
Chronic overexpression of membrane-bound flt3 ligand by T lymphocytes in severe aplastic anaemia
10
British Journal of Haematology 2000, 109, 211–220 Chronic overexpression of membrane-bound flt3 ligand by T lymphocytes in severe aplastic anaemia O TMAR P FISTER , 1 E LENA C HKLOVSKAIA, 1 WENDY J ANSEN, 1 K INGA ME ´ SZA ´ ROS , 1 C ATHERINE N ISSEN, 1 C HRISTOPH R AHNER , 2 N INA H URWITZ , 3 NATALIA B OGATCHEVA, 4 S TEWART D. LYMAN 5 and A LEKSANDRA WODNAR- F ILIPOWICZ 11 Department of Research, University Hospital Basle, Switzerland, 2 Institute of Anatomy, University of Basle, Switzerland, 3 Institute of Pathology, University Hospital Basle, Switzerland, 4 Institute of Paediatric Haematology, Moscow, Russia, and 5 Immunex R & D Seattle, WA, USA Received 7 October 1999; accepted for publication 29 December 1999 Summary. Aplastic anaemia (AA) is an immune-mediated bone marrow failure associated with high serum levels of flt3 ligand (FL). We examined expression of the membrane- bound isoform of FL in peripheral blood and bone marrow cells from AA patients at diagnosis (n 16) and after immunosuppressive (IS) treatment (n 36). Flow cytome- try demonstrated strongly increased FL levels on the cell surface of T lymphocytes in AA relative to normal controls (P , 0·0001). T-cell-specific expression of membrane- bound FL was confirmed by confocal microscopy. FL mRNA and total cellular FL protein levels were increased about threefold. Overexpression of FL in AA was observed for up to 20 years after IS treatment. FL levels correlated inversely with CD34 1 cell numbers and the colony-forming ability of AA bone marrow (R 20·68 and 20·85 respectively). Histological examination of spleen specimens and bone marrow biopsies gave no evidence of degeneration or fibrosis due to prolonged exposure to high FL. Levels of membrane-bound FL were not increased in autoimmune diseases (n 23), including rheumatoid arthritis and lupus erythematosus, nor in graft-versus-host disease (n 8). Chronic overexpression of FL on the surface of T lympho- cytes in AA, but not in other T-cell-mediated disorders, suggests that membrane-bound FL plays a role in cell–cell interactions in bone marrow failure and may be important for long-term haemopoietic recovery. Keywords: aplastic anaemia, flt3 ligand, T lymphocytes, membrane-bound cytokines, bone marrow and spleen fibrosis. Flt3 ligand (FL) is a haemopoietic growth factor specific for a tyrosine kinase receptor flt3 (flt3R; Lyman et al, 1993; Hannum et al, 1994). This receptor is structurally related to c-kit, the receptor for stem cell factor (SCF), and c-fms, the receptor for macrophage colony-stimulating factor (M-CSF). Signalling through flt3R is important in very early human haemopoiesis, as indicated by the restricted pattern of receptor expression found predominantly on primitive haemopoietic precursors (Matthews et al, 1991; Rosnet et al, 1993). Accordingly, FL stimulates proliferation and differentiation of most primitive lineage-negative popula- tions of CD34 1 progenitor cells from human cord blood, peripheral blood (PB), bone marrow (BM) and fetal liver (reviewed in Lyman & Jacobsen, 1998). Like several other haemopoietic cytokines, including SCF and M-CSF, FL exists in membrane-bound and soluble forms. Analysis of the alternatively spliced FL transcripts in human cells revealed an abundance of the species encoding membrane-bound molecules (Lyman et al, 1994), and pulse-chase studies demonstrated that these primary products undergo subse- quent processing to yield soluble FL (McClanahan et al, 1996). Biological properties of FL are recognized by the activity of its soluble form, whereas the function of membrane-bound FL is not known. In analogy to SCF, of which the membrane-bound form is essential for normal haemopoietic development in the mouse (Flanagan et al, 1991) and is more active than soluble SCF in stroma- supported haemopoiesis in vitro (Toksoz et al, 1992; Slanicka Krieger et al, 1998), membrane-bound FL may play an equally important role in cell–cell interactions within the early progenitor cell compartment. Although experimental results in vitro and in vivo have 211 q 2000 Blackwell Science Ltd Correspondence: Aleksandra Wodnar-Filipowicz, Department of Research, University Hospital Basel, Hebelstr. 20, CH-4031 Basel, Switzerland. E-mail: [email protected]
Transcript of Chronic overexpression of membrane-bound flt3 ligand by T lymphocytes in severe aplastic anaemia

British Journal of Haematology 2000, 109, 211±220
Chronic overexpression of membrane-bound flt3 ligand by
T lymphocytes in severe aplastic anaemia
OTMAR PFIS TER,1 ELENA CHKLOVSKAIA,1 WENDY JANSEN,1 KINGA MEÂ SZAÂ ROS,1 CATHERIN E NISSEN,1
CHRISTOPH RAHNER,2 NINA HURW ITZ,3 NATA LIA BOGATCHEVA,4 STEWART D. LYMAN5 and
ALEKS ANDRA WO DNA R -FILIPOW ICZ1 1Department of Research, University Hospital Basle, Switzerland, 2Institute of
Anatomy, University of Basle, Switzerland, 3Institute of Pathology, University Hospital Basle, Switzerland, 4Institute of
Paediatric Haematology, Moscow, Russia, and 5Immunex R & D Seattle, WA, USA
Received 7 October 1999; accepted for publication 29 December 1999
Summary. Aplastic anaemia (AA) is an immune-mediatedbone marrow failure associated with high serum levels offlt3 ligand (FL). We examined expression of the membrane-bound isoform of FL in peripheral blood and bone marrowcells from AA patients at diagnosis (n � 16) and afterimmunosuppressive (IS) treatment (n � 36). Flow cytome-try demonstrated strongly increased FL levels on the cellsurface of T lymphocytes in AA relative to normal controls(P , 0´0001). T-cell-specific expression of membrane-bound FL was confirmed by confocal microscopy. FLmRNA and total cellular FL protein levels were increasedabout threefold. Overexpression of FL in AA was observedfor up to 20 years after IS treatment. FL levels correlatedinversely with CD341 cell numbers and the colony-formingability of AA bone marrow (R � 20´68 and 20´85
respectively). Histological examination of spleen specimensand bone marrow biopsies gave no evidence of degenerationor fibrosis due to prolonged exposure to high FL. Levels ofmembrane-bound FL were not increased in autoimmunediseases (n � 23), including rheumatoid arthritis and lupuserythematosus, nor in graft-versus-host disease (n � 8).Chronic overexpression of FL on the surface of T lympho-cytes in AA, but not in other T-cell-mediated disorders,suggests that membrane-bound FL plays a role in cell±cellinteractions in bone marrow failure and may be importantfor long-term haemopoietic recovery.
