
Temporal dynamics of biogeochemical processes at the Norman Landfill site

This article was downloaded by: [Gabriella Caruso]On: 17 April 2013, At: 07:01Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Advances in Oceanography andLimnologyPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/taol20
Microbial enzymatic activities andprokaryotic abundance in the upwellingsystem of the Straits of Messina(Sicily): distribution, dynamics andbiogeochemical considerationsG. Caruso a , F. Azzaro a , R. La Ferla a , F. De Pasquale a , F. Raffaa & F. Decembrini aa Istituto per l’Ambiente Marino Costiero, Consiglio Nazionaledelle Ricerche (IAMC-CNR), Messina, ItalyVersion of record first published: 17 Apr 2013.
To cite this article: G. Caruso , F. Azzaro , R. La Ferla , F. De Pasquale , F. Raffa & F. Decembrini(2013): Microbial enzymatic activities and prokaryotic abundance in the upwelling system of theStraits of Messina (Sicily): distribution, dynamics and biogeochemical considerations, Advances inOceanography and Limnology, DOI:10.1080/19475721.2012.755568
To link to this article: http://dx.doi.org/10.1080/19475721.2012.755568
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representationthat the contents will be complete or accurate or up to date. The accuracy of anyinstructions, formulae, and drug doses should be independently verified with primarysources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

Microbial enzymatic activities and prokaryotic abundance in the
upwelling system of the Straits of Messina (Sicily): distribution,
dynamics and biogeochemical considerations
G. Caruso�, F. Azzaro, R. La Ferla, F. De Pasquale, F. Raffa and F. Decembrini
Istituto per l’Ambiente Marino Costiero, Consiglio Nazionale delle Ricerche (IAMC-CNR),Spianata S. Raineri 86, 98122 Messina, Italy
(Received 27 June 2012; final version received 29 November 2012)
Three microbial enzymatic activities involved in organic matter transformation(Leucine AminoPeptidase, LAP, b-glucosidase, b-GLU and Alkaline Phosphatase,AP) and prokaryotic abundance were studied in the Straits of Messina upwelling sys-tem. Samplings were performed monthly, between June 2003 and March 2004, at threestations with different hydrodynamic patterns. Potential hydrolysis rates were estimat-ed using specific fluorogenic substrates. Unlike total prokaryotic abundances, quitespatially homogeneous, enzymatic values described different scenarios for each exam-ined station. At the southern Sicilian side (Scaletta) the highest average activity rateswere recorded (LAP: 72.19 � 19.14 nmol Cleu dm
�3h�1, b-GLU, 12.53 � 3.15 nmolCglu dm
�3h�1, AP, 113.59 � 35.47 nmol PO4dm�3h�1), while at the Calabrian side
(Pellaro) the minimum values of LAP, b-GLU and AP were measured. Enzymatic ac-tivity values showed seasonal patterns everywhere, with summer values which wereone order of magnitude higher than those recorded in other seasons and autumn valueswhich sharply decreased. During winter, the enhanced rates of b-GLU and APsuggested the occurrence of active polysaccharide decomposition and phosphorus re-generation. The variability in the enzymatic activity patterns recorded in the Straitsarea was indicative of biogeochemical features differing among the examined stations;this resulted in changes in the balance between production and decompositionprocesses.
Keywords: microorganisms; enzymatic activity; biogeochemistry; upwelling area;Straits of Messina
1. Introduction
In natural ecosystems, microorganisms are recognised to play a key role in organic matter
processing and biogeochemical cycles, through their enzymatic activities on both dis-
solved and particulate polymers; in this way, they regulate the fluxes of matter and energy
through the trophic web [1–3]. Several studies [4–6 and references herein] have reviewed
the enzymatic patterns in different aquatic ecosystems, pointing out their significance for
the comprehension of ecological processes. Microbial metabolism closely reflects
changes occurring in environmental conditions [5,6], and, in particular, heterotrophic bac-
teria may adapt their metabolism depending on organic carbon supply [3]. As a conse-
quence, microbial enzymes involved in organic matter transformation are expected to
vary their activity patterns to respond to the time-scale variability of environmental
parameters. This happens particularly in upwelling regions, that are highly productive
*Corresponding author. Email: [email protected]
� 2013 Taylor & Francis
Advances in Oceanography and Limnology, 2013
http://dx.doi.org/10.1080/19475721.2012.755568
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

systems, due to the richness in inorganic nutrients, which are recycled through organic
matter mineralisation and fertilise surface waters. Heterotrophic bacterioplankton has
shown to be an important component within the prokaryotic community inhabiting up-
welling ecosystems, which is able to quickly respond to the pulsed production cycles typi-
cal of these areas [7,8] by channelling a significant amount of primary production [9–11].
In the Straits of Messina, the importance of microbial activities in organic matter
processing and in nutrient cycles has been neglected till now. Located at the centre of the
Mediterranean Sea, the Straits connect two adjacent basins, the Ionian and the Tyrrhe-
nian, with different hydrographic properties. Their condition of ‘transitional area’ is relat-
ed to a great complexity in terms of hydrological structures, even on a short spatial scale
(20 km length). For many decades, the Straits have been the object of oceanographic stud-
ies due to the occurrence of the upwelling and water turbulence phenomena, consequent
to strong tidal currents, which mix warmer Tyrrhenian Surface Waters (TSW) with cooler
Levantine Intermediate Waters (LIW) [12–14]. Several researches concerning the influ-
ence of the divergence (which pumps nutrient-rich waters in the euphotic layer) on bio-
logical components as well as on the primary production process were largely performed
in not too recent times [14–19]. Conversely, the pelagic microbial community, and its dy-
namics and ecological significance, were scarcely investigated [20–22] and are still most-
ly unknown.
Hydrodynamic phenomena occurring in the Messina Straits are supposed to affect sig-
nificantly the prokaryotes’ metabolism, thus suggesting the possible use of this ecosystem
as a natural mesocosm to study the effects of water divergence on microbes–organic mat-
ter interactions, and in general, on biogeochemical processes.
In order to estimate the importance of the role played by microorganisms in the organ-
ic matter decomposition in the Straits of Messina, three microbial enzymes (Leucine
AminoPeptidase [LAP], b-glucosidase, [b-GLU], and Alkaline Phosphatase [AP]) and
the abundance of total prokaryotes were investigated. The ecological significance of these
enzymes is widely known in different aquatic environments: LAP and b-GLU are in-
volved in the decomposition of proteins and polysaccharides, respectively, and therefore
in the carbon cycle. AP regulates the decomposition of organic phosphates, controlling
phosphorus regeneration and then the P cycle [4,5,23]; as a consequence, this enzyme is
conventionally used as an indicator of P limitation [24–26]. The reciprocal ratios among
the three measured enzymes provide further information on organic matter quality
(LAP/b-GLU) as well as on the nutritional status of a biological system (AP/LAP). In par-
ticular, high LAP/b-GLU ratios indicate that the flux of organic matter occurs preferen-
tially through proteins, which are more readily available to bacteria, instead of
polysaccharides [27,28]. The AP/LAP ratios provide insights on the relative importance
of P mineralisation compared to N mobilisation. Sala et al. [29] suggested the use of this
ratio for evaluating ‘in situ’ P versus N limitation of microbial communities. Moreover,
the molar ratios of enzymatic activity rates give an indication of the potential biogeo-
chemical implications of the microbial activities over organic compounds in terms of mo-
lar C/P and N/P ratios.
The aims of the present study were: 1) to evaluate changes occurring in the decom-
position rates of some organic substrates by analysing the distribution of the microbial
enzymatic activities at three coastal stations of the Straits of Messina, differently affect-
ed by hydrodynamic phenomena, in order; 2) to follow the time evolution of enzymatic
activities in relation to the main environmental parameters; 3) to provide insights
on the biogeochemical implications of the enzymatic dynamics for organic matter
processing.
2 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

2. Materials and methods
2.1. Samplings
From June 2003 to March 2004, seawater samples were collected monthly from three
coastal stations of the Straits, at depths within the upper 100 m (Figure 1). The stations
were selected to be representative of different environmental conditions: Guardia is a sta-
tion located where the formation of ‘Messina Mixed Water’ (MMW) takes place and is
affected by water upwelling; Pellaro is a station located at the boundary of the
‘divergence area’ and has got a steady water column; Scaletta is a station situated in the
output zone of the system where MMW exits from the Straits flowing southward [30–33].
To obtain a comparable data set of this highly dynamic ecosystem, the time-series sam-
pling was carried out during the period between high and low tidal phases during spring-
tide. In such a condition, the tidal phases could have strong effects at Guardia, due to the
effects of vertical water mixing, whereas they had a limited role at the other stations.
Samplings were carried out using 10-litre sterile Niskin bottles at surface (0.5 m
depth) and at three depths: above (10 m), within (30 m, in both Scaletta and Guardia;
50 m, in Pellaro) and below (100 m) the Chlorophyll Maximum (CM).
2.2. Physical and chemical measurements
Temperature (T), salinity (S) and dissolved oxygen (DO) were measured using a SeaBird
Electronics SBE-911þCTOD profiler. The density dataset was analysed in order to calcu-
late the difference in density between surface and bottom layers during each sampling,
according to [34]. The median of the differences was used as the threshold value to dis-
criminate stratified (density difference > median value) from mixed (density difference <
median value) water columns.
Figure 1. Map of the study area showing the three sampling stations.
Advances in Oceanography and Limnology 3
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

