
l-Arginine modulates CD3ζ expression and T cell function in activated human T lymphocytes
11
Cellular Immunology 232 (2004) 21–31 www.elsevier.com/locate/ycimm 0008-8749/$ - see front matter 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.cellimm.2005.01.004 Immunology ellular L-Arginine modulates CD3 expression and T cell function in activated human T lymphocytes Arnold H. Zea a,b,¤ , Paulo C. Rodriguez a,h , Kirk S. Culotta c , Claudia P. Hernandez a , Joanna DeSalvo d , Juan B. Ochoa e , Hae-Joon Park f , Jovanny Zabaleta g , Augusto C. Ochoa a,h a Stanley S. Scott Cancer Center, LSUHSC, New Orleans, LA, USA b Microbiology Immunology and Parasitology, LSUHSC, New Orleans, LA, USA c UT M.D. Anderson Cancer Center-PDC, Houston, TX, USA d Tulane University, New Orleans, LA, USA e Department of Surgery, University of Pittsburgh, Pittsburgh, PA, USA f MOGAM Biotechnology Research Institute, Yogin-City, Republic of Korea g Department of Pathology, LSUHSC, New Orleans, USA h Department of Pediatrics, LSUHSC, New Orleans, USA Received 1 September 2004; accepted 10 January 2005 Available online 23 February 2005 Abstract Engagement of the T cell receptor (TCR) by antigen or anti-CD3 antibody results in a cycle of internalization and re-expression of the CD3. Following internalization, CD3 is degraded and replaced by newly synthesized CD3 on the cell surface. Here, we provide evidence that availability of the amino acid L-arginine modulates the cycle of internalization and re-expression of CD3 and cause T cell dysfunction. T cells stimulated and cultured in presence of L-arginine, undergo the normal cycle of internalization and re-expression of CD3. In contrast, T cells stimulated and cultured in absence of L-arginine, present a sustained down-regula- tion of CD3 preventing the normal expression of the TCR, exhibit a decreased proliferation, and a signiWcantly diminished pro- duction of IFN, IL5, and IL10, but not IL2. The replenishment of L-arginine recovers the expression of CD3. The decreased expression of CD3 is not caused by a decreased CD3 mRNA, an increased CD3 degradation or T cell apoptosis. 2005 Elsevier Inc. All rights reserved. Keywords: CD3; L-Arginine; Lymphocytes; T cell function; T cell receptor; IL-2 1. Introduction L-Arginine is a semi-essential amino acid important in diVerent biological and metabolic systems including the immune system. In macrophages, L-arginine is metabolized by nitric oxide synthase (NOS) 1 to pro- duce nitric oxide (NO) [1,2] or by arginase to produce urea and ornithine, the latter being a precursor of poly- amines [3–5]. NO is one of the principal cytolytic mech- anisms in macrophages while polyamines are essential for cell proliferation [6–8]. Various conditions includ- ing liver transplantation and trauma, can lead to the depletion of L-arginine in vivo, resulting in a profound decrease in T cell function [9,10]. L-Arginine supple- mentation in these patients results in the recovery of normal T cell responses [11,12], suggesting that L-argi- nine may play an important role in regulating T cell function. ¤ Corresponding author: Fax: +1 504 599 0864. E-mail address: [email protected] (A.H. Zea). 1 Abbreviations used: C-RPMI, conventional RPMI; Arg-free-RP- MI, RPMI media without L-arginine; CD3, CD3 zeta chain; NO, ni- tric oxide; NOS, nitric oxide synthase; IL2R, interleukin 2 receptor.
Transcript of l-Arginine modulates CD3ζ expression and T cell function in activated human T lymphocytes

Cellular Immunology 232 (2004) 21–31
www.elsevier.com/locate/ycimm
Immunologyellular
L-Arginine modulates CD3� expression and T cell functionin activated human T lymphocytes
Arnold H. Zeaa,b,¤, Paulo C. Rodrigueza,h, Kirk S. Culottac, Claudia P. Hernandeza,Joanna DeSalvod, Juan B. Ochoae, Hae-Joon Parkf, Jovanny Zabaletag,
Augusto C. Ochoaa,h
a Stanley S. Scott Cancer Center, LSUHSC, New Orleans, LA, USAb Microbiology Immunology and Parasitology, LSUHSC, New Orleans, LA, USA
c UT M.D. Anderson Cancer Center-PDC, Houston, TX, USAd Tulane University, New Orleans, LA, USA
e Department of Surgery, University of Pittsburgh, Pittsburgh, PA, USAf MOGAM Biotechnology Research Institute, Yogin-City, Republic of Korea
g Department of Pathology, LSUHSC, New Orleans, USAh Department of Pediatrics, LSUHSC, New Orleans, USA
Received 1 September 2004; accepted 10 January 2005Available online 23 February 2005
Abstract
Engagement of the T cell receptor (TCR) by antigen or anti-CD3 antibody results in a cycle of internalization and re-expressionof the CD3�. Following internalization, CD3� is degraded and replaced by newly synthesized CD3� on the cell surface. Here, weprovide evidence that availability of the amino acid L-arginine modulates the cycle of internalization and re-expression of CD3�and cause T cell dysfunction. T cells stimulated and cultured in presence of L-arginine, undergo the normal cycle of internalizationand re-expression of CD3�. In contrast, T cells stimulated and cultured in absence of L-arginine, present a sustained down-regula-tion of CD3� preventing the normal expression of the TCR, exhibit a decreased proliferation, and a signiWcantly diminished pro-duction of IFN�, IL5, and IL10, but not IL2. The replenishment of L-arginine recovers the expression of CD3�. The decreasedexpression of CD3� is not caused by a decreased CD3� mRNA, an increased CD3� degradation or T cell apoptosis. 2005 Elsevier Inc. All rights reserved.
Keywords: CD3�; L-Arginine; Lymphocytes; T cell function; T cell receptor; IL-2
1. Introduction
L-Arginine is a semi-essential amino acid importantin diVerent biological and metabolic systems includingthe immune system. In macrophages, L-arginine ismetabolized by nitric oxide synthase (NOS)1 to pro-
¤ Corresponding author: Fax: +1 504 599 0864.E-mail address: [email protected] (A.H. Zea).
1 Abbreviations used: C-RPMI, conventional RPMI; Arg-free-RP-MI, RPMI media without L-arginine; CD3�, CD3 zeta chain; NO, ni-tric oxide; NOS, nitric oxide synthase; IL2R, interleukin 2 receptor.
0008-8749/$ - see front matter 2005 Elsevier Inc. All rights reserved.doi:10.1016/j.cellimm.2005.01.004
duce nitric oxide (NO) [1,2] or by arginase to produceurea and ornithine, the latter being a precursor of poly-amines [3–5]. NO is one of the principal cytolytic mech-anisms in macrophages while polyamines are essentialfor cell proliferation [6–8]. Various conditions includ-ing liver transplantation and trauma, can lead to thedepletion of L-arginine in vivo, resulting in a profounddecrease in T cell function [9,10]. L-Arginine supple-mentation in these patients results in the recovery ofnormal T cell responses [11,12], suggesting that L-argi-nine may play an important role in regulating T cellfunction.

22 A.H. Zea et al. / Cellular Immunology 232 (2004) 21–31
The T cell receptor � chain (CD3�) is the principalsignal transduction element of the TCR and is the rate-limiting step in its assembly and membrane expression[13,14]. The decreased expression of CD3� and adecreased in vitro response to antigens or mitogens hasbeen demonstrated in patients with cancer [15–19],chronic infectious diseases [20,21] and autoimmunity[22]. The mechanisms leading to the decreased expres-sion of CD3� and T cell dysfunction are poorly under-stood. We recently demonstrated that the depletion ofL-arginine from the tissue culture medium blocked pro-liferation of the Jurkat T cell line and induced the lossof CD3� due mostly to a decreased CD3� mRNA stabil-ity [23,24]. However, the eVect of L-arginine depletionon normal human T cells was signiWcantly diVerent. Thedata presented here demonstrates that L-arginine deple-tion does not aVect the expression of CD3� in resting Tcells. Instead, it appears to impair the cycle of internali-zation and re-expression of CD3� after antigen stimula-tion. The absence of L-arginine also blocked cytokineproduction and cell proliferation. However, this eVectwas not caused by T cell apoptosis a decreased expres-sion in the CD3� mRNA or to increased CD3� degrada-tion. Instead, preliminary data suggest that the absenceof L-arginine prevents the synthesis of new CD3�. Thechanges in signal transduction and T cell function werereversible by the replenishment of L arginine.