Keywords: aplastic anaemia, flt3 ligand, T lymphocytes,membrane-bound cytokines, bone marrow and spleenfibrosis.
Flt3 ligand (FL) is a haemopoietic growth factor specific for atyrosine kinase receptor flt3 (flt3R; Lyman et al, 1993;Hannum et al, 1994). This receptor is structurally related toc-kit, the receptor for stem cell factor (SCF), and c-fms, thereceptor for macrophage colony-stimulating factor (M-CSF).Signalling through flt3R is important in very early humanhaemopoiesis, as indicated by the restricted pattern ofreceptor expression found predominantly on primitivehaemopoietic precursors (Matthews et al, 1991; Rosnetet al, 1993). Accordingly, FL stimulates proliferation anddifferentiation of most primitive lineage-negative popula-tions of CD341 progenitor cells from human cord blood,peripheral blood (PB), bone marrow (BM) and fetal liver(reviewed in Lyman & Jacobsen, 1998). Like several other
haemopoietic cytokines, including SCF and M-CSF, FL existsin membrane-bound and soluble forms. Analysis of thealternatively spliced FL transcripts in human cells revealedan abundance of the species encoding membrane-boundmolecules (Lyman et al, 1994), and pulse-chase studiesdemonstrated that these primary products undergo subse-quent processing to yield soluble FL (McClanahan et al,1996). Biological properties of FL are recognized by theactivity of its soluble form, whereas the function ofmembrane-bound FL is not known. In analogy to SCF, ofwhich the membrane-bound form is essential for normalhaemopoietic development in the mouse (Flanagan et al,1991) and is more active than soluble SCF in stroma-supported haemopoiesis in vitro (Toksoz et al, 1992; SlanickaKrieger et al, 1998), membrane-bound FL may play anequally important role in cell±cell interactions within theearly progenitor cell compartment.
Although experimental results in vitro and in vivo have
211 q 2000 Blackwell Science Ltd
Correspondence: Aleksandra Wodnar-Filipowicz, Department ofResearch, University Hospital Basel, Hebelstr. 20, CH-4031 Basel,
Switzerland. E-mail: [email protected]

provided evidence for the role of the FL±flt3R interaction innormal haemopoiesis, the involvement of this ligand±receptor pair in haematological diseases is less well under-stood. The flt3R-encoding gene is frequently mutated (Xuet al, 1999), and flt3R expression was found in a largepercentage of human acute leukaemias of myeloid andlymphoid lineages (Birg et al, 1992; Carow et al, 1996). FL-dependent growth of leukaemic blasts in vitro was demon-strated in some cases (Piacibello et al, 1995; McKenna et al,1996) but there is no evidence that either the receptor orthe ligand play a causal role in the pathogenesis ofleukaemic transformation. Our recent studies suggest thatFL might be important for haemopoietic reconstitution inBM failure. Serum levels of FL, which are normally14 ^ 31 pg/ml (Lyman et al, 1995), increase up to7000 pg/ml in patients with aplastic anaemia (AA) and incancer patients with chemotherapy and/or irradiation-induced BM failure (Wodnar-Filipowicz et al, 1996). Themembrane-bound form of FL is also transiently over-expressed by T lymphocytes during cytoreductive che-motherapy (Chklovskaia et al, 1999).
Immune-mediated mechanisms are implicated in thepathogenesis of BM failure in AA (Young, 1995). Clinicalevidence is provided by high response rates to immunosup-pressive (IS) therapy (Marsh & Gordon-Smith, 1998). Theproportion of activated cytotoxic T lymphocytes is elevatedin the circulation and in BM infiltrates (Maciejewski et al,1994; Melenhorst et al, 1997) and it has been postulatedthat T cells play a role in BM destruction by overproducinginterferon-gamma and tumour necrosis factor, both potentinhibitors of haemopoiesis (Maciejewski et al, 1995; Nagafuiet al, 1995). In this work, we investigated expression of FLby T lymphocytes in AA before treatment and after IS
therapy. Levels of membrane-bound FL were also determinedin classical autoimmune diseases and in graft-versus-hostdisease (GVHD). As necrosis and fibrosis of the spleen andother organs were described in mice overexpressing FL (Juanet al, 1997), we examined splenectomy specimens and BMbiopsies of patients with treatment-refractory AA forevidence of degenerative changes resulting in fibre increase.Chronic overexpression of membrane-bound FL was foundonly in AA, reflecting a long-lasting defect in haemopoiesisand arguing for the significance of T-cell-derived mem-brane-bound FL in the pathophysiology of BM failure. Notissue damage was detected in the spleen or the BM frompatients exposed to high endogenous FL, both membranebound and soluble, providing an argument against fibrosisas an adverse effect of in vivo treatment with FL.
MATERIALS AND METHODS
Patients and controls. Sixteen patients were analysed atdiagnosis of severe idiopathic AA. All patients presentedwith acute previously untreated disease, hypoplastic bonemarrow, low blood cell values (see Table I) and weretransfusion dependent. Thirty-six AA patients were ana-lysed at 3 months to 20 years after IS treatment withantilymphocyte globulin (ALG) and cyclosporin A (CyA). Assome patients were analysed repeatedly, a total of 51measurements were performed after IS. Patients withautoimmune diseases (n � 23) included 13 patients withrheumatoid arthritis (RA; disease duration 1±33 years;nine patients with significant seropositivity), six patientswith systemic lupus erythematosus (SLE; three patients withactive disease) and patients with chronic CREST, acute Still'sdisease, acute vasculitis and cryoglobulinaemia. Eight
Table I. Characteristics of AA patients and FL expression at diagnosis.
Patient Age/sex Diagnosis Hb (g; %) Platelets (� 109/l) Neutrophils (� 109/l) FL* (MFI)
1 9/M SAAi/PNH 9´0 23 650 8´6
2 13/M SAAi 8´9 20 150 5´0
3 13/F SAAi 5´5 2 80 6´0
4 26/F SAAi 6´2 32 400 2´75 9/M SAAi 7´0 4 0 4´0
6 11/F SAAi 6´3 4 120 3´0
7 40/F SAAi 10´5 53 180 8´3
8 73/F SAAi 8´5 13 100 6´09 2/F SAAi 6´5 2 200 4´0
10 61/F SAAi 7´7 18 315 8´4
11 10/F SAAi 10´5 9 180 8´012 57/M SAAi 10´0 26 340 8´7
13 16/M SAAi 7´8 13 500 4´3
14 6/F SAAi 8´7 10 100 2´5
15 39/M SAAi 7´6 19 1115² 4´816 44/M SAAi 5´5 34 200 4´8
*FL expression levels on the cell surface of PB T lymphocytes, as determined in mean fluorescence intensity (MFI) by FACS (see Materials and
methods).