2.3. Nutrients
For the determination of ammonia (NH4), nitrite þ nitrate (NO2,3) and orthophosphate
(PO4) contents, water samples were filtered through Whatman GF/F glass-fibre filters and
kept frozen (�20 �C). NH4 concentration was estimated according to Aminot and Chausse-
pied [35], while NO2,3 and PO4 concentrations were estimated according to Strickland and
Parsons [36].
2.4. Chlorophyll a (Chl-a)
For the determination of Chl-a, as an index of phytoplankton biomass, water samples
(0.8–1.0 l) were filtered through Whatman GF/F glass-fibre filters, according to Lazzara
et al. [37]. After filtration, the filters were immediately stored at �20 �C. Photosyntheticpigments were extracted in 90% acetone and read before and after acidification. Determi-
nations were carried out with a Varian Eclipse spectrofluorometer. Maximum excitation
and emission wavelengths (431 and 667 nm respectively) were selected on a prescan with
a solution of chlorophyll a from Anacystis nidulans (by Sigma Co).
2.5. Microbial enzymatic activities
Leucine AminoPeptidase (LAP), b-glucosidase (b-GLU) and Alkaline Phosphatase (AP)
activities were determined according to Hoppe [38], using the fluorogenic substrates L-
leucine-7-amido-4-methylcoumarine hydrochloride (Leu-MCA), 4-methylumbelliferyl-
b-D-glucopyranoside (MUF-glu) and 4-methylumbelliferyl phosphate (MUF-phosphate)
(Sigma), for LAP, b-GLU and AP, respectively. All the enzymatic activities were mea-
sured with a Hitachi F-2000 spectrofluorometer in terms of potential activities (maximum
hydrolysis velocity V max, in nanomoles of Cleu or Cglu or PO4 released per dm3 and per
hour, nmol dm�3h�1), using a multi-concentration method based on the addition of in-
creasing amounts (0.2 to 20 mM) of a 5 mM substrate stock solution to 10 ml sub-samples
and incubation at in situ temperature for 3 hours. Measurements were performed at
380/440 nm excitation/emission wavelengths, for LAP, and at 365/445 nm for b-GLU
and AP, respectively [39]. The nanomoles of leucine potentially released by LAP were
converted into nanomoles of C and N taking into consideration that leucine contains 6
atoms of C and 1 atom of N; on the contrary, one mole of AP potentially releases 1 mole
of PO4. As a consequence, a factor of 14 was used to convert the nanomoles of leucine re-
leased into nanograms of N, while a factor of 31 was used to convert the nanomoles of P
released by AP into nanograms of P [40].
For later comparative analyses among the stations, the enzymatic activity rates were also
normalised to prokaryotic abundance values to calculate the cell-specific activity values.
2.6. Total prokaryotic abundance (PA)
To estimate the abundance of total prokaryotes (Bacteria and Archaea), volumes (1 to
5 ml) of seawater were filtered through 0.22 mm porosity polycarbonate Black mem-
branes (Nuclepore); the filters were stained with a solution of 40,6-diamidino-2-
phenylindole (DAPI, according to Porter and Feig [41]) and observed under a Zeiss AXI-
OPLAN 2 Imaging epifluorescence microscope. The counts were expressed in cells ml�1.
2.7. Statistical analysis of data
The statistical significance of the differences among the values measured for the parame-
ters was checked through a three-way analysis of variance (ANOVA), considering the
4 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

following three factors as the major sources of variation: ‘station’ (at three levels, i.e. Sca-
letta, Pellaro and Guardia); ‘months’ (at ten levels, June to March) and ‘depths’ (at four
levels, i.e. surface, above, within and below the CM), factors were tested for their simple
effects and interactions (stations X months, stations X depths, months X depths, stations
X months X depths). Prior to their analysis, the few missing data were replaced by the
mean for their group data. A normality test was performed on the measured data, to assess
their compliance with normal distribution; data that did not meet this assumption were
logarithmically transformed to attain homogeneity of variance.
The Pearson’s r correlation coefficients were calculated to determine the degree of as-
sociation between microbial and environmental parameters at each station separately.
The ANOVA test was performed using SIGMA-STAT software, version 3.1 (SYSTAT
Software, Inc.).
The whole physico-chemical and biological dataset was further subjected to Principal
Component Analysis (PCA), in order to identify the main factors driving the microbial
community dynamics at each station separately. This multivariate analysis shows the dis-
tribution of the different variables in new linear combinations (principal components,
PC). Only PC with Eigenvalues > 1 were considered. PCA was carried out using the
PRIMER 6 software, version 6b R6 (Plymouth Marine Laboratory, United Kingdom).
According to the results yielded by the density dataset analysis, the enzymatic dataset
was shared in two groups, assigned to ‘stratified’ and ‘mixed’ water regimes; they under-
went a Mann-Whitney test to verify whether the differences related to the two different
hydrological conditions were statistically significant.
3. Results
The whole original data set of physical, chemical and biological parameters is reported in
Table 1, where the annual average, minimum and maximum values are shown.
3.1. Hydrological characterisation
During the annual cycle, the waters of the Straits of Messina were characterised by a wide
variability in T (13.96–27.22 �C), S (37.52–38.94) and st values (25.5–29.1). T and S pat-
terns (Figure 2) varied dramatically among the three stations examined, showing high
variability in the layers comprised between isopycnal 25.5 and 28.5 st. At Guardia, salini-
ty followed a stratified distribution, increasing from 37.52 to 38.77 (25.27 �C < T <
14 �C), whereas at Pellaro, stratification was evident in the thermic values which de-
creased from 27.2 �C to 14.1 �C (38.55 < S < 38.94). At Scaletta, T (range 13.96–
25.76 �C) and S (range 37.65–38.76) values fell between those recorded in Guardia and
Pellaro (Table 1). The thermo-haline differences among the stations were negligible at
the 100 m depth, where a water mass close to 29.0 st was observed. In autumn (data not
shown), the surface waters were characterised by a prominent thermocline which intensi-
fied the stratification over the whole area (DT� 5–10 �C).From the thermo-haline structures observed at the three examined stations (Figure 2),
two main water masses were identified, the Tyrrhenian Surface Water (TSW, S < 37.9)
and the Ionian Water (IW, S > 38.1). Considering the salinity distribution, the hydrody-
namic regimen of the Straits was as follows: TSW (less saline) entered from the north
close to Guardia (DS ¼ 1.1) and underwent mixing with the IW (more saline) coming
from the south and a horizontal diffusion took place along the Straits. Therefore, this
modified water, characterised by a subsurface salinity minimum, spread southward close
Advances in Oceanography and Limnology 5
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

Table1.
Annualaveragevalues,x,andstandarddeviation(inbrackets)ofphysical,chem
icalandbiologicalparam
eters,withtheirranges
ofvariation,measured
attheexam
ined
stations.
TDO
PO4
NH4
NO2,3
Stations
(�C)
S(m
ll�
1)
(mM)
(mM)
(mM)
Scaletta
x(s.d.)
16.77(2.84)
38.43(0.24)
5.25(0.58)
0.23(0.13)
0.92(0.97)
3.55(1.63)
Min-m
ax13.96–25.76
37.65–38.76
4.35–6.44
0.03–0.58
0.09–3.82
0.30–6.43
Guardia
x(s.d.)
16.89(2.84)
38.21(0.34)
5.34(0.63)
0.35(0.23)
0.75(0.79)
3.48(1.63)
Min-m
ax14.00–25.27
37.52–38.77
4.62–6.92
0.09–0.87
0.02–3.81
1.59–7.31
Pellaro
x(s.d.)
18.16(4.44)
38.75(0.10)
5.29(0.61)
0.45(0.37)
0.89(0.91)
3.00(1.38)
Min-m
ax14.12–27.22
38.55–38.94
4.55–6.66
0.07–1.47
0.07–3.55
0.67–6.35
Stations
Chl-a
LAP
b-G
LU
AP
PA
(mgl�
1)
(nmolCleudm
�3h�1)
(nmolCgludm
�3h�1)
(nmolPO4dm
�3h�1)
(x106cellsml�
1)
Scaletta
x(s.d.)
0.107(0.05)
72.19(19.14)
12.53(3.15)
113.59(35.47)
4.7(6.3)
Min-m
ax0.02–0.33
0.006–935.21
0.005–173.31
0.007–1994.34
1.0–34.0
Guardia
x(s.d.)
0.075(0.03)
43.44(13.02)
9.71(1.83)
92.07(16.40)
6.3(1.4)
Min-m
ax0.01–0.15
0.002–874.83
0.03–87.05
0.02–700.05
0.1–94.0
Pellaro
x(s.d.)
0.128(0.09)
26.84(7.43)
9.30(1.77)
63.04(12.49)
4.8(7.8)
Min-m
ax0.04–0.49
0.003–484.15
0.0003–89.41
0.01–500.21
0.1–47.2
Abbreviations:temperature
(T),salinity(S),dissolved
oxygen
(DO),inorganicphosphate(PO4),am
monium
(NH4),nitriteþ
nitrate(N
O2, 3),chlorophyll-a
(Chl-a),LeucineAmino-
Peptidase(LAP),b-glucosidase(b-G
LU),AlkalinePhosphatase(A
P),prokaryoticabundance
(PA).
6 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

to the Sicilian coast and flowed through Scaletta (DS ¼ 0.5), going away from the Straits
system. Along the Calabrian side, Pellaro showed haline homogeneity (DS ¼ 0.3), being
located at the boundary of the horizontal shifts of water masses typical of Scaletta and not
affected by the vertical transport of water masses observed in the upwelling station
Guardia.
Annual average DO values oscillated between 5.25 and 5.34 ml l�1, detected in Sca-
letta and in Guardia, respectively (Table 1).
3.2. Nutrients distribution
Annual average PO4 concentration ranged from a maximum of 0.45 � 0.37 mM in Pellaro
to a minimum of 0.23 � 0.13 mM recorded in Scaletta, while in Guardia intermediate val-
ues were measured (Table 1). NH4 values ranged between a maximum of 0.92 � 0.97 mMmeasured in Scaletta and a minimum of 0.75 � 0.79 mM in Guardia, while NO2,3 concen-
tration ranged from a maximum of 3.55 � 1.63 mM recorded in Scaletta to a minimum of
3.00 � 1.38 mM recorded in Pellaro. The high variability of nutrient concentrations, as
shown by high standard deviation values, suggested the presence of an intermittent nutri-
ent source, referable to water upwelling. Vertical distribution of PO4 and NO2,3 (not
shown in Figure) showed that their concentration increased with depth in Pellaro, with
peak values (1.47 and 6.35 mM, respectively) measured at 100 m (below CM) in February
and July, respectively. Conversely, in Scaletta the highest values of PO4 (0.58 mM,
Figure 2. T/S diagram of the three stations (�, Scaletta,&, Guardia,~, Pellaro).
Advances in Oceanography and Limnology 7
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

