
time frames preceding sampling dates. Briefly, we used temperatures from the 2 and 7 days
before each sampling date (T2_mn / T2_SD and T7_mn / T7_SD). We averaged diversity
values per month since no significant differences within months were found and, consequently,
decreased the impact of serial correlation in the analyses. We assessed the influence of
monthly temperature using samples grouped by month and temperature variables derived
from the periods preceding each of the two sampling points in a given month (T4_mn / T4_SD
and T14_mn / T14_SD). Additionally, we tested the effects of monthly mean temperature and
respective standard deviation (T_month_mn / T_month_SD) and of a broader categorical
time-frame created by dividing the year into cold or warm months based on the daily annual
average temperature (18.6 °C, see Supplementary File S4.1 for more details).
We used generalized least squares models (gls) from the nlme R package (Pinheiro
et al., 2021) to estimate the relation between bacterial diversity and the abundance of PP
genera with temperature. For this test, matrices containing pairwise distances for diversity
metrics or abundance of PP genera were correlated with corresponding matrices of
temperature differences (see Supplementary File S4.1 for more details). Due to the high inter-

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
130
individual variability of alpha-diversity estimates observed within some dates and months, and
to test whether temperature could prompt such variability, the impact of temperature on the
standard deviations of Shannon (Shannon SD) and Faith's PD (PD SD) indices was also
assessed for skin and gill microbiota using the same set of tests (lm and PERMANOVA, as
well as gls for pairwise matrices).
The metagenomic Phylogenetic Investigation of Communities by Reconstruction of
Unobserved States software (PICRUSt2) embedded in QIIME2 (Douglas et al., 2019) was
used to predict the putative microbial metabolism, applying a weighted nearest taxon index
(NSTI) cutoff of 0.03. Predicted metagenomes were collapsed using the Kyoto Encyclopaedia
of Genes and Genomes (KEGG) Pathway metadata (Kanehisa et al., 2019). We identified
differentially abundant predicted metabolic pathways between cold/warm months in the skin
and gill microbiota using linear discriminant analysis (LDA) in LEfSe (Segata et al., 2011). To
this end, we used a P-value cut-off of 0.05 and a LDA effect size cut-off of 2 (Segata et al.,
2011).
A more detailed description of the Materials and Methods above is provided in
Supplementary File S4.1.
4.4 Results
4.4.1 Bacterial composition and temporal dynamics of the microbiota Six bacterial phyla and 28 genera were categorized as the most abundant taxa (≥5%)
in European seabass skin and gill, and associated water, microbiota throughout the 12 month
study period (Figure S4.2, Table S4.1). Bacteroidota and Proteobacteria were consistently
abundant across all months in the fish microbiota and water, and Verrucomicrobiota was highly
abundant in the water for all months. Glaciecola, NS3a marine group, Pseudoalteromonas,
Psychrobacter, Rubritalea, and an unidentified genus belonging to the Flavobacteriaceae
family were the most abundant genera across all studied microbiota. However, none of the 28
genera were consistently highly abundant across all months in the fish mucosa or water.
Nevertheless, there were a total of 73 core ASVs present in all three studied microbiota
throughout the studied months and from these 44, 42 and 46 ASVs were present in the
microbiota of the skin, gill and water, respectively (Figure 4.1, Table S4.2). The three
microbiota communities analyzed shared 25% of the core ASVs, while skin and gill shared
37%, skin and water shared 43%, and gill and water shared 26% (Figure 4.1). Out of the 73
monthly core ASVs, three belonged to potentially pathogenic (PP) genera: a Photobacterium
ASV in the skin, and two Vibrio ASVs, one present in the skin, and the other in the water (Table
S4.2).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
131
Monthly estimates of microbial alpha-diversity of the skin and gills followed the same
trends (increase/decrease) throughout the sampling period, showing significant monthly
fluctuations (Kruskal-Wallis, P ≤ 0.05; Table S4.3). Pairwise comparisons showed significant
shifts in alpha-diversity mainly between warm months and between cold to warm or warm to
cold (hereafter termed cold/warm) transitions. Chronologically, significant decreases of
Shannon and Faith’s PD indices in both fish mucosae occurred between March and April
(coinciding with the transition from cold to warm months); this was followed by a significant
increase of the Shannon index between April and May in the skin microbiota and a significant
increase of the Faith PD index between May and June in the gill microbiota; between June
and July both alpha-diversity indices significantly decreased in both mucosae (P ≤ 0.05, Table
S4.3). Additionally, between October and November, significant decreases occurred in the
Shannon index for the skin microbiota and in Faith’s PD for the gill microbiota, coinciding with
the transition between warm to cold months (P ≤ 0.05, Table S4.3). Finally, a significant
increase in the Shannon diversity occurred between November and December in the gill
microbiota (P ≤ 0.05, Table S4.3). It is noteworthy that aforementioned significant decreases
in alpha-diversity occurred in parallel with an increase in the abundance of some PP genera
in the skin in April and in both tissues in November (Figure 4.2).
Figure 4.1 Venn diagram showing the number and percentage of shared core ASVs between the skin and gill
microbiota (N=10 x 12 months x tissue) of the seabass Dicentrarchus labrax and the surrounding water (N=2 x 11
months) throughout the 12 sampling months.
Although microbial structure showed significant differences between months
(PERMANOVA, P ≤ 0.05; Table S4.3), pairwise comparisons showed no significant
differences between consecutive months (corrected P ≥ 0.05, Table S4.3). Moreover, there
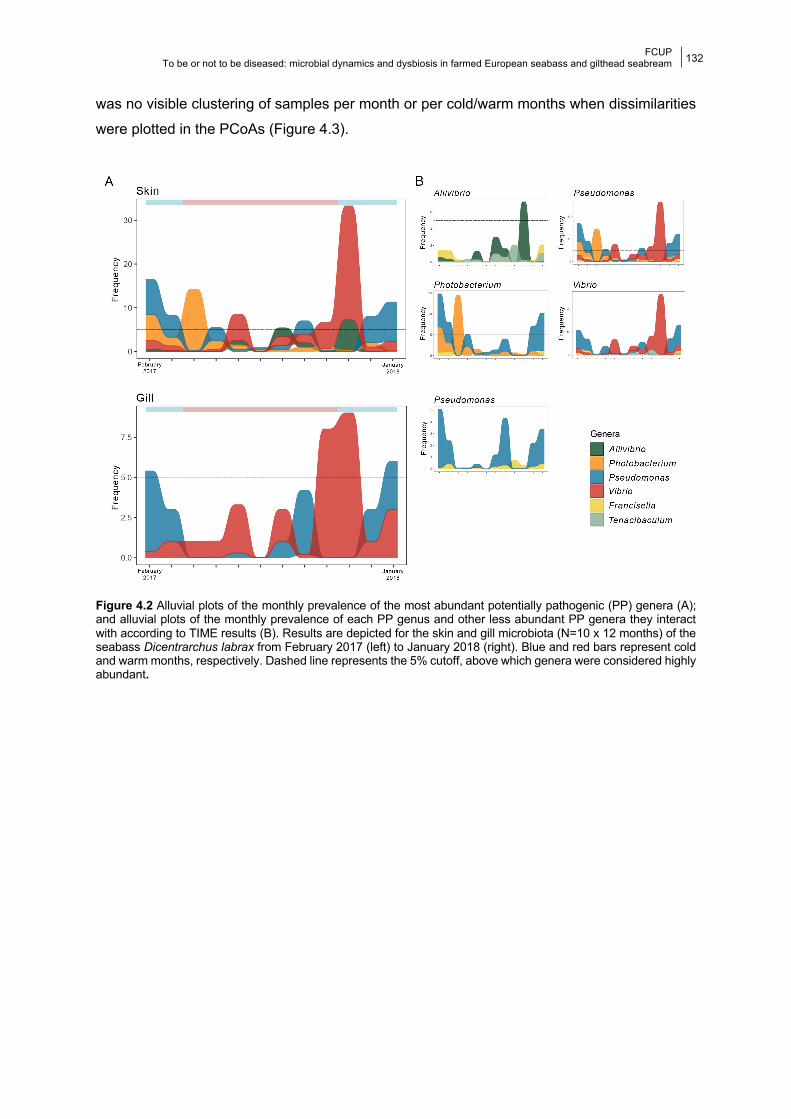
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
132
was no visible clustering of samples per month or per cold/warm months when dissimilarities
were plotted in the PCoAs (Figure 4.3).
Figure 4.2 Alluvial plots of the monthly prevalence of the most abundant potentially pathogenic (PP) genera (A); and alluvial plots of the monthly prevalence of each PP genus and other less abundant PP genera they interact with according to TIME results (B). Results are depicted for the skin and gill microbiota (N=10 x 12 months) of the seabass Dicentrarchus labrax from February 2017 (left) to January 2018 (right). Blue and red bars represent cold and warm months, respectively. Dashed line represents the 5% cutoff, above which genera were considered highly abundant.
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
133
Figure 4.3 Temperature models (A), and beta-diversity (B) and alpha-diversity (C) estimates. The temperature models were calculated for each sampling date and month. Beta-diversity is represented by PCoA plots computed using Unifrac weighted distance for the skin and gill microbiota (N=10 x 12 months x tissue) of the seabass Dicentrarchus labrax and the surrounding water (N=2 x 11 months) throughout the 12 sampling months. Each dot represents a microbiota sample. Alpha-diversity estimates are represented by boxplots depicting mean values and standard deviations of Shannon and Faith’s PD estimates for the skin and gill microbiota (N=10 x 12 months x tissue) of the seabass Dicentrarchus labrax and the surrounding water (N=2 x 11 months) from February 2017 (left) to January 2018 (right). T_month_mn = mean monthly temperature; T_month_SD = standard deviation of T_month_mn; T14_mn = mean temperature from the 7 days previous to both sampling dates in a given month; T14_SD = standard deviation of T14_mn; T4_mn = mean temperature from the 2 days previous to both sampling dates in a given month; T4_SD = standard deviation of T4_mn; T7_mn = mean temperature from the 7 days previous to a sampling date; T7_SD = standard deviation of T7_mn; T2_mn = mean temperature from the 2 days previous to a sampling date; T2_SD = standard deviation of T2_mn.
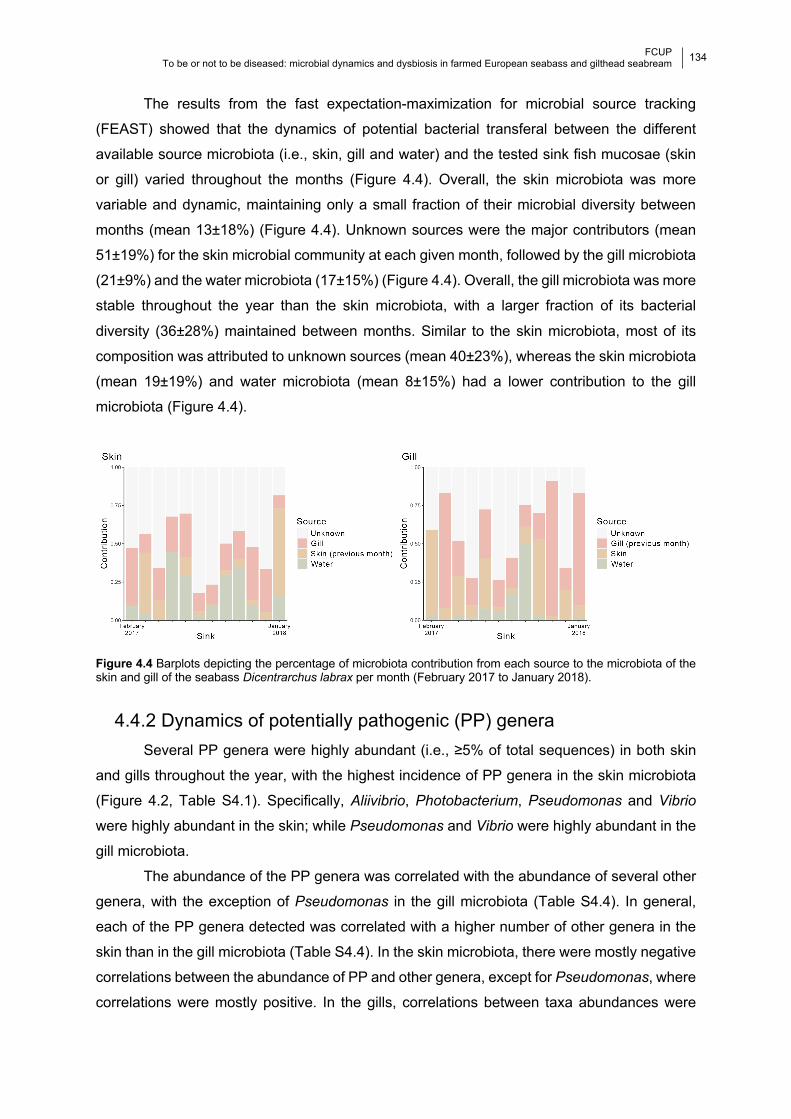
FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
134 133
The results from the fast expectation-maximization for microbial source tracking
(FEAST) showed that the dynamics of potential bacterial transferal between the different
available source microbiota (i.e., skin, gill and water) and the tested sink fish mucosae (skin
or gill) varied throughout the months (Figure 4.4). Overall, the skin microbiota was more
variable and dynamic, maintaining only a small fraction of their microbial diversity between
months (mean 13±18%) (Figure 4.4). Unknown sources were the major contributors (mean
51±19%) for the skin microbial community at each given month, followed by the gill microbiota
(21±9%) and the water microbiota (17±15%) (Figure 4.4). Overall, the gill microbiota was more
stable throughout the year than the skin microbiota, with a larger fraction of its bacterial
diversity (36±28%) maintained between months. Similar to the skin microbiota, most of its
composition was attributed to unknown sources (mean 40±23%), whereas the skin microbiota
(mean 19±19%) and water microbiota (mean 8±15%) had a lower contribution to the gill
microbiota (Figure 4.4).
Figure 4.4 Barplots depicting the percentage of microbiota contribution from each source to the microbiota of the skin and gill of the seabass Dicentrarchus labrax per month (February 2017 to January 2018).
4.4.2 Dynamics of potentially pathogenic (PP) genera Several PP genera were highly abundant (i.e., ≥5% of total sequences) in both skin
and gills throughout the year, with the highest incidence of PP genera in the skin microbiota
(Figure 4.2, Table S4.1). Specifically, Aliivibrio, Photobacterium, Pseudomonas and Vibrio
were highly abundant in the skin; while Pseudomonas and Vibrio were highly abundant in the
gill microbiota.
The abundance of the PP genera was correlated with the abundance of several other
genera, with the exception of Pseudomonas in the gill microbiota (Table S4.4). In general,
each of the PP genera detected was correlated with a higher number of other genera in the
skin than in the gill microbiota (Table S4.4). In the skin microbiota, there were mostly negative
correlations between the abundance of PP and other genera, except for Pseudomonas, where
correlations were mostly positive. In the gills, correlations between taxa abundances were
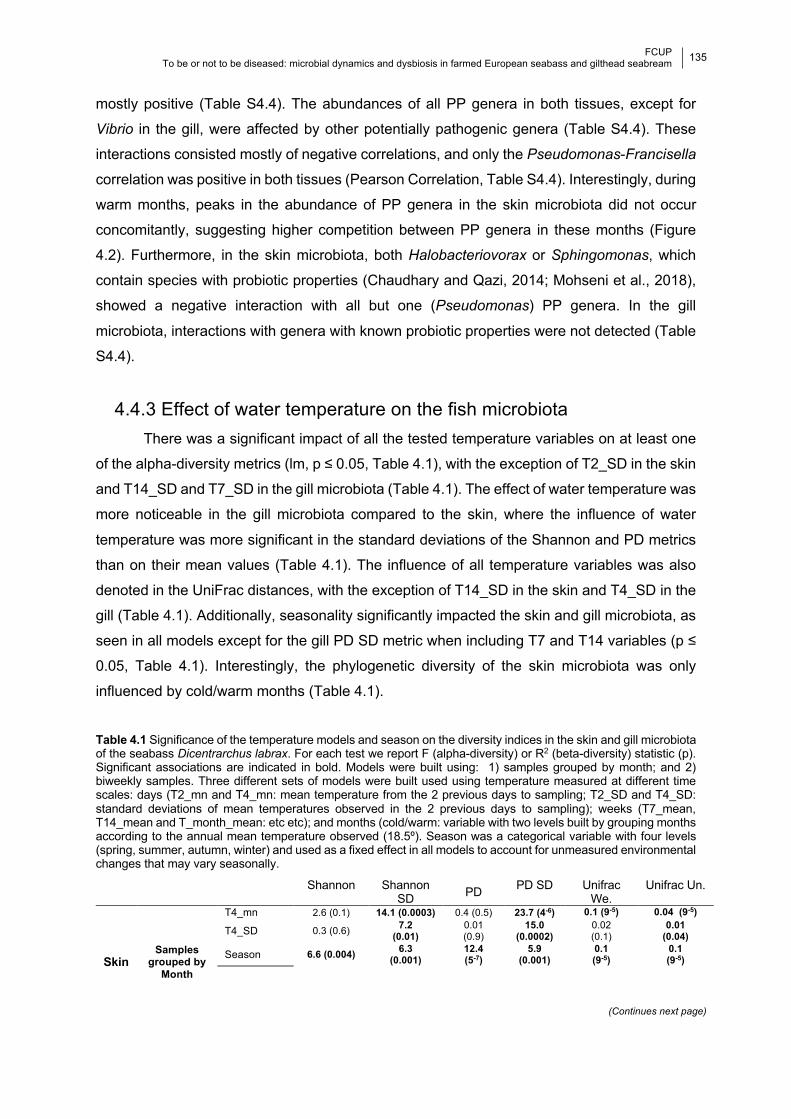
FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
135 133
mostly positive (Table S4.4). The abundances of all PP genera in both tissues, except for
Vibrio in the gill, were affected by other potentially pathogenic genera (Table S4.4). These
interactions consisted mostly of negative correlations, and only the Pseudomonas-Francisella
correlation was positive in both tissues (Pearson Correlation, Table S4.4). Interestingly, during
warm months, peaks in the abundance of PP genera in the skin microbiota did not occur
concomitantly, suggesting higher competition between PP genera in these months (Figure
4.2). Furthermore, in the skin microbiota, both Halobacteriovorax or Sphingomonas, which
contain species with probiotic properties (Chaudhary and Qazi, 2014; Mohseni et al., 2018),
showed a negative interaction with all but one (Pseudomonas) PP genera. In the gill
microbiota, interactions with genera with known probiotic properties were not detected (Table
S4.4).
4.4.3 Effect of water temperature on the fish microbiota There was a significant impact of all the tested temperature variables on at least one
of the alpha-diversity metrics (lm, p ≤ 0.05, Table 4.1), with the exception of T2_SD in the skin
and T14_SD and T7_SD in the gill microbiota (Table 4.1). The effect of water temperature was
more noticeable in the gill microbiota compared to the skin, where the influence of water
temperature was more significant in the standard deviations of the Shannon and PD metrics
than on their mean values (Table 4.1). The influence of all temperature variables was also
denoted in the UniFrac distances, with the exception of T14_SD in the skin and T4_SD in the
gill (Table 4.1). Additionally, seasonality significantly impacted the skin and gill microbiota, as
seen in all models except for the gill PD SD metric when including T7 and T14 variables (p ≤
0.05, Table 4.1). Interestingly, the phylogenetic diversity of the skin microbiota was only
influenced by cold/warm months (Table 4.1).
Table 4.1 Significance of the temperature models and season on the diversity indices in the skin and gill microbiota of the seabass Dicentrarchus labrax. For each test we report F (alpha-diversity) or R2 (beta-diversity) statistic (p). Significant associations are indicated in bold. Models were built using: 1) samples grouped by month; and 2) biweekly samples. Three different sets of models were built used using temperature measured at different time scales: days (T2_mn and T4_mn: mean temperature from the 2 previous days to sampling; T2_SD and T4_SD: standard deviations of mean temperatures observed in the 2 previous days to sampling); weeks (T7_mean, T14_mean and T_month_mean: etc etc); and months (cold/warm: variable with two levels built by grouping months according to the annual mean temperature observed (18.5º). Season was a categorical variable with four levels (spring, summer, autumn, winter) and used as a fixed effect in all models to account for unmeasured environmental changes that may vary seasonally.
Shannon Shannon SD PD PD SD Unifrac
We. Unifrac Un.
Skin Samples
grouped by Month
T4_mn 2.6 (0.1) 14.1 (0.0003) 0.4 (0.5) 23.7 (4-6) 0.1 (9-5) 0.04 (9-5)
T4_SD 0.3 (0.6) 7.2
(0.01) 0.01
(0.9) 15.0
(0.0002) 0.02
(0.1) 0.01
(0.04)
Season 6.6 (0.004) 6.3 (0.001)
12.4 (5-7)
5.9 (0.001)
0.1 (9-5)
0.1 (9-5)
(Continues next page)
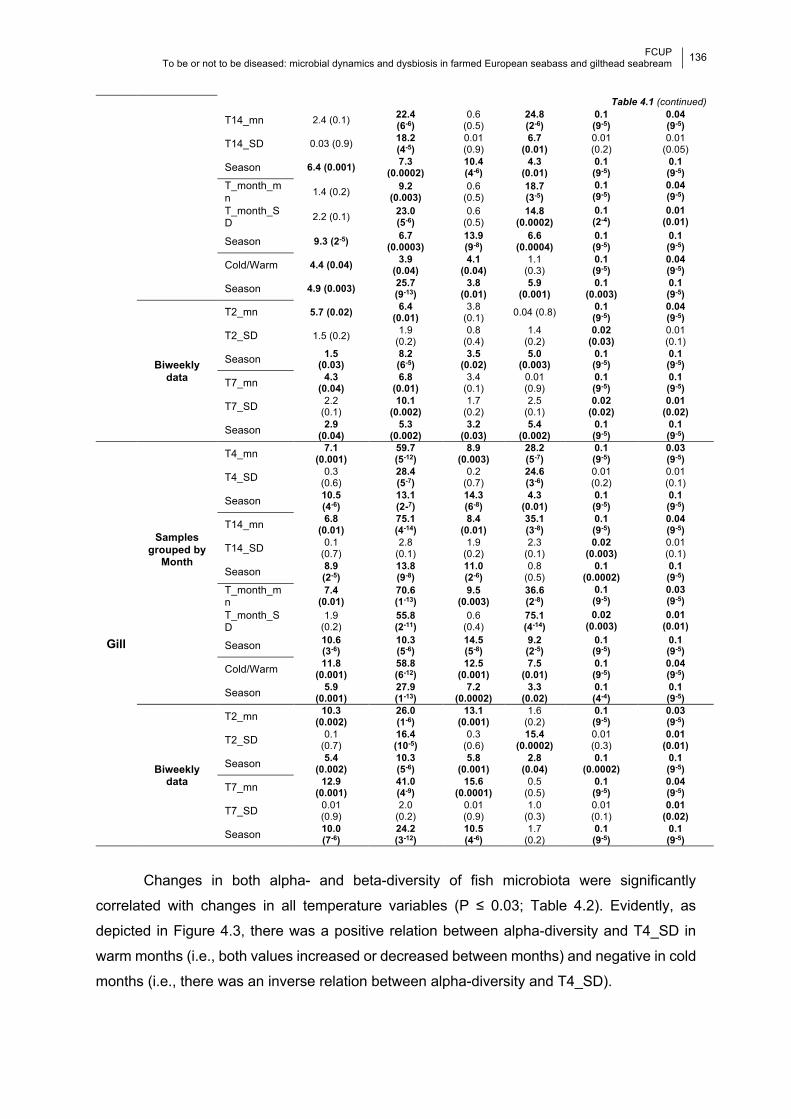
FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
136 133
Table 4.1 (continued)
T14_mn 2.4 (0.1) 22.4 (6-6)
0.6
(0.5) 24.8 (2-6)
0.1 (9-5)
0.04 (9-5)
T14_SD 0.03 (0.9) 18.2 (4-5)
0.01
(0.9)
6.7 (0.01)
0.01
(0.2)
0.01
(0.05)
Season 6.4 (0.001) 7.3 (0.0002)
10.4 (4-6)
4.3 (0.01)
0.1 (9-5)
0.1 (9-5)
T_month_mn 1.4 (0.2)
9.2 (0.003)
0.6
(0.5) 18.7 (3-5)
0.1 (9-5)
0.04 (9-5)
T_month_SD 2.2 (0.1)
23.0 (5-6)
0.6
(0.5) 14.8
(0.0002) 0.1 (2-4)
0.01 (0.01)
Season 9.3 (2-5) 6.7 (0.0003)
13.9 (9-8)
6.6 (0.0004)
0.1 (9-5)
0.1 (9-5)
Cold/Warm 4.4 (0.04) 3.9 (0.04)
4.1 (0.04)
1.1
(0.3) 0.1 (9-5)
0.04 (9-5)
Season 4.9 (0.003) 25.7 (9-13)
3.8 (0.01)
5.9 (0.001)
0.1 (0.003)
0.1 (9-5)
Biweekly data
T2_mn 5.7 (0.02) 6.4 (0.01)
3.8
(0.1) 0.04 (0.8) 0.1 (9-5)
0.04 (9-5)
T2_SD 1.5 (0.2) 1.9
(0.2) 0.8
(0.4) 1.4
(0.2) 0.02
(0.03) 0.01
(0.1)
Season 1.5 (0.03)
8.2 (6-5)
3.5 (0.02)
5.0 (0.003)
0.1 (9-5)
0.1 (9-5)
T7_mn 4.3 (0.04)
6.8 (0.01)
3.4
(0.1) 0.01
(0.9)
0.1 (9-5)
0.1 (9-5)
T7_SD 2.2
(0.1) 10.1
(0.002) 1.7
(0.2) 2.5
(0.1) 0.02
(0.02) 0.01
(0.02)
Season 2.9 (0.04)
5.3 (0.002)
3.2 (0.03)
5.4 (0.002)
0.1 (9-5)
0.1 (9-5)
Gill
Samples grouped by
Month
T4_mn 7.1 (0.001)
59.7 (5-12)
8.9 (0.003)
28.2 (5-7)
0.1 (9-5)
0.03 (9-5)
T4_SD 0.3
(0.6) 28.4 (5-7)
0.2
(0.7) 24.6 (3-6)
0.01
(0.2)
0.01
(0.1)
Season 10.5 (4-6)
13.1 (2-7)
14.3 (6-8)
4.3 (0.01)
0.1 (9-5)
0.1 (9-5)
T14_mn 6.8 (0.01)
75.1 (4-14)
8.4 (0.01)
35.1 (3-8)
0.1 (9-5)
0.04 (9-5)
T14_SD 0.1
(0.7) 2.8
(0.1) 1.9
(0.2)
2.3
(0.1)
0.02 (0.003)
0.01
(0.1)
Season 8.9 (2-5)
13.8 (9-8)
11.0 (2-6)
0.8
(0.5)
0.1 (0.0002)
0.1 (9-5)
T_month_mn
7.4 (0.01)
70.6 (1-13)
9.5 (0.003)
36.6 (2-8)
0.1 (9-5)
0.03 (9-5)
T_month_SD
1.9
(0.2) 55.8 (2-11)
0.6
(0.4) 75.1 (4-14)
0.02 (0.003)
0.01 (0.01)
Season 10.6 (3-6)
10.3 (5-6)
14.5 (5-8)
9.2 (2-5)
0.1 (9-5)
0.1 (9-5)
Cold/Warm 11.8
(0.001) 58.8 (6-12)
12.5 (0.001)
7.5 (0.01)
0.1 (9-5)
0.04 (9-5)
Season 5.9 (0.001)
27.9 (1-13)
7.2 (0.0002)
3.3 (0.02)
0.1 (4-4)
0.1 (9-5)
Biweekly data
T2_mn 10.3 (0.002)
26.0 (1-6)
13.1 (0.001)
1.6
(0.2) 0.1 (9-5)
0.03 (9-5)
T2_SD 0.1
(0.7) 16.4 (10-5)
0.3
(0.6) 15.4
(0.0002) 0.01
(0.3)
0.01 (0.01)
Season 5.4 (0.002)
10.3 (5-6)
5.8 (0.001)
2.8 (0.04)
0.1 (0.0002)
0.1 (9-5)
T7_mn 12.9 (0.001)
41.0 (4-9)
15.6 (0.0001)
0.5
(0.5)
0.1 (9-5)
0.04 (9-5)
T7_SD 0.01
(0.9)
2.0
(0.2)
0.01
(0.9)
1.0
(0.3)
0.01
(0.1)
0.01 (0.02)
Season 10.0 (7-6)
24.2 (3-12)
10.5 (4-6)
1.7
(0.2)
0.1 (9-5)
0.1 (9-5)
Changes in both alpha- and beta-diversity of fish microbiota were significantly
correlated with changes in all temperature variables (P ≤ 0.03; Table 4.2). Evidently, as
depicted in Figure 4.3, there was a positive relation between alpha-diversity and T4_SD in
warm months (i.e., both values increased or decreased between months) and negative in cold
months (i.e., there was an inverse relation between alpha-diversity and T4_SD).
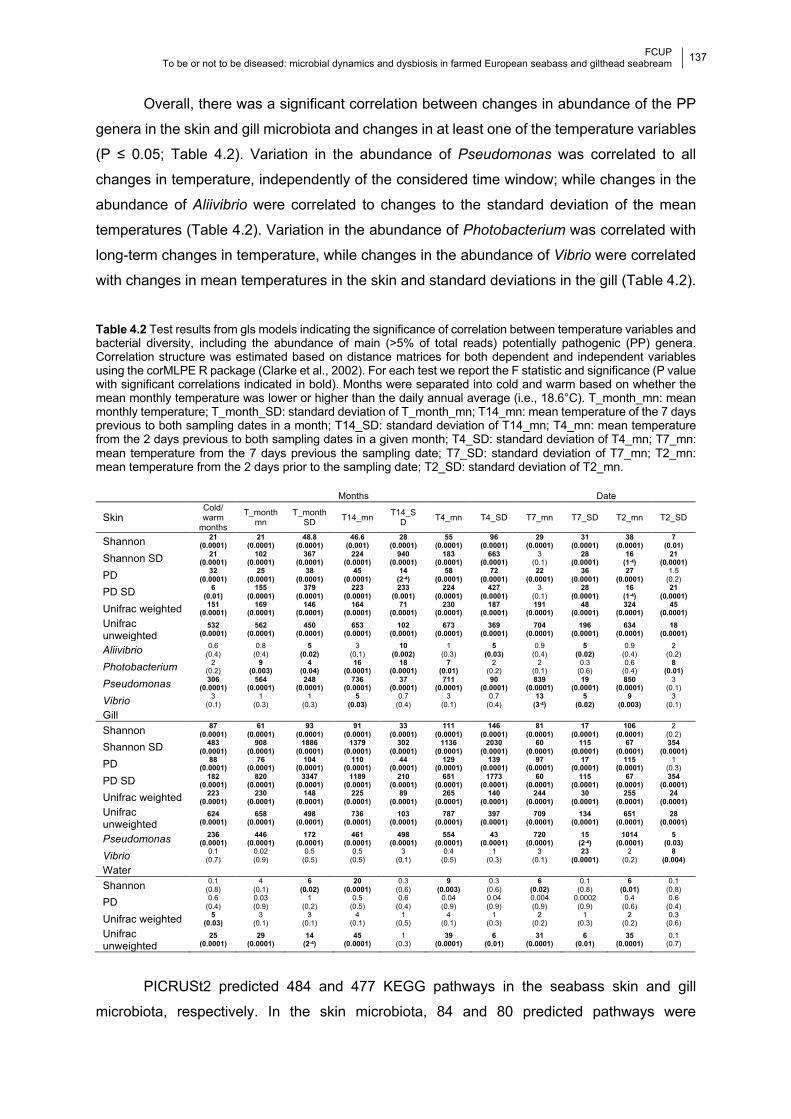
FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
137 133
Overall, there was a significant correlation between changes in abundance of the PP
genera in the skin and gill microbiota and changes in at least one of the temperature variables
(P ≤ 0.05; Table 4.2). Variation in the abundance of Pseudomonas was correlated to all
changes in temperature, independently of the considered time window; while changes in the
abundance of Aliivibrio were correlated to changes to the standard deviation of the mean
temperatures (Table 4.2). Variation in the abundance of Photobacterium was correlated with
long-term changes in temperature, while changes in the abundance of Vibrio were correlated
with changes in mean temperatures in the skin and standard deviations in the gill (Table 4.2).
Table 4.2 Test results from gls models indicating the significance of correlation between temperature variables and bacterial diversity, including the abundance of main (>5% of total reads) potentially pathogenic (PP) genera. Correlation structure was estimated based on distance matrices for both dependent and independent variables using the corMLPE R package (Clarke et al., 2002). For each test we report the F statistic and significance (P value with significant correlations indicated in bold). Months were separated into cold and warm based on whether the mean monthly temperature was lower or higher than the daily annual average (i.e., 18.6°C). T_month_mn: mean monthly temperature; T_month_SD: standard deviation of T_month_mn; T14_mn: mean temperature of the 7 days previous to both sampling dates in a month; T14_SD: standard deviation of T14_mn; T4_mn: mean temperature from the 2 days previous to both sampling dates in a given month; T4_SD: standard deviation of T4_mn; T7_mn: mean temperature from the 7 days previous the sampling date; T7_SD: standard deviation of T7_mn; T2_mn: mean temperature from the 2 days prior to the sampling date; T2_SD: standard deviation of T2_mn.
Months Date
Skin Cold/ warm
months
T_month mn
T_month SD T14_mn T14_S
D T4_mn T4_SD T7_mn T7_SD T2_mn T2_SD
Shannon 21 (0.0001)
21 (0.0001)
48.8 (0.0001)
46.6 (0.001)
28 (0.0001)
55 (0.0001)
96 (0.0001)
29 (0.0001)
31 (0.0001)
38 (0.0001)
7 (0.01)
Shannon SD 21 (0.0001)
102 (0.0001)
367 (0.0001)
224 (0.0001)
940 (0.0001)
183 (0.0001)
663 (0.0001)
3 (0.1)
28 (0.0001)
16 (1-4)
21 (0.0001)
PD 32 (0.0001)
25 (0.0001)
38 (0.0001)
45 (0.0001)
14 (2-4)
58 (0.0001)
72 (0.0001)
22 (0.0001)
36 (0.0001)
27 (0.0001)
1.5 (0.2)
PD SD 6 (0.01)
155 (0.0001)
379 (0.0001)
223 (0.0001)
233 (0.001)
224 (0.0001)
427 (0.0001)
3 (0.1)
28 (0.0001)
16 (1-4)
21 (0.0001)
Unifrac weighted 151 (0.0001)
169 (0.0001)
146 (0.0001)
164 (0.0001)
71 (0.0001)
230 (0.0001)
187 (0.0001)
191 (0.0001)
48 (0.0001)
324 (0.0001)
45 (0.0001)
Unifrac unweighted
532 (0.0001)
562 (0.0001)
450 (0.0001)
653 (0.0001)
102 (0.0001)
673 (0.0001)
369 (0.0001)
704 (0.0001)
196 (0.0001)
634 (0.0001)
18 (0.0001)
Aliivibrio 0.6 (0.4)
0.8 (0.4)
5 (0.02)
3 (0.1)
10 (0.002)
1 (0.3)
5 (0.03)
0.9 (0.4)
5 (0.02)
0.9 (0.4)
2 (0.2)
Photobacterium 2
(0.2) 9
(0.003) 4
(0.04) 16
(0.0001) 18
(0.0001) 7
(0.01) 2
(0.2) 2
(0.1) 0.3
(0.6) 0.6
(0.4) 8
(0.01)
Pseudomonas 306 (0.0001)
564 (0.0001)
248 (0.0001)
736 (0.0001)
37 (0.0001)
711 (0.0001)
90 (0.0001)
839 (0.0001)
19 (0.0001)
850 (0.0001)
3 (0.1)
Vibrio 3 (0.1)
1 (0.3)
1 (0.3)
5 (0.03)
0.7 (0.4)
3 (0.1)
0.7 (0.4)
13 (3-4)
5 (0.02)
9 (0.003)
3 (0.1)
Gill
Shannon 87 (0.0001)
61 (0.0001)
93 (0.0001)
91 (0.0001)
33 (0.0001)
111 (0.0001)
146 (0.0001)
81 (0.0001)
17 (0.0001)
106 (0.0001)
2 (0.2)
Shannon SD 483 (0.0001)
908 (0.0001)
1886 (0.0001)
1379 (0.0001)
302 (0.0001)
1136 (0.0001)
2030 (0.0001)
60 (0.0001)
115 (0.0001)
67 (0.0001)
354 (0.0001)
PD 88 (0.0001)
76 (0.0001)
104 (0.0001)
110 (0.0001)
44 (0.0001)
129 (0.0001)
139 (0.0001)
97 (0.0001)
17 (0.0001)
115 (0.0001)
1 (0.3)
PD SD 182 (0.0001)
820 (0.0001)
3347 (0.0001)
1189 (0.0001)
210 (0.0001)
651 (0.0001)
1773 (0.0001)
60 (0.0001)
115 (0.0001)
67 (0.0001)
354 (0.0001)
Unifrac weighted 223 (0.0001)
230 (0.0001)
148 (0.0001)
225 (0.0001)
89 (0.0001)
265 (0.0001)
140 (0.0001)
244 (0.0001)
30 (0.0001)
255 (0.0001)
24 (0.0001)
Unifrac unweighted
624 (0.0001)
658 (0.0001)
498 (0.0001)
736 (0.0001)
103 (0.0001)
787 (0.0001)
397 (0.0001)
709 (0.0001)
134 (0.0001)
651 (0.0001)
28 (0.0001)
Pseudomonas 236 (0.0001)
446 (0.0001)
172 (0.0001)
461 (0.0001)
498 (0.0001)
554 (0.0001)
43 (0.0001)
720 (0.0001)
15 (2-4)
1014 (0.0001)
5 (0.03)
Vibrio 0.1 (0.7)
0.02 (0.9)
0.5 (0.5)
0.5 (0.5)
3 (0.1)
0.4 (0.5)
1 (0.3)
3 (0.1)
23 (0.0001)
2 (0.2)
8 (0.004)
Water
Shannon 0.1 (0.8)
4 (0.1)
6 (0.02)
20 (0.0001)
0.3 (0.6)
9 (0.003)
0.3 (0.6)
6 (0.02)
0.1 (0.8)
6 (0.01)
0.1 (0.8)
PD 0.6 (0.4)
0.03 (0.9)
1 (0.2)
0.5 (0.5)
0.6 (0.4)
0.04 (0.9)
0.04 (0.9)
0.004 (0.9)
0.0002 (0.9)
0.4 (0.6)
0.6 (0.4)
Unifrac weighted 5 (0.03)
3 (0.1)
3 (0.1)
4 (0.1)
1 (0.5)
4 (0.1)
1 (0.3)
2 (0.2)
1 (0.3)
2 (0.2)
0.3 (0.6)
Unifrac unweighted
25 (0.0001)
29 (0.0001)
14 (2-4)
45 (0.0001)
1 (0.3)
39 (0.0001)
6 (0.01)
31 (0.0001)
6 (0.01)
35 (0.0001)
0.1 (0.7)
PICRUSt2 predicted 484 and 477 KEGG pathways in the seabass skin and gill
microbiota, respectively. In the skin microbiota, 84 and 80 predicted pathways were

