
Hypermethylation of the GABRE miR-452 miR-224 Promoter in Prostate Cancer Predicts Biochemical...
14
Imaging, Diagnosis, Prognosis Hypermethylation of the GABREmiR-452miR-224 Promoter in Prostate Cancer Predicts Biochemical Recurrence after Radical Prostatectomy Helle Kristensen 1 , Christa Haldrup 1 , Siri Strand 1 , Kamilla Mundbjerg 1 , Martin M. Mortensen 1,2 , Kasper Thorsen 1 , Marie Stampe Ostenfeld 1 , Peter J. Wild 4 , Christian Arsov 5 , Wolfgang Goering 5 , Tapio Visakorpi 6 , Lars Egevad 7 , Johan Lindberg 8 , Henrik Gr€ onberg 8 , Søren Høyer 3 , Michael Borre 2 , Torben F. rntoft 1 , and Karina D. Sørensen 1 Abstract Purpose: Available tools for prostate cancer diagnosis and prognosis are suboptimal and novel biomarkers are urgently needed. Here, we investigated the regulation and biomarker potential of the GABREmiR-452miR-224 genomic locus. Experimental Design: GABRE/miR-452/miR-224 transcriptional expression was quantified in 80 nonmalignant and 281 prostate cancer tissue samples. GABREmiR-452miR-224 promoter methyl- ation was determined by methylation-specific qPCR (MethyLight) in 35 nonmalignant, 293 prostate cancer [radical prostatectomy (RP) cohort 1] and 198 prostate cancer tissue samples (RP cohort 2). Diagnostic/prognostic biomarker potential of GABREmiR-452miR-224 methylation was evaluated by ROC, Kaplan–Meier, uni- and multivariate Cox regression analyses. Functional roles of miR-224 and miR-452 were investigated in PC3 and DU145 cells by viability, migration, and invasion assays and gene-set enrichment analysis (GSEA) of posttransfection transcriptional profiling data. Results: GABREmiR-452miR-224 was significantly downregulated in prostate cancer compared with nonmalignant prostate tissue and had highly cancer-specific aberrant promoter hypermethylation (AUC ¼ 0.98). Functional studies and GSEA suggested that miR-224 and miR-452 inhibit proliferation, migration, and invasion of PC3 and DU145 cells by direct/indirect regulation of pathways related to the cell cycle and cellular adhesion and motility. Finally, in uni- and multivariate analyses, high GABREmiR-452miR-224 promoter methylation was significantly associated with biochemical recurrence in RP cohort 1, which was successfully validated in RP cohort 2. Conclusion: The GABREmiR-452miR-224 locus is downregulated and hypermethylated in prostate cancer and is a new promising epigenetic candidate biomarker for prostate cancer diagnosis and prognosis. Tumor-suppressive functions of the intronic miR-224 and miR-452 were demonstrated in two prostate cancer cell lines, suggesting that epigenetic silencing of GABREmiR-452miR-224 may be selected for in prostate cancer. Clin Cancer Res; 20(8); 2169–81. Ó2014 AACR. Introduction Prostate cancer is a common malignancy among males in Western countries. Despite its widespread use, prostate- specific antigen (PSA) testing has suboptimal specificity and sensitivity for prostate cancer detection. Another major challenge in prostate cancer management is to distinguish between indolent and aggressive tumors due to the limited accuracy of currently available prognostic indicators (main- ly PSA, TNM stage, and Gleason score). Thus, novel bio- markers for prostate cancer are needed to improve the accuracy of diagnosis and prognosis (1). Molecular biomar- kers with important biologic functions in prostate cancer could also have potential as therapeutic targets. In this study, we investigated the regulation, function, and biomarker potential for prostate cancer of a genomic Authors' Affiliations: Departments of 1 Molecular Medicine and 2 Urology and 3 Institute of Pathology, Aarhus University Hospital, Aarhus, Denmark; 4 Institute of Surgical Pathology, University Hospital Zurich, Zurich, Swit- zerland; 5 Department of Urology, Medical Faculty, Heinrich Heine Uni- versity, D€ usseldorf, Germany; 6 Institute of Biomedical Technology and BioMediTech, University of Tampere and Tampere University Hospital, Tampere, Finland; Departments of 7 Oncology and Pathology and 8 Med- ical Epidemiology and Biostatistics, Karolinska Institute, Stockholm, Sweden Note: Supplementary data for this article are available at Clinical Cancer Research Online (http://clincancerres.aacrjournals.org/). Corresponding Author: Karina Dalsgaard Sørensen, Department of Molecular Medicine, Aarhus University Hospital, Brendstrupgaardsvej 100, DK-8200 Aarhus N, Aarhus, Denmark. Phone: (+45) 78455316; Fax: (+45) 86782108; E-mail: [email protected] doi: 10.1158/1078-0432.CCR-13-2642 Ó2014 American Association for Cancer Research. Clinical Cancer Research www.aacrjournals.org 2169 on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Hypermethylation of the GABRE miR-452 miR-224 Promoter in Prostate Cancer Predicts Biochemical...

Imaging, Diagnosis, Prognosis
Hypermethylation of the GABRE�miR-452�miR-224Promoter in Prostate Cancer Predicts BiochemicalRecurrence after Radical Prostatectomy
Helle Kristensen1, Christa Haldrup1, Siri Strand1, Kamilla Mundbjerg1, Martin M. Mortensen1,2,Kasper Thorsen1, Marie Stampe Ostenfeld1, Peter J. Wild4, Christian Arsov5, Wolfgang Goering5,Tapio Visakorpi6, Lars Egevad7, Johan Lindberg8, Henrik Gr€onberg8, Søren Høyer3, Michael Borre2,Torben F. �rntoft1, and Karina D. Sørensen1
AbstractPurpose: Available tools for prostate cancer diagnosis and prognosis are suboptimal and novel
biomarkers are urgently needed. Here, we investigated the regulation and biomarker potential of the
GABRE�miR-452�miR-224 genomic locus.
Experimental Design: GABRE/miR-452/miR-224 transcriptional expression was quantified in 80
nonmalignant and 281 prostate cancer tissue samples. GABRE�miR-452�miR-224 promoter methyl-
ation was determined by methylation-specific qPCR (MethyLight) in 35 nonmalignant, 293 prostate
cancer [radical prostatectomy (RP) cohort 1] and 198 prostate cancer tissue samples (RP cohort 2).
Diagnostic/prognostic biomarker potential of GABRE�miR-452�miR-224methylation was evaluated by
ROC, Kaplan–Meier, uni- and multivariate Cox regression analyses. Functional roles of miR-224 and
miR-452 were investigated in PC3 and DU145 cells by viability, migration, and invasion assays and
gene-set enrichment analysis (GSEA) of posttransfection transcriptional profiling data.
Results: GABRE�miR-452�miR-224 was significantly downregulated in prostate cancer compared with
nonmalignant prostate tissue and had highly cancer-specific aberrant promoter hypermethylation (AUC¼0.98). Functional studies and GSEA suggested that miR-224 and miR-452 inhibit proliferation, migration,
and invasion of PC3 and DU145 cells by direct/indirect regulation of pathways related to the cell cycle and
cellular adhesion and motility. Finally, in uni- and multivariate analyses, high GABRE�miR-452�miR-224
promoter methylation was significantly associated with biochemical recurrence in RP cohort 1, which was
successfully validated in RP cohort 2.
Conclusion: The GABRE�miR-452�miR-224 locus is downregulated and hypermethylated in prostate
cancer and is a new promising epigenetic candidate biomarker for prostate cancer diagnosis and prognosis.
Tumor-suppressive functions of the intronic miR-224 and miR-452 were demonstrated in two prostate
cancer cell lines, suggesting that epigenetic silencing of GABRE�miR-452�miR-224 may be selected for in
prostate cancer. Clin Cancer Res; 20(8); 2169–81. �2014 AACR.
IntroductionProstate cancer is a commonmalignancy amongmales in
Western countries. Despite its widespread use, prostate-specific antigen (PSA) testing has suboptimal specificityand sensitivity for prostate cancer detection. Another majorchallenge in prostate cancer management is to distinguishbetween indolent and aggressive tumors due to the limitedaccuracy of currently available prognostic indicators (main-ly PSA, TNM stage, and Gleason score). Thus, novel bio-markers for prostate cancer are needed to improve theaccuracy of diagnosis and prognosis (1).Molecular biomar-kers with important biologic functions in prostate cancercould also have potential as therapeutic targets.
In this study, we investigated the regulation, function,and biomarker potential for prostate cancer of a genomic
Authors' Affiliations: Departments of 1Molecular Medicine and 2Urologyand 3Institute of Pathology, AarhusUniversity Hospital, Aarhus, Denmark;4Institute of Surgical Pathology, University Hospital Zurich, Zurich, Swit-zerland; 5Department of Urology, Medical Faculty, Heinrich Heine Uni-versity, D€usseldorf, Germany; 6Institute of Biomedical Technology andBioMediTech, University of Tampere and Tampere University Hospital,Tampere, Finland; Departments of 7Oncology and Pathology and 8Med-ical Epidemiology and Biostatistics, Karolinska Institute, Stockholm,Sweden
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
Corresponding Author: Karina Dalsgaard Sørensen, Department ofMolecular Medicine, Aarhus University Hospital, Brendstrupgaardsvej100, DK-8200 Aarhus N, Aarhus, Denmark. Phone: (+45) 78455316;Fax: (+45) 86782108; E-mail: [email protected]
doi: 10.1158/1078-0432.CCR-13-2642
�2014 American Association for Cancer Research.