2. Materials and methods
2.1. T cell preparations
The peripheral blood mononuclear cells (PBMCs)from normal donors to be used for in vitro experimentswere purchased from the Blood Center of New Orleans.The use of samples from normal donors is currentlyapproved under the LSU-IRB protocols 4761 and 4867that are currently active. The PBMCs were separatedover Ficoll–Paque (Amersham Biosciences, Uppsala,Sweden) and passed through T cell enrichment columns(R&D Systems, Minneapolis, MN). After a 10 min incu-bation the columns were washed. The resulting enrichedT cells were counted and tested for surface markers byXow cytometry. The resulting T cell preparation con-tained >95% CD3+ (Clone HIT3a) cells, <3% CD16+
(Clone 3G8) cells, and <1% each of B (CD19+) (CloneHIB19) cells and monocytes (CD14+) (Clone ME52).
2.2. Tissue culture media and cell culture
Tissue culture dishes (100 mm) (Corning, Corning,NY) were coated overnight at 4 °C with PBS containing10 �g/ml of anti-CD3 (OKT-3 Ortho Pharmaceutical,Raritan, NJ) and then carefully washed once with coldphosphate-buVered saline (PBS). Ten million enriched
T cells were then added to each dish, resuspended in10 ml of conventional RPMI-1640 (C-RPMI) whichcontains 1140�M L-arginine (Cambrex, Walkersville,MD) or in RPMI-1640 without L-arginine (Arg-free-RPMI) (Gibco-Invitrogen, Carlsbad, CA) or RPMI-1640media without L-glutamine or L-glycine. Anti-CD28(Becton–Dickinson, San Jose, CA) at 100 ng/ml wasadded to each dish. Then the T cells were cultured forthe diVerent time periods at 37 °C in 5% CO2. As a neg-ative control, T cells were plated under the same mediaculture conditions but without OKT-3 and CD28. Allcultures were supplemented with 10% fetal calf serum(FCS) (Hyclone, Logan, UT), 25 mM Hepes buVer(Invitrogen, Carlsbad, CA), and 4 mM L-glutamine(Cambrex).
2.3. Flow cytometry
To measure CD3� expression, puriWed T cells wereincubated for 15 min at 4 °C with anti-CD3-FITC(Clone UCHT1) or the isotype control (Beckman-Coul-ter, Miami, FL) at 1 �g of antibody/106 cells. Cells werewashed and re-suspended in PBS containing digitonin at500 �g/ml (Wako, BioProducts, Richmond, VA) and2.5 �g of anti-CD3�-PE (Clone2H2D9) antibody (Beck-man-Coulter, Miami, FL). Cells were incubated for8 min at 4 °C, washed, and re-suspended in PBS for anal-ysis. For surface markers, 3 £ 105 T cells were plated on a96-well/U-bottom plates (Corning, Corning, NY) andincubated with 1 �g of isotype control, CD3 (CloneHIT3a), CD4 (Clone RPA-T4), CD8 (Clone HIT8a),CD14 (Clone M5E2), CD45RO (Clone UCHL1),CD45RA (Clone HI100), CD69 (Clone FN50) or CD25(Clone M-A251) antibodies (Becton–Dickinson, SanJose, CA) for 15 min at 4 °C, followed by two washeswith PBS containing 2% bovine serum albumin (BSA)(Sigma, St. Louis, MO), then Wxed in 1% paraformalde-hyde. For apoptosis, T cells were stained with Annexin VFITC apoptosis detection kit (Oncogene, San Diego,CA). Fluorescence analysis was done using a Coulter-EPICS Xow cytometer (Beckman-Coulter, Miami, FL).
2.4. Western blots and protein tyrosine phosphorylation
For Western blot, 10 £ 106 stimulated T cells wereharvested after culture at diVerent time points in thepresence or absence of L-arginine and lysed in TritonX-100 buVer with protease inhibitors as described before[20]. Lysates were electrophoresed in 14% Tris–glycinegels (Invitrogen, Carlsbad, CA), transfer to PVDF mem-branes (Invitrogen), immunoblotted with the diVerentantibodies and detected by horseradish peroxidase con-jugated antibodies and ECL (Amersham Biosciences,Little Chalfont, England). Gels were autoradiographedon X-OMAT AR Wlms (Eastman Kodak, Rochester,NY).

A.H. Zea et al. / Cellular Immunology 232 (2004) 21–31 23
For protein tyrosine phosphorylation, 5 £ 106 stimu-lated T cells were cultured in C-RPMI or in Arg-free-RPMI for 12, 24, and 48 h harvested and washed withcold PBS. As a negative control unstimulated T cellswere cultured in C-RPMI for the same time periods. Totrigger protein phosphorylation the cells were re-sus-pended in 200 �l of media, incubated with 1 �g of OKT-3(Ortho Pharmaceutical) on ice for 10 min, followed bycross-linking with 1.25 �g of goat anti-mouse (GAM)(Kirkegaard and Perry Labs., Gaithersburg, MD) for2 min at 37 °C. The reaction was stopped by washing thecells in cold PBS containing 400 �M sodium orthovana-date and 1 mM EDTA (Sigma, St. Louis, MO). The cellswere then lysed and electrophoresed in 10% polyacryl-amide gels, transferred to PVDF membranes and immu-noblotted with anti-phosphotyrosine 4G10-antibody(Upstate Biotechnology, Lake Placid, NY).
2.5. Calcium Xux
One million of stimulated T cells cultured in C-RPMIor in Arg-free-RPMI for 12 and 24 h were harvested andloaded with 2 mM Flou-3 AM (Molecular Probes,Eugene, OR) and 0.02% Pluronic 127F (MolecularProbes) for 30 min at 37 °C. The cells were then washedonce and incubated in calcium buVer (HBSS, 1 mMCaCl2, 1 mM MgCl2) for 20 min at room temperature inthe dark. Calcium Xux was measured at 488 nm as afunction of time (up to 450 s) in response to 1�g ofOKT-3 as a positive control or PBS as a negative control30 s after initial monitoring using a Coulter-EPICS Xowcytometer (Beckman-Coulter, Miami, FL). The Xowcytometry data were analyzed using WinMDI 2.8 soft-ware (TSRI, La Jolla, CA).
2.6. Cytokine production
Supernatants from the T cells cultures were collected at48, 72, and 96 h and tested for IL2, IFN�, IL5, and IL10production by ELISA. BrieXy, 96-well plates (ImmulonIV, Dynatech, Burlington, MA) were coated with therespective capture monoclonal antibody (Biosource, Cam-arillo, CA) and the culture supernatants incubated for30 min. The reaction was detected by biotin–streptavidinconjugated with horseradish peroxidase (BD-Pharmingen,San Diego, CA) using 3,3�,5,5�-tetramethylbenzidine(Roche, Indianapolis, IN) as a substrate. The reaction wasstopped with 0.8 M sulfuric acid and the absorbances wereread at 450 nm. The minimum level of cytokines detect-able by the assay was 30pg/ml.
2.7. Proliferation assay
Unstimulated or stimulated cells were plated at1 £ 105 per well and cultured in RPMI with or withoutarginine for 24, 48, and 72 h. [3H]Thymidine (0.5 �Ci)
(Perkin-Elmer Life Sciences, Boston, MA) was added toeach well and allowed to incubate for an additional 18 hat 37 °C. Each condition was tested in triplicate. Cellswere lysed by freezing and thawing, harvested onto aUniWlter-96 GF/B (Packard, Meriden, CT) and countedusing a TOPCOUNT Microplate Scintillation Counter(Packard, Meriden, CT).
2.8. Lysosome and proteasome inhibition
To inhibit lysosome function 2 £ 106 T cells culturedin the presence or absence of L-arginine for 24 h, weretreated with 1�M bafylomycin A1 (Calbiochem, SanDiego, CA). Similar treatment was done at 48 and 72 h inculture. To inhibit proteasome activity 2.5�M lactacy-stin (Calbiochem) was added to the cultures at the sametime points. DMSO was used as a control vehicle in allthe cases. The CD3� expression was tested by Xowcytometry 24 and 48 h after the addition of the inhibi-tors.
