
Radiofrequency radiation does not induce stress response in human T-lymphocytes and rat primary...
11
Bioelectromagnetics 27:578 ^ 588 (2006) Radiofrequency Radiation Does Not Induce Stress Response in HumanT-Lymphocytes and Rat Primary Astrocytes Jae-Seon Lee, 1{ Tai-Qin Huang, 1 Tae-Hyung Kim, 1 Jin Young Kim, 2 Hyun Jeong Kim, 2 Jeong-Ki Pack, 3 and Jeong-Sun Seo 1 * 1 Department of Biochemistry and Molecular Biology, ILCHUN Molecular Medicine Institute MRC, Seoul National University College of Medicine, Seoul, Korea 2 Department of Dental Anesthesiology and Dental Research Institute, Seoul National University College of Dentistry, Seoul, Korea 3 Department of Radio Sciences & Engineering, Chungnam National University, Daejeon, Korea Heat shock proteins (HSPs) are rapidly induced by a variety of stressors, including heat shock, ethanol, heavy metals, UV, and g-radiation. Mitogen-activated protein kinases (MAPKs) are also involved in the stress transduction pathways in all eukaryotes. In this study, we attempted to determine whether radiofrequency (RF) radiation is able to induce a non-thermal stress response. Human T-lymphocyte Jurkat cells and rat primary astrocytes were exposed to 1763 MHz of RF radiation at an average specific absorption rate (SAR) of either 2 W/kg or 20 W/kg, for 30 min or 1 h. Temperature was completely controlled at 37 0.2 8C throughout the exposure period. The sham exposures were performed under exactly identical experimental conditions without exposure to RF radiation. We assessed alterations in the expression of HSPs and the activation of MAPKs in the RF-exposed cells. No detectable difference was observed in the expression levels of HSP90, HSP70, and HSP27. The phosphorylation status of MAPKs, extracellular signal-regulated kinases (ERK1/2), c-Jun N-terminal protein kinases (JNK1/2), or p38, did not change significantly. In order to determine whether RF radiation can promote the effects of 12-O-tetradecanoylphorbol 13-acetate (TPA) on stress response, cells were exposed to RF radiation coupled with TPA treatment. When TPA alone was applied, the MAPKs were found to be phosphorylated in a dose-dependent manner. However, RF radiation did not result in any enhancement of TPA-induced MAPK phosphorylation. Neither TPA nor RF radiation exerted any detectable effect on the induction of HSPs. These results indicate that 1763 MHz RF radiation alone did not elicit any stress response, nor did it have any effect on TPA-induced MAPK phosphorylation, under our experimental conditions. Bioelectromagnetics 27:578–588, 2006. ȣ 2006 Wiley-Liss, Inc. Key words: RF radiation; mobile phone; HSP; MAPK INTRODUCTION The use of mobile phones has increased contin- uously over recent years, and the mobile phone has become one of the principal sources of environmental radiofrequency (RF) radiation. A number of studies have been performed in order to determine whether RF radiation triggers stress responses [Cleary et al., 1997; Fritze et al., 1997; Leszczynski et al., 2002]. Although some available evidence suggests that RF radiation induces stress responses, many contradictory findings have also been reported; and the available evidence on both sides appears equally compelling. Therefore, further research will be required in order to determine whether RF radiation could be an environmental stressor. ȣ 2006 Wiley-Liss, Inc. —————— Grant sponsor: The Ministry of Information and Communication of Korea; Grant number: 2003-X-004. { Jae-Seon Lee’s present address is Laboratory of Functional Genomics, Korean Institute of Radiological & Medical Sciences, Seoul, Korea. *Correspondence to: Jeong-Sun Seo, ILCHUN Molecular Med- icine Institute MRC, Seoul National University College of Medicine, Seoul 110-799, Korea. E-mail: [email protected] Received for review 1 July 2005; Final revision received 17 February 2006 DOI 10.1002/bem.20235 Published online 12 July 2006 in Wiley InterScience (www.interscience.wiley.com).
-
Upload
menninconsulting -
Category
Documents
-
view
0 -
download
0
Transcript of Radiofrequency radiation does not induce stress response in human T-lymphocytes and rat primary...

Bioelectromagnetics 27:578^588 (2006)
Radiofrequency Radiation Does Not InduceStress Response in HumanT-Lymphocytes and
Rat Primary Astrocytes
Jae-Seon Lee,1{ Tai-Qin Huang,1 Tae-Hyung Kim,1 JinYoung Kim,2 Hyun Jeong Kim,2
Jeong-Ki Pack,3 and Jeong-Sun Seo1*1Department of Biochemistry andMolecular Biology, ILCHUNMolecular
Medicine InstituteMRC, SeoulNational University College ofMedicine, Seoul, Korea2Department of DentalAnesthesiology andDental Research Institute,
SeoulNational University College of Dentistry, Seoul, Korea3Department of Radio Sciences & Engineering, ChungnamNational University,
Daejeon, Korea
Heat shock proteins (HSPs) are rapidly induced by a variety of stressors, including heat shock, ethanol,heavy metals, UV, and g-radiation. Mitogen-activated protein kinases (MAPKs) are also involved inthe stress transduction pathways in all eukaryotes. In this study, we attempted to determine whetherradiofrequency (RF) radiation is able to induce a non-thermal stress response. Human T-lymphocyteJurkat cells and rat primary astrocytes were exposed to 1763 MHz of RF radiation at an averagespecific absorption rate (SAR) of either 2 W/kg or 20 W/kg, for 30 min or 1 h. Temperature wascompletely controlled at 37� 0.2 8C throughout the exposure period. The sham exposures wereperformed under exactly identical experimental conditions without exposure to RF radiation. Weassessed alterations in the expression of HSPs and the activation of MAPKs in the RF-exposed cells.No detectable difference was observed in the expression levels of HSP90, HSP70, and HSP27. Thephosphorylation status ofMAPKs, extracellular signal-regulated kinases (ERK1/2), c-Jun N-terminalprotein kinases (JNK1/2), or p38, did not change significantly. In order to determine whether RFradiation can promote the effects of 12-O-tetradecanoylphorbol 13-acetate (TPA) on stress response,cells were exposed to RF radiation coupled with TPA treatment. When TPA alone was applied, theMAPKs were found to be phosphorylated in a dose-dependent manner. However, RF radiation did notresult in any enhancement of TPA-induced MAPK phosphorylation. Neither TPA nor RF radiationexerted any detectable effect on the induction of HSPs. These results indicate that 1763 MHz RFradiation alone did not elicit any stress response, nor did it have any effect on TPA-induced MAPKphosphorylation, under our experimental conditions. Bioelectromagnetics 27:578–588, 2006.� 2006 Wiley-Liss, Inc.