ClinicalCancer
Research
www.aacrjournals.org 2169
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

locus at Xq28, encompassing the GABRE gene and twointronic microRNAs (miR-224 and miR-452). GABREencodes the epsilon subunit of gamma-aminobutyric acid(GABA) A receptor (2) and has a promoter-associated CpGisland (CGI). Hypermethylation of promoter CGIs is closelylinked with gene silencing in cancer, and aberrant promoterhypermethylation of specific genes, such as GSTP1 andPITX2, has shown promising diagnostic and prognosticbiomarker potential for prostate cancer (3–7). Here, wefocused on the GABRE�miR-452�miR-224 locus, becauseintegrative analysis of mRNA and miRNA expression andDNA methylation profiling data available in-house indicat-ed significant downregulation accompanied by aberrantpromoter hypermethylation in prostate cancer. Moreover,results from several studies (see below) suggestedpotentiallyimportant functional role(s) for this locus in prostate cancer.
Deregulation ofGABRE expression in prostate cancer hasnot been reported before this work. The GABAA receptor is aheteropentameric chloride channel that transmits the fast-est inhibitory signal of the central nervous system (CNS)with its ligand GABA (8). The GABAA receptor family con-sists of 19homologous genes, encoding subunitsa1-a6,b1-b3, g1-g3, d, r1-r3, q, p, and e (GABRE). The dominantreceptor subtype is believed to be 2a:2b:1g . GABRE resem-bles the g-subunit andmay replace this to form a functionalreceptor (2, 9). Outside the CNS, elevated GABA levels havebeen associated with prostate cancer metastasis (10) andGABA agonist isoguvacine has been shown to stimulateproliferation of four prostate cancer cell lines (11). Incontrast, GABA signaling has been found to inhibit breastcancer cell migration (12), suggesting a highly context-dependent function.
MicroRNAs (miRNA) are endogenous small noncodingRNAs (�22 nt) that regulate gene expression at the post-transcriptional level, typically by binding to sequence-spe-cific sites in the 30UTRof targetmRNAs to repress translationand/or initiate mRNA decay (13). A single miRNA mayfunction either as a tumor suppressor or an oncogene,depending on the cellular context and its specific mRNAtarget(s) (13). miRNAs have shown promising potential ascandidate cancer biomarkers, therapeutic targets, and anti-cancer drugs (13). Downregulation of miR-224 in prostatecancer tissue samples has been reported in two recentstudies (14, 15), but another study reported upregulationof miR-224 in perineural invasion (PNI) positive versusnegative prostate cancer (16). miR-452 expression in pros-tate cancer is also poorly characterized, although a study hassuggested that miR-452 is overexpressed in prostate cancerprogenitor cells (17). Downregulation ofmiR-224 has beenreported in ovarian, breast, and lung cancer (18–20), whileupregulation has been reported in medullablastoma, thy-roid, liver, renal, and colorectal cancer (21–25), suggestingcell type-specific expression and function. Although miR-224 has been linked to proliferation, migration, and inva-sion in several cancers (21, 22), its function remains to beinvestigated in prostate cancer. miR-452 has been found tobe upregulated in medulloblastoma (21) and in urothelialcarcinomas with lymph node metastasis, suggesting a pos-sible role in progression of these malignancies (26); how-ever, its potential function in prostate cancer has not yetbeen studied.
Here, we found that the entire GABRE�miR-452�miR-224 locus was downregulated in prostate cancer comparedwith nonmalignant prostate tissue samples, and that silenc-ing was associated with frequent aberrant promoter hyper-methylation. We also found miR-224 and miR-452 toinhibit viability, migration, and invasion of two prostatecancer cell lines, suggesting that they hold tumor suppressorfunctions in prostatic cells. Finally, high GABRE�miR-452�miR-224 methylation was a significant independentadverse prognostic factor for biochemical recurrence afterradical prostatectomy in a large prostate cancer patientcohort (n ¼ 293, training cohort), which was successfullyconfirmed in an independent validation cohort (n ¼ 198).
Materials and MethodsClinical samples
Fresh-frozen tissue-tek (Bayer Corporation) embeddedradical prostatectomy and transurethral resection of theprostate (TURP) tissue specimens were collected at Depart-ment of Urology, Aarhus University Hospital, Denmark(Supplementary Table S1) and used for bisulfite sequencingand Infinium analyses.
For MethyLight analyses, two radical prostatectomycohorts were used (Table 1). RP cohort 1 (training) con-sisted of consecutive curatively intended radical prostatec-tomies of histologically verified clinically localized prostatecancer from patients treated at Departments of Urology,Aarhus University Hospital (Aarhus, Denmark; collected1997–2005), and University Hospital Zurich (Zurich,
Translational RelevanceManagement of clinically localized prostate cancer is
highly challenging due to absence of accurate prognostictools. Here, using two independent prostate cancerpatient cohorts, including a total of 491 patients fromfive countries, we found that patients with high GAB-RE�miR-452�miR-224 promoter methylation were atsignificantly higher risk of biochemical recurrence afterradical prostatectomy than patients with low methyla-tion. Although further validation is needed, our resultsindicate that a simple PCR-based test for GABRE�miR-452�miR-224 promotermethylation can add significantprognostic value to currently used routine predictors forprostate cancer patient outcome. We note that our studyused radical prostatectomy specimens; however, if trans-ferred to diagnostic prostate biopsies, or urine or bloodsamples, such a test could be used to aid in treatmentdecisions, for example, between active surveillance andsurgery. Furthermore, we found that hypermethylationwas highly cancer-specific, suggesting that GABRE�miR-452�miR-224 may also have potential as a diagnosticbiomarker for prostate cancer.
Kristensen et al.
Clin Cancer Res; 20(8) April 15, 2014 Clinical Cancer Research2170
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

Switzerland; collected 1993–2001). Out of a total of 633radical prostatectomies, we could retrieve formalin-fixedand paraffin-embedded (FFPE) tissue blocks from 457patients (see Supplementary Fig. S1A for flow chart ofinclusion/exclusion criteria according to REMARK guide-lines). All specimens were evaluated by a trained patholo-gist, representative regions with >90% tumor were markedon hematoxylin and eosin (H&E)-stained sections, andpunch biopsies were taken from the corresponding FFPEblocks for DNA and RNA extraction. Of the 457 patients,164 were excluded due to either lack of follow-up (n¼ 56),pre-/postoperative endocrine treatment (n ¼ 40), or poorDNA quality (n ¼ 68). Final analyses included 293 radicalprostatectomy patients (Table 1).RP cohort 2 (validation) consisted of 310 curatively
intended radical prostatectomies of histologically verifiedclinically localized prostate cancer from Departmentsof Urology, Heinrich Heine University (D€usseldorf, Ger-many; collected 1993–2002), Tampere University Hospital
(Finland; collected 1992–2003), and Karolinska UniversityHospital (Stockholm, Sweden; collected 2003–2007). Atrained pathologist dissected the prostate immediately afterradical prostatectomy and tumor sampleswere snap-frozen.On the basis of H&E-stained sections, DNA was extractedfrom samples with >70% tumor content. Inclusion/exclu-sion criteria were employed as for cohort 1 (SupplementaryFig. S1A). Final analyses comprised 198 radical prostatec-tomy samples (Table 1).
FFPE adjacent nonmalignant (AN), benign prostatichyperplasia (BPH), prostatic intraepithelial neoplasia (PIN),hormone-na€�ve metastatic prostate cancer (MPC, primarytumor), and castration-resistant metastatic prostate cancer(CRPC, primary tumor)were collected, evaluated, andbiop-sied as described for cohort 1 (Table 1; SupplementaryFig. S1B).
The studywas approved by relevant ethical committees ineach country. Written informed consent was obtained fromall patients.
Table 1. Clinicopathologic characteristics of patient sample sets used for MethyLight analyses
Prostate cancer samplesRP cohort 1,training (n ¼ 293)
RP cohort 2,validation (n ¼ 198) MPC (n ¼ 26) CRPC (n ¼ 13) LMN (n ¼ 15)
Median age, range 63 (46–73) 63 (47–75) 71 (52–89) 64 (49–77) 63 (54–71)PSA at diagnosis, n (%)�10 ng/mL 114 (38.9) 121 (61.1) 2 (7.7) 1 (3.6) 2 (13.3)>10 ng/mL 178 (60.8) 69 (34.8) 24 (92.3) 12 (32.1) 11 (73.3)Unknown 1 (0.3) 8 (4.0) — — 2 (13.3)
Pathologic T stage, n (%)T1 0 (0.0) 1 (0.5) NA NA NAT2a-c 184 (62.8) 112 (56.6) NA NA NAT3a-b 106 (36.2) 83 (41.9) NA NA NAT4 3 (1.0) 0 (0.0) NA NA NAUnknown 0 (0.0) 2 (1.0)
Gleason score, n (%)3–6 113 (38.6) 72 (36.4) 2 (7.7) 0 (0.0) NA7 143 (48.8) 90 (45.5) 5 (19.2) 3 (23.1) NA8–10 36 (12.3) 32 (16.2) 19 (73.1) 10 (76.9) NAUnknown 1 (0.3) 4 (2.0) — —
Nodal status, n (%)pN0 253 (86.3) 40 (20.2) NA NA —
pN1 5 (1.7) 1 (0.5) NA NA 15 (100)Unknown 35 (11.9) 157 (79.3) NA NA —
Surgical margin status, n (%)Negative 196 (66.9) 71 (35.9) NA NA NAPositive 93 (31.7) 57 (28.8) NA NA NAUnknown 4 (1.4) 70 (35.4) NA NA NA
Recurrence status, n (%)No recurrence 170 (58.0) 103 (52.0) NA NA NARecurrence 123 (42.0) 95 (48.0) NA NA NA
Mean follow-up time, months (range) 49 (2–151) 60.4 (1–181) NA NA NA
Nonmalignant samples AN (n ¼ 18) BPH (n ¼ 17) PIN (n ¼ 11) — —
Median age (range) 63 (56–72) 67 (56–83) 63 (54–68) — —
Abbreviation: NA, data not applicable/available.