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
138 133
differentially abundant in warm and cold months, respectively; while 70 and 78 predicted
pathways were differentially abundant in the gill microbiota during warm and cold months,
respectively (Table S4.5). Interestingly, most of the differently abundant potential pathways
were shared between the skin and gill microbiota in cold (51%) and warm (44%) months.
Predicted pathways related to biosynthesis, degradation/utilization/assimilation and
generation of precursor metabolite and energy were enriched in cold and warm months in the
microbiota of both mucosae (Figure 4.5, Table S4.5). Predicted pathways related to
detoxification and superpathways were only enriched in cold months in both mucosae. In the
skin microbiota, there was a decrease in the frequency of predicted pathways related to
biosynthesis and an increase in degradation/utilization/assimilation related pathways from
warm to cold months (76% to 47% and 13% to 43%, respectively) (Figure 4.5, Table S4.5).
The microbiota of both tissues showed a decrease in the frequency of predicted pathways
related to generation of precursor metabolite and energy from warm to cold months (10% to
5% in the skin; 12% to 1% in the gill) (Figure 4.5, Table S4.5).
Figure 4.5 Relative frequency of the differentially enriched potential pathways in the skin and gill microbiota of the seabass Dicentrarchus labrax during cold and warm months grouped by broader (A) or smaller (B) KEGG categories. Circles are coloured according to broader categories.
At a finer scale, the microbiota of both tissues presented a decrease in frequency of
potential pathways related to amino acid biosynthesis, cell structure biosynthesis, nucleoside

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
139 133
and nucleotide biosynthesis, and TCA cycle from warm to cold months (Figure 4.5, Table
S4.5). In contrast, potential pathways related to fatty acid and lipid biosynthesis and aromatic
compound degradation increased in frequency from warm to cold months in both tissues. In
the microbiota of both tissues, amine and polyamine degradation, C1 compound utilization
and assimilation and pentose phosphate predicted pathways were only enriched in warm
months; whereas secondary metabolite degradation, antibiotic resistance and the super
pathway of histidine, purine, and pyrimidine biosynthesis predicted pathway categories were
only enriched in the cold months (Figure 4.5, Table S4.5).
4.5 Discussion The skin and gill microbiota of the farmed European seabass were highly dynamic,
being significantly affected by water temperature. Although previous studies reported an effect
of water temperature on fish microbiota, they focused on testing the effects of single
temperatures (Horlick et al., 2020; Kokou et al., 2018; Sugita et al., 1989) or established an
indirect link with temperature based on different geographic locations (Krotman et al., 2020;
Vasemägi et al., 2017). To the best of our knowledge, the only other longitudinal study which
specifically tested the effects of natural fluctuations of sea water temperatures on fish
microbiota was Minich et al.'s (2020) on chub mackerel. In that study, although temperature
significantly affected the skin and gill microbial composition on each sampling date, an effect
on beta-diversity was less clear (Minich et al., 2020). Our results show that mean water
temperature and associated standard deviations occurring at different time scales,
significantly affected the composition, structure and function of the skin and gill microbiota of
the European seabass Dicentrarchus labrax. Additionally, temperature seems to be a factor
triggering dysbiosis.
4.5.1 Temporal dynamics of the microbiota The skin and gill microbiota of farmed European seabass varied significantly over a
12 month period, particularly at lower taxonomic levels (most abundant genera and core
ASVs), while maintaining phylogenetic relatedness at higher taxonomic levels (most abundant
phyla). Similar long-term patterns have been reported in the microbiota of other teleost
species, including the Atlantic charr (gut; Element et al., 2020) and tench (gut; Dulski et al.,
2020), gulf killifish (skin; Larsen et al., 2015) and the largemouth bass, bluegill and spotted
gar (skin and gut; Arias et al., 2019).
The skin and gill microbiomes of healthy European seabass can be very distinct
(Rosado et al., 2019a), responding asymmetrically to disease (photobacteriousis) and with
different recovery trajectories after antibiotic treatment (Rosado et al., 2019b). Here, despite

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
140 133
such intrinsic differences, the overall bacterial composition of both mucosae followed the same
trends, i.e., concomitantly increasing or decreasing in diversity throughout the year. This
suggests they are likely modulated by the same host and/or environmental factors.
Furthermore, our analysis showed that the dynamics of the skin and gill microbiota are
complex, with only a small portion of the microbiota remaining stable over time (ca. 13% in the
skin and ca. 36% in the gill on average over the 12 months). Skin and particularly the gills
seem to be highly selective habitats, and bacterial variation did not seem to arise due to
substantial outsourcing of bacteria from the water column. This result is in line with previous
studies where water microbiota was shown to have a minor influence on the mucosal
microbiota of adult fish (Bledsoe et al., 2016; Chiarello et al., 2015; Legrand et al., 2018;
Llewellyn et al., 2016; Pratte et al., 2018; Rosado et al., 2021). Instead, bacterial recruitment
from the neighbouring environments seems to be more important. These results suggest that
there is a continuous microbial exchange of communities in the skin and gill, most likely due
to constant abiotic changes in the water environment (e.g., temperature) prompting complex
microbial dynamics.
The high frequency of dysbiotic events, coupled with the dynamics observed in the
abundances of potentially pathogenic (PP) genera in both fish mucosae, seems to be in line
with what has been reported in farmed fish, where stress is often associated with a decrease
in microbial diversity and increased susceptibility to secondary bacterial opportunists (Kelly
and Salinas, 2017; Legrand et al., 2018; Silva et al., 2020). Furthermore, the negative
correlation observed between abundance of PP genera and genera with probiotic properties
confirms direct competition between taxa has an important role in controlling the abundance
of PP genera and potential disease outcome (Legrand et al., 2020). Specifically,
Halobacteriovorax and Sphingomonas had a negative impact on the abundance of Aliivibrio
and Vibrio or Photobacterium, respectively, in the skin microbiota. The probiotic properties of
Halobacteriovorax were previously demonstrated against Vibrio parahaemolyticus in mussels
(Ottaviani et al., 2020). Additionally, Sphingomonas species were described as having
antagonistic properties against the fish pathogen Vibrio anguillarum in roho fingerlings
(Chaudhary and Qazi, 2014).
4.5.2 Water temperature effects in the diversity of fish microbiota Here we show that seasonal patterns, which influence physio-chemical properties of
water and are highly influenced by temperature, contribute significantly to the high variability
of farmed European seabass microbiota composition and structure over a year. Seasonal
changes in the composition and structure of fish microbiota have been reported for the skin
and gut (e.g., Arias et al., 2019; Dulski et al., 2020). However, such studies are commonly

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
141 133
based on sparsely sampled timepoints (e.g., May, August and November, Arias et al., 2019;
Autumn 2017 and Summer 2018, Dulski et al., 2020). Additionally, the reported effect of
season in the microbiomes of wild fish is mostly linked to differences in prey availability (e.g.,
Ringø et al., 2016) or water properties like chlorophyll a, salinity and temperature (e.g., Minich
et al., 2020). Understanding seasonal variations in fish microbiota can be very important in an
aquaculture context, for example to understand disease dynamics and outbreaks. However,
the lack of longitudinal studies is still hampering such insights.
The bacterial composition and structure of farmed European seabass microbiota were
significantly affected by changes in temperature occurring over both short (two days) and
longer (several months) time frames. This is not surprising given the high sensitivity of bacteria
to temperature (Corkrey et al., 2012). Microbial shifts occurring at small temporal scales, from
just a few hours to days are not unprecedented for fish. Such events occurred, for example,
after cold temperature shock (tropical tilapia, Kokou et al., 2018), acclimation (e.g., to the wild
in the common snook, Tarnecki et al., 2019; from freshwater to seawater in the Atlantic
salmon, Dehler et al., 2017); chemical exposure (e.g., in zebrafish, Gaulke et al., 2016;
channel catfish, Mohammed and Arias, 2015) or parasitism (e.g., in Atlantic salmon, Llewellyn
et al., 2017). In the present work, temperature was a significant factor influencing high variation
in bacterial composition within samples. Differences in bacterial diversity measures between
individuals were significantly correlated with the difference in temperatures experienced by
individuals across short and long time frames. In light of these results, we suggest that
changes in water temperature played a major role in the high temporal variability observed in
the microbiota of several teleost species (e.g., Larsen et al., 2015; Pratte et al., 2018; Uren
Webster et al., 2018), including the European seabass (Rosado et al., 2021, 2019a).
Another emerging trend in our analysis was that the microbiota of the European seabass
was more dynamic during the warmer months and more prone to dysbiosis during cold/warm
transition periods. Microbial diversity significantly decreased on three occasions during the
sampled year, signaling bacterial imbalance. During cold/warm transitions, decreases in
diversity were accompanied by an increase in the abundance of PP genera (i.e., Alivibrio,
Photobacterium or Vibrio), highlighting the opportunistic nature of these bacteria. Seasonal
occurrence of Vibriosis and Photobacteriosis in fish was previously observed (e.g., Bellos et
al., 2015; Habiba et al., 2015), with higher abundances of Vibrio and Photobacterium reported
during warmer months, even if fish remained asymptomatic (e.g., Horlick et al., 2020; Minich
et al., 2020; Sugita et al., 1989). On the other hand, higher incidence of severe outbreaks of
both Aliivibrio (e.g., Guijarro et al., 2015; Khider et al., 2018) and Pseudomonas (e.g., Huang
et al., 2019; Tao et al., 2016) species usually occur at lower temperatures, explaining the high
abundances of these PP genera in cold months. Temperature ranges observed here during
cold/warm transition periods (15-20°C) are known to impact European sea bass homeostasis,

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
142 133
reducing adaptive immune response and increasing susceptibility to disease (17-23 °C;
Varsamos et al., 2006). Moreover, the adaptive immune response of European seabass is
intrinsically linked to temperature, being delayed at low temperatures (≤18°C) and stimulated
at higher temperatures (≥24°C) (Cecchini and Saroglia, 2002; Varsamos et al., 2006). It is
possible that the cumulative effect of temperature changes occurring at cold/warm transition
periods and changes in host homeostasis could have prompted dysbiosis. Nevertheless,
dysbiotic events did not lead to visible signs of disease. Aspects of fish immunity are also
correlated with age; for example, in rainbow trout, resistance to bacterial infections increases
with host age (Castro et al., 2015). Since the European seabass included in our study were
adults (26 to 32 months), age might explain their apparent resilience to PP genera upgrowth.
4.5.3 Water temperature effects in the predicted microbiota function Changes in microbial composition and structure between cold/warm periods were
accompanied by significant changes in the predicted metabolic function of bacteria. It is
important to note that these results should be interpreted with caution since PICRUSt2
analysis is biased towards human health-related microorganisms (Choi et al., 2017).
Nevertheless, differentially enriched predicted metabolic pathways occurring in the skin and
gill microbiota of the European seabass during warm periods were mostly related to
biosynthesis processes, indicating high energetic expenditure. In particular, predicted
pathways related to amino acid, and nucleoside and nucleotide biosynthesis are considered
essential for bacterial growth (Samant et al., 2008), and were strongly enriched in both
mucosae during this period. Similarly, more predicted pathways related to the generation of
precursor metabolites and energy were enriched in both fish mucosae during warm months.
During the cold period there was a shift, particularly in the skin, where biosynthesis and
metabolic energy generation were reduced and degradation/utilization/assimilation predicted
pathways increased by almost 4-fold. In the gill microbiota, changes between differentially
expressed predicted metabolic pathways were less evident, also pointing to a more stable
microbiota. This suggests that broad time scale temperature shifts elicit microbial composition
changes in order to harbour species with different predicted metabolic functions. Finally, it is
worth noting that antimicrobial resistance (a detoxification mechanism) was significantly
enriched in both tissues only in cold months. Suboptimal temperature conditions can favour
the fitness of antibiotic resistant bacterial strains; for example, non-optimal temperature can
increase the fitness of antibiotic resistant Escherichia coli (see Trindade et al., 2012). Similarly,
increased salinity and decreased pH can select for antibiotic resistant phenotypes in several
bacterial food-related pathogens (McMahon et al., 2007).

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
143 133
4.6 Conclusion Changes in water temperature have a major impact on teleosts, leading to altered
physiology and behaviour (Alfonso et al., 2020; Almeida et al., 2015; Islam et al., 2020a,
2020b), contributing to parasitism resistance and tolerance (Jackson et al., 2020). We
demonstrate here that water temperature oscillation also impacts the composition, structure
and function of the skin and gill microbiota of farmed European seabass. In particular,
dysbiosis seems to be more frequent in warm months and during cold/warm transitions, and
the abundance of several PP genera is also affected by changes in water temperature.
Furthermore, the microbiota of fish external mucosae behaved in similar ways with regards to
their compositional diversity, suggesting they may be modulated by similar host and
environmental stressors. These results highlight the need for further longitudinal studies
examining the full thermal spectrum experienced by a species, as well as other environmental
factors.
4.7 Acknowledgements This work was funded by the European Regional Development Fund (ERDF) through the
COMPETE program and by National Funds through FCT - Foundation for Science and
Technology (project PTDC/BIA-MIC/27995/2017 POCI-01-0145- FEDER-027995); DR, MP-L
and RX were supported by FCT under the Programa Operacional Potencial Humano – Quadro
de Referência Estratégico Nacional funds from the European Social Fund and Portuguese
Ministério da Educação e Ciência (DR doctoral grant SFRH/ BD/117943/2016; MP-L:
IF/00764/2013; RX: IF/00359/2015; and 2020.00854.CEECIND; PT:
DL57/2016/CP1440/CT0008).
4.8 References Alfonso, S., Gesto, M., Sadoul, B., 2020. Temperature increase and its effects on fish stress
physiology in the context of global warming. J. Fish Biol. jfb.14599.
https://doi.org/10.1111/jfb.14599
Almeida, J.R., Gravato, C., Guilhermino, L., 2015. Effects of Temperature in Juvenile Seabass
(Dicentrarchus labrax L.) Biomarker Responses and Behaviour: Implications for
Environmental Monitoring. Estuaries and Coasts 38, 45–55.
https://doi.org/10.1007/s12237-014-9792-7
Anderson, M.J., 2017. Permutational Multivariate Analysis of Variance (PERMANOVA), in:
Wiley StatsRef: Statistics Reference Online. John Wiley & Sons, Ltd, Chichester, UK, pp.
1–15. https://doi.org/10.1002/9781118445112.stat07841
Arias, C.R., Ray, C.L., Cai, W., Willmon, E., 2019. Fish are not alone: characterization of the

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
144 133
gut and skin microbiomes of Largemouth Bass (Micropterus salmoides), Bluegill
(Lepomis macrochirus), and Spotted Gar (Lepisosteus oculatus). J. Aquac. Fish. Fish
Sci. 2. https://doi.org/10.25177/JAFFS.2.2.RA.459
Baker-Austin, C., Oliver, J.D., Alam, M., Ali, A., Waldor, M.K., Qadri, F., Martinez-Urtaza, J.,
2018. Vibrio spp. infections. Nat. Rev. Dis. Prim. 4, 1–19. https://doi.org/10.1038/s41572-
018-0005-8
Baker-Austin, C., Trinanes, J., Gonzalez-Escalona, N., Martinez-Urtaza, J., 2017. Non-
Cholera Vibrios: The Microbial Barometer of Climate Change. Trends Microbiol.
https://doi.org/10.1016/j.tim.2016.09.008
Baker-Austin, C., Trinanes, J.A., Taylor, N.G.H., Hartnell, R., Siitonen, A., Martinez-Urtaza, J.,
2013. Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim.
Chang. 3, 73–77. https://doi.org/10.1038/nclimate1628
Baksi, K.D., Kuntal, B.K., Mande, S.S., 2018. ‘TIME’: A Web Application for Obtaining Insights
into Microbial Ecology Using Longitudinal Microbiome Data. Front. Microbiol. 9, 9–36.
https://doi.org/10.3389/fmicb.2018.00036
Beck, B.H., Peatman, E., 2015. Mucosal health in aquaculture. Academic Press, UK.
Bellos, G., Angelidis, P., Miliou, H., 2015. Effect of temperature and seasonality principal
epizootiological risk factor on vibriosis and photobacteriosis outbreaks for European sea
bass in Greece (1998-2013). ournal Aquac. Res. Dev. 6.
Bledsoe, J.W., Peterson, B.C., Swanson, K.S., Small, B.C., 2016. Ontogenetic
Characterization of the Intestinal Microbiota of Channel Catfish through 16S rRNA Gene
Sequencing Reveals Insights on Temporal Shifts and the Influence of Environmental
Microbes. PLoS One 11, e0166379. https://doi.org/10.1371/journal.pone.0166379
Boutin, S., Bernatchez, L., Audet, C., Derôme, N., 2013. Network Analysis Highlights Complex
Interactions between Pathogen, Host and Commensal Microbiota. PLoS One 8, e84772.
https://doi.org/10.1371/journal.pone.0084772
Califano, G., Castanho, S., Soares, F., Ribeiro, L., Cox, C.J., Mata, L., Costa, R., 2017.
Molecular Taxonomic Profiling of Bacterial Communities in a Gilthead Seabream (Sparus
aurata) Hatchery. Front. Microbiol. 8, 204. https://doi.org/10.3389/fmicb.2017.00204
Callahan, B.J., McMurdie, P.J., Rosen, M.J., Han, A.W., Johnson, A.J.A., Holmes, S.P., 2016.
DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13,
581–583. https://doi.org/10.1038/nmeth.3869
Castro, R., Jouneau, L., Tacchi, L., Macqueen, D.J., Alzaid, A., Secombes, C.J., Martin,
S.A.M., Boudinot, P., 2015. Disparate developmental patterns of immune responses to
bacterial and viral infections in fish. Sci. Rep. 5, 1–13. https://doi.org/10.1038/srep15458
Cecchini, S., Saroglia, M., 2002. Antibody response in sea bass (Dicentrarchus labrax L.) in
relation to water temperature and oxygenation. Aquac. Res. 33, 607–613.

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
145 133
https://doi.org/10.1046/j.1365-2109.2002.00698.x
Chaudhary, A., Qazi, J.I., 2014. Probiotic Antagonism of Sphingomonas sp. against Vibrio
anguillarum Exposed Labeo rohita Fingerlings. Adv. Life Sci. 2014, 156–165.
https://doi.org/10.5923/j.als.20140403.11
Chen, Q., Yan, Q., Wang, K., Zhuang, Z., Wang, X., 2008. Portal of entry for pathogenic Vibrio
alginolyticus into large yellow croaker Pseudosciaena crocea, and characteristics of
bacterial adhesion to mucus. Dis. Aquat. Organ. 80, 181–188.
https://doi.org/10.3354/dao01933
Chiarello, M., Villéger, S., Bouvier, C., Bettarel, Y., Bouvier, T., 2015. High diversity of skin-
associated bacterial communities of marine fishes is promoted by their high variability
among body parts, individuals and species. FEMS Microbiol. Ecol. 91.
https://doi.org/10.1093/femsec/fiv061
Choi, J., Yang, F., Stepanauskas, R., Cardenas, E., Garoutte, A., Williams, R., Flater, J.,
Tiedje, J.M., Hofmockel, K.S., Gelder, B., Howe, A., 2017. Strategies to improve
reference databases for soil microbiomes. ISME J. 11, 829–834.
https://doi.org/10.1038/ismej.2016.168
Corkrey, R., Olley, J., Ratkowsky, D., McMeekin, T., Ross, T., 2012. Universality of
thermodynamic constants governing biological growth rates. PLoS One 7, e32003.
https://doi.org/10.1371/journal.pone.0032003
Dash, S., Das, S.K., Samal, J., Thatoi, H.N., 2018. Epidermal mucus, a major determinant in
fish health: A review. Iran. J. Vet. Res. https://doi.org/10.22099/ijvr.2018.4849
Dehler, C.E., Secombes, C.J., Martin, S.A.M., 2017. Seawater transfer alters the intestinal
microbiota profiles of Atlantic salmon (Salmo salar L.). Sci. Rep. 7, 1–11.
https://doi.org/10.1038/s41598-017-13249-8
Douglas, G.M., Maffei, V.J., Zaneveld, J., Yurgel, S.N., Brown, J.R., Taylor, C.M.,
Huttenhower, C., Langille, M.G.I., 2019. PICRUSt2: An improved and extensible
approach for metagenome inference. bioRxiv. https://doi.org/10.1101/672295
Duarte, L.N., Coelho, F.J.R.C., Cleary, D.F.R., Bonifácio, D., Martins, P., Gomes, N.C.M.,
2019. Bacterial and microeukaryotic plankton communities in a semi-intensive
aquaculture system of sea bass (Dicentrarchus labrax): A seasonal survey. Aquaculture
503, 59–69. https://doi.org/10.1016/j.aquaculture.2018.12.066
Dulski, T., Kozłowski, K., Ciesielski, S., 2020. Habitat and seasonality shape the structure of
tench (Tinca tinca L.) gut microbiome. Sci. Rep. 10, 1–11.
https://doi.org/10.1038/s41598-020-61351-1
Eissa, I.A.M., Derwa, H.I., Ismail, M., El-lamie, M., Dessouki, A.A., Elsheshtawy, H., Bayoumy,
E.M., 2018. Molecular and phenotypic characterization of Photobacterium damselae
among some marine fishes in Lake Temsah. Microb. Pathog. 114, 315–322.

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
146 133
https://doi.org/10.1016/j.micpath.2017.12.006
Element, G., Engel, K., Neufeld, J.D., Casselman, J.M., Coeverden de Groot, P., Greer, C.W.,
Walker, V.K., 2020. Seasonal habitat drives intestinal microbiome composition in
anadromous Arctic char (Salvelinus alpinus). Environ. Microbiol. 22, 3112–3125.
https://doi.org/10.1111/1462-2920.15049
FAO, 2016. FAO Fisheries & Aquaculture - Cultured Aquatic Species Information Programme
- Dicentrarchus labrax (Linnaeus, 1758) [WWW Document]. URL
http://www.fao.org/fishery/culturedspecies/Dicentrarchus_labrax/en (accessed 2.12.21).
Gaulke, C.A., Barton, C.L., Proffitt, S., Tanguay, R.L., Sharpton, T.J., 2016. Triclosan
Exposure Is Associated with Rapid Restructuring of the Microbiome in Adult Zebrafish.
PLoS One 11, e0154632. https://doi.org/10.1371/journal.pone.0154632
Gomez, D., Sunyer, J.O., Salinas, I., 2013. The mucosal immune system of fish: The evolution
of tolerating commensals while fighting pathogens. Fish Shellfish Immunol.
https://doi.org/10.1016/j.fsi.2013.09.032
Guijarro, J.A., Cascales, D., García-Torrico, A.I., García-Domínguez, M., Méndez, J., 2015.
Temperature-dependent expression of virulence genes in fish-pathogenic bacteria.
Front. Microbiol. https://doi.org/10.3389/fmicb.2015.00700
Habiba, M.E., Tayel, S.I., Torky, H.A., 2015. Occurrence and Characterization of
Photobacteria damselae in Cultured Marine Fish. Alexandria J. Vet. Sci. 44, 103–110.
https://doi.org/10.5455/ajvs.129
Haenen, O., 2017. Major bacterial diseases affecting aquaculture, Aquatic AMR Workshop.
Mangalore, India.
Horlick, J., Booth, M.A., Tetu, S.G., 2020. Alternative dietary protein and water temperature
influence the skin and gut microbial communities of yellowtail kingfish (Seriola lalandi).
PeerJ 2020, e8705. https://doi.org/10.7717/peerj.8705
Huang, L., Zuo, Y., Jiang, Q., Su, Y., Qin, Y., Xu, X., Zhao, L., Yan, Q., 2019. A metabolomic
investigation into the temperature-dependent virulence of Pseudomonas plecoglossicida
from large yellow croaker (Pseudosciaena crocea). J. Fish Dis. 42, 431–446.
https://doi.org/10.1111/jfd.12957
Islam, M.J., Kunzmann, A., Bögner, M., Meyer, A., Thiele, R., James Slater, M., 2020a.
Metabolic and molecular stress responses of European seabass, Dicentrarchus labrax at
low and high temperature extremes. Ecol. Indic. 112, 106118.
https://doi.org/10.1016/j.ecolind.2020.106118
Islam, M.J., Slater, M.J., Bögner, M., Zeytin, S., Kunzmann, A., 2020b. Extreme ambient
temperature effects in European seabass, Dicentrarchus labrax: Growth performance
and hemato-biochemical parameters. Aquaculture 522, 735093.
https://doi.org/10.1016/j.aquaculture.2020.735093

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
147 133
Jackson, J.A., Friberg, I.M., Hablützel, P.I., Masud, N., Stewart, A., Synnott, R., Cable, J.,
2020. Partitioning the environmental drivers of immunocompetence. Sci. Total Environ.
747, 141152. https://doi.org/10.1016/j.scitotenv.2020.141152
Kanehisa, M., Sato, Y., Furumichi, M., Morishima, K., Tanabe, M., 2019. New approach for
understanding genome variations in KEGG. Nucleic Acids Res. 47, D590–D595.
https://doi.org/10.1093/nar/gky962
Kayansamruaj, P., Pirarat, N., Hirono, I., Rodkhum, C., 2014. Increasing of temperature
induces pathogenicity of Streptococcus agalactiae and the up-regulation of inflammatory
related genes in infected Nile tilapia (Oreochromis niloticus). Vet. Microbiol. 172, 265–
271. https://doi.org/10.1016/j.vetmic.2014.04.013
Kelly, C., Salinas, I., 2017. Under pressure: Interactions between commensal microbiota and
the teleost immune system. Front. Immunol. https://doi.org/10.3389/fimmu.2017.00559
Khider, M., Willassen, N.P., Hansen, H., 2018. The alternative sigma factor RpoQ regulates
colony morphology, biofilm formation and motility in the fish pathogen Aliivibrio
salmonicida. BMC Microbiol. 18, 1–13. https://doi.org/10.1186/s12866-018-1258-9
Kokou, F., Sasson, G., Nitzan, T., Doron-Faigenboim, A., Harpaz, S., Cnaani, A., Mizrahi, I.,
2018. Host genetic selection for cold tolerance shapes microbiome composition and
modulates its response to temperature. Elife 7. https://doi.org/10.7554/eLife.36398
Kozich, J.J., Westcott, S.L., Baxter, N.T., Highlander, S.K., Schloss, P.D., 2013. Development
of a dual-index sequencing strategy and curation pipeline for analyzing amplicon
sequence data on the miseq illumina sequencing platform. Appl. Environ. Microbiol. 79,
5112–5120. https://doi.org/10.1128/AEM.01043-13
Krotman, Y., Yergaliyev, T.M., Alexander Shani, R., Avrahami, Y., Szitenberg, A., 2020.
Dissecting the factors shaping fish skin microbiomes in a heterogeneous inland water
system. Microbiome 8, 9. https://doi.org/10.1186/s40168-020-0784-5
Kruskal, W.H., Wallis, W.A., 1952. Use of Ranks in One-Criterion Variance Analysis. J. Am.
Stat. Assoc. 47, 583–621. https://doi.org/10.1080/01621459.1952.10483441
Larsen, A.M., Bullard, S.A., Womble, M., Arias, C.R., 2015. Community Structure of Skin
Microbiome of Gulf Killifish, Fundulus grandis, Is Driven by Seasonality and Not Exposure
to Oiled Sediments in a Louisiana Salt Marsh. Microb. Ecol. 70, 534–544.
https://doi.org/10.1007/s00248-015-0578-7
Legrand, T.P.R.A., Catalano, S.R., Wos-Oxley, M.L., Stephens, F., Landos, M., Bansemer,
M.S., Stone, D.A.J., Qin, J.G., Oxley, A.P.A., 2018. The Inner Workings of the Outer
Surface: Skin and Gill Microbiota as Indicators of Changing Gut Health in Yellowtail
Kingfish. Front. Microbiol. 8, 2664. https://doi.org/10.3389/fmicb.2017.02664
Legrand, T.P.R.A., Wynne, J.W., Weyrich, L.S., Oxley, A.P.A., 2020. A microbial sea of
possibilities: current knowledge and prospects for an improved understanding of the fish

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
148 133
microbiome. Rev. Aquac. 12, 1101–1134. https://doi.org/10.1111/raq.12375
Llewellyn, M.S., Leadbeater, S., Garcia, C., Sylvain, F.E., Custodio, M., Ang, K.P., Powell, F.,
Carvalho, G.R., Creer, S., Elliot, J., Derome, N., 2017. Parasitism perturbs the mucosal
microbiome of Atlantic Salmon. Sci. Rep. 7, 1–10. https://doi.org/10.1038/srep43465
Llewellyn, M.S., McGinnity, P., Dionne, M., Letourneau, J., Thonier, F., Carvalho, G.R., Creer,
S., Derome, N., 2016. The biogeography of the atlantic salmon (Salmo salar) gut
microbiome. ISME J. 10, 1280–1284. https://doi.org/10.1038/ismej.2015.189
Martinez Arbizu, P., 2017. pairwiseAdonis: Pairwise multilevel comparison using adonis.
Martins, P., Coelho, F.J.R.C., Cleary, D.F.R., Pires, A.C.C., Marques, B., Rodrigues, A.M.,
Quintino, V., Gomes, N.C.M., 2018. Seasonal patterns of bacterioplankton composition
in a semi-intensive European seabass (Dicentrarchus labrax) aquaculture system.
Aquaculture 490, 240–250. https://doi.org/10.1016/j.aquaculture.2018.02.038
Matanza, X.M., Osorio, C.R., 2018. Transcriptome changes in response to temperature in the
fish pathogen Photobacterium damselae subsp. damselae: Clues to understand the
emergence of disease outbreaks at increased seawater temperatures. PLoS One 13,
e0210118. https://doi.org/10.1371/journal.pone.0210118
McMahon, M.A.S., Xu, J., Moore, J.E., Blair, I.S., McDowell, D.A., 2007. Environmental stress
and antibiotic resistance in food-related pathogens. Appl. Environ. Microbiol. 73, 211–
217. https://doi.org/10.1128/AEM.00578-06
McMurdie, P.J., Holmes, S., 2013. phyloseq: An R Package for Reproducible Interactive
Analysis and Graphics of Microbiome Census Data. PLoS One 8, e61217.
https://doi.org/10.1371/journal.pone.0061217
Minich, J., Petrus, S., Michael, J.D., Michael, T.P., Knight, R., Allen, E.E., 2020. Temporal,
Environmental, and Biological Drivers of the Mucosal Microbiome in a Wild Marine Fish,
Scomber japonicus. Msphere. 5. https://doi.org/10.1128/mSphere.00401-20
Mohammed, H.H., Arias, C.R., 2015. Potassium permanganate elicits a shift of the external
fish microbiome and increases host susceptibility to columnaris disease. Vet. Res. 46,
82. https://doi.org/10.1186/s13567-015-0215-y
Mohseni, M., Mohammadhosseinzadeh, N., Keramat, A., 2018. A predator Halobacteriovorax
isolated from the Caspian Sea and the investigation of its ability to control some gram
negative pathogenic bacteria. J. Cell. Mol. Res. Iranian J. Biol. 31, 422–435.
Moon, T.K., 1996. The expectation-maximization algorithm. IEEE Signal Process. Mag. 13,
47–60. https://doi.org/10.1109/79.543975
Mouquet, N., Hoopes, M.F., Amarasekare, P., 2005. The world is patchy and heterogeneous.
Trade-off and source-sink dynamics in competitive metacommunities, in: Holyoak, M.,
Leibold, M.A., Holt, R.D. (Eds.), Metacommunities: Spatial Dynamics and Ecological
Communities. The University of Chicago Press, Chicago, pp. 237–262.

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
149 133
Muniesa, A., Basurco, B., Aguilera, C., Furones, D., Reverté, C., Sanjuan-Vilaplana, A.,
Jansen, M.D., Brun, E., Tavornpanich, S., 2020. Mapping the knowledge of the main
diseases affecting sea bass and sea bream in Mediterranean. Transbound. Emerg. Dis.
67, 1089–1100. https://doi.org/10.1111/tbed.13482
Oksanen, J., Kindt, R., Legendre, P., O’Hara, B., Simpson, G.L., Stevens, M.H.H., Wagner,
H., 2008. The vegan package: community ecology package, version 1.13-1 [WWW
Document].
Ottaviani, D., Pieralisi, S., Chierichetti, S., Rocchegiani, E., Hattab, J., Mosca, F., Tiscar, P.G.,
Leoni, F., Angelico, G., 2020. Vibrio parahaemolyticus control in mussels by a
Halobacteriovorax isolated from the Adriatic sea, Italy. Food Microbiol. 92, 103600.
https://doi.org/10.1016/j.fm.2020.103600
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., Team, R.C., 2021. _nlme: Linear and Nonlinear
Mixed Effects Models_.
Pratte, Z.A., Besson, M., Hollman, R.D., Stewart, F.J., 2018. The Gills of Reef Fish Support a
Distinct Microbiome Influenced by Host-Specific Factors. https://doi.org/10.1128/AEM
Pridgeon, J.W., Klesius, P.H., 2012. Major bacterial diseases in aquaculture and their vaccine
development. Anim. Sci. Rev. 7, 1–16.
Ringø, E., Zhou, Z., Vecino, J.L.G., Wadsworth, S., Romero, J., Krogdahl, Olsen, R.E.,
Dimitroglou, A., Foey, A., Davies, S., Owen, M., Lauzon, H.L., Martinsen, L.L., De
Schryver, P., Bossier, P., Sperstad, S., Merrifield, D.L., 2016. Effect of dietary
components on the gut microbiota of aquatic animals. A never-ending story? Aquac. Nutr.
22, 219–282. https://doi.org/10.1111/anu.12346
Rosado, D., Pérez-Losada, M., Pereira, A., Severino, R., Xavier, R., 2021. Effects of aging on
the skin and gill microbiota of farmed seabass and seabream. Anim. Microbiome 3.
https://doi.org/10.1186/s42523-020-00072-2
Rosado, D., Pérez-Losada, M., Severino, R., Cable, J., Xavier, R., 2019a. Characterization of
the skin and gill microbiomes of the farmed seabass (Dicentrarchus labrax) and
seabream (Sparus aurata). Aquaculture 500, 57–64.
https://doi.org/10.1016/j.aquaculture.2018.09.063
Rosado, D., Xavier, R., Severino, R., Tavares, F., Cable, J., Pérez-Losada, M., 2019b. Effects
of disease, antibiotic treatment and recovery trajectory on the microbiome of farmed
seabass (Dicentrarchus labrax). Sci. Rep. 9, 1–11. https://doi.org/10.1038/s41598-019-
55314-4
Samant, S., Lee, H., Ghassemi, M., Chen, J., Cook, J.L., Mankin, A.S., Neyfakh, A.A., 2008.
Nucleotide Biosynthesis Is Critical for Growth of Bacteria in Human Blood. PLoS Pathog.
4, e37. https://doi.org/10.1371/journal.ppat.0040037
Schade, F.M., Raupach, M.J., Mathias Wegner, K., 2016. Seasonal variation in parasite

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
150 133
infection patterns of marine fish species from the Northern Wadden Sea in relation to
interannual temperature fluctuations. J. Sea Res. 113, 73–84.
https://doi.org/10.1016/j.seares.2015.09.002
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W.S., Huttenhower, C.,
2011. Metagenomic biomarker discovery and explanation. Genome Biol. 12, R60.
https://doi.org/10.1186/gb-2011-12-6-r60
Shenhav, L., Thompson, M., Joseph, T.A., Briscoe, L., Furman, O., Bogumil, D., Mizrahi, I.,
Pe’er, I., Halperin, E., 2019. FEAST: fast expectation-maximization for microbial source
tracking. Nat. Methods 16, 627–632. https://doi.org/10.1038/s41592-019-0431-x
Silva, B.R. dos S., Derami, M.S., Paixão, D.A., Persinoti, G.F., Dias da Silveira, W., Maluta,
R.P., 2020. Comparison between the intestinal microbiome of healthy fish and fish
experimentally infected with Streptococcus agalactiae. Aquac. Res. 51, 3412–3420.
https://doi.org/10.1111/are.14676
Solem, S.T., Stenvik, J., 2006. Antibody repertoire development in teleosts - A review with
emphasis on salmonids and Gadus morhua L. Dev. Comp. Immunol. 30, 57–76.
https://doi.org/10.1016/j.dci.2005.06.007
Sugita, H., Iwata, J., Miyajima, C., Kubo, T., Noguchi, T., Hashimoto, K., Deguchi, Y., 1989.
Changes in microflora of a puffer fish Fugu niphobles, with different water temperatures.
Mar. Biol. 101, 299–304. https://doi.org/10.1007/BF00428125
Tao, Z., Zhou, T., Zhou, S., Wang, G., 2016. Temperature-regulated expression of type VI
secretion systems in fish pathogen Pseudomonas plecoglossicida revealed by
comparative secretome analysis. FEMS Microbiol. Lett. 363, fnw261.
https://doi.org/10.1093/femsle/fnw261
Tarnecki, A.M., Brennan, N.P., Schloesser, R.W., Rhody, N.R., 2019. Shifts in the Skin-
Associated Microbiota of Hatchery-Reared Common Snook Centropomus undecimalis
During Acclimation to the Wild. Microb. Ecol. 77, 770–781.
https://doi.org/10.1007/s00248-018-1252-7
Team, R.C., 2012. R: A language and environment for statistical computing.
Trindade, S., Sousa, A., Gordo, I., 2012. Antibiotic resistance and stress in the light of Fisher’s
model. Evolution (N. Y). 66, 3815–3824. https://doi.org/10.1111/j.1558-
5646.2012.01722.x
Trivedi, B., 2012. Microbiome: the surface brigade. Nature 492, S60–S61.
Uren Webster, T.M., Consuegra, S., Hitchings, M., Garcia de Leaniz, C., 2018. Interpopulation
Variation in the Atlantic Salmon Microbiome Reflects Environmental and Genetic
Diversity. Appl. Environ. Microbiol. 84. https://doi.org/10.1128/aem.00691-18
Varsamos, S., Flik, G., Pepin, J.F., Bonga, S.E.W., Breuil, G., 2006. Husbandry stress during
early life stages affects the stress response and health status of juvenile sea bass,

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
151 133
Dicentrarchus labrax. Fish Shellfish Immunol. 20, 83–96.
https://doi.org/10.1016/j.fsi.2005.04.005
Vasemägi, A., Visse, M., Kisand, V., 2017. Effect of Environmental Factors and an Emerging
Parasitic Disease on Gut Microbiome of Wild Salmonid Fish. mSphere 2, 418–435.
https://doi.org/10.1128/msphere.00418-17
Yukgehnaish, K., Kumar, P., Sivachandran, P., Marimuthu, K., Arshad, A., Paray, B.A.,
Arockiaraj, J., 2020. Gut microbiota metagenomics in aquaculture: factors influencing gut
microbiome and its physiological role in fish. Rev. Aquac. 12, raq.12416.
https://doi.org/10.1111/raq.12416
Zhang, Xiaoting, Ding, L., Yu, Y., Kong, W., Yin, Y., Huang, Z., Zhang, Xuezhen, Xu, Z., 2018.
The Change of Teleost Skin Commensal Microbiota Is Associated With Skin Mucosal
Transcriptomic Responses During Parasitic Infection by Ichthyophthirius multifillis. Front.
Immunol. 9, 2972. https://doi.org/10.3389/fimmu.2018.02972

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
152 133
4.9 Supplementary material Supplementary File S4.1 – Extended Material and methods Experimental design, sampling and processing
European seabass were sampled in a Portuguese fish farm, from a rearing tank
containing a single age cohort of mature adult seabass in order to reduce any possible
maturation effects which are known to affect studied microbiota (e.g., Rosado et al., 2021).
All fish were 26 months old at the beginning of the study and 38 months old at the last sampling
date. We swabbed the right upper lateral part of the fish skin from head to tail and the right gill
filaments between the first and second arch; fish were then released unharmed. Swabs and
filters were immediately frozen at -20°C and then transported in dry ice to the CIBIO-InBIO
laboratory where they were stored at -80°C until further processing.
Data processing and statistical analysis
DNA extraction kit (n=10) and PCR blanks (n=4) were used to control laboratory
contamination. Additionally, four identical mock communities (ZymoBIOMICS Microbial
Community DNA Standard) were sequenced and analyzed together with our fish samples. All
fish and water samples were successfully amplified and sequenced.
Standard filtering parameters for DADA2 were used, with forward and reverse reads
truncated at 220 nt and 200 nt respectively, and with a maximum of two expected errors per
read. Default settings were also used for amplicon sequence variants (ASV) inference and
chimera detection. Taxonomic inferences were made against the SSU SILVA Release 138
reference database (Quast et al., 2012). Approximately 4 336 483, 2 990 334 and 563 545
16S rRNA sequences were retrieved from the skin and gill microbiotas of commercially farmed
European seabass and associated water samples, respectively. The number of sequences
per sample ranged from 2 033 to 6 150 in the skin, 2 077 to 63 711 in the gill and 15 355 to
47 549 in the water samples. ASV abundances were normalized using the negative binomial
distribution (McMurdie and Holmes, 2014). A midpoint rooted tree of ASVs was estimated
using the Quantitative Insights Into Microbial Ecology 2 package (QIIME2; release 2019.7).
After normalization and removal of non-bacterial reads, 5 559, 4 141 and 1 584 ASVs were
assigned to the skin, gill and water microbiotas, respectively. ASVs present in negative
controls (extraction kits and PCR) that were represented by >1% of the reads were removed
from downstream analysis. These corresponded to <0.3% of the reads present in fish and
water samples. Concomitantly, none of the abundant ASVs (>1%) in the fish samples were
present in the controls. Diversity and bacterial abundances of the mock communities
corresponded to that described by the manufacturer.