²Under G-CSF treatment.
Only AA patients before IS treatment are included in this Table.
SAAi, idiopathic severe AA; PNH, paroxysmal nocturnal haemoglobinuria.
212 O. Pfister et al
q 2000 Blackwell Science Ltd, British Journal of Haematology 109: 211±220

patients had GVHD, grades II±IV, developed after allogeneicstem cell transplantation. Control PB and BM was from 20healthy donors. Informed consent was obtained frompatients and donors and the study was approved by theEthical Committee of the University Hospital Basel.
Cell purification. Mononuclear cells from heparinized PB orBM (PBMC and BMMC respectively) were isolated by Ficoll-Hypaque density gradient centrifugation (d � 1´077) andused immediately without prior freezing for flow cytometry(FACS), confocal microscopy, RNA preparation and proteinextraction (see below).
Measurement of soluble FL. Serum was collected fromnative PB after centrifugation for 10 min at 3000 r.p.m.,aliquoted and stored at 2708C. Soluble FL was measured byan enzyme-linked immunosorbent assay (ELISA) (Lymanet al, 1995). Samples were analysed in duplicate; thereproducibility of the assay was 2±5%.
FACS analysis. PBMCs and BMMCs were washed inphosphate-buffered saline (PBS) containing 0´5% bovineserum albumin (BSA), and incubated for 20 min on icewith the appropriate monoclonal antibodies (mAbs). Mem-brane-bound FL was stained with mAb M5 against human FL(rat IgG2a; Lyman et al, 1995) at 20 mg/ml, followed byfluorescein isothiocyanate (FITC)-conjugated goat anti-ratIgG (Jackson Immunoresearch Laboratories, West Grove, PA,USA). CD34 was stained with phycoerythrin (PE)-labelledHPCA2 mAb (Becton Dickinson, Heidelberg, Germany) andCD3 with mouse anti-human CD3±PE mAbs (BectonDickinson). For the control staining, goat anti-rat IgG±FITC(Jackson Laboratories) and PE-labelled mouse IgG1 (BectonDickinson) were used. Acquisition of 10 000±25 000 eventswas performed with FACScan (Becton Dickinson). Dead cellswere stained with propidium iodide and were excluded fromthe analysis. Analysis was performed using CellQuest soft-ware (Becton Dickinson). Expression level of membrane-bound FL on the surface of CD31 T lymphocytes wasquantified as the mean fluorescence intensity (MFI) ratio ofstaining with FL-specific and isotype-matched control mAbs.
Confocal microscopy. Slides with PBMCs were prepared asdescribed previously (Chklovskaia et al, 1999). Cells wereincubated for 1 h at room temperature with M5 mAb at40 mg/ml and mouse anti-human CD3 at 20 mg/ml,washed in PBS containing 0´5% BSA and 0´1% saponin,and stained for 1 h in the dark with goat anti-rat IgG±FITC(Becton Dickinson) and goat anti-mouse IgG±Cy3 (Amer-sham, Pittsburgh, PA, USA). Confocal microscopy wascarried out with a TCS4D (Leica, Glattbrugg, Switzerland)operating in the sequential acquisition mode with 488(FITC) and 568 (Cy3) excitation lines.
Reverse transcription polymerase chain reaction (RT-PCR).An aliquot (1 mg) of total cellular RNA extracted fromPBMCs and BMMCs (Chomczynski & Sacchi, 1987) wastranscribed into complementary DNA and amplified by PCRunder the conditions of the linear accumulation of FL andb-actin PCR products (Chklovskaia et al, 1999), using thefollowing programmes: 948C for 1 min; 608C for 1 min;728C for 1 min; 25 cycles in a Perkin Elmer Cetus thermalcycler 8930 (Norwalk, CN, USA). DNA was electrophoresedin 1´2% agarose gels, transferred onto nylon membranes
and hybridized with [g-32P]-ATP-labelled oligonucleotidescorresponding to the internal sequences of amplifiedfragments. Determination of FL mRNA levels was based onthe ratio of signals given by FL and b-actin PCR products,analysed using Phosphoimager (Bio-Rad, Hercules, CA,USA). Autoradiography was with X-OMAT Kodak X-rayfilm and amplifying screens.
Western blot analysis. Extracts from 1 � 106 PBMCs(Chklovskaia et al, 1999) or recombinant human FL(rhFL; 5 ng) in Laemmli's sample buffer were electrophor-esed on reducing SDS polyacrylamide gels (12%) andtransferred to nitrocellulose. Western blot analysis wasperformed with M5 mAb (3 mg/ml) followed by secondaryhorseradish peroxidase-conjugated rabbit anti-rat IgG(Southern Biotechnology, Birmingham, AL, USA) anddetection with an enhanced chemiluminescent SuperSignal-substrate (Pierce, Rockford, IL, USA). For the controlimmunoblotting, isotype-matched rat IgG2a (PharMingen,San Diego, CA, USA) was used.
Haemopoietic colony formation assay. BMMCs (2 � 105)were plated in 1 ml methylcellulose cultures (Wodnar-Filipowicz et al, 1992). After 14 d at 378C, colonies of morethan 50 cells derived from myeloid and erythroid colony-forming units (CFU) were scored microscopically.
Histological analysis. Paraffin-embedded tissue blocks fromsplenectomy specimens (n � 5) and BM trephine biopsies(n � 6) were stained with haematoxylin and eosin (H & E)for demonstration of spleen architecture and with NovotnySilverstain for demonstration of reticulin fibres. Specimens fromspleen removed because of traumatic rupture served as normalcontrols. Splenectomy specimens and BM biopsies with fibrosisdue to a myeloproliferative disorder were used for comparison.
Statistical analyses. The Mann±Whitney test was used tocompare expression levels of membrane-bound FL by Tlymphocytes from healthy donors, AA and other patients.The Spearman rank test was used to analyse the relation-ship of FL levels with the content of CD341 cells, CFU andthe PB neutrophil count.