Prognostic Value of GABRE�miR-452�miR-224 Methylation in Prostate Cancer
www.aacrjournals.org Clin Cancer Res; 20(8) April 15, 2014 2171
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

DNA extraction and methylation analysisBisulfite sequencing (biSeq) was performed as described
previously (27, 28). A 441 nt region (containing 52 CpGsites) of the GABRE�miR-452�miR-224 promoter wasamplified using PCR primers: 50-AGATGTTTAGGAGGATTG-GAAGAAT-30 and 50-ACCCCACATCTTATCCCTAAAACAA-30. At least 7 clones were sequenced per sample.
MethyLight. For Danish and Swiss FFPE samples, DNAwas extracted using gDNA Eliminator columns from themiRneasy FFPE Kit (Qiagen) and the blood and cell cultureDNA kit (Qiagen), respectively. For Swedish and Finnishsamples, DNA extraction was conducted using the AllPrepMini DNA/RNA kit (Qiagen), and for German samplesthe Blood and cell culture DNA kit (Qiagen). In all cases,DNA was bisulfite converted using the EZ-96 DNA Meth-ylation-Gold Kit (Zymo Research). The following primers/probes were used for quantitative methylation-specificPCR (MethyLight) analysis of the GABRE�miR-452�miR-224 promoter: 50-GATGTTTAGGAGGATTGGAAGA-30 and50-CTCCGCGCAAATAATCG-30 plus FAM-50-ATATTTTCG-CGGAGATCGGC-30-BHQ1. For normalization and test ofinput DNA quality and quantity, a CpG-free region ofMYOD1 was analyzed in parallel using primers/probe: 50-CCAACTCCAAATCCCCTCTCTAT-30 and 50-TGGTTTTTTT-AGGGAGTAAGTTTGTT-30 plus FAM-50-TCCCTTCCTATT-CCTAAATCCAACCTAAATACCTCC-30-BHQ1. All reactionswere run in triplicates in 384-well plates using the AppliedBiosystems 7900HT Sequence Detection System and Taq-Man Universal PCR Master Mix without AmpErase UNG(Applied Biosystems). Samples with low input DNA (CT
(MYOD1) >36 in at least 2/3 reactions) were excluded fromfurther analysis. The relativeGABRE�miR-452�miR-224pro-moter methylation level in each sample was determined asthe GABRE�miR-452�miR-224/MYOD1 ratio. The accuracyof the GABRE�miR-452�miR-224/MYOD1 assay was vali-dated by direct comparison with biSeq data from severalprostate (cancer) tissue samples andcell lines, andby analysisof dilution series of fully methylated/unmethylated controlDNA (data not shown).
All primers and probes were purchased from EurofinsMWG Operon.
Infinium arrays. One microgram of DNA was bisulfiteconverted, whole genome amplified, and applied to Illumi-na’s Infinium HumanMethylation27 Beadchip v1.2 (29) orHumanMethylation450 BeadChip (Strand and colleagues,unpublished) according to protocols from the manufacturer.
Cell cultureProstatic cell lines PNT1A (European Collection of Cell
Cultures), BPH1 (German Collection of Microorganismsand Cell Cultures), DuCaP and VCaP (kindly provided byDr. Kenneth Pienta, University of Michigan, Ann Arbor,MI), 22rv1, LNCaP, PC3, and DU145 (American TypeCulture Collection) were propagated in RPMI-1640 withL-glutamine (Invitrogen) supplemented with 10% heat-inactivated fetal calf serum, 100 U/mL penicillin, and100 mg/mL streptomycin at 37�C in humidified air atmo-sphere at 5% CO2. PrEC cells were cultured in Prostate
Epithelial Cell BasalMediumwith added Prostate EpithelialGrowth Kit (all from American Type Culture Collection).The authenticity of all cell lineswas verified by short tandemrepeat analysis (http://IdentiCell.eu). PC3 andDU145 cellswere reverse transfected with 20 nmol/L of each miRNA(miRNA mimics purchased from Applied Biosystems/Ambion) using Lipofectamine 2000 reagent (Invitrogen).Medium was changed 6 hours posttransfection. Transfec-tion efficiencies were examined by fluorescencemicroscopyafter transfection of Cy3-conjugated scrambled miRNA(Ambion) and/or verified by qRT-PCR (see below). Cellviability, migration, and invasion assays were performed asdescribed in Supplementary Methods.
Gene Set Enrichment Analysis RNA extraction, qRT-PCR, and ChIP analyses
For details, see Supplementary Methods
Statistical analysesSTATA version 11 (StataCorp) was used for all statistical
analyses. P values <0.05 were considered statistically signif-icant. Phenotypic effects in transfected cell lines and geneexpression differences were assessed by two-sided Studentt tests, and methylation differences by the Mann–WhitneyU test. Correlations between expression and methylationwere analyzed by Spearman rank test. Associations betweenGABRE�miR-452�miR-224 methylation and clinicopath-ologic variables were assessed using Spearman rank test orthe Mann–Whitney U test. The diagnostic potential ofGABRE�miR-452�miR-224/MYOD1 methylation wasevaluated by receiver operator characteristics (ROC) curveanalysis.
For survival analysis, biochemical/PSA recurrence (Dan-ish, Swiss, Swedish, and German samples cutoff �0.2ng/mL, Finnish samples cutoff �0.5 ng/mL, based on localclinical practice) was used as endpoint. Patients not havingexperienced biochemical recurrence were censored at theirlast normal PSA measurement. The prognostic value ofGABRE�miR-452�miR-224/MYOD1methylationwas eval-uatedbyKaplan–Meier analysis and two-sided log-rank test,and by univariate and multivariate Cox regression analysisas a continuous as well as a dichotomous variable. Foranalysis of GABRE�miR-452�miR-224/MYOD1 methyla-tion as a dichotomous variable, patients in cohort 1 weredivided intohigh and lowmethylation groups using a cutoffvalue determined by ROC analysis of recurrence/nonrecur-rence status optimized for high specificity. For validation,patients in cohort 2 were divided into two groups using thecutoff (fraction) defined in cohort 1. The proportionalhazards assumption was verified by the log-negative-logsurvival distribution function for all variables. Prognosticaccuracy was estimated using Harrell Concordance Index.
ResultsSilencing of the GABRE/miR-452/miR-224 locus inprostate cancer
By microarray and qRT-PCR analyses, we observed sig-nificant downregulation of GABRE, miR-224, and miR-452
Kristensen et al.
Clin Cancer Res; 20(8) April 15, 2014 Clinical Cancer Research2172
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
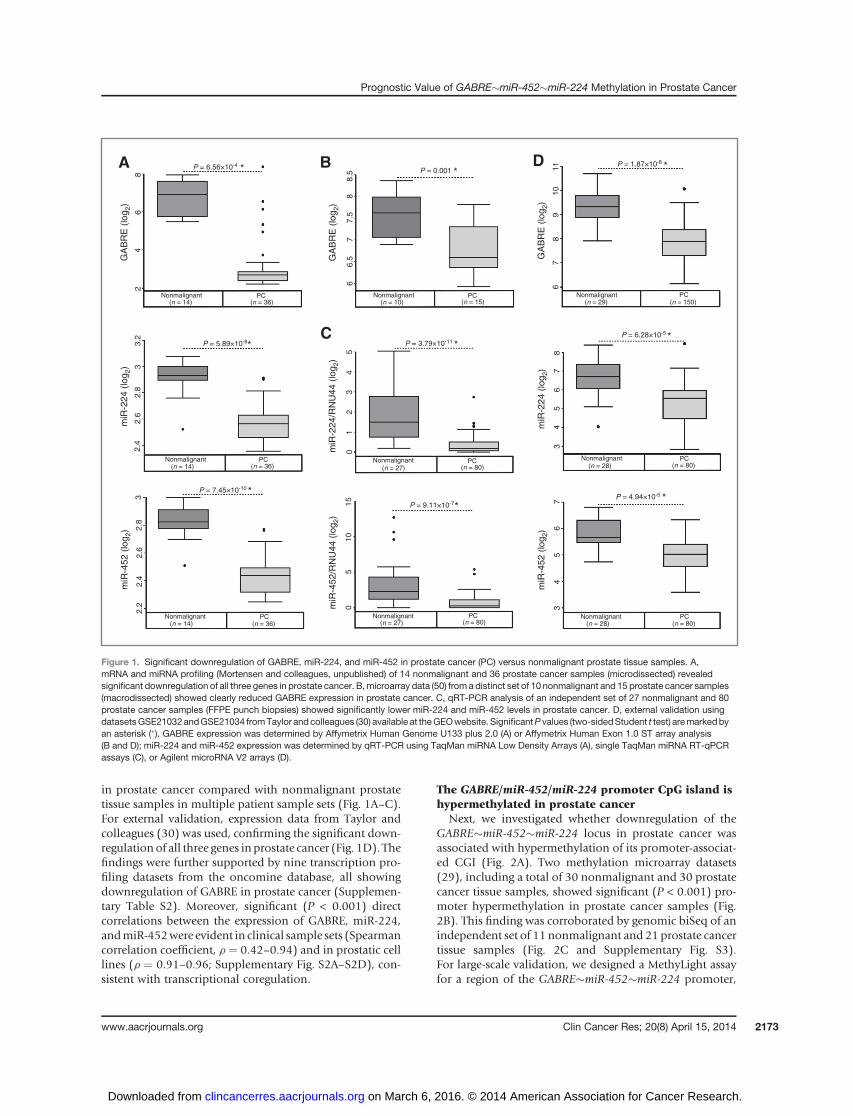
in prostate cancer compared with nonmalignant prostatetissue samples in multiple patient sample sets (Fig. 1A–C).For external validation, expression data from Taylor andcolleagues (30) was used, confirming the significant down-regulationof all three genes in prostate cancer (Fig. 1D). Thefindings were further supported by nine transcription pro-filing datasets from the oncomine database, all showingdownregulation of GABRE in prostate cancer (Supplemen-tary Table S2). Moreover, significant (P < 0.001) directcorrelations between the expression of GABRE, miR-224,andmiR-452were evident in clinical sample sets (Spearmancorrelation coefficient, r ¼ 0.42–0.94) and in prostatic celllines (r ¼ 0.91–0.96; Supplementary Fig. S2A–S2D), con-sistent with transcriptional coregulation.