2.9. Isolation of RNA for Northern blots and ribonuclease protection assay
Total RNA was extracted from 107 T cells by lysiswith TRIzol (Invitrogen, Carlsbad, CA). Ten micro-grams of total RNA was electrophoresed under dena-turing conditions, blotted onto nytran membranes(Schleicher & Schuell, Keene, NH), and cross-linked byUV irradiation. Membranes were prehybridized at42 °C in ULTRAhyb buVer (Ambion, Austin, TX) andhybridized overnight with 1 £ 106 cpm/ml of 32P-labeled speciWc probes. Membranes were washed threetimes, and auto-radiographed at ¡70 °C using KodakBIOMAX-MR Wlms (Eastman Kodak, Rochester,NY). The murine cDNA glyceraldehyde-3 phosphatedehydrogenase (GAPDH) (Clontech, Palo Alto, CA)and the human CD3� (a kind gift from Dr. AllanWeissman, National Institutes of Health, Bethesda,MD) were labeled by random priming using a Redi-Prime Kit (Amersham Biosciences) and [�-32P]dCTP3,000 Ci/mmol (Perkin-Elmer Life Sciences, Boston,MA). All signal intensities were normalized toGAPDH.
For the ribonuclease protection assay (RPA), 5�g ofRNA was mixed with the templates (BD-Pharmingen,San Diego, CA) and incubated Wrst at 90 °C allowing thetemperature to decrease slowly to 56 °C. The sampleswere treated with RNAse followed by proteinase K.After extraction with phenol–chlorophorm, the samplewas precipitated in 100% ethanol for 30 min at ¡70 °Cand recovered by centrifugation at 12,000 rpm, resus-pended in loading buVer and separated on a polyacryl-amide gel containing 8 M Urea. Gels were dried andexposed to BIO-MAX Wlms (Eastman Kodak, Roches-ter, NY).
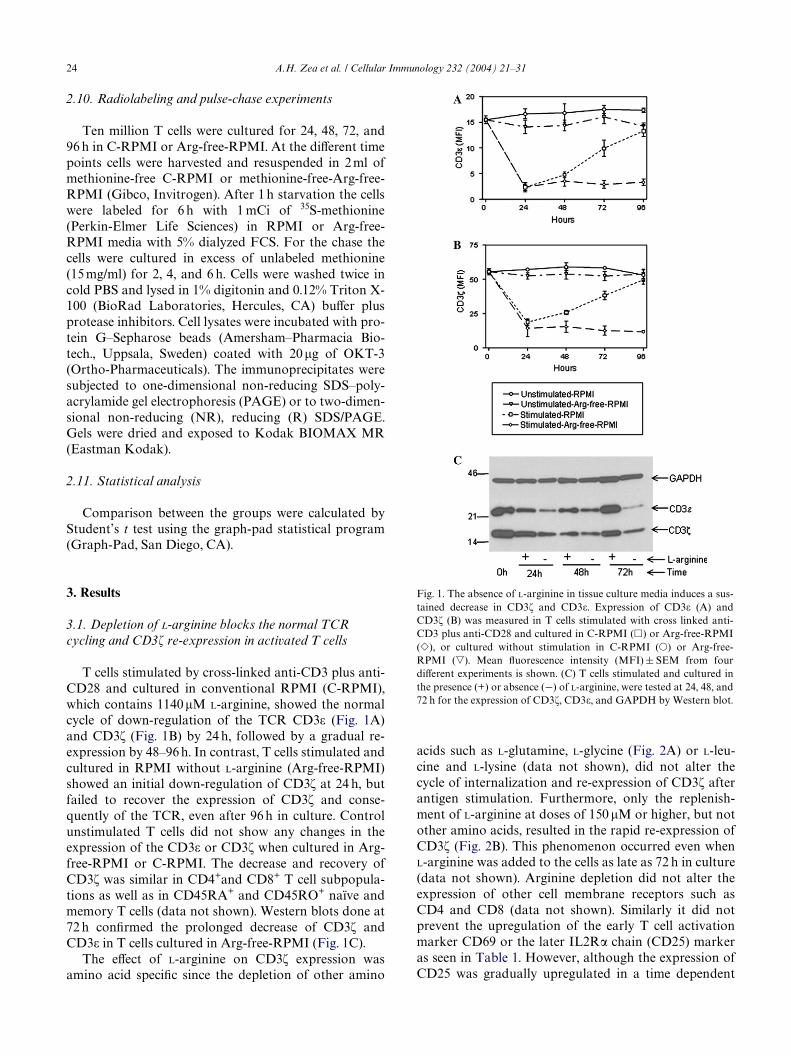
24 A.H. Zea et al. / Cellular Immunology 232 (2004) 21–31
2.10. Radiolabeling and pulse-chase experiments
Ten million T cells were cultured for 24, 48, 72, and96 h in C-RPMI or Arg-free-RPMI. At the diVerent timepoints cells were harvested and resuspended in 2 ml ofmethionine-free C-RPMI or methionine-free-Arg-free-RPMI (Gibco, Invitrogen). After 1 h starvation the cellswere labeled for 6 h with 1 mCi of 35S-methionine(Perkin-Elmer Life Sciences) in RPMI or Arg-free-RPMI media with 5% dialyzed FCS. For the chase thecells were cultured in excess of unlabeled methionine(15 mg/ml) for 2, 4, and 6 h. Cells were washed twice incold PBS and lysed in 1% digitonin and 0.12% Triton X-100 (BioRad Laboratories, Hercules, CA) buVer plusprotease inhibitors. Cell lysates were incubated with pro-tein G–Sepharose beads (Amersham–Pharmacia Bio-tech., Uppsala, Sweden) coated with 20�g of OKT-3(Ortho-Pharmaceuticals). The immunoprecipitates weresubjected to one-dimensional non-reducing SDS–poly-acrylamide gel electrophoresis (PAGE) or to two-dimen-sional non-reducing (NR), reducing (R) SDS/PAGE.Gels were dried and exposed to Kodak BIOMAX MR(Eastman Kodak).
2.11. Statistical analysis
Comparison between the groups were calculated byStudent’s t test using the graph-pad statistical program(Graph-Pad, San Diego, CA).
3. Results
3.1. Depletion of L-arginine blocks the normal TCR cycling and CD3� re-expression in activated T cells
T cells stimulated by cross-linked anti-CD3 plus anti-CD28 and cultured in conventional RPMI (C-RPMI),which contains 1140�M L-arginine, showed the normalcycle of down-regulation of the TCR CD3� (Fig. 1A)and CD3� (Fig. 1B) by 24 h, followed by a gradual re-expression by 48–96 h. In contrast, T cells stimulated andcultured in RPMI without L-arginine (Arg-free-RPMI)showed an initial down-regulation of CD3� at 24 h, butfailed to recover the expression of CD3� and conse-quently of the TCR, even after 96 h in culture. Controlunstimulated T cells did not show any changes in theexpression of the CD3� or CD3� when cultured in Arg-free-RPMI or C-RPMI. The decrease and recovery ofCD3� was similar in CD4+and CD8+ T cell subpopula-tions as well as in CD45RA+ and CD45RO+ naïve andmemory T cells (data not shown). Western blots done at72 h conWrmed the prolonged decrease of CD3� andCD3� in T cells cultured in Arg-free-RPMI (Fig. 1C).
The eVect of L-arginine on CD3� expression wasamino acid speciWc since the depletion of other amino
acids such as L-glutamine, L-glycine (Fig. 2A) or L-leu-cine and L-lysine (data not shown), did not alter thecycle of internalization and re-expression of CD3� afterantigen stimulation. Furthermore, only the replenish-ment of L-arginine at doses of 150 �M or higher, but notother amino acids, resulted in the rapid re-expression ofCD3� (Fig. 2B). This phenomenon occurred even whenL-arginine was added to the cells as late as 72 h in culture(data not shown). Arginine depletion did not alter theexpression of other cell membrane receptors such asCD4 and CD8 (data not shown). Similarly it did notprevent the upregulation of the early T cell activationmarker CD69 or the later IL2R� chain (CD25) markeras seen in Table 1. However, although the expression ofCD25 was gradually upregulated in a time dependent
Fig. 1. The absence of L-arginine in tissue culture media induces a sus-tained decrease in CD3� and CD3�. Expression of CD3� (A) andCD3� (B) was measured in T cells stimulated with cross linked anti-CD3 plus anti-CD28 and cultured in C-RPMI (�) or Arg-free-RPMI(�), or cultured without stimulation in C-RPMI (�) or Arg-free-RPMI (�). Mean Xuorescence intensity (MFI) § SEM from fourdiVerent experiments is shown. (C) T cells stimulated and cultured inthe presence (+) or absence (¡) of L-arginine, were tested at 24, 48, and72 h for the expression of CD3�, CD3�, and GAPDH by Western blot.