Key words: RF radiation; mobile phone; HSP; MAPK
INTRODUCTION
The use of mobile phones has increased contin-uously over recent years, and the mobile phone hasbecome one of the principal sources of environmentalradiofrequency (RF) radiation. A number of studieshave been performed in order to determine whether RFradiation triggers stress responses [Cleary et al., 1997;Fritze et al., 1997; Leszczynski et al., 2002]. Althoughsome available evidence suggests that RF radiationinduces stress responses, many contradictory findingshave also been reported; and the available evidenceon both sides appears equally compelling. Therefore,further research will be required in order to determinewhether RF radiation could be an environmentalstressor.
�2006Wiley-Liss, Inc.
——————Grant sponsor: The Ministry of Information and Communicationof Korea; Grant number: 2003-X-004.
{Jae-Seon Lee’s present address is Laboratory of FunctionalGenomics, Korean Institute of Radiological & Medical Sciences,Seoul, Korea.
*Correspondence to: Jeong-Sun Seo, ILCHUN Molecular Med-icine Institute MRC, Seoul National University College ofMedicine, Seoul 110-799, Korea. E-mail: [email protected]
Received for review 1 July 2005; Final revision received 17February 2006
DOI 10.1002/bem.20235Published online 12 July 2006 in Wiley InterScience(www.interscience.wiley.com).

Stress responses are generally characterized bythe induction of heat shock proteins (HSPs), which areubiquitously distributed and highly conserved frombacteria to mammals. HSPs can be induced by a varietyof stressors, such as heat, ethanol, amino acid analogs,infections, heavy metals, UV, and g-radiation [Mor-imoto, 1992]. In addition to their chaperoning functionsin protein folding and assembly, HSP family membersprotect cells against harmful environments. HSP90,HSP70, and HSP27 have been reported as the mostinducible molecules among HSP family membersunder the stressful conditions [Morimoto, 1992; Aliet al., 1996; Musch et al., 2004]. Therefore, we havechosen those proteins to examine whether RF radiationinduce stress response in this study.
The mitogen-activated protein kinases (MAPKs)are major participants in stress responses. MAPKactivation plays a pivotal role in cell recovery,proliferation, differentiation, and death [Minden andKarin, 1997; Ip and Davis, 1998; Schaeffer and Weber,1999]. These kinases phosphorylate and activate tran-scription factors via kinase cascades in response tostresses [Johnson and Lapadat, 2002].
The present study was conducted to determine theeffects of the 1763 MHz radiation emitted from mobilephones on stress response. The expression of HSPsand the phosphorylation of MAPKs were monitored,after the exposure of human T-lymphocytes and ratprimary astrocytes after RF radiation exposure.We alsoattempted to determine whether RF radiation exerted asynergistic effect on the status of MAPK phosphor-ylation induced by 12-O-tetradecanoylphorbol 13-acetate (TPA).
MATERIALS AND METHODS
Materials
Antibodies against c-Jun N-terminal kinase(JNK), extracellular signal-regulated kinase (ERK),p38, phospho-JNK (Thr183/Tyr185), phospho-ERK(Thr202/yr204), and phospho-p38 (Thr180/Tyr182)were purchased from Cell Signaling (Beverly, MA,USA). Anti-HSP27, anti-HSP70, anti-HSP90, anti-actin, anti-glial fibrillary acidic protein (GFAP), andhorseradish peroxidase-conjugated secondary antibod-ies were obtained from Santa Cruz Biotechnology(Santa Cruz, CA, USA). TPA was purchased fromSigma-Aldrich (St. Louis, MO, USA). The molecularweight standards for sodium dodecyl sulfate (SDS)–polyacrylamide gel electrophoresis were obtained fromBio-Rad Laboratories (Hercules, CA, USA). Bicincho-ninic acid (BCA) Protein Assay Reagents wereobtained from PIERCE (Rockford, IL). The enhanced
chemiluminescence (ECL) detection kit was fromAmersham Biosciences (Little Chalfont, UK).Sprague–Dawley rats for the preparation of rat corticalastrocytes were purchased from Daehan Biolink Co.,Korean Branch of Harlan Laboratories (Seoul, Korea).Jurkat cell line was purchased form the American TypeCulture Collection (ATCC; Manassas, VA, USA).
Preparation of Cortical Astrocytesand Cell Culture
Cortical astrocyte cells of 1- or 2-day-oldSprague–Dawley rat pups were cultured according tothe method of Galve-Roperh et al. [1997]. Briefly,mechanically dissociated cortical cells were seeded inpoly-D-lysine-coated 24 wells at the density of 2� 105
cells/ml in minimal essential medium (MEM) supple-mented with 20 mM glucose, 5% fetal bovine serum(FBS), 5% horse serum, 2 mM glutamine, 100 mg/mlstreptomycin, and 100U/ml penicillin.When cellsweregrown to confluence in 5% CO2 incubator at 37 8C,cortical glial cultures were characterized by anti-GFAPantibody as an astrocyte marker. Astrocyte-enrichedcultures comprised more than 95% GFAP-positivecells. Most experiments were carried out 14 days afterseeding on the plate. For the preparation of rat corticalastrocytes, animal experiments were performed afterreceiving approval of the Institutional Animal Care andUse Committee (IACUC) in Seoul National UniversityCollege of Medicine, and National Research Council(NRC) guideline for the care and use of laboratoryanimals were observed.
Human T-lymphocyte Jurkat cells were grownin RPMI 1640 medium supplemented with 10%heat-inactivated FBS, 100 mg/ml streptomycin, and100 U/ml penicillin in a 5% CO2 incubator at 37 8C.For the treatment of TPA, Jurkat cells were treatedwith 1–50 nM concentration of TPA dissolved inacetone.
Heat Shock Treatment
Culture plates containing cells were tightlywrapped with parafilm and immersed in a water bathat 43� 0.2 8C for 30min, and then recovered atCO2 cellculture incubator at 37� 0.2 8C for the indicated timeperiods or prepared cell lysate immediately.
RF Exposure System
The exposure system specifically designed for thisstudy is a rectangular-cavity-type. For the PCS system,a real CDMA signal at 1762.5 MHz (channel 250) wasapplied. A real CMDA signal is applied to the chamberafter amplification as shown in Figure 1A.We utilized amicroprocessor unit (MPU) chip in which the CDMAformat code is written. The MPU, in turn, controls a
RFand Stress Responses 579

CPU to generate the real CDMA test signal. Themaximum input power is 30 W. Exposure level andschedule can be controlled by computer manipulation.The exposure system is fed with a lg/8 monopoleantenna at the lg/4 position of the top plate of the cavityin order to excite only the fundamental TE102mode, andit allows the uniform exposure of one 100 mm (innerdiameter: 85mm) culture plate. The external dimensionof the chamber is 115� 80� 240 mm and monopoleantenna length is 22 mm. The chamber is made ofaluminum,which acts as an electromagnetic shield. Theexposure system is especially designed for properenvironmental conditions like ventilation, humidity,and temperature. TomaintainCO2density and humidityinside the chamber, the gas from an incubator iscirculated through the chamber. Also a cooling devicefor circulating cooling water through the bottom of thecavity is used to prevent the temperature rise of culturemedium.