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
153 133
Phyla and genera with mean relative proportion ≥5% of sequence counts were
considered the most abundant in each fish mucosa (skin and gills) and water for each month.
To determine the core microbiota throughout the year, ASVs present in all months were
identified in each mucosa and water samples.
For the fast expectation-maximization for microbial source tracking (FEAST; Shenhav
et al., 2019) analysis, we used a monthly average ASV table as input. Then the skin/gill
microbiota in a given month was considered as the sink with three potential environmental
sources tested: i) the microbiota of the same mucosa obtained from the previous month (to
infer the fraction of the microbiota that remains stable across consecutive months); ii) the
alternative mucosal microbiota obtained in the same month (to infer microbial exchange
between tissues); and iii) the water microbiota obtained in the same month (to infer microbial
recruitment from the surrounding water). Finally, FEAST also reports the fraction of the sink
microbiota that could potentially be attributed to other unknown sources.
To perform the Temporal Insights into Microbial Ecology (TIME; Baksi et al., 2018)
analysis, an ASV table was used as input. Causality networks were inspected to identify
specific genera that were affected by or were responsible for affecting the temporal changes
of the most abundant PP genera. Secondly, Pearson correlation tests were used to determine
whether there was any correlation between PP genera and the other genera; negative
correlations indicating competition.
The standard deviation of Shannon (Shannon SD) and Faith’s PD (PD SD) indices
were calculated using all 5 samples of a given sampling date or using all 10 samples of a
given month, whether the temperature model to use was based on sampling date or monthly
measurements, respectively.
To evaluate the effect of water temperature, we built four different temperature models
encompassing different time windows to test which explained most of the variation in the
studied microbiota. We first used a longer time frame, where a categorical model was created
by dividing the year into cold (November, December, January, February, and March) and
warm (April, May, June, July, August, September and October). A month was considered cold
or warm whether its mean monthly temperature was lower or higher than the daily annual
average (i.e., 18.6°C, Figure S4.1). The second set of models were based on intermediate
time frames and included the mean monthly temperature measurements across all days of a
month and their respective standard deviation (T_month_mn and T_month_SD). We also
tested four shorter frame models, using the mean temperatures and their respective standard
deviations from 2 and 7 days prior to each sampling date (T2_mn / T2_SD and T7_mn /
T7_SD). For monthly analyses we combined these measurements for the two sampling points
of a given month, in a total of 4 and 14 days per month (T4_mn / T4_SD and T14_mn /
T14_SD). Linear models (lm) were used to assess variation in microbial composition and

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
154 133
included each set of mean and standard deviation temperature models plus season, in order
to account for seasonal variability. Season was categorized into winter (January, February,
March), spring (April, May, June), summer (July, August, September), and fall (October,
November, December). The final general lm formula was expressed as: microbial diversity ~
T_mn + T_SD + Season; or microbial diversity ~ cold/warm + Season. We also analyzed the
relations between diversity and temperature in terms of distance between samples.
Correlation structure between distance matrices was estimated by the generalized least
squares models (gls) We used a maximum likelihood population effects model, as
implemented in corMLPE R package (Clarke et al., 2002), to account for the lack of
independence between pairwise samples in a distance matrix. Distance matrices were
constructed for each continuous temperature variable, alpha-diversity measures and PP
genera abundances using Euclidean distances, while the Gower’s distance (Tuerhong and
Kim, 2014) was used for the categorical temperature variable (cold/warm months). For beta-
diversity, we used the unifrac unweighted and weighted distance matrices.
Bibliography
Baksi, K.D., Kuntal, B.K., Mande, S.S., 2018. ‘TIME’: A Web Application for Obtaining Insights
into Microbial Ecology Using Longitudinal Microbiome Data. Front. Microbiol. 9, 9–36.
https://doi.org/10.3389/fmicb.2018.00036
Clarke, R.T., Rothery, P., Raybould, A.F., 2002. Confidence limits for regression relationships
between distance matrices: Estimating gene flow with distance. J. Agric. Biol. Environ.
Stat. 7, 361–372. https://doi.org/10.1198/108571102320
McMurdie, P.J., Holmes, S., 2014. Waste Not, Want Not: Why Rarefying Microbiome Data Is
Inadmissible. PLoS Comput. Biol. 10, e1003531.
https://doi.org/10.1371/journal.pcbi.1003531
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., Peplies, J., Glöckner,
F.O., 2013. The SILVA ribosomal RNA gene database project: Improved data processing
and web-based tools. Nucleic Acids Res. 41, D590–D596.
https://doi.org/10.1093/nar/gks1219
Rosado, D., Pérez-Losada, M., Pereira, A., Severino, R., Xavier, R., 2021. Effects of aging on
the skin and gill microbiota of farmed seabass and seabream. Anim. Microbiome 3.
https://doi.org/10.1186/s42523-020-00072-2
Shenhav, L., Thompson, M., Joseph, T.A., Briscoe, L., Furman, O., Bogumil, D., Mizrahi, I.,
Pe’er, I., Halperin, E., 2019. FEAST: fast expectation-maximization for microbial source
tracking. Nat. Methods 16, 627–632. https://doi.org/10.1038/s41592-019-0431-x
Tuerhong, G., Kim, S.B., 2014. Gower distance-based multivariate control charts for a mixture
of continuous and categorical variables. Expert Syst. Appl. 41, 1701–1707.
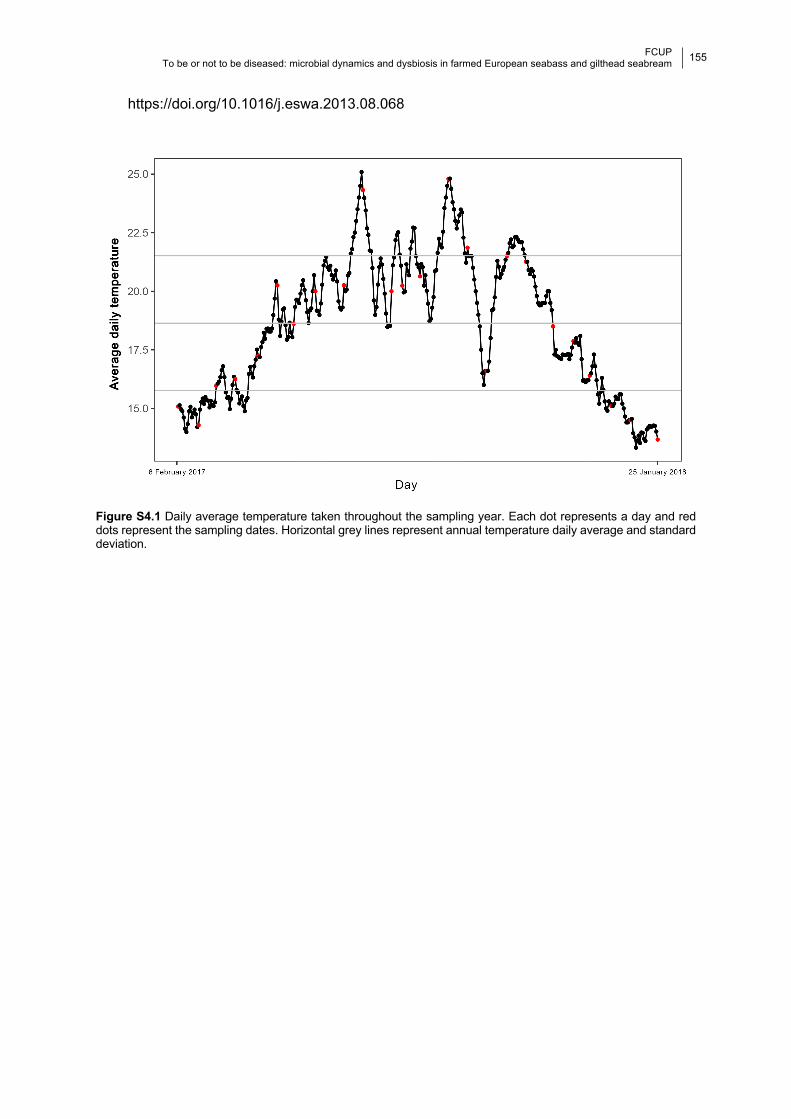
FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
155 133
https://doi.org/10.1016/j.eswa.2013.08.068
Figure S4.1 Daily average temperature taken throughout the sampling year. Each dot represents a day and red dots represent the sampling dates. Horizontal grey lines represent annual temperature daily average and standard deviation.
Ave
rage
dai
ly te
mpe
ratu
re
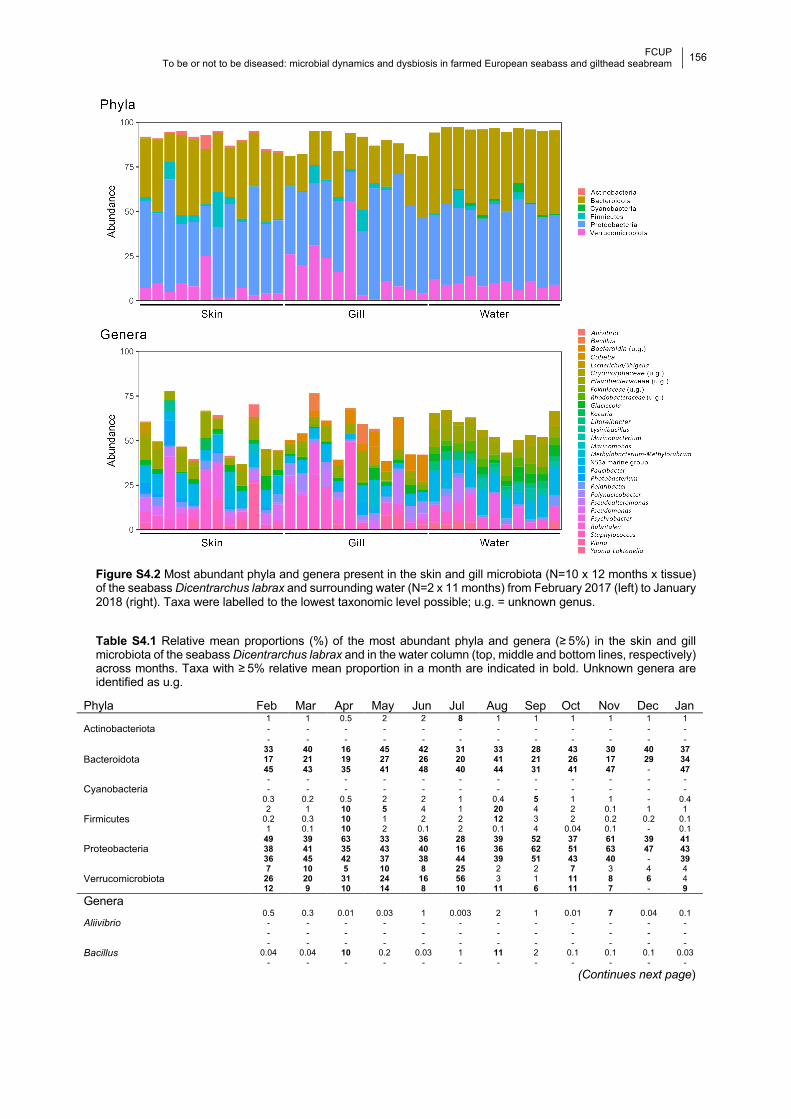
FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
156 133
Figure S4.2 Most abundant phyla and genera present in the skin and gill microbiota (N=10 x 12 months x tissue) of the seabass Dicentrarchus labrax and surrounding water (N=2 x 11 months) from February 2017 (left) to January 2018 (right). Taxa were labelled to the lowest taxonomic level possible; u.g. = unknown genus. Table S4.1 Relative mean proportions (%) of the most abundant phyla and genera (≥ 5%) in the skin and gill microbiota of the seabass Dicentrarchus labrax and in the water column (top, middle and bottom lines, respectively) across months. Taxa with ≥ 5% relative mean proportion in a month are indicated in bold. Unknown genera are identified as u.g.
Phyla Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan
Actinobacteriota
1 1 0.5 2 2 8 1 1 1 1 1 1 - - - - - - - - - - - - - - - - - - - - - - - -
Bacteroidota
33 40 16 45 42 31 33 28 43 30 40 37 17 21 19 27 26 20 41 21 26 17 29 34 45 43 35 41 48 40 44 31 41 47 - 47
Cyanobacteria
- - - - - - - - - - - - - - - - - - - - - - - -
0.3 0.2 0.5 2 2 1 0.4 5 1 1 - 0.4
Firmicutes
2 1 10 5 4 1 20 4 2 0.1 1 1 0.2 0.3 10 1 2 2 12 3 2 0.2 0.2 0.1 1 0.1 10 2 0.1 2 0.1 4 0.04 0.1 - 0.1
Proteobacteria
49 39 63 33 36 28 39 52 37 61 39 41 38 41 35 43 40 16 36 62 51 63 47 43 36 45 42 37 38 44 39 51 43 40 - 39
Verrucomicrobiota
7 10 5 10 8 25 2 2 7 3 4 4 26 20 31 24 16 56 3 1 11 8 6 4 12 9 10 14 8 10 11 6 11 7 - 9
Genera
Aliivibrio 0.5 0.3 0.01 0.03 1 0.003 2 1 0.01 7 0.04 0.1 - - - - - - - - - - - - - - - - - - - - - - - -
Bacillus - - - - - - - - - - - -
0.04 0.04 10 0.2 0.03 1 11 2 0.1 0.1 0.1 0.03 - - - - - - - - - - - -
(Continues next page)
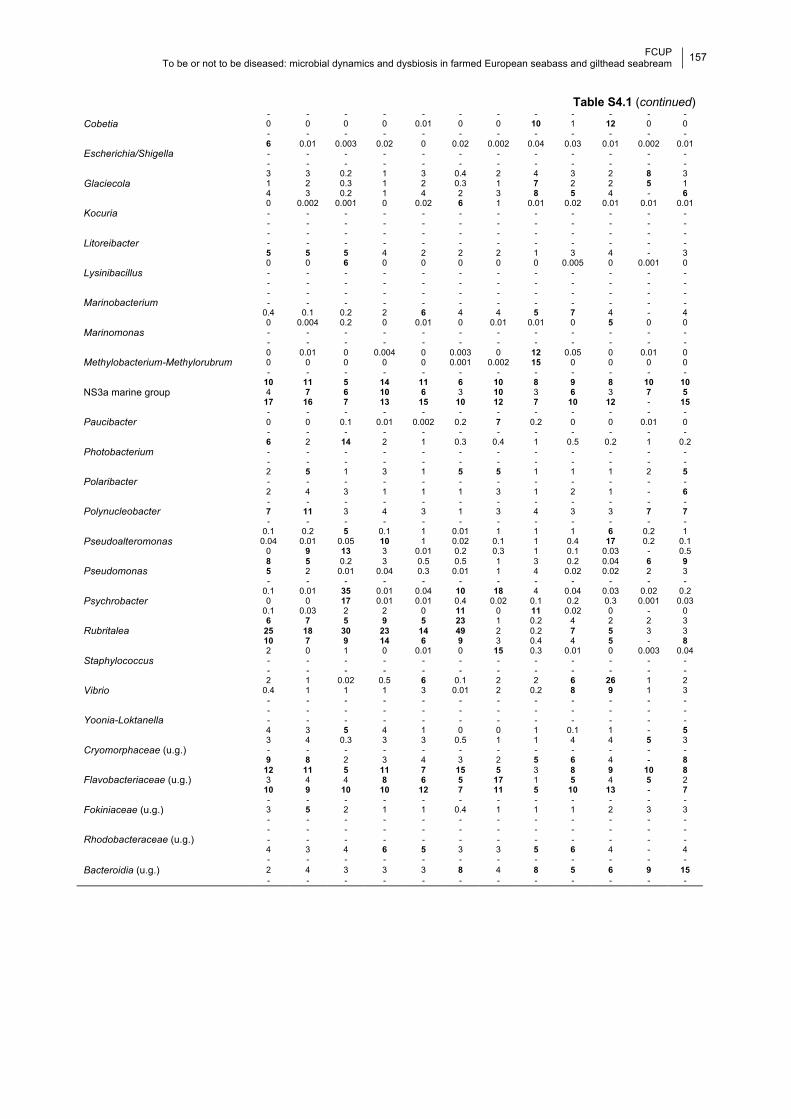
FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
157 133
Table S4.1 (continued)
Cobetia - - - - - - - - - - - - 0 0 0 0 0.01 0 0 10 1 12 0 0 - - - - - - - - - - - -
Escherichia/Shigella 6 0.01 0.003 0.02 0 0.02 0.002 0.04 0.03 0.01 0.002 0.01 - - - - - - - - - - - - - - - - - - - - - - - -
Glaciecola 3 3 0.2 1 3 0.4 2 4 3 2 8 3 1 2 0.3 1 2 0.3 1 7 2 2 5 1 4 3 0.2 1 4 2 3 8 5 4 - 6
Kocuria 0 0.002 0.001 0 0.02 6 1 0.01 0.02 0.01 0.01 0.01 - - - - - - - - - - - - - - - - - - - - - - - -
Litoreibacter - - - - - - - - - - - - - - - - - - - - - - - - 5 5 5 4 2 2 2 1 3 4 - 3
Lysinibacillus 0 0 6 0 0 0 0 0 0.005 0 0.001 0 - - - - - - - - - - - - - - - - - - - - - - - -
Marinobacterium - - - - - - - - - - - - - - - - - - - - - - - -
0.4 0.1 0.2 2 6 4 4 5 7 4 - 4
Marinomonas 0 0.004 0.2 0 0.01 0 0.01 0.01 0 5 0 0 - - - - - - - - - - - - - - - - - - - - - - - -
Methylobacterium-Methylorubrum 0 0.01 0 0.004 0 0.003 0 12 0.05 0 0.01 0 0 0 0 0 0 0.001 0.002 15 0 0 0 0 - - - - - - - - - - - -
NS3a marine group
10 11 5 14 11 6 10 8 9 8 10 10 4 7 6 10 6 3 10 3 6 3 7 5
17 16 7 13 15 10 12 7 10 12 - 15
Paucibacter - - - - - - - - - - - - 0 0 0.1 0.01 0.002 0.2 7 0.2 0 0 0.01 0 - - - - - - - - - - - -
Photobacterium 6 2 14 2 1 0.3 0.4 1 0.5 0.2 1 0.2 - - - - - - - - - - - - - - - - - - - - - - - -
Polaribacter 2 5 1 3 1 5 5 1 1 1 2 5 - - - - - - - - - - - - 2 4 3 1 1 1 3 1 2 1 - 6
Polynucleobacter - - - - - - - - - - - - 7 11 3 4 3 1 3 4 3 3 7 7 - - - - - - - - - - - -
Pseudoalteromonas 0.1 0.2 5 0.1 1 0.01 1 1 1 6 0.2 1
0.04 0.01 0.05 10 1 0.02 0.1 1 0.4 17 0.2 0.1 0 9 13 3 0.01 0.2 0.3 1 0.1 0.03 - 0.5
Pseudomonas 8 5 0.2 3 0.5 0.5 1 3 0.2 0.04 6 9 5 2 0.01 0.04 0.3 0.01 1 4 0.02 0.02 2 3 - - - - - - - - - - - -
Psychrobacter 0.1 0.01 35 0.01 0.04 10 18 4 0.04 0.03 0.02 0.2 0 0 17 0.01 0.01 0.4 0.02 0.1 0.2 0.3 0.001 0.03
0.1 0.03 2 2 0 11 0 11 0.02 0 - 0
Rubritalea 6 7 5 9 5 23 1 0.2 4 2 2 3
25 18 30 23 14 49 2 0.2 7 5 3 3 10 7 9 14 6 9 3 0.4 4 5 - 8
Staphylococcus 2 0 1 0 0.01 0 15 0.3 0.01 0 0.003 0.04 - - - - - - - - - - - - - - - - - - - - - - - -
Vibrio 2 1 0.02 0.5 6 0.1 2 2 6 26 1 2
0.4 1 1 1 3 0.01 2 0.2 8 9 1 3 - - - - - - - - - - - -
Yoonia-Loktanella - - - - - - - - - - - - - - - - - - - - - - - - 4 3 5 4 1 0 0 1 0.1 1 - 5
Cryomorphaceae (u.g.)
3 4 0.3 3 3 0.5 1 1 4 4 5 3 - - - - - - - - - - - - 9 8 2 3 4 3 2 5 6 4 - 8
Flavobacteriaceae (u.g.)
12 11 5 11 7 15 5 3 8 9 10 8 3 4 4 8 6 5 17 1 5 4 5 2
10 9 10 10 12 7 11 5 10 13 - 7
Fokiniaceae (u.g.)
- - - - - - - - - - - - 3 5 2 1 1 0.4 1 1 1 2 3 3 - - - - - - - - - - - -
Rhodobacteraceae (u.g.)
- - - - - - - - - - - - - - - - - - - - - - - - 4 3 4 6 5 3 3 5 6 4 - 4
Bacteroidia (u.g.)
- - - - - - - - - - - - 2 4 3 3 3 8 4 8 5 6 9 15 - - - - - - - - - - - -
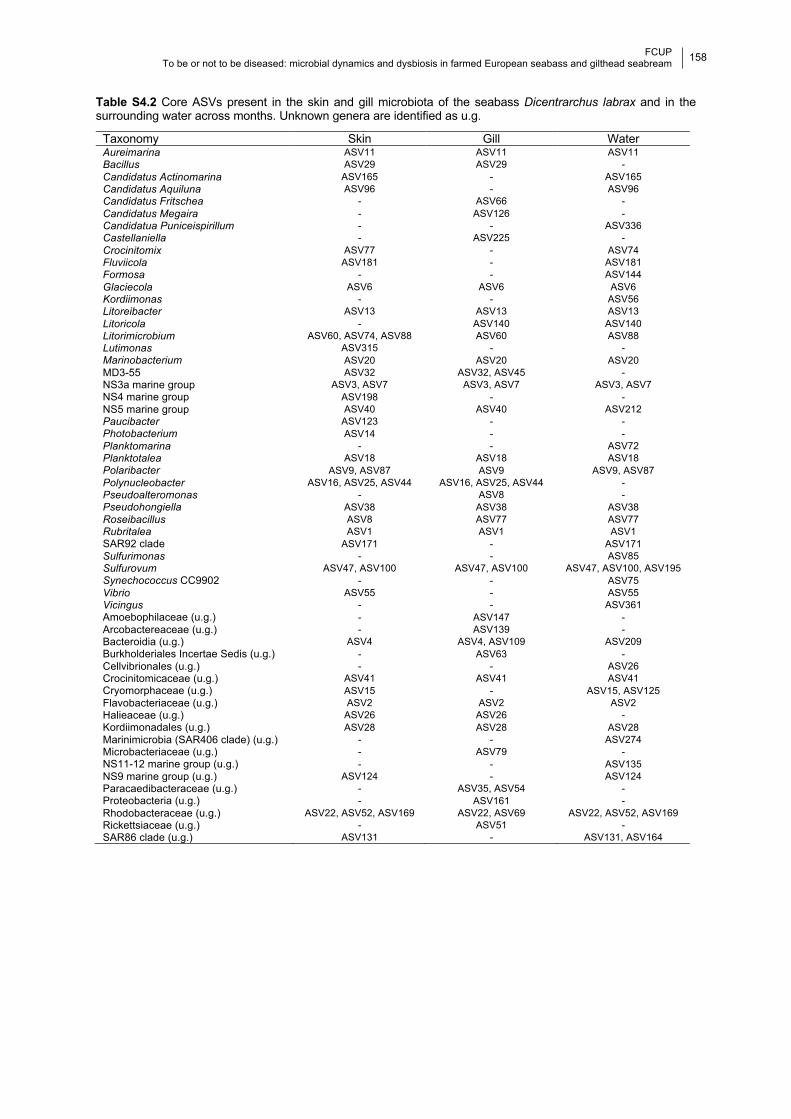
FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
158 133
Table S4.2 Core ASVs present in the skin and gill microbiota of the seabass Dicentrarchus labrax and in the surrounding water across months. Unknown genera are identified as u.g.
Taxonomy Skin Gill Water Aureimarina ASV11 ASV11 ASV11
Bacillus ASV29 ASV29 -
Candidatus Actinomarina ASV165 - ASV165
Candidatus Aquiluna ASV96 - ASV96
Candidatus Fritschea - ASV66 -
Candidatus Megaira - ASV126 -
Candidatua Puniceispirillum - - ASV336
Castellaniella - ASV225 -
Crocinitomix ASV77 - ASV74
Fluviicola ASV181 - ASV181
Formosa - - ASV144
Glaciecola ASV6 ASV6 ASV6
Kordiimonas - - ASV56
Litoreibacter ASV13 ASV13 ASV13
Litoricola - ASV140 ASV140
Litorimicrobium ASV60, ASV74, ASV88 ASV60 ASV88
Lutimonas ASV315 - -
Marinobacterium ASV20 ASV20 ASV20
MD3-55 ASV32 ASV32, ASV45 -
NS3a marine group ASV3, ASV7 ASV3, ASV7 ASV3, ASV7
NS4 marine group ASV198 - -
NS5 marine group ASV40 ASV40 ASV212
Paucibacter ASV123 - -
Photobacterium ASV14 - -
Planktomarina - - ASV72
Planktotalea ASV18 ASV18 ASV18
Polaribacter ASV9, ASV87 ASV9 ASV9, ASV87
Polynucleobacter ASV16, ASV25, ASV44 ASV16, ASV25, ASV44 -
Pseudoalteromonas - ASV8 -
Pseudohongiella ASV38 ASV38 ASV38
Roseibacillus ASV8 ASV77 ASV77
Rubritalea ASV1 ASV1 ASV1
SAR92 clade ASV171 - ASV171
Sulfurimonas - - ASV85
Sulfurovum ASV47, ASV100 ASV47, ASV100 ASV47, ASV100, ASV195
Synechococcus CC9902 - - ASV75
Vibrio ASV55 - ASV55
Vicingus - - ASV361
Amoebophilaceae (u.g.) - ASV147 -
Arcobactereaceae (u.g.) - ASV139 -
Bacteroidia (u.g.) ASV4 ASV4, ASV109 ASV209
Burkholderiales Incertae Sedis (u.g.) - ASV63 -
Cellvibrionales (u.g.) - - ASV26
Crocinitomicaceae (u.g.) ASV41 ASV41 ASV41
Cryomorphaceae (u.g.) ASV15 - ASV15, ASV125
Flavobacteriaceae (u.g.) ASV2 ASV2 ASV2
Halieaceae (u.g.) ASV26 ASV26 -
Kordiimonadales (u.g.) ASV28 ASV28 ASV28
Marinimicrobia (SAR406 clade) (u.g.) - - ASV274
Microbacteriaceae (u.g.) - ASV79 -
NS11-12 marine group (u.g.) - - ASV135
NS9 marine group (u.g.) ASV124 - ASV124
Paracaedibacteraceae (u.g.) - ASV35, ASV54 -
Proteobacteria (u.g.) - ASV161 -
Rhodobacteraceae (u.g.) ASV22, ASV52, ASV169 ASV22, ASV69 ASV22, ASV52, ASV169
Rickettsiaceae (u.g.) - ASV51 -
SAR86 clade (u.g.) ASV131 - ASV131, ASV164
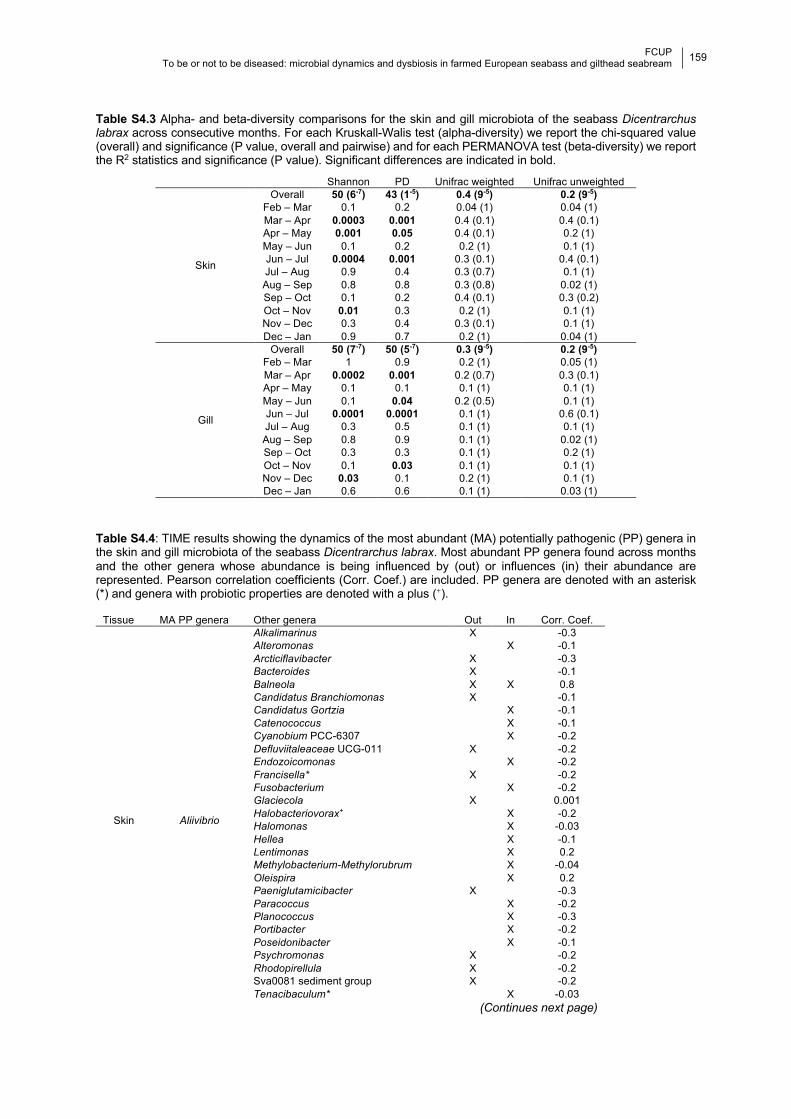
FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
159 133
Table S4.3 Alpha- and beta-diversity comparisons for the skin and gill microbiota of the seabass Dicentrarchus labrax across consecutive months. For each Kruskall-Walis test (alpha-diversity) we report the chi-squared value (overall) and significance (P value, overall and pairwise) and for each PERMANOVA test (beta-diversity) we report the R2 statistics and significance (P value). Significant differences are indicated in bold.
Shannon PD Unifrac weighted Unifrac unweighted
Skin
Overall 50 (6-7) 43 (1-5) 0.4 (9-5) 0.2 (9-5) Feb – Mar 0.1 0.2 0.04 (1) 0.04 (1) Mar – Apr 0.0003 0.001 0.4 (0.1) 0.4 (0.1) Apr – May 0.001 0.05 0.4 (0.1) 0.2 (1) May – Jun 0.1 0.2 0.2 (1) 0.1 (1) Jun – Jul 0.0004 0.001 0.3 (0.1) 0.4 (0.1) Jul – Aug 0.9 0.4 0.3 (0.7) 0.1 (1) Aug – Sep 0.8 0.8 0.3 (0.8) 0.02 (1) Sep – Oct 0.1 0.2 0.4 (0.1) 0.3 (0.2) Oct – Nov 0.01 0.3 0.2 (1) 0.1 (1) Nov – Dec 0.3 0.4 0.3 (0.1) 0.1 (1) Dec – Jan 0.9 0.7 0.2 (1) 0.04 (1)
Gill
Overall 50 (7-7) 50 (5-7) 0.3 (9-5) 0.2 (9-5) Feb – Mar 1 0.9 0.2 (1) 0.05 (1) Mar – Apr 0.0002 0.001 0.2 (0.7) 0.3 (0.1) Apr – May 0.1 0.1 0.1 (1) 0.1 (1) May – Jun 0.1 0.04 0.2 (0.5) 0.1 (1) Jun – Jul 0.0001 0.0001 0.1 (1) 0.6 (0.1) Jul – Aug 0.3 0.5 0.1 (1) 0.1 (1) Aug – Sep 0.8 0.9 0.1 (1) 0.02 (1) Sep – Oct 0.3 0.3 0.1 (1) 0.2 (1) Oct – Nov 0.1 0.03 0.1 (1) 0.1 (1) Nov – Dec 0.03 0.1 0.2 (1) 0.1 (1) Dec – Jan 0.6 0.6 0.1 (1) 0.03 (1)
Table S4.4: TIME results showing the dynamics of the most abundant (MA) potentially pathogenic (PP) genera in the skin and gill microbiota of the seabass Dicentrarchus labrax. Most abundant PP genera found across months and the other genera whose abundance is being influenced by (out) or influences (in) their abundance are represented. Pearson correlation coefficients (Corr. Coef.) are included. PP genera are denoted with an asterisk (*) and genera with probiotic properties are denoted with a plus (+).
Tissue MA PP genera Other genera Out In Corr. Coef.
Skin Aliivibrio
Alkalimarinus X -0.3 Alteromonas X -0.1 Arcticiflavibacter X -0.3 Bacteroides X -0.1 Balneola X X 0.8 Candidatus Branchiomonas X -0.1 Candidatus Gortzia X -0.1 Catenococcus X -0.1 Cyanobium PCC-6307 X -0.2 Defluviitaleaceae UCG-011 X -0.2 Endozoicomonas X -0.2 Francisella* X -0.2 Fusobacterium X -0.2 Glaciecola X 0.001 Halobacteriovorax+ X -0.2 Halomonas X -0.03 Hellea X -0.1 Lentimonas X 0.2 Methylobacterium-Methylorubrum X -0.04 Oleispira X 0.2 Paeniglutamicibacter X -0.3 Paracoccus X -0.2 Planococcus X -0.3 Portibacter X -0.2 Poseidonibacter X -0.1 Psychromonas X -0.2 Rhodopirellula X -0.2 Sva0081 sediment group X -0.2 Tenacibaculum* X -0.03
(Continues next page)

FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
160 133
Table S4.4 (continued) Thalassotalea X -0.04
Photobacterium
Acholeplasma X -0.1 Alkalimarinus X -0.3 Amylibacter X X 0.7 Aureispira X -0.2 Candidatus Branchiomonas X -0.1 Cetobacterium X -0.2 Clostridium sensu stricto X -0.1 Defluviitaleaceae UCG-011 X -0.2 Desulforhopalus X -0.3 Devosia X -0.02 Fransicella* X -0.1 Hypnocyclicus X -0.2 Ilumatobacter X -0.01 Labilibacter X -0.1 Lentisphaera X -0.1 Lewinella X -0.2 Litoricola X -0.05 Maribacter X -0.3 Marinobacter X -0.1 Marinoscillum X -0.2 Massilia X 0.04 MD3-55 X 0.03 Moritella X -0.2 OM43 clade X -0.03 Pantoea X -0.1 Parahaliea X -0.2 Phaeocystidibacter X -0.2 Propionigenium X -0.2 Pseudochrobactrum X -0.1 Pseudofulvibacter X X 0.1 fffPseudomonas* X -0.2 Psychrilyobacter X -0.1 Rhodopirellula X -0.2 RS62 marine group X 0.3 Salinirepens X -0.2 Sanguibacter X -0.02 SAR92 clade X -0.2 Sediminispirochaeta X -0.2 SEEP-SRB1 X -0.1 Sphingomonas+ X -0.03 Sulfurimonas X -0.2
Sulfurovum X -0.2 Thiomicrorhabdus X -0.2 Ulvibacter X -0.1 Wenyingzhuangia X -0.2
Pseudomonas
Alkalimarinus X 0.7 Bacteroides X 0.1 BD1-7 clade X 0.2 Candidatus Fritschea X 0.7 Candidatus Omnitrophus X -0.04 Defluviitaleaceae UCG-011 X 0.6 Desulfobulbus X -0.3 Desulforhopalus X 0.2 Francisella* X 0.8 Marinomonas X -0.3 Micrococcus X -0.1 Muricauda X -0.1 Phaeocystidibacter X 0.6 Photobacterium* X -0.2 Psychrilyobacter X 0.2 RBG-16-49-21 X 0.1 Sva0081 sediment group X 0.2 Thiomicrorhabdus X 0.4 Vibrio* X -0.3
Vibrio
Alkalimarinus X -0.2 Alteromonas X -0.1 Arcticiflavibacter X -0.2 Bacteroides X -0.1
(Continues next page)
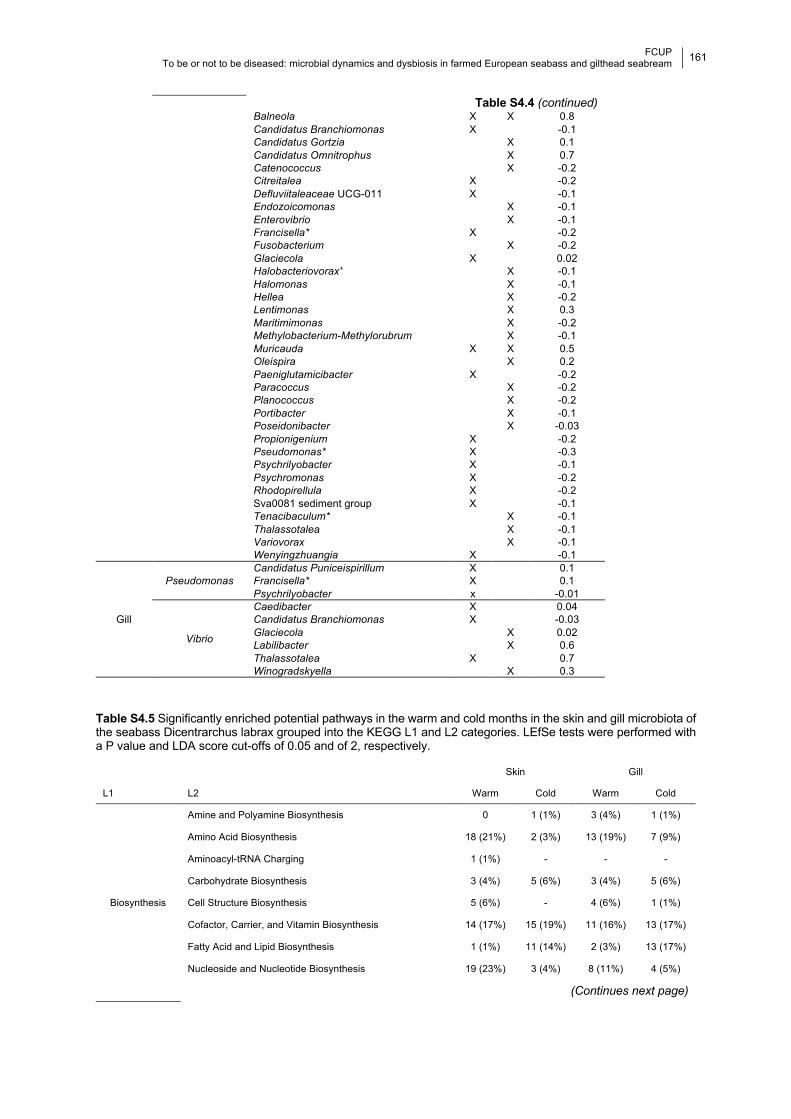
FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
161 133
Table S4.4 (continued) Balneola X X 0.8 Candidatus Branchiomonas X -0.1 Candidatus Gortzia X 0.1 Candidatus Omnitrophus X 0.7 Catenococcus X -0.2 Citreitalea X -0.2 Defluviitaleaceae UCG-011 X -0.1 Endozoicomonas X -0.1 Enterovibrio X -0.1 Francisella* X -0.2 Fusobacterium X -0.2 Glaciecola X 0.02 Halobacteriovorax+ X -0.1 Halomonas X -0.1 Hellea X -0.2 Lentimonas X 0.3 Maritimimonas X -0.2 Methylobacterium-Methylorubrum X -0.1 Muricauda X X 0.5 Oleispira X 0.2 Paeniglutamicibacter X -0.2 Paracoccus X -0.2 Planococcus X -0.2 Portibacter X -0.1 Poseidonibacter X -0.03 Propionigenium X -0.2 Pseudomonas* X -0.3 Psychrilyobacter X -0.1 Psychromonas X -0.2 Rhodopirellula X -0.2 Sva0081 sediment group X -0.1 Tenacibaculum* X -0.1 Thalassotalea X -0.1 Variovorax X -0.1 Wenyingzhuangia X -0.1
Gill
Pseudomonas Candidatus Puniceispirillum X 0.1 Francisella* X 0.1 Psychrilyobacter x -0.01
Vibrio
Caedibacter X 0.04 Candidatus Branchiomonas X -0.03 Glaciecola X 0.02 Labilibacter X 0.6 Thalassotalea X 0.7 Winogradskyella X 0.3
Table S4.5 Significantly enriched potential pathways in the warm and cold months in the skin and gill microbiota of the seabass Dicentrarchus labrax grouped into the KEGG L1 and L2 categories. LEfSe tests were performed with a P value and LDA score cut-offs of 0.05 and of 2, respectively.
Skin Gill
L1 L2 Warm Cold Warm Cold
Biosynthesis
Amine and Polyamine Biosynthesis 0 1 (1%) 3 (4%) 1 (1%)
Amino Acid Biosynthesis 18 (21%) 2 (3%) 13 (19%) 7 (9%)
Aminoacyl-tRNA Charging 1 (1%) - - -
Carbohydrate Biosynthesis 3 (4%) 5 (6%) 3 (4%) 5 (6%)
Cell Structure Biosynthesis 5 (6%) - 4 (6%) 1 (1%)
Cofactor, Carrier, and Vitamin Biosynthesis 14 (17%) 15 (19%) 11 (16%) 13 (17%)
Fatty Acid and Lipid Biosynthesis 1 (1%) 11 (14%) 2 (3%) 13 (17%)
Nucleoside and Nucleotide Biosynthesis 19 (23%) 3 (4%) 8 (11%) 4 (5%)
(Continues next page)
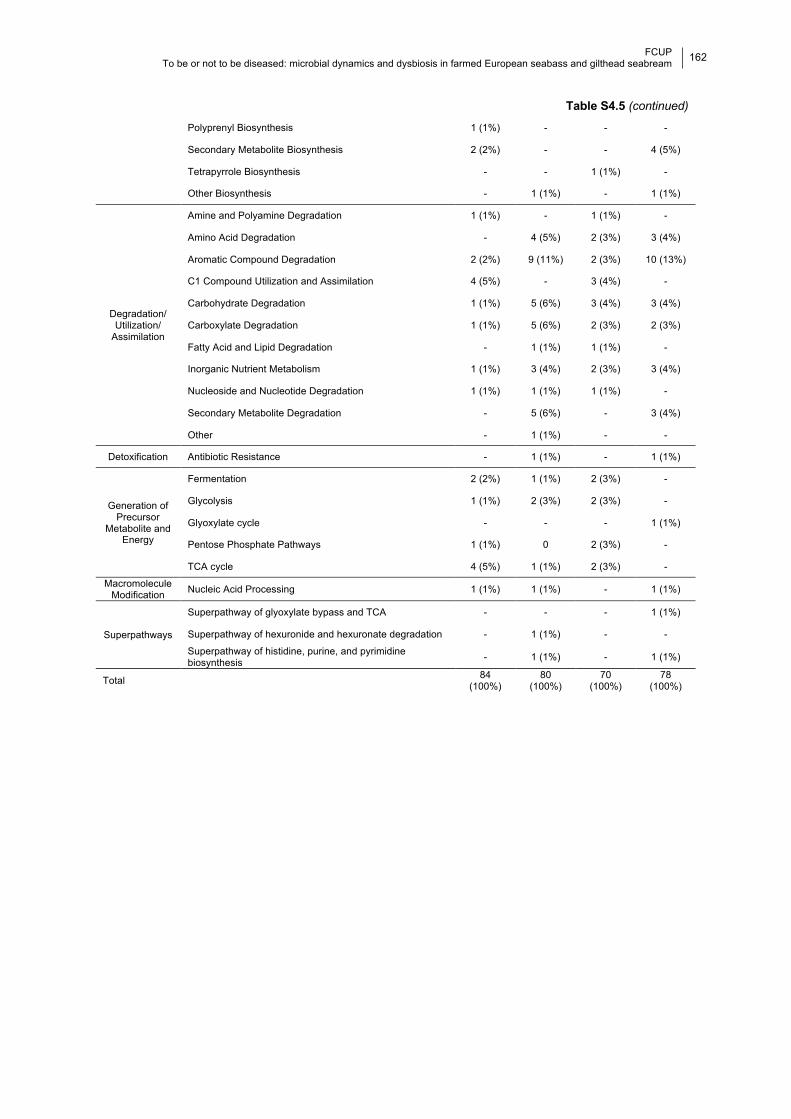
FCUP
To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
162 133
Table S4.5 (continued)
Polyprenyl Biosynthesis 1 (1%) - - -
Secondary Metabolite Biosynthesis 2 (2%) - - 4 (5%)
Tetrapyrrole Biosynthesis - - 1 (1%) -
Other Biosynthesis - 1 (1%) - 1 (1%)
Degradation/
Utilization/
Assimilation
Amine and Polyamine Degradation 1 (1%) - 1 (1%) -
Amino Acid Degradation - 4 (5%) 2 (3%) 3 (4%)
Aromatic Compound Degradation 2 (2%) 9 (11%) 2 (3%) 10 (13%)
C1 Compound Utilization and Assimilation 4 (5%) - 3 (4%) -
Carbohydrate Degradation 1 (1%) 5 (6%) 3 (4%) 3 (4%)
Carboxylate Degradation 1 (1%) 5 (6%) 2 (3%) 2 (3%)
Fatty Acid and Lipid Degradation - 1 (1%) 1 (1%) -
Inorganic Nutrient Metabolism 1 (1%) 3 (4%) 2 (3%) 3 (4%)
Nucleoside and Nucleotide Degradation 1 (1%) 1 (1%) 1 (1%) -
Secondary Metabolite Degradation - 5 (6%) - 3 (4%)
Other - 1 (1%) - -
Detoxification Antibiotic Resistance - 1 (1%) - 1 (1%)
Generation of
Precursor
Metabolite and
Energy
Fermentation 2 (2%) 1 (1%) 2 (3%) -
Glycolysis 1 (1%) 2 (3%) 2 (3%) -
Glyoxylate cycle - - - 1 (1%)
Pentose Phosphate Pathways 1 (1%) 0 2 (3%) -
TCA cycle 4 (5%) 1 (1%) 2 (3%) -
Macromolecule
Modification Nucleic Acid Processing 1 (1%) 1 (1%) - 1 (1%)
Superpathways
Superpathway of glyoxylate bypass and TCA - - - 1 (1%)
Superpathway of hexuronide and hexuronate degradation - 1 (1%) - -
Superpathway of histidine, purine, and pyrimidine
biosynthesis - 1 (1%) - 1 (1%)
Total 84
(100%)
80
(100%)
70
(100%)
78
(100%)

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
163
CHAPTER 5
Effects of disease and antibiotic treatment on the
microbiota of juvenile and adult seabass
5.1 Effects of disease, antibiotic treatment and
recovery trajectory on the microbiome of farmed
seabass (Dicentrarchus labrax)
Daniela Rosado, Raquel Xavier, Ricardo Severino, Fernando Tavares, Jo
Cable, Marcos Pérez-Losada
2019. Scientific Reports 9, 18946. https://doi.org/10.1038/s41598-019-55314-4.
5.1.1 Abstract The mucosal surfaces of fish harbour microbial communities that can act as the first-
line of defense against pathogens. Infectious diseases are one of the main constraints to
aquaculture growth leading to huge economic losses. Despite their negative impacts on
microbial diversity and overall fish health, antibiotics are still the method of choice to treat
many such diseases. Here, we use 16 rRNA V4 metataxonomics to study over a 6 week period
the dynamics of the gill and skin microbiomes of farmed seabass before, during and after a
natural disease outbreak and subsequent antibiotic treatment with oxytetracycline.
Photobacterium damselae was identified as the most probable causative agent of disease.
Both infection and antibiotic treatment caused significant, although asymmetrical, changes in
the microbiome composition of the gills and skin. The most dramatic changes in microbial
taxonomic abundance occurred between healthy and diseased fish. Disease led to a decrease
in the bacterial core diversity in the skin, whereas in the gills there was both an increase and
a shift in core diversity. Oxytetracycline caused a decrease in core diversity in the gill and an
increase in the skin. Severe loss of core diversity in fish mucosae demonstrates the disruptive
impact of disease and antibiotic treatment on the microbial communities of healthy fish.
Keywords: aquaculture, 16S rRNA, oxytetracycline, Photobacterium damselae, dysbiosis

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
164
5.1.2 Introduction Mucosal surfaces of animals harbour microbial communities (i.e., microbiomes), which
can act as the first-line of defense against pathogens, either through competition or production
of antibiotic compounds (Gómez and Balcázar, 2008; Kelly and Salinas, 2017; Trivedi, 2012).
Furthermore, microbiomes are thought to have evolved to optimize the immune response of
each organ and promote homeostasis (Kelly and Salinas, 2017; Lee and Mazmanian, 2010).
Usually a diverse microbiome is associated with healthy phenotypes, but disruptions to this
equilibrium can lead to an increase in abundance of opportunistic pathogens and disease
susceptibility (Mohammed and Arias, 2015; Reid et al., 2017).
Many factors can shape the composition of the fish microbiomes, including host
species (Larsen et al., 2013), stress (Boutin et al., 2013; Zha et al., 2018), diet (Chiarello et
al., 2018), water quality (Galbraith et al., 2018), host physiology (Pratte et al., 2018; Ye et al.,
2014) and infection (Legrand et al., 2018; Llewellyn et al., 2017; Reid et al., 2017). Importantly,
healthy mucosal surfaces, such as the skin and gills, are naturally colonized by pathogens
(Givens et al., 2015; Li et al., 2018; Rosado et al., 2019) from the surrounding waters that can
integrate into the host’s microbial community (Califano et al., 2017; Rud et al., 2017). A shift
in the abundance of such pathogens on the fish mucosae can lead to microbial imbalance (i.e.
dysbiosis) and disease (Hess et al., 2015), which is usually accompanied by a reduction in
bacterial diversity (Legrand et al., 2018; Llewellyn et al., 2014; Reid et al., 2017).
Stress imposed by fish farming conditions can also result in changes in microbiome
composition that may lead to an increase in disease susceptibility (Boutin et al., 2013). As
infectious disease is one of the main constraints to aquaculture growth and profitability, it is
crucial to have a better understanding of the host-symbiont-pathogen nexus. The European
seabass Dicentrarchus labrax is one of the main farmed fish species in southern Europe,
totaling 103.476 tons in landings (10% of global aquaculture production) between 2002 and
2011 (FAO, 2016). This important food resource is susceptible to several bacterial pathogens:
Photobacterium damselae, which causes photobacteriosis, Vibrio spp. causing vibriosis, and
Tenacibaculum maritimum causing tenacibaculosis, just to name a few (Toranzo et al., 2005).
All of these pathogens can induce bacterial septicemia resulting in high mortalities in fish farms
(Faílde et al., 2014; Romalde et al., 2008; Zlotkin et al., 1998). Photobacterium damselae in
particular, is increasingly being reported as the main etiologic agent affecting fish farms
worldwide and has been also described in molluscs, crustaceans and mammals (Labella et
al., 2011; Pedersen et al., 2009; Rivas et al., 2013, 2011; Tao et al., 2018; Terceti et al., 2018,
2016). Control of photobacteriosis in fish farms is challenging, and mortality can reach 60–
80% rates in farmed European seabass (Bakopoulos et al., 2003; Essam et al., 2016).
Although vaccination is available, immunization is still not fully effective (Bakopoulos et al.,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
165
2003; Byadgi et al., 2018), and in many cases antibiotic treatment remains the preferred option
to control such pathogens (e.g., Oxytetracycline, Abdel-Aziz et al., 2013; Labella et al., 2011).
Most commonly, the impact of antibiotic use on fish health is assessed through
toxicological studies (Enis Yonar et al., 2011; Rodrigues et al., 2017). The few studies that
have investigated the effects of antibiotics on the microbiome of fish have focused on gut
dysbiosis (Carlson et al., 2017, 2015; Liu et al., 2012; Narrowe et al., 2015; Pindling et al.,
2018; Zhou et al., 2018). Not surprisingly, a decrease in microbial diversity was detected
(Carlson et al., 2017; Zhou et al., 2018), along with an increased susceptibility to secondary
infection, reduced host growth (Carlson et al., 2015; Liu et al., 2012) and higher mortality
(Pindling et al., 2018). Importantly, these studies also reported bacterial pathogens acquiring
resistance after antibiotic treatment, suggesting that farmed fish microbiomes could become
reservoirs for antibiotic resistant genes (Carlson et al., 2017; Liu et al., 2012; Pindling et al.,
2018). In fact, several studies showed an increase in resistance to tetracycline and
streptomycin antibiotics in strains of P. damselae sampled from both wild and farmed fish
hosts (Abdel-Aziz et al., 2013; Chiu et al., 2013; Essam et al., 2016; Nonaka et al., 2012).
In the present study, we characterized the dynamics of the gill and skin microbiomes
of the seabass Dicentrarchus labrax before, during and after a disease outbreak potentially
caused by Photobacterium damselae, and subsequent antibiotic treatment with
oxytetracycline (Rigos and Troisi, 2005). We describe the dysbiosis caused by disease and
antibiotic treatment in microbial diversity of both mucosae over 3 weeks. Towards this aim, we
used 16S rRNA high-throughput sequencing (metataxonomics) and amplicon sequence
variance analysis to examine changes in both alpha- and beta-diversity, as well as differences
in taxa proportion over time.
5.1.3 Material and methods
5.1.3.1 Ethical statement This study monitored a natural infection and subsequent antibiotic treatment as part
of routine procedures in a commercial fish farm. All animals were handled by the fish farm
employees, our sampling through swabbing was non-invasive and fish were released
unharmed with no mortalities observed. According to the Portuguese legislation DL N°
113/2013, our work does not involve animal experimentation and therefore is exempted from
the need of ethical approval.
5.1.3.2 Experimental design, sample collection and preparation Ten individuals of seabass were collected once a week between August 21 and
October 3, 2016, from the same rearing tank in a commercial fish farm located in the estuarine

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
166
environment of the Ria Formosa (Portimão), southern Portugal. Fish were hatched at
September 26, 2014 and entered the growth facility at March 6, 2015. Fish were kept in an
open water circulation system in a semi-intensive farming facility, where water is supplied to
each tank from the estuary. Fish were kept at a density of ca. 3 kg/m3 corresponding to roughly
100 fish/tank with fish weighting on average 281 g. Given that it was not possible to tag
individual fish and the unlikelihood of re-sampling the same individuals every week, a subset
of samples believed to be representative of the population was chosen, i.e., 10 individuals (~
10%), and for statistical purposes individuals were considered as pseudo-replicates. All fish
were fed with the same commercial feed and they shared the same clinical history. Individuals
were randomly caught using a fishing pole and skin and gill swabs were collected immediately
using tubed sterile dry swabs (Medical Wire & Equipment, UK). Skin samples were taken by
swabbing several times along the right upper lateral part of the fish from head to tail, while gill
swabs were taken from the right filaments between the first and second arch. Due to the non-
invasive nature of our sampling procedure, it was not possible to ascertain the sex of the
individuals sampled; however, we do not expect this to impact our conclusions since, to the
best of our knowledge, no gender bias in microbiome composition has ever been reported for
skin or gill of piscine hosts. Swabs were immediately stored at −20 °C until transported on dry
ice to the CIBIO-InBIO laboratory by airmail where they were kept at −80 °C until further
processing.
To assess gill and skin microbiome dynamics in the seabass during a disease outbreak
and under oxytetracycline treatment after infection, fish were sampled in 4 different states:
healthy, diseased, treatment and recovery (Figure 5.1.1). During the healthy state (August 21
and 29), all fish specimens were considered healthy due to a lack of visible disease symptoms,
such as external lesions or behavioural alterations. On September 8 fish began to die in the
farming tanks, showing symptoms of disease, and treatment with oxytetracycline antibiotic (a
broad-spectrum tetracycline) was initiated, being administrated at 35 g/Kg through commercial
feed for at lasted 8 days. On the same day, smears from spleen and kidney were collected for
culture using Bionor kits DE020, MONO-VA-50 for Vibrio anguillarum and DL020, MONO-Pp-
50 for Photobacterium piscicida. Agglutination essays were not conclusive and, at that stage,
the causative agent of the disease was unknown. We do not have samples from September
8, hence we used the samples from our closest time point, September 5, which we classified
as potentially diseased (i.e., diseased state). Antibiotic treatment lasted until September 16
and fish were sampled on September 12; this sample point corresponded to the treatment
state. Then, three additional time points were sampled (September 19 and 26, and October
3), when fish were no longer dying or presented signs of infection; these three time points
corresponded to the recovery state.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
167
Total DNA from 140 fish samples (70 skin and 70 gills) was extracted using the
PowerSoil DNA Isolation Kit (QIAGEN, Netherlands), following the manufacturer’s protocol.
DNA extractions were shipped in dry ice to the University of Michigan Medical School (USA)
for amplification and sequencing on a single run of the Illumina MiSeq platform according to
the protocol of Kozich et al. (2013). Each sample was amplified for the V4 (~250 bp)
hypervariable region of the 16S rRNA gene, using the primers in Caporaso et al. (2011). This
region has been widely used to characterize microbiomes from vertebrates (Earth Microbiome
Project, Gilbert et al., 2014), including fish (Carlson et al., 2017; Llewellyn et al., 2016; Nielsen
et al., 2017; Wang et al., 2017).
5.1.3.3 Data and statistical analyses Raw FASTQ files were analyzed using the Quantitative Insights Into Microbial Ecology
2 (QIIME2; release 2018.4) platform. Clean sequences were aligned against the SILVA (132
release) reference database (Quast et al., 2013) using the DADA2 pipeline (Callahan et al.,
2016). A feature table containing amplicon sequence variants (ASVs) was constructed and
normalized using the negative binomial distribution (McMurdie and Holmes, 2014). The core
microbiome was assessed at the ASV level for the gill and skin of seabass for each state
(healthy, diseased, treatment and recovery) separately. An ASV was considered as part of the
core microbiome if present in 100% of the samples from each state. Core diversity is here
defined as number of ASVs represented in a given group.
Microbial alpha-diversity (intra-sample) was calculated using Shannon, ACE, Fisher
and Faith’s phylogenetic diversity (PD) indices as implemented in the R package phyloseq
(McMurdie and Holmes, 2013). Microbial beta-diversity (inter-sample) was estimated using
phylogenetic Unifrac (unweighted and weighted) and Bray-Curtis distances. Dissimilarity
between samples was assessed by principal coordinates analysis (PCoA). Variation in
microbial alpha-diversity and the mean proportions of the most abundant taxa (with more than
4% of all reads) were assessed using linear models with randomized residuals in a
permutation procedure (RRPP). Differences in community composition (beta-diversity) were
tested using permutational multivariate analysis of variance (PERMANOVA) with 1,000
permutations as implemented in the adonis function of the R vegan package (Oksanen et al.,
2008). All statistical analyses were carried out separately for the gills and skin. All statistical
analyses were performed in R-studio v1.0.143 (Studio, 2012).
5.1.4 Results Approximately, a total of 3.6 million raw reads were generated, while the number of
sequences per sample ranged from 2,354 to 50,564. A total of 6,485 unique ASVs were

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
168
detected, but after normalization and depletion of Archaea and Algae ASVs, a total of 3,827
ASVs (1,560,279 sequences) and 3,741 ASVs (1,904,115 sequences) were analyzed for the
gill and skin microbiomes, respectively. Taxa showing a mean proportion ≥4% in any state
were considered the most abundant taxa. Analyses of alpha- and beta-diversity showed no
significant differences (P > 0.05; data not shown) between samples from the two healthy time
points as well as between the samples from the three recovery time points (Figure 5.1.1) for
both gill and skin microbiomes. Therefore, in all our subsequent analyses, samples from Aug
21 (Healthy 1) and Aug 29 (Healthy 2) were combined into the “healthy” state; and samples
from Sep 19 (Recovery 1), Sep 26 (Recovery 2) and Oct 3 (Recovery 3) were combined into
the “recovery” state in order to increase sample size (Figure 5.1.1).
Figure 5.1.1 Schematic illustration of the experimental design and health status of each sampling point. Ten fish were sampled for gill and skin microbial communities at each sampling point, totaling 70 fish sampled in this experiment.
5.1.4.1 Gill bacterial composition and diversity No significant differences were detected in alpha-diversity across all states (RRPP,
P > 0.5), with the exception of the Shannon index (RRPP, P = 0.04) (Table 5.1.1, Figure
5.1.2A). There were, however, significant differences between healthy and recovery states for
all alpha-diversity indices (RRPP, P ≤ 0.03; Table 5.1.1). Beta-diversity also varied greatly
between states (PCoA, Figure 5.1.3A), with significant differences both in overall and pairwise
comparisons in almost all the tests (Adonis, P < 0.05; Table 5.1.1).
Bacteroidetes, Proteobacteria and Verrucomicrobia were the most abundant phyla
retrieved from the seabass gill microbiome across states, accounting for 83% to 93% of the
sequences altogether (Table 5.1.2). The most abundant genera were NS3a marine group,
Polaribacter 4, Pseudomonas and Rubritalea, which were present in all four states (Table
5.1.2, Figure S5.1.1A). Other relatively abundant genera were: (a) Polynucleobacter, which
accounted for 4–7% of the sequences in the diseased, treatment and recovery states, but only
0.2% in the healthy state; (b) Stenotrophomonas, represented by 5% of the sequences in the
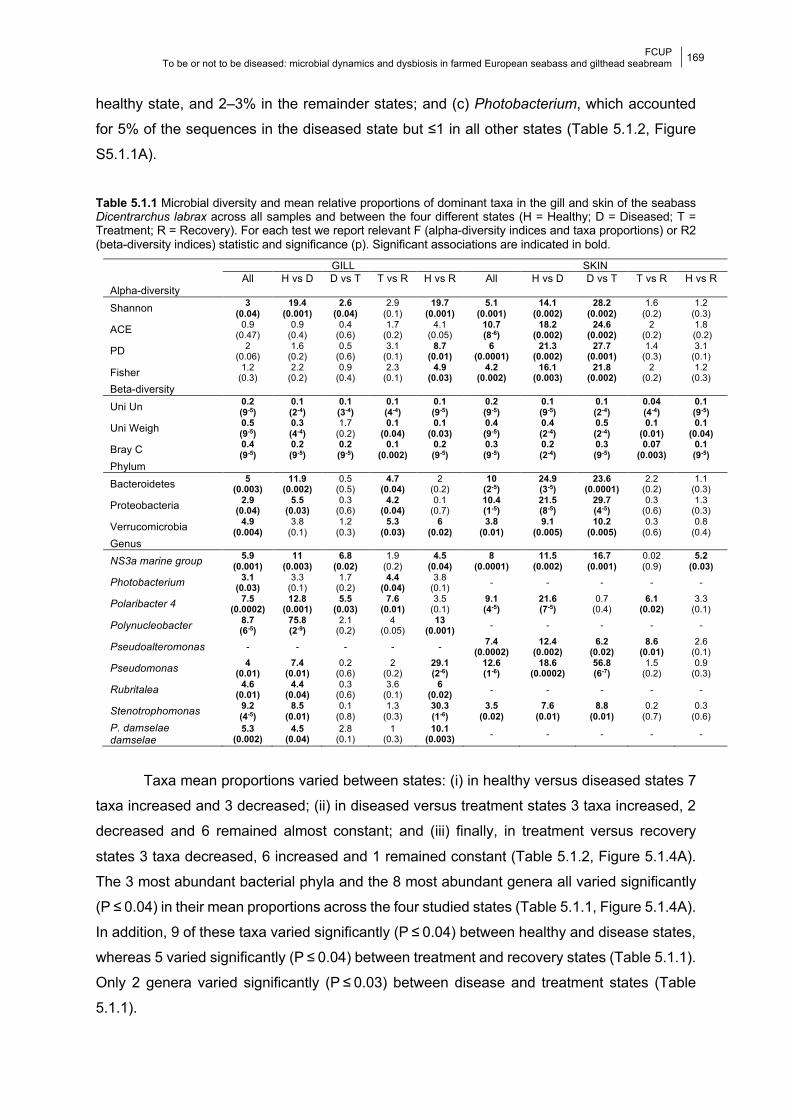
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
169
healthy state, and 2–3% in the remainder states; and (c) Photobacterium, which accounted
for 5% of the sequences in the diseased state but ≤1 in all other states (Table 5.1.2, Figure
S5.1.1A).
Table 5.1.1 Microbial diversity and mean relative proportions of dominant taxa in the gill and skin of the seabass Dicentrarchus labrax across all samples and between the four different states (H = Healthy; D = Diseased; T = Treatment; R = Recovery). For each test we report relevant F (alpha-diversity indices and taxa proportions) or R2 (beta-diversity indices) statistic and significance (p). Significant associations are indicated in bold.
GILL SKIN All H vs D D vs T T vs R H vs R All H vs D D vs T T vs R H vs R
Alpha-diversity Shannon 3
(0.04) 19.4
(0.001) 2.6
(0.04) 2.9
(0.1) 19.7
(0.001) 5.1
(0.001) 14.1
(0.002) 28.2
(0.002) 1.6
(0.2) 1.2
(0.3)
ACE 0.9 (0.47)
0.9 (0.4)
0.4 (0.6)
1.7 (0.2)
4.1 (0.05)
10.7 (8-6)
18.2 (0.002)
24.6 (0.002)
2 (0.2)
1.8 (0.2)
PD 2 (0.06)
1.6 (0.2)
0.5 (0.6)
3.1 (0.1)
8.7 (0.01)
6 (0.0001)
21.3 (0.002)
27.7 (0.001)
1.4 (0.3)
3.1 (0.1)
Fisher 1.2 (0.3)
2.2 (0.2)
0.9 (0.4)
2.3 (0.1)
4.9 (0.03)
4.2 (0.002)
16.1 (0.003)
21.8 (0.002)
2 (0.2)
1.2 (0.3)
Beta-diversity
Uni Un 0.2 (9-5)
0.1 (2-4)
0.1 (3-4)
0.1 (4-4)
0.1 (9-5)
0.2 (9-5)
0.1 (9-5)
0.1 (2-4)
0.04 (4-4)
0.1 (9-5)
Uni Weigh 0.5 (9-5)
0.3 (4-4)
1.7 (0.2)
0.1 (0.04)
0.1 (0.03)
0.4 (9-5)
0.4 (2-4)
0.5 (2-4)
0.1 (0.01)
0.1 (0.04)
Bray C 0.4 (9-5)
0.2 (9-5)
0.2 (9-5)
0.1 (0.002)
0.2 (9-5)
0.3 (9-5)
0.2 (2-4)
0.3 (9-5)
0.07 (0.003)
0.1 (9-5)
Phylum
Bacteroidetes 5 (0.003)
11.9 (0.002)
0.5 (0.5)
4.7 (0.04)
2 (0.2)
10 (2-5)
24.9 (3-5)
23.6 (0.0001)
2.2 (0.2)
1.1 (0.3)
Proteobacteria 2.9 (0.04)
5.5 (0.03)
0.3 (0.6)
4.2 (0.04)
0.1 (0.7)
10.4 (1-5)
21.5 (8-5)
29.7 (4-5)
0.3 (0.6)
1.3 (0.3)
Verrucomicrobia 4.9 (0.004)
3.8 (0.1)
1.2 (0.3)
5.3 (0.03)
6 (0.02)
3.8 (0.01)
9.1 (0.005)
10.2 (0.005)
0.3 (0.6)
0.8 (0.4)
Genus
NS3a marine group 5.9 (0.001)
11 (0.003)
6.8 (0.02)
1.9 (0.2)
4.5 (0.04)
8 (0.0001)
11.5 (0.002)
16.7 (0.001)
0.02 (0.9)
5.2 (0.03)
Photobacterium 3.1 (0.03)
3.3 (0.1)
1.7 (0.2)
4.4 (0.04)
3.8 (0.1) - - - - -
Polaribacter 4 7.5 (0.0002)
12.8 (0.001)
5.5 (0.03)
7.6 (0.01)
3.5 (0.1)
9.1 (4-5)
21.6 (7-5)
0.7 (0.4)
6.1 (0.02)
3.3 (0.1)
Polynucleobacter 8.7 (6-5)
75.8 (2-9)
2.1 (0.2)
4 (0.05)
13 (0.001) - - - - -
Pseudoalteromonas - - - - - 7.4 (0.0002)
12.4 (0.002)
6.2 (0.02)
8.6 (0.01)
2.6 (0.1)
Pseudomonas 4 (0.01)
7.4 (0.01)
0.2 (0.6)
2 (0.2)
29.1 (2-6)
12.6 (1-6)
18.6 (0.0002)
56.8 (6-7)
1.5 (0.2)
0.9 (0.3)
Rubritalea 4.6 (0.01)
4.4 (0.04)
0.3 (0.6)
3.6 (0.1)
6 (0.02) - - - - -
Stenotrophomonas 9.2 (4-5)
8.5 (0.01)
0.1 (0.8)
1.3 (0.3)
30.3 (1-6)
3.5 (0.02)
7.6 (0.01)
8.8 (0.01)
0.2 (0.7)
0.3 (0.6)
P. damselae damselae
5.3 (0.002)
4.5 (0.04)
2.8 (0.1)
1 (0.3)
10.1 (0.003) - - - - -
Taxa mean proportions varied between states: (i) in healthy versus diseased states 7
taxa increased and 3 decreased; (ii) in diseased versus treatment states 3 taxa increased, 2
decreased and 6 remained almost constant; and (iii) finally, in treatment versus recovery
states 3 taxa decreased, 6 increased and 1 remained constant (Table 5.1.2, Figure 5.1.4A).
The 3 most abundant bacterial phyla and the 8 most abundant genera all varied significantly
(P ≤ 0.04) in their mean proportions across the four studied states (Table 5.1.1, Figure 5.1.4A).
In addition, 9 of these taxa varied significantly (P ≤ 0.04) between healthy and disease states,
whereas 5 varied significantly (P ≤ 0.04) between treatment and recovery states (Table 5.1.1).
Only 2 genera varied significantly (P ≤ 0.03) between disease and treatment states (Table
5.1.1).
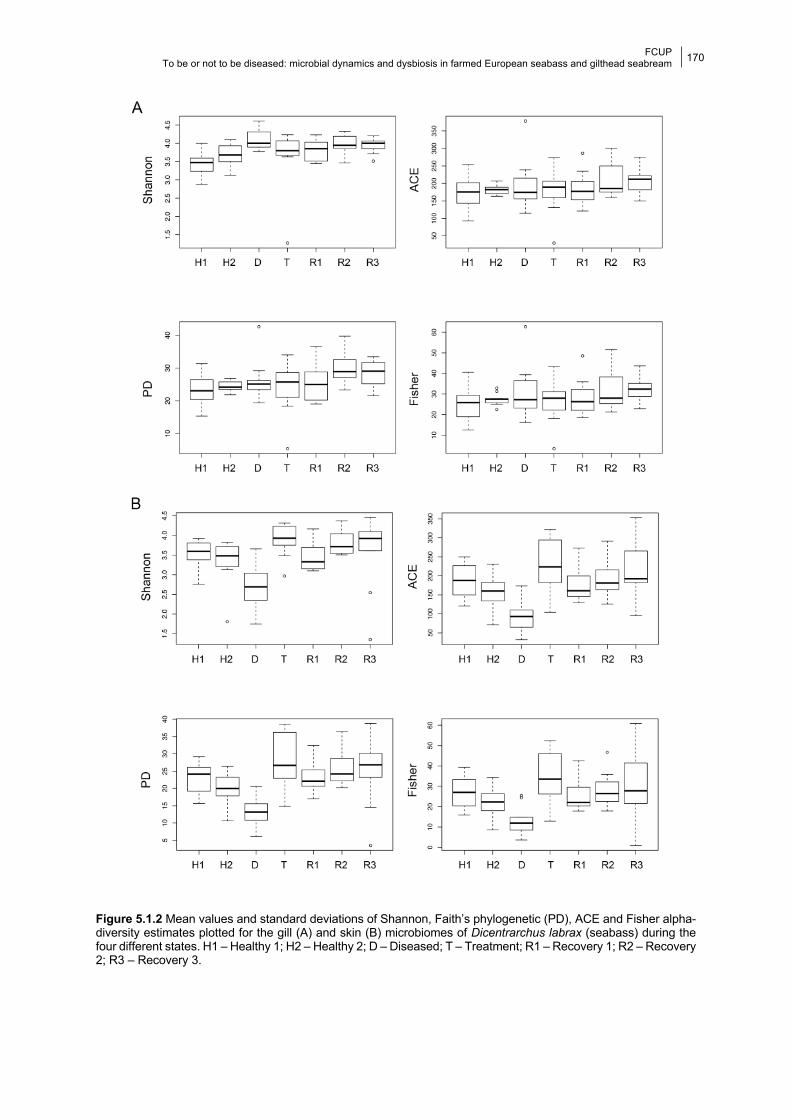
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
170
Figure 5.1.2 Mean values and standard deviations of Shannon, Faith’s phylogenetic (PD), ACE and Fisher alpha-diversity estimates plotted for the gill (A) and skin (B) microbiomes of Dicentrarchus labrax (seabass) during the four different states. H1 – Healthy 1; H2 – Healthy 2; D – Diseased; T – Treatment; R1 – Recovery 1; R2 – Recovery 2; R3 – Recovery 3.
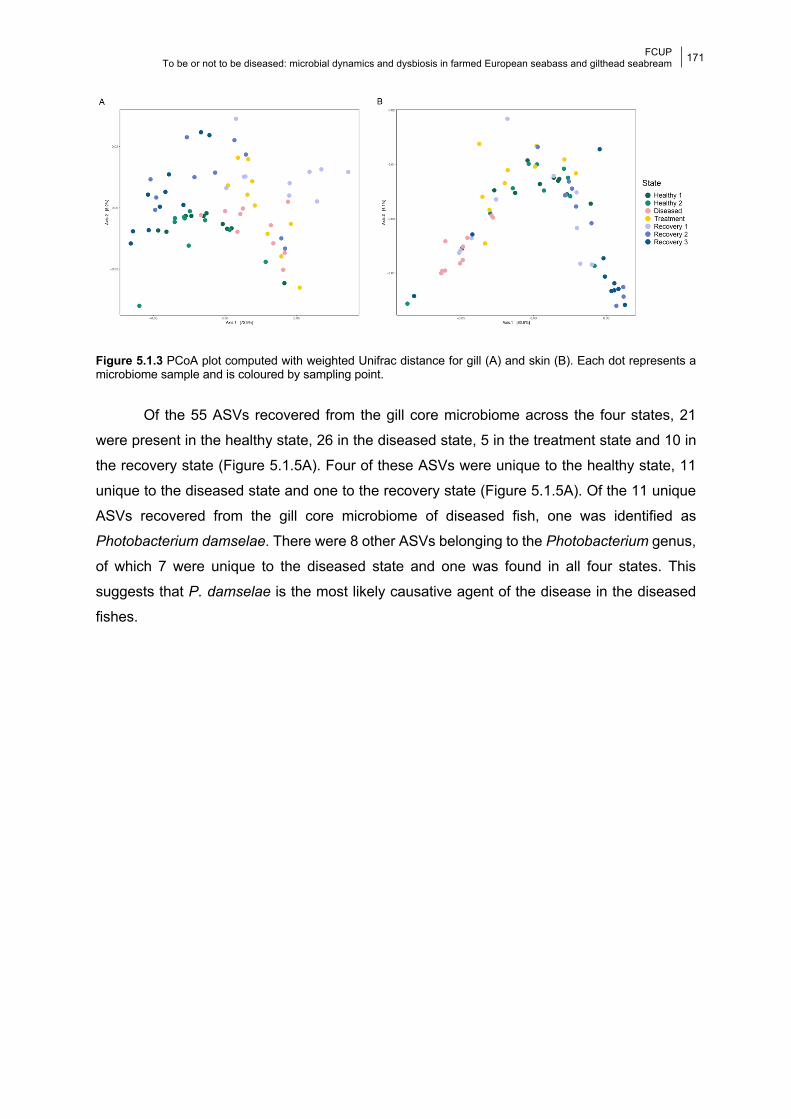
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
171
Figure 5.1.3 PCoA plot computed with weighted Unifrac distance for gill (A) and skin (B). Each dot represents a microbiome sample and is coloured by sampling point.
Of the 55 ASVs recovered from the gill core microbiome across the four states, 21
were present in the healthy state, 26 in the diseased state, 5 in the treatment state and 10 in
the recovery state (Figure 5.1.5A). Four of these ASVs were unique to the healthy state, 11
unique to the diseased state and one to the recovery state (Figure 5.1.5A). Of the 11 unique
ASVs recovered from the gill core microbiome of diseased fish, one was identified as
Photobacterium damselae. There were 8 other ASVs belonging to the Photobacterium genus,
of which 7 were unique to the diseased state and one was found in all four states. This
suggests that P. damselae is the most likely causative agent of the disease in the diseased
fishes.
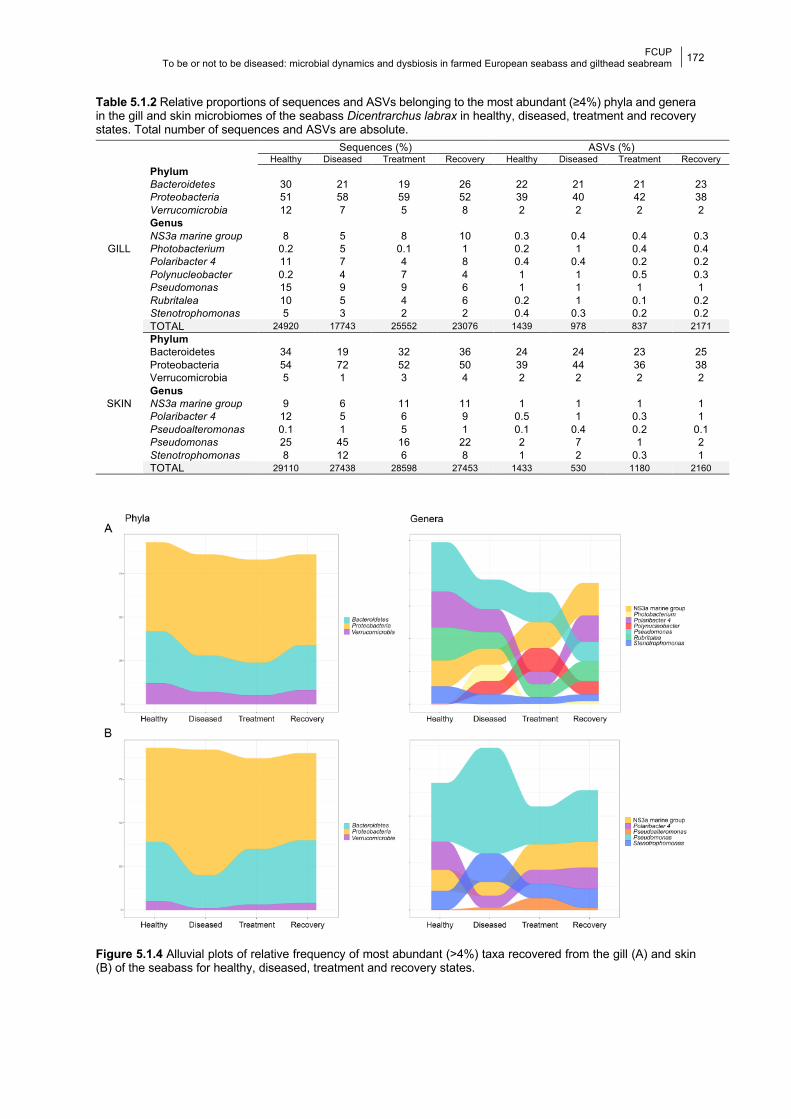
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
172
Table 5.1.2 Relative proportions of sequences and ASVs belonging to the most abundant (≥4%) phyla and genera in the gill and skin microbiomes of the seabass Dicentrarchus labrax in healthy, diseased, treatment and recovery states. Total number of sequences and ASVs are absolute.
Figure 5.1.4 Alluvial plots of relative frequency of most abundant (>4%) taxa recovered from the gill (A) and skin (B) of the seabass for healthy, diseased, treatment and recovery states.
Sequences (%) ASVs (%) Healthy Diseased Treatment Recovery Healthy Diseased Treatment Recovery
GILL
Phylum Bacteroidetes 30 21 19 26 22 21 21 23 Proteobacteria 51 58 59 52 39 40 42 38 Verrucomicrobia 12 7 5 8 2 2 2 2 Genus NS3a marine group 8 5 8 10 0.3 0.4 0.4 0.3 Photobacterium 0.2 5 0.1 1 0.2 1 0.4 0.4 Polaribacter 4 11 7 4 8 0.4 0.4 0.2 0.2 Polynucleobacter 0.2 4 7 4 1 1 0.5 0.3 Pseudomonas 15 9 9 6 1 1 1 1 Rubritalea 10 5 4 6 0.2 1 0.1 0.2 Stenotrophomonas 5 3 2 2 0.4 0.3 0.2 0.2 TOTAL 24920 17743 25552 23076 1439 978 837 2171
SKIN
Phylum Bacteroidetes 34 19 32 36 24 24 23 25 Proteobacteria 54 72 52 50 39 44 36 38 Verrucomicrobia 5 1 3 4 2 2 2 2 Genus NS3a marine group 9 6 11 11 1 1 1 1 Polaribacter 4 12 5 6 9 0.5 1 0.3 1 Pseudoalteromonas 0.1 1 5 1 0.1 0.4 0.2 0.1 Pseudomonas 25 45 16 22 2 7 1 2 Stenotrophomonas 8 12 6 8 1 2 0.3 1 TOTAL 29110 27438 28598 27453 1433 530 1180 2160