found in February at 50 m) and NH4 (2.63 mM, found in June at 50 m) were recorded
below CM. In Guardia the highest NO2,3 value (3.67 mM, measured in June) was found
at the surface layer, confirming that the spatial distribution of peak values of nutrients
greatly reflected the different hydrological characteristics of the three stations.
3.3. Chlorophyll a
Chl-a concentration showed generally low values during the year, suggesting the presence
of moderate phytoplankton biomass. The highest average value was found in Pellaro
(0.128 � 0.09 mg m�3) and the lowest one in Guardia (0.075 � 0.03 mg m�3); intermedi-
ate values were recorded in Scaletta (0.107 � 0.05 mg m�3) (Table 1). The maximum of
Chl-a (CM) oscillated in a sub-surface layer (10–30 m) in both Scaletta and Guardia,
sinking to deeper layers (30–50 m) in Pellaro. The uplifting of the CM depth, similarly to
those of phosphacline and nitracline, was observed mainly in Guardia, proving the occur-
rence of the upwelling at this station, with consequent water column instability.
3.4. Microbial enzymatic activities and total prokaryotic abundance (PA)
Enzymatic activity values displayed evident spatial and temporal differences at each sta-
tion (Table 1); temporal variations were indicated by the high standard deviations. All the
enzymatic activities prevailed in Scaletta, where the annual average activity rates were
higher than those measured at the other stations. Intermediate rates of LAP, b-GLU and
AP were found in Guardia, whereas the lowest ones occurred in Pellaro. In Guardia and
Pellaro, similar levels of b-GLU were recorded.
The vertical distribution of LAP (Figure 3) showed high potential activity rates at
the surface (annual average values, 149.62 and 22.47 nmol Cleu dm�3h�1, in Scaletta
and Pellaro) and a general decrease with increasing depth. Activity levels sometimes in-
creased at intermediate depths, such as in Guardia (annual average LAP value: 13.77 nmol
Cleu dm�3h�1) or at the 100 m depth, as observed in Pellaro (annual average value:
41.94 nmol Cleu dm�3h�1) probably due to organic matter enrichment by the upwelled
waters. At all the stations, the peak of proteolytic activity was associated with the highest
PA.
As regards the CM layer (Figure 4), peaks of LAP activity were found at the surface
and above the CM in Scaletta, whereas they moved progressively above and below the
CM in Guardia and in Pellaro, respectively. At Pellaro, the detection of the maximum
LAP activity at deep layers indicated higher occurrence of labile organic matter with
increasing depth.
Of the three measured enzymes, b-GLU was recorded at the lowest levels in the
Messina Straits; maximum b-GLU activity was measured mainly at 10 m in Scaletta (on
average, 30.98 nmol Cglu dm�3h�1), while it was located at lower depths (30–50 m) in
Guardia and Pellaro (10.32 and 11.75 nmol Cglu dm�3h�1, respectively) (Figure 3). This
enzymatic activity prevailed at all the stations in the water mass above the CM (Figure 4),
suggesting a high amount of polysaccharides in this layer.
AP values were high at the surface in both Scaletta and Guardia (Figures 3 and 4, aver-
age AP value: 242.59 and 407.69 nmol PO4 dm�3h�1), and decreased along the water col-
umn, except for some peaks detected close to the 100 m depth (average value, 152.27 nmol
PO4 dm�3h�1 in Scaletta). Moreover, AP prevailed within the CM in Pellaro, where this
enzyme allowed a fast regeneration of P, therefore supporting the phytoplankton growth.
8 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013
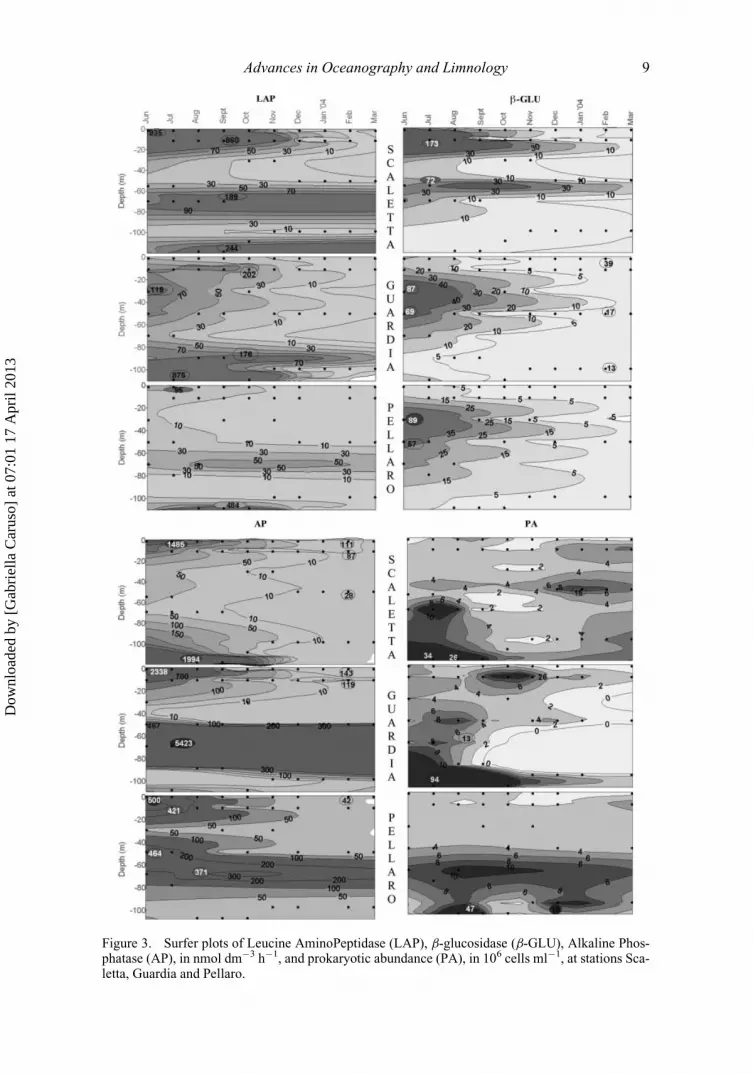
Figure 3. Surfer plots of Leucine AminoPeptidase (LAP), b-glucosidase (b-GLU), Alkaline Phos-phatase (AP), in nmol dm�3 h�1, and prokaryotic abundance (PA), in 106 cells ml�1, at stations Sca-letta, Guardia and Pellaro.
Advances in Oceanography and Limnology 9
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

Three-way ANOVA computed on overall data revealed no significant differences in enzy-
matic activity rates due to the factors ‘station’ or ‘depth’, while the factor ‘months’
accounted significantly for their variability (F ¼ 56.28, 39.44, 43.17, P < 0.01, for LAP,
b-GLU and AP, respectively).
PA was in the order of magnitude of 106 cells ml�1 and its spatial distribution was
quite homogeneous (Table 1). Conversely, seasonal differences were found, with the
highest densities detected in the deepest layer in summer (July to September), particularly
in Guardia (Figure 3).
3.5. Seasonal decomposition and production patterns
Seasonal patterns of the enzymatic activities and PA are reported in Table 2. At all the
studied stations, an abrupt decrease of hydrolysis rates from summer to autumn was ob-
served. Thereafter, LAP activity rates underwent a progressive decreasing trend towards
winter, whilst AP and b-GLU increased from autumn to winter, and their winter values
were about one magnitude order lower than in summer. In winter, AP contributed
84.10% of the total enzymatic activity in Scaletta at the surface (data not shown).
Seasonal distribution of the integrated and depth-normalised enzymatic activity
rates, Chl-a and PA values, obtained using the trapezoid method, confirmed that both
the decomposition and production processes were particularly enhanced in summer
(Figure 5). During this season, the decomposition process driven by microbial enzymes
was high everywhere, dropping to levels one order of magnitude lower in the successive
seasons. Also the phytoplankton production, as reflected by the Chl-a content, was at its
maximum in Pellaro (average value: 0.152 � 0.131 mg m�2); in autumn and winter its
intensity decreased. In winter, in Scaletta and Guardia a further increase was observed
in both autotrophic biomass and heterotrophic activities, with respect to AP and
b-GLU, leading to active phosphorus regeneration and polysaccharide decomposition,
respectively. The seasonal behaviour of PA matched quite well with that described by
the enzymatic activity rates, and the highest PA values were recorded in Guardia in
summer-autumn.
Figure 4. Average values of potential Leucine AminoPeptidase (LAP, &, b-glucosidase (b-GLU,& and Alkaline Phosphatase (AP, ) recorded at each station and per layer (surface: 0.5 m; abovethe Chlorophyll Maximum, CM: 10 m; within the CM: 30–50m; below the CM: 100 m). They areexpressed as nmol Cleu dm
�3 h�1, nmol Cglu dm�3 h�1, nmol PO4 dm
�3 h�1 respectively.
10 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

Table 2. Seasonal enzymatic activity and abundance values, obtained from overall data, reportedas average values � standard deviation. The number of samples analysed is reported in brackets.
Station Summer Autumn Winter
LAP (nmol Cleu dm�3h�1)(n ¼ 112)
Scaletta 199.5 ± 30.4 2.7 ± 2.4 0.7 ± 0.5Guardia 108.1 ± 21.3 3.3 ± 2.9 0.9 ± 0.8Pellaro 58.0 ± 12.1 2.6 ± 2.3 0.3 ± 0.2
b-GLU(nmol Cglu dm�3h�1)(n ¼ 110)
Scaletta 26.4 ± 4.6 1.3 ± 1.2 2.3 ± 3.4Guardia 18.5 ± 2.5 0.6 ± 0.4 6.0 ± 1.2Pellaro 18.3 ± 2.4 0.5 ± 0.4 3.4 ± 3.2
AP (nmol PO4 dm�3h�1)(n ¼ 113)
Scaletta 298.1 ± 59.9 2.9 ± 2.6 20.1 ± 3.8Guardia 183.3 ± 21.9 4.5 ± 0.7 41.8 ± 6.1Pellaro 137.0 ± 18.4 9.5 ± 1.1 14.2 ± 1.9
PA (x106 cells ml�1)(n ¼ 104)Scaletta 6.7 ± 1.0 1.9 ± 1.8 5.5 ± 4.0Guardia 10.1 ± 2.3 4.7 ± 0.7 1.9 ± 1.4Pellaro 6.7 ± 1.2 2.9 ± 2.7 3.1 ± 2.2
Abbreviations: Leucine AminoPeptidase (LAP), b-glucosidase (b-GLU), alkaline phosphatase (AP) andprokaryotic abundance (PA).
µ
µ
µ
Figure 5. Box plots of integrated and depth-normalised seasonal enzymatic activity rates (LAP,Leucine AminoPeptidase; b-GLU, b-glucosidase; AP, alkaline phosphatase), Chlorophyll-a (Chl-a)and Prokaryotic Abundance (PA) measured at Scaletta, Guardia and Pellaro stations. In each boxplot, the bold dash represents the median, the small dashes enclose the first and third quartiles, andthe vertical bars indicate the maximum and minimum of the data. Abbreviations: S, summer; A, au-tumn; W, winter.
Advances in Oceanography and Limnology 11
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