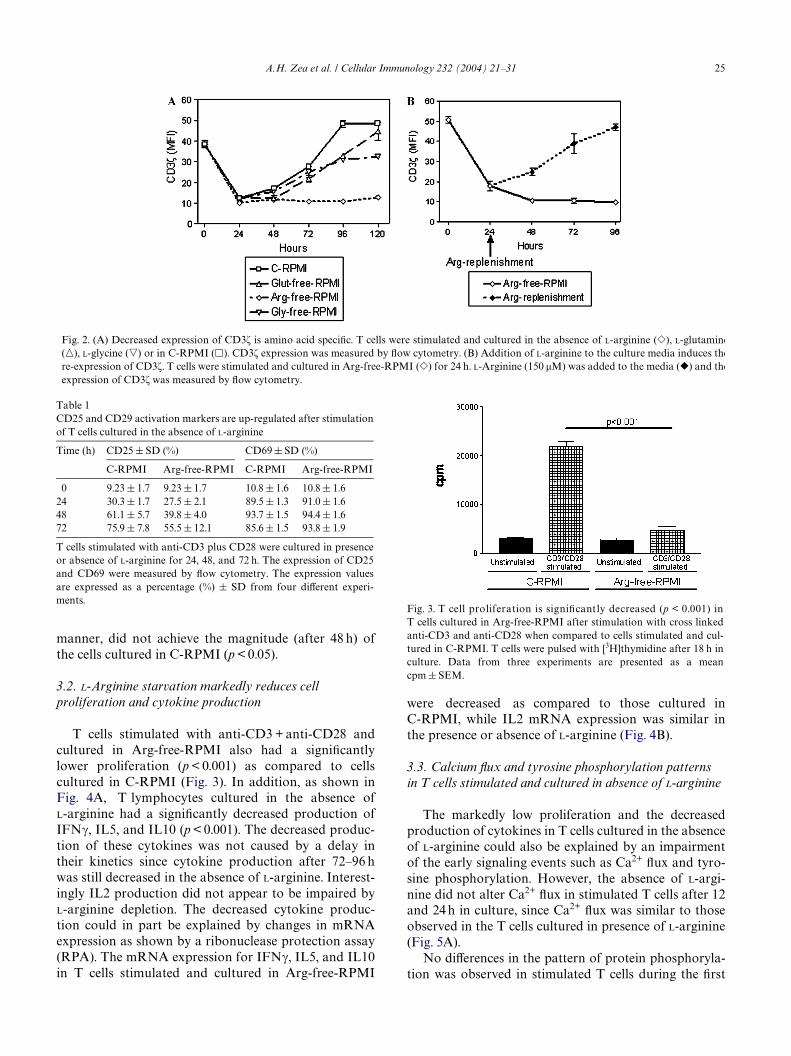
A.H. Zea et al. / Cellular Immunology 232 (2004) 21–31 25
manner, did not achieve the magnitude (after 48 h) ofthe cells cultured in C-RPMI (p < 0.05).
3.2. L-Arginine starvation markedly reduces cell proliferation and cytokine production
T cells stimulated with anti-CD3 + anti-CD28 andcultured in Arg-free-RPMI also had a signiWcantlylower proliferation (p < 0.001) as compared to cellscultured in C-RPMI (Fig. 3). In addition, as shown inFig. 4A, T lymphocytes cultured in the absence ofL-arginine had a signiWcantly decreased production ofIFN�, IL5, and IL10 (p < 0.001). The decreased produc-tion of these cytokines was not caused by a delay intheir kinetics since cytokine production after 72–96 hwas still decreased in the absence of L-arginine. Interest-ingly IL2 production did not appear to be impaired byL-arginine depletion. The decreased cytokine produc-tion could in part be explained by changes in mRNAexpression as shown by a ribonuclease protection assay(RPA). The mRNA expression for IFN�, IL5, and IL10in T cells stimulated and cultured in Arg-free-RPMI
Table 1CD25 and CD29 activation markers are up-regulated after stimulationof T cells cultured in the absence of L-arginine
T cells stimulated with anti-CD3 plus CD28 were cultured in presenceor absence of L-arginine for 24, 48, and 72 h. The expression of CD25and CD69 were measured by Xow cytometry. The expression valuesare expressed as a percentage (%) § SD from four diVerent experi-ments.
Time (h) CD25 § SD (%) CD69 § SD (%)
C-RPMI Arg-free-RPMI C-RPMI Arg-free-RPMI
0 9.23 § 1.7 9.23 § 1.7 10.8 § 1.6 10.8 § 1.624 30.3 § 1.7 27.5 § 2.1 89.5 § 1.3 91.0 § 1.648 61.1 § 5.7 39.8 § 4.0 93.7 § 1.5 94.4 § 1.672 75.9 § 7.8 55.5 § 12.1 85.6 § 1.5 93.8 § 1.9
were decreased as compared to those cultured inC-RPMI, while IL2 mRNA expression was similar inthe presence or absence of L-arginine (Fig. 4B).
3.3. Calcium Xux and tyrosine phosphorylation patterns in T cells stimulated and cultured in absence of L-arginine
The markedly low proliferation and the decreasedproduction of cytokines in T cells cultured in the absenceof L-arginine could also be explained by an impairmentof the early signaling events such as Ca2+ Xux and tyro-sine phosphorylation. However, the absence of L-argi-nine did not alter Ca2+ Xux in stimulated T cells after 12and 24 h in culture, since Ca2+ Xux was similar to thoseobserved in the T cells cultured in presence of L-arginine(Fig. 5A).
No diVerences in the pattern of protein phosphoryla-tion was observed in stimulated T cells during the Wrst
Fig. 3. T cell proliferation is signiWcantly decreased (p < 0.001) inT cells cultured in Arg-free-RPMI after stimulation with cross linkedanti-CD3 and anti-CD28 when compared to cells stimulated and cul-tured in C-RPMI. T cells were pulsed with [3H]thymidine after 18 h inculture. Data from three experiments are presented as a meancpm § SEM.
Fig. 2. (A) Decreased expression of CD3� is amino acid speciWc. T cells were stimulated and cultured in the absence of L-arginine (�), L-glutamine(�), L-glycine (�) or in C-RPMI (�). CD3� expression was measured by Xow cytometry. (B) Addition of L-arginine to the culture media induces there-expression of CD3�. T cells were stimulated and cultured in Arg-free-RPMI (�) for 24 h. L-Arginine (150 �M) was added to the media (�) and theexpression of CD3� was measured by Xow cytometry.