RESULTS
Elevated expression of membrane-bound FL by T lymphocytes inAAOur previous studies have demonstrated that membrane-bound FL is not detectable on the surface of normalmononuclear cells from healthy donors, or is expressed atvery low levels (Chklovskaia et al, 1999). In this study, weexamined the expression of FL in AA, using PB and BM from16 untreated patients at diagnosis and 36 patients after IStreatment with ALG and CyA. FACS analysis withFL-specific mAbs and antibodies against mononuclear cell-surface markers demonstrated that expression ofmembrane-bound FL was highly elevated in CD31 cells inAA compared with normal levels (Fig 1). Overexpression ofFL was equally pronounced in CD41 and CD81 subpopula-tions of T lymphocytes (not shown). The level of FL in severeuntreated AA, quantified as the mean fluorescence intensity(MFI), was 5´6 ^ 0´5 (mean ^ s.e.m.; range 2´5±8´7;Fig 2), significantly higher than normal values of
q 2000 Blackwell Science Ltd, British Journal of Haematology 109: 211±220
Expression of Membrane-bound flt3 Ligand in Aplastic Anaemia 213

1´4 ^ 0´04 (range 1´2±1´7; P , 0´0001; Fig 2). Forindividual patients, the correlation between FL expressionlevel and severity of AA at diagnosis was not obviousbecause all patients included in the analysis presented withsevere disease (see Table I). After IS, cell-surface levels of FLdecreased to 2´7 ^ 0´26 within the first year, but remainedelevated even 20 years after treatment (1´95 ^ 0´09;P , 0´0001; Fig 2). Notably, normal values in the rangeof 1´2±1´7 were observed in only 30% of all treated patients. FLlevels correlated inversely with the PB neutrophil count, used asa transfusion-independent parameter of haemopoiesis(R � 20´60; P , 0´0001). The level of membrane-boundFL expressed by BM T lymphocytes of AA patients at diagnosiswas 5´7 ^ 1´1 (compared with 1´7 ^ 0´1 in control BMcells; not shown), and thus corresponded closely to FL levels inPB (R � 0´96). For all measured samples, there was areciprocal correlation between membrane-bound FL and thecontent of CD341 progenitor cells in the BM (R � 20´68;P , 0´001; Fig 3A) and the number of haemopoieticcolonies formed by AA progenitors in vitro (R � 20´85;P , 0´0001; Fig 3B). These results suggest that cell-surface
Fig 1. FACS analysis of membrane-bound FL. Normal (N) and AA cells from PB and BM were double-stained with anti-CD3 and FL-specificmAb M5 (see Materials and methods). Staining of CD32 cell populations with mAb M5, equally pronounced with N and AA cells, was due to
unspecific binding of the antibody to Fc receptors on CD141 monocytic and CD191 B cells, which was not abolished by preincubation of M5
mAb with 50-fold molar excess of rhFL (not shown).
Fig 2. Effect of IS treatment on expression of membrane-bound FL
by T cells in AA. Results show mean fluorescence intensity (MFI) ofFL expression determined by FACS analysis of PB. Time after IS
therapy and the number of samples (n) are indicated; N, normal
controls. Asterisk, statistical significance of FL levels in AA
compared with N by Mann±Whitney correlation test wasP , 0´0001 before treatment, and P � 0´0001±0´002 at the
analysed time intervals after IS treatment.
214 O. Pfister et al
q 2000 Blackwell Science Ltd, British Journal of Haematology 109: 211±220

expression of FL reflects in an inverse manner the haemo-poietic activity of the BM and the clinical evolution in AA.
To determine whether overexpression of membrane-bound FL is specific for AA, we examined FL levels inclassic autoimmune diseases, including acute and chronicRA, SLE, CREST syndrome, Still's disease, vasculitis andcryoglobulinaemia and in GVHD developed after stem celltransplantation, i.e. in disorders associated with abnormalT-cell activation but without severe pancytopenia. FACSanalysis of PB for all these cases demonstrated that FLexpression was in the normal range (Table II). Similarly, FLserum levels measured by ELISA in 17 patients with SLEwere normal (not shown). This indicates that elevatedexpression of membrane-bound FL by T cells in AA reflectsfailure of haemopoietic function of the BM rather thanantigen-dependent T-cell activation.
Mechanism of FL expression in AATo examine the mechanism underlying the regulation of FL
expression in AA, we compared levels of FL mRNA andprotein in PBMCs and BMMCs from normal controls andfrom patients with severe AA before treatment (Fig 4).Semiquantitative RT-PCR detected constitutively expressed
Fig 3. Correlation between levels of membrane-bound FL (mean fluorescence intensity, MFI) with the content of (A) CD341 cells, and (B) thecolony-forming units (CFU) in AA BM. Results were obtained at 0±9 years after IS treatment of 31 and 23 patients respectively. Statistical
analysis with the Spearman rank test.
Table II. Expression of membrane-bound FL by T lymphocytes inautoimmune diseases and GVHD.
Patient group n FL (MFI ^ s.e.m.)
Normal controls 12 1´4 ^ 0´04
AA 16 5´6 ^ 0´50
RA 13 1´4 ^ 0´05
SLE 6 1´6 ^ 0´11Other AID* 4 1´6 ^ 0´31
GVHD 8 1´3 ^ 0´05
*AID, autoimmune diseases: CREST syndrome, Still's disease,
vasculitis and cryoglobulinaemia.
FL expression levels on the cell surface of PB T lymphocytes were
determined as mean fluorescence intensity (MFI) by FACS (see
Materials and methods). AA patients, included for a comparison,
were before IS treatment (see Table I). RA, rheumatoid arthritis;SLE, systemic lupus erythematosus.
For patients' description, see Materials and methods. FL levels in
AA compared with normal levels (Mann±Whitney test) was
P , 0´0001 (see also legend to Fig 2); RA, SLE, other AID andGVHD compared with normal levels P . 0´05.
Fig 4. Expression of FL mRNA and FL protein in AA. (A)
Semiquantitative RT-PCR analysis of FL- and b-actin-specific
transcripts. Twenty-five PCR cycles were performed with cDNAproducts from PB and BM from untreated AA patients and normal
(N) controls. Southern blots were hybridized to 32P-labelled internal
probes of FL and b-actin; autoradiography was for 4 h and 30 min
respectively. Each lane represents one patient or healthy donor. (B)Western analysis of FL protein in PB cells. Lane 1, rhFL, 5 ng; lanes
2±5 and 11, cells from normal donors; lanes 6±10, cells from AA
patients. Immunoblotting was with anti-FL mAb M5 (lanes 1±10)and with control IgG2a antibody (lane 11; see Materials and
methods). Arrows indicate 30- and 36-kDa FL proteins. Western
blot analysis with antiactin antibodies was performed as a loading
control.
q 2000 Blackwell Science Ltd, British Journal of Haematology 109: 211±220
Expression of Membrane-bound flt3 Ligand in Aplastic Anaemia 215

FL mRNA (Fig 4A). The signal intensity of FL-specifictranscripts was 2´5 ^ 0´4 times higher in AA (n � 17)than in controls (n � 18). Western blot analysis of PB cellsdetected constitutively expressed FL protein bands of about30 and 36 kDa, thought to represent the heterogeneouslyglycosylated FL species (McClanahan et al, 1996;Chklovskaia et al, 1999). The total content of FL proteinwas 2´9 ^ 0´4 times higher in AA (n � 5) than innormal levels (n � 4).