ANOVA computed per each station separately (Table 3) confirmed that seasonal LAP
variations in Scaletta were statistically significant (F ¼ 4.80, P < 0.01); a similar
result was obtained for b-GLU in Guardia and in Pellaro (F ¼ 4.15, P ¼ 0.02 and F ¼4.10, P ¼ 0.03, respectively). In Guardia and Pellaro AP values differed seasonally too
(F ¼ 5.82, P ¼ 0.01 and F ¼ 4.51, P ¼ 0.02, respectively). Conversely, seasonal differen-
ces detected in PA values never reached levels of statistical significance.
3.6. Statistical relationships between enzymatic activities and environmental
parameters
Pearson correlation coefficients, calculated at each station (Table 4), showed that enzy-
matic activity rates correlated differently at each station with the main hydrological
Table 3. Results of Analysis of variance (ANOVA) (F-test and P) performed on environmentaland microbial variables taking into account the factors ‘depth’ and ‘seasons’. Statistically signifi-cant differences at P < 0.01 are reported in bold, whilst at lower P in italics.
Among seasons Among depth
F P F P
SCALETTA LAP 4.80 0.01 1.06 n.s.b-GLU 2.77 n.s. 0.86 n.s.AP 2.71 n.s. 0.50 n.s.PA 1.79 n.s. 2.01 n.s.T 7.95 0.001 12.07 2.0�10�5
S 0.27 n.s. 0.65 n.s.DO 14.14 2.9�10�5 0.05 n.s.NH4 3.53 0.04 1.75 n.s.NO2.3 3.53 0.04 3.71 0.03PO4 3.70 0.03 0.77 n.s.Chl-a 2.31 n.s. 4.08 0.03
GUARDIA LAP 2.92 n.s. 0.36 n.s.b-GLU 4.15 0.022 0.56 n.s.AP 5.82 0.005 0.63 n.s.PA 0.75 n.s. 0.43 n.s.T 7.01 <0.002 21.89 0.0001S 0.16 n.s. 5.87 0.02DO 8.10 0.001 0.47 n.s.NH4 5.10 0.016 0.11 n.s.NO2.3 0.29 n.s. 0.02 n.s.PO4 1.70 n.s. 3.85 0.041Chl-a 1.62 n.s. 5.23 0.01
PELLARO LAP 2.13 n.s. 0.31 n.s.b-GLU 4.10 0.026 0.82 n.s.AP 4.51 0.018 0.79 n.s.PA 0.73 0.49 1.13 n.s.T 5.21 0.011 17.55 6.0�10�7
S 2.70 n.s. 4.48 0.01DO 6.06 0.006 0.69 n.s.NH4 4.49 0.023 0.90 n.s.NO2,3 0.93 n.s. 2.96 n.s.PO4 0.70 n.s. 0.83 n.s.Chl-a 0.69 n.s. 4.69 0.02
12 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

Table 4. Significant Pearson coefficients detected between all the data of physical, chemical andbiological parameters measured at Scaletta, Pellaro and Guardia stations.
Scaletta
T S DO Chl-a PO4 NH4 NO2,3 LAP b-GLU AP PA
TSDOChl-a 0.496��
PO4
NH4 0.554�� 0.595��
NO2,3
LAP 0.630�� �0.672��
b-GLU �0.411�� 0.531�� 0.723��
AP 0.841�� 0.718��
PA 0.684�� 0.811�� 0.388�
n 39 39 39 30 30 30 30 37 35 39 38
Guardia
T S DO Chl-a PO4 NH4 NO2,3 LAP b-GLU AP PA
TS �0.384�
DO �0.342�
Chl-a �0.480��
PO4
NH4 �0.358� 0.386�
NO2,3 0.585�� �0.710��
LAP �0.743��
b-GLU 0.625�� 0.649��
AP 0.611�� 0.493��
PA 0.908�� 0.908��
n 37 37 37 30 30 30 30 40 41 38 35
Pellaro
T S DO Chl-a PO4 NH4 NO2,3 LAP b-GLU AP PA
TSDO �0.349�
Chl-a �0.433��
PO4 �0.369�
NH4 �0.525�� 0.587��
NO2,3 �0.399� 0.342�
LAP �0.726��
b-GLU �0.455�� 0.596�� 0.531��
AP 0.565�� 0.549�� 0.718�� �0.627�� 0.310�
PA
n 36 36 36 27 27 27 27 35 34 36 31
Abbreviations: temperature (T), salinity (S), dissolved oxygen (DO), chlorophyll-a (Chl-a), phosphate (PO4),ammonia (NH4), nitrite þ nitrate (NO2,3), Leucine AminoPeptidase (LAP), b-glucosidase (b-GLU), AlkalinePhosphatase (AP), prokaryotic abundance (PA).n ¼ number of samples; level of statistical significance: �, P < 0.05; ��, P < 0.01.
Advances in Oceanography and Limnology 13
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

characteristics as well as with the parameters (nutrients and organic inputs) related to the
biochemistry of the water column. T affected positively AP in both Scaletta and Pellaro,
and LAP in Scaletta. S influenced inversely b-GLU in Scaletta and in Pellaro. An inverse
relationship between NO2,3 and LAP was observed at all the stations; this suggested that
the microbial decomposition of the protein substrates was inversely regulated by nitrogen
availability. Low PO4 concentration stimulated AP synthesis in Pellaro only. Trophic
inputs of organic polymers, rich in polysaccharides and produced by phytoplankton,
explained the positive relationships occurring between Chl-a and b-GLU in Scaletta and
in Guardia.
The relationships between PA and the enzymatic rates detected in Scaletta, indicated
the importance of prokaryotes in the polymers decomposition, mostly in the phosphoric
and glucidic compounds (Table 4). This community also played an important role in the
production of LAP in Guardia. The direct correlation between AP and Chl-a, found in
Scaletta as well as in Pellaro, suggested that the phytoplanktonic biomass accounted sig-
nificantly for the synthesis of this enzyme.
Concerning the separation into the different depths (above, within and below CM), the
contribution of the phytoplankton biomass to AP was more evident within the CM in Sca-
letta (AP versus Chl-a, all data, r ¼ 0.775, P < 0.01, n ¼ 10, data not shown). In Scaletta
and Pellaro, total prokaryotes accounted for AP synthesis, particularly below the CM (AP
versus PA, all data, r ¼ 0.558, P < 0.05, n ¼ 15; r ¼ 0.561, P < 0.05, n ¼ 13 respectively,
data not shown).
Pearson correlation coefficients computed on data grouped on a seasonal scale
(Table 5) generally reflected the behaviour shown by the annual statistical analysis
(Table 4), confirming the seasonal importance of environmental factors (T, nutrients,
Chl-a) in controlling enzymatic expression.
The reciprocal influence among the physical, chemical and biological parameters has
been analysed by PCA for each station separately, which has yielded the plots shown in
Figure 6. PCA outputs reveal that, globally, the two PCs explain as much as 38.3%,
46.6% and 45.7% of data variability at Scaletta, Guardia and Pellaro, respectively.
Table 5. Significant Pearson coefficients detected at each station between enzymatic data andphysical and chemical data, pooled on a seasonal basis.
Summer n Autumn n Winter n
Scaletta LAP-NO2,3(�0.59)� 12 AP-T(0.855��) 12 LAP-T(0.670�) 11LAP-NO2,3(�0.679�) 9 b-GLU-Chl-a(0.797��) 9b-GLU-PA(0.813��) 9 AP-Chl-a(0.760�) 9
Guardia LAP- NO2,3(�0.749��) 9LAP-PA(0.943��) 15 AP-PA(0.924��) 11 LAP-PA(0.587�) 12
Pellaro b-GLU-S(�0.645��) 16 AP-T(0.597�) 12LAP- NO2,3(�0.788��) 9AP-PO4(�0.662�) 9AP-Chl-a(0.704�) 9
Abbreviations: temperature (T), salinity (S), chlorophyll-a (Chl-a), phosphate (PO4), nitrite þ nitrate (NO2,3),Leucine AminoPeptidase (LAP), b-glucosidase (b-GLU), Alkaline Phosphatase (AP), prokaryotic abundance(PA).n ¼ number of samples; level of statistical significance: �, P < 0.05; ��, P < 0.01
14 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

Figure 6. Outputs of Principal Component Analysis performed at each station. S ¼ summer; A ¼autumn; W ¼ winter.
Advances in Oceanography and Limnology 15
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