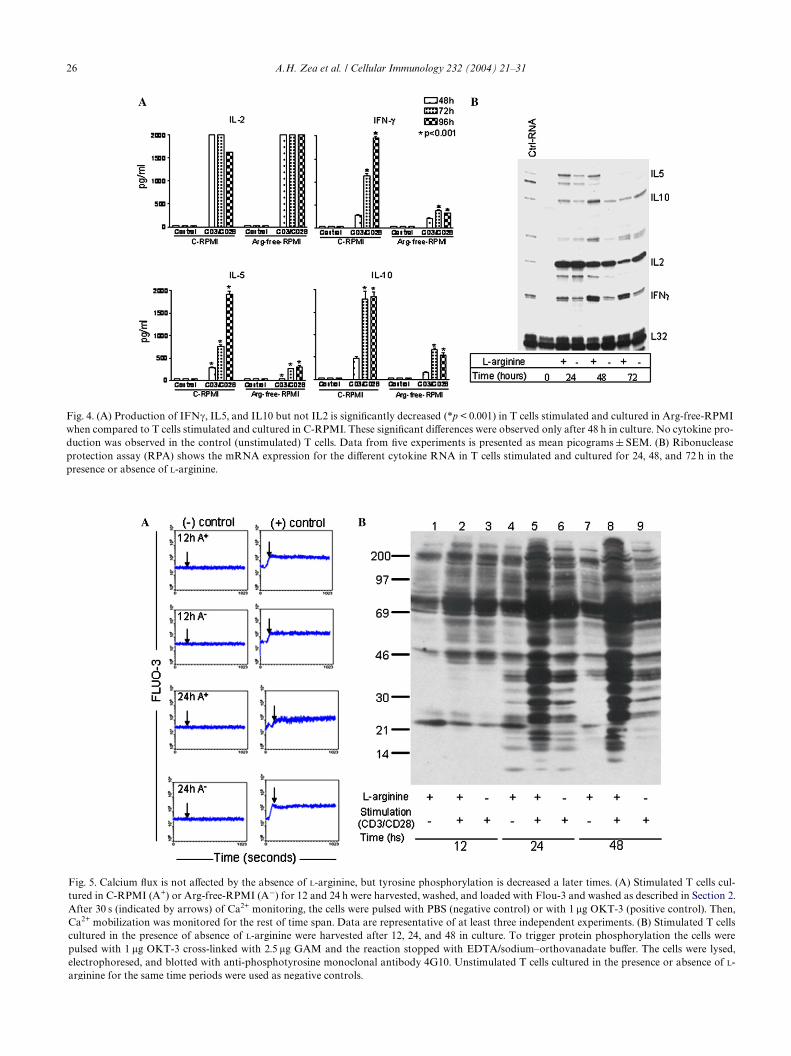
26 A.H. Zea et al. / Cellular Immunology 232 (2004) 21–31
arginine for the same time periods were used as negative controls.
Fig. 4. (A) Production of IFN�, IL5, and IL10 but not IL2 is signiWcantly decreased (*p < 0.001) in T cells stimulated and cultured in Arg-free-RPMIwhen compared to T cells stimulated and cultured in C-RPMI. These signiWcant diVerences were observed only after 48 h in culture. No cytokine pro-duction was observed in the control (unstimulated) T cells. Data from Wve experiments is presented as mean picograms § SEM. (B) Ribonucleaseprotection assay (RPA) shows the mRNA expression for the diVerent cytokine RNA in T cells stimulated and cultured for 24, 48, and 72 h in thepresence or absence of L-arginine.
Fig. 5. Calcium Xux is not aVected by the absence of L-arginine, but tyrosine phosphorylation is decreased a later times. (A) Stimulated T cells cul-tured in C-RPMI (A+) or Arg-free-RPMI (A¡) for 12 and 24 h were harvested, washed, and loaded with Flou-3 and washed as described in Section 2.After 30 s (indicated by arrows) of Ca2+ monitoring, the cells were pulsed with PBS (negative control) or with 1 �g OKT-3 (positive control). Then,Ca2+ mobilization was monitored for the rest of time span. Data are representative of at least three independent experiments. (B) Stimulated T cellscultured in the presence of absence of L-arginine were harvested after 12, 24, and 48 in culture. To trigger protein phosphorylation the cells werepulsed with 1 �g OKT-3 cross-linked with 2.5 �g GAM and the reaction stopped with EDTA/sodium–orthovanadate buVer. The cells were lysed,electrophoresed, and blotted with anti-phosphotyrosine monoclonal antibody 4G10. Unstimulated T cells cultured in the presence or absence of L-

A.H. Zea et al. / Cellular Immunology 232 (2004) 21–31 27
12 h of culture in C-RPMI (lane 2) or Arg-free-RPMI(lane 3). However, after 24 h in culture, the overall phos-phorylation patterns were markedly decreased in thecells cultured in Arg-free-RPMI (lane 6) as compared tothose seen in the T cells cultured in C-RPMI (lane 5).These diVerences in phosphorylation patterns were moreprominent after 48 hours in culture (C-RPMI lane 9 vs.Arg-free-RPMI lane 8). At the diVerent time points, theunstimulated T cells used as control presented a quitesimilar decreased phosphorylation patterns as observedin stimulated T cells cultured in absence of L-arginine(lanes 1, 4, and 7).
3.4. The decreased expression of CD3� it is not caused by apoptosis of T cells, an increased protein degradation or a decreased mRNA expression
The inability to re-express CD3� in T cells cultured inthe absence of L-arginine could be caused by severalmechanisms including an increased T cell apoptosis, anincrease in the degradation of CD3� protein, a decreasein CD3� mRNA expression or a decrease in CD3� chainsynthesis. Viability of T cells (by trypan blue exclusion)cultured in the absence of L-arginine was >92% over the96 h of culture in these experiments. Furthermore, apop-tosis, tested by annexin V expression was not increasedin T cells cultured in the absence of L-arginine (Table 2).We then tested whether an increased degradation ofCD3� could explain the inability to recover the expres-sion of this TCR chain. Experiments blocking lysosomeand proteasome function with baWlomycin and lactacy-stin respectively, failed to show the re-expression ofCD3� in T cells cultured in the absence of L-arginine(Fig. 6A). We then tested whether the inhibition of CD3�could be caused by a decrease in CD3� mRNA. North-ern blots showed no diVerences in CD3� mRNA expres-sion at 1, 12, 24, 48, and 72 h in culture (Fig. 6B), and didnot show a decrease in mRNA half life (data not shown).
We then tested the possibility that the absence ofL-arginine impaired the synthesis of CD3� or other com-ponents of the TCR. Fig. 7A shows pulse experiments of
Table 2The down-regulation of CD3� is not due to an increase in T cellapoptosis
Stimulated T cells were cultured in presence or absence of L-argininefor 24, 48, and 72 h. Annexin V-FITC apoptosis detection kit (Onco-gene, San Diego, CA) was used to measure apoptosis at the diVerenttime points of the experiments. The results are expressed as a percent-age (%) § SD of four diVerent experiments.
Time (h) Percentage § SD
C-RPMI Arg-free-RPMI
0 1.36 § 0.6 1.08 § 0.624 3.12 § 1.2 1.80 § 1.648 2.04 § 0.5 1.14 § 0.672 1.68 § 1.0 1.58 § 0.9
T cells labeled with 35S-methionine after being culturedin C-RPMI or Arg-free-RPMI for 24, 48, 72, and 96 h.CD3� and other TCR chains were isolated by immuno-precipitation with anti-CD3 monoclonal antibody or anirrelevant control antibody. Immunoprecipitates wereanalyzed by one-dimensional SDS–PAGE under nonreducing conditions or by two-dimensional SDS–PAGEunder non-reducing (NR), reducing (R) conditions asseen in Fig. 7B. T cells stimulated and cultured in C-RPMI normally synthesized CD3� and CD3�. Incontrast, T cells stimulated and cultured in the absenceof L-arginine, showed an inability to synthesize newCD3�, although the synthesis of CD3� was normal. The��, �, and � chains of the TCR were similarly synthe-sized in C-RPMI and Arg-free-RPMI. Chase experi-ments done in T cells cultured in C-RPMI showed a longhalf life of the newly synthesized CD3� and CD3�,respectively (data not shown). Therefore, initial datasuggests that the depletion of L-arginine may speciWcallyimpair the process of CD3� synthesis.
4. Discussion
L-Arginine is a semi-essential amino acid that playsan important role in the immune response. In macro-phages, L-arginine can be metabolized by the induciblenitric oxide synthase (iNOS) to produce nitric oxide(NO), important in the cytotoxic mechanism of thesecells [25], or by arginase I or arginase II to produce urea
Fig. 6. Decrease expression of CD3� in absence of L-arginine it is notdue to protein degradation or to a decreased mRNA expression. (A)EVect of lysosome (baWlomycin) and proteasome (lactasystin) inhibi-tors on T cells stimulated and cultured in C-RPMI or Arg-free-RPMIfor 24 h. CD3� expression was tested 24 and 48 h later by Xow cytome-try. (B) Stimulated T cells were cultured in conventional RPMI orArg-free-RPMI for 1, 12, 24, 48, and 72 h. Northern blot analysis wasused to test for CD3� RNA expression using 10 �g of total RNA.GAPDH was used as housekeeping gene.