The GABRE/miR-452/miR-224 promoter CpG island ishypermethylated in prostate cancer
Next, we investigated whether downregulation of theGABRE�miR-452�miR-224 locus in prostate cancer wasassociated with hypermethylation of its promoter-associat-ed CGI (Fig. 2A). Two methylation microarray datasets(29), including a total of 30 nonmalignant and 30 prostatecancer tissue samples, showed significant (P < 0.001) pro-moter hypermethylation in prostate cancer samples (Fig.2B). This finding was corroborated by genomic biSeq of anindependent set of 11 nonmalignant and 21 prostate cancertissue samples (Fig. 2C and Supplementary Fig. S3).For large-scale validation, we designed a MethyLight assayfor a region of the GABRE�miR-452�miR-224 promoter,
Nonmalignant Nonmalignant Nonmalignant
Nonmalignant
NonmalignantNonmalignantNonmalignant
Nonmalignant Nonmalignant
(n = 14) (n = 10) (n = 29)
(n = 28)
(n = 28)(n = 27)(n = 14)
(n = 14) (n = 27) (n = 80)(n = 36)
(n = 36) (n = 80) (n = 80)
(n = 80)
(n = 150)(n = 36) (n = 15)PC PC PC
PC
PCPCPC
PC PC
P = 6.56×10-4
P = 5.89×10-8
P = 7.45×10-10
P = 9.11×10-7P = 4.94×10-5
P = 3.79×10-11
P = 0.001
P = 6.28×10-5
P = 1.87×10-8
* *
*
**
*
*
*
*
2
6 67
89
10
11
34
56
78
02.4
2.2 0
5
34
56
7
10
15
2.4
2.6
2.8
32
.62.8
33
.2
12
34
56
.57
7.5
88
.5
46
8
GA
BR
E (
log
2)
GA
BR
E (
log
2)
GA
BR
E (
log
2)
miR
-224 (
log
2)
miR
-45
2 (
log
2)
miR
-45
2/R
NU
44
(lo
g2)
miR
-45
2 (
log
2)
miR
-224 (
log
2)
miR
-224/R
NU
44 (
log
2)
A B D
C
Figure 1. Significant downregulation of GABRE, miR-224, and miR-452 in prostate cancer (PC) versus nonmalignant prostate tissue samples. A,mRNA and miRNA profiling (Mortensen and colleagues, unpublished) of 14 nonmalignant and 36 prostate cancer samples (microdissected) revealedsignificant downregulation of all three genes in prostate cancer. B,microarray data (50) from a distinct set of 10 nonmalignant and 15 prostate cancer samples(macrodissected) showed clearly reduced GABRE expression in prostate cancer. C, qRT-PCR analysis of an independent set of 27 nonmalignant and 80prostate cancer samples (FFPE punch biopsies) showed significantly lower miR-224 and miR-452 levels in prostate cancer. D, external validation usingdatasetsGSE21032andGSE21034 fromTaylor and colleagues (30) available at theGEOwebsite. SignificantP values (two-sidedStudent t test) aremarkedbyan asterisk (�). GABRE expression was determined by Affymetrix Human Genome U133 plus 2.0 (A) or Affymetrix Human Exon 1.0 ST array analysis(B and D); miR-224 and miR-452 expression was determined by qRT-PCR using TaqMan miRNA Low Density Arrays (A), single TaqMan miRNA RT-qPCRassays (C), or Agilent microRNA V2 arrays (D).
Prognostic Value of GABRE�miR-452�miR-224 Methylation in Prostate Cancer
www.aacrjournals.org Clin Cancer Res; 20(8) April 15, 2014 2173
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

including 7 CpG sites also analyzed by biSeq (Fig. 2A). ACpG-free region of MYOD1 was used for normalization(represents total input of bisulfite-converted DNA). GAB-RE�miR-452�miR-224/MYOD1 methylation was deter-mined in 291 FFPE tissue samples (191 LPC/RP samplesfrom cohort 1, plus several nonmalignant, premalignant,
and advanced prostate cancer samples; Table 1). Overall,GABRE�miR-452�miR-224 was significantly (P ¼ 3.56 �10�19; Mann–Whitney U test) hypermethylated in prostatecancer compared with nonmalignant tissue samples(Fig. 2D, left). Also,GABRE�miR-452�miR-224was hyper-methylated in 3 of 11 PIN samples, suggesting it could be an
–351
–270
Bisulfite sequencingA
B C
D
MethyLight
(7 CpGs)
(52 CpGs)
–351
+144
+470–404 CpG island
(83 CpGs)
+14967
+15051
+16021
+16101
TSS
miR-452 miR-224+1 (ATG)
Exon 1
Intron 6
GABRE
27K dataset
Non-
malignant
PC
(n = 9)(n = 9)
450K dataset
Nonmalignant (n = 21)
100
80
60
40
20
0
PC (n = 21)P = 1.00 P = 0.26
P = 6.98 × 10–7
cg08
7830
90
cg25
5286
46
cg07
0538
80
CpG islandcg12204574
Nonmalignant
(n = 35)
PC
(n = 245)AN
(n = 18)
BPH
(n = 17)
PIN
(n = 11)
LPC
(n = 191)
CRPC
(n = 13)
MPC
(n = 26)
LNM
(n = 15)
Se
nsitiv
ity
MethyLight dataset
AUC = 0.98
0.00 0.25 0.50 0.75 1.00
Sensitivity = 95.5%
1 – Specificity
Specificity = 94.3%
cg01
4805
50
cg12
2045
74
BiSeq dataset
AN BPH LPC MPC(n = 7) (n = 4) (n = 10) (n = 11)
0.6
0.4
0.2
0
P = 0.006
P = 3.56 × 10–19
β-va
lue
β-va
lue
1.0
0.8
0.6
0.4
0.2
0.0
% G
AB
RE
~miR
-452
~miR
-224
met
hyla
tio
n
Me
thyla
tio
n
GA
BR
E~
miR
-224
~m
iR-4
52/M
YO
D1
Me
thyla
tio
n
GA
BR
E~
miR
-224
~m
iR-4
52/M
YO
D1
01
23
4
01
23
4
0.0
00.2
50.5
00.7
51.0
0
NS
Figure 2. A, schematic structure of GABRE�miR-224�miR-452 genomic locus (not to scale). miR-224 and miR-452 are located in intron 6. Location ofpromoter-associated CpG island, containing 83 CpG sites, is shown by a gray box. Regions analyzed by bisulfite sequencing and MethyLight are markedwith arrows, indicating PCRprimer positions. TSSmarks the putative transcription start site. B–D,GABRE�miR-452�miR-224promotermethylation levels infour independent clinical sample sets determined by (B) Illumina 27 K and 450 Kmicroarray analysis, (C) bisulfite sequencing (percent methylated CpG sites/total number of CpG sites; see Supplementary Fig. S3 for more details), or (D) MethyLight. The number of patients in each group is given in brackets.B, an asterisk (�) indicates significant P values (P < 0.001; Mann–Whitney U Test) for Illumina probes covering GABRE�miR-452�miR-224. C andD, horizontal gray lines mark median methylation levels in each group and significant P values (P < 0.05 vs. ANþBPH; Mann–Whitney U test) aremarked by an asterisk (�). D, ROCcurve analysis (right) showing accurate discrimination between nonmalignant and prostate cancer tissue samples based onGABRE�miR-452�miR-224 methylation. AUC, area under the curve. AN, adjacent nonmalignant prostate; BPH, benign prostatic hyperplasia; LPC,clinically localized prostate cancer (radical prostatectomy specimen); MPC, hormone-naïve metastatic prostate cancer (primary tumor); CRPC, castration-resistant prostate cancer (primary tumor); LMN, lymph node metastasis. NS, not significant.
Kristensen et al.