We used confocal microscopy to visualize the subcellulardistribution of FL in T cells. In normal PB cells stained withanti-CD3 and anti-FL antibodies, most of the FL protein wasdetected as a strong clustered signal in the cell cytoplasm(Fig 5B), in agreement with our recent data demonstratingthe existence of intracellular FL stores (Chklovskaia et al,1999). In AA, the FL-specific staining was found predomi-nantly on the cell surface, largely overlapping with stainingof CD3 surface antigen (Fig 5C). These results confirm thatcirculating T lymphocytes in severe AA express high levelsof membrane-bound FL on the cell surface.
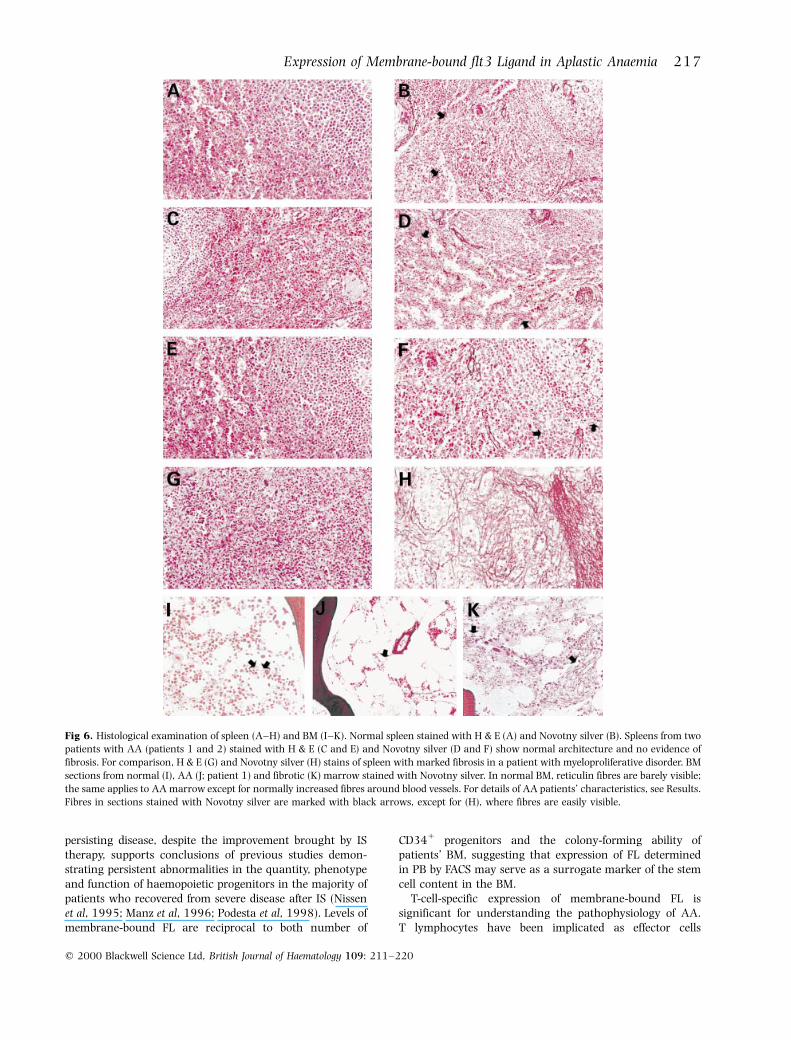
Absence of tissue fibrosis in spleen and BM from patients withAAAs overexpression of FL leads to tissue fibrosis in a mousemodel (Juan et al, 1997), we were interested whether long-term exposure to high endogenous FL, in both its solubleand membrane-bound forms, causes fibrosis of tissuesaccessible for examination in patients with chronic severeAA. Tissue from splenectomized patients and BM biopsiesfrom patients with treatment-refractory disease wereexamined histologically for the presence of fibrotic degen-eration (Fig 6). None of the reviewed splenectomy speci-mens from patients with high FL showed evidence offibrosis. Patient 1 (Fig 6C and D) underwent splenectomy12 years after the initial diagnosis, having been exposed toserum FL of 2500±4000 pg/ml throughout the entire 12-years period. In patient 2 (Fig 6E and F), splenectomy wasperformed 6 months after diagnosis; both membrane-boundand soluble FL were measured and found to be highlyelevated (4´7 MFI and 4000 pg/ml respectively). BMbiopsies from these patients showed severe reduction ofhaemopoietic cells and an increase in fat spaces typical forAA, but no fibre increase (Fig 6J). In summary, these resultsargue against major organ toxicity of high FL in humans.
DISCUSSION
In this paper, we have demonstrated that the membrane-bound form of FL was highly overexpressed in PB and BM ofpatients with severe AA compared with normal controls.Cell-surface expression of FL is specific for T lymphocytesand has not been found in other types of mononuclear cells.To date, AA represents the only known human diseaseassociated with elevated membrane-bound FL. We found noincrease in FL in T cells from patients with autoimmunedisorders, such as RA and SLE, or in patients with GVHDafter stem cell transplantation. Membrane-bound FL wasalso not increased in two patients with acute T-cellleukaemia (data not shown). These observations indicatethat surface expression of FL takes place in response tohaemopoietic failure and is not due to pathologicalactivation of T cells in response to allo- or autoantigens.As membrane-bound FL is thought to represent a precursorto the soluble isoform (Lyman et al, 1993; Hannum et al,1994), its overexpression in AA is in accordance withpreviously reported high serum levels of soluble FL in thisdisease (Lyman et al, 1995; Wodnar-Filipowicz et al, 1996).In studies on the mechanism responsible for increased FLlevels in AA, we have shown that the increase in FL mRNAexpression and cellular protein content is relatively modest,given the strongly pronounced increase in membrane-bound and soluble FL at onset of severe AA. These resultsconfirm our conclusions from a study on the regulation ofFL expression in chemotherapy-induced aplasia, whichdemonstrated that mobilization of preformed intracellularFL and its translocation to the T-lymphocyte cell surface,rather than enhanced FL gene transcription, are responsiblefor increased FL levels in BM failure (Chklovskaia et al,1999). The nature of the signal which triggers theaccelerated release of FL from intracellular stores uponloss of haemopoietic progenitors is not known.
FL levels decrease in response to IS treatment, ashaemopoietic function improves. However, expression ofmembrane-bound FL by T cells in AA rarely normalizes andremains elevated in most patients even 20 years aftertreatment. This corresponds to results on soluble FL, serumconcentrations of which decrease significantly in responseto treatment, but remain at levels exceeding normal values(Wodnar-Filipowicz et al, 1996). Chronic overexpression ofFL, both membrane bound and soluble, as a sign of
Fig 5. Localization of FL in T lymphocytes from AA patients. Two-colour immunofluorescence confocal microscopy was performed withnormal and AA blood cells stained with anti-FL M5 mAb followed by secondary FITC-labelled Ab and with anti-CD3 followed by secondary
Cy3-labelled Ab. (A) Normal T cell stained for CD3 (red); (B) normal T cells stained for FL (green) and CD3 (red); (C) AA T cells stained for FL
(green) and CD3 (red). Exact overlap of red and green signals is highlighted in white.