Specific absorption rate (SAR) inside the exposedmedium has been characterized by numerical simu-lations, using the finite difference time domain (FDTD)method (XFDTD, Remcom, Inc., State College, PA,USA), and also by measurement, using the LUXTRONfluoroptic thermal probe 790 with thermal resolution of0.1 8C. In themeasurement, probewas located and at thenine points shownFigure 1B.Wemeasured temperaturerise at nine positions inside of the Petri dish, usingfluoroptic temperature probes (model 790, LuxtronCorp., Santa Clara, CA, USA). Figure 1C,D show theSAR measurements using a temperature probe and thegraphs of temperature rise in a cell culture, respectively.The SAR values are calculated from the temperaturerise using the well-known equation given below.
SAR ¼ Cp
DTDt
� Cp
dT
dtð1Þ
where Cp is specific heat [J/kg�8C], T is rise intemperature by exposure [8C], and t is exposuretime [s].
For FDTD simulation, cell size wasDx¼Dy¼Dz¼ 1mm, and in the source region, subgridof Dx¼Dy¼Dz¼ 0.3 mm was used. The innerdiameter of the 100 mm Petri dish is 85 mm, and thethickness of the wall is 1 mm. The inner diameter of thelid is 91 mm. The quantity of culture medium is 18 mland it corresponds to 4 mm height in the Petri dish. The
electrical properties of the materials are given inTable 1. Some details of the modeling are given inFigure 1E. The results of measurement and simulationare summarized in Table 2. All the values in Table 2 arethe SAR values at each position normalized to the inputpower at the feeding point, that is the input connector.The results of FDTD simulation reveal an excellentagreement with the measurement data. Thus, SARvalues for exposure experiment can be obtained by thenormalized SAR values given in Table 2 by the inputpower. The reported SAR value (2 and 20 W/kg)corresponds to the mean of SAP values that wereobtained at three points (Fig. 1B) in radial directionfrom center to edge.
RF Radiaiton Exposure
Cells were exposed to 1763 MHz RF radiation,either alone or in combination with TPA, in 100 mmPetri dishes, at a cell concentration of 5� 105 cells/ml.Petri disheswere placedwithin the exposure chamber atthe position shown in Figure 1B. The exposure systemwas then warmed up for 30 min, to equilibrate it beforeexposure toRF. RF radiation exposurewas conducted atSAR values of 2 W/kg or 20 W/kg. During exposure,the temperature in the chamber was maintained at37� 0.2 8Cby circulatingwater within the cavity. Afterexposure of 30 min or 1 h, the cells were immediatelyharvested or transferred to a cell culture incubator forthe indicated time and then harvested. Harvested cellswere washed with ice-cold phosphate buffered saline(PBS), then stored at�70 8C until further analysis. Forthe sham exposure, the cells were incubated in the RFradiation device, but were not exposed to RF radiation.
Western Blotting
After washing with chilled PBS, cells weresuspended with extraction buffer (1% Triton X-100,0.5% sodium deoxycholate, 20 mM Tris-HCl, pH 7.5,12 mM b-glycerophosphate, 150 mM NaCl, 5 mMEGTA, 10 mM NaF, 3 mM DTT, 1 mM sodiumorthovanadate, 1 mM phenymethysulfonyl fluoride,20 mg/ml aprotinin) and incubated on ice for 30 min.The lysate was centrifuged and quantitated with BCAProtein Assay Reagents. The equal amounts of totalproteins were separated on a 10% SDS–PAGE gel.After transfer of proteins to nitrocellulose membrane,the membrane was blocked with 5% (w/v) non-fat dry
Fig. 1. Exposuresystem.A:Blockdiagramoftheexposuresystem.B:Cavitystructureandmeasure-ment points in the culture plate.C: The photos of the exposure chamber for PCS frequency.The leftphoto is the set up for the SARmeasurement using a temperature probe. a:Exposure chamber, b:Signalgenerator, c:Signalamplifier, d:Computer, e: Temperatureprobe.The right photo shows theexposure chamber with a Petri dish in it. D: The graphs of the temperature rise in a cell culture.E:Cross-sectionalviewof theexposuresystemfornumericalsimulation.
580 Lee et al.
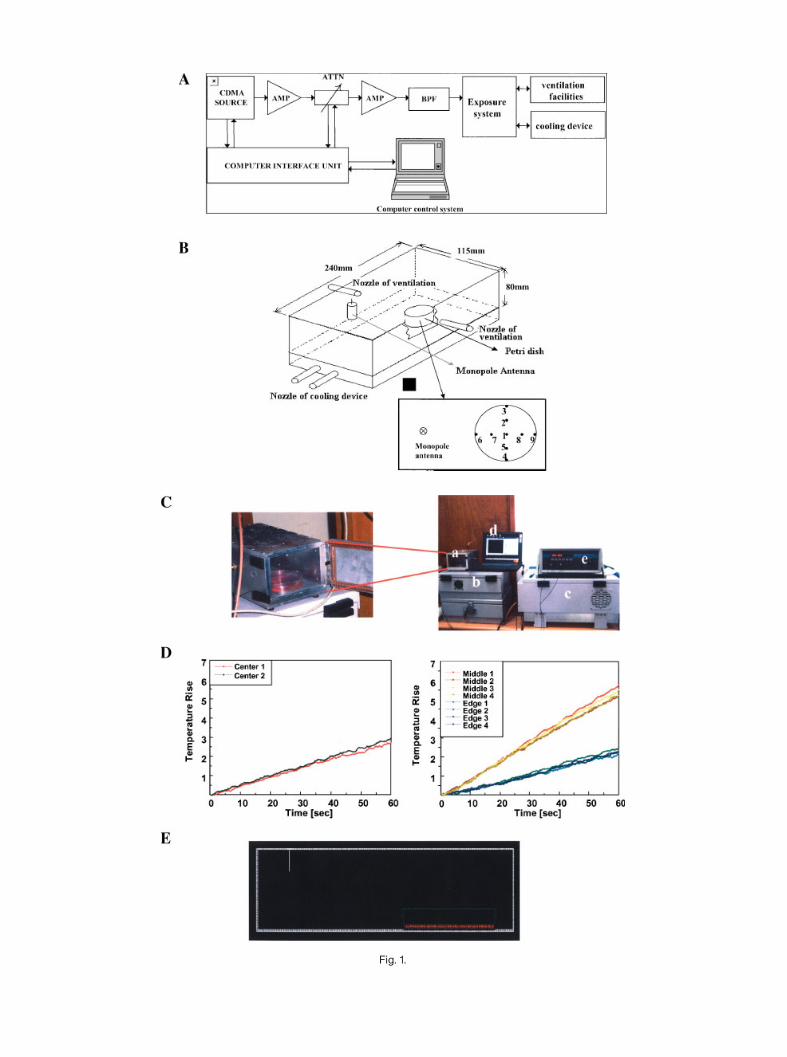
Fig. 1.