In Scaletta, a great dispersion characterised the summer samples, in contrast with the
close grouping of the autumn-winter ones; PCA allowed discrimination between the upper
layers (surface and above CM) in summer-autumn and the deepest layers (within and be-
low CM) during autumn-winter. T has significantly negative weightings on
PC2, while PC1 is dominated by salinity (Sal.); there is a close association between PO4
and PA, b-GLU and DO, and between production (Chl-a) and decomposition (LAP)
processes.
In Guardia, the deepest samples, collected below CM, group together unless of their
sampling season, suggesting that the spatial arrangement of data obtained at this station is
unaffected by the variable month. Upper samples, collected in summer-autumn, group to-
gether too. Positive weightings are observed on PC1 for b-GLU, DO and AP; LAP and
PA were also in close association along this PC. On PC2, salinity and NO3 have a positive
influence on data dispersion, while they have inverse projections with Chl-a.
Like Guardia, in Pellaro, samples collected from deeper layers (below CM) are spa-
tially separated from those collected from the upper layers (surface and above CM) during
summer. On PC1, salinity has significant negative weightings, in opposite projection with
the positive weightings found for DO, nutrients and b-GLU. On PC2, T has a positive in-
fluence, and is projected in the same direction as AP.
Results of the Mann-Whitney test applied to the enzymatic activity dataset for mixed
and stratified regimes at each station separately are shown in Table 6. Stratification was
observed during five months in Scaletta and Guardia, while in Pellaro this hydrological
condition lasted longer (seven months). Although ANOVA performed per each month
showed no significant influence of depth on enzymatic activity, the Mann-Whitney test
revealed that water column stratification affected significantly all the enzymatic activity
rates, while the mixed water column condition did not. The most significant differences
were detected above the CM layer in Pellaro and Scaletta, while in Guardia at the surface
layer. During the mixed water condition, significant differences were found at Scaletta
only.
3.7. Biogeochemical implications
The reciprocal ratios computed between the three enzymes measured, as well as between
the amounts of nutrients potentially released from the microbial activity, are reported in
Table 7. In particular, the molar C/P ratio between the amounts of C potentially released
from LAPþb-GLU and the amounts of P potentially released from AP was calculated, as
well as the molar N/P ratio between N released from LAP and P released from AP. The
time required by microbes for the mobilisation of dissolved inorganic nitrogen (DIN ¼NH4þNO2,3) by LAP and of PO4 by AP was also computed.
The LAP/b-GLU ratio displayed the highest values during summer months at all the
stations, followed by those recorded in Pellaro during autumn; conversely, in winter the
values were lower, particularly in Scaletta. Low AP/LAP ratios (less than 5.2 � 0.7) were
observed during autumn, increasing in summer and in winter, when they reached 36.8 �5.3 in Pellaro. In summer the values of the DIN/LAP ratio were lower than those obtained
in autumn and winter (ranges: 62.8–116.1 and 170.7–293.8 d�1 respectively); a similar
result was observed for the summer values of the PO4/AP ratio, ranging from 0.8 in Guar-
dia to 1.4 d�1 in Scaletta. The seasonal distribution of the C/P and N/P molar ratios calcu-
lated for each station, showed that, for both the ratios, summer values were one order of
magnitude higher than those calculated in autumn, while minimum values occurred in
winter, thus reflecting the seasonal patterns of enzymatic activities. C/P ratios in Pellaro
16 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

Table 6. Results of the Mann-Whitney test on enzymatic data measured at different depths.
Stratified water column Mixed water column
Scaletta Scaletta
LAP SurfaceAboveCM
WithinCM
BelowCM Surface
AboveCM
WithinCM
BelowCM
Surface / /Above CM 2.1� / ns /Within CM ns 2.1� / ns ns /Below CM ns 4.0� ns / ns ns ns /b-GLUSurface / /Above CM 2.2� / ns /Within CM ns 2.1� / ns ns /Below CM ns 0.1� ns / ns 4.1� ns /APSurface / /Above CM 2.5� / ns /Within CM ns ns / ns ns /Below CM 6.0� 4.1� ns / ns ns ns /
Guardia Guardia
LAP SurfaceAboveCM
WithinCM
BelowCM Surface
AboveCM
WithinCM
BelowCM
Surface / /Above CM 2.2� / ns /Within CM 2.1� ns / ns ns /Below CM 2.1� ns ns / ns ns ns /b-GLUSurface / /Above CM 2.2� / ns /Within CM ns ns / ns ns /Below CM 6.1� ns ns / ns ns ns /APSurface / /Above CM ns / ns /Within CM ns ns / ns ns /Below CM 6.0� ns ns / ns ns ns /
Pellaro Pellaro
LAP SurfaceAboveCM
WithinCM
BelowCM Surface
AboveCM
WithinCM
BelowCM
Surface / /Above CM 6.0� / ns /Within CM ns 6.1� / ns ns /Below CM ns 6.0� ns / ns ns ns /b-GLUSurface / /Above CM 5.2� / ns /Within CM ns 6.0� / ns ns /Below CM ns 8.1� ns / ns ns ns /APSurface / /Above CM 6.1� / ns /Within CM ns 8.1� / ns ns /Below CM ns 7.0� ns / ns ns ns /
�, P < 0.05.ns = not significantLAP ¼ Leucine AminoPeptidase; b-GLU ¼ b-glucosidase; AP ¼ Alkaline Phosphatase; CM ¼ ChlorophyllMaximum.
Advances in Oceanography and Limnology 17
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

were mostly lower than those found in Scaletta and Guardia and displayed similar values
in autumn and in winter.
4. Discussion
4.1. Hydrological characteristics and relationships with enzymatic patterns
The first evident feature related to upwelling was the detection of higher average nutrient
values in the Straits compared to the adjacent Tyrrhenian and Ionian basins [42,43]. The
heterogeneous enzymatic activity patterns found in the Straits of Messina reflected the
complex hydrological characteristics existing within the study area. The circulation pat-
tern of the water masses indicated Scaletta as a point of exit of waters (MMW) from the
Straits flowing southward, thus leading us to hypothesise a possible concentration of or-
ganic inputs at this station. Upwelling filaments, such as those observed by Barbosa et al.
[44] along the NW Iberian margin, can promote horizontal transport of organic matter
produced during upwelling events. This consideration could justify the high rates of enzy-
matic activity recorded in Scaletta. According to their values, we hypothesise that at this
station the microbial community was able to metabolise quickly environmental inputs of
organic substrates, providing an important N, C and P source to the environment. Com-
pared to Scaletta, the upwelling Guardia exhibited lower enzymatic activity rates; this re-
duction in hydrolysis rates agreed with that found by Bianchi et al. [45] in near bottom
waters in the upwelling area off the Malaga coast. At Pellaro, located at the boundary of
the divergence area, the steady-state hydrological conditions favoured a higher phyto-
plankton development, compared to the other stations, while the availability of labile or-
ganic substrates due to the shallow depth was hypothesised to inhibit the polymer
hydrolysis process. Studies performed in other upwelling regions have displayed a close
relationship between primary production and bacterial metabolism [7–10]. In the Straits
of Messina, the link between photosynthetically produced organic inputs and the decom-
position process by microbial enzymatic activity was evident during summer (Figure 5),
confirming that heterotrophic microbes at the studied coastal stations were responsive to
carbon compounds released during phytoplankton bloom, as previously observed by
Table 7. Reciprocal ratios between Leucine AminoPeptidase (LAP), b-glucosidase (b-GLU) andAlkaline Phosphatase (AP), potential mobilisation ratios (PO4/AP and DIN/LAP) and molar rela-tionships (C/P and N/P) calculated as seasonal average value � standard deviation, per each station.Total C mobilised was calculated from the amounts of C released by LAPþb-GLU, total N and Pmobilised were estimated from the amounts of N and P released by LAP and AP, respectively.
summer autumn winter summer autumn winter
LAP/b-GLU AP/LAPScaletta 196.5 ± 34.4 4.1 ± 3.6 2.5 ± 0.3 6.61 ± 1.5 2.7 ± 0.4 29.7 ± 3.6Guardia 70.1 ± 1.8 7.8 ± 1.8 3.2 ± 0.5 15.7 ± 4.3 1.9 ± 0.4 32.9 ± 3.7Pellaro 54.7 ± 17.9 44.6 ± 20.7 3.4 ± 1.2 8.6 ± 1.1 5.2 ± 0.7 36.8 ± 5.3
DIN/LAP (d�1) PO4/AP (d�1)Scaletta 4.8 ± 4.5 90.4 ± 7.3 170.7 ± 7.3 1.4 ± 0.3 23.0 ± 2.9 27.2 ± 3.4Guardia 10.3 ± 1.3 116.1 ± 9.2 293.8 ± 3.7 0.8 ± 0.1 20.2 ± 2.7 31.4 ± 2.7Pellaro 21.3 ± 3.4 62.8 ± 4.5 232.8 ± 9.9 1.2 ± 0.2 9.3 ± 1.1 52.2 ± 4.9
C/P N/PScaletta 24.5 ± 0.7 10.9 ± 1.5 1.4 ± 0.2 25.6 ± 7.3 8.1 ± 1.1 1.1 ± 0.2Guardia 6.9 ± 0.7 2.7 ± 0.3 1.9 ± 0.4 6.8 ± 0.7 2.1 ± 0.3 0.3 ± 0.1Pellaro 17.3 ± 6.5 1.2 ± 1.1 1.1 ± 0.9 16.8 ± 6.5 0.9 ± 0.2 0.4 ± 0.1
18 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

Middelboe et al. [27]. On the other hand, the outputs of coupled density-enzymatic data-
set elaboration pointed out the presence of a stratified water column for at least one half
of the study period, which lasted a bit longer in correspondence of Pellaro station; this
favoured phytoplankton growth. During the whole time period, all the microbial degrada-
tion rates were sensibly affected by stratification, similarly to what was found at a shallow
NE Adriatic coastal site [34].
We hypothesise that the interference by different factors (i.e. circulation of water
masses, availability and quality of organic inputs, less or more prone to microbial degra-
dation) could explain the occurrence of microbially mediated biogeochemical processes
that were different among the stations studied. The uncoupling between organic inputs
and microbial utilisation, frequently observed in other coastal environments – where
allochthonous inputs play a greater role compared with autochthonous ones [46], – may
reflect the role played by heterotrophic bacterioplankton as a relatively more stable com-
ponent, compared to phytoplankton, in the food web of upwelling systems. Bacterio-
plankton dynamics may not follow changes in hydrodynamic conditions as strictly as
phytoplankton, whose dynamics closely depends on nutrient distribution. In addition, mi-
crobial metabolism is generally variable, depending on the physiological state of single
community components, while prokaryotic abundance is more homogeneous, so often
resulting in the uncoupling between activity and abundance patterns, as found by Zaccone
et al. [47]. In fact, substantial variations on short time scales (< 30 days) have been
detected in the enzymatic patterns of marine microbial communities in coastal southern
Californian waters, confirming that microbial metabolism is strongly affected by the pop-
ulation dynamics of enzyme-producing organisms [48].
Comparing the enzymatic activity rates measured in the Straits of Messina with esti-
mates reported from other marine environments (Table 8), LAP levels were found to be
similar to those described in other upwelling regions, but lower than those measured in
coastal waters. AP rates fell within a range by 1–2 orders of magnitude higher than that
reported for other upwelling regions and were similar to those recorded in eutrophic lakes.
AP was the most abundant enzyme among those measured, suggesting high potential P re-
generation in the study area. Vertical LAP profiles increased with increasing depth, in asso-
ciation with increased PA (see Figure 3); this finding indicated the presence of
metabolically active microorganisms in the waters close to the 100 m depth, particularly at
Pellaro.
To better compare the three stations according to their levels of microbial metabolism,
the enzymatic activity rates were divided by the PA measured in each sample, yielding the
vertical profiles of cell-specific enzymatic activity shown in Figure 7. Specific enzymatic ac-
tivity rates pointed out the predominance of AP compared to LAP and b-GLU in the whole
area. Scaletta was characterised by the highest cell-specific LAP, b-GLU and AP activities,
measured mainly in summer at surface depths. Within CM, discrete cell-specific activity
rates were detected for all the three enzymes too. In Guardia, both cell-specific AP and
b-GLU activities reached their maxima at surface in winter. At this same depth, high cell-
specific b-GLU activity rates were also found in autumn, suggesting that the prokaryotic me-
tabolism was sustained by the decomposition of polysaccharidic substrates. Conversely, the
highest cell-specific LAP activity rates were measured in summer above CM. High enzymat-
ic activity rates in the sub-surface layers have been reported as an evident sign of the
transfer of organic matter to depth, consequent to particle sinking processes [49]. In Pellaro,
cell-specific LAP activity followed a vertical profile similar to that found in Guardia, but
with a peak within CM; very low cell-specific AP and b-GLU activity rates were found.
Advances in Oceanography and Limnology 19
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