216 O. Pfister et al
q 2000 Blackwell Science Ltd, British Journal of Haematology 109: 211±220
persisting disease, despite the improvement brought by IStherapy, supports conclusions of previous studies demon-strating persistent abnormalities in the quantity, phenotypeand function of haemopoietic progenitors in the majority ofpatients who recovered from severe disease after IS (Nissenet al, 1995; Manz et al, 1996; Podesta et al, 1998). Levels ofmembrane-bound FL are reciprocal to both number of
CD341 progenitors and the colony-forming ability ofpatients' BM, suggesting that expression of FL determinedin PB by FACS may serve as a surrogate marker of the stemcell content in the BM.
T-cell-specific expression of membrane-bound FL issignificant for understanding the pathophysiology of AA.T lymphocytes have been implicated as effector cells
Fig 6. Histological examination of spleen (A±H) and BM (I±K). Normal spleen stained with H & E (A) and Novotny silver (B). Spleens from twopatients with AA (patients 1 and 2) stained with H & E (C and E) and Novotny silver (D and F) show normal architecture and no evidence of
fibrosis. For comparison, H & E (G) and Novotny silver (H) stains of spleen with marked fibrosis in a patient with myeloproliferative disorder. BM
sections from normal (I), AA (J; patient 1) and fibrotic (K) marrow stained with Novotny silver. In normal BM, reticulin fibres are barely visible;
the same applies to AA marrow except for normally increased fibres around blood vessels. For details of AA patients' characteristics, see Results.Fibres in sections stained with Novotny silver are marked with black arrows, except for (H), where fibres are easily visible.
q 2000 Blackwell Science Ltd, British Journal of Haematology 109: 211±220
Expression of Membrane-bound flt3 Ligand in Aplastic Anaemia 217

responsible for the destruction of haemopoietic progenitors.However, the culprit antigen on the progenitors remainsunknown and elucidation of the mechanism of haemopoie-tic suppression is further complicated by the fact that thefrequency of phenotypic and functional abnormalities foundin the T-cell compartment was relatively low, and none wascommon to all patients (Maciejewski et al, 1994; Nakao et al,1995; Zeng et al, 1999). In contrast, strong overexpressionof FL on the surface of T cells is an invariable finding in allpatients with severe untreated or treatment-refractory AA.Membrane-bound FL is equally present on CD41 helper andCD81 cytotoxic T-cell subsets, irrespective of their propor-tions in patients' PB and BM, suggesting that FL expressiondoes not correlate with the cytotoxic function of T cells.Also, the decrease in FL levels after IS therapy was equallypronounced in both cell subsets.
Properties of the membrane-bound FL isoform have notyet been characterized. Soluble FL has a wide array ofactivities, including potent effects on haemopoiesis and onthe immune system via its ability to stimulate thedevelopment of dendritic cells and natural killer cells(Fernandez et al, 1999). The latter property raises theprovocative hypothesis that FL, greatly elevated as aconsequence of stem cell deficiency, may stimulate out-growth of dendritic cells from the residual haemopoieticprogenitors. This may lead to enhanced presentation of theputative offending (auto)antigens to T cells, and thusaggravate AA. On the other hand, the haemopoieticproperties of FL justify speculations that up-regulation ofFL in response to BM failure reflects physiological require-ments and represents a haemopoietic rescue mechanism. Aslymphoid cells are spared in the majority of AA patients,availability of FL should not be limiting. Therefore, selectiveinfiltration of the pancytopenic BM with T cells, so farconsidered detrimental because of overproduction of cyto-kines toxic to haemopoiesis, may also have a positive effectderiving from the ability of infiltrating cells to supply locallymembrane-bound and soluble FL. The biological importanceof membrane-bound cytokines has been emphasized for SCFby haematological deficiencies in murine Steel-Dickiemutants genetically lacking membrane-bound SCF (Flanaganet al, 1991). Other cytokines, such as tumour necrosis factorand Fas ligand, are also more potent in their membrane-bound form than as soluble proteins (Grell et al, 1995;Tanaka et al, 1998). Accordingly, chronic overexpression ofmembrane-bound FL may play a role in local compensatorygrowth factor supply in BM failure, and thus help tomaintain blood production from the reduced haemopoieticreservoir in AA.
Potential clinical applications of FL include haemopoieticstem cell transplantation and cancer immunotherapy basedon in vivo properties of FL such as mobilization ofhaemopoietic stem cells to the circulation, radioprotectionand anti-tumour effects (Lynch et al, 1997; Papayannopoulouet al, 1997; Gratwohl et al, 1998). However, a potential riskof in vivo administration of FL has recently been raised byreports on the occurrence of severe splenic fibrosis andleukaemic predisposition in mice transplanted with retro-virally transduced BM cells expressing high levels of FL
(Juan et al, 1997; Hawley et al, 1998). Chronically elevatedFL in AA offers a chance to test these questions in humans.The development of clonal malignancies, including leukae-mia, is a known late complication of AA (Tichelli et al,1988). That chronically elevated FL is involved in thistransformation process cannot be ruled out; however, theexpression level of flt3R on AA precursors is not higher thannormal (our unpublished data). The absence of fibrotictissue degeneration in spleen and BM of AA patients exposedfor decades to chronically elevated soluble and membrane-bound FL did not confirm the results obtained in anexperimental mouse transplantation model. A possibleexplanation for the divergent findings is an exceedinglyhigh expression level of FL by the engineered cells, althoughno measurements of serum FL in transplanted mice (Juan et al,1997) were reported. In healthy volunteers given FL over2 weeks, serum levels reach about 1000 pg/ml (Lebsack et al,1997), below the levels measured in patients with severe BMfailure. Therefore, lack of tissue fibrosis in these patientsargues against this potential hazard with clinical applicationof FL.
ACKNOWLEDGMENTS
This work was supported by grants from the Swiss NationalScience Foundation (32-045926.95 and 055694.98),Roche Research Foundation, Basler Krebsliga, EmiliaGuggenheim Foundation, Lichtenstein Stiftung and UardaFrutiger Stiftung. We are indebted to A. Gratwohl, J.Passweg, G. Favre, A. Tyndall and D. Maclachlan forproviding PB and BM samples from patients with AA,autoimmune diseases and GVHD.
REFERENCES
Birg, F., Courcoul, M., Rosnet, O., Bardin, F., Pebusque, M.J.,Marchetto, S., Tabilio, A., Mannoni, P. & Birnbaum, D. (1992)
Expression of the FMS/KIT-like gene FLT3 in human acute
leukemias of the myeloid and lymphoid lineages. Blood, 80,2584±2593.
Carow, C.E., Levenstein, M., Kaufmann, S.H., Chen, J., Amin, S.,
Rockwell, P., Witte, L., Borowitz, M.J., Civin, C.I. & Small, D.