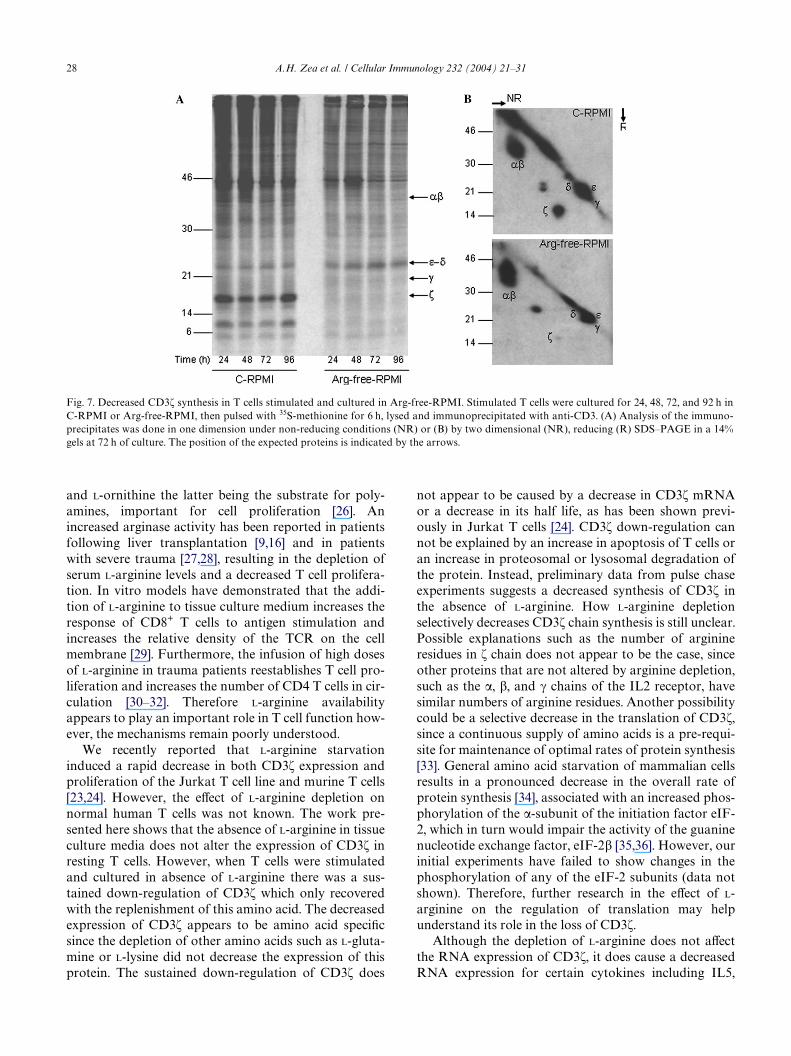
28 A.H. Zea et al. / Cellular Immunology 232 (2004) 21–31
and L-ornithine the latter being the substrate for poly-amines, important for cell proliferation [26]. Anincreased arginase activity has been reported in patientsfollowing liver transplantation [9,16] and in patientswith severe trauma [27,28], resulting in the depletion ofserum L-arginine levels and a decreased T cell prolifera-tion. In vitro models have demonstrated that the addi-tion of L-arginine to tissue culture medium increases theresponse of CD8+ T cells to antigen stimulation andincreases the relative density of the TCR on the cellmembrane [29]. Furthermore, the infusion of high dosesof L-arginine in trauma patients reestablishes T cell pro-liferation and increases the number of CD4 T cells in cir-culation [30–32]. Therefore L-arginine availabilityappears to play an important role in T cell function how-ever, the mechanisms remain poorly understood.
We recently reported that L-arginine starvationinduced a rapid decrease in both CD3� expression andproliferation of the Jurkat T cell line and murine T cells[23,24]. However, the eVect of L-arginine depletion onnormal human T cells was not known. The work pre-sented here shows that the absence of L-arginine in tissueculture media does not alter the expression of CD3� inresting T cells. However, when T cells were stimulatedand cultured in absence of L-arginine there was a sus-tained down-regulation of CD3� which only recoveredwith the replenishment of this amino acid. The decreasedexpression of CD3� appears to be amino acid speciWcsince the depletion of other amino acids such as L-gluta-mine or L-lysine did not decrease the expression of thisprotein. The sustained down-regulation of CD3� does
not appear to be caused by a decrease in CD3� mRNAor a decrease in its half life, as has been shown previ-ously in Jurkat T cells [24]. CD3� down-regulation cannot be explained by an increase in apoptosis of T cells oran increase in proteosomal or lysosomal degradation ofthe protein. Instead, preliminary data from pulse chaseexperiments suggests a decreased synthesis of CD3� inthe absence of L-arginine. How L-arginine depletionselectively decreases CD3� chain synthesis is still unclear.Possible explanations such as the number of arginineresidues in � chain does not appear to be the case, sinceother proteins that are not altered by arginine depletion,such as the �, �, and � chains of the IL2 receptor, havesimilar numbers of arginine residues. Another possibilitycould be a selective decrease in the translation of CD3�,since a continuous supply of amino acids is a pre-requi-site for maintenance of optimal rates of protein synthesis[33]. General amino acid starvation of mammalian cellsresults in a pronounced decrease in the overall rate ofprotein synthesis [34], associated with an increased phos-phorylation of the �-subunit of the initiation factor eIF-2, which in turn would impair the activity of the guaninenucleotide exchange factor, eIF-2� [35,36]. However, ourinitial experiments have failed to show changes in thephosphorylation of any of the eIF-2 subunits (data notshown). Therefore, further research in the eVect of L-arginine on the regulation of translation may helpunderstand its role in the loss of CD3�.
Although the depletion of L-arginine does not aVectthe RNA expression of CD3�, it does cause a decreasedRNA expression for certain cytokines including IL5,
Fig. 7. Decreased CD3� synthesis in T cells stimulated and cultured in Arg-free-RPMI. Stimulated T cells were cultured for 24, 48, 72, and 92 h inC-RPMI or Arg-free-RPMI, then pulsed with 35S-methionine for 6 h, lysed and immunoprecipitated with anti-CD3. (A) Analysis of the immuno-precipitates was done in one dimension under non-reducing conditions (NR) or (B) by two dimensional (NR), reducing (R) SDS–PAGE in a 14%gels at 72 h of culture. The position of the expected proteins is indicated by the arrows.

A.H. Zea et al. / Cellular Immunology 232 (2004) 21–31 29
IL10, and IFN�. The inability to up-regulate the genesfor these cytokines could in part be explained by thedecreased tyrosine phosphorylation observed in theabsence of L-arginine after 24 h. Interestingly, IL2 pro-duction was not aVected by the depletion of L-arginine.One way to explain this is that enough intracellular argi-nine allowed the activation of CD69 and the IL2 recep-tor � chain-CD25 that are synthesized and expressedsoon after activation of T cells. Similarly, early T cellfunctions such as Ca2+ Xux and tyrosine phosphoryla-tion were not aVected in the Wrst 24 h of culture. How-ever, phosphorylation patterns changed signiWcantlyafter 48 h in the absence of L-arginine. It is possible thatcertain cytokines genes such as IL2, which is expressedsoon after activation, is not impaired. Instead, othercytokines genes such as IL4, IL5, and IL10 may require alonger sustained signaling which is impaired after 48 h ofculture in the absence of L-arginine. It is also possiblethat the availability of amino acids regulates the tran-scription of cytokine genes [37–39]. Recent publicationshave shown that the depletion of the amino acid trypto-phan by cells producing indoleamine 2,3-dioxygenase(IDO) can severely impair T cell gene expression andfunction [40]. It is also possible that other amino acidsmay also play an important role in the regulation ofcytokine production. L-Glutamine enhances the produc-tion of IL2, IL10, and IFN� in T lymphocytes [41,42]and a recent study demonstrated that adequate concen-trations of glutamine increase a Th1 responses [43].However, the eVect seen here is caused exclusively by thedepletion of L-arginine since glutamine was present in allcultures. It is diYcult to determine with the resultsshown here whether or not there is a link between theabsence of L-arginine and their eVect on cytokine pro-duction as an independent eVect on CD3� synthesis.More research targeting these questions need to be done.