milk in 1� PBS overnight at 4 8C. Each of protein wasdetected with its specific antibody. The antibody-antigen complex was detected using horseradishperoxidase-conjugated secondary antibody, and visual-ized by an ECL detection kit according to themanufacturer’s instruction. Signal intensities werequantified by scanning densitometry. The signal wasscanned with EPSON Perfection 1660 Photo and thesignal intensity was determined with ImageJ Program(http://rsb.info.nih.gov/ij/). Figures show the level ofeach density as a ratio after normalization with internalcontrol density.
Statistical Analysis
Data are expressed as mean� standard deviation(M� SD). Differences between control and exposedgroups were analyzed using a Wilcoxon’s ranktest. Values of P<.05 were considered statisticallysignificant.
RESULTS
HSP Expression in Human T-Lymphocytes
As a positive control experiment, human T-lymphocyte Jurkat cells were exposed to heat shock(43� 0.2 8C, 30 min), which is known to be the mostgeneral stressor for the induction of HSPs. HSP70 andHSP27 expression levels in the heat-treated cells werefound to have dramatically increased after 12 h and 24 hrecovery time intervals, as compared to those in theunstressed negative control cells (Fig. 2A). There wasno detectable change in the levels of HSP90 expressionin the cells. To determinewhether RF exposure inducesthe expression of HSPs, the cells were exposed to1763MHzRF radiation at a SARof 2W/kg for 1 h, thenrecovered in a 37 8C cell culture incubator for theindicated time periods. HSP levels were assessed viaWestern blot analyses after recovery. No detectablealterations were observed in the levels of HSP27,HSP70, and HSP90 expression, as compared to thelevels measured in the sham-exposed cells, at 6, 12, or24 h after recovery (Fig. 2A).
To evaluate the possibility that HSPs expressionoccurred as the result of exposure to RF radiation ofhigher SAR values, the cells were exposed to 20 W/kgof RF radiations for 1 h. No apparent effect of RFexposure at a SAR of 20 W/kg on HSPs induction was
observed in the RF-exposed Jurkat cells (Fig. 2A). Thisexperiment was performed three times independentlywith identical results. Figure 2 provides the representa-tive data. b-actin was used as an internal loadingcontrol. For quantitative analysis, the level of proteinexpressionwasmeasured by scanning densitometry andnormalized with internal control protein. Each bar inFigure 2A shows the mean of the ratio of threeindependent experiments together with the standarddeviation (SD). No significant difference among groupswas indicated using Wilcoxon’s rank test (P>.05). Inorder to discern any non-thermal effects of RFexposure, all experiments in this study were conductedunder carefully controlled temperature conditions(37� 0.2 8C).
Phosphorylation of ERK, JNK, and p38 inHuman T-Lymphocytes
The phosphorylation of MAPKs was assessedto determine whether MAPKs were involved in anypossible RF-induced stress responses. Jurkat cells wereexposed for 0.5 or 1 h, either with or without a 1 or 2 hrecovery period. We prepared lysates from the shamand RF exposed cells and conducted Westernblot analyses using anti-phospho MAPK antibodies.ERK1/2, JNK1/2, and p38 phosphorylations were notdetected in the sham- or RF-exposed cells (Fig. 2B).This experiment was conducted independently threetimes, andMAPKs phosphorylationwas not detected inthe sham and RF exposed cells. Representative data areshown in Figure 2B. The whole cell lysates of the heat-treated Jurkat cells (43� 0.2 8C, 30 min) wereemployed as positive controls in the detection ofERK, JNK, and p38 phosphorylation.
HSPs Induction and MAPKs Activation inRat Primary Astrocytes
To determinewhether RF-induced stress responsecould be detected in different cell types, we conductedan identical set of experiments using rat primaryastrocytes. Levels of HSPs expression and MAPKsphosphorylation in the RF-exposed rat primary astro-cytes were analyzed, as shown in Figure 3. We detectedno changes in the level of HSP90,HSP70, andHSP27 in
TABLE 1. Electrical Properties of the Materials
1763 MHz Polystyrene (Petri dish) Culture medium
er 2.6 79.5s 0 2.0
TABLE 2. Comparison of Simulation and MeasurementResults at PCS Band (1763 MHz)
Position SAR [W/kg/W]
ClassificationCenter(point 1)
Middle (points2, 5, 7, 8)
Edge (points3, 4, 6, 9)
Simulation 7.9 10.4 5.2Measurement 5.1 10.7 5.5
582 Lee et al.

Fig. 2. Effects of RF exposure on HSPs level and MAPKs phos-phorylation in Jurkat cells. A: Western blot analysis of HSPsexpression in Jurkat cells exposed to RF radiation at either 2 or20W/kg for1h, thenrecovered for 6,12, or 24hina 37 8Ccellcultureincubator. Representative data from three independent experi-ments are shown. HS (positive control) consisted of heat-treated(43 8C,30min)cellswith12or24hofrecoveryat37 8C.Thenegativecontrol group (NC) received no treatment.S and E stand for shamandexposed, respectively.B:Westernblot analysisofphosphory-lated MAPKs in lysates prepared immediately after exposure orafter the indicated recovery periods. E 0.5 h and R 0 h indicateextracts prepared immediately after 30 min or1h of RF exposurewithout recovery, respectively. R1h and R 2 h extracts were pre-pared fromrecovered cells for1hand 2 hafter1hof RF-exposure,respectively.Representative data from three independent experi-mentsareshown.TheHS (positivecontrol) lysates,werepreparedimmediatelyafter theapplicationofheat shock (43 8C,30min).Thenegative control (NC) receivedno treatment.SandEdenoteshamandexposedcells, respectively.