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
173
Figure 5.1.5 Core microbiota of seabass gill (A) and skin (B) at the ASV level. Distinctive bars represent relative abundance of each ASV for healthy, diseased, treatment and recovery states, labeled to the lowest taxonomic level possible.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
174
5.1.4.2 Skin bacterial composition and diversity Alpha-diversity estimates varied significantly across all states (RRPP, P ≤ 0.002; Table
5.1.1, Figure 5.1.2B) and between states in the skin microbiome. They decreased significantly
between healthy and diseased fish and increased significantly between diseased and
treatment states (RRPP, P ≤ 0.003; Table 5.1.1, Figure 5.1.2B). Beta-diversity estimates show
significant differences across all states and between states (Adonis, P < 0.05; Table 5.1.1).
As for the skin microbiome, Bacteroidetes, Proteobacteria and Verrucomicrobia were
the most abundant phyla retrieved across states, accounting for 87% to 93% of the sequences
altogether (Table 5.1.2). The genera NS3a marine group, Polaribacter 4, Pseudomonas and
Stenotrophomonas were the most abundant in all four states (Table 5.1.2, Figure S5.1.1B).
Moreover, Pseudoalteromonas accounted for 5% of the sequences in the treatment state, but
only 0.1–1% in the remaining states (Table 5.1.2, Figure S5.1.1B).
Mean proportions of the bacterial taxa varied significantly between states: (i) in healthy
versus diseased states 4 taxa increased and 4 decreased; (ii) in diseased versus treatment
states 5 taxa increased and 3 decreased; and (iii) in treatment versus recovery states 5 taxa
increased, 2 decreased and 1 remained constant (Table 5.1.2, Figure 5.1.4B). The 3 most
abundant phyla and 5 most abundant genera all varied significantly (P ≤ 0.03) across the four
states (Table 5.1.1, Figure 5.1.4B). All taxa varied significantly between healthy and diseased
states (P ≤ 0.01); all except one varied significantly between diseased and treatment states
(P ≤ 0.02); and 2 genera varied significantly (P ≤ 0.02) between treatment and recovery states
(Table 5.1.1).
A total of 43 ASVs formed the core microbiome of all four states, 17 were present in
the healthy state, 8 were present in the diseased state, 33 in the treatment state and 8 in the
recovery state (Figure 5.1.5B). It is worth noticing that 2 ASVs were unique to the healthy state
and 16 ASVs were unique to the treatment state.
5.1.5 Discussion In this study, we investigated the dynamics of the gill and skin microbiomes in 140
samples of the farmed seabass Dicentrarchus labrax during a natural disease outbreak and
subsequent antibiotic treatment with oxytetracycline. We used high-throughput sequencing
technology to generate 16S rRNA bacterial ASVs and examine changes in microbial
composition and diversity over six weeks. We identified Photobacterium damselae as the most
probable causative agent of disease.
The most abundant taxa found in the gill and skin microbiomes of healthy farmed
seabass (Dicentrarchus labrax) belonged to the Proteobacteria, Bacteroidetes and
Verrucomicrobia phyla. These phyla have been previously described as the most abundant in

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
175
the gill and skin microbiomes of several teleosts (Boutin et al., 2014; Legrand et al., 2018;
Lowrey et al., 2015; Tapia-Paniagua et al., 2018), including the seabass (Chiarello et al., 2015;
Pimentel et al., 2017; Rosado et al., 2019). At the genus level, the most abundant taxa were
the NS3a marine group, Polaribacter 4, Pseudomonas, and Stenotrophomonas in the gills and
skin (Table 5.1.2, Figure 5.1.4, Figure S5.1.1), and Rubritalea in the gills (Table 5.1.2, Figure
5.1.4, Figure S5.1.1). These results are mainly in accordance with previously described
microbiomes of healthy seabass (Pimentel et al., 2017; Rosado et al., 2019), including fish
retrieved from the same farmed population during winter months (Rosado et al., 2019).
However, one of the most abundant genera in the healthy seabass gill microbiome was
Polynucleobacter (Rosado et al., 2019), which in the present study only accounted for 0.2%
of the sequences in the healthy state, and 4–7% in the other three studied states (Table 5.1.2).
Another compositional difference was the high abundance of Stenotrophomonas found in both
tissues in apparently healthy individuals (Table 5.1.2) in this study, but not in Rosado et al.
(2019). Several environmental factors known to impact microbiome composition, such as
seasonality (de Bruijn et al., 2018; Larsen et al., 2015) and water temperature (Lokesh and
Kiron, 2016), could be driving these differences between our two studies.
The composition and diversity of the gill and skin seabass microbiomes varied
differently during infection. Whereas in the skin there was a significant decrease in alpha-
diversity between healthy and diseased fish, there were no significant differences in the gill
microbiome. An overall decrease in microbial richness was also reported for the skin of Atlantic
salmon as a result of infection with salmonid alphavirus (Reid et al., 2017) and sea lice
(Llewellyn et al., 2017); but interestingly, as in the present study, Legrand et al. (2018) reported
significant differences in microbial richness between the skin of healthy and enteritis-infected
yellowtail kingfish, but not in the gills.
Significant changes in beta-diversity occurred in both gills and skin, showing clear
signs of dysbiosis in both tissues. In the skin microbiome of diseased fish, the abundance of
taxa from the non-pathogenic NS3a marine group and Polaribacter 4 decreased, whereas the
pathogenic Pseudomonas and Stenotrophomonas significantly increased. Pseudomonas spp.
almost doubled their abundance and largely dominated the skin microbiome of diseased fish.
While the genus Stenotrophomonas contains important globally emergent fish pathogens (e.g.
Stenotrophomonas maltophilia; Abraham and Adikesavalu, 2016; Brooke, 2012),
Pseudomonas harbors both opportunist fish pathogens (e.g. P. baetica, P. chlororaphis, Hatai
et al., 1975; López et al., 2012; amongst others, Boutin et al., 2013; Pridgeon and Klesius,
2012) and taxa with known antimicrobial activity against fish pathogens (e.g. Flavobacterium
psychrophilum, Korkea-aho et al., 2011). For example, P. fluorescens is an important
pathogen of carp and salmon (Austin and Austin, 2016; Loch et al., 2012), but is also known
to inhibit the growth of Saprolegnia, an oomycete that causes huge losses in aquaculture (Liu

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
176
et al., 2015; van West, 2006). Importantly, a ten-fold increase of Pseudoalteromonas, which
was not amongst the most abundant taxa in healthy fish, occurred in the skin of diseased fish.
Species from this genus can inhibit the growth of both Vibrio spp. and Photobacterium
damselae (Lloyd and Pespeni, 2018; Offret et al., 2016; Papaleo et al., 2012; Reid et al., 2017;
Richards et al., 2017), hence an increase of Pseudoalteromonas could lead to a decrease of
the other two genera, as we have seen in the skin microbiomes of seabass transitioning from
healthy to diseased states (from 2% to 0.7% and from 0.3% to 0.2%, respectively). In the gills
of diseased fish, the majority of the most abundant bacterial genera in the healthy state (NS3a
marine group, Polaribacter 4, Pseudomonas, Rubritalea and Stenotrophomonas) decreased
significantly in abundance during infection, with the exception of Polynucleobacter. Amongst
the most abundant taxa in the gill, only Photobacterium spp. was exclusively associated with
diseased fish, where it showed a 25-fold increase. Similarly, all studies addressing the effects
of parasitic infection on fish microbiomes reported significant changes in microbial composition
(Legrand et al., 2018; Llewellyn et al., 2017; Reid et al., 2017). Importantly, all of these studies
reported an increase of potentially pathogenic taxa, which highlights the opportunistic nature
of such pathogens (Legrand et al., 2018; Llewellyn et al., 2017; Reid et al., 2017). Although
Photobacterium damselae was only highly abundant in the diseased gill microbiome, the
tissue with more significant shifts in overall bacterial composition (alpha-diversity) between
healthy and diseased states was the skin. This is not totally unexpected, since it has been
shown that this pathogen can unequally affect the microbiome of distinctive mucosal surfaces
such as the skin and gill (Legrand et al., 2018).
The effects of the disease in the core microbiomes were also significant and again
different between tissues, with a shift of core species in the gill and a decrease of core diversity
in the skin from healthy to diseased states. A shift of the microbial assemblages with
enrichment of specific groups was also described for the gill microbiome of the yellowtail
kingfish as a result of enteritis (Legrand et al., 2018).
Antibiotics administration can negatively impact host physiology in different ways (e.g.,
inhibiting mitochondrial gene expression, Morgun et al., 2015; decreasing enzymatic activity,
Chiu et al., 2013; leading to dysbiosis and the emergence of antibiotic resistant bacteria,
Carlson et al., 2017; Essam et al., 2016; Gaulke et al., 2016; Liu et al., 2012; Pindling et al.,
2018). Specifically, the reported effects of oxytetracycline in the gut microbiome of the Atlantic
salmon showed a clear reduction in taxonomic diversity, becoming almost exclusively
composed of the oxytetracycline resistant Aeromonas spp., which include the salmon
pathogens Aeromonas sobria and A. salmonicida (Navarrete et al., 2008). Similarly, in
zebrafish, long-term exposure (6 weeks) to environmental concentrations of oxytetracycline,
prompted both a decrease in gut microbial diversity and higher mortality when fish were
challenged with the pathogen A. hydrophila (Zhou et al., 2018). The impact of broad-spectrum

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
177
antibiotics in the skin microbiome of Gambusia affinis have also been assessed (Carlson et
al., 2017, 2015). In this case, the use of rifampicin led to a decrease of diversity in the skin
microbiome after 2.6 days of antibiotic administration. Additionally, as reported for zebrafish
and Atlantic salmon, fish subjected to rifampicin antibiotic administration were more
susceptible to infection due to osmotic stress and exhibited less growth compared to the
control group, an effect that lasted one month after treatment (Carlson et al., 2017, 2015). A
key difference with the present study is that the fish used by Carlson et al. (2017, 2015) were
healthy before antibiotic administration. Importantly, our results showed that skin core diversity
was higher in healthy than in recovery individuals, indicating a negative effect of disease and
antibiotic use.
In the present study, administration of oxytetracycline resulted in a dramatic reduction
of Photobacterium abundance in the gill microbiome, with this genus no longer being one of
the most abundant taxa in the treatment and recovery states. This was expected given the
reported sensitivity of P. damselae to several antibiotics, including oxytetracycline (Abdel-Aziz
et al., 2013). Pseudoalteromonas, however, remained one of the most abundant taxa in the
skin microbiome during treatment perhaps due to the host innate immune response mediated
by the skin microbiome, given the ability of this genus to produce antimicrobial metabolites
that are correlated with host homeostasis (Offret et al., 2016).
Previous studies on the impact of antibiotics on fish skin microbiomes showed that,
even though stabilization of bacterial communities during recovery occurs, neither diversity
nor composition returns to healthy-like values in the short term (after 1 week, Carlson et al.,
2017, 2015). Here the relative frequency of the most abundant taxa found in the skin
microbiome of the seabass during the recovery period, which corresponded to 3 weeks, was
similar to that in healthy individuals (P ≥ 0.1 for all taxa except the NS3a marine group). In the
gill microbiome, however, differences in taxa proportions between the healthy and recovery
states were significant for almost all of the most abundant taxa. Hence, although dysbiosis
due to infection was more noticeable in the skin than in the gill, the microbial communities
present in the skin seem to be more resilient than those of the gill. Importantly, although the
abundance of Photobacterium damselae in the gill seemed to have been controlled through
antibiotic administration, it increased significantly in the recovery state, surpassing its initial
proportion in the healthy state.
5.1.6 Conclusion In summary, the mucosal surfaces of fish, such as the gill and the skin, are constantly
exposed to several pathogens in the aquatic environment and are crucial to prevent and/or
control disease (Trivedi, 2012). It has been shown that both infectious diseases and antibiotic

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
178
treatment lead to a decrease in microbial diversity, which translates into a decrease in host
immunity (Carlson et al., 2017; Legrand et al., 2018). Here we described microbial changes in
the gill and skin of adult seabass in response to a natural disease outbreak followed by a
succeeding treatment with oxytetracycline. We showed that the gill and skin microbiomes are
highly disturbed by both infection and antibiotic treatment, ultimately decreasing their diversity.
5.1.7 Acknowledgements This work was funded by the European Regional Development Fund (ERDF) through the
COMPETE program and by National Funds through FCT - Foundation for Science and
Technology (project PTDC/MAR-BIO/0902/2014 -POCI-01-0145-FEDER-016550; project
PTDC/BIA-MIC/27995/2017 POCI-01-0145-FEDER-027995; and by a “Projecto de
Investigação Exploratória”: IF/00764/2013); the Welsh Government and Higher Education
Funding Council for Wales (HEFCW) AquaWales Project through the Sêr Cymru National
Research Network for Low Carbon Energy and Environment (NRN-LCEE). DR, MP-L and RX
were supported by FCT under the Programa Operacional Potencial Humano – Quadro de
Referência Estratégico Nacional funds from the European Social Fund and Portuguese
Ministério da Educação e Ciência (DR doctoral grant SFRH/BD/117943/2016; MPL:
IF/00764/2013; RX: IF/00359/2015).
5.1.8 References Abdel-Aziz, M., Eissa, A.E., Hanna, M., Okada, M.A., 2013. Identifying some pathogenic
Vibrio/Photobacterium species during mass mortalities of cultured Gilthead seabream
(Sparus aurata) and European seabass (Dicentrarchus labrax) from some Egyptian
coastal provinces. Int. J. Vet. Sci. Med. 1, 87–95.
https://doi.org/10.1016/j.ijvsm.2013.10.004
Abraham, T.J., Adikesavalu, H., 2016. Association of Stenotrophomonas maltophilia in African
Catfish, Clarias gariepinus (Burchell, 1822) Fry Mortalities with Dropsy. Int. J. Aquac. 6.
https://doi.org/10.5376/ija.2016.06.0013
Austin, B., Austin, D.A., 2016. Bacterial fish pathogens, 6th ed. Springer, Stirling, UK.
https://doi.org/10.1007/978-3-319-32674-0
Bakopoulos, V., Volpatti, D., Gusmani, L., Galeotti, M., Adams, A., Dimitriadis, G.J., 2003.
Vaccination trials of sea bass, Dicentrarchus labrax (L.), against Photobacterium
damsela subsp. piscicida, using novel vaccine mixtures. J. Fish Dis. 26, 77–90.
https://doi.org/10.1046/j.1365-2761.2003.00438.x
Boutin, S., Bernatchez, L., Audet, C., Derôme, N., 2013. Network Analysis Highlights Complex
Interactions between Pathogen, Host and Commensal Microbiota. PLoS One 8, e84772.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
179
https://doi.org/10.1371/journal.pone.0084772
Boutin, S., Sauvage, C., Bernatchez, L., Audet, C., Derome, N., 2014. Inter Individual
Variations of the Fish Skin Microbiota: Host Genetics Basis of Mutualism? PLoS One 9,
e102649. https://doi.org/10.1371/journal.pone.0102649
Brooke, J.S., 2012. Stenotrophomonas maltophilia: An emerging global opportunistic
pathogen. Clin. Microbiol. Rev. https://doi.org/10.1128/CMR.00019-11
Byadgi, O., Uyen, N.H.N., Chou, R.L., Guo, J.J., Lee, Y.H., Lee, J.W., Cheng, T.C., 2018.
Immunogenicity of inactivated formalin-killed Photobacterium damselae subsp. piscicida
combined with Toll-like receptor 9 agonist in Cobia Rachycentron canadum. Aquaculture
492, 369–378. https://doi.org/10.1016/j.aquaculture.2018.04.035
Califano, G., Castanho, S., Soares, F., Ribeiro, L., Cox, C.J., Mata, L., Costa, R., 2017.
Molecular Taxonomic Profiling of Bacterial Communities in a Gilthead Seabream (Sparus
aurata) Hatchery. Front. Microbiol. 8, 204. https://doi.org/10.3389/fmicb.2017.00204
Callahan, B.J., McMurdie, P.J., Rosen, M.J., Han, A.W., Johnson, A.J.A., Holmes, S.P., 2016.
DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13,
581–583. https://doi.org/10.1038/nmeth.3869
Caporaso, J.G., Lauber, C.L., Walters, W.A., Berg-Lyons, D., Lozupone, C.A., Turnbaugh,
P.J., Fierer, N., Knight, R., 2011. Global patterns of 16S rRNA diversity at a depth of
millions of sequences per sample. Proc. Natl. Acad. Sci. U. S. A. 108, 4516–4522.
https://doi.org/10.1073/pnas.1000080107
Carlson, J.M., Hyde, E.R., Petrosino, J.F., Manage, A.B.W., Primm, T.P., 2015. The host
effects of Gambusia affinis with an antibiotic-disrupted microbiome. Comp. Biochem.
Physiol. Part - C Toxicol. Pharmacol. 178, 163–168.
https://doi.org/10.1016/j.cbpc.2015.10.004
Carlson, J.M., Leonard, A.B., Hyde, E.R., Petrosino, J.F., Primm, T.P., 2017. Microbiome
disruption and recovery in the fish Gambusia affinis following exposure to broad-
spectrum antibiotic. Infect. Drug Resist. 10, 143–154.
https://doi.org/10.2147/IDR.S129055
Chiarello, M., Auguet, J.C., Bettarel, Y., Bouvier, C., Claverie, T., Graham, N.A.J.,
Rieuvilleneuve, F., Sucré, E., Bouvier, T., Villéger, S., 2018. Skin microbiome of coral
reef fish is highly variable and driven by host phylogeny and diet. Microbiome 6, 147.
https://doi.org/10.1186/s40168-018-0530-4
Chiarello, M., Villéger, S., Bouvier, C., Bettarel, Y., Bouvier, T., 2015. High diversity of skin-
associated bacterial communities of marine fishes is promoted by their high variability
among body parts, individuals and species. FEMS Microbiol. Ecol. 91.
https://doi.org/10.1093/femsec/fiv061
Chiu, T.-H., Kao, L.-Y., Chen, M.-L., 2013. Antibiotic resistance and molecular typing of

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
180
Photobacterium damselae subsp. damselae , isolated from seafood. J. Appl. Microbiol.
114, 1184–1192. https://doi.org/10.1111/jam.12104
de Bruijn, I., Liu, Y., Wiegertjes, G.F., Raaijmakers, J.M., 2018. Exploring fish microbial
communities to mitigate emerging diseases in aquaculture. FEMS Microbiol. Ecol. 94,
161. https://doi.org/10.1093/femsec/fix161
Enis Yonar, M., Mişe Yonar, S., Silici, S., 2011. Protective effect of propolis against oxidative
stress and immunosuppression induced by oxytetracycline in rainbow trout
(Oncorhynchus mykiss, W.). Fish Shellfish Immunol. 31, 318–325.
https://doi.org/10.1016/j.fsi.2011.05.019
Essam, H.M., Abdellrazeq, G.S., Tayel, S.I., Torky, H.A., Fadel, A.H., 2016. Pathogenesis of
Photobacterium damselae subspecies infections in sea bass and sea bream. Microb.
Pathog. 99, 41–50. https://doi.org/10.1016/j.micpath.2016.08.003
Faílde, L.D., Bermúdez, R., Losada, A.P., Riaza, A., Santos, Y., Quiroga, M.I., 2014.
Immunohistochemical diagnosis of tenacibaculosis in paraffin-embedded tissues of
Senegalese sole Solea senegalensis Kaup, 1858. J. Fish Dis. 37, 959–968.
https://doi.org/10.1111/jfd.12199
FAO, 2016. FAO Fisheries & Aquaculture - Cultured Aquatic Species Information Programme
- Dicentrarchus labrax (Linnaeus, 1758) [WWW Document]. URL
http://www.fao.org/fishery/culturedspecies/Dicentrarchus_labrax/en (accessed 2.12.21).
Galbraith, H., Iwanowicz, D., Spooner, D., Iwanowicz, L., Keller, D., Zelanko, P., Adams, C.,
2018. Exposure to synthetic hydraulic fracturing waste influences the mucosal bacterial
community structure of the brook trout (Salvelinus fontinalis) epidermis. AIMS Microbiol.
4, 413–427. https://doi.org/10.3934/microbiol.2018.3.413
Gaulke, C., Martins, M., Watral, V., Kent, M., Sharpton, T., 2016. Parasitic Infection by
Pseudocapillaria tomentosa is Associated with a Longitudinal Restructuring of the
Zebrafish Gut Microbiome. bioRxiv 076596. https://doi.org/10.1101/076596
Gilbert, J.A., Jansson, J.K., Knight, R., 2014. The Earth Microbiome project: Successes and
aspirations. BMC Biol. https://doi.org/10.1186/s12915-014-0069-1
Givens, C., Ransom, B., Bano, N., Hollibaugh, J., 2015. Comparison of the gut microbiomes
of 12 bony fish and 3 shark species. Mar. Ecol. Prog. Ser. 518, 209–223.
https://doi.org/10.3354/meps11034
Gómez, G.D., Balcázar, J.L., 2008. A review on the interactions between gut microbiota and
innate immunity of fish: Table 1. FEMS Immunol. Med. Microbiol. 52, 145–154.
https://doi.org/10.1111/j.1574-695X.2007.00343.x
Hatai, K., Egusa, S., Nakajima, M., Chikahata, H., 1975. Pseudomonas chlororaphis as a fish
pathogen. Bull. Japanese Soc. Sci. Fish. 41, 1203.
https://doi.org/10.2331/suisan.41.1203

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
181
Hess, S., Wenger, A.S., Ainsworth, T.D., Rummer, J.L., 2015. Exposure of clownfish larvae to
suspended sediment levels found on the Great Barrier Reef: Impacts on gill structure and
microbiome. Sci. Rep. 5, 1–8. https://doi.org/10.1038/srep10561
Kelly, C., Salinas, I., 2017. Under pressure: Interactions between commensal microbiota and
the teleost immune system. Front. Immunol. https://doi.org/10.3389/fimmu.2017.00559
Korkea-aho, T.L., Heikkinen, J., Thompson, K.D., von Wright, A., Austin, B., 2011.
Pseudomonas sp. M174 inhibits the fish pathogen Flavobacterium psychrophilum. J.
Appl. Microbiol. 111, 266–277. https://doi.org/10.1111/j.1365-2672.2011.05044.x
Kozich, J.J., Westcott, S.L., Baxter, N.T., Highlander, S.K., Schloss, P.D., 2013. Development
of a dual-index sequencing strategy and curation pipeline for analyzing amplicon
sequence data on the miseq illumina sequencing platform. Appl. Environ. Microbiol. 79,
5112–5120. https://doi.org/10.1128/AEM.01043-13
Labella, A., Berbel, C., Manchado, M., Castro, D., Borrego J J, 2011. Photobacterium
damselae subsp. damselae, an emerging pathogen affecting new cultured marine fish
species in southern Spain., in: Recent Advances in Fish Farms. InTech, pp. 135–152.
Larsen, A., Tao, Z., Bullard, S.A., Arias, C.R., 2013. Diversity of the skin microbiota of fishes:
evidence for host species specificity. FEMS Microbiol. Ecol. 85, 483–494.
https://doi.org/10.1111/1574-6941.12136
Larsen, A.M., Bullard, S.A., Womble, M., Arias, C.R., 2015. Community Structure of Skin
Microbiome of Gulf Killifish, Fundulus grandis, Is Driven by Seasonality and Not Exposure
to Oiled Sediments in a Louisiana Salt Marsh. Microb. Ecol. 70, 534–544.
https://doi.org/10.1007/s00248-015-0578-7
Lee, Y.K., Mazmanian, S.K., 2010. Has the microbiota played a critical role in the evolution of
the adaptive immune system? Science. 330, 6012.
https://doi.org/10.1126/science.1195568
Legrand, T.P.R.A., Catalano, S.R., Wos-Oxley, M.L., Stephens, F., Landos, M., Bansemer,
M.S., Stone, D.A.J., Qin, J.G., Oxley, A.P.A., 2018. The Inner Workings of the Outer
Surface: Skin and Gill Microbiota as Indicators of Changing Gut Health in Yellowtail
Kingfish. Front. Microbiol. 8, 2664. https://doi.org/10.3389/fmicb.2017.02664
Li, E., Xu, C., Wang, X., Wang, S., Zhao, Q., Zhang, M., Qin, J.G., Chen, L., 2018. Gut
Microbiota and its Modulation for Healthy Farming of Pacific White Shrimp Litopenaeus
vannamei. Rev. Fish. Sci. Aquac. 26, 381–399.
https://doi.org/10.1080/23308249.2018.1440530
Liu, Y., Rzeszutek, E., van der Voort, M., Wu, C.-H., Thoen, E., Skaar, I., Bulone, V.,
Dorrestein, P.C., Raaijmakers, J.M., de Bruijn, I., 2015. Diversity of Aquatic
Pseudomonas Species and Their Activity against the Fish Pathogenic Oomycete
Saprolegnia. PLoS One 10, e0136241. https://doi.org/10.1371/journal.pone.0136241

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
182
Liu, Y., Zhou, Z., Wu, N., Tao, Y., Xu, L., Cao, Y., Zhang, Y., Yao, B., 2012. Gibel carp
Carassius auratus gut microbiota after oral administration of trimethoprim/
sulfamethoxazole. Dis. Aquat. Organ. 99, 207–213. https://doi.org/10.3354/dao02477
Llewellyn, M.S., Boutin, S., Hoseinifar, S.H., Derome, N., 2014. Teleost microbiomes: The
state of the art in their characterization, manipulation and importance in aquaculture and
fisheries. Front. Microbiol. https://doi.org/10.3389/fmicb.2014.00207
Llewellyn, M.S., Leadbeater, S., Garcia, C., Sylvain, F.E., Custodio, M., Ang, K.P., Powell, F.,
Carvalho, G.R., Creer, S., Elliot, J., Derome, N., 2017. Parasitism perturbs the mucosal
microbiome of Atlantic Salmon. Sci. Rep. 7, 1–10. https://doi.org/10.1038/srep43465
Llewellyn, M.S., McGinnity, P., Dionne, M., Letourneau, J., Thonier, F., Carvalho, G.R., Creer,
S., Derome, N., 2016. The biogeography of the atlantic salmon (Salmo salar) gut
microbiome. ISME J. 10, 1280–1284. https://doi.org/10.1038/ismej.2015.189
Lloyd, M.M., Pespeni, M.H., 2018. Microbiome shifts with onset and progression of Sea Star
Wasting Disease revealed through time course sampling. Sci. Rep. 8, 16476.
https://doi.org/10.1038/s41598-018-34697-w
Loch, T.P., Scribner, K., Tempelman, R., Whelan, G., Faisal, M., 2012. Bacterial infections of
Chinook salmon, Oncorhynchus tshawytscha (Walbaum), returning to gamete collecting
weirs in Michigan. J. Fish Dis. 35, 39–50. https://doi.org/10.1111/j.1365-
2761.2011.01322.x
Lokesh, J., Kiron, V., 2016. Transition from freshwater to seawater reshapes the skin-
associated microbiota of Atlantic salmon. Sci. Rep. 6, 1–10.
https://doi.org/10.1038/srep19707
López, J.R., Diéguez, A.L., Doce, A., de la Roca, E., de la Herran, R., Navas, J.I., Toranzo,
A.E., Romalde, J.L., 2012. Pseudomonas baetica sp. nov., a fish pathogen isolated from
wedge sole, Dicologlossa cuneata (Moreau). Int. J. Syst. Evol. Microbiol. 62, 874–882.
https://doi.org/10.1099/ijs.0.030601-0
Lowrey, L., Woodhams, D.C., Tacchi, L., Salinas, I., 2015. Topographical mapping of the
rainbow trout (Oncorhynchus mykiss) microbiome reveals a diverse bacterial community
with antifungal properties in the skin. Appl. Environ. Microbiol. 81, 6915–6925.
https://doi.org/10.1128/AEM.01826-15
McMurdie, P.J., Holmes, S., 2014. Waste Not, Want Not: Why Rarefying Microbiome Data Is
Inadmissible. PLoS Comput. Biol. 10, e1003531.
https://doi.org/10.1371/journal.pcbi.1003531
McMurdie, P.J., Holmes, S., 2013. phyloseq: An R Package for Reproducible Interactive
Analysis and Graphics of Microbiome Census Data. PLoS One 8, e61217.
https://doi.org/10.1371/journal.pone.0061217
Mohammed, H.H., Arias, C.R., 2015. Potassium permanganate elicits a shift of the external

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
183
fish microbiome and increases host susceptibility to columnaris disease. Vet. Res. 46,
82. https://doi.org/10.1186/s13567-015-0215-y
Morgun, A., Dzutsev, A., Dong, X., Greer, R.L., Sexton, D.J., Ravel, J., Schuster, M., Hsiao,
W., Matzinger, P., Shulzhenko, N., 2015. Uncovering effects of antibiotics on the host
and microbiota using transkingdom gene networks. Gut 64, 1732–1743.
https://doi.org/10.1136/gutjnl-2014-308820
Narrowe, A.B., Albuthi-Lantz, M., Smith, E.P., Bower, K.J., Roane, T.M., Vajda, A.M., Miller,
C.S., 2015. Perturbation and restoration of the fathead minnow gut microbiome after low-
level triclosan exposure. Microbiome 3, 6. https://doi.org/10.1186/s40168-015-0069-6
Navarrete, P., Mardones, P., Opazo, R., Espejo, R., Romero, J., 2008. Oxytetracycline
treatment reduces bacterial diversity of intestinal microbiota of Atlantic salmon. J. Aquat.
Anim. Health 20, 177–183. https://doi.org/10.1577/H07-043.1
Nielsen, S., Walburn, J.W., Vergés, A., Thomas, T., Egan, S., 2017. Microbiome patterns
across the gastrointestinal tract of the rabbitfish Siganus fuscescens. PeerJ 2017, e3317.
https://doi.org/10.7717/peerj.3317
Nonaka, L., Maruyama, F., Miyamoto, M., Miyakoshi, M., Kurokawa, K., 2012. Novel
conjugative transferable multiple drug resistance plasmid pAQU1 from Photobacterium
damselae subsp. damselae isolated from marine aquaculture environment. Microbes
Environ. 27, 263–272.
Offret, C., Desriac, F., Le Chevalier, P., Mounier, J., Jégou, C., Fleury, Y., 2016. Spotlight on
Antimicrobial Metabolites from the Marine Bacteria Pseudoalteromonas: Chemodiversity
and Ecological Significance. Mar. Drugs 14, 129. https://doi.org/10.3390/md14070129
Oksanen, J., Kindt, R., Legendre, P., O’Hara, B., Simpson, G.L., Stevens, M.H.H., Wagner,
H., 2008. The vegan package: community ecology package, version 1.13-1].
Papaleo, M.C., Fondi, M., Maida, I., Perrin, E., Lo Giudice, A., Michaud, L., Mangano, S.,
Bartolucci, G., Romoli, R., Fani, R., 2012. Sponge-associated microbial Antarctic
communities exhibiting antimicrobial activity against Burkholderia cepacia complex
bacteria. Biotechnol. Adv. 30, 272–293. https://doi.org/10.1016/j.biotechadv.2011.06.011
Pedersen, K., Skall, H.F., Lassen-Nielsen, A.M., Bjerrum, L., Olesen, N.J., 2009.
Photobacterium damselae subsp. damselae , an emerging pathogen in Danish rainbow
trout, Oncorhynchus mykiss (Walbaum), mariculture. J. Fish Dis. 32, 465–472.
https://doi.org/10.1111/j.1365-2761.2009.01041.x
Pimentel, T., Marcelino, J., Ricardo, F., Soares, A.M.V.M., Calado, R., 2017. Bacterial
communities 16S rDNA fingerprinting as a potential tracing tool for cultured seabass
Dicentrarchus labrax. Sci. Rep. 7, 1–10. https://doi.org/10.1038/s41598-017-11552-y
Pindling, S., Azulai, D., Zheng, B., Dahan, D., Perron, G.G., 2018. Dysbiosis and early
mortality in zebrafish larvae exposed to subclinical concentrations of streptomycin. FEMS

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
184
Microbiol. Lett. 365, 188. https://doi.org/10.1093/femsle/fny188
Pratte, Z.A., Besson, M., Hollman, R.D., Stewart, F.J., 2018. The Gills of Reef Fish Support a
Distinct Microbiome Influenced by Host-Specific Factors. https://doi.org/10.1128/AEM
Pridgeon, J.W., Klesius, P.H., 2012. Major bacterial diseases in aquaculture and their vaccine
development. Anim. Sci. Rev. 7, 1–16.
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., Peplies, J., Glöckner,
F.O., 2013. The SILVA ribosomal RNA gene database project: Improved data processing
and web-based tools. Nucleic Acids Res. 41, D590–D596.
https://doi.org/10.1093/nar/gks1219
Reid, K.M., Patel, S., Robinson, A.J., Bu, L., Jarungsriapisit, J., Moore, L.J., Salinas, I., 2017.
Salmonid alphavirus infection causes skin dysbiosis in Atlantic salmon (Salmo salar L.)
post-smolts. PLoS One 12, e0172856. https://doi.org/10.1371/journal.pone.0172856
Richards, G.P., Watson, M.A., Needleman, D.S., Uknalis, J., Boyd, E.F., Fay, J.P., 2017.
Mechanisms for Pseudoalteromonas piscicidainduced killing of vibrios and other bacterial
pathogens. Appl. Environ. Microbiol. 83, 175–192. https://doi.org/10.1128/AEM.00175-
17
Rigos, G., Troisi, G.M., 2005. Antibacterial agents in Mediterranean finfish farming: A synopsis
of drug pharmacokinetics in important euryhaline fish species and possible environmental
implications. Rev. Fish Biol. Fish. https://doi.org/10.1007/s11160-005-7850-8
Rivas, A.J., Balado, M., Lemos, M.L., Osorio, C.R., 2011. The Photobacterium damselae
subsp. damselae hemolysins damselysin and HlyA are encoded within a new virulence
plasmid. Infect. Immun. 79, 4617–4627. https://doi.org/10.1128/IAI.05436-11
Rivas, A.J., Lemos, M.L., Osorio, C.R., 2013. Photobacterium damselae subsp. damselae, a
bacterium pathogenic for marine animals and humans. Front. Microbiol. 4, 283.
https://doi.org/10.3389/fmicb.2013.00283
Rodrigues, S., Antunes, S.C., Correia, A.T., Nunes, B., 2017. Rainbow trout (Oncorhynchus
mykiss) pro-oxidant and genotoxic responses following acute and chronic exposure to
the antibiotic oxytetracycline. Ecotoxicology 26, 104–117.
https://doi.org/10.1007/s10646-016-1746-3
Romalde, J.L., Ravelo, C., Valdés, I., Magariños, B., de la Fuente, E., Martín, C.S., Avendaño-
Herrera, R., Toranzo, A.E., 2008. Streptococcus phocae, an emerging pathogen for
salmonid culture. Vet. Microbiol. 130, 198–207.
https://doi.org/10.1016/j.vetmic.2007.12.021
Rosado, D., Pérez-Losada, M., Severino, R., Cable, J., Xavier, R., 2019. Characterization of
the skin and gill microbiomes of the farmed seabass (Dicentrarchus labrax) and
seabream (Sparus aurata). Aquaculture 500, 57–64.
https://doi.org/10.1016/j.aquaculture.2018.09.063

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
185
Rud, I., Kolarevic, J., Holan, A.B., Berget, I., Calabrese, S., Terjesen, B.F., 2017. Deep-
sequencing of the bacterial microbiota in commercial-scale recirculating and semi-closed
aquaculture systems for Atlantic salmon post-smolt production. Aquac. Eng. 78, 50–62.
https://doi.org/10.1016/j.aquaeng.2016.10.003
Studio, R., 2012. RStudio: integrated development environment for R.
Tao, Z., Shen, C., Zhou, S.M., Yang, N., Wang, G.L., Wang, Y.J., Xu, S.L., 2018. An outbreak
of Photobacterium damselae subsp. damselae infection in cultured silver pomfret
Pampus argenteus in Eastern China. Aquaculture 492, 201–205.
https://doi.org/10.1016/j.aquaculture.2018.04.013
Tapia-Paniagua, S.T., Ceballos-Francisco, D., Balebona, M.C., Esteban, M.Á., Moriñigo,
M.Á., 2018. Mucus glycosylation, immunity and bacterial microbiota associated to the
skin of experimentally ulcered gilthead seabream (Sparus aurata). Fish Shellfish
Immunol. 75, 381–390. https://doi.org/10.1016/j.fsi.2018.02.006
Terceti, M.S., Ogut, H., Osorio, C.R., 2016. Photobacterium damselae subsp. damselae, an
emerging fish pathogen in the Black Sea: Evidence of a multiclonal origin. Appl. Environ.
Microbiol. 82, 3736–3745. https://doi.org/10.1128/AEM.00781-16
Terceti, M.S., Vences, A., Matanza, X.M., Dalsgaard, I., Pedersen, K., Osorio, C.R., 2018.
Molecular Epidemiology of Photobacterium damselae subsp. damselae Outbreaks in
Marine Rainbow Trout Farms Reveals Extensive Horizontal Gene Transfer and High
Genetic Diversity. Front. Microbiol. 9, 2155. https://doi.org/10.3389/fmicb.2018.02155
Toranzo, A.E., Magariños, B., Romalde, J.L., 2005. A review of the main bacterial fish
diseases in mariculture systems. Aquaculture 246, 37–61.
https://doi.org/10.1016/j.aquaculture.2005.01.002
Trivedi, B., 2012. Microbiome: the surface brigade. Nature 492, S60–S61.
van West, P., 2006. Saprolegnia parasitica, an oomycete pathogen with a fishy appetite: new
challenges for an old problem. Mycologist 20, 99–104.
https://doi.org/10.1016/j.mycol.2006.06.004
Wang, J., Tao, Q., Wang, Z., Mai, K., Xu, W., Zhang, Y., Ai, Q., 2017. Effects of fish meal
replacement by soybean meal with supplementation of functional compound additives on
intestinal morphology and microbiome of Japanese seabass (Lateolabrax japonicus).
Aquac. Res. 48, 2186–2197. https://doi.org/10.1111/are.13055
Ye, L., Amberg, J., Chapman, D., Gaikowski, M., Liu, W.T., 2014. Fish gut microbiota analysis
differentiates physiology and behavior of invasive Asian carp and indigenous American
fish. ISME J. 8, 541–551. https://doi.org/10.1038/ismej.2013.181
Zha, Y., Eiler, A., Johansson, F., Svanbäck, R., 2018. Effects of predation stress and food
ration on perch gut microbiota. Microbiome 6, 1–12. https://doi.org/10.1186/s40168-018-
0400-0