(1996) Expression of the hematopoietic growth factor receptor
flt3 (stk-1/flk-2) in human leukemias. Blood, 87, 1089±1096.
Chklovskaia, E., Jansen, W., Nissen, C., Lyman, S.D., Rahner, C.,Landmann, L. & Wodnar-Filipowicz, A. (1999) Mechanism of flt3
ligand expression in bone marrow failure: translocation from
intracellular stores to the surface of T lymphocytes after
chemotherapy-induced suppression of hematopoiesis. Blood, 93,2595±2604.
Chomczynski, P. & Sacchi, N. (1987) Single-step method of RNA
isolation by acid guanidinium thiocyanate±phenol±chloroform
extraction. Analytical Biochemistry, 162, 156±159.
Fernandez, N.C., Lozier, A., Flament, C., Ricciardi-Castagnoli, P.,Bellet, D., Sutter, M., Perricaudet, M., Tursz, T., Maraskovsky, E. &
Zitvogel, L. (1999) Dendritic cells directly trigger NK functions:
cross-talk relevant in innate anti-tumor immune responses in
vivo. Nature Medicine, 5, 405±411.
Flanagan, J.G., Chan, D.C. & Leder, P. (1991) Transmembrane formof the kit ligand growth factor is determined by alternative
splicing and is missing in the Sld mutant. Cell, 64, 1025±1035.
218 O. Pfister et al
q 2000 Blackwell Science Ltd, British Journal of Haematology 109: 211±220

Gratwohl, A., John, L., Baldomero, H., Roth, J., Tichelli, A., Nissen,
C., Lyman, S.D. & Wodnar-Filipowicz, A. (1998) Flt3 ligand
provides hematopoietic protection from total body irradiation inrabbits. Blood, 92, 765±769.
Grell, M., Lohden, M., Maxeiner, B., Lesslauer, W. & Pfizenmeier, K.
(1995) The transmembrane form of tumor necrosis factor is theprime activating ligand of the 80 kDa tumor necrosis factor
receptor. Cell, 83, 795±805.
Hannum, C., Culpepper, J., Campbell, D., McClanahan, T., Zurawski,S., Bazan, J.F., Kastelein, R., Hudak, S., Wagner, J., Mattson, J.,
Luh, J., Duda, G., Martina, N., Peterson, D., Menon, S., Shanafelt,
A., Muench, M., Kelner, G., Namikawa, R., Rennick, D.,
Roncarolo, M.-G., Zlotnik, A., Rosnet, O., Dubreuil, P., Birnbaum,D. & Lee, F. (1994) Ligand for FLT3/FLK2 receptor tyrosine kinase
regulates growth of haematopoietic stem cells and is encoded by
variant RNAs. Nature, 368, 643±648.
Hawley, T.S., Fong, A.Z., Griesser, H., Lyman, S.D. & Hawley, R.G.
(1998) Leukemic predisposition of mice transplanted with gene-
modified hematopoietic precursors expressing flt3 ligand. Blood,
92, 2003±2011.
Juan, T.S.C., Van McNiece, I.G., Lacey, D., Hartley, C., McElroy, P.,
Sun, Y., Argento, J., Hill, D., Yan, X.Q. & Fletcher, F.A. (1997)
Chronic expression of murine flt3 ligand in mice results inincreased circulating white blood cell levels and abnormal
cellular infiltrates associated with splenic fibrosis. Blood, 90,
76±84.
Lebsack, M.E., McKenna, H.J., Hoek, J.A., Hanna, R., Feng, A.,
Maraskovsky, E. & Hayes, F.A. (1997) Safety of flt3 ligand in
healthy volunteers. Blood, 90, 170a.
Lyman, S. & Jacobsen, S. (1998) c-kit ligand and Flt3 ligand: stem/
progenitor cell factors with overlapping yet distinct activities.
Blood, 91, 1101±1134.
Lyman, S.D., James, L., Vanden, B.T., de Vries, P., Brasel, K., Gliniak,
B., Hollingsworth, L.T., Picha, K.S., McKenna, H.J., Splett, R.R.,
Fletcher, F.F., Maraskovsky, E., Farrach, F., Foxworthe, D.,
Williams, D.E. & Beckmann, M.P. (1993) Molecular cloning of aligand for the flt3/flk-2 tyrosine kinase receptor: a proliferative
factor for primitive hematopoietic cells. Cell, 75, 1157±1167.
Lyman, S.D., James, L., Johnson, L., Brasel, K., de Vries, P., Escobar,S.S., Downey, H., Splett, R.R., Beckmann, M.P. & McKenna, H.J.
(1994) Cloning of the human homologue of the murine flt3
ligand: a growth factor for early hematopoietic progenitor cells.
Blood, 83, 2795±2801.
Lyman, S.D., Seaberg, M., Hanna, R., Zappone, J., Brasel, K.,
Abkowitz, J.L., Prchal, J., Schultz, J. & Shahidi, N.T. (1995) Flt3
ligand plasma/serum levels are low in normal individuals and arehighly elevated in patients with Fanconi anemia and acquired
aplastic anemias. Blood, 86, 4091±4096.
Lynch, D.H., Andreasen, A.E.M., Whitmore, J., Miller, R.E. & Schuh,J.C.L. (1997) Flt3 ligand induces tumor regression and antitumor
immune responses in vivo. Nature Medicine, 3, 625±631.
Maciejewski, J.P., Hibbs, J.R., Anderson, S., Katevas, P. & Young, N.S.(1994) Bone marrow and peripheral blood lymphocyte pheno-
type in patients with bone marrow failure. Experimental
Hematology, 22, 1102±1110.
Maciejewski, J., Selleri, C., Anderson, S. & Young, N.S. (1995) Fas
antigen expression on CD341 human marrow cells is induced by
interferon g and tumor necrosis factor a and potentiates
cytokine-mediated hematopoietic suppression in vitro. Blood, 85,3138±3190.
Manz, C.Y., Nissen, C. & Wodnar-Filipowicz, A. (1996) Deficiency of
CD341c-kit1 and CD341382 hematopoietic precursors inaplastic anemia after immunosuppressive treatment. American
Journal of Hematology, 52, 264±274.
Marsh, J.C. & Gordon-Smith, E.C. (1998) Treatment options in
severe aplastic anaemia. Lancet, 351, 1830±1831.
Matthews, W., Jordan, C.T., Gavin, M., Jenkins, N.A., Copeland, N.G.
& Lemischka, I.R. (1991) A receptor tyrosine kinase cDNA
isolated from a population of enriched primitive hematopoietic
cells and exhibiting close genetic linkage to c-kit. Proceedings of theNational Academy of Sciences of the United States of America, 88,
9026±9030.