Fig. 3. Effects of RF exposure on HSPs levels and MAPKs phos-phorylation in rat primary astrocytes. A: Western blot analysis ofHSP expression in cells exposed to RF radiation at either 2 or20W/kg for1h, thenallowedtorecover for 6,12, or 24 hina 37 8Ccellculture incubator.Therepresentative data from three independentexperimentsareshown.SandEstandforshamandexposedcells,respectively.The HS (positive control) were heat-treated lysates(43 8C,30 min) prepared following 24 h of recovery ina cell cultureincubatorat 37 8C.The controlgroup (Con) receivedno treatment.B:Westernblot analysisof phosphorylatedMAPKsin lysatespre-paredimmediatelyafterRFexposure, orafterrecovery for theindi-cated periods. E 0.5 h and R 0 h extracts were preparedimmediatelyafter 30minor1hof RFexposurewithout recovery.R1handR2hextractswerepreparedfromcellsthatrecoveredfor1hand2hafter1hofRF-exposure, respectively.Representativedatafrom three independent experiments are shown.The HS (positivecontrol)washeat-treated (43 8C,30min).SandEdenoteshamandexposedcells.
RFand Stress Responses 583

the RF-exposed cells at either 2 W/kg or 20 W/kg(Fig. 3A). No significant alteration in the phosphor-ylation status of ERK1/2, JNK1/2, and p38 wasdetected in any of the RF-exposed samples (2 W/kgand 20 W/kg) (Fig. 3B). In the case of the rat primaryastrocytes, we observed basal ERK1/2 phosphorylationin the sham-exposed cells. However, in contrast to whatwas observed in the heat-treated astrocytes (43� 0.2 8C,30 min) used as positive controls, we detected noadditional phosphorylation in the RF-exposed cells, ateither of the tested SAR values.
Figure 3 shows the representative Western blotsand quantitative analyses obtained in three independentexperiments. For quantitative analysis, the level ofprotein expression was measured by scanning densi-tometry and normalized with internal control protein(actin for HSPs, unphosphorylated ERK for phosphory-lated ERK). Each bar in graphs denotes the mean of theratio of three independent experiments together withthe SD. No significant difference among groups wasindicated using Wilcoxon’s rank test (P>.05).
Co-Treatment With RF and TPA on MAPKActivation and HSP Expression in Jurkat Cells
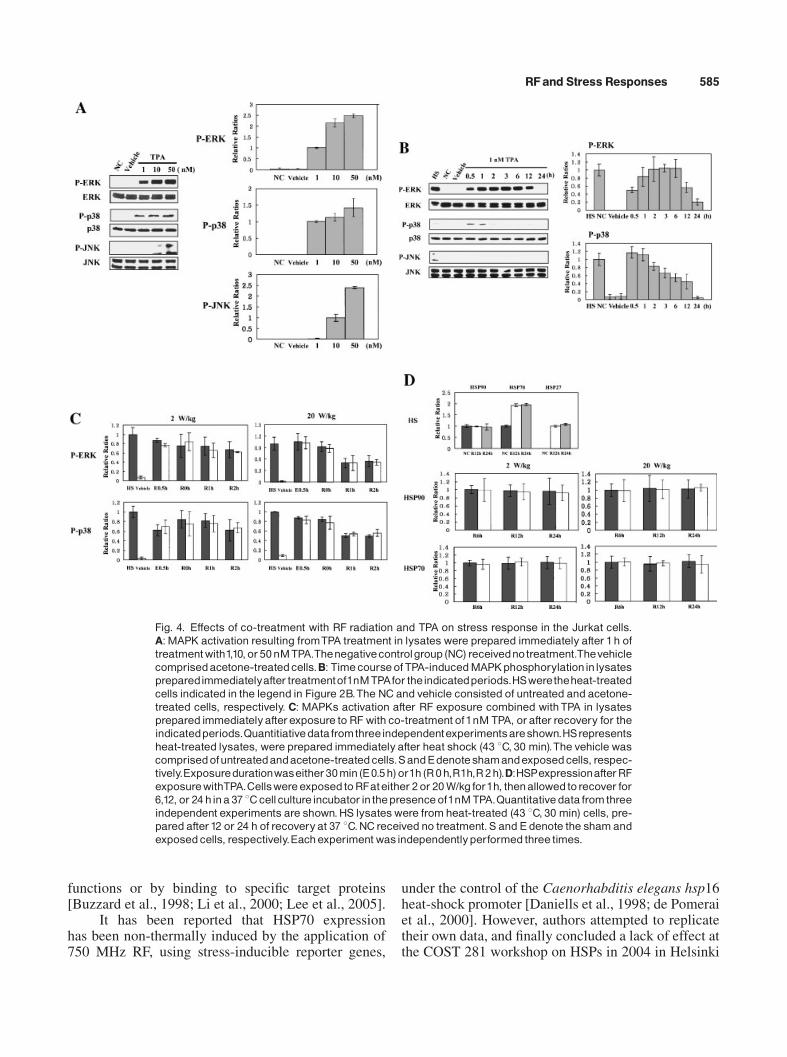
Since we detected no difference in the levels ofHSPs expression and MAPKs phosphorylation underour experimental conditions,we attempted to determinewhether RF exposure exerts a synergistic effect onTPA-induced stress responses. First, we assessed thephosphorylation status of MAPKs in the TPA-treatedJurkat cells at several concentrations (Fig. 4A).ERK1/2, JNK1/2, and p38were found to be phosphory-lated in a dose-dependent manner. MAPK phosphory-lation was not found to be saturated at 1 nM of TPA.Therefore, in order to detect the synergistic effects ofRF exposure on TPA-induced MAPK phosphorylation,we decided to expose the cells to RF radiation, coupledwith 1 nM TPA.
When we observed the status of MAPK phos-phorylation for up to 24 h in 1 nM TPA-treated cells,ERK1/2 and p38 phosphorylation achieved maximumlevels within 1 h (Fig. 4B). JNK1/2 phosphorylationwas not detected after treatment with 1 nM TPA. TheJurkat cells were exposed to 2 W/kg or 20 W/kg ofRF radiation, coupled with 1 nM TPA, and MAPKphosphorylation was analyzed in the sham and RFexposed cells. We noted no discernable alteration inthe status of MAPK phosphorylation (Fig. 4C). Thisindicated that RF exposure exerts no synergistic effectson TPA-induced MAPKs phosphorylation. NeitherTPA alone nor RF exposure coupled with TPA resultedin any discernible changes in the levels of HSP90,HSP70, and HSP27 expression (Fig. 4D). In contrastheat-treated (43 8C, 30min) positive control cells (HS),
neither sham exposed nor RF exposed samples inducedany HSP27 expression, that is, no basal level ofexpression. We did not take the quantitative data ofHSP27. These experiments were repeated three timesindependently.
For quantitative analysis, the level of proteinexpressionwasmeasured by scanning densitometry andnormalized with the internal control protein. Each bardenotes the mean of the ratio of three independentexperiments together with the SD. No significantdifference among groups was indicated using Wilcox-on’s rank test (P>.05).