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
186
Zhou, L., Limbu, S.M., Shen, M., Zhai, W., Qiao, F., He, A., Du, Z.Y., Zhang, M., 2018.
Environmental concentrations of antibiotics impair zebrafish gut health. Environ. Pollut.
235, 245–254. https://doi.org/10.1016/j.envpol.2017.12.073
Zlotkin, A., Hershko, H., Eldar, A., 1998. Possible transmission of Streptococcus iniae from
wild fish to cultured marine fish. Appl. Environ. Microbiol. 64, 4065–4067.
https://doi.org/10.1128/aem.64.10.4065-4067.1998
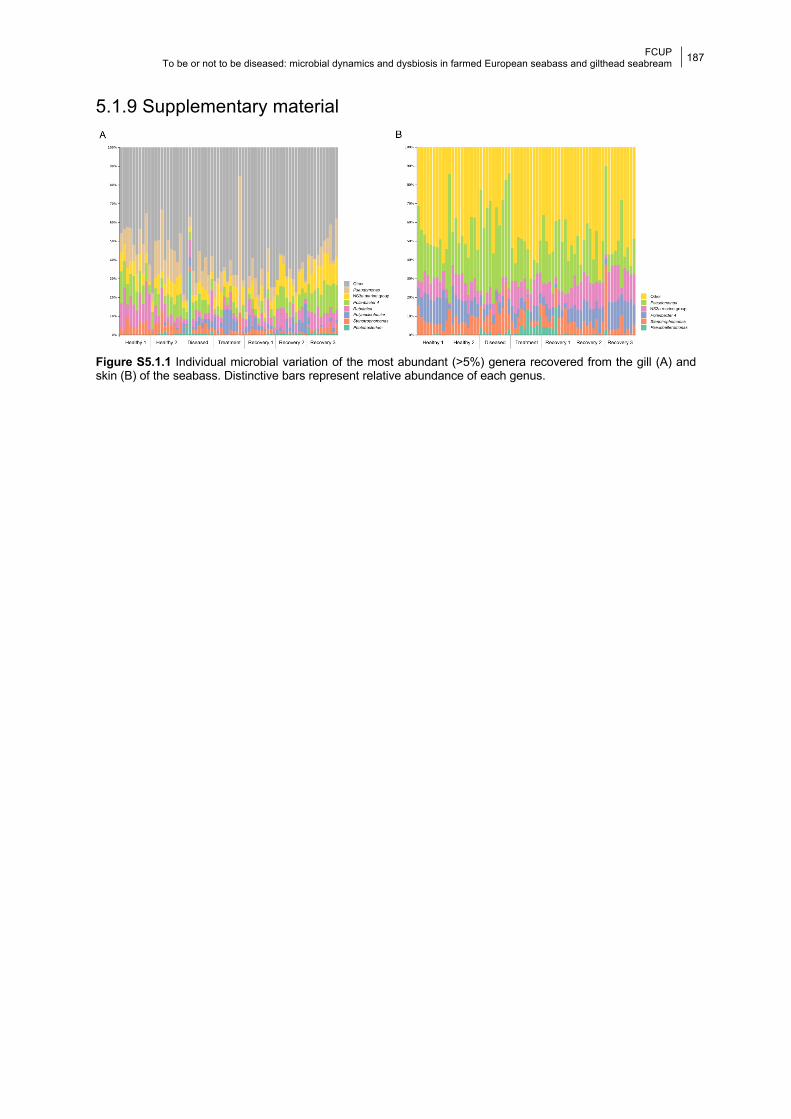
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
187
5.1.9 Supplementary material
Figure S5.1.1 Individual microbial variation of the most abundant (>5%) genera recovered from the gill (A) and skin (B) of the seabass. Distinctive bars represent relative abundance of each genus.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
188
5.2 Monitoring disease and antibiotic treatment in
the skin microbiota of farmed seabass fingerlings
Daniela Rosado, Marcos Pérez-Losada, Ricardo Severino, Raquel Xavier
2021. Microbial Ecology. https://doi.org/10.1007/s00248-021-01795-8.
5.2.1 Abstract The microbiota of fish skin, the primary barrier against disease, is highly dynamic and
modulated by several factors. In fish aquaculture, disease outbreaks occur mainly during
early-life stages, with associated high economic losses. Antibiotic treatments sometimes
remain the best option to control bacterial diseases, despite many reported negative impacts
of its use on fish and associated microbiota. Notwithstanding, studies monitoring the effects
of disease and antibiotic treatment on the microbiota of fingerlings are scarce. We sequenced
the bacterial 16S rRNA V4 gene region using a metabarcoding approach to assess the impact
of a mixed infection with Photobacterium damselae ssp. piscicida and Vibrio harveyi and
subsequent antibiotic treatment with flumequine, on the skin microbiota of farmed seabass
(Dicentrarchus labrax) fingerlings. Both infection and antibiotic treatment led to a significant
increase in bacterial diversity and core microbial communities and impacted microbiome
structure. Dysbiosis was confirmed by changes in the abundance of potential pathogenic and
opportunistic bacterial taxa. Skin bacterial metabolic function was also significantly affected
by flumequine administration, suggesting a detriment to fish skin health. Our results add to an
increasing body of literature, showing how fish microbiome response to infection and
antibiotics cannot be easily predicted.
Keywords: Flumequine, Dicentrarchus labrax, fish microbiota, Photobacterium damselae
ssp. piscicida, Vibrio harveyi, dysbiosis
5.2.2 Introduction Commensal microbiota are an essential part of the immune response of animals,
including fish, with a crucial role in disease prevention (Kelly and Salinas, 2017; Trivedi, 2012).
Perturbations to the homeostasis of the commensal organisms is termed dysbiosis and can
occur through three not mutually exclusive events linked to the occurrence of diseases
(Petersen and Round, 2014): i) pathobiont expansion, ii) reduced diversity, iii) and loss of

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
189
beneficial microbes. Most of the microbiome research in vertebrates is focused on the gut
microbiota due to their recognized role in sustaining the gut-brain axis and in disease outcome
(Osadchiy et al., 2019). However, in the case of fish, because pathogens are ubiquitous in the
aquatic environment, fish skin and associated mucus are considered the primary barrier
against diseases, with increasing numbers of studies focusing on the skin microbiota (e.g.,
Legrand et al., 2018; Llewellyn et al., 2017; Rosado et al., 2019). Evidence shows that fish
skin bacteria are highly dynamic, with composition and diversity being sensitive to both biotic
(e.g., ontogeny, Rosado et al., 2021) and abiotic factors (e.g., infection, Reid et al., 2017).
Although aquaculture is the fastest growing food-producing industry, disease
outbreaks due to pathogenic bacteria are one of the biggest constraints for its sustainability
(Borges et al., 2020). The impact of disease on the microbiota of several fish species has been
assessed, albeit being directed mainly at adult populations (e.g., skin and gill, Legrand et al.,
2018; Llewellyn et al., 2017; Reid et al., 2017; Rosado et al., 2019; gut and stomach,
Parshukov et al., 2019; Tran et al., 2018). It is well established that disease often leads to
dysbiosis, through a decrease in bacterial diversity and/or increases in the abundance of
pathogenic taxa other than the main etiological agent of disease (e.g., Le Luyer et al., 2021;
Legrand et al., 2018). Frequently, dysbiosis was also seen to lead to changes in predicted
microbial function (e.g., Llewellyn et al., 2017; Tran et al., 2018; Zhang et al., 2018). Although
disease and mortality incidences in aquaculture are higher on fry or young fingerlings, the
impact of disease on the microbiota during these early life stages is still poorly known (Borges
et al., 2020). Nevertheless, existing reports show that diversity, structure and potential function
of the microbiotas of early-life stages are also affected by disease (skin, Le Luyer et al., 2021;
gut, Vasemägi et al., 2017; gut, Miyake et al., 2020). The European seabass Dicentrarchus
labrax is susceptible to several bacterial pathogens, two of the most threatening being
Photobacterium damselae and Vibrio spp. (FAO, 2016). Both bacteria genera are usually
reported from the skin of healthy fish, and dysbiosis is usually accompanied by increases in
their abundances (e.g., European seabass, Rosado et al., 2021b; perl gentian grouper, Deng
et al., 2020). These pathogens cause photobacteriosis and vibriosis and infections can be
systemic, affecting multiple organs (Essam et al., 2016; Mohamad et al., 2019).
Although vaccines are available against major diseases, procedures are costly and
cause substantial stress and mortality, and they mostly confer short term immunity
(Bakopoulos et al., 2003). For these reasons, antibiotics are still widely used to control disease
in aquaculture. Several studies have reported the negative impacts of antibiotic use on fish,
which include behavioral changes (Almeida et al., 2019a), microbial diversity decrease
(Almeida et al., 2019b, 2019a; Rosado et al., 2019), increased susceptibility to secondary
infections (Carlson et al., 2017), changes in predicted microbial function (Almeida et al.,
2019b) and mortality (Pindling et al., 2018). Flumequine is a fluoroquinolone antibiotic active

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
190
against Gram-negative bacteria and widely used in aquaculture (De Liguoro et al., 2019).
Depending on water temperature, it can persist in fish skin and muscle up to 4-14 days since
the last administration (FAO, 1997). To the best of our knowledge, the impact of flumequine
in the microbiota of fish has never been assessed.
Here, we used a metataxonomic approach (Marchesi and Ravel, 2015) to characterize
the skin microbiota of the seabass Dicentrarchus labrax fingerlings before, during and after a
natural disease outbreak of Photobacterium damselae ssp. piscicida and Vibrio harveyi, and
subsequent antibiotic treatment with flumequine. Our goal was to describe changes in
composition, structure and potential function of the skin microbiota caused by disease and
subsequent antibiotic treatment. We predict that: i) disease will cause dysbiosis through a
decrease in the microbial diversity and core microbiota, while antibiotic treatment will have the
opposite effect; and ii) both disease and antibiotic treatment will lead to changes in the most
abundant bacterial taxa and differences in microbiome structure.
5.2.3 Material and methods
5.2.3.1 Experimental design, sampling and preparation Fish were reared in an open water circulation system in a semi-intensive farming
facility, where water is supplied from the Alvor Estuary (Southern Portugal). Sampled fish
belonged to the same age cohort and were sampled from the same rearing pond. Fish were
6 months old on the first sampling date, weighing on average 57 g, and 7 months old on the
last sampling date, weighing on average 70 g. Individuals were randomly caught using a
fishing pole, and skin samples were taken using tubed sterile swabs (Medical Wire &
Equipment, UK). We swabbed the right upper lateral part of the fish skin from head to tail.
Swabs were immediately stored at -20 °C until transported on dry ice to the CIBIO-InBIO
laboratory where they were kept at -80 °C until processed.
Individuals were collected once a week between August 23 and September 13, 2016,
encompassing four different sampling dates corresponding to four different fish health states:
healthy (N=30), diseased (N=30), treatment (N=30) and recovery (N=15) (Figure 5.2.1).
During the “healthy” state (sampled on August 23), all individuals were considered healthy due
to lack of visible disease symptoms. The second sampling point occurred on August 30, and
on August 31 fish began to die. Hence, samples collected on August 30 were categorized as
“diseased” (although still asymptomatic). Treatment with flumequine antibiotic was initiated on
August 31, being administered at 35 g/tns of fish through commercial feed until September 6.
Bacterial isolates from the liver, kidney and spleen of diseased fish were collected prior to the
start of antibiotic treatment and pathogens were identified via PCR by a commercial company
(Acuipharma, Spain). PCR amplification showed that the etiological agents of infection were

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
191
Photobacterium damselae ssp. piscicida and Vibrio harveyi. Fish were again sampled on the
last day of antibiotic administration (September 6) and these samples were categorized as
“treatment”. A final sampling point occurred on September 13 when fish were asymptomatic,
corresponding to the “recovery” state.
Total DNA from 105 fish samples and 6 controls (extraction kit negative controls) was
extracted using the PowerSoil DNA Isolation Kit (QIAGEN, Netherlands), following the
manufacturer’s protocol. Extraction kit negative controls were pooled into one single sample.
DNA extractions were shipped in dry ice to the University of Michigan Medical School (USA)
for amplification and sequencing on a single run of the Illumina MiSeq platform according to
the protocol of Kozich et al. (2013). Each sample plus 4 PCR blanks and 4 identical mock
communities (ZymoBIOMICS Microbial Community DNA Standard) were amplified and
sequenced for the V4 hyper-variable region of the 16S rRNA gene (~ 250 bp).
In total, 3,389,081 16S rRNA sequences were retrieved for the skin of the seabass
fingerlings, and the number of sequences per sample ranged from 12,891 to 48,211. ASVs
present in negative controls (extraction kit and PCR) were removed from downstream
analysis. After removal of contaminants and non-bacterial sequences, 6,163 ASVs (3,300,989
sequences) were assigned to the skin microbiota of the seabass fingerlings. Diversity and
bacterial abundances of the mock communities corresponded to those described by the
manufacturer. Microbial taxa showing a mean relative proportion ≥ 1% were considered as
part of the most abundant taxa in the microbiota.
5.2.3.2 Data and statistical analyses Raw FASTQ files were denoised using the DADA2 pipeline in R (Callahan et al., 2016).
We estimated a midpoint rooted tree of ASVs using the Quantitative Insights Into Microbial
Ecology 2 package (QIIME2; release 2020.11). We constructed a table containing amplicon
sequence variants (ASVs) and made taxonomic inferences against the SILVA (138 release)
reference database (Quast et al., 2013). We normalized ASV abundances using the negative
binomial distribution, which accounts for library size differences and biological variability
(McMurdie and Holmes, 2014). The core microbiota was assessed at the ASV level for each
health state (i.e., sampling date) separately. An ASV was considered as part of the core
microbiota if present in 100% of the samples from each state.
Microbial alpha-diversity was calculated using Shannon and Faith’s phylogenetic (PD)
diversity indices as implemented in the R phyloseq package (McMurdie and Holmes, 2013).
Additionally, Pielou’s evenness was calculated as implemented in the R microbiome package
(Lahti and Shetty, 2018). Microbiome structure (beta-diversity) was estimated using
phylogenetic UniFrac (weighted and unweighted) and Bray-Curtis distances. We assessed

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
192
variation in microbiome diversity and structure using Kruskal-Wallis and PERMANOVA
(adonis function of the R vegan package, Oksanen et al., 2008), respectively. Dissimilarity
between samples was visually assessed through a principal coordinates analysis (PCoA) and
dendrograms. A heatmap was built to depict changes in the abundance of the most abundant
phyla and genera (≥1% of all reads). All analyses were performed in R-studio v1.2.500.
Predicted bacterial metabolic functions were estimated using the metagenomic
Phylogenetic Investigation of Communities by Reconstruction of Unobserved States software
(PICRUSt2) embedded in QIIME2 (Douglas et al., 2019), applying a weighted nearest
sequenced taxon index (NSTI) cutoff of 0.03. Predicted metagenomes were collapsed using
the Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway metadata (Kanehisa et al.,
2019). We used linear discriminant analysis (LDA) in LEfSe to identify differentially abundant
metabolic pathways in the skin microbiota of the seabass fingerlings using state as a class, a
P-value cut-off of 0.05 and a LDA effect size cut-off of 2 (Segata et al., 2011).
5.2.4 Results
5.2.4.1 Microbial diversity and composition Alpha-diversity estimates varied significantly between states (P ≤ 3-3, Table 5.2.1),
with the exception of the PD index between treatment and recovery states, and the Pielou’s
evenness between healthy and recovery states (P = 0.2, Table 5.2.1). There was a significant
increase in diversity and evenness from healthy to diseased states as well as from diseased
to treatment states (Figure 5.2.2). Between treatment and recovery states, a significant
decrease in diversity and evenness occurred (Figure 5.2.2).
Figure 5.2.1 Schematic illustration of the experimental design and health status of each sampling point.
There were a total of 49 bacterial phyla and 926 bacterial genera identified across all
samples. Of those, 6 phyla and 32 genera were found in high abundance in at least one of the
health states (Figure 5.2.3, Table S5.2.1). The abundance of these taxa varied with the onset
of disease and antibiotic treatment (Figure 5.2.3, Table S5.2.1). It is noteworthy that the
abundance of Photobacterium (identified as P. damselae) in the skin remained stable between

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
193
healthy and disease states, having decreased with antibiotic treatment and then again in
recovery (Figure 5.2.3, Figure 5.2.4, Table S5.2.1). On the other hand, the abundance of Vibrio
increased from healthy to the disease state, having decreased with antibiotic treatment and
increased again in recovery (Figure 5.2.3, Figure 5.2.4, Table S5.2.1).
Table 5.2.1 Microbial diversity comparisons for the skin of the seabass Dicentrarchus labrax across all samples and between the four different states. For each Kruskall-Walis test (alpha-diversity) we report the chi-squared value (overall) and significance (P value, overall and pairwise) and for each PERMANOVA test (beta-diversity) we report the R2 statistics (overall) and significance (P value, overall and pairwise). Significant associations are indicated in bold.
Metric Overall Healthy/ Diseased
Diseased/ Treatment
Treatment/ Recovery
Healthy/ Recovery
Shannon 66 (3-14) 4-7 2-6 1-6 4-7 PD 55 (9-12) 6-7 3-3 0.2 3-9 Pielou’s 53 (2-11) 3-3 8-7 1-7 0.2 UniFrac weighted 0.3 (9-5) 7-2 6-3 6-3 0.1 UniFrac unweighted 0.1 (9-5) 6-3 6-3 6-3 6-3 Bray-Curtis 0.3 (9-5) 2-3 1-3 1-3 1-3
Figure 5.2.2 Alpha- and beta-diversity estimates plotted for the skin microbiota of the seabass Dicentrarchus labrax fingerlings during the different states. Alpha-diversity estimates are represented by boxplots depicting mean values and standard deviations of the Shannon, Faith’s phylogenetic (PD) and Pielou’s evenness estimates. Beta-diversity is represented by PCoA plots computed using the UniFrac (weighted and unweighted) and Bray-Curtis distances. Microbial structure based on the UniFrac unweighted distance (Ward method) is further represented by a dendrogram. Each dot in the PCoA and each line in the dendrogram represents a microbiota sample. All graphs are colored by state.
Of the 74 ASVs recovered from the core microbiota across the four states, 6 were
present in the healthy state, 20 in the diseased state, 30 in the treatment state and 67 in the
recovery state (Table S5.2.2). This corresponded to 0.4, 0.7, 1 and 4% of the total ASVs found
in the healthy, diseased, treatment and recovery states, respectively. There were 2 unique
core ASVs in the diseased and treatment states each, while 38 were unique to the recovery
state (Table S5.2.2). Of the 2 unique core ASVs recovered from the microbiota of diseased
fish, one was identified as Photobacterium damselae (Table S5.2.2). Additionally, there were
2 ASVs belonging to Vibrio that were part of the core microbiota during the diseased state, but

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
194
not during the healthy or treatment states. In the recovery state, those two and four other
Vibrio ASVs were part of the core microbiota of the skin.
Figure 5.2.3 Heatmaps of the most abundant (³ 1%) phyla and genera recovered from the skin of the seabass Dicentrarchus labrax fingerlings for healthy, diseased, treatment and recovery states. Unknown genera are labeled as u.g.
Beta-diversity estimates show significant differences between states (p ≤ 6-3, Table
5.2.1), except for the UniFrac weighted distance between healthy and recovery states (P =
0.1, Table 5.2.1). On the other hand, differences between states showed relatively low
dispersal (R < 0.3, Table 5.2.1). Analyses of the PCoA of Bray-Curtis distances showed that
samples from the healthy, treatment and recovery states clustered separately, while no
apparent structural cluster of samples between states was observed from the UniFrac
distances (PCoA, Figure 5.2.2). Moreover, the dendrogram constructed from the UniFrac
unweighted distance showed two main clusters separating healthy fish from the remainder
samples. Within the latter cluster, treatment and recovery states were more closely related
(Figure 5.2.2). Dendrograms constructed from the UniFrac unweighted and Bray-Curtis
distances showed no structure between the health states (Figure S5.2.1).
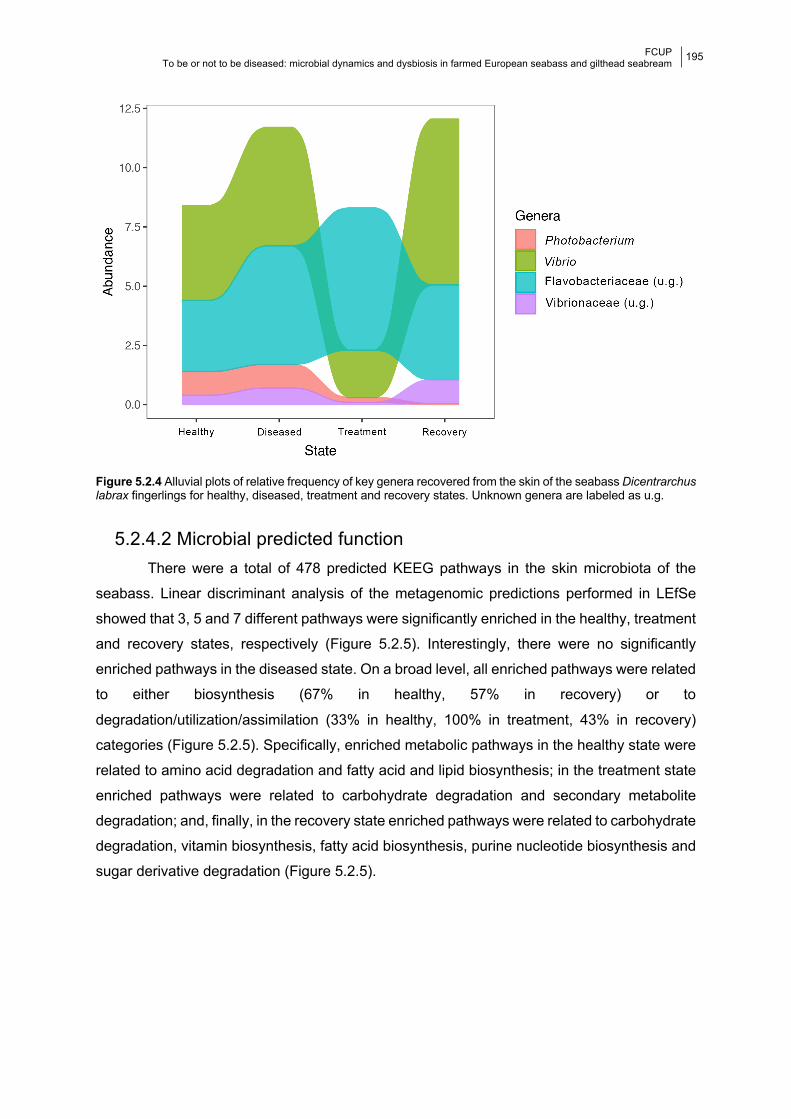
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
195
Figure 5.2.4 Alluvial plots of relative frequency of key genera recovered from the skin of the seabass Dicentrarchus labrax fingerlings for healthy, diseased, treatment and recovery states. Unknown genera are labeled as u.g.
5.2.4.2 Microbial predicted function There were a total of 478 predicted KEEG pathways in the skin microbiota of the
seabass. Linear discriminant analysis of the metagenomic predictions performed in LEfSe
showed that 3, 5 and 7 different pathways were significantly enriched in the healthy, treatment
and recovery states, respectively (Figure 5.2.5). Interestingly, there were no significantly
enriched pathways in the diseased state. On a broad level, all enriched pathways were related
to either biosynthesis (67% in healthy, 57% in recovery) or to
degradation/utilization/assimilation (33% in healthy, 100% in treatment, 43% in recovery)
categories (Figure 5.2.5). Specifically, enriched metabolic pathways in the healthy state were
related to amino acid degradation and fatty acid and lipid biosynthesis; in the treatment state
enriched pathways were related to carbohydrate degradation and secondary metabolite
degradation; and, finally, in the recovery state enriched pathways were related to carbohydrate
degradation, vitamin biosynthesis, fatty acid biosynthesis, purine nucleotide biosynthesis and
sugar derivative degradation (Figure 5.2.5).
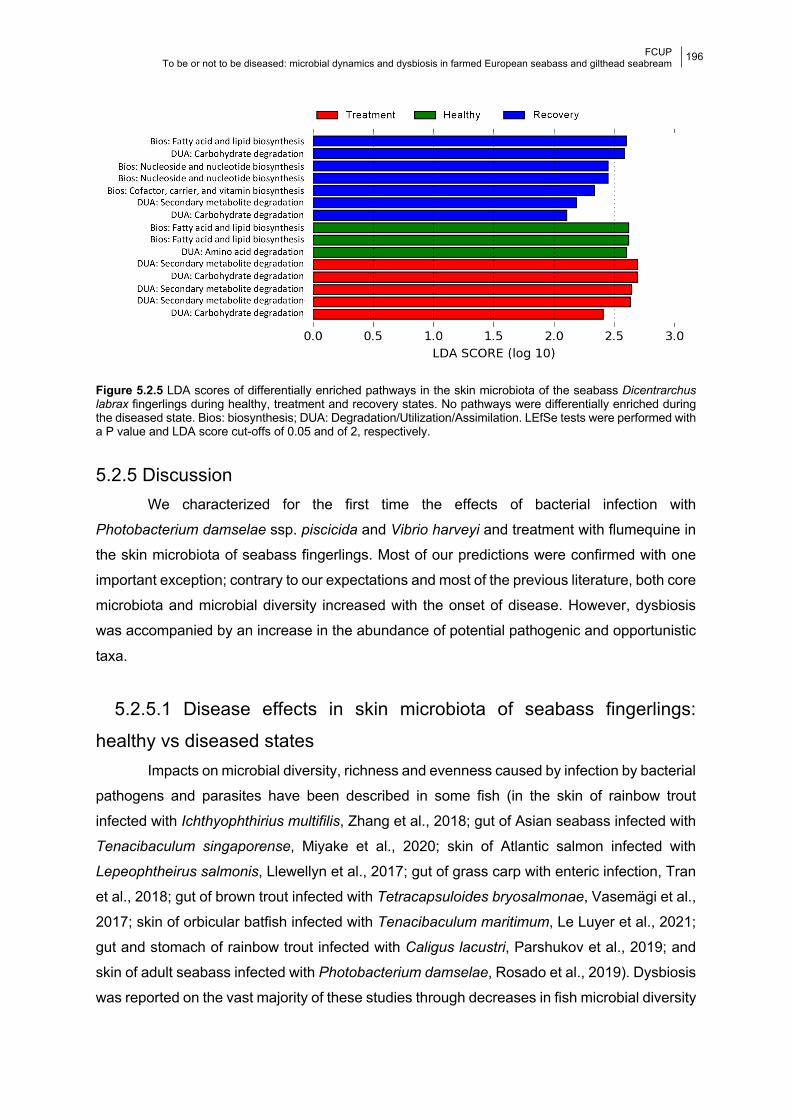
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
196
Figure 5.2.5 LDA scores of differentially enriched pathways in the skin microbiota of the seabass Dicentrarchus labrax fingerlings during healthy, treatment and recovery states. No pathways were differentially enriched during the diseased state. Bios: biosynthesis; DUA: Degradation/Utilization/Assimilation. LEfSe tests were performed with a P value and LDA score cut-offs of 0.05 and of 2, respectively.
5.2.5 Discussion We characterized for the first time the effects of bacterial infection with
Photobacterium damselae ssp. piscicida and Vibrio harveyi and treatment with flumequine in
the skin microbiota of seabass fingerlings. Most of our predictions were confirmed with one
important exception; contrary to our expectations and most of the previous literature, both core
microbiota and microbial diversity increased with the onset of disease. However, dysbiosis
was accompanied by an increase in the abundance of potential pathogenic and opportunistic
taxa.
5.2.5.1 Disease effects in skin microbiota of seabass fingerlings:
healthy vs diseased states Impacts on microbial diversity, richness and evenness caused by infection by bacterial
pathogens and parasites have been described in some fish (in the skin of rainbow trout
infected with Ichthyophthirius multifilis, Zhang et al., 2018; gut of Asian seabass infected with
Tenacibaculum singaporense, Miyake et al., 2020; skin of Atlantic salmon infected with
Lepeophtheirus salmonis, Llewellyn et al., 2017; gut of grass carp with enteric infection, Tran
et al., 2018; gut of brown trout infected with Tetracapsuloides bryosalmonae, Vasemägi et al.,
2017; skin of orbicular batfish infected with Tenacibaculum maritimum, Le Luyer et al., 2021;
gut and stomach of rainbow trout infected with Caligus lacustri, Parshukov et al., 2019; and
skin of adult seabass infected with Photobacterium damselae, Rosado et al., 2019). Dysbiosis
was reported on the vast majority of these studies through decreases in fish microbial diversity

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
197
and increases in pathobionts. Even though an increase in diversity was observed in the
present study, the direction of the changes in the abundance of key microbial taxa indicates
dysbiosis occurred in the skin microbiome of seabass fingerlings. In the present study, Vibrio
(that encompasses one of the etiological agents of infection), and two other unidentified
genera belonging to families with opportunistic taxa, Flavobacteriaceae and Vibrionaceae
(Austin and Austin, 2016), increased their abundance in the diseased state. Another genus
that increased in abundance in diseased fish was Aureispira, previously found to be highly
abundant in the intestinal microbiota of grouper juveniles after iridovirus infections (Joe et al.,
2021). Similar increases in bacterial diversity after disease have been already described in
other fish (e.g., Llewellyn et al., 2017; Tran et al., 2018; Vasemägi et al., 2017; Zhang et al.,
2018), indicating that changes in microbial diversity cannot be readily expected and growth or
decline of specific taxa easily predicted.
Alterations to the core microbiota in response to infection were also previously reported
in the skin of adult European seabass infected with Photobacterium damselae (Rosado et al.,
2019), and in the yellowtail kingfish infected with with enteritis (Legrand et al., 2018). In the
present study, a Photobacterium damselae ASV was 100% prevalent in the diseased state.
However, its mean abundance remained unaltered between the healthy and disease states,
suggesting the skin was only indirectly affected by this pathogen. These results are in line with
our previous study showing that infection caused by P. damselae can lead to dysbiosis of skin
microbiota of farmed seabass despite no increase in abundance (Rosado et al., 2019). Similar
results were also obtained by Legrand et al. (2018), who described skin and gill dysbiosis in
the yellowtail kingfish during enteritis, a gut disease.
Microbiome community structure of the skin microbiota was also significantly affected
by infection. Although samples of diseased fish were collected on the day prior to disease
onset, only a few samples from the diseased state (7 out of 30) clustered within the healthy
group, confirming that significant taxonomic changes had occurred in most individuals
analyzed. These results suggest that some properties of the skin mucous that allowed certain
phylogenetically related taxa to thrive in the skin may have been altered by disease,
consequently affecting resident microbiota. However, despite the increase in microbial
diversity and changes in structure driven by disease, microbial metabolic functions remained
unaltered. This suggests that the increase in diversity observed between healthy and diseased
states was due to colonization by bacteria with similar functions (Kelly and Salinas, 2017).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
198
5.2.5.2 Flumequine effects in skin microbiota of seabass fingerlings:
diseased vs treatment states In the present study, microbial diversity was observed to increase on the 8th day of
treatment with flumequine. However, as expected, administration of flumequine resulted in a
decrease in abundance of both etiological agents of disease in this study. This is unsurprising
given the reported sensitivity of both species to this antibiotic (Laganà et al., 2011).
Importantly, this treatment led to an increase of potentially harmful Flavobacteriaceae (Austin
and Austin, 2016). Interestingly, the genus Alteromonas, which has been shown to exhibit
antibacterial activity against fish pathogens, including Photobacterium damselae and several
Vibrio spp. (Stevens et al., 2016), and resistance against amoxicillin, erythromycin and
gentamicin (Wu et al., 2019), increased in abundance during antibiotic treatment. Microbial
disruptions have been reported after oxytetracycline and rifampicin treatment in microbiota of
adult fish (e.g., Almeida et al., 2019b, 2019a; Carlson et al., 2017; Rosado et al., 2019), and
after streptomycin, ciprofloxacin or oxytetracycline treatment on earlier life fish stages (e.g.,
zebrafish larvae, López Nadal et al., 2018; Pindling et al., 2018). Although an increase in fish
microbial diversity caused by antibiotic administration is less common, it has been reported
before (e.g., López Nadal et al., 2018; Rosado et al., 2019). Indeed, in the studies where
diversity decreased after antibiotic exposure, there were no pre-existing health conditions. On
the other hand, in this study as well as in Rosado et al. (2019), disease had occurred, and in
the study by López Nadal et al. (2018), fish were immersed with the anti-nutritional compound
saponin before antibiotic treatment. This suggests pre-existing disease/microbiota disruption
and antibiotics may have a compound effect on microbial diversity.
Significant changes in the potential function of the skin microbiota were detected after
antibiotic treatment. Specifically, the degradation of carbohydrates and secondary metabolites
were significantly enriched during antibiotic treatment. However, the production of
carbohydrates and secondary metabolites is linked to the protective role of the microbiota
(e.g., Hooper and Gordon, 2001; Kelly and Salinas, 2017). For example, carbohydrates are
directly related to specific cell-cell adhesion, modulating microbial binding to the mucus
(Brandley and Schnaar, 1986). It is suggested that carbohydrate synthesis by the human
microbiota helps establish symbiosis with microbial commensals and aids pathogenic evasion
(Hooper and Gordon, 2001). Furthermore, production of secondary metabolites is one of the
mechanisms by which commensal microbiota fight against pathogens (Kelly and Salinas,
2017). These results suggest a microbial response to antibiotics, which may ultimately have a
negative effect on fish immunity.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
199
5.2.5.3 Recovery of the skin microbiota of seabass fingerlings: healthy
vs recovery states Previous studies reported that short-term recovery of the microbiota of fish after
antibiotic treatment does not lead to the diversity levels observed in the healthy state (e.g., 1-
week recovery, Carlson et al., 2017). In the present study, with the exception of Pielou’s
evenness, diversity significantly increased between healthy and recovery states. Importantly,
the abundance of Vibrio increased in the recovery state, indicating microbial balance may not
have been fully obtained.
Microbiome structure was also significantly different between the recovery and healthy
states. However, closely related microbial structuring was found in fish from treatment and
recovery states. Almost half of the enriched metabolic pathways during the recovery state
were related to the same categories of pathways enriched during the treatment state
(carbohydrate and secondary metabolite degradation), with significant differences from the
healthy state. To the best of our knowledge, only the study by Brumlow et al. (2019) has
effectively measured the effects of 3-day antibiotic treatments (Tetracycline and Rifampicin)
in the biochemical profile of the skin of Gambusia affinis. In this study the authors also report
changes in community composition relative to pre-treatment, after 8 day recovery. However,
unlike the present results, where significant changes to microbial function were predicted, the
results of Brumlow et al. (2019) indicated that the biochemical functions of microbiota were
mostly reestablished after the 8 day recovery. Flumequine is a highly persistent antibiotic and
it can take several weeks to be fully depleted from the blood and tissues of fish (Hansen et al.,
2001; Sohlberg et al., 2002). This antibiotic has a slower depletion rate in the skin than in
muscle or liver, and can be present in the skin 20 days after oral administration (Malvisi et al.,
1997). Although a longer time frame would be necessary to evaluate whether full functional
recovery of the skin microbiota does occur, our results highlight the high susceptibility of skin
microbiota to antibiotic exposure.
5.2.6 Conclusion Homeostasis of the microbial communities in the mucosal surfaces of fish is central to
control pathogen abundances (Trivedi, 2012). Here, we describe a dysbiosis episode caused
by a disease outbreak induced by Photobacterium damselae ssp. piscicida and Vibrio harveyi,
and subsequent antibiotic treatment with flumequine. Although, overall, antibiotic treatment
appeared to have a greater impact on the skin microbiota when compared with the infection,
this could be a result of a cumulative effect of both disease and antibiotic treatment. Moreover,
the microbial profile of the fish in the recovery state differed from that of the fish in the healthy
state, as well as their predicted metabolic function. The results from this study highlight that