The Wndings described here could be important inexplaining the diminished expression of CD3� and otherT cell signal transduction proteins described in patientswith cancer [16,17,44,45], chronic infections [20,21] andmore recently in trauma [46]. The similarity in T cell sig-nal transduction alterations in diseases with diVerentpathophysiology suggests the possibility of a commonmechanism as the cause for such changes. Several mech-anisms have been postulated to cause a decrease inCD3� chain expression in cancer, including Fas–FasLinteractions [47,48], the production of H2O2 by macro-phages and neutrophils [49,50] and more recently thedepletion of L-arginine by macrophages [51]. L-Argininelevels are regulated in vivo by dietary intake and bythree enzymatic pathways in macrophages namelyiNOS, arginase I or II. Increased arginase activity, withthe depletion of L-arginine and an impaired T cell func-tion has been reported in clinical conditions such assevere trauma [27] or following liver transplantation[9,52]. Increased arginase production has been reported
in colon and gastric cancer [53,54] some leukemias [55]and in chronic infections such as leprosy [56]. Therefore,it is possible that the depletion of L-arginine in themicroenvironment (at the tumor or sites of infection) orsystemically (in metastatic disease) may decrease theexpression of speciWc signal transduction proteins in Tcells that are activated by antigen, resulting in animpaired T cell response. Despite the diVerent patho-physiology of these diseases the regulation of L-arginineavailability could be a common mechanism responsiblefor immune dysfunction. Additional clinical researchwill test this possibility and determine its impact on theoutcome of disease and the potential for the develop-ment of new therapies that may overcome them.
Acknowledgments
This work was supported in part by Grants RO1-CA82689, RO1-CA88885, and R21-CA83198 from theNational Cancer Institute, National Institutes of Health-(NIH-NCI), Bethesda MD. We thank Drs. MichaelHagensee, Diego Aviles, Alison Quayle, and ImtiazKhan for their comments to this paper. We are gratefulMr. Kevin Zwezdaryk for his technical assistance as wellas Ms. Sandra Lee and Ms. Nicole Barron for their assis-tance in preparing the manuscript.
References
[1] G. Wu, S.M. Morris, Arginine metabolism: Nitric oxide andbeyond, Biochem. J. 336 (1998) 1–17.
[2] J.E. Albina, M.D. Caldwell, W.L. Henry, C.D. Mills, Regulation ofmacrophage functions by L-arginine, J. Exp. Med. 169 (1989)1021–1029.
[3] C.I. Chang, J.C. Liao, L. Kuo, Arginase modulates nitric oxideproduction in activated macrophages, Am. J. Physiol. 274 (1998)H342–H348.
[4] S. Moncada, R.M. Palmer, E.A. Higgs, Biosynthesis of nitric oxidefrom L-arginine, a pathway for the regulation of cell function andcommunication, Biochem. Pharmacol. 38 (1989) 1709–1715.
[5] C.W. Tabor, H. Tabor, Polyamines, Annu. Rev. Biochem. 53(1984) 749–790.
[6] J.C. Chuang, C.L. Yu, S.R. Wang, Modulation of lymphocyte pro-liferation by enzymes that degrade amino acids, Clin. Exp. Immu-nol. 82 (1990) 469–472.
[7] D.T. Efron, S.J. Kirk, M.C. Regan, H.L. Wasserkrug, A. Barbul,Nitric oxide generation from L-arginine is required for optimalhuman peripheral blood lymphocyte DNA synthesis, Surgery 110(1991) 327–334.
[8] M. Hurson, M.C. Regan, S.J. Kirk, H.L. Wasserkrug, A. Barbul,Metabolic eVects of arginine in a healthy elderly population, J.Parenter. Enteral Nutr. 19 (1995) 227–230.
[9] E. Roth, R. Steininger, S. Winkler, F. Langle, T. Grunberger, R.Fugger, F. Muhlbacher, L-arginine deWciency after liver transplan-tation as an eVect of arginase eZux from the graft. InXuence onnitric oxide metabolism, Transplantation 57 (1994) 665–669.
[10] J.B. Ochoa, A.O. Udekwu, T.R. Billiar, R.D. Curran, F.B. Cerra,R.L. Simmons, A.B. Peitzman, Nitrogen oxide levels in patientsafter trauma and during sepsis, Ann. Surg. 214 (1991) 621–626.

30 A.H. Zea et al. / Cellular Immunology 232 (2004) 21–31
[11] A. Barbul, H.L. Wasserkrug, N. Yoshimura, R. Tao, G. Efron, Higharginine levels in intravenous hyperalimentation abrogate post-traumatic immune suppression, J. Surg. Res. 36 (1984) 620–624.
[12] S.J. Kirk, A. Barbul, Role of arginine in trauma, sepsis, and immu-nity, J. Parenter. Enteral Nutr. 14 (1990) S226–S229.
[13] Y. Minami, A.M. Weissman, L.E. Samelson, R.D. Klausner, Build-ing a multichain receptor: Synthesis, degradation, and assembly ofthe T-cell antigen receptor, Proc. Natl. Acad. Sci. USA 84 (1987)2688–2692.
[14] A.M. Weissman, P. Ross, E.T. Luong, P. Garcia-Morales, M.L.Jelachich, W.E. Biddison, R.D. Klausner, L.E. Samelson, Tyrosinephosphorylation of the human T cell antigen receptor zeta-chain:Activation via CD3 but not CD2, J. Immunol. 141 (1988) 3532–3536.
[15] Y. Gunji, S. Hori, T. Aoe, T. Asano, T. Ochiai, K. Isono, T. Saito,High frequency of cancer patients with abnormal assembly of theT cell receptor-CD3 complex in peripheral blood T lymphocytes,Jpn. J. Cancer Res. 85 (1994) 1189–1192.
[16] J.H. Finke, A.H. Zea, J. Stanley, D.L. Longo, H. Mizoguchi, R.R.Tubbs, R.H. Wiltrout, J.J. O’Shea, S. Kudoh, E. Klein, A.C.Ochoa, Loss of T-cell receptor zeta chain and p56lck in T-cellsinWltrating human renal cell carcinoma, Cancer Res. 53 (1993)5613–5616.
[17] C.G. Healy, J.W. Simons, M.A. Carducci, T.L. DeWeese, M. Bart-kowski, K.P. Tong, W.E. Bolton, Impaired expression and func-tion of signal-transducing zeta chains in peripheral T cells andnatural killer cells in patients with prostate cancer, Cytometry 32(1998) 109–119.
[18] H. Rabinowich, Y. Suminami, T.E. Reichert, P. Crowley-Nowick,M. Bell, R. Edwards, T.L. Whiteside, Expression of cytokine genesor proteins and signaling molecules in lymphocytes associatedwith human ovarian carcinoma, Int. J. Cancer 68 (1996) 276–284.
[19] E. Rossi, E. Matutes, R. Morilla, K. Owusu-Ankomah, A.M.HeVernan, D. Catovsky, Zeta chain and CD28 are poorlyexpressed on T lymphocytes from chronic lymphocytic leukemia,Leukemia 10 (1996) 494–497.
[20] A.H. Zea, M.T. Ochoa, P. Ghosh, D.L. Longo, W.G. Alvord, L.Valderrama, R. Falabella, L.K. Harvey, N. Saravia, L.H. Moreno,A.C. Ochoa, Changes in expression of signal transduction proteinsin T lymphocytes of patients with leprosy, Infect. Immun. 66(1998) 499–504.
[21] I. Stefanova, M.W. Saville, C. Peters, F.R. Cleghorn, D. Schwartz,D.J. Venzon, K.J. Weinhold, N. Jack, C. Bartholomew, W.A. Blatt-ner, R. Yarchoan, J.B. Bolen, I.D. Horak, HIV infection-inducedposttranslational modiWcation of T cell signaling molecules asso-ciated with disease progression, J. Clin. Invest. 98 (1996) 1290–1297.
[22] S.N. Liossis, X.Z. Ding, G.J. Dennis, G.C. Tsokos, Altered patternof TCR/CD3-mediated protein-tyrosyl phosphorylation in T cellsfrom patients with systemic lupus erythematosus. DeWcientexpression of the T cell receptor zeta chain, J. Clin. Invest. 101(1998) 1448–1457.
[23] F. Taheri, J.B. Ochoa, Z. Faghiri, K. Culotta, H.J. Park, M.S. Lan,A.H. Zea, A.C. Ochoa, L-arginine regulates the expression of theT-cell receptor zeta chain (CD3zeta) in Jurkat cells, Clin. CancerRes. 7 (2001) 958s–965s.