Clin Cancer Res; 20(8) April 15, 2014 Clinical Cancer Research2174
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

early event in prostate carcinogenesis. Methylation levelswere similar between subgroups of nonmalignant (AN/BPH) and malignant (LPC/MPC/CRPC/LNM) samples,respectively (Fig. 2D, middle). Together, these resultsstrongly indicate that aberrant promoter hypermethylationis associated with coordinated downregulation of the entireGABRE�miR-452�miR-224 locus in prostate cancer.Consistentwithepigeneticsilencing,asignificant(P<0.001)
inverse correlation between promoter methylation and miR-224 (Spearman rho, r ¼ �0.41) or miR-452 expression (r ¼�0.43), respectively, was seen in the clinical samples (Supple-mentary Fig. S2C). Likewise, expression of GABRE, miR-224,and miR-452 was strongly inversely correlated with pro-moter methylation (r ¼ �0.83 to�0.88) in 9 prostatic celllines (Supplementary Fig. S2E). GABRE�miR-452�miR-224was hypermethylated in 4 of 6 prostate cancer cell linesand in 0 of 3 nonmalignant prostate cell lines (Supplemen-tary Fig. S4A–S4C). Treatment with the demethylating drug5-aza-dCwas insufficient for reactivation of transcription inhypermethylated DU145 and PC3 cells, which may beexplained by presence of high levels of the repressiveH3K27me3 mark at the inactive GABRE�miR-452�miR-224 promoter (Supplementary Fig. S4D and S4E).To assess the diagnostic potential of GABRE�miR-
452�miR-224methylation, we performed ROC curve anal-ysis using the large MethyLight dataset (Fig. 2D, left). Thepreliminary sensitivity and specificity of GABRE�miR-452�miR-224 methylation for prostate cancer detectionwas 95.5% and 94.3%, respectively (Fig. 2D, right). Togeth-er, our findings strongly indicate that aberrant promoterhypermethylation of the GABRE�miR-452�miR-224 pro-moter-associated CGI is a frequent event in clinical prostatecancer samples as well as in prostate cancer cell lines. Wenote that theGABRE�miR-452�miR-224 locus is located atXq28 within a cluster of MAGE (cancer-testis antigens)genes known to be hypomethylated in prostate cancer andrecently described as a long range epigenetic activationregion (31). Focal hypermethylation of the GABRE�miR-452�miR-224 promoterwithin this otherwise activated andhypomethylated region (Supplementary Fig. S5) suggeststhat epigenetic silencing of GABRE�miR-452�miR-224 ishighly specific and may be selected for during prostatecancer development and/or progression.
miR-224 and miR-452 inhibit viability, migration, andinvasion of prostate cancer cellsNext,we investigated thepotential functional rolesofmiR-
224 and miR-452 in prostate cancer (GABRE was not ana-lyzed aswewere unable to validate a good antibody; datanotshown). miR-224 or miR-452 mimics were transfected intotwo prostate cancer cell lines (PC3 and DU145) with lowendogenous expression (Supplementary Fig. S4B). As posi-tive controls,weusedmiR-145 in cell viability assays (knownto reducePC3andDU145 cell viability; ref. 32) andmiR-101in cell migration/invasion assays (known to inhibit migra-tion and invasion of PC3 and DU145 cells; ref. 33). Scram-bled miRNA was used as a negative control. Transfectionefficiencies were confirmed by qRT-PCR (data not shown).
Reintroduction of either miR-224 or miR-452 had amoderate (�25%) and statistically significant (P < 0.05)inhibitory effect on cell viability in both PC3 and DU145cells, as evident at 72 hours posttransfection but not at 48hours (Fig. 3A). Furthermore, ectopic expression of miR-224 or miR-452 significantly (P < 0.05) reduced migrationand invasion of PC3 and DU145 cells at 48 hours post-transfection (Fig. 3B and C), and hence was not simplyascribed to miRNA-induced reduction in cell viability, asthis occurred at 72 hours. Cotransfection of miR-224 and
Figure 3. Ectopic expression of miR-224 or miR-452 inhibited viability,migration, and invasion in two prostate cancer cell lines. A, cell viability at48 and 72 hours posttransfection of miR-224 or miR-452 mimics (20nmol/L) in PC3 and DU145 cells, as determined by an MTT assay. Allresults are presented relative to scrambled (scr) miRNA transfected cells.B and C, Trevigen Cultrex Migration and Invasion assays performed 48hours posttransfection. Background migration levels for nontransfectedcells without chemoattractant (no serum) are also given (B). miR-145 andmiR-101 were used as positive controls for inhibition of cell viability (A)and migration/invasion (B and C), respectively. The presented results(A-C) are representative of a minimum of three experiments performed intriplicate; all significant phenotypic effects, that is, P < 0.05 (two-sidedStudent t test) and >25% reduction, are marked by an asterisk (�).
Prognostic Value of GABRE�miR-452�miR-224 Methylation in Prostate Cancer
www.aacrjournals.org Clin Cancer Res; 20(8) April 15, 2014 2175
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

miR-452 did not cause further inhibition of cell viability,migration, or invasion (Fig. 3A–C), suggesting at least somefunctional redundancy between these miRNAs. Subsequent-ly, we performed GSEA of microarray transcriptional profil-ing data obtained at 48 hours posttransfection. For genes(mRNA) downregulated upon miR-224 or miR-452 trans-fection, the top ten most significantly enriched "Canon-ical Pathways" in both DU145 and PC3 cells (Supple-mentary Fig. S6A and S6B) were primarily associated withpositive regulation of the cell cycle, DNA replication, andmitosis, consistent with the observed negative effect ofthese miRNAs on prostate cancer cell viability (Fig. 3A).GSEA was also performed for genes significantly upregu-lated upon miR-224 or miR-452 transfection, likely repre-senting indirect effects. Several "Canonical Pathways"related to cellular adhesion and motility were enrichedfor both miR-224 and miR-452 (Supplementary. Fig. S6Cand S6D) in agreement with their effects on migrationand invasion (Fig. 3B and C).
Among the genesmost heavily downregulated in both celllines aftermiR-224 andmiR-452 overexpression, respective-ly (Supplementary Tables S3 and S4), we shortlisted poten-tial target genes using the following criteria: (i) mRNAscontaining at least one 7mer target site predicted by bothTargetScan (www.targetscan.org) and PicTar (http://pictar.mdc-berlin.de), and (ii)mRNAs significantly upregulated inprostate cancer versus nonmalignant prostate tissue samplesin multiple patient sample sets (Supplementary Table S5).Among eight shortlisted potential miR-224 target genes,expression ofC1orf116 (chromosome1 open reading frame116, also known as specifically androgen-regulated gene,SARG), GOLM1 (golgi membrane protein 1), and FAM64A(family with sequence similarity 64, member A) was signif-icantly inversely correlated with miR-224 expression in thedatasets fromTaylor and colleagues (30) andMortensenandcolleagues (Spearman r: �0.49 to �0.25; P < 0.05; Supple-mentary Table S5), suggesting that these genes could beclinically relevant.Moreover, among eight shortlistedpoten-tialmiR-452 target genes, expressionofDR1 (downregulatorof transcription 1) and IGF2BP2 [insulin-like growth factor2mRNAbinding protein 2 (p62)]was significantly inverselycorrelated with miR-452 in the dataset fromMortensen andcolleagues (r: �0.47 to �0.30; P < 0.05), but no significantcorrelations were seen in the Taylor and colleagues dataset(Supplementary Table S5).
Prognostic value of GABRE/miR-452/miR-224methylation and association with clinicopathologicalparameters
Next, we investigated possible correlations between clin-icopathologic variables and GABRE�miR-452�miR-224methylation in two independent radical prostatectomycohorts (Table 1; Supplementary Fig. S1A). There was nosignificant correlation betweenGABRE�miR-452�miR-224methylation and age at diagnosis (Supplementary. Fig.S7A), but high methylation was significantly (P < 0.05)associated with the established adverse prognostic factorshigh PSA, high T-stage, high Gleason score, and positive
surgicalmargin status in at least one cohort (SupplementaryFig. S7B–S7E).
To assess the prognostic value ofGABRE�miR-452�miR-224 methylation, we performed PSA-based recurrence-freesurvival (RFS) analysis. By univariate Cox regression anal-ysis of GABRE�miR-452�miR-224 methylation (as a con-tinuous variable) in RP cohort 1 (training cohort, n ¼ 293;see Table 1), short RFS timewas significantly associatedwithhigh GABRE�miR-452�miR-224 methylation, as well aswith the established prognostic factors high PSA, highGleason score, advanced T-stage, and positive surgical mar-gin status (P < 0.001; Table 2). All variables remainedsignificant in a multivariate model (Table 2), indicatingthat GABRE�miR-452�miR-224 methylation is a signifi-cant independent prognostic factor for prostate cancerpatients treated by radical prostatectomy. The prognosticvalue of GABRE�miR-452�miR-224 methylation was suc-cessfully validated by univariate as well as multivariateanalysis in cohort 2 (validation cohort, n ¼ 198;see Table 1; Table 2). In cohort 2, surgical margin statuswas left out in the final model, because of missing data for70 patients. Predictive accuracies were estimated by HarrellC-index, which increased from0.73 to 0.74 in cohort 1, andfrom 0.75 to 0.77 in cohort 2 by addition of GABRE�miR-452�miR-224 methylation, suggesting moderatelyimproved model performance. The contribution of GAB-RE�miR-452�miR-224 methylation to the multivariatemodel was comparable with that of a single clinicopatho-logic variable: Harrell C-index of the multivariate model(cohort 1/cohort 2) when leaving out either PSA (0.72/0.76), T-stage (0.72/0.74), Gleason score (0.73/0.73), orsurgical margin status (0.73/not applicable).
Similar results were obtained when GABRE�miR-452�miR-224methylation was analyzed as a dichotomousvariable in uni- and multivariate Cox regression analyses(Supplementary Table S6). For these analyses, patients incohort 1were divided into low/highmethylation subgroupsusing a cutoff value determined by ROC curve analysis(optimized for high specificity). Likewise, patients in cohort2 were divided into low/high methylation subgroups usingthe cutoff (fraction) defined in cohort 1. Finally, by Kaplan–Meier analysis, high GABRE�miR-452�miR-224 methyla-tion was significantly (P < 0.001, log-rank test) associatedwith short RFS in cohort 1 (Fig. 4A), which was successfullyvalidated (P < 0.001) in cohort 2 (Fig. 4B). In summary, wefound GABRE�miR-452�miR-224 methylation to be a sig-nificant independent prognostic predictor of biochemicalrecurrence in two radical prostatectomy patient cohorts.
Finally, to assess the possible prognostic value of GABRE,miR-452, andmiR-224 expression, we used the large radicalprostatectomy patient cohort from Taylor and colleagues(30). A multigene model including the expression of allthree genes was a significant independent predictor ofbiochemical recurrence after radical prostatectomy (HR,2.37; P ¼ 0.013; Supplementary Table S7). Expression ofeither GABRE, miR-224, or miR-452 individually was notsignificant in multivariate analysis, but GABRE and miR-224 were significant in univariate analysis (Supplementary
Kristensen et al.