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
200
the response of fish commensal bacteria to disease and antibiotics are complex and not easily
predicted.
5.2.7 Acknowledgements This work was funded by the European Regional Development Fund (ERDF) through the
COMPETE program and by National Funds through FCT - Foundation for Science and
Technology (project PTDC/BIA-MIC/27995/2017 POCI-01-0145- FEDER-027995); DR, MP-L
and RX were supported by FCT under the Programa Operacional Potencial Humano – Quadro
de Referência Estratégico Nacional funds from the European Social Fund and Portuguese
Ministério da Educação e Ciência (DR doctoral grant SFRH/ BD/117943/2016; MP-L:
IF/00764/2013; RX: IF/00359/2015; and 2020.00854.CEECIND).
5.2.8 References Almeida, A.R., Alves, M., Domingues, I., Henriques, I., 2019a. The impact of antibiotic
exposure in water and zebrafish gut microbiomes: A 16S rRNA gene-based
metagenomic analysis. Ecotoxicol. Environ. Saf. 186, 109771.
https://doi.org/10.1016/j.ecoenv.2019.109771
Almeida, A.R., Tacão, M., Machado, A.L., Golovko, O., Zlabek, V., Domingues, I., Henriques,
I., 2019b. Long-term effects of oxytetracycline exposure in zebrafish: A multi-level
perspective. Chemosphere 222, 333–344.
https://doi.org/10.1016/j.chemosphere.2019.01.147
Austin, B., Austin, D.A., 2016. Bacterial fish pathogens, 6th ed. Springer, Stirling, UK.
https://doi.org/10.1007/978-3-319-32674-0
Bakopoulos, V., Volpatti, D., Gusmani, L., Galeotti, M., Adams, A., Dimitriadis, G.J., 2003.
Vaccination trials of sea bass, Dicentrarchus labrax (L.), against Photobacterium
damselae subsp. piscicida, using novel vaccine mixtures. J. Fish Dis. 26, 77–90.
https://doi.org/10.1046/j.1365-2761.2003.00438.x
Borges, N., Keller-Costa, T., Sanches-Fernandes, G.M.M., Louvado, A., Gomes, N.C.M.,
Costa, R., 2020. Bacteriome Structure, Function, and Probiotics in Fish Larviculture: The
Good, the Bad, and the Gaps. Annu. Rev. Anim. Biosci. 9, 423–452.
https://doi.org/10.1146/annurev-animal-062920
Brandley, B.K., Schnaar, R.L., 1986. Cell-Surface Carbohydrates in Cell Recognition and
Response. J. Leukoc. Biol. 40, 97–111. https://doi.org/10.1002/jlb.40.1.97
Brumlow, C.E., Luna, R.A., Hollister, E.B., Gomez, J.A., Burcham, L.A., Cowdrey, M.B.,
Primm, T.P., 2019. Biochemical but not compositional recovery of skin mucosal
microbiome communities after disruption. Infect. Drug Resist. 12, 399–416.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
201
https://doi.org/10.2147/IDR.S185992
Callahan, B.J., McMurdie, P.J., Rosen, M.J., Han, A.W., Johnson, A.J.A., Holmes, S.P., 2016.
DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13,
581–583. https://doi.org/10.1038/nmeth.3869
Carlson, J.M., Leonard, A.B., Hyde, E.R., Petrosino, J.F., Primm, T.P., 2017. Microbiome
disruption and recovery in the fish Gambusia affinis following exposure to broad-
spectrum antibiotic. Infect. Drug Resist. 10, 143–154.
https://doi.org/10.2147/IDR.S129055
De Liguoro, M., Maraj, S., Merlanti, R., 2019. Transgenerational toxicity of flumequine over
four generations of Daphnia magna. Ecotoxicol. Environ. Saf. 169, 814–821.
https://doi.org/10.1016/j.ecoenv.2018.11.077
Deng, Y., Zhang, Y., Chen, H., Xu, L., Wang, Q., Feng, J., 2020. Gut–Liver Immune Response
and Gut Microbiota Profiling Reveal the Pathogenic Mechanisms of Vibrio harveyi in
Pearl Gentian Grouper (Epinephelus lanceolatus ♂ × E. fuscoguttatus ♀). Front.
Immunol. 11, 1. https://doi.org/10.3389/fimmu.2020.607754
Douglas, G.M., Maffei, V.J., Zaneveld, J., Yurgel, S.N., Brown, J.R., Taylor, C.M.,
Huttenhower, C., Langille, M.G.I., 2019. PICRUSt2: An improved and extensible
approach for metagenome inference. bioRxiv. https://doi.org/10.1101/672295
Essam, H.M., Abdellrazeq, G.S., Tayel, S.I., Torky, H.A., Fadel, A.H., 2016. Pathogenesis of
Photobacterium damselae subspecies infections in sea bass and sea bream. Microb.
Pathog. 99, 41–50. https://doi.org/10.1016/j.micpath.2016.08.003
FAO, 2016. FAO Fisheries & Aquaculture - Cultured Aquatic Species Information Programme
- Dicentrarchus labrax (Linnaeus, 1758) [WWW Document]. URL
http://www.fao.org/fishery/culturedspecies/Dicentrarchus_labrax/en (accessed 2.12.21).
FAO, 1997. Residues of some veterinary drugs in animals and foods [WWW Document].
Hansen, M., Ingebrigtsen, K., Hayton, W., Horsberg, T., 2001. Disposition of 14C-flumequine
in eel Anguilla anguilla, turbot Scophthalmus maximus and halibut Hippoglossus
hippoglossus after oral and intravenous administration. Dis. Aquat. Organ. 47, 183–191.
https://doi.org/10.3354/dao047183
Hooper, L. V., Gordon, J.I., 2001. Glycans as legislators of host-microbial interactions:
spanning the spectrum from symbiosis to pathogenicity. Glycobiology 11, 1R-10R.
https://doi.org/10.1093/glycob/11.2.1R
Joe, J.T.X., Tseng, Y.C., Wu, J.L., Lu, M.W., 2021. The alteration of intestinal microbiota
profile and immune response in Epinephelus coioides during pathogen infection. Life 11,
1–18. https://doi.org/10.3390/life11020099
Kanehisa, M., Sato, Y., Furumichi, M., Morishima, K., Tanabe, M., 2019. New approach for
understanding genome variations in KEGG. Nucleic Acids Res. 47, D590–D595.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
202
https://doi.org/10.1093/nar/gky962
Kelly, C., Salinas, I., 2017. Under pressure: Interactions between commensal microbiota and
the teleost immune system. Front. Immunol. https://doi.org/10.3389/fimmu.2017.00559
Kozich, J.J., Westcott, S.L., Baxter, N.T., Highlander, S.K., Schloss, P.D., 2013. Development
of a dual-index sequencing strategy and curation pipeline for analyzing amplicon
sequence data on the miseq illumina sequencing platform. Appl. Environ. Microbiol. 79,
5112–5120. https://doi.org/10.1128/AEM.01043-13
Laganà, P., Caruso, G., Minutoli, E., Zaccone, R., Delia, S., 2011. Susceptibility to antibiotics
of Vibrio spp. and Photobacterium damselae ssp. piscicida strains isolated from Italian
aquaculture farms. New Microbiol. 34, 53–63.
Lahti, L., Shetty, S., 2018. Introduction to the microbiome R package [WWW Document].
Le Luyer, J., Schull, Q., Auffret, P., Lopez, P., Crusot, M., Belliard, C., Basset, C., Carradec,
Q., Poulain, J., Planes, S., Saulnier, D., 2021. Dual RNAseq Highlights the Kinetics of
Skin Microbiome and Fish Host Responsiveness to Bacterial Infection.
https://doi.org/10.21203/rs.3.rs-108248/v1
Legrand, T.P.R.A., Catalano, S.R., Wos-Oxley, M.L., Stephens, F., Landos, M., Bansemer,
M.S., Stone, D.A.J., Qin, J.G., Oxley, A.P.A., 2018. The Inner Workings of the Outer
Surface: Skin and Gill Microbiota as Indicators of Changing Gut Health in Yellowtail
Kingfish. Front. Microbiol. 8, 2664. https://doi.org/10.3389/fmicb.2017.02664
Llewellyn, M.S., Leadbeater, S., Garcia, C., Sylvain, F.E., Custodio, M., Ang, K.P., Powell, F.,
Carvalho, G.R., Creer, S., Elliot, J., Derome, N., 2017. Parasitism perturbs the mucosal
microbiome of Atlantic Salmon. Sci. Rep. 7, 1–10. https://doi.org/10.1038/srep43465
López Nadal, A., Peggs, D., Wiegertjes, G.F., Brugman, S., 2018. Exposure to Antibiotics
Affects Saponin Immersion-Induced Immune Stimulation and Shift in Microbial
Composition in Zebrafish Larvae. Front. Microbiol. 9, 2588.
https://doi.org/10.3389/fmicb.2018.02588
Malvisi, J., Della Rocca, G., Anfossi, P., Giorgetti, G., 1997. Tissue distribution and depletion
of flumequine after in-feed administration in sea bream (Sparus aurata). Aquaculture 157,
197–204. https://doi.org/10.1016/S0044-8486(97)00160-9
Marchesi, J.R., Ravel, J., 2015. The vocabulary of microbiome research: a proposal.
Microbiome 3, 31. https://doi.org/10.1186/s40168-015-0094-5
McMurdie, P.J., Holmes, S., 2014. Waste Not, Want Not: Why Rarefying Microbiome Data Is
Inadmissible. PLoS Comput. Biol. 10, e1003531.
https://doi.org/10.1371/journal.pcbi.1003531
McMurdie, P.J., Holmes, S., 2013. phyloseq: An R Package for Reproducible Interactive
Analysis and Graphics of Microbiome Census Data. PLoS One 8, e61217.
https://doi.org/10.1371/journal.pone.0061217

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
203
Miyake, S., Soh, M., Nursyafiq Azman, M., Yan Ngoh, S., Orbán, L., Seedorf, H., 2020.
Insights into the microbiome of farmed Asian sea bass (Lates calcarifer) with symptoms
of tenacibaculosis and description of Tenacibaculum singaporense sp. nov. Antonie Van
Leeuwenhoek 113, 737–752. https://doi.org/10.1007/s10482-020-01391-9
Mohamad, N., Amal, M.N.A., Yasin, I.S.M., Zamri Saad, M., Nasruddin, N.S., Al-saari, N.,
Mino, S., Sawabe, T., 2019. Vibriosis in cultured marine fishes: a review. Aquaculture
512, 734289. https://doi.org/10.1016/j.aquaculture.2019.734289
Oksanen, J., Kindt, R., Legendre, P., O’Hara, B., Simpson, G.L., Stevens, M.H.H., Wagner,
H., 2008. The vegan package: community ecology package, version 1.13-1 [WWW
Document].
Osadchiy, V., Martin, C.R., Mayer, E.A., 2019. The Gut–Brain Axis and the Microbiome:
Mechanisms and Clinical Implications. Clin. Gastroenterol. Hepatol.
https://doi.org/10.1016/j.cgh.2018.10.002
Parshukov, A.N., Kashinskaya, E.N., Simonov, E.P., Hlunov, O.V., Izvekova, G.I., Andree,
K.B., Solovyev, M.M., 2019. Variations of the intestinal gut microbiota of farmed rainbow
trout, Oncorhynchus mykiss (Walbaum), depending on the infection status of the fish. J.
Appl. Microbiol. 127, 379–395. https://doi.org/10.1111/jam.14302
Petersen, C., Round, J.L., 2014. Defining dysbiosis and its influence on host immunity and
disease. Cell. Microbiol. 16, 1024–1033. https://doi.org/10.1111/cmi.12308
Pindling, S., Azulai, D., Zheng, B., Dahan, D., Perron, G.G., 2018. Dysbiosis and early
mortality in zebrafish larvae exposed to subclinical concentrations of streptomycin. FEMS
Microbiol. Lett. 365, 188. https://doi.org/10.1093/femsle/fny188
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., Peplies, J., Glöckner,
F.O., 2013. The SILVA ribosomal RNA gene database project: Improved data processing
and web-based tools. Nucleic Acids Res. 41, D590–D596.
https://doi.org/10.1093/nar/gks1219
Reid, K.M., Patel, S., Robinson, A.J., Bu, L., Jarungsriapisit, J., Moore, L.J., Salinas, I., 2017.
Salmonid alphavirus infection causes skin dysbiosis in Atlantic salmon (Salmo salar L.)
post-smolts. PLoS One 12, e0172856. https://doi.org/10.1371/journal.pone.0172856
Rosado, D., Pérez-Losada, M., Pereira, A., Severino, R., Xavier, R., 2021a. Effects of aging
on the skin and gill microbiota of farmed seabass and seabream. Anim. Microbiome 3.
https://doi.org/10.1186/s42523-020-00072-2
Rosado, D., Xavier, R., Cable, J., Severino, R., Tarroso, P., Pérez-Losada, M., 2021b.
Longitudinal sampling of external mucosae in farmed European seabass reveals the
impact of water temperature on bacterial dynamics. ISME Commun.
Rosado, D., Xavier, R., Severino, R., Tavares, F., Cable, J., Pérez-Losada, M., 2019. Effects
of disease, antibiotic treatment and recovery trajectory on the microbiome of farmed

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
204
seabass (Dicentrarchus labrax). Sci. Rep. 9, 1–11. https://doi.org/10.1038/s41598-019-
55314-4
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W.S., Huttenhower, C.,
2011. Metagenomic biomarker discovery and explanation. Genome Biol. 12, R60.
https://doi.org/10.1186/gb-2011-12-6-r60
Sohlberg, S., Ingebrigtsen, K., Hansen, M., Hayton, W., Horsberg, T., 2002. Flumequine in
Atlantic salmon Salmo salar: disposition in fish held in sea water versus fresh water. Dis.
Aquat. Organ. 49, 39–44. https://doi.org/10.3354/dao049039
Stevens, J., Jackson, R., Olson, J., 2016. Bacteria associated with lionfish (Pterois
volitans/miles complex) exhibit antibacterial activity against known fish pathogens. Mar.
Ecol. Prog. Ser. 558, 167–180. https://doi.org/10.3354/meps11789
Tran, N.T., Zhang, J., Xiong, F., Wang, G.T., Li, W.X., Wu, S.G., 2018. Altered gut microbiota
associated with intestinal disease in grass carp (Ctenopharyngodon idellus). World J.
Microbiol. Biotechnol. 34, 71. https://doi.org/10.1007/s11274-018-2447-2
Trivedi, B., 2012. Microbiome: the surface brigade. Nature 492, S60–S61.
Vasemägi, A., Visse, M., Kisand, V., 2017. Effect of Environmental Factors and an Emerging
Parasitic Disease on Gut Microbiome of Wild Salmonid Fish. mSphere 2, 418–435.
https://doi.org/10.1128/msphere.00418-17
Wu, J., Mao, C., Deng, Y., Guo, Z., Liu, G., Xu, L., Bei, L., Su, Y., Feng, J., 2019. Diversity
and abundance of antibiotic resistance of bacteria during the seedling period in marine
fish cage-culture areas of Hainan, China. Mar. Pollut. Bull. 141, 343–349.
https://doi.org/10.1016/j.marpolbul.2019.02.069
Zhang, Xiaoting, Ding, L., Yu, Y., Kong, W., Yin, Y., Huang, Z., Zhang, Xuezhen, Xu, Z., 2018.
The Change of Teleost Skin Commensal Microbiota Is Associated With Skin Mucosal
Transcriptomic Responses During Parasitic Infection by Ichthyophthirius multifillis. Front.
Immunol. 9, 2972. https://doi.org/10.3389/fimmu.2018.02972
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
205
5.2.9 Supplementary material Table S5.2.1 Relative mean proportions of sequences (%) belonging to the most abundant (≥ 1%) phyla and genera in the skin microbiota of the seabass Dicentrarchus labrax in healthy (N=30), diseased (N=30), treatment (N=30) and recovery states (N=15). Taxa with ≥ 1% relative mean proportion in any state are indicated in bold. Unknown genera are labeled as u.g.
Healthy Diseased Treatment Recovery Phyla Actinobacteriota 1 0,7 0,6 0,2 Bacteroidota 21 29 37 28 Chloroflexi 0,4 0,4 1 1 Fusobacteriota 0,2 0,3 0,8 1 Proteobacteria 74 63 52 63 Verrucomicrobiota 1 2 4 3 Genera Alkalimarinus 1 1 1 1 Alteromonas 2 2 5 3 Arcticiflavibacter 0,9 1 2 1 Aureispira 0,3 2 0,2 0,4 Fabibacter 0,4 0,5 1 0,3 Formosa 0,2 0,9 1 0,6 Maribacter 0,3 1 2 1 Marinomonas 1 0,9 2 1 NS10 marine group 0,8 3 3 0,5 NS3a marine group 0,5 0,2 0,8 1 Oleispira 8 9 3 13 Photobacterium 1 1 0,2 0,1 Polaribacter 5 3 4 7 Pseudoalteromonas 17 12 12 19 Pseudomonas 19 11 6 1 Psychrobium 1 1 0,2 0,3 Psychrosphaera 1 0,9 2 0,9 Rubritalea 0,3 0,2 0,9 1 Ruegeria 0,2 0,5 1 0,5 Stenotrophomonas 5 3 2 0.4 Tenacibaculum 2 2 1 0,4 Thalassotalea 3 1 2 2 Vibrio 4 5 2 7 Winogradskyella 0,2 0,5 0,9 0,4 Alteromonadaceae (u.g.) 0,1 0,1 2 0,4 Cryomorphaceae (u.g.) 3 2 2 2 Flavobacteriaceae (u.g.) 3 5 6 4 Rhodobacteraceae (u.g.) 0,5 1 1 0,7 Saprospiraceae (u.g.) 0,4 0,8 1 0,6 Vibrionaceae (u.g.) 0,4 0,7 0,1 1 Bacteroidia (u.g.) 0,3 0,5 1 0,7 Gammaproteobacteria (u.g.) 0,3 1 1 0,2
Table S5.2.2 Core ASVs present in the skin microbiota of the seabass Dicentrarchus labrax across states. Unknown genera are labeled as u.g.
Taxonomy Healthy Diseased Treatment Recovery Actibacter - - - ASV48 Alkalimarinus - ASV16 ASV16 ASV16 Alteromonas - ASV7 ASV7 ASV7 Arcticiflavibacter - - ASV19 ASV19; ASV38 Aureispira - - - ASV20 DBS1 - - ASV54 - Fabibacter - - ASV22 ASV22 Formosa - - ASV29 ASV29 Fusobacterium - - - ASV133 Hypnocyclicus - - - ASV99 Labilibacter - - - ASV78 Lacinutrix - - ASV40 ASV40 Lentisphaera - - - ASV36 Leucothrix mucor - - - ASV62 Lewinella - - - ASV50 Maribacter - - ASV32 ASV28; ASV32 Marinicella - - - ASV55 Marinifilum - - ASV59 ASV59 Marinomonas ASV15 ASV15 ASV15 ASV15
(Continues next page)
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
206
(Table S5.2.2 Continued) Neiella marina - - - ASV142 NS1 marine group - ASV9 ASV9 ASV69 NS3a marine group - - - ASV31 Oceanospirillum - - - ASV68 Oleiphilus - - - ASV85 Oleispira antarctica ASV3 ASV3 ASV3 ASV3 Photobacterium damselae - ASV18 - - Polaribacter - ASV4 ASV4 ASV4 Porticoccus - - - ASV88 Propionigenium - - - ASV102 Pseudoalteromonas ASV1 ASV1 ASV1 ASV1 Pseudoalteromonas phenolica - ASV12 ASV12 ASV12
Pseudomonas ASV14; ASV2 ASV14; ASV2 ASV14;
ASV2 ASV2
Psychrilyobacter atlanticus - - - ASV111 Psychrobium - ASV21 - ASV21 Psychromonas - - - ASV176 Psychrosphaera saromensis - ASV13 ASV13 ASV13 Reinekea - - - ASV94 Rubritalea - - ASV35 ASV35 Ruegeria - - ASV26 ASV26 Salinirepens - - ASV98 - Sanguibacter - ASV24 - - Spongiibacterium - - - ASV57 Stenotrophomonas ASV5 ASV5 ASV5 Tamlana - ASV37 - ASV37 Tenacibaculum - - ASV17 ASV17 Thalassotalea - ASV11 ASV11 ASV11 Thalassotalea agariperforans - - ASV43 ASV43
Vibrio - ASV23; ASV27 - ASV129; ASV23; ASV27; ASV33; ASV63;
ASV82 Vibrio fortis - ASV10 ASV10 ASV10 Vicingus - - - ASV83 Winogradskyella echinorum - - ASV34 ASV34 Alteromonadaceae (u.g.) - - ASV25 ASV25 Cellvibrionaceae (u.g.) - - - ASV47 Cryomorphaceae (u.g.) - - ASV8 ASV8 Flavobacteriaceae (u.g.) - - ASV6 ASV6 Marinifilaceae (u.g.) - - - ASV208 Rhodobacteraceae (u.g.) - - - ASV71 Saprospiraceae (u.g.) - - - ASV97 Spongiibacteraceae (u.g.) - - - ASV156; ASV89 Vibrionaceae (u.g.) - - - ASV30 Kordiimonadales (u.g.) - - - ASV53 Verrucomicrobiales (u.g.) - - - ASV116 Lentisphaeria (u.g.) - - - ASV140; ASV233
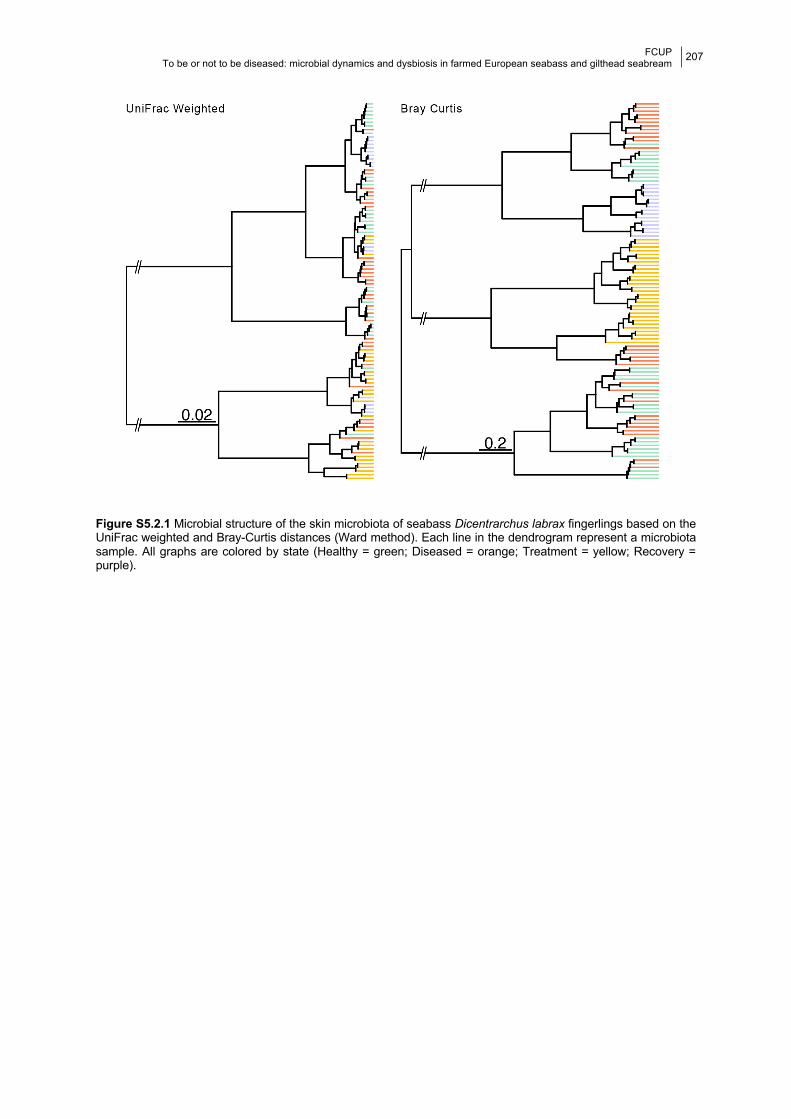
FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
207
Figure S5.2.1 Microbial structure of the skin microbiota of seabass Dicentrarchus labrax fingerlings based on the UniFrac weighted and Bray-Curtis distances (Ward method). Each line in the dendrogram represent a microbiota sample. All graphs are colored by state (Healthy = green; Diseased = orange; Treatment = yellow; Recovery = purple).

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
208
CHAPTER 6
General Discussion This doctoral thesis was developed in close collaboration with an European seabass
and gilthead seabream farm, located in the south of Portugal. The natural settings of a fish
farm presented several advantages to the study of fish microbiomes in relation to studies
developed in the wild, namely information provided on fish health, age and diet, and to some
extent the environment. The sampling scheme allowed me to take advantage of regular
access to fish, which encompassed weekly sampling between 2016 and 2018 of the skin and
gill microbiota of the seabass and seabream. This allowed me to generate important insights
about the biotic and abiotic factors shaping the microbiota of farmed seabass and seabream.
Specifically, I was able to test the impact of: a) host taxonomy (Chapter 2); b) mucosal tissue
(skin and gill, Chapters 2, 3, 4 and 5.1); c) host age (Chapter 3); d) water temperature
(Chapter 4); and e) water microbial communities (Chapters 3 and 4) on shaping the skin and
gill microbiota of the European seabass Dicentrarchus labrax and gilthead seabream Sparus
aurata. The occurrence of natural bacterial disease outbreaks and consequent antibiotic
treatment in seabass, also allowed me to shed light on the role of infection and antibiotics on
seabass dysbiosis and disease (Chapter 5).
6.1 Variability associated with the skin and gill microbiota in farmed
seabass and seabream The microbial communities of the two studied fish mucosae, skin and gill, presented
differences in diversity (seabass and seabream, Chapters 2 and 3) and response to external
stressors (seabass, Chapter 5.1). Generally, the skin microbiota had higher diversity than the
gill microbiota. Such microbial differences between and within body habitats are well
documented in teleost species (e.g., Legrand et al., 2020a; Lowrey et al., 2015; Pratte et al.,
2018; Zhang et al., 2019). It is known that commensal microbial communities have coevolved
with each body habitat, being highly adapted to specific environment (Lee and Mazmanian,
2010). This adaptation is the result of each niche selective pressures related with physiology
and immunity (Kelly and Salinas, 2017). In fish, these could be the result of different mucosa
associated lymphoid tissues (MALTs, Kelly and Salinas, 2017), physicochemical properties
(Egerton et al., 2018), osmo- and iono-regulatory function (Fu et al., 2010; LeBlanc et al.,
2015), antimicrobial peptides (Noga et al., 2010), mucous histology (Dang et al., 2020), to
mention a few. To the best of my knowledge, there are no comparative studies between fish

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
209
tissue and microbiome function under homeostasis (e.g., through dual-transcriptomics). I
hypothesize that different physiological requirements within each tissue will translate into
singular microbial communities optimized to provide specific functional needs at each body
site. Nevertheless, the longitudinal analysis of microbial diversity of the skin and gill of the
seabass across one year (in Chapter 4) followed the same trends, suggesting modulation by
the same factors. In that study I observed that under healthy conditions, diversity of the gill
microbiota of the seabass was more stable, i.e., less variable across months, while the skin
microbiota was more variable throughout the year (Chapter 4).
Dysbiosis, observed in Chapters 4 and 5.1, was also more pronounced in the skin
microbiota of the seabass, which presented higher susceptibility to dysbiosis as well as higher
incidences of potentially pathogenic genera. On the other hand, the gill microbiota of the
seabass was less resilient than the skin microbiota in presence of disease and antibiotic
treatment, with significant differences in diversity between healthy and recovery states
(Chapter 5.1). These results suggest that, although the variation observed in the skin
microbiota was higher than in the gill and may underlie a higher susceptibility to dysbiosis, it
may also be responsible for a faster recovery to healthy-like diversity values after microbial
disruption. It has been hypothesized that physiological plasticity, i.e., the ability to change in
response to changes in the environment, confer the microbial community resilience to
disturbances (e.g., Seebacher et al., 2014; Shade et al., 2012). Hence, the skin, perhaps due
to its larger area, offers more opportunities for colonisation by different organisms when
compared to the gill. This could explain its higher bacterial richness and, to some extent, its
increased variability over time due to increasing microbial interactions (e.g., competition). The
differences in microbial response to stressors observed here between tissues can also be a
manifest of the different immunological characteristics of each body niche. For example, a
study showed that cortisol related signatures of stress in Atlantic salmon were related to
changes in the microbial diversity and structure of the feces but not the skin (Uren Webster et
al., 2020). It is known that fish microbial communities regulate host adaptive immune system
and its response, influencing the resilience to stress and disease (Kelly and Salinas, 2017).
Given that microbial diversity is highly adapted to each tissue, it is only logical that its role in
the immune response will also change between body habitats.
Although significantly different, skin microbial structure of both seabass and seabream
were more similar to the water microbial structure, when compared to the gill, especially in
younger age groups (Chapter 3). A recent study showed that skin and gut microbiota interact
more with the surrounding environment, such as water and biofilms formed in fish tanks, than
the gill microbiota (Minich et al., 2020b). Nonetheless, predicted monthly microbial dispersion
was higher between the skin and gill microbiota of adult seabass than between both tissues
and water (Chapter 4). This further suggests that continuous microbial exchange of

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
210
communities between skin and gill may be creating colonizing opportunities in response to
changes in the environment (Chapter 4).
6.2 Host related factors shaping the skin and gill microbiota in farmed
seabass and seabream Baseline microbial characterization of the skin and gill of farmed seabass and
seabream revealed structural differences between the two species (Chapter 2), corroborating
the effect of host taxonomy on microbial diversity reported for fish (e.g., Larsen et al., 2013;
Pratte et al., 2018; Reverter et al., 2017; Sylvain et al., 2020). However, differences in
microbial composition between seabass and seabream were not seen in the gill. This indicates
that when seabass and seabream share the same environmental conditions, host taxonomy
mostly influences microbiota at the structural level. Similar results have been reported for the
gut microbiota of the phylogenetically related species blue and channel catfish (Bledsoe et al.,
2018). This is likely related to species' traits that exert specific selective pressures. For
example, different species can have distinctive physiological and immune requirements at
homeostasis and respond differently to altered conditions (e.g., Ellison et al., 2020). Despite
the high inter-individual microbial variation commonly observed in fish (e.g., Burns et al., 2017;
Chiarello et al., 2015), and also verified in our studies, seabream microbiota were overall less
diverse than those of the seabass (Chapters 2 and 3). Results also showed that differences
between skin and gill were more pronounced in the seabass, with significant differences in
microbial composition and structure, when compared to the seabream, where only structure
was significantly different (Chapter 2).
The longitudinal study in Chapter 3 showed different effects of aging in seabass and
seabream, although for technical reasons the results for the two species were not directly
comparable. Individuals of farmed seabass and seabream belonging to distinct age groups
(corresponding to different levels of sexual maturation and growth), presented differences in
microbial structure and predicted function. However, only seabass showed significant
differences in microbial composition across age groups, indicating a higher stability of
seabream microbiota during development. The few longitudinal studies evaluating the
ontogenetic effect on teleost microbiota showed that fish experience diversity shifts at different
times of the development depending on the species (e.g., Bledsoe et al., 2016; Lokesh et al.,
2019; Nikouli et al., 2019; Stephens et al., 2016; Wilkes Walburn et al., 2019; Yan et al., 2016).
On the other hand, major life events such as diet transitions (Wilkes Walburn et al., 2019),
environmental transitions (Lokesh et al., 2019), sexual differentiation (Stephens et al., 2016),
or sexual maturation (Chapter 3), seem to be an important factor triggering microbial diversity
shifts through time across teleost species. It is known that sexual maturation in fish is

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
211
accompanied by physiological changes that will allocate energy from survival to gamete
production and reproductive behavior (Saborido-Rey and Kjesbu, 2005). Growth and sexual
maturation can be characterized by changes in metabolism (e.g., Persson et al., 2005),
morphology (e.g., Walleser et al., 2014), hormones (e.g., Hatef and Unniappan, 2019),
behavior (e.g., Davis and Kassel, 1975), among others. Such extreme changes lead to
alterations in the physicochemical characteristics of the fish tissues (e.g., Saborido-Rey and
Kjesbu, 2005), that can trigger shifts in the microbial communities in order to adapt to the
altered environment as well as functionally aid any supplementary needs. In our study, the
trend of an increased diversity with age observed in the seabass was translated into an
enrichment of the predicted functional diversity. In the seabream, although bacterial diversity
remained seemingly stable throughout development, an increase in functional diversity was
also predicted.
6.3 Water related factors shaping the skin and gill microbiota in farmed
seabass and seabream Besides the effects of host age, significant microbial shifts in fish were also previously
suggested to reflect changes in a multitude of environmental variables, the vast majority
related to water characteristics, such as temperature, pH, ionic composition, dissolved
oxygen, conductivity or chlorophyll a (Hovda et al., 2012; Krotman et al., 2020; Minich et al.,
2020a; Vasemägi et al., 2017; Zarkasi et al., 2014). In fact, in the present thesis a pronounced
effect of water temperature on seabass skin and gill microbial diversity and predicted functions
were seen throughout the year (Chapter 4). Such impact was evident both over short (couple
of days) and long (several weeks) periods of time and was patent in both microbial composition
and structure. Microbial shifts due to temperature occurring at small temporal scales have
been reported for abrupt transitions (e.g., Kokou et al., 2018; Tarnecki et al., 2019), while long
term microbial changes were often associated with season (e.g., Minich et al., 2020a). Albeit
the observed strong effect of temperature, other environmental characteristics might have also
been responsible for the high microbial variability observed in the microbiota of farmed
seabass over the studied year. These results highlight the importance of longitudinal studies
and repeated methodical measurements of environmental variables.
Overall, water microbiota had higher compositional diversity than host associated
communities, independently of their age group (Chapter 3), in accordance to what is usually
described in the literature (Thompson et al., 2017). However, this was not always the case
when analyzing the monthly diversity of the microbiota over a year (Chapter 4). Excluding
months where dysbiosis occurred, there were some months where water microbial diversity
was lower than those of the skin or gill, demonstrating the importance of assessing longitudinal

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
212
variation. This work also demonstrated a significant structural distinction of the water
microbiota from the skin and gill microbiota of the seabass and seabream (Chapter 3), in
agreement with the majority of the studies (e.g., Chiarello et al., 2019; Pratte et al., 2018; Wu
et al., 2020; Zhang et al., 2019). Nevertheless, at least to some extent, microbial communities
of the surrounding water seem to contribute to the microbial diversity of the skin and gill, since
several ASVs were shared between water and all studied microbiotas in both species
(Chapter 3). Similar findings are not uncommon and suggests that body microbiota
composition is a subset selection of the environment microbes (e.g., Nikouli et al., 2019; Pratte
et al., 2018; Zhang et al., 2018). In fact, water microbial communities were found to be one of
the source contributors to the monthly skin and gill microbial diversity of adult seabass;
however, that contribution was the lowest when compared to the other sources (Chapter 4).
On the other hand, it is interesting to notice that the potentially pathogenic genera, whose
increase in abundance promoted dysbiosis in the skin and gill microbiota of the seabass, were
not found in high abundances in the water microbial community (Chapter 4). This suggests
that other potential sources may contribute to the dispersal of potentially pathogenic genera.
One such source might be the biofilms established in rearing tanks, which are known
reservoirs of pathogenic bacteria (e.g., Cai and Arias, 2017; King et al., 2004). Moreover,
biofilms were seen to have greater impact than the surrounding water in mucoase microbial
diversity (Minich et al., 2020b). Additionally, potentially pathogenic genera can enter
aquaculture settings through feed or broodstock, among others (Saksida et al., 2014). On the
other hand, it is known that healthy mucosal surfaces are naturally colonized by potentially
pathogenic bacteria that, under specific environmental conditions, can become pathogenic to
the host (e.g., Givens et al., 2015; Hess et al., 2015; Legrand et al., 2018).
6.4 Dysbiosis in the skin and gill microbiota of farmed seabass and
seabream There were several dysbiotic events observed in the skin and gills of farmed European
seabass populations during the analyzed period. In all but one occasion, dysbiosis was
associated with an increase in the abundance of potentially pathogenic (PP) genera; however,
disease was not always confirmed (Chapter 4 and 5).
In the longitudinal study performed in Chapter 4, despite seabass individuals showing
no signs of disease, three dysbiotic events were evident. Dysbiosis was characterized by a
significant decrease in diversity, which on two occasions was accompanied by an increase in
the abundance of potentially pathogenic genera (Aliivibrio, Pseudomonas, Photobacterium
and Vibrio). Even though pathogenic genera can naturally be part of the healthy microbiota
(e.g., Califano et al., 2017; Givens et al., 2015), such as Vibrio in seabass and seabream

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
213
(Chapter 2), an increase in their abundance can lead to dysbiosis. In Chapter 4, we showed
that changes in the abundance of potentially pathogenic bacteria were also correlated to
changes in the water temperature, indicating that it is a factor inducing dysbiosis. Additionally,
these dysbiotic events occurred in warmer months and during cold/warm transitions, further
demonstrating the importance of changes in water temperature for the onset of such events.
It is especially important that the relationship between temperature and commensal microbial
dynamics is further investigated given the emergence of climate change and ocean warming
(e.g., Baker-Austin et al., 2017). For example, it has been demonstrated that environmental
factors influence the proportion of bacteria with probiotic or pathogenic properties in fish gut
(Zarkasi et al., 2014; Zhao et al., 2020).
Monitoring the skin and gill microbiota of healthy seabass allowed me to understand
how bacterial infection was linked to microbial imbalance in adults (Chapter 5.1) and
fingerlings (Chapter 5.2). Microbial response to disease was asymmetrical between tissues
in the adult population (Chapter 5.1), although representing what constitutes a standard sign
of dysbiosis (Petersen and Round, 2014). Additionally, the increase of abundance of the genus
Pseudoalteromonas in the skin microbial community suggests that this genus could represent
a biomarker for seabass health. The probiotic activity of several Pseudoalteromonas species
has been described against Vibrio spp. and Photobacterium damselae (e.g., Offret et al., 2016;
Richards et al., 2017). Additionally, differences in response between different age groups of
seabass (Chapter 5; Cámara-Ruiz et al., 2021) reinforce a possible ontogenetic effect, likely
related to the development of the immune system. However, it is also likely that different
diseases (photobacteriousis vs vibriosis + photobacteriousis) could cause the observed
differences in response between age groups.
It is important to notice that within the past 5 years no signs of infection were detected
in the seabream populations of the tanks monitored during this thesis. This could attest for a
stronger immune system, including features within the microbiome (e.g., higher stability
throughout time, Chapter 3); however, studies outside the scope of this thesis would be
necessary to fully understand this question.
In both studies included in Chapter 5 treatment with antibiotics, a standard procedure
in fish farms (e.g., Cabello, 2006), was initiated after disease diagnosis. The two antibiotics
used were oxytetracycline in adults (Chapter 5.1) and flumequine in fingerlings (Chapter 5.2).
Results revealed that both antibiotics significantly impacted microbial diversity, with both
oxytetracycline and flumequine leading to an increase in diversity in the skin microbiota
(Chapter 5). On the other hand, oxytetracycline prompted a decrease in core diversity in the
gill microbiota of adult seabass, although differences in composition were not significant
(Chapter 5.1). Differences in response to external stressors are most likely related to
differences in the overall diversity and stability observed between tissues. Overall, gill

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
214
microbial community was more stable than that of the skin. Flumequine treatment also
impacted the predicted metabolic function of the skin microbiota of seabass fingerlings. These
results add to an increasing body of literature showing the harmful effects of antibiotics on fish
microbial homeostasis and consequently, fish health (e.g., Almeida et al., 2019; Carlson et al.,
2017; Kim et al., 2019; Legrand et al., 2020a; Pindling et al., 2018). These studies further
reveal how unpredictable these effects can be and reaffirm the urgent need for alternatives to
antibiotic administration in aquaculture.
Short term microbial recovery of both adults (3 weeks, Chapter 5.1) and fingerlings (1
week, Chapter 5.2) after antibiotic administration was also assessed. Significant differences
in composition and structure between healthy and recovery states were in line with previous
studies revealing the lasting impacts of antibiotic exposure on skin microbial diversity of fish
(e.g., Carlson et al., 2017, 2015). Interestingly, core diversity decreased in adult fish and
increased in fingerlings during recovery, when compared to the core diversity presented by
healthy individuals of both age groups, exhibiting another asymmetrical response that could
be related to development. Additionally, the abundance of the etiological agents of disease in
both studies (Photobacterium, Chapter 5.1 and Vibrio, Chapter 5.2) increased again during
the recovery state, indicating microbial homeostasis had not been reached yet.
6.5 Microbiome monitoring implications for aquaculture The microbiome of fish can be responsible by a great number of beneficial functions,
such as digestion nutrition, growth, reproduction and immunity (Diwan et al., 2021), granting
the host with the phenotypic plasticity needed to adapt to a particular habitat and living
conditions (Rajeev et al., 2021). In aquaculture settings, infectious diseases are one of the
main causes of economic loss with an overexploitation of antimicrobials having developed into
the emergence of antimicrobial resistance (Schar et al., 2020; Rajeev et al., 2021). In this
regard, microbiome research can be explored to optimize fish farm practices, since
commensal microbes play a role in immune protection (Luna et al., 2022; Rajeev et al., 2021).
There are already several preventive and treatment approaches in aquaculture directed
towards microbial modulation, such as the use of probiotics (e.g., Sun et al., 2014), prebiotics
(e.g., Torrecillas et al., 2013), synbiotics (e.g., Rodriguez-Estrada et al., 2013), fecal
microbiota transplant (FMT, e.g., Smith et al., 2017), biofloc technology (e.g., Emerenciano et
al., 2013), or RAS technology (e.g., Espinal and Matulić, 2019) (see Chapter 1 for details).
Thus, it is well established that microbial manipulation is essential to guarantee a profitable
and sustainable aquaculture industry by helping prevent as well as mitigate infectious
diseases (Diwan et al., 2021; Luna et al., 2022; Yılmaz et al., 2021). However, in order to
maximize the efficiency of microbial therapy, it is critical to know what constitutes a healthy