DISCUSSION
We face a continuously increasing amount ofenvironmental and pathophysiological stresses in ourmodern world. As the number of mobile phone usershas increased substantially during the last 10 years, RFradiation from mobile phones and base stations hasbecome a primary source of environmental RFradiation. Therefore, there has been growing publicconcern with regard to whether RF radiation frommobile phones might constitute an environmentalstressor.
When cells are exposed to stressful conditions, thecells trigger appropriate protective mechanisms inresponse to the stress. This conserved cellular responseto harmful stimuli is generally termed as ‘stressresponse.’ Stress response modulates a variety ofintracellular processes, via the alteration of geneexpressions and protein activities, in a process ofadaptation to new stressful conditions. The induction ofHSPs and the activation of MAPKs are the best-characterized stress responses.
HSPs perform important functions as chaperoningmolecules, which are essential for proper proteinfolding and assembly. HSPs have also been employedas sensitive stress response markers due to theirimmediate induction upon exposure to a variety ofstressors [Morimoto, 1992; Lee and Seo, 2002]. Therapidly-induced HSPs play cytoprotective roles againstcellular damage.HSP90,HSP70, andHSP27 are knownparticularly for their apoptosis inhibitory qualities.Apoptosis can be induced by a variety of differentsignals, including the activation of death receptors,excessive DNA damage, or stresses including heatshock, hyperosmotic shock, or UV irradiation [Weiet al., 1994; Martin et al., 1995]. Apoptotic malfunc-tions have been previously implicated in humandiseases, including cancer, neurodegenerative disorders,and ischemic stroke [Jacobson et al., 1997; Price et al.,1998]. HSPs can prevent apoptotic cell death at severalsteps of apoptosis, via the exertion of their chaperoning
584 Lee et al.
functions or by binding to specific target proteins[Buzzard et al., 1998; Li et al., 2000; Lee et al., 2005].
It has been reported that HSP70 expressionhas been non-thermally induced by the application of750 MHz RF, using stress-inducible reporter genes,
under the control of the Caenorhabditis elegans hsp16heat-shock promoter [Daniells et al., 1998; de Pomeraiet al., 2000]. However, authors attempted to replicatetheir own data, and finally concluded a lack of effect atthe COST 281 workshop on HSPs in 2004 in Helsinki
Fig. 4. Effects of co-treatment with RF radiation and TPA on stress response in the Jurkat cells.A:MAPK activation resulting fromTPA treatment in lysateswere prepared immediately after 1h oftreatmentwith1,10,or50nMTPA.Thenegativecontrolgroup (NC) receivednotreatment.Thevehiclecomprisedacetone-treatedcells.B: TimecourseofTPA-inducedMAPKphosphorylationinlysatespreparedimmediatelyafter treatmentof1nMTPAfor theindicatedperiods.HSweretheheat-treatedcells indicated in the legend in Figure 2B.The NC and vehicle consisted of untreated and acetone-treated cells, respectively. C: MAPKs activation after RF exposure combined withTPA in lysatesprepared immediatelyafterexposure to RF with co-treatment of1nM TPA, orafter recovery for theindicatedperiods.Quantitiativedatafromthreeindependentexperimentsareshown.HSrepresentsheat-treated lysates, were prepared immediatelyafter heat shock (43 8C, 30 min).The vehicle wascomprisedofuntreatedandacetone-treatedcells.SandEdenoteshamandexposedcells, respec-tively.Exposuredurationwaseither30min(E0.5h)or1h(R0h,R1h,R2h).D:HSPexpressionafterRFexposurewithTPA.CellswereexposedtoRFateither 2or 20W/kg for1h, thenallowedtorecover for6,12, or 24 hina 37 8Ccellcultureincubator inthepresenceof1nMTPA.Quantitativedata fromthreeindependent experiments are shown.HS lysateswere fromheat-treated (43 8C, 30 min) cells, pre-pared after12 or 24 h of recoveryat 37 8C.NCreceived no treatment.S and E denote the shamandexposedcells, respectively.Eachexperiment wasindependentlyperformed three times.
RFand Stress Responses 585

(http://www.Cost281.org/). Weisbrot et al. [2003]demonstrated that non-thermal radiation from GSMmobile phones elicited a rapid elevation in HSP70 levelwith Drosophila melanogaster. The level of HSP70 inchick embryos was found to increase after exposure to915 MHz radiation [Shallom et al., 2002].
However, no HSP induction or decreases in thelevel of HSP70 has been reported as the result of EMFexposure as in other reports. Fritze et al. [1997]suggested that transient and slight induction of hsp70and c-fos mRNAs might be related with the restraint ofrats, rather than to the effects of RF-EMF. Cleary et al.[1997] observed that RF EMF exerted no discernableeffect on the induction of HSPs in mammalian cells.Exposure to 1800 MHz RF radiation exerted nodiscernable effect on the level of HSP70 in humanblood cells [Capri et al., 2004]. In another study, nosignificant difference in HSP70 and HSP27 expressionswas detected after exposure to 900 MHz RF fields inhuman leukocytes [Lim et al., 2005].
In contrast, HSP70 has been induced by theexposure of human cells to a 2.45 GHz EMF at a highSAR value of more than 20 W/kg [Tian et al., 2002].These reports indicate that questions regarding thepositive, negative, or null effects of RF radiation remaindebatable. A number of tumor classes have beenreported to exhibit altered HSP expression profiles[Lazaris et al., 1997; Morino et al., 1997; Jolly andMorimoto, 2000]. The possibility of a direct associationbetween the continuous expression of HSP70 andcancer has been suggested [French et al., 2001].Therefore, it is vital to determine whether the non-thermal effects of RF radiation are sufficient to inducethe expression of HSP70.
MAPKs can be activated by a variety of stressors,including radiation, free radicals, heat, osmotica, andcytokines. These kinases carry stress signals to thenucleus, via the activation of transcription factors. Thisultimately results in the alteration of intracellularstatus, allowing the cell to defend against or adapt toextracellular stresses [Johnson and Lapadat, 2002]. IfRF radiation induces stress responses, MAPK activitiesmight change as the result of exposure to RF radiation.Weisbrot et al. [2003] reported an increase of severalfolds in the phosphorylation of ELK-1, the downstreamkinase of ERK1/2, upon the exposure of D. mela-nogaster to RF radiation. A significant decrease in PKClevels was also observed in rats exposed to RF EMF intheir brain [Paulraj and Behari, 2004]. Transientincreases in HSP27 phosphorylation and expression,as well as the expression of p38, have also been recentlyreported to be elicited by exposure to mobile phoneradiation in human endothelial cells [Leszczynski et al.,2002]. There have also been some reports concerning
other kinases involved in signaling pathways. MAPKkinase 3 expression was increased upon exposure to theRF radiation emitted by aGSMcellular phone in humanskin fibroblasts [Pacini et al., 2002].