Clin Cancer Res; 20(8) April 15, 2014 Clinical Cancer Research2176
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

Table S7). Finally, among the shortlistedmiR-224 andmiR-452 potential target genes (Supplementary Table S5), highexpression of OAZ2 (Ornithine Decarboxylase Antizyme 2;
predicted miR-452 target) had significant adverse prognos-tic value inmultivariate analysis in theTaylor and colleaguesdataset (Supplementary Table S8). A multigene model
Cohort 1A B Cohort 2
Low meth.
High meth.
P < 0.001
Low meth.
High meth.
P < 0.001
0.0
00
.25
0.5
00
.75
1.0
0
0.0
00
.25
0.5
00
.75
1.0
0
0 25 50 75 100 125
Recurrence-free survival time (months)
0 25 50 75 100 125
Recurrence-free survival time (months)
Cu
mu
lative
su
rviv
al
Cu
mu
lative
su
rviv
al
Number at risk
Low meth. 193
100
141
67
93
40
33
18
21
5
8
1
130
68
107
40
88
29
64
16
27
1
12
0High meth.
Figure 4. Kaplan–Meier survival analysis of RFS based onGABRE�miR-452�miR-224/MYOD1methylation levels (low vs. high) in radical prostatectomy samples.A, patients in cohort 1 (training)weredivided intohighversus lowmethylationgroupsafterROCanalysis.B,patients invalidationcohort2weredivided into low/highmethylation groupsaccording to thecutoff (fraction)defined incohort 1.Significant (�)P values for two-sided log-rank test aregiven.HighGABRE�miR-452�miR-224 methylation was significantly associated with early biochemical recurrence after radical prostatectomy in two independent cohorts.
Table 2. Uni- and multivariate Cox regression analyses of biochemical recurrence-free survival in two RPcohorts
Univariate Multivariate
Variable Characteristics HR (95% CI) P C-indexa HR (95% CI) P C-indexa
Cohort 1 (training),n ¼ 293
Age at diagnosis Continuous 0.98 (0.95–1.01) 0.211 0.55 — —
Preoperative PSA �10 vs. >10 ng/mL 2.67 (1.75–408) <0.001 0.61 2.37 (1.53–3.69) <0.001 0.74 0.73Tumor stage pT2a-c vs. pT3a-c 3.37 (2.35- 4.84) <0.001 0.65 2.06 (1.37–3.10) 0.001Surgical margin status Negative vs. positive 3.03 (2.10–4.36) <0.001 0.63 1.99 (1.33–2.98) 0.001Gleason score Continuous 1.48 (1.22–1.80) <0.001 0.59 1.27 (1.02–1.58) 0.033GABRE�miR-452�miR-224methylation
Continuous 1.75 (1.37–2.23) <0.001 0.61 1.38 (1.06–1.81) 0.019
Cohort 2 (validation),n ¼ 198
Age at diagnosis Continuous 1.02 (0.99–1.06) 0.206 0.53 — —
Preoperative PSA �10 vs. >10 ng/mL 2.20 (1.45–3.34) <0.001 0.59 2.22 (1.45–3.41) <0.001 0.77 0.75Tumor stage pT2a-c vs. pT3a-c 3.60 (2.36–5.49) <0.001 0.66 2.58 (1.63–4.08) <0.001Surgical margin statusb Negative vs. positive 2.50 (1.44–4.37) 0.001 0.62 — —
Gleason score Continuous 1.84 (1.50–2.25) <0.001 0.69 1.71 (1.45–3.41) <0.001GABRE�miR-452�miR-224methylation
Continuous 2.99 (1.71–5.21) <0.001 0.65 2.45 (1.26–4.75) 0.008
aPredictive accuracy, estimated by the Harrell concordance index (c-index). Left column, C-index based on all variables; right column,C-index based on clinicopathological variables only (i.e. excluding GABRE�miR-452�miR-224 methylation).bMargin status was excluded from the final multivariate analysis, because of missing data for 70 of 198 patients in cohort 2.Significant P values (P < 0.05) are marked in bold.
Prognostic Value of GABRE�miR-452�miR-224 Methylation in Prostate Cancer
www.aacrjournals.org Clin Cancer Res; 20(8) April 15, 2014 2177
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

including GABRE,miR-224, miR452, andOAZ2 expressionalso remained significant in a multivariate model (Supple-mentary Table S8), further supporting a clinically relevantrole of theGABRE�miR-224�miR-452 locus and its expres-sion circuit in prostate cancer.
DiscussionThis study showed that the GABRE�miR-452�miR-224
locus is downregulated in prostate cancer and that aberrantGABRE�miR-452�miR-224 promoter hypermethylation is avery common event in prostate cancer, suggesting it is a keymolecularmechanism for coordinated silencing of this locusin prostate cancer cells. Moreover, GABRE�miR-452�miR-224 hypermethylation was found to be a significant inde-pendent predictor of biochemical recurrence in two radicalprostatectomy patient cohorts from five European countries.To the best of our knowledge, this is the first comprehensivestudy of GABRE�miR-452�miR-224 expression in prostatecancer as well as the first report to identify GABRE�miR-452�miR-224 as a promising diagnostic and prognosticmethylation marker candidate for prostate cancer.
Consistent with our results, downregulation of miR-224in prostate cancer tissue samples has been reported in tworecent studies (14, 15). Another study has reportedmiR-224upregulation inPNI-positive versus negative prostate cancer(16), but used only a small sample set without nonmalig-nant prostate samples for comparison. PNI status was notdetermined in our sample set. Moreover, although oneearlier study has reported upregulation of miR-452 inprostate cancer progenitor cells (17), differential expressionin prostate cancer patient samples has not been previouslydescribed. There are no prior reports ofGABREderegulationin prostate cancer, but upregulation has been reported innon–small cell lung cancer (34).
In this study,GABRE�miR-452�miR-224 promoter meth-ylation was found to be highly cancer-specific (AUC¼ 0.98)and thereby comparable to previously reported top candidatemethylationmarkers for prostate cancer detection, includingGSTP1 (5). Furthermore, our findings indicated that GAB-RE�miR-452�miR-224methylation has significant indepen-dent prognostic value for prediction of biochemical recur-rence after radical prostatectomy, beyond routine clinico-pathologic predictors. The prognostic potential of GAB-RE�miR-452�miR-224 methylation was identified in atraining cohort fromDenmark and Switzerland and success-fully validated in an independent patient set from Germany,Finland, and Sweden, supporting the validity of our finding.Before the present study,PITX2 (3, 4) andC1orf114 (29)werethe only single genes with independent prognostic valuereported in tworadicalprostatectomycohorts. Further studiesincluding large prospective cohorts are needed to assess theclinical utility of GABRE�miR-452�miR-224methylation asa prognostic candidate marker alone and in combinationwith other candidate molecular markers.
The significant prognostic value ofGABRE�miR-452�miR-224 methylation was also reflected at the expression level,when we analyzed the radical prostatectomy cohort fromTaylor and colleagues (30). Thus, a three-gene GABRE/
miR-224/miR-452expressionsignaturewasa significant inde-pendent predictor of PSA recurrence after radical prostatec-tomy in this cohort. Low expression was associated withhigher recurrence risk, consistent with an important roleof coordinated epigenetic silencing of the GABRE�miR-452�miR-224 locus in prostate cancer.
Our work showed that miR-224 and miR-452 inhibitproliferation, migration, and invasion of two prostate can-cer cell lines. The GABRE�miR-452�miR-224 locus has notbeen functionally linked to prostate cancer before, butearlier studies have shown involvement of miR-224 in bothrepression and stimulation of cell viability, migration, and/or invasion in other cancer types (20, 22, 35). None of thevalidatedmiR-224 targets from these studies (e.g., KLK10 inovarian cancer and RKIP in breast cancer; refs. 20, 35) weresignificantly affected, when we overexpressed miR-224 inPC3 orDU145 cells (data not shown), consistent with a celltype and context-dependent function of thismiRNA. To ourknowledge, there arenoprevious reports of a functional rolenor validated target for miR-452. Using an integrativebioinformatic approach, we shortlisted 16 predicted miR-224ormiR-452 target genesof potential clinical importancein prostate cancer (Supplementary Table S5). Among these,we identified the three predicted miR-224 target genesC1orf116, GOLM1, and FAM64A that were not only down-regulated in PC3 and DU145 cells upon miR-224 over-expression, but also significantly upregulated in prostatecancer tissue versus nonmalignant tissue samples in mul-tiple patient sets. Similarly, we identified predictedmiR-452target genes, including IGF2BP2, DR1, and OAZ2.
These genes have all been previously linked to cancer andin some cases prostate cancer specifically. Thus, GOLM1hasbeen suggested as a putative tissue and urine biomarker ofprostate cancer (36–39) and shown to induce migrationand invasion of prostate cancer cells (40). C1orf116 hasbeen reported as highly expressed in prostate cancer tissue(41), whereas FAM64A is highly expressed in leukemia andlymphoma, and known to promote cell division (42, 43).Moreover, we found that high expression of the predictedmiR-452 target gene OAZ2 was associated with high recur-rence risk after radical prostatectomy in the patient cohortfrom Taylor and colleagues. Expression of OAZ2 has pre-viously been associated with aggressive behavior in neuro-blastoma (44). IGF2BP2 expression has been associatedwith liver cancer, liposarcoma, and endometrial adenocar-cinomas, and with promotion of cell migration and metas-tasis (45, 46). High expression of DR1 is associated withbasal cell carcinoma in renal transplant patients (47).Another interesting potential miR-452 target gene, ARF4(ADP-ribosylation factor 4), has been shown to promotebreast cancer cell migration (48) and to inhibit apoptosis inglioblastoma (49).