[24] P.C. Rodriguez, A.H. Zea, K.S. Culotta, J. Zabaleta, J.B. Ochoa,A.C. Ochoa, Regulation of T cell receptor CD3zeta chain expres-sion by L-arginine, J. Biol. Chem. 277 (2002) 21123–21129.
[25] P.K Lala, SigniWcance of nitric oxide in carcinogenesis, tumor pro-gression and cancer therapy, Cancer Metastasis Rev. 17 (1998) 91–106.
[26] T. Gotoh, T. Sonoki, A. Nagasaki, K. Terada, M. Takiguchi, M.Mori, Molecular cloning of cDNA for nonhepatic mitochondrialarginase (arginase II) and comparison of its induction with nitricoxide synthase in a murine macrophage-like cell line, FEBS Lett.395 (1966) 119–122.
[27] A.C. Bernard, S.K. Mistry, S.M. Morris Jr., W.E. O’Brien, B.J.Tsuei, M.E. Maley, L.A. Shirley, P.A. Kearney, B.R. Boulanger,J.B. Ochoa, Alterations in arginine metabolic enzymes in trauma,Shock 15 (2001) 215–219.
[28] M.K. Angele, N. Smail, A. Ayala, W.G. CioY, K.I. Bland, I.H.Chaudry, L-Arginine: A unique amino acid for restoring thedepressed macrophage functions after trauma-hemorrhage, J.Trauma 46 (1999) 34–41.
[29] J.B. Ochoa, J. Strange, P. Kearney, G. Gellin, E. Endean, E. Fitz-patrick, EVects of L-arginine on the proliferation of T lymphocytesubpopulations, J. Parenter. Enteral Nutr. 25 (2001) 23–29.
[30] A. Barbul, S.A. Lazarou, D.T. Efron, H.L. Wasserkrug, G. Efron,Arginine enhances wound healing and lymphocyte immuneresponses in humans, Surgery 108 (1990) 331–336.
[31] J. Brittenden, S.D. Heys, J. Ross, K.G. Park, O. Eremin, Nutri-tional pharmacology: EVects of L-arginine on host defences,response to trauma and tumour growth, Clin. Sci. 86 (1994) 123–132.
[32] R.J. Beale, D.J. Bryg, D.J. Bihari, Immunonutrition in the criticallyill: A systematic review of clinical outcome, Crit. Care Med. 27(1999) 2799–2805.
[33] S.R. Kimball, Regulation of global and speciWc mRNA transla-tion by amino acids, J. Nutr. 132 (2002) 883–886.
[34] S.R. Kimball, L.S. JeVerson, Control of protein synthesis by aminoacid availability, Curr. Opin. Clin. Nutr. Metab. Care 5 (2002) 63–67.
[35] V.M. Pain, Translational control during amino acid starvation,Biochimie 76 (1994) 718–728.
[36] L.S. JeVerson, S.R. Kimball, Amino acid regulation of geneexpression, J. Nutr. 131 (2001) 2460S–2466S.
[37] P. Fafournoux, A. Bruhat, C. Jousse, Amino acid regulation ofgene expression, Biochem. J. 351 (2000) 1–12.
[38] A.L. Mellor, D.B. Keskin, T. Johnson, P. Chandler, D.H. Munn,Cells expressing indoleamine 2,3-dioxygenase inhibit T cellresponses, J. Immunol. 168 (2002) 3771–3776.
[39] G.K. Lee, H.J. Park, M. Macleod, P. Chandler, D.H. Munn, A.L.Mellor, Tryptophan deprivation sensitizes activated T cells toapoptosis prior to cell division, Immunology 107 (2002) 452–460.
[40] D.H. Munn, E. ShaWzadeh, J.T. Attwood, I. Bondarev, A. Pashine,A.L. Mellor, Inhibition of T cell proliferation by macrophagetryptophan catabolism, J. Exp. Med. 189 (1999) 1363–1372.
[41] P. Yaqoob, P.C. Calder, Glutamine requirement of proliferating Tlymphocytes, Nutrition 13 (1997) 646–651.
[42] P. Yaqoob, P.C. Calder, Cytokine production by human periphe-ral blood mononuclear cells: DiVerential sensitivity to glutamineavailability, Cytokine 10 (1998) 790–794.
[43] W.K. Chang, K.D. Yang, M.F. Shaio, EVect of glutamine on Th1and Th2 cytokine responses of human peripheral blood mononu-clear cells, Clin. Immunol. 93 (1999) 294–301.
[44] A.H. Zea, B.D. Curti, D.L. Longo, W.G. Alvord, S.L. Strobl, H.Mizoguchi, S.P. Creekmore, J.J. O’Shea, G.C. Powers, W.J. Urba,A.C. Ochoa, Alterations in T cell receptor and signal transductionmolecules in melanoma patients, Clin. Cancer Res. 1 (1995) 1327–1335.
[45] M. Massaia, C. Attisano, E. Beggiato, A. Bianchi, A. Pileri, Cor-relation between disease activity and T-cell CD3 zeta chainexpression in a B-cell lymphoma, Br. J. Haematol. 88 (1994) 886–888.
[46] F. Ichihara, K. Kono, T. Sekikawa, Y. Matsumoto, Surgical stressinduces decreased expression of signal-transducing zeta moleculesin T cells, Eur. Surg. Res. 31 (1999) 138–146.
[47] B.R. Gastman, D.E. Johnson, T.L. Whiteside, H. Rabinowich,Tumor-induced apoptosis of T lymphocytes: Elucidation of intra-cellular apoptotic events, Blood 95 (2000) 2015–2023.
[48] B.R. Gastman, Y. Atarshi, T.E. Reichert, T. Saito, L. Balkir, H.Rabinowich, T.L. Whiteside, Fas ligand is expressed on human

A.H. Zea et al. / Cellular Immunology 232 (2004) 21–31 31
squamous cell carcinomas of the head and neck, and it promotesapoptosis of T lymphocytes, Cancer Res. 59 (1999) 5356–5364.
[49] M. Otsuji, Y. Kimura, T. Aoe, Y. Okamoto, T. Saito, Oxidativestress by tumor-derived macrophages suppresses the expression ofCD3 zeta chain of T-cell receptor complex and antigen-speciWc T-cell responses, Proc. Natl. Acad. Sci. 93 (1966) 13119–13124.
[50] J. Schmielau, O.J. Finn, Activated granulocytes and granulocyte-derived hydrogen peroxide are the underlying mechanism of sup-pression of T-cell function in advanced cancer patients, CancerRes. 61 (2001) 4756–4760.
[51] V. Bronte, P. SeraWni, C. De Santo, I. Marigo, V. Tosello, A. Mazz-oni, D.M. Segal, C. Staib, M. Lowel, G. Sutter, M.P. Colombo, P.Zanovello, IL-4-induced arginase 1 suppresses alloreactive T cellsin tumor-bearing mice, J. Immunol. 170 (2003) 270–278.
[52] M. Ikemoto, S. Tsunekawa, K. Tanaka, A. Tanaka, Y. Yamaoka,K. Ozawa, Y. Fukuda, F. Moriyasu, M. Totani, Y. Kasai, T. Mori,
K. Ueda, Liver-type arginase in serum during and after liver trans-plantation: A novel index in monitoring conditions of the livergraft and its clinical signiWcance, Clin. Chim. Acta 271 (1998)11–23.
[53] S.Y. Leu, S.R. Wang, Clinical signiWcance of arginase in colorectalcancer, Cancer 70 (1992) 733–736.
[54] C.W. Wu, W.W. Chung, C.W. Chi, H.L. Kao, W.Y. Lui, F.K. Peng,S.R. Wang, Immunohistochemical study of arginase in cancer ofthe stomach, Virchows Arch. 428 (1996) 325–331.
[55] L. Konarska, I. Widzynska, H. Zienkiewicz, K. Sulek, Arginaseactivity alterations in peripheral blood lymphocytes in the humanchronic lymphocytic leukemia, Acta Biochim. Pol. 40 (1993) 160–163.
[56] C.S. Babu, K.B. Kannan, V.P. Bharadwaj, V.M. Katoch, Lympho-cyte arginase activity in leprosy—a preliminary report, Indian J.Med. Res. 91 (1990) 193–196.




