Table8.
Enzymaticactivityvalues
recorded
inthewater
columnofcoastalandupwellingareas.
Area
Tim
eperiod
LAPnmolh�1
b-G
LUnmolh�1
APnmolh�1
Reference
Coastalenvironments
Plubseelake
bloom
49.6–413.3
1.37–8.47
10
[60]
breakdown
422.8–541.9
48.9
Plubseelake
bloom
160
16
15–55(<
3mm)
[27]
breakdown
309
32
15–187(>
3mm)
AdriaticSeatrophic
gradient(0.5m)
pre-bloom
164
[61]
bloom
1712
postbloom
298.5
summer
133.7
NEAdriaticSea
annualstudy
9.7–72.4
1.8–9.2
4.1–37.6
[34]
NorthernAdriaticSea
42.92–138.47
[40]
NorthernAdriaticSea
0.1–39
0.1–107.5
[51]
Oliveri-Tindarilagoon(Sicily)
annualstudy
22.22–4358.33
0–76.39
110.32–414.84
[62]
SchleiFjord
176–3041
32.2–76.4
[63]
KielFjord
12.92–311.11
0.004–0.16
[64]
PomeranianBight(SouthernBalticSea)
12.4–704.4
0.8–12.6
5.7–270.0
[65]
MediterraneanSea
June
0.62–12.6
[55]
Sargasso
Sea
spring-summer
1.39–2.70
[66]
VillefrancheBay
(Mediterraneancoast)
23–33
[54]
Ligurian
Sea
springto
autumn(0–200m
integration)
1.78–4.08
0.04–0.19
[49]
Upwellingenvironments
northernIndianOcean
(ArabianSea)
June-July
5–30
1–2.5
2–5
[58]
NorthAfrican
upwelling(M
oroccocoast)
September
70
1.1�
0.2
[55]
CentralAtlanticOcean,includingNW
African
equatorialconvergence
May
andNovem
ber
4–50
[26]
NW
African
upwelling
August
<2.1
[59]
CentralChile(ItataRiver)
australspring-summer
1.2–182
0.08–61
[67]
StraitsofMessina
annualstudy
26.84–72.19
9.30–12.53
63.04–113.59
Thisstudy
20 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

4.2. Seasonal ecosystem functioning
Both enzymatic dynamics and environmental factors, responsible for the regulation of en-
zyme expression, underwent seasonal changes. Enzymatic activity rates displayed season-
al patterns at each station, with summer values one order of magnitude higher than those
Figure 7. Cell specific enzymatic activity rates (in fentongrams per cell per hour) calculated ateach station and per each water depth (surface, above, within and below Chlorophyll Maximum,CM). Rhombus, summer; square, autumn; circle, winter.
Advances in Oceanography and Limnology 21
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

recorded in other seasons and a sharp decrease in autumn. PA values showed that micro-
bial growth increased during summer months, when water masses become enriched with
monomers or dissolved compounds produced by hydrolysis. During autumn, LAP was in-
versely related to NO2,3 everywhere, while AP was affected positively by T in Scaletta
and Pellaro. In winter, enhanced b-GLU and AP rates were associated with high Chl-a
concentrations, particularly in Scaletta, suggesting that fresh organic substrates from phy-
toplankton production stimulated the microbial metabolism.
Positive relationships found between b-GLU and PA in Scaletta and between LAP
and PA in Guardia suggested that prokaryotes played an important role in organic poly-
mers decomposition; in turn, monomers released through the hydrolysis processes sup-
ported microbial growth. In both Scaletta and Pellaro, the phytoplanktonic component
had a significant role in the synthesis of AP, as suggested by the positive correlation be-
tween AP and Chl-a.
During summer, the decomposition processes driven by heterotrophic microorgan-
isms were particularly enhanced everywhere. In this season the amount of P potentially
remineralised by AP was estimated to range from 101.9 to 221.8 mg P dm�3 day�1, mea-
sured in Pellaro and Scaletta, respectively. At the same stations, the amount of dissolved
organic N potentially released by LAP varied from 19.5 to 67.0 mg dm�3 day�1, respec-
tively. Microbial decomposition predominated over the productive process still in au-
tumn. During this season, the microbial community replied to nutrient consumption
performed by autotrophs via a stimulated production of enzymes, which allowed a quick
nutrient recycling. Production and decomposition processes performed by autotrophic
biomass and heterotrophic activities were more reciprocally balanced in winter.
4.3. Biogeochemical implications
Considering the microbial enzymatic activities with implications on the Carbon cycle, in
the Straits of Messina LAP always prevailed over b-GLU, particularly during summer.
This indicated that fresh labile organic substrates such as proteins, both of autochthonous
and allochthonous origin, occurred in high amounts during this season; conversely, lower
LAP/b-GLU ratios recorded in winter suggested that organic substrates more refractory
than proteins were preferentially decomposed by microorganisms. High LAP/b-GLU val-
ues are typical of many temperate coastal environments, where a higher potential degra-
dation of proteins compared to that of polysaccharides is found [40,50,51]; on the other
hand, the greater amount of LAP compared to b-GLU may be explained by the capability
of released amino acids to provide a source of both C and N for bacteria [51]. Other than
Carbon, LAP is also involved in the Nitrogen cycle. High LAP hydrolysis rates recorded
in summer were also reflected by very low values of the DIN/LAP ratio, suggesting that
during the summer season the time required by LAP to mobilise the N pool was shorter
compared to other seasons.
Microorganisms contributed significantly to the Phosphorus cycle in the Straits
through AP, whose activity values exceeded those of LAP. P limitation has been widely
reported for the Mediterranean Sea [25,52–57], but in the Straits of Messina PO4 was al-
ways detected in discrete amounts (up to 0.64 mM in Pellaro in winter), suggesting that
no P limitation exists in this ecosystem. Therefore, the detection of an intense mineralisa-
tion activity of the organic phosphates in spite of the availability of P confirmed that in
the Straits of Messina AP could provide an important source of C, other than P
[23,58,59]. In other words, AP was probably regulated by C-limitation instead of
P-limitation. The high efficiency of microbial hydrolysis on organophosphoric
22 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

compounds in P recycling, particularly in summer, was confirmed by the very low
PO4/AP ratio values, underlining the importance of AP activity in the regeneration of the
P pool. In fact, the phytoplankton growth occurring in both Scaletta and Pellaro during
summer months, explained the high consumption and demand for this element.
Estimates of the molar ratios between the inorganic elements (C, N, P) released from
the combined action of LAP, b-GLU and AP, may provide further information on biogeo-
chemical implications of microbial-related activities taking place in the Straits of Messina
ecosystem. The molar C/P ratios indicated that C decomposition prevailed over P miner-
alisation everywhere, particularly in summer. In addition, the high molar N/P ratios found
in summer confirmed that during this season N supplied by the high N decomposition
exceeded the amounts of P released by AP. Conversely, the low molar N/P ratio values
estimated in winter, suggested a greater importance of P regeneration during this season.
The molar N/P ratios calculated from nutrient data followed a trend similar to that de-
scribed by enzymatic activity measurements, showing the highest values in summer
(55.05, 27.56, 17.60 in Scaletta, Guardia and Pellaro, respectively), that progressively de-
creased towards the normalisation (Redfield ratio N:P 16:1) in winter (16.73, 19.83, 9.82
at the same stations, respectively). These values, higher than the Redfield ratio, suggested
that our ecosystem was generally characterised by the excess of the DIN pool with respect
to PO4.
Overall, the ratios between the pool of inorganic nutrients (DIN and PO4) and the mi-
crobial activities involved in their release (LAP and AP) underlined the importance of the
microbial metabolism in the flux of nutrients from biotic to abiotic compartments. Sea-
sonal and spatial variations in these ratios reflected changes in the capability of the micro-
bial community to metabolise the available organic substrates.
5. Conclusion
This study focused on microbial enzymatic activity rates in organic matter processing and
biogeochemical cycles in the upwelling area of the Straits of Messina. Although the mea-
sured activity rates were only ‘potential’ – and consequently they could be overestimated
compared to ‘in situ’ rates – they pointed out significant differences in biogeochemical
processes and microbial metabolism among the stations and seasons.
The pronounced seasonality of hydrolysis rates, with higher values in summer at the
surface and above the CM, led us to presume that the increased availability of organic
polymers produced by phytoplankton was the main factor accounting for high enzymatic
levels. The relative contribution of the three enzymatic activities suggested that, among
the biogenic substrates occurring in water, proteins and organophosphoric compounds
were the most important in sustaining the microbial metabolism. In spite of P availability,
the high AP rates recorded particularly in summer, indicated the high demand for this ele-
ment, that was quickly regenerated, being potentially released in amounts sufficient to
sustain both bacteria and phytoplankton needs (from 101.9 to 221.8 mg P dm�3 day). In
autumn, activity patterns seemed to respond more closely to inorganic nutrient levels: as
an example, LAP was inversely related to NO2,3 concentration everywhere, suggesting
the synthesis of this enzyme was regulated by nitrogen availability. On the other hand,
the uncoupling between microbial activity patterns and PA, whose spatial distribution
was relatively homogeneous, supported the hypothesis that the variations noticed in the
activity rates could depend on changes in the specific community composition or in the
physiological status of cell components, rather than on variations in PA.
Advances in Oceanography and Limnology 23
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