Additional studies are needed to investigate the molec-ular pathways and target genes regulated by miR-224 andmiR-452 in prostatic cells in more details. Moreover, thepotential functional role of the host geneGABRE remains tobe investigated. Despite several attempts to overexpressGABRE in prostate cancer cell lines, we observed no
Kristensen et al.
Clin Cancer Res; 20(8) April 15, 2014 Clinical Cancer Research2178
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

phenotypic effects (data not shown); however, we wereunable to validate GABRE expression at the protein leveldue to lack of a specific antibody. ElevatedGABA levels havepreviously been associated with prostate cancer metastasis(10) and GABA agonist isoguvacine has been shown tostimulate proliferation of some prostate cancer cell lines(11), but the exact composition(s) of the GABA receptor(s)mediating these effects are not known. Furthermore, GABAsignaling has been found to inhibit breast cancer cellmigration (12), suggesting a context-dependent function.We note that all methylation analyses were performed
using radical prostatectomy tissue specimens. A prognostictest based on GABRE�miR-452�miR-224 methylationwould currently have limited clinical utility at this stage,as there are no established adjuvant therapies after radicalprostatectomy. In future studies, it will therefore be impor-tant to investigate if GABRE�miR-452�miR-224 methyla-tion levels measured in prostate biopsies, or in a urine orblood sample, taken at the time of diagnosis can predictprostate cancer aggressiveness and thus guide treatmentdecisions. Conceivably, a high level of GABRE�miR-452�miR-224 methylation in a biopsy may favor interven-tion (e.g., surgery) over active surveillance. Likewise, anincrease in GABRE�miR-452�miR-224 methylation inbiopsies taken during active surveillance could indicatedisease progression, and hence the need for intervention.Similarly, to assess the clinical utility of GABRE�miR-
452�miR-224 methylation for prostate cancer diagnosis,future studies should investigate whether detection oftumor-derived hypermethylated GABRE�miR-452�miR-224 DNA in biofluids, such as urine or blood, can be usedto identify patientswith prostate cancer. Given the very highfrequency and cancer specificity of GABRE�miR-452�miR-224promoter hypermethylation thatwe observed in radicalprostatectomy tissue samples, it is possible that a methyl-ation test forGABRE�miR-452�miR-224 in biofluids couldbe used as a supplement to for example, a PSA test (highsensitivity, low specificity) to increase the accuracy of diag-nosis. Furthermore, premalignant and/or cancer field effectsthat are not evident histopathologically may be detected byDNA methylation analysis of prostate biopsies, as demon-strated by others for e.g. GSTP1 (6, 7). Accordingly, GAB-RE�miR-452�miR-224 methylation analysis could alsopotentially be used to guide repeat biopsy in men withinitial cancer-negative biopsies.The use of PSA recurrence as clinical endpoint is also a
limitation of our study. Notably, due to lower sensitivity ofthe specific PSAkit applied, a higher cutoff valuewas used todefine biochemical recurrence in the Finnish patients (0.5ng/mL) compared with other patients (0.2 ng/mL). Thus, itcannot be excluded that detection of PSA recurrence wasslightly delayed for some of the Finnish patients. However,
time to recurrence is also influenced by the frequency andexact timing of PSA testing after radical prostatectomy,which despite regulation by clinical guidelines also mayvary between patients in real life. Future studies shouldinvestigate other clinical endpoints such as cancer-specificand overall survival. Given the long disease course ofprostate cancer this would require large cohorts with atleast 10 to 20 years of follow-up.
In conclusion, our results showed that the GAB-RE�miR-452�miR-224 locus is epigenetically silenced inprostate cancer compared with nonmalignant prostatetissue samples. Furthermore, our findings indicated thatmiR-224 and miR-452 hold important tumor suppressorfunctions in prostate cancer. Finally, we identified theGABRE�miR-452�miR-224 promoter as a promising newcandidate methylation marker for prostate cancer detec-tion and for prediction of biochemical recurrence afterradical prostatectomy.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: H. Kristensen, S. Høyer, M. Borre, T.F. �rntoft,K.D. SørensenDevelopment of methodology: H. Kristensen, C. Haldrup, S. Høyer, K.D.SørensenAcquisitionofdata (provided animals, acquired andmanagedpatients,provided facilities, etc.): H. Kristensen, S. Strand, K. Mundbjerg, M.M.Mortensen, P.J. Wild, C. Arsov, T. Visakorpi, L. Egevad, J. Lindberg,H. Gr€onberg, S. Høyer, M. Borre, K.D. SørensenAnalysis and interpretation of data (e.g., statistical analysis, biosta-tistics, computational analysis): H. Kristensen, K. Mundbjerg, M.M.Mortensen, K. Thorsen,M.S. Ostenfeld, S. Høyer, T.F.�rntoft, K.D. SørensenWriting, review, and/or revision of the manuscript: H. Kristensen,C. Haldrup, K. Mundbjerg, M.S. Ostenfeld, P.J. Wild, C. Arsov, W. Goering,T. Visakorpi, H. Gr€onberg, S. Høyer, M. Borre, T.F. �rntoft, K.D. SørensenAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): C. Haldrup, M.M. Mortensen, P.J.Wild, W. Goering, J. LindbergStudy supervision: C. Haldrup, M.M. Mortensen, K.D. Sørensen
AcknowledgmentsThe authors thank Anne Slotsdal, Pamela Celis, Maria Mark, Gitte Høj,
Mette Rasmussen, Conni Sørensen and Susanne Skou for excellent technicalassistance and Dr. Wolfgang A. Schulz for comments on the article. TheDanishCancer Biobank (DCB) is acknowledged for biologicmaterial and forinformation regarding handling and storage.
Grant SupportThis work was supported by The Danish Cancer Society, The Lundbeck
Foundation, The John andBirtheMeyer Foundation, TheDanishCouncil forStrategic Research, The Danish Advanced Technology Foundation, and TheDeutsche Forschungsgemeinschaft (Schu604/16-3).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received September 25, 2013; revised January 6, 2014; accepted February5, 2014; published online April 15, 2014.
References1. Prensner JR, RubinMA,Wei JT, ChinnaiyanAM.BeyondPSA: the next
generation of prostate cancer biomarkers. Sci Transl Med 2012;4:127rv3.
2. Wilke K, Gaul R, Klauck SM, Poustka A. A gene in human chromosomeband Xq28 (GABRE) defines a putative new subunit class of theGABAA neurotransmitter receptor. Genomics 1997;45:1–10.
Prognostic Value of GABRE�miR-452�miR-224 Methylation in Prostate Cancer
www.aacrjournals.org Clin Cancer Res; 20(8) April 15, 2014 2179
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

3. Weiss G, Cottrell S, Distler J, Schatz P, Kristiansen G, Ittmann M, et al.DNA methylation of the PITX2 gene promoter region is a strong inde-pendent prognostic marker of biochemical recurrence in patients withprostate cancer after radical prostatectomy. J Urol 2009;181:1678–85.
4. Banez LL, Sun L, van Leenders GJ, Wheeler TM, Bangma CH, Freed-land SJ, et al. Multicenter clinical validation of PITX2 methylation as aprostate specific antigen recurrence predictor in patients with post-radical prostatectomy prostate cancer. J Urol 2010;184:149–56.
5. GoeringW, KlothM, SchulzWA. DNAmethylation changes in prostatecancer. Methods Mol Biol 2012;863:47–66.
6. TrockBJ,BrotzmanMJ,MangoldLA,Bigley JW,Epstein JI,McLeodD,et al. Evaluation of GSTP1 and APC methylation as indicators forrepeat biopsy in a high-risk cohort of men with negative initial prostatebiopsies. BJU Int 2012;110:56–62.
7. Troyer DA, Lucia MS, de Bruine AP, Mendez-Meza R, Baldewijns MM,Dunscomb N, et al. Prostate cancer detected by methylated genemarkers in histopathologically cancer-negative tissues from men withsubsequent positive biopsies. Cancer Epidemiol Biomarkers Prev2009;18:2717–22.
8. D'Hulst C, Atack JR, Kooy RF. The complexity of the GABAA receptorshapes unique pharmacological profiles. Drug Discov Today 2009;14:866–75.
9. Neelands TR, Fisher JL, Bianchi M, Macdonald RL. Spontaneous andgamma-aminobutyric acid (GABA)-activated GABA(A) receptor chan-nels formed by epsilon subunit-containing isoforms. Mol Pharmacol1999;55:168–78.
10. AzumaH, Inamoto T,SakamotoT,KiyamaS,Ubai T, ShinoharaY, et al.Gamma-aminobutyric acid as apromoting factor of cancermetastasis;induction of matrix metalloproteinase production is potentially itsunderlying mechanism. Cancer Res 2003;63:8090–6.
11. Abdul M, McCray SD, Hoosein NM. Expression of gamma-aminobu-tyric acid receptor (subtype A) in prostate cancer. Acta Oncol2008;47:1546–50.
12. Drell TLt, Joseph J, Lang K, Niggemann B, Zaenker KS, Entschladen F.Effects of neurotransmitters on the chemokinesis and chemotaxis ofMDA-MB-468 humanbreast carcinoma cells. Breast Cancer Res Treat2003;80:63–70.
13. Jansson MD, Lund AH. MicroRNA and cancer. Mol Oncol 2012;6:590–610.
14. Mavridis K, StravodimosK,Scorilas A.Downregulation andprognosticperformance of microRNA 224 expression in prostate cancer. ClinChem 2013;59:261–9.
15. Martens-Uzunova ES, Jalava SE, Dits NF, van Leenders GJ, Moller S,Trapman J, et al. Diagnostic and prognostic signatures from the smallnon-coding RNA transcriptome in prostate cancer. Oncogene2012;31:978–91.