As a research group in the field of stress biology,we previously elucidated the cytoprotective functionsof HSP70 as a natural apoptosis inhibitor [Li et al.,2000; Lee et al., 2005]. We also generated hsp70.1-deficient mice and reported cellular damage in the renaltissue to the response of osmotic stress [Shim et al.,2002]. We conducted a series of experiments todetermine whether RF radiation exerts adverse healtheffects, using hsp70.1-deficient mice.
In order to address the issue as to whether RFradiation functions as an environmental stressor, weattempted in this study to assess the induction ofstress responses upon exposure to 1763 MHz RFradiation. Human T-lymphocyte Jurkat cells and ratprimary astrocytes were exposed to SAR levels ofeither 2 W/kg or 20 W/kg. The observed discrepanciesin HSPs induction andMAPKs activation referenced inprevious reports might be attributable to the thermaleffects associated with the administration of highlevels of SAR. In order to eliminate the possibilitythat high SAR levels elicit stress responses viathermal effects, this study was conducted underisothermal conditions. Cooling water was circulatedthroughout the cavity during exposure, in order tomaintain a temperature of 37� 0.2 8C. Temperature ofthe culture media was monitored twice a secondthroughout the exposure period, at either 2 W/kg or20 W/kg. Under our experimental conditions, when wemonitored the induction of HSPs in the RF-exposedcells, we observed no detectable alteration in theexpression of HSPs.
We also assessed the levels of phosphorylation ofMAPKs, that is, ERK1/2, JNK1/2, and p38, in the RF-exposed cells. No MAPKs exhibited any discernablealterations in activity. TPA, a well-known activator ofprotein kinase C, regulates a variety of cell signalingmolecules, including NF-KappaB and MAPKs, andalso mediates cell transformation, apoptosis, and cellproliferation [Afaq et al., 2005]. We attempted todetermine whether RF radiation synergistically affectsthe status of TPA-induced MAPK phosphorylation. RFexposure, coupled with TPA treatment, exerted noadditive effect on the phosphorylation of MAPKs. Weobserved no detectable changes in the levels of anystress response molecules in the human T-lymphocytesand rat primary astrocytes due to the exposure toRF radiation.
As we failed to find any detectable change in theexpression of stress proteins and the activation ofstress-activated protein kinases, we are currently
586 Lee et al.

conducting an experiment involving oligonucleotidechip assays to profile gene expression patterns whichhave been altered by exposure to RF radiation. The useof high-throughput screening techniques might proveuseful with regard to the analysis of global changes ingene expression due to RF radiation, as was previouslysuggested by Leszczynski et al. [2004].
In conclusion, the present work shows that RFradiation exposure did not affect onHSP expression andMAPK phosphorylation in both human T-lymphocytesand rat primary astrocytes. Different results have beenobtained regarding the effect of RF exposure on stressresponse as shown in Table 3. There is still an obviousdemand for exploring this issue. For further inves-tigation, researchers have to pay more attention to theexperimental and exposure conditions.
REFERENCES
Afaq F, Saleem M, Krueger CG, Reed JD, Mukhtar H. 2005.Anthocyanin- and hydrolyzable tannin-rich pomegranatefruit extract modulatesMAPK andNF-kappaB pathways andinhibits skin tumorigenesis in CD-1 mice. Int J Cancer113(3):423–433.
Ali A, Krone PH, Pearson DS, Heikkila JJ. 1996. Evaluation ofstress-inducible hsp90 gene expression as a potentialmolecular biomarker in Xenopus laevis. Cell Stress Chap-erones 1:62–69.
Buzzard KA, Giaccia AJ, Killender M, Anderson RL. 1998. Heatshock protein 72 modulates pathways of stress-inducedapoptosis. J Biol Chem 273:17147–17153.
Capri M, Scarcella E, Bianchi E, Fumelli C, Mesirca P, Agostini C,Remondini D, Schuderer J, Kuster N, Franceschi C, BersaniF. 2004. 1800MHz radiofrequency (mobile phones, differentglobal system for mobile communication modulations) doesnot affect apoptosis and heat shock protein 70 level inperipheral blood mononuclear cells from young and olddonors. Int J Radiat Biol 80:389–397.
Cleary SF, Cao G, Liu L-M, Egle PM, Shelton KR. 1997. Stressproteins are not induced in mammalian cells exposed toradiofrequencyormicrowave radiation. Bioelectromagnetics18:499–505.
Daniells C, Duce I, Thomas D, Sewell P, Tattersall J, de Pomerai D.1998. Transgenic nematodes as biomonitors of microwave-induced stress. Mutat Res 399:55–64.
de Pomerai D, Daniells C, David H, Allan J, Duce I, Mutwakil M,Thomas D, Sewell P, Tattersall J, Jones D, Candido P. 2000.Non-thermal heat-shock response to microwaves. Nature405: 417–418.
Di Carlo A, White N, Guo F, Garrett P, Litovitz T. 2002.Chronic electromagnetic field exposure decreases HSP70levels and lowers cytoprotection. J Cell Biochem 84:447–454.
French PW, Penny R, Laurence JA, McKenzie DR. 2001. Mobilephones, heat shock proteins and cancer. Differentiation 67:93–97.
Fritze K, Wiessner C, Kuster N, Sommer C, Gass P, Hermann DM,Kiessling M, Hossmann KA. 1997. Effect of global systemfor mobile communication microwave exposure on thegenomic response of the rat brain. Neuroscience 81:627–639.T
ABLE3.Summary
ofStudiesRegardingRFRadiation-InducedStressResponses
System
employed
RF-radiationfrequency
SARorpower
Durationofexposure
Observed
molecule
Effectonstress
response
Reference
Invivo
Rat
GSM
0.3,1.5,7.5
W/kg
4h
Hsp70,c-jun,c-fos
Negative
Fritzeet
al.[1997]
C.elegans
750MHz
27dBm
0–16h
HSP16
Positivea
Daniellset
al.[1998]
C.elegans
750MHz
0.5
W18h
HSP16
Positivea
dePomerai
etal.
[2000]
Chickem
bryo
915MHz
1.75,2.5
W/kg
30min
HSP70
Positive
Shallom
etal.[2002]
Chickem
bryo
60Hzþ915MHz
8mT
þ3.5
mW
20,30,60min
HSP70
Positive
DiCarlo
etal.[2002]
D.melanogaster
900/1,900MHz
�1.4
W/kg
HSP70,ELK-1
Positive
Weisbrotetal.[2003]
Rat
112MHz
1.48W/kg
2h/day
for35days
Protein
kinaseC
Positive
Paulraj
andBehari
[2004]
Invitro
Mam
maliancells
27/2450MHz
25,100W/kg
2h
HSP70
Negative
Clearyet
al.[1997]
Human
skin
fibroblasts
GSM
1h
MAPkinasekinase3
Positive
Paciniet
al.[2002]
Human
gliomacells
2450MHz
25,78W/kg
2h
HSP70
Negative
Tianet
al.[2002]
Human
endothelialcells
900MHz
2W/kg
1h
HSP27,p38MAPK
Positive
Leszczynskiet
al.