Overall, this study confirmed that enzymatic activity dynamics respond to a lot of dif-
ferent environmental factors (temperature, salinity, current conditions, availability of la-
bile polymers and inorganic nutrients) that undergo reciprocal complex interactions. This
reason may explain the difficulties in identifying the major controls over enzymatic syn-
thesis. We suppose that productivity pulses driven by upwelling resulted in rapid changes
in the nature and composition of organic inputs transported by water masses, determining,
in turn, changes in microbial enzymatic patterns.
The different hydrodynamic characteristics of the three stations and the particular wa-
ter masses circulation both acted as important sources of variability for enzymatic pat-
terns; in fact, the high enzymatic rates detected in Scaletta could result from the organic
matter enrichment of coastal Sicilian waters which flow southward and exit from the
Straits system. Moreover, a different balance between production and decomposition
pathways characterised the stations examined: at Pellaro, more saline and nutrient-rich
waters showed low activity rates and high phytoplankton biomass values, demonstrating
that photosynthesis was the prevailing physiological process at the boundary of the up-
welling area. Conversely, the high potential enzymatic rates recorded along the Sicilian
side, mainly in Scaletta, indicated the occurrence of more intense mineralisation process-
es and a shift of the community’s metabolism towards heterotrophy.
Acknowledgements
The study was carried out within the MURST (Italian Ministry for University and Scientific Re-search) Cluster 10-SAM (Advanced Monitoring Systems) Project. The authors are grateful to A.Marini and M.G. Galletta (CNR- IAMC, Messina) for their assistance with nutrient analyses.
References
[1] B.C. Cho and F. Azam, Major role of bacteria in biogeochemical fluxes in the ocean’s interior,Nature 332 (1988), pp. 441–442.
[2] F. Azam, D.C. Smith, G.F. Steward, and A. Hagstrom, Bacteria-organic matter coupling andits significance for oceanic carbon cycling, Microb. Ecol. 28 (1993), pp. 167–179.
[3] F. Azam, Microbial control of oceanic carbon flux: The plot thickens, Science 280 (1998),pp. 694–696.
[4] R.J. Chrost, Microbial ectoenzymes in aquatic environments, in Aquatic Microbial Ecology:Biochemical and Molecular Approaches, J. Overbeck and R.J. Chrost, eds., Springer-Verlag,New York, 1990, pp. 47–78.
[5] H.G. Hoppe, C. Arnosti, and G. Herndl, Ecological significance of bacterial enzymes in themarine environment, in Enzymes in the Environment, Activity, Ecology and Applications, R.G. Burns and R.P. Dick, eds., Marcel Dekker, New York, 2002, pp. 73–107.
[6] A. Cunha, A. Almeida, F.J.R.C. Coelho, N.C.M. Gomes, V. Oliveira, and A.L. Santos, Bacte-rial extracellular enzymatic activity in globally changing aquatic ecosystems, in Current Re-search, Technology and Education Topics in Applied Microbiology and MicrobialBiotechnology, Microbiology Book Series no. 2, A. Mendez-Vila, ed., Formatex, Spain,2010, pp. 124–135.
[7] G.B. McManus and W.T. Peterson, Bacterioplankton production in the nearshore zone duringupwelling off central Chile, Mar. Ecol. Progr. Ser. 43 (1988), pp.11–17.
[8] C.J. Wiebinga, M.J.W. Veldhuis, and H.J.W. Baar, Abundance and productivity of bacterio-plankton in relation to seasonal upwelling in the northwest Indian Ocean, Deep-Sea Res. 44(1997), pp. 451–476.
[9] V.A. Troncoso, G. Daneri, L.A. Cuevas, B. Jacob, and P. Montero, Bacterial carbon flow inthe Humboldt Current System off Chile, Mar. Ecol. Progr. Ser. 250 (2003), pp.1–12.
[10] L.A. Cuevas, G. Daneri, B. Jacob, and P. Montero, Microbial abundance and activity in theseasonal upwelling area off Concepci�on ( 36�S), central Chile: A comparison of upwellingand non-upwelling conditions, Deep-Sea Res. 51 (2004), pp. 2427–2440.
24 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

[11] P. Montero, G. Daneri, L.A. Cuevas, H.E. Gonzalez, B. Jacob, L. Lizarraga, and E. Menschel,Productivity cycles in the coastal upwelling area off Concepci�on: The importance of diatomsand bacterioplankton in the organic carbon flux, Progr. Oceanogr. 75 (2007), pp. 518–530.
[12] E. De Domenico, Caratteristiche fisiche e chimiche delle acque nello Stretto di Messina:[Physical and chemical characteristics of the waters of the Straits of Messina], Documents etTravaux de l’ IGAL, Paris, 11 (1987), pp. 225–235.
[13] E. De Domenico, G. Cortese, and G. Pulican�o, Chemical characteristics of waters in the Straitsof Messina, in The Strait of Messina Ecosystem, L. Gugliemo, A. Manganaro, and E. DeDomenico, eds., Universita degli Studi, Messina, 1995, pp. 31–41.
[14] G. Magazz�u, M. Aubert, and F. Decembrini, The effect of tidal movements on planktonictransfer through the Straits of Messina, in The Strait of Messina Ecosystem, L. Gugliemo, A.Manganaro, and E. De Domenico, eds., Universita degli Studi, Messina, 1995, pp. 191–202.
[15] F. Azzaro, F. Decembrini, and E. Crisafi, Continuous survey of upwelling in the Straits ofMessina, Rapp. Comm. Int. Mer Medit. 34 (1995), pp. 167.
[16] B. Cescon, F. Azzaro, S. Creazzo, F. Decembrini, and G. Magazz�u, Processes affecting up-welling and primary productivity of the Straits of Messina, Boll. Geofisica Teor. Appl. 38(1997), pp. 137–147.
[17] F. Decembrini, F. Azzaro, E. Paschini, and G. Magazz�u, Influenza dei fenomeni di upwellingsulla produzione e biomassa fitoplanctoniche nello Stretto di Messina [Influence of upwellingphenomena on the phytoplankton production and biomass in the Straits of Messina], in Atti12� Congresso Associazione Italiana di Oceanologia e Limnologia, M. Picazzo, ed., Associa-zione Italiana di Oceanologia e Limnologia, Genova, 1998, pp. 159–171.
[18] F. Decembrini, F. Azzaro, and M. Leonardi, Variabilita a breve e lungo termine delle caratter-istiche idrologiche nelle acque costiere di un sistema ad elevato idrodinamismo (Stretto diMessina [Short- and long-term variability of the hydrological characteristics in the coastalwaters of a system under high hydrodynamism (Straits of Messina)], in Atti 13� CongressoAssociazione Italiana di Oceanologia e Limnologia, M. Picazzo, ed., Associazione Italiana diOceanologia e Limnologia, Genova, 1999, pp. 151–158.
[19] F. Azzaro, F. Decembrini, F. Raffa, and E. Crisafi, Seasonal variability of phytoplankton fluo-rescence in relation to the Straits of Messina (Sicily) tidal upwelling, Ocean Sci. 3 (2007),pp. 451–460.
[20] S. Genovese, G. Cortese, E. Crisafi, E. De Domenico, M. De Domenico, L. Genovese, R. LaFerla, and G. Pulican�o, Caratterizzazione delle popolazioni microbiche in acque ioniche e tir-reniche [Characterization of the microbial populations in the Ionian and Tyrrhenian waters],Nova Thalassia, 6 (1984), pp. 425–432.
[21] M. De Domenico, G. Cortese, G. Pulican�o, and E. De Domenico, Variazione spazio-temporaledi nutrienti, clorofilla e carica microbica nelle acque dello Stretto di Messina [Spatial and tem-poral variation of nutrients, chlorophyll and microbial density in the waters of the Straits ofMessina], in Atti 8� Congresso Associazione Italiana di Oceanologia e Limnologia, M. Pic-azzo, ed., Associazione Italiana di Oceanologia e Limnologia, Genova, 1988, pp. 337–355.
[22] M. De Domenico, Distribution of microbial heterotrophic communities in the Straits of Mes-sina, in The Strait of Messina Ecosystem, L. Gugliemo, A. Manganaro, and E. De Domenico,eds., Universita degli Studi, Messina, 1995, pp. 145–154.
[23] H.G. Hoppe, Phosphatase activity in the sea, Hydrobiologia 493 (2003), pp. 187–200.[24] M. Nausch, Alkaline phosphatase activities and the relationship to inorganic phosphate in the
Pomeranian Bight (southern Baltic Sea), Aquat. Microb. Ecol. 16 (1993), pp. 87–94.[25] T. Zohary and R.D. Robarts, Experimental study of microbial P limitation in the eastern Medi-
terranean, Limnol. Oceanogr. 43 (1998), pp. 387–395.[26] M. Vidal, C.M. Duarte, S. Agust�ı, J.M. Gasol, and D. Vaque, Alkaline phosphatase activities
in the central Atlantic Ocean indicate large areas with phosphorus deficiency, Mar. Ecol.Progr. Ser. 262 (2003), pp. 43–53.
[27] M. Middelboe, M. Sondergaard, Y. Letarte, and N.H. Borch, Attached and free-living bacte-ria: Production and polymer hydrolysis during a diatom bloom, Microb. Ecol. 29 (1995), pp.231–248.
[28] G.T. Taylor, J. Way, Y. Yu, and M.I. Scranton, Ectohydrolase activity in surface waters of theHudson River and western Long Island Sound estuaries, Mar. Ecol. Progr. Ser. 263 (2003),pp. 1–15.
Advances in Oceanography and Limnology 25
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