McClanahan, T., Culpepper, J., Campbell, D., Wagner, J., Franz-Bacon, K., Mattson, J., Tsai, S., Luh, J., Guimaraes, J., Mattei, M.-
G., Rosnet, O., Birnbaum, D. & Hannum, C.H. (1996) Biochemical
and genetic characterisation of multiple splice variants of the flt3
ligand. Blood, 88, 3371±3382.
McKenna, H.J., Smith, F.O., Brasel, K., Hirschstein, D., Bernstein,
I.D., Williams, D.E. & Lyman, S.D. (1996) Effects of flt3 ligand on
acute myeloid and lymphocytic blast cells from children.Experimental Hematology, 24, 378±385.
Melenhorst, J.J., Van Krieken, J.H., Dreef, E., Landegent, J.E.,
Willemze, R. & Fibbe, W.E. (1997) T cells selectively infiltratebone marrow areas with residual haemopoiesis of patients with
acquired aplastic anaemia. British Journal of Haematology, 99,
517±519.
Nagafui, K., Shibuya, T., Harada, M., Mizuno, S.I., Takenaka, K.,
Miyamoto, T., Okamura, T., Gondo, H. & Niho, Y. (1995)
Functional expression of Fas antigen (CD95) on hematopoietic
progenitor cells. Blood, 86, 883±889.
Nakao, S., Takamatsu, H., Yachie, A., Itoh, T., Yamaguchi, M.,
Ueda, M., Shiobara, S. & Matsuda, T. (1995) Establishment of a
CD41 T cell clone recognizing autologous hematopoieticprogenitor cells from a patient with immune-mediated aplastic
anaemia. Experimental Hematology, 23, 433±438.
Nissen, C., Wodnar-Filipowicz, A., Slanicka Krieger, M., Gratwohl,A., Tichelli, A. & Speck, B. (1995) Persistent growth impairment
of bone marrow stroma after antilymphocyte globulin treatment
for severe aplastic anemia and its association with relapse.
European Journal of Haematology, 55, 255±261.
Papayannopoulou, T., Nakamoto, B., Andrews, R.G., Lyman, S.D. &
Lee, M.Y. (1997) In vivo effects of flt3/flk2 ligand on mobilization
of hematopoietic progenitors in primates and potent synergisticenhancement with granulocyte colony-stimulating factor. Blood,
90, 620±629.
Piacibello, W., Fubini, L., Sanavio, F., Brizzi, M.F., Severino, A.,Garetto, L., Stacchini, A., Pegoraro, L. & Aglietta, M. (1995)
Effects of human flt3 ligand on myeloid leukemias cell growth:
heterogeneity in response and synergy with other hematopoietic
growth factors. Blood, 85, 4105±4114.
Podesta, M., Piaggio, G., Frassoni, F., Pitto, A., Zikos, P., Sessarego,
M., Abate, M.M.T.V.L., Berisso, G. & Bacigalupo, A. (1998) The
assessment of the hematopoietic reservoir after immunosuppres-sive therapy or bone marrow transplantation in severe aplastic
anemia. Blood, 91, 1959±1965.
Rosnet, O., Schiff, C., Pebusque, M.J., Marchetto, S., Tonnelle, C.,Toiron, Y., Birg, F. & Birnbaum, D. (1993) Human FLT3/FLK2
gene: cDNA cloning and expression in hematopoietic cells. Blood,
82, 1110±1119.
Slanicka Krieger, M., Nissen, C., Manz, C.Y., Toksoz, D.S.D., Lyman,
S.D. & Wodnar-Filipowicz, A. (1998) The membrane-bound
isoform of stem cell factor synergizes with soluble flt3 ligand in
supporting early hematopoiesis in long-term cultures of normaland aplastic anemia bone marrow. Experimental Hematology, 26,
365±373.
Tanaka, M., Itai, T., Adachi, M. & Nagata, S. (1998)Downregulation of Fas ligand by shedding. Nature Medicine, 4,
31±36.
q 2000 Blackwell Science Ltd, British Journal of Haematology 109: 211±220
Expression of Membrane-bound flt3 Ligand in Aplastic Anaemia 219

Tichelli, A., Gratwohl, A., WuÈ rsch, A., Nissen, C. & Speck, B. (1988)
Late haematological complications in severe aplastic anaemia.
British Journal of Haematology, 69, 413±418.Toksoz, D., Zsebo, K.M., Smith, K.A., Hu, S., Brankow, D., Suggs,
S.V., Martin, F.H. & Williams, D.A. (1992) Support of human
hematopoiesis in long-term bone marrow cultures by murinestromal cells selectively expressing the membrane-bound and
secreted forms of the human homolog of the steel gene product,
stem cell factor. Proceedings of the National Academy of Sciences of
the United States of America, 89, 7350±7354.Wodnar-Filipowicz, A., Tichelli, A., Zsebo, K.M., Speck, B. & Nissen,
C. (1992) Stem cell factor stimulates the in vitro growth of bone
marrow cells from aplastic anemia patients. Blood, 79, 3196±
3202.Wodnar-Filipowicz, A., Lyman, S., Gratwohl, A., Tichelli, A., Speck,
B. & Nissen, C. (1996) Flt3 ligand level reflects hematopoietic
progenitor cell function in aplastic anemia and chemotherapy-
induced bone marrow aplasia. Blood, 88, 4493±4499.
Xu, F., Taki, T., Yang, H.W., Hanada, R., Hongo, T., Ohnishi, H.,Kobayashi, M., Bessho, F., Yaniagisawa, M. & Hayashi, Y. (1999)
Tandem duplication of the flt3 gene is found in acute
lymphoblastic leukaemia as well as acute myeloid leukaemiabut not in myelodysplastic syndrome or juvenile chronic
myelogenous leukaemia in children. British Journal of Haematol-
ogy, 105, 155±162.
Young, N.S. (1995) Aplastic anaemia. Lancet, 346, 228±232.Zeng, W., Nakao, S., Takamatsu, H., Yachie, A., Takami, A., Kondo,
Y., Sugimori, N., Yamazaki, H., Miura, Y., Shiobara, S. & Matsuda,
T. (1999) Characterization of T-cell repertoire of the bone
marrow in immune-mediated aplastic anemia: evidence for theinvolvement of antigen-driven T-cell response in cyclosporin-
dependent aplastic anemia. Blood, 93, 3008±3016.
220 O. Pfister et al
q 2000 Blackwell Science Ltd, British Journal of Haematology 109: 211±220









![Optimization of Imidazo[4,5- b ]pyridine-Based Kinase Inhibitors: Identification of a Dual FLT3/Aurora Kinase Inhibitor as an Orally Bioavailable Preclinical Development Candidate](https://static.fdokumen.com/doc/165x107/6345ee68596bdb97a909280b/optimization-of-imidazo45-b-pyridine-based-kinase-inhibitors-identification.jpg)