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
215
commensal community for a species or population and how it can be affected by disturbances
such as disease (Rajeev et al., 2021).
In this thesis, longitudinal monitoring of the skin and gill microbiota of fish revealed
consistent abundant taxa across time, uncovering the predominant bacteria of adult seabass
(Chapters 2, 3 and 4) and seabream (Chapter 2 and 3). Proteobacteria and Bacteroidota
were the dominant phyla, in both fish species and across tissues, time of sampling and health
status, in accordance to what has been described for teleost species (Legrand et al., 2020b;
Llewellyn et al., 2014), including the skin of seabass and seabream (Cámara-Ruiz et al., 2021;
Chiarello et al., 2015; Pimentel et al., 2017; Tapia-Paniagua et al., 2018). Microbial
composition at the genus level was more variable and differed between the sampling periods
analyzed in this thesis. Nevertheless, the genus NS3a marine group was highly abundant in
healthy tissues and species analysed in this thesis. Additionally, Rubritalea and
Polynucleobacter were highly abundant in seabass gill, and Vibrio in seabream skin across
time (Chapters 2, 3 and 4). Despite the variability of abundant genera in time, several ASVs
were 100% prevalent and established the core microbiota of the populations analyzed during
winter (seabass and seabream, Chapter 2) and across one year (seabass, Chapter 4). This
resilient and healthy functional core is composed by genetic strains capable of resisting host
and habitat disturbances (Rajeev et al., 2021).
Monitoring farmed seabass populations also allowed me to explore microbial dynamics
during dysbiotic events (Chapters 4 and 5). For example, I was able to see how the
abundance of potentially pathogenic genera can be correlated with the abundance of potential
probiotics (Chapter 4), or how certain genera increase their abundance during infection
(Chapter 5), representing potential biomarkers. In microbiome research, a biomarker is a
microbial species or a group of species that represent an altered state of homeostasis due to
changes in the biotic and/or abiotic conditions (Llewellyn et al., 2014). For example, the
commensal microbiota will shift depending on host behavioral and physiological response to
stress (Llewellyn et al., 2014). This imbalance of the microbiome, or dysbiosis, is often a
precursor of disease, being crucial to detect the microbial shift through the known biomarkers
before the onset of disease (Llewellyn et al., 2014). In the future, a deeper thorough study of
the complex microbial dynamics occurring during the onset of dysbiosis will be very insightful
to detect all possible microbial biomarkers for stress and disease resistance. Biomarkers can
be further studied to find healthier alternatives to antibiotics for farmed European seabass and
gilthead seabream, such as probiotics.
The results of this thesis supplemented the body of literature revealing not only the
baseline healthy microbial communities of farmed seabass and seabream but also potential
probiotic genera. These results can be taken further to develop alternatives to vaccination
and/or antibiotics to combat infection, helping decrease antimicrobial resistance and

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
216
production losses. Indeed, such practice is already been recommended to be implemented for
the thriving of the aquaculture industry without putting at risk entire populations of aquatic
animals (Rajeev et al., 2021). Additionally, the results of this thesis regarding fish development
(Chapters 3 and 5) highlight the importance of considering age in fish microbiome studies.
Information regarding the different healthy microbial communities and different microbial
response to infection can refine the research on potential probiotics to be used in aquaculture
settings at each developmental stage. Such extremely valuable information for the successful
application of microbiome research in aquaculture was only unveiled due to the frequent
monitoring of the microbial communities in these farmed populations. Combining
environmental and microbial data using regular monitoring within aquaculture settings coupled
with machine learning algorithms can be very powerful for biomonitoring programs and early
detection of microbial imbalance and prevention of disease (Cordier et al., 2019; Luna et al.,
2022).
6.6 Future perspectives The methodology applied in this thesis to achieve the goals was carefully deliberated
in order to make the most of the available resources. All data were collected non-invasively
and all work presented here is the result of natural occurrences, i.e., no condition was ever
manipulated. Although this strategy might have limited the results in terms of the amount of
information gathered (for example only temperature was regularly measured), it provided
“real” data. Overall, my goals were accomplished and there were very few limitations to sample
collection and sequencing success. On the other hand, in the process of answering the initial
questions, many others germinated.
One of the main caveats in this research was the impossibility to record additional
environmental data besides temperature, that could help explain much of the variability
observed. Fish were reared in an exterior coastal circulation system situated in an estuary,
where water is renewed with every tide and where the environment presents extreme turbidity
and is highly variable throughout the day. For these reasons, specific probes would have been
needed to take accurate, repeated, short-term (e.g., hourly) measurements in order to have
reliable data on other environmental variables. Future work will definitely address these
issues, which will complement our results and help describe to what extent environmental
variables shape microbial diversity. In this regard, it will also allow us to identify other variables
triggering dysbiosis, eventually helping to generate prediction models for dysbiosis in
aquaculture settings. It will also be critical to assess other sources of microbial dispersal,
including potential pathogens, like biofilms, nets, soil or feed. Lastly, in order to evaluate the

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
217
true resilience of the microbial communities, assessment of the recovery period will need to
be further extended beyond three weeks after infection or antibiotic treatment.
By selecting to collect non-invasive samples, our work also lacked analyses of other
tissues, mainly the gut, as well as other host related factors that could explain microbial
variation, such as sex or weight. Designing experimental studies within the laboratory or in
mesocosm settings under controlled environments will be important to measure other
explanatory factors and compare immune response and dysbiosis. For example, in order to
better understand the effect of ontogeny in response to disease, controlled experiments should
be done using the same aetiological agent in fish populations comprising different age groups.
All data produced in this thesis was based on a metataxonomics approach using the
16S rRNA marker gene, allowing identification of the microbial communities present in the skin
and gill of farmed seabass and seabream under different conditions. However, to gain
additional knowledge regarding the role of factors shaping microbial diversity and function,
such as the ones studied here, and to explore host-microbiome interactions, other
approaches, such as shotgun, metatranscriptomic or dual-transcriptomic sequencing should
be employed.
The microbiota comprises all the microbes (bacteria, archaea, fungi, viruses and
protozoa) living in an environment; however, fish microbiome research has mainly focused
on the bacterial microbiota. In order to fully understand microbe-host-environment
interactions, further studies should also include other microbial organisms and widen the
questions.
6.7 Conclusions The study of the microbiome was initially focused on humans and model organisms,
but recently expanded to many other taxa. This doctoral thesis substantially increased our
knowledge on factors shaping the microbiota of farmed fish as well the factors triggering
dysbiosis. Aquaculture is the most promising food-production industry worldwide that is bound
to accompany human population growth (FAO, 2018). However, there are several constraints
to its further development, the biggest being infectious diseases, urgently requiring improved
practices and health management (Béné et al., 2015; Lafferty et al., 2015). In this regard,
studying the microbiome of fish and the surrounding environment can unveil many answers,
given the role of the microbiome in health and immunity (Kelly and Salinas, 2017; Trivedi,
2012).
Our research showed a high variability of the skin and gill microbiota across time
(Chapters 2-4) governed by host (Chapters 2-3) and environmental (Chapter 4) factors. We
also showed that regular monitoring of the microbial communities of farmed species can yield

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
218
extremely valuable insights that can ultimately help detect, prevent or mitigate disease
outbreaks in aquaculture settings (Chapter 5). Microbiomes are intimately linked to the
function and health of ecosystems (Sehnal et al., 2021). Due to their sensitivity to
environmental disturbances, ubiquitous nature as well as their high-throughput low cost
sequencing, versatile and informative analysis, microbiomes can be perfect indicators of
health and ecological status of aquatic systems (Sehnal et al., 2021). In this regard, ecosystem
bioassessment and ecological quality status can be done through microbiomes, especially
when other traditional biomonitoring tools are not available or effective (Sehnal et al., 2021).
The questions in this field of research are virtually endless and its potential applications
are fundamental for the future of all living beings. Results from this thesis substantially added
to the existing knowledge and can pave the way to fundamental improvements in the health
of European seabass and gilthead seabream in aquaculture settings.
6.8 References Almeida, A.R., Alves, M., Domingues, I., Henriques, I., 2019. The impact of antibiotic exposure
in water and zebrafish gut microbiomes: A 16S rRNA gene-based metagenomic analysis.
Ecotoxicol. Environ. Saf. 186, 109771. https://doi.org/10.1016/j.ecoenv.2019.109771
Baker-Austin, C., Trinanes, J., Gonzalez-Escalona, N., Martinez-Urtaza, J., 2017. Non-
Cholera Vibrios: The Microbial Barometer of Climate Change. Trends Microbiol.
https://doi.org/10.1016/j.tim.2016.09.008
Béné, C., Barange, M., Subasinghe, R., Pinstrup-Andersen, P., Merino, G., Hemre, G.I.,
Williams, M., 2015. Feeding 9 billion by 2050 – Putting fish back on the menu. Food
Secur. 7, 261–274. https://doi.org/10.1007/s12571-015-0427-z
Bier, R.L., Bernhardt, E.S., Boot, C.M., Graham, E.B., Hall, E.K., Lennon, J.T., Nemergut,
D.R., Osborne, B.B., Ruiz-González, C., Schimel, J.P., Waldrop, M.P., Wallenstein, M.D.,
2015. Linking microbial community structure and microbial processes: An empirical and
conceptual overview. FEMS Microbiol. Ecol. https://doi.org/10.1093/femsec/fiv113
Bledsoe, J.W., Peterson, B.C., Swanson, K.S., Small, B.C., 2016. Ontogenetic
Characterization of the Intestinal Microbiota of Channel Catfish through 16S rRNA Gene
Sequencing Reveals Insights on Temporal Shifts and the Influence of Environmental
Microbes. PLoS One 11, e0166379. https://doi.org/10.1371/journal.pone.0166379
Bledsoe, J.W., Waldbieser, G.C., Swanson, K.S., Peterson, B.C., Small, B.C., 2018.
Comparison of Channel Catfish and Blue Catfish Gut Microbiota Assemblages Shows
Minimal Effects of Host Genetics on Microbial Structure and Inferred Function. Front.
Microbiol. 9, 1073. https://doi.org/10.3389/fmicb.2018.01073
Burns, A.R., Miller, E., Agarwal, M., Rolig, A.S., Milligan-Myhre, K., Seredick, S., Guillemin,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
219
K., Bohannan, B.J.M., 2017. Interhost dispersal alters microbiome assembly and can
overwhelm host innate immunity in an experimental zebrafish model. Proc. Natl. Acad.
Sci. U. S. A. 114, 11181–11186. https://doi.org/10.1073/pnas.1702511114
Cabello, F.C., 2006. Heavy use of prophylactic antibiotics in aquaculture: a growing problem
for human and animal health and for the environment. Environ. Microbiol. 8, 1137–1144.
https://doi.org/10.1111/j.1462-2920.2006.01054.x
Cai, W., Arias, C.R., 2017. Biofilm formation on aquaculture substrates by selected bacterial
fish pathogens. J. Aquat. Anim. Health 29, 95–104.
https://doi.org/10.1080/08997659.2017.1290711
Califano, G., Castanho, S., Soares, F., Ribeiro, L., Cox, C.J., Mata, L., Costa, R., 2017.
Molecular Taxonomic Profiling of Bacterial Communities in a Gilthead Seabream (Sparus
aurata) Hatchery. Front. Microbiol. 8, 204. https://doi.org/10.3389/fmicb.2017.00204
Cámara-Ruiz, M., Cerezo, I.M., Guardiola, F.A., García-Beltrán, J.M., Balebona, M.C.,
Moriñigo, M.Á., Esteban, M.Á., 2021. Alteration of the Immune Response and the
Microbiota of the Skin during a Natural Infection by Vibrio harveyi in European Seabass
(Dicentrarchus labrax). Microorganisms 9, 964.
https://doi.org/10.3390/microorganisms9050964
Carlson, J.M., Hyde, E.R., Petrosino, J.F., Manage, A.B.W., Primm, T.P., 2015. The host
effects of Gambusia affinis with an antibiotic-disrupted microbiome. Comp. Biochem.
Physiol. Part - C Toxicol. Pharmacol. 178, 163–168.
https://doi.org/10.1016/j.cbpc.2015.10.004
Carlson, J.M., Leonard, A.B., Hyde, E.R., Petrosino, J.F., Primm, T.P., 2017. Microbiome
disruption and recovery in the fish Gambusia affinis following exposure to broad-
spectrum antibiotic. Infect. Drug Resist. 10, 143–154.
https://doi.org/10.2147/IDR.S129055
Chiarello, M., Paz-Vinas, I., Veyssière, C., Santoul, F., Loot, G., Ferriol, J., Boulêtreau, S.,
2019. Environmental conditions and neutral processes shape the skin microbiome of
European catfish (Silurus glanis) populations of Southwestern France. Environ. Microbiol.
Rep. 11, 605–614. https://doi.org/10.1111/1758-2229.12774
Chiarello, M., Villéger, S., Bouvier, C., Bettarel, Y., Bouvier, T., 2015. High diversity of skin-
associated bacterial communities of marine fishes is promoted by their high variability
among body parts, individuals and species. FEMS Microbiol. Ecol. 91.
https://doi.org/10.1093/femsec/fiv061
Cordier, T., Lanzén, A., Apothéloz-Perret-Gentil, L., Stoeck, T., Pawlowski, J., 2019.
Embracing Environmental Genomics and Machine Learning for Routine Biomonitoring.
Trends Microbiol. https://doi.org/10.1016/j.tim.2018.10.012
Dang, M., Pittman, K., Sonne, C., Hansson, S., Bach, L., Søndergaard, J., Stride, M., Nowak,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
220
B., 2020. Histological mucous cell quantification and mucosal mapping reveal different
aspects of mucous cell responses in gills and skin of shorthorn sculpins (Myoxocephalus
scorpius). Fish Shellfish Immunol. 100, 334–344.
https://doi.org/10.1016/J.FSI.2020.03.020
Davis, R.E., Kassel, J., 1975. The ontogeny of agonistic behavior and the onset of sexual
maturation in the paradise fish, Macropodus opercularis (linnaeus). Behav. Biol. 14, 31–
39. https://doi.org/10.1016/S0091-6773(75)90290-4
Diwan, A.D., Harke, S.N., Gopalkrishna, Panche, A.N., 2021. Aquaculture industry
prospective from gut microbiome of fish and shellfish: An overview. J. Anim. Physiol.
Anim. Nutr. (Berl). https://doi.org/10.1111/JPN.13619
Ellison, A.R., Uren Webster, T.M., Rodriguez-Barreto, D., de Leaniz, C.G., Consuegra, S.,
Orozco-terWengel, P., Cable, J., 2020. Comparative transcriptomics reveal conserved
impacts of rearing density on immune response of two important aquaculture species.
Fish Shellfish Immunol. 104, 192–201. https://doi.org/10.1016/J.FSI.2020.05.043
Emerenciano, M., Gaxiola, G., Cuzon, G., 2013. Biofloc technology (BFT): a review for
aquaculture application and animal food industry., in: Matovic, M.D. (Ed.), Biomass Now:
Cultivation and Utilization. BoD–Books on Demand, pp. 301–328.
Espinal, C.A., Matulić, D., 2019. Recirculating Aquaculture Technologies, in: Goddek, S.,
Joyce, A., Kotzen, B., Burnell, G.M. (Eds.), Aquaponics Food Production Systems:
Combined Aquaculture and Hydroponic Production Technologies for the Future. Springer
Open, pp. 35–55
FAO, 2018. The state of world fisheries and aquaculture 2018 - Meeting the sustainable
development goals. Rome.
Givens, C., Ransom, B., Bano, N., Hollibaugh, J., 2015. Comparison of the gut microbiomes
of 12 bony fish and 3 shark species. Mar. Ecol. Prog. Ser. 518, 209–223.
https://doi.org/10.3354/meps11034
Hatef, A., Unniappan, S., 2019. Metabolic hormones and the regulation of spermatogenesis
in fishes. Theriogenology 134, 121–128.
https://doi.org/10.1016/J.THERIOGENOLOGY.2019.05.021
Hess, S., Wenger, A.S., Ainsworth, T.D., Rummer, J.L., 2015. Exposure of clownfish larvae to
suspended sediment levels found on the Great Barrier Reef: Impacts on gill structure and
microbiome. Sci. Rep. 5, 1–8. https://doi.org/10.1038/srep10561
Hovda, M.B., Fontanillas, R., Mcgurk, C., Obach, A., Rosnes, J.T., 2012. Seasonal variations
in the intestinal microbiota of farmed Atlantic salmon (Salmo salar L.) 43, 154–159.
https://doi.org/10.1111/j.1365-2109.2011.02805.x
Kelly, C., Salinas, I., 2017. Under pressure: Interactions between commensal microbiota and
the teleost immune system. Front. Immunol. https://doi.org/10.3389/fimmu.2017.00559

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
221
Kim, A., Kim, N., Roh, H.J., Chun, W.K., Ho, D.T., Lee, Y., Kim, D.H., 2019. Administration of
antibiotics can cause dysbiosis in fish gut. Aquaculture 512, 734330.
https://doi.org/10.1016/j.aquaculture.2019.734330
King, R.K., Flick, G.J., Pierson, M.D., Smith, S.A., Boardman, G.D., Coale, C.W., 2004.
Identification of bacterial pathogens in biofilms of recirculating aquaculture systems. J.
Aquat. Food Prod. Technol. 13, 125–133. https://doi.org/10.1300/J030v13n01_11
Kokou, F., Sasson, G., Nitzan, T., Doron-Faigenboim, A., Harpaz, S., Cnaani, A., Mizrahi, I.,
2018. Host genetic selection for cold tolerance shapes microbiome composition and
modulates its response to temperature. Elife 7. https://doi.org/10.7554/eLife.36398
Krotman, Y., Yergaliyev, T.M., Alexander Shani, R., Avrahami, Y., Szitenberg, A., 2020.
Dissecting the factors shaping fish skin microbiomes in a heterogeneous inland water
system. Microbiome 8, 9. https://doi.org/10.1186/s40168-020-0784-5
Lafferty, K.D., Harvell, C.D., Conrad, J.M., Friedman, C.S., Kent, M.L., Kuris, A.M., Powell,
E.N., Rondeau, D., Saksida, S.M., 2015. Infectious Diseases Affect Marine Fisheries and
Aquaculture Economics. Ann. Rev. Mar. Sci. 7, 471–496.
https://doi.org/10.1146/annurev-marine-010814-015646
Larsen, A., Tao, Z., Bullard, S.A., Arias, C.R., 2013. Diversity of the skin microbiota of fishes:
evidence for host species specificity. FEMS Microbiol. Ecol. 85, 483–494.
https://doi.org/10.1111/1574-6941.12136
Lee, Y.K., Mazmanian, S.K., 2010. Has the microbiota played a critical role in the evolution of
the adaptive immune system? Science (80). https://doi.org/10.1126/science.1195568
Legrand, T.P.R.A., Catalano, S.R., Wos-Oxley, M.L., Stephens, F., Landos, M., Bansemer,
M.S., Stone, D.A.J., Qin, J.G., Oxley, A.P.A., 2018. The Inner Workings of the Outer
Surface: Skin and Gill Microbiota as Indicators of Changing Gut Health in Yellowtail
Kingfish. Front. Microbiol. 8, 2664. https://doi.org/10.3389/fmicb.2017.02664
Legrand, T.P.R.A., Catalano, S.R., Wos-Oxley, M.L., Wynne, J.W., Weyrich, L.S., Oxley,
A.P.A., 2020a. Antibiotic-induced alterations and repopulation dynamics of yellowtail
kingfish microbiota. Anim. Microbiome 2, 26. https://doi.org/10.1186/s42523-020-00046-
4
Legrand, T.P.R.A., Wynne, J.W., Weyrich, L.S., Oxley, A.P.A., 2020b. A microbial sea of
possibilities: current knowledge and prospects for an improved understanding of the fish
microbiome. Rev. Aquac. 12, 1101–1134. https://doi.org/10.1111/raq.12375
Llewellyn, M.S., Boutin, S., Hoseinifar, S.H., Derome, N., 2014. Teleost microbiomes: The
state of the art in their characterization, manipulation and importance in aquaculture and
fisheries. Front. Microbiol. https://doi.org/10.3389/fmicb.2014.00207
Lokesh, J., Kiron, V., Sipkema, D., Fernandes, J.M.O., Moum, T., 2019. Succession of
embryonic and the intestinal bacterial communities of Atlantic salmon (Salmo salar)

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
222
reveals stage-specific microbial signatures. Microbiologyopen 8, e00672.
https://doi.org/10.1002/mbo3.672
Lowrey, L., Woodhams, D.C., Tacchi, L., Salinas, I., 2015. Topographical mapping of the
rainbow trout (Oncorhynchus mykiss) microbiome reveals a diverse bacterial community
with antifungal properties in the skin. Appl. Environ. Microbiol. 81, 6915–6925.
https://doi.org/10.1128/AEM.01826-15
Luna, G.M., Quero, G.M., Kokou, F., Kormas, K., 2022. Time to integrate biotechnological
approaches into fish gut microbiome research. Curr. Opin. Biotechnol. 73, 121–127.
https://doi.org/10.1016/J.COPBIO.2021.07.018
Minich, J., Petrus, S., Michael, J.D., Michael, T.P., Knight, R., Allen, E.E., 2020a. Temporal,
Environmental, and Biological Drivers of the Mucosal Microbiome in a Wild Marine Fish,
Scomber japonicus Downloaded from. https://doi.org/10.1128/mSphere.00401-20
Minich, J., Poore, G.D., Jantawongsri, K., Johnston, C., Bowie, K., Bowman, J., Knight, R.,
Nowak, B., Allen, E.E., 2020b. Microbial ecology of atlantic salmon (Salmo salar)
hatcheries: Impacts of the built environment on fish mucosal microbiota. Appl. Environ.
Microbiol. 86. https://doi.org/10.1128/AEM.00411-20
Nikouli, Meziti, Antonopoulou, Mente, Kormas, 2019. Host-Associated Bacterial Succession
during the Early Embryonic Stages and First Feeding in Farmed Gilthead Sea Bream
(Sparus aurata). Genes (Basel). 10, 483. https://doi.org/10.3390/genes10070483
Noga, E.J., Borron, P.J., Hinshaw, J., Gordon, W.C., Gordon, L.J., Seo, J.-K., 2010.
Identification of histones as endogenous antibiotics in fish and quantification in rainbow
trout (Oncorhynchus mykiss) skin and gill. Fish Physiol. Biochem. 2010 371 37, 135–152.
https://doi.org/10.1007/S10695-010-9422-7
Offret, C., Desriac, F., Le Chevalier, P., Mounier, J., Jégou, C., Fleury, Y., 2016. Spotlight on
Antimicrobial Metabolites from the Marine Bacteria Pseudoalteromonas: Chemodiversity
and Ecological Significance. Mar. Drugs 14, 129. https://doi.org/10.3390/md14070129
Persson, P., Sundell, K., Björnsson, B.T., Lundqvist, H., 2005. Calcium metabolism and
osmoregulation during sexual maturation of river running Atlantic salmon. J. Fish Biol.
52, 334–349. https://doi.org/10.1111/J.1095-8649.1998.TB00801.X
Petersen, C., Round, J.L., 2014. Defining dysbiosis and its influence on host immunity and
disease. Cell. Microbiol. 16, 1024–1033. https://doi.org/10.1111/cmi.12308
Pimentel, T., Marcelino, J., Ricardo, F., Soares, A.M.V.M., Calado, R., 2017. Bacterial
communities 16S rDNA fingerprinting as a potential tracing tool for cultured seabass
Dicentrarchus labrax. Sci. Rep. 7, 1–10. https://doi.org/10.1038/s41598-017-11552-y
Pindling, S., Azulai, D., Zheng, B., Dahan, D., Perron, G.G., 2018. Dysbiosis and early
mortality in zebrafish larvae exposed to subclinical concentrations of streptomycin. FEMS
Microbiol. Lett. 365, 188. https://doi.org/10.1093/femsle/fny188

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
223
Pratte, Z.A., Besson, M., Hollman, R.D., Stewart, F.J., 2018. The Gills of Reef Fish Support a
Distinct Microbiome Influenced by Host-Specific Factors. https://doi.org/10.1128/AEM
Rajeev, R., Adithya, K.K., Kiran, G.S., Selvin, J., 2021. Healthy microbiome: a key to
successful and sustainable shrimp aquaculture. Rev. Aquac. 13, 238–258.
https://doi.org/10.1111/RAQ.12471
Reverter, M., Sasal, P., Tapissier-Bontemps, N., Lecchini, D., Suzuki, M., 2017.
Characterisation of the gill mucosal bacterial communities of four butterflyfish species: a
reservoir of bacterial diversity in coral reef ecosystems. FEMS Microbiol. Ecol. 93, 51.
https://doi.org/10.1093/femsec/fix051
Richards, G.P., Watson, M.A., Needleman, D.S., Uknalis, J., Boyd, E.F., Fay, J.P., 2017.
Mechanisms for Pseudoalteromonas piscicida induced killing of vibrios and other
bacterial pathogens. Appl. Environ. Microbiol. 83, 175–192.
https://doi.org/10.1128/AEM.00175-17
Rodriguez-Estrada, U., Satoh, S., Haga, Y., Fushimi, H., Sweetman, J., 2013. Effects of
inactivated enterococcus faecalis and mannan oligosaccharide and their combination on
growth, immunity, and disease protection in rainbow trout. N. Am. J. Aquac. 75, 416–428.
https://doi.org/10.1080/15222055.2013.799620
Saborido-Rey, F., Kjesbu, O.S., 2005. Growth and maturation dynamics.
Saksida, S.M., Gardner, I., Lent, M.L., 2014. Transmission of infected agents between wild
and farmed fish, in: Woo, P.T.K., Bruno, D.W. (Eds.), Saksida SM, Gardner I, Kent ML.
2014. Transmission of Infectious Agents between Wild and Farmed Fish. In Diseases
and Disorders of Finfish in Cage Culture. CABI, Wallingford, UK, p. 313.
Schar, D., Klein, E.Y., Laxminarayan, R., Gilbert, M., Van Boeckel, T.P., 2020. Global trends
in antimicrobial use in aquaculture. Sci. Rep. 10, 21878. https://doi.org/10.1038/s41598-
020-78849-3
Seebacher, F., White, C.R., Franklin, C.E., 2015. Physiological plasticity increases resilience
of ectothermic animals to climate change. Nat. Clim. Chang. 5, 61–66.
https://doi.org/10.1038/nclimate2457
Sehnal, L., Brammer-Robbins, E., Wormington, A.M., Blaha, L., Bisesi, J., Larkin, I., Martyniuk,
C.J., Simonin, M., Adamovsky, O., 2021. Microbiome Composition and Function in
Aquatic Vertebrates: Small Organisms Making Big Impacts on Aquatic Animal Health.
Front. Microbiol. https://doi.org/10.3389/fmicb.2021.567408
Shade, A., Peter, H., Allison, S.D., Baho, D.L., Berga, M., Bürgmann, H., Huber, D.H.,
Langenheder, S., Lennon, J.T., Martiny, J.B.H., Matulich, K.L., Schmidt, T.M.,
Handelsman, J., 2012. Fundamentals of microbial community resistance and resilience.
Front. Microbiol. 3, 417. https://doi.org/10.3389/fmicb.2012.00417
Smith, P., Willemsen, D., Popkes, M., Metge, F., Gandiwa, E., Reichard, M., Valenzano, D.R.,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
224
2017. Regulation of life span by the gut microbiota in the short-lived african turquoise
killifish. Elife 6. https://doi.org/10.7554/ELIFE.27014
Stephens, W.Z., Burns, A.R., Stagaman, K., Wong, S., Rawls, J.F., Guillemin, K., Bohannan,
B.J.M., 2016. The composition of the zebrafish intestinal microbial community varies
across development. ISME J. 10, 644–654. https://doi.org/10.1038/ismej.2015.140
Sun, Y.Z., Xia, H.Q., Yang, H.L., Wang, Y.L., Zou, W.C., 2014. TLR2 signaling may play a key
role in the probiotic modulation of intestinal microbiota in grouper Epinephelus coioides.
Aquaculture 430, 50–56. https://doi.org/10.1016/j.aquaculture.2014.03.042
Sylvain, F.-É., Holland, A., Bouslama, S., Audet-Gilbert, É., Lavoie, C., Val, A.L., Derome, N.,
2020. Fish Skin and Gut Microbiomes Show Contrasting Signatures of Host Species and
Habitat Downloaded from. https://doi.org/10.1128/AEM.00789-20
Tapia-Paniagua, S.T., Ceballos-Francisco, D., Balebona, M.C., Esteban, M.Á., Moriñigo,
M.Á., 2018. Mucus glycosylation, immunity and bacterial microbiota associated to the
skin of experimentally ulcered gilthead seabream (Sparus aurata). Fish Shellfish
Immunol. 75, 381–390. https://doi.org/10.1016/j.fsi.2018.02.006
Tarnecki, A.M., Brennan, N.P., Schloesser, R.W., Rhody, N.R., 2019. Shifts in the Skin-
Associated Microbiota of Hatchery-Reared Common Snook Centropomus undecimalis
During Acclimation to the Wild. Microb. Ecol. 77, 770–781.
https://doi.org/10.1007/s00248-018-1252-7
Thompson, L.R., Sanders, J.G., McDonald, D., Amir, A., Ladau, J., Locey, K.J., Prill, R.J.,
Tripathi, A., Gibbons, S.M., Ackermann, G., Navas-Molina, J.A., Janssen, S., Kopylova,
E., Vázquez-Baeza, Y., González, A., Morton, J.T., Mirarab, S., Xu, Z.Z., Jiang, L.,
Haroon, M.F., Kanbar, J., Zhu, Q., Song, S.J., Kosciolek, T., Bokulich, N.A., Lefler, J.,
Brislawn, C.J., Humphrey, G., Owens, S.M., Hampton-Marcell, J., Berg-Lyons, D.,
McKenzie, V., Fierer, N., Fuhrman, J.A., Clauset, A., Stevens, R.L., Shade, A., Pollard,
K.S., Goodwin, K.D., Jansson, J.K., Gilbert, J.A., Knight, R., Agosto Rivera, J.L., Al-
Moosawi, L., Alverdy, J., Amato, K.R., Andras, J., Angenent, L.T., Antonopoulos, D.A.,
Apprill, A., Armitage, D., Ballantine, K., Bárta, J., Baum, J.K., Berry, A., Bhatnagar, A.,
Bhatnagar, M., Biddle, J.F., Bittner, L., Boldgiv, B., Bottos, E., Boyer, D.M., Braun, J.,
Brazelton, W., Brearley, F.Q., Campbell, A.H., Caporaso, J.G., Cardona, C., Carroll, J.L.,
Cary, S.C., Casper, B.B., Charles, T.C., Chu, H., Claar, D.C., Clark, R.G., Clayton, J.B.,
Clemente, J.C., Cochran, A., Coleman, M.L., Collins, G., Colwell, R.R., Contreras, M.,
Crary, B.B., Creer, S., Cristol, D.A., Crump, B.C., Cui, D., Daly, S.E., Davalos, L.,
Dawson, R.D., Defazio, J., Delsuc, F., Dionisi, H.M., Dominguez-Bello, M.G., Dowell, R.,
Dubinsky, E.A., Dunn, P.O., Ercolini, D., Espinoza, R.E., Ezenwa, V., Fenner, N., Findlay,
H.S., Fleming, I.D., Fogliano, V., Forsman, A., Freeman, C., Friedman, E.S., Galindo, G.,
Garcia, L., Garcia-Amado, M.A., Garshelis, D., Gasser, R.B., Gerdts, G., Gibson, M.K.,

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
225
Gifford, I., Gill, R.T., Giray, T., Gittel, A., Golyshin, P., Gong, D., Grossart, H.P., Guyton,
K., Haig, S.J., Hale, V., Hall, R.S., Hallam, S.J., Handley, K.M., Hasan, N.A., Haydon,
S.R., Hickman, J.E., Hidalgo, G., Hofmockel, K.S., Hooker, J., Hulth, S., Hultman, J.,
Hyde, E., Ibáñez-Álamo, J.D., Jastrow, J.D., Jex, A.R., Johnson, L.S., Johnston, E.R.,
Joseph, S., Jurburg, S.D., Jurelevicius, D., Karlsson, A., Karlsson, R., Kauppinen, S.,
Kellogg, C.T.E., Kennedy, S.J., Kerkhof, L.J., King, G.M., Kling, G.W., Koehler, A. V.,
Krezalek, M., Kueneman, J., Lamendella, R., Landon, E.M., Lanede Graaf, K., LaRoche,
J., Larsen, P., Laverock, B., Lax, S., Lentino, M., Levin, I.I., Liancourt, P., Liang, W., Linz,
A.M., Lipson, D.A., Liu, Y., Lladser, M.E., Lozada, M., Spirito, C.M., MacCormack, W.P.,
MacRae-Crerar, A., Magris, M., Martín-Platero, A.M., Martín-Vivaldi, M., Martínez, L.M.,
Martínez-Bueno, M., Marzinelli, E.M., Mason, O.U., Mayer, G.D., McDevitt-Irwin, J.M.,
McDonald, J.E., McGuire, K.L., McMahon, K.D., McMinds, R., Medina, M., Mendelson,
J.R., Metcalf, J.L., Meyer, F., Michelangeli, F., Miller, K., Mills, D.A., Minich, J., Mocali,
S., Moitinho-Silva, L., Moore, A., Morgan-Kiss, R.M., Munroe, P., Myrold, D., Neufeld,
J.D., Ni, Y., Nicol, G.W., Nielsen, S., Nissimov, J.I., Niu, K., Nolan, M.J., Noyce, K.,
O’Brien, S.L., Okamoto, N., Orlando, L., Castellano, Y.O., Osuolale, O., Oswald, W.,
Parnell, J., Peralta-Sánchez, J.M., Petraitis, P., Pfister, C., Pilon-Smits, E., Piombino, P.,
Pointing, S.B., Pollock, F.J., Potter, C., Prithiviraj, B., Quince, C., Rani, A., Ranjan, R.,
Rao, S., Rees, A.P., Richardson, M., Riebesell, U., Robinson, C., Rockne, K.J.,
Rodriguezl, S.M., Rohwer, F., Roundstone, W., Safran, R.J., Sangwan, N., Sanz, V.,
Schrenk, M., Schrenzel, M.D., Scott, N.M., Seger, R.L., Seguinorlando, A., Seldin, L.,
Seyler, L.M., Shakhsheer, B., Sheets, G.M., Shen, C., Shi, Y., Shin, H., Shogan, B.D.,
Shutler, D., Siegel, J., Simmons, S., Sjöling, S., Smith, D.P., Soler, J.J., Sperling, M.,
Steinberg, P.D., Stephens, B., Stevens, M.A., Taghavi, S., Tai, V., Tait, K., Tan, C.L.,
Taş, N., Taylor, D.L., Thomas, T., Timling, I., Turner, B.L., Urich, T., Ursell, L.K., Van Der
Lelie, D., Van Treuren, W., Van Zwieten, L., Vargas-Robles, D., Thurber, R.V., Vitaglione,
P., Walker, D.A., Walters, W.A., Wang, S., Wang, T., Weaver, T., Webster, N.S., Wehrle,
B., Weisenhorn, P., Weiss, S., Werner, J.J., West, K., Whitehead, A., Whitehead, S.R.,
Whittingham, L.A., Willerslev, E., Williams, A.E., Wood, S.A., Woodhams, D.C., Yang,
Y., Zaneveld, J., Zarraonaindia, I., Zhang, Q., Zhao, H., 2017. A communal catalogue
reveals Earth’s multiscale microbial diversity. Nature 551, 457–463.
https://doi.org/10.1038/nature24621
Torrecillas, S., Makol, A., Betancor, M.B., Montero, D., Caballero, M.J., Sweetman, J.,
Izquierdo, M., 2013. Enhanced intestinal epithelial barrier health status on European sea
bass (Dicentrarchus labrax) fed mannan oligosaccharides. Fish Shellfish Immunol. 34,
1485–1495. https://doi.org/10.1016/j.fsi.2013.03.351
Trivedi, B., 2012. Microbiome: the surface brigade. Nature 492, S60–S61.

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream
226
Uren Webster, T.M., Rodriguez-Barreto, D., Consuegra, S., Garcia de Leaniz, C., 2020.
Cortisol-Related Signatures of Stress in the Fish Microbiome. Front. Microbiol. 0, 1621.
https://doi.org/10.3389/FMICB.2020.01621
Vasemägi, A., Visse, M., Kisand, V., 2017. Effect of Environmental Factors and an Emerging
Parasitic Disease on Gut Microbiome of Wild Salmonid Fish. mSphere 2, 418–435.
https://doi.org/10.1128/msphere.00418-17
Walleser, L.R., Sandheinrich, M.B., Howard, D.R., Gaikowski, M.P., Amberg, J.J., 2014.
Spatial and Temporal Variation of the Gill Rakers of Gizzard Shad and Silver Carp in
Three Midwestern Rivers. North Am. J. Fish. Manag. 34, 875–884.
https://doi.org/10.1080/02755947.2014.920740
Wilkes Walburn, J., Wemheuer, B., Thomas, T., Copeland, E., O’Connor, W., Booth, M.,
Fielder, S., Egan, S., 2019. Diet and diet-associated bacteria shape early microbiome
development in Yellowtail Kingfish (Seriola lalandi). Microb. Biotechnol. 12, 275–288.
https://doi.org/10.1111/1751-7915.13323
Wu, Y., Xiao, F., Wang, C., Shu, L., Zheng, X., Xu, K., Yu, X., Zhang, K., Luo, H., Yang, Y.,
He, Z., Yan, Q., 2020. The Beta-Diversity of Siganus fuscescens-Associated Microbial
Communities From Different Habitats Increases With Body Weight. Front. Microbiol. 11,
1562. https://doi.org/10.3389/fmicb.2020.01562
Yan, Q., Li, J., Yu, Y., Wang, J., He, Z., Van Nostrand, J.D., Kempher, M.L., Wu, L., Wang,
Y., Liao, L., Li, X., Wu, S., Ni, J., Wang, C., Zhou, J., 2016. Environmental filtering
decreases with fish development for the assembly of gut microbiota. Environ. Microbiol.
18, 4739–4754. https://doi.org/10.1111/1462-2920.13365
Yılmaz, S., Yılmaz, E., Dawood, M.A.O., Ringø, E., Ahmadifar, E., Abdel-Latif, H.M.R., 2021.
Probiotics, prebiotics, and synbiotics used to control vibriosis in fish: A review.
Aquaculture 737514. https://doi.org/10.1016/J.AQUACULTURE.2021.737514
Zarkasi, K.Z., Abell, G.C.J., Taylor, R.S., Neuman, C., Hatje, E., Tamplin, M.L., Katouli, M.,
Bowman, J.P., 2014. Pyrosequencing-based characterization of gastrointestinal bacteria
of Atlantic salmon (Salmo salar L.) within a commercial mariculture system. J. Appl.
Microbiol. 117, 18–27. https://doi.org/10.1111/jam.12514
Zhang, Xiaoting, Ding, L., Yu, Y., Kong, W., Yin, Y., Huang, Z., Zhang, Xuezhen, Xu, Z., 2018.
The Change of Teleost Skin Commensal Microbiota Is Associated With Skin Mucosal
Transcriptomic Responses During Parasitic Infection by Ichthyophthirius multifillis. Front.
Immunol. 9, 2972. https://doi.org/10.3389/fimmu.2018.02972
Zhang, Z., Li, D., Xu, W., Tang, R., Li, L., 2019. Microbiome of Co-cultured Fish Exhibits Host
Selection and Niche Differentiation at the Organ Scale. Front. Microbiol. 10, 2576.
https://doi.org/10.3389/fmicb.2019.02576

FCUP To be or not to be diseased: microbial dynamics and dysbiosis in farmed European seabass and gilthead seabream 227
APPENDIX A
List of additional publications
List of additional publications that are within the scope of fish microbial research and that I
participated in as a co-author.
Xavier, R., Mazzei, R., Pérez-Losada, M., Rosado, D., Santos, J.L., Veríssimo, A., Soares,
M.C., 2019. A Risky Business? Habitat and Social Behavior Impact Skin and Gut
Microbiomes in Caribbean Cleaning Gobies. Front. Microbiol. 10, 716.
https://doi.org/10.3389/fmicb.2019.00716
Xavier, R., Pereira, A., Pagan, A., Hendrick, G.C., Nicholson, M.D., Rosado, D., Soares, M.C.,
Pérez-Losada, M., Sikkel, P.C., 2020. The effects of environment and ontogeny on the
skin microbiome of two Stegastes damselfishes (Pomacentridae) from the eastern
Caribbean Sea. Mar. Biol. 167, 102. https://doi.org/10.1007/s00227-020-03717-7




