[2002]
Human
peripheral
bloodmononuclearcells
1800MHz
1.4,2.0
W/kg
44h(10on/20off)
HSP70
Negative
Capriet
al.[2004]
Human
endothelialcells
900MHz
2.4
W/kg
1h
HSP27
Positive
Leszczynskiet
al.
[2004]
Human
leukocytes
900MHz
0.4,2.0,3.6
W/kg
20min,1h,4h
HSP27,HSP70
Negative
Lim
etal.[2005]
aNotreproducible(http://www.Cost281.org).
RFand Stress Responses 587

Galve-Roperh I, Haro A, Dı́az-Laviada I. 1997. Induction of nervegrowth factor synthesis by sphingomyelinase and ceramidein primary astrocyte cultures. Brain Res Mol Brain Res 52:90–97.
Ip YT, Davis RJ. 1998. Signal transduction by the c-Jun N-terminalkinase (JNK)—from inflammation to development. CurrOpin Cell Biol 10:205–219.
Jacobson MD, Weil M, Raff MC. 1997. Programmed cell death inanimal development. Cell 88:347–354.
Johnson GL, Lapadat R. 2002. Mitogen-activated protein kinasepathways mediated by ERK, JNK, and p38 protein kinases.Science 298:1911–1912.
Jolly C, Morimoto RI. 2000. Role of the heat shock response andmolecular chaperones in oncogenesis and cell death. J NatlCancer Inst 92:1564–1572.
Lazaris ACh, Chatzigianni EB, Panoussopoulos D, Tzimas GN,Davaris PS, Golematis BCh. 1997. Proliferating cell nuclearantigen and heat shock protein 70 immunolocalization ininvasive ductal breast cancer not otherwise specified. BreastCancer Res Treat 43:43–51.
Lee J-S, Seo J-S. 2002. Differential expression of two stress-inducible hsp70 genes by various stressors. Exp Mol Med34:131–136.
Lee J-S, Lee J-J, Seo J-S. 2005. HSP70 deficiency results inactivation of c-jun N-terminal kinase, extracellular signal-regulated kinase, and caspase-3 in hyperosmolarity-inducedapoptosis. J Biol Chem 280:6634–6641.
Leszczynski D, Joenvaara S, Reivinen J, Kuokka R. 2002. Non-thermal activation of the hsp27/p38MAPK stress pathway bymobile phone radiation in human endothelial cells: Molec-ular mechanism for cancer- and blood-brain barrier-relatedeffects. Differentiation 70:120–129.
Leszczynski D, Nylund R, Joenvaara S, Reivinen J. 2004.Applicability of discovery science approach to determinebiological effects of mobile phone radiation. Proteomics 4:426–431.
Li C-Y, Lee J-S, KoY-G, Kim J-I, Seo J-S. 2000. Heat shock protein70 inhibits apoptosis downstream of cytochrome c releaseand upstream of caspase-3 activation. J Biol Chem 275:25665–25671.
Lim HB, Cook GG, Barker AT, Coulton LA. 2005. Effect of 900MHz electromagnetic fields on nonthermal induction of heat-shock proteins in human leukocytes. Radiat Res 163:45–52.
Martin SJ, Newmeyer DD, Mathias S, Farschon DM, Wang HG,Reed JC, Kolesnick RN, Green DR. 1995. Cell-freereconstitution of Fas-, UV radiation- and ceramide-inducedapoptosis. EMBO J 14:5191–5200.
Minden A, Karin M. 1997. Regulation and function of the JNKsubgroup of MAP kinases. Biochim Biophys Acta 1333:F85–F104.
Morimoto RI. 1992. Transcriptional regulation of heat shock genes:A paradigm for inducible genomic responses. J Biol Chem267:21987–21990.
Morino M, Tsuzuki T, Ishikawa Y, Shirakami T, Yoshimura M,Kiyosuke Y, Matsunaga K, Yoshikumi C, Saijo N. 1997.Specific expression of HSP27 in human tumor cell linesin vitro. In Vivo 11:179–184.
MuschMW, Petrof EO, Kojoma K, Ren H,McKay DM, Chang EB.2004. Bacterial superantigen-treated intestinal epithelialcells upregulate heat shock proteins 25 and 72 and areresistant to oxidant cytotoxicity. Infect Immun 72:3187–3194.
Pacini S, Ruggiero M, Sardi I, Aterini S, Gulisano F, Gulisano M.2002. Exposure to global system for mobile communication(GSM) cellular phone radiofrequency alters gene expression,proliferation, and morphology of human skin fibroblasts.Oncol Res 13:19–24.
Paulraj R, Behari J. 2004. Radio frequency radiation effects onprotein kinase C activity in rats’ brain. Mutat Res 545:127–130.
Price DL, Sisodia SS, Borchelt DR. 1998. Genetic neurodegener-ative diseases: The human illness and transgenic models.Science 282:1079–1083.
Schaeffer HJ, Weber MJ. 1999. Mitogen-activated protein kinases:Specific messages from ubiquitous messengers. Mol CellBiol 19:2435–2444.
Shallom JM,DiCarloAL,KoD, PenafielLM,NakaiA, Litovitz TA.2002. Microwave exposure induces Hsp70 and confersprotection against hypoxia in chick embryos. J Cell Biochem86:490–496.
Shim E-H, Kim J-I, Bang E-S, Heo J-S, Lee J-S, Kim E-Y, Lee J-E,Park W-Y, Kim S-H, Kim H-S, Smithies O, Jang J-J, Jin D-I,Seo J-S. 2002. Targeted disruption of hsp70.1 sensitizes toosmotic stress. EMBO Rep 3:857–861.
Tian F, Nakahara T, Wake K, Taki M, Miyakoshi J. 2002. Exposureto 2.45 GHz electromagnetic fields induces hsp70 at ahigh SAR of more than 20W/Kg but not at 5 W/kg in humanglioma MO54 cells. Int J Radiat Biol 78:433–440.
Wei YQ, Zhao X, Kariya Y, Fukata H, Teshigawara K, Uchida A.1994. Induction of apoptosis by quercetin: Involvement ofheat shock protein. Cancer Res 54:4952–4957.
Weisbrot D, Lin H, Ye L, Blank M, Goodman R. 2003. Effectsof mobile phone radiation on reproduction and developmentin Drosophila melanogaster. J Cell Biochem 89:48–55.
588 Lee et al.




