[29] M.M. Sala, M. Karner, L. Arin, and C. Marrase, Measurement of ectoenzyme activities as anindication of inorganic nutrient imbalance in microbial communities, Aquat. Microb. Ecol. 23(2001), pp. 301–311.
[30] G. Grancini and G. Magazz�u, A note on the oceanographic property of the Eastern Sicilycoast, Mem. Biol. Mar. Oceanogr. 3 (1973), pp. 27–49.
[31] T.S. Hopkins, E. Salusti, and D. Settimi, Tidal forcing of the water mass interface in the Straitof Messina, J. Geophys. Res. 89 (1984), pp. 2013–2024.
[32] E. Bohm, G. Magazz�u, L. Wald, and M.L. Zoccolotti, Coastal currents on the Sicilian shelfsouth of Messina, Oceanologica Acta 10 (1987), pp. 137–142.
[33] A. Adragna and E. Salusti, Observation of small scale shelf-trapped dipolar vortices near theEastern Sicilian coast, J. Phys. Oceanogr. 20 (1990), pp. 1105–1112.
[34] M. Celussi and P. Del Negro, Microbial degradation at a shallow coastal site: Long-term spec-tra and rates of exoenzymatic activities in the NE Adriatic Sea, Estuar. Coast. Shelf Sci.(2012), doi: 10.1016/j.ecss.2012.02.002.
[35] A. Aminot and M. Chaussepied, Manuel des analyses chimiques en milieu marin [Manual ofchemical analyses for the marine environment], Centre National pour l’exploitation desOceans, CNEXO, Brest, 1983.
[36] J.F.H. Strickland and T.R. Parsons, A practical handbook of seawater analysis, Bull. Fish. Res.Bd. Canada 167 (1972), pp. 1–311.
[37] L. Lazzara, F. Bianchi, M. Falcucci, V. Hull, M. Modigh, and M. Ribera D’Alcala, Pigmenticlorofilliani [Chlorophyll Pigments], Nova Thalassia 11 (1990), pp. 207–223.
[38] H.G. Hoppe, Use of fluorogenic model substrates for extracellular enzyme activity (EEA)measurement of bacteria, in Current Methods in Aquatic Microbial Ecology, P.F. Kemp., B.F.Sherr., E.B. Sherr, and J.J. Cole, eds., CRC Press, Boca Raton, 1993, pp. 423–431.
[39] R. La Ferla, R. Zaccone, G. Caruso, and M. Azzaro, Enzyme activities and carbon fluxthrough the microbial compartment in the Adriatic Sea, in Structure and Processes in the Med-iterranean Ecosystems, F.M. Faranda, L. Guglielmo, and G. Spezie, eds., Springer-Verlag Ita-lia, Milano, 2001, pp. 485–493.
[40] G. Caruso, Leucine Aminopeptidase, b-Glucosidase and Alkaline Phosphatase activity ratesand their significance in nutrient cycles in some coastal Mediterranean sites, Mar. Drugs 8(2010), pp. 916–940.
[41] K.G. Porter and Y.S. Feig, The use of DAPI for identifying and counting aquatic microflora,Limnol. Oceanogr. 25 (1980), pp. 943–948.
[42] F. Azzaro, F. Decembrini, F. Raffa, and S. Greco, Caratteristiche idrobiologiche dello stratoeufotico nel Tirreno Sud Orientale [Hydrobiological characteristics of the euphotic layer inthe South-Eastern Tyrrhenian Sea], Biol. Mar. Medit. 10 (2003), pp. 631–635.
[43] F. Decembrini, T.S. Hopkins, and F. Azzaro, Variability and sustenance of the deep-chloro-phyll maximum over a narrow shelf, Augusta Gulf (Sicily), Chem. Ecol. 20 (2004), pp. 231–247.
[44] A.B. Barbosa, H.M. Galv~ao, P.A. Mendes, X.A. Alvarez-Salgado, F.G. Figueiras, and I. Joint,Short-term variability of heterotrophic bacterioplankton during upwelling off the NW Iberianmargin, Progr. Oceanogr. 51 (2001), pp. 339–359.
[45] A. Bianchi, A. Calafat, R. De Wit, J. Garcin, O. Tholosan, I. Cacho, M. Canals, J. Fabres, H.Grout, P. Masque, J-A. Sanchez-Cabeza, and R. Sempere, Microbial activity at the deep watersediment boundary layer in two highly productive systems in the Western Mediterranean: TheAlmeria-Oran front and the Malaga upwelling, Oceanologica Acta 25 (2003), pp. 315–324.
[46] J.M. Gasol, M.D. Doval, J. Pinhassi, J.I. Calder�on- Paz, N. Guixa-Boixareu, D. Vaque, and C.Pedr�os-Ali�o, Diel variations in the bacterial heterotrophic activity and growth in the north-western Mediterranean Sea, Mar. Ecol. Progr. Ser. 164 (1998), pp. 107–124.
[47] R. Zaccone, A. Boldrin, G. Caruso, R. La Ferla, G. Maimone, C. Santinelli, and M. Turchetto,Enzymatic activities and prokaryotic abundance in relation to organic matter along a west–east Mediterranean transect (TRANSMED Cruise), Microb. Ecol. 64 (2012), pp. 54–66.
[48] S.D. Allison, Y. Chao, J.D. Farrara, S. Hatosy, and A.C. Martiny, Fine-scale temporal varia-tion in marine extracellular enzymes of coastal southern California, Front. Microbiol. 3(2012), pp. 1–10.
[49] C. Misic and M. Fabiano, Ectoenzymatic activity and its relationship to chlorophyll-a and bac-teria in the Gulf of Genoa (Ligurian Sea, NW Mediterranean), J. Mar. Syst. 60 (2006),pp. 193–206.
26 G. Caruso et al.
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013

[50] G. Caruso and R. Zaccone, Estimates of leucine-aminopeptidase activity in different marineand brackish environments, J. Appl. Microbiol. 89 (2000), pp. 951–959.
[51] R. Zaccone and G. Caruso, Microbial hydrolysis of polysaccharides and organic phosphates inthe northern Adriatic Sea, Chem. Ecol. 18 (2002), pp. 85–94.
[52] D.J. Bonin, M.C. Bonin, and T. Berman, Mise en evidence experimentale des facteurs nutritifslimitants de la production du micro-nanoplancton et de l’ultraplancton dans une eau coti�ere dela Mediterranee orientale (Ha€ıfa, Isra€el) [Experimental detection of nutrients limiting micro-,nano- and ultra-plankton production in a coastal water of the Eastern Mediterranean Sea(Ha€ıfa, Isra€el)], Aquat. Sci. 51 (1989), pp. 129–152.
[53] M.D. Krom, N. Kress, S. Brenner, and L.I. Gordon, Phosphorus limitation of primary produc-tivity in the eastern Mediterranean Sea, Limnol. Oceanogr. 36 (1991), pp. 424–432.
[54] T.F. Thingstad, U.L. Zweifel, and F. Rassoulzadegan, P limitation of heterotrophic bacteriaand phytoplankton in the northwestern Mediterranean, Limnol. Oceanogr. 43 (1998), pp. 88–94.
[55] F. Van Wambeke, U. Christaki, A. Giannakourou, T. Moutin, and K. Souvemerzoglou, Longi-tudinal and vertical trends of bacterial limitation by phosphorus and carbon in the Mediterra-nean Sea, Microb. Ecol. 43 (2002), pp. 119–133.
[56] M.D. Krom, B. Herut, and R.F.C. Mantoura, Nutrient budget for the Eastern Mediterranean:Implications for P limitation, Limnol. Oceanogr. 49 (2004), pp.1582–1592.
[57] T.F. Thingstad, M.D. Krom, R.F.C. Mantoura, G.A.F. Flaten, S. Groom, B. Herut, N. Kress,C.S. Law, A. Pasternak, P. Pitta, S. Psarra, F. Rassoulzadegan, T. Tanaka, A. Tselepides, P.Wassmann, E.M.S. Woodward, C. Wexels Riser, G. Zodiatis, and T. Zohary, Nature of Phos-phorus limitation in the ultraoligotrophic Eastern Mediterranean, Science 309 (2005),pp. 1068–1071.
[58] H.G. Hoppe and S. Ullrich, Profiles of ectoenzymes in the Indian Ocean: Phenomena of phos-phatase activity in the mesopelagic zone, Aquat. Microb. Ecol. 19 (1999), pp. 139–148.
[59] M. Sebastian, J. Aristegui, M.F. Montero, and F.X. Niell, Kinetics of alkaline phosphatase ac-tivity, and effect of phosphate enrichment: A case study in the NW African upwelling region,Mar. Ecol. Progr. Ser. 270 (2004), pp. 1–13.
[60] R.J. Chrost, U. Munster, H. Rai, D. Albrecht, P.K. Witzel, and J. Overbeck, Photosyntheticproduction and exoenzymatic degradation of organic matter in the euphotic zone of a eutro-phic lake, J. Plankton Res. 11 (1989), pp. 223–242.
[61] M. Karner, D. Fuks, and G. Herndl, Bacterial activity along a trophic gradient, Microb. Ecol.24, (1992), pp. 243–257.
[62] G. Caruso, L. Monticelli, F. Azzaro, M. Azzaro, F. Decembrini, R. La Ferla, M. Leonardi, andR. Zaccone, Dynamics of extracellular enzymatic activities in a shallow Mediterranean eco-system (Tindari ponds, Sicily), Mar. Freshwater Res. 56 (2005), pp. 173–188.
[63] H.G. Hoppe, H.C. Giesenhagen, and K. Gocke, Changing patterns of bacterial substrate de-composition in a eutrophication gradient, Aquat. Microb. Ecol. 15 (1998), pp. 1–13.
[64] H.G. Hoppe, Relations between bacterial extracellular enzyme activities and heterotrophicsubstrate uptake in a brackish water environment, 2�eme GERBAM – CNRS Colloque Interna-tional de Bacteriologie marine, Ifremer, Plouzane, France, 1986, pp. 119–128.
[65] M. Nausch, F. Pollehne, and E. Kerstan, Extracellular enzyme activities in relation to hydro-dynamics in the Pomeranian Bight (Southern Baltic Sea), Microb. Ecol. 36 (1998), pp. 251–258.
[66] J.B. Cotner, J.W. Ammerman, E.R. Peele, and E. Bentzen, Nutrient-limited bacterioplanktongrowth in the Sargasso Sea, Aquat. Microb. Ecol. 13 (1997), pp. 141–149.
[67] S. Pantoja, M.H. Gutierrez, P. Ampuero, and E. Tejos, Degradation capability of the coastalenvironment adjacent to the Itata River in central Chile (36.5�S), Biogeosci. 8 (2011),pp. 2063–2074.
Advances in Oceanography and Limnology 27
Dow
nloa
ded
by [
Gab
riel
la C
arus
o] a
t 07:
01 1
7 A
pril
2013
Copyright © 2022 FDOKUMEN




