16. Prueitt RL, Yi M, Hudson RS, Wallace TA, Howe TM, Yfantis HG, et al.Expression of microRNAs and protein-coding genes associated withperineural invasion in prostate cancer. Prostate 2008;68:1152–64.
17. Liu C, Kelnar K, Vlassov AV, Brown D, Wang J, Tang DG. DistinctmicroRNA expression profiles in prostate cancer stem/progenitor cellsand tumor-suppressive functionsof let-7.CancerRes2012;72:3393–404.
18. Giricz O, Reynolds PA, Ramnauth A, LiuC,Wang T, Stead L, et al. Hsa-miR-375 is differentially expressed during breast lobular neoplasia andpromotes loss of mammary acinar polarity. J Pathol 2012;226:108–19.
19. Yanaihara N, Caplen N, Bowman E, Seike M, Kumamoto K, Yi M, et al.Unique microRNA molecular profiles in lung cancer diagnosis andprognosis. Cancer Cell 2006;9:189–98.
20. White NM, Chow TF, Mejia-Guerrero S, Diamandis M, Rofael Y,Faragalla H, et al. Three dysregulated miRNAs control kallikrein 10expression and cell proliferation in ovarian cancer. Br J Cancer2010;102:1244–53.
21. Gokhale A, Kunder R, Goel A, Sarin R, Moiyadi A, Shenoy A, et al.Distinctive microRNA signature of medulloblastomas associated withthe WNT signaling pathway. J Cancer Res Ther 2010;6:521–9.
22. Li Q, Wang G, Shan JL, Yang ZX, Wang HZ, Feng J, et al. MicroRNA-224 is upregulated inHepG2cells and involved in cellularmigration andinvasion. J Gastroenterol Hepatol 2010;25:164–71.
23. Boguslawska J, Wojcicka A, Piekielko-Witkowska A, Master A, Nau-man A. MiR-224 targets the 30UTR of type 1 50-iodothyronine deiodi-
nase possibly contributing to tissue hypothyroidism in renal cancer.PloS ONE 2011;6:e24541.
24. Arndt GM, Dossey L, Cullen LM, Lai A, Druker R, Eisbacher M, et al.Characterization of global microRNA expression reveals oncogenicpotential of miR-145 in metastatic colorectal cancer. BMC Cancer2009;9:374.
25. Liu H, Brannon AR, Reddy AR, Alexe G, Seiler MW, Arreola A, et al.Identifying mRNA targets of microRNA dysregulated in cancer: withapplication to clear cell Renal Cell Carcinoma. BMCSyst Biol 2010;4:51.
26. Veerla S, Lindgren D, Kvist A, Frigyesi A, Staaf J, Persson H, et al.MiRNA expression in urothelial carcinomas: important roles of miR-10a, miR-222, miR-125b, miR-7 and miR-452 for tumor stage andmetastasis, and frequent homozygous losses of miR-31. Int J Cancer2009;124:2236–42.
27. Vestergaard EM, Nexo E, Torring N, BorreM, Orntoft TF, Sorensen KD.Promoter hypomethylation and upregulation of trefoil factors in pros-tate cancer. Int J Cancer 2010;127:1857–65.
28. Sorensen KD, Abildgaard MO, Haldrup C, Ulhoi BP, Kristensen H,Strand S, et al. Prognostic significance of aberrantly silenced ANPEPexpression in prostate cancer. Br J Cancer 2013;108:420–8.
29. Haldrup C,Mundbjerg K, Vestergaard EM, Lamy P,Wild P, SchulzWA,et al. DNA methylation signatures for prediction of biochemical recur-rence after radical prostatectomy of clinically localized prostate can-cer. J Clin Oncol 2013;31:3250–8.
30. Taylor BS, Schultz N, Hieronymus H, Gopalan A, Xiao Y, Carver BS,et al. Integrative genomic profiling of human prostate cancer. CancerCell 2010;18:11–22.
31. Bert SA, RobinsonMD, Strbenac D, StathamAL, Song JZ, Hulf T, et al.Regional activation of the cancer genome by long-range epigeneticremodeling. Cancer Cell 2013;23:9–22.
32. Fuse M, Nohata N, Kojima S, Sakamoto S, Chiyomaru T, Kawakami K,et al. Restoration of miR-145 expression suppresses cell proliferation,migration and invasion in prostate cancer by targeting FSCN1. Int JOncol 2011;38:1093–101.
33. CaoP,DengZ,WanM,HuangW,Cramer SD, Xu J, et al.MicroRNA-101negatively regulates Ezh2 and its expression is modulated by androgenreceptor and HIF-1alpha/HIF-1beta. Mol Cancer 2010;9:108.
34. Zhang X, Zhang R, Zheng Y, Shen J, Xiao D, Li J, et al. Expression ofgamma-aminobutyric acid receptors on neoplastic growth and pre-diction of prognosis in non-small cell lung cancer. J Transl Med2013;11:102.
35. Huang L, Ting D, Lin X, Zhao XH, Chen XT, Wang CJ, et al. MicroRNA-224 targets RKIP to control cell invasion and expression of metastasisgenes in human breast cancer cells. Biochem Biophys Res Commun2012;24;425:127–33.
36. Li W, Wang X, Li B, Lu J, Chen G. Diagnostic significance of over-expression of Golgi membrane protein 1 in prostate cancer. Urology2012;80:952 e1–7.
37. Varambally S, LaxmanB,Mehra R, CaoQ, Dhanasekaran SM, TomlinsSA, et al. Golgi protein GOLM1 is a tissue and urine biomarker ofprostate cancer. Neoplasia 2008;10:1285–94.
38. Laxman B, Morris DS, Yu J, Siddiqui J, Cao J, Mehra R, et al. A first-generationmultiplex biomarker analysis of urine for the early detectionof prostate cancer. Cancer Res 2008;68:645–9.
39. Kristiansen G, Fritzsche FR, Wassermann K, Jager C, Tolls A, Lein M,et al. GOLPH2 protein expression as a novel tissue biomarker forprostate cancer: implications for tissue-based diagnostics. Br J Can-cer 2008;99:939–48.
40. Kojima S, Enokida H, Yoshino H, Itesako T, Chiyomaru T, Kinoshita T,et al. The tumor-suppressive microRNA-143/145 cluster inhibits cellmigration and invasion by targeting GOLM1 in prostate cancer. J HumGenet 2014;59:78–87.
41. SteketeeK, Ziel-van derMadeAC, vanderKorputHA,Houtsmuller AB,Trapman J. A bioinformatics-based functional analysis shows that thespecifically androgen-regulated gene SARG contains an active directrepeat androgen response element in the first intron. J Mol Endocrinol2004;33:477–91.
42. Zhao JJ, Yang J, Lin J, Yao N, Zhu Y, Zheng J, et al. Identification ofmiRNAs associated with tumorigenesis of retinoblastoma by miRNAmicroarray analysis. Childs Nerv Syst 2009;25:13–20.
Kristensen et al.
Clin Cancer Res; 20(8) April 15, 2014 Clinical Cancer Research2180
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

43. Archangelo LF, Greif PA, Holzel M, Harasim T, Kremmer E, PrzemeckGK, et al. The CALM and CALM/AF10 interactor CATS is a marker forproliferation. Mol Oncol 2008;2:356–67.
44. Geerts D, Koster J, Albert D, Koomoa DL, Feith DJ, Pegg AE, et al. Thepolyamine metabolism genes ornithine decarboxylase and antizyme 2predict aggressive behavior in neuroblastomas with and withoutMYCN amplification. Int J Cancer 2010;126:2012–24.
45. Bell JL, Wachter K, Muhleck B, Pazaitis N, Kohn M, Lederer M, et al.Insulin-like growth factor 2 mRNA-binding proteins (IGF2BPs): post-transcriptional drivers of cancer progression? Cell Mol Life Sci2013;70:2657–75.
46. Png KJ, Halberg N, YoshidaM, Tavazoie SF. AmicroRNA regulon thatmediates endothelial recruitment and metastasis by cancer cells.Nature 2012;481:190–4.
47. de Carvalho AV, Bonamigo RR, da Silva CM, Pinto AC. Positivity forHLA DR1 is associated with basal cell carcinoma in renal transplantpatients in southern Brazil. Int J Dermatol 2012;51:1448–53.
48. Jang SY, Jang SW, Ko J. Regulation of ADP-ribosylation factor 4expression by small leucine zipper protein and involvement in breastcancer cell migration. Cancer Lett 2012;314:185–97.
49. Woo IS, Eun SY, Jang HS, Kang ES, Kim GH, Kim HJ, et al.Identification of ADP-ribosylation factor 4 as a suppressor of N-(4-hydroxyphenyl)retinamide-induced cell death. Cancer Lett 2009;276:53–60.
50. Thorsen K, Sorensen KD, Brems-Eskildsen AS, Modin C, GaustadnesM, Hein AM, et al. Alternative splicing in colon, bladder, and prostatecancer identified by exon array analysis. Mol Cell Proteomics2008;7:1214–24.
www.aacrjournals.org Clin Cancer Res; 20(8) April 15, 2014 2181
Prognostic Value of GABRE�miR-452�miR-224 Methylation in Prostate Cancer
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from

2014;20:2169-2181. Clin Cancer Res Helle Kristensen, Christa Haldrup, Siri Strand, et al. ProstatectomyProstate Cancer Predicts Biochemical Recurrence after Radical
Promoter inmiR-224∼miR-452∼GABREHypermethylation of the
Updated version
http://clincancerres.aacrjournals.org/content/20/8/2169
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2015/07/17/20.8.2169.DC1.html
Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/20/8/2169.full.html#ref-list-1
This article cites 50 articles, 10 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/20/8/2169.full.html#related-urls
This article has been cited by 5 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications Department at
on March 6, 2016. © 2014 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from




















