
BAGE: a new gene encoding an antigen recognized on human melanomas by cytolytic T lymphocytes
9
Immunity, Vol. 2, 167-175, February, 1995, Copyright © 1995 by Cell Press BAGE: a New Gene Encoding an Antigen Recognized on Human Melanomas by Cytolytic T Lymphocytes Pascale BoiH, * Claude Wildmann,* Maria Luisa Sensi,t Robert Brasseur,~ Jean-Christophe Renauld,* Pierre Coulie,* Thierry Boon,* and Pierre van der Bruggen* • Ludwig Institute for Cancer Research Brussels Branch 74 avenue Hippocrate Universite Catholique de Louvain 74.59 and Cellular Genetics Unit Universite Catholique de Louvain B1200 Brussels Belgium flstituto Nazionale per Io Studio e la Cura dei Tumori Division of Experimental Oncology D 20133 Milano Italy $Centre de Biophysique Moleculaire Numer[que Facult~ des Sciences Agronomiques de Gembloux B5030 Gembloux, Belgium Summary Several tumor antigens are recognized by autologous cytolytic T lymphocytes (CTL) on human melanoma MZ2-MEL. Some of them are encoded by genes MAGE.1 and MAGE.3, which are not expressed in normal tis- sues except in testis. Here, we report the identification of a new gene that codes for another of these antigens. This gene, named BAGE, codes for a putative protein of 43 aa and seems to belong to a family of several genes. The antigen recognized by the autologous CTL consists of BAGE-encoded peptide AARAVFLAL bound to an HLA-Cw*1601 molecule. Gene BAGE is expressed in 22% of melanomas, 30% of infiltrating bladder carcinomas, 1'0% of mammary carcinomas, 8% of head and neck squamous cell carcinomas, and 6% of non-small cell lung carcinomas. Like the MAGE genes, it is silent in normal tissues with the exception of testis. Because of its tumor-specific expression, the BAGE-encoded antigen may prove useful for cancer immunotherapy. Introduction Cytolytic T lymphocytes (CTL) directed against human tu- rnor cells can often be obtained by culture of irradiated tumor cells with autologous iymphocytes (reviewed by Boon et al., 1994). From such a responder T cell population obtained against melanoma cell line MZ2-MEL, it was pos- sible to derive a large number of stable anti-tumor CTL clones, which showed specificity for the tumor cells inso- far as they did not lyse natural killer target cells, such as K562, autologous fibroblasts, and autologous Epstein- Barr virus (EBV)-transformed lymphobtastoid cells (Herin et al., 1987). These CTL clones were used to select resis- tant variants of MZ2-MEL and the analysis of these anti- gen-loss variants led to the conclusion that several distinct antigens were recognized on MZ2-MEL by autologous CTL (Van den Eynde et al., 1989). A gene transfection approach led to the identification of the gene coding for one of these antigens, namely MZ2-E. This gene, named MAGE-1, is not expressed in normal tissues except testis and it is expressed in approximately '37% of melanoma tumor samples (van der Bruggen et al., 1991; De Smet et al., 1994). Antigen MZ2-E consists of a MAGE-l-encoded nonapeptide presented by an HLA-A1 molecule (Traversari et al., 1992a). Another antigen recog- nized by CTL on melanoma MZ2-MEL is also encoded by MAGE-1 and it is presented by an HLA-Cw* 1601 molecule (van der Bruggen et al., 1994b). Gene MAGE-1 belongs to a family of at least 12 closely related genes, and another member of this gene family, namely MAGE-3, codes for a third antigen present on MZ2-M EL. Like MZ2-E, this anti- gen is presented by HLA-A1 (De Plaen et al., 1994; Gaugler et al., 1994). We report here the identification of a new gene, which codes for yet another antigen recognized on MZ2-MEL by autologous CTL. This gene is not related to the MAGE family. Results CTL 82•82 Recognizes an Antigen Presented by HLA-Cw* 1601 Melanoma cell line MZ2-MEL was derived from an abdomi- nal metastasis of patient MZ2 and clonal line MZ2-MEL.3.0 was obtained by limiting dilution. After more than 150 gen- erations in culture, this subline was found to be resistant to a large fraction of autologo us CTL clones that had strong lytic activity on MZ2-MEL.3.0. This resistant subline was named MZ2-MEL.3.1 (Figure 1A; Van den Eynde et al., 1989). The target antigen of the CTL that failed to lyse MZ2-MEL.3.1 was named MZ2-B, under the erroneous assumption that a single antigen had been lost by this subline. Subsequent analysis indicated that subline MZ2- MEL.3.1 had lost the genes coding for HLA-A29, B44, and Cw'1601 (van der Bruggen et al., 1994b). Transfection experiments with HLA-Cw* 1601 indicated that several of the anti-B CTL recognized an antigen presented by the product of this HLA-C allele (Figure 1A). Some of these CTL recognized an antigen, named Bb, that turned out to be encoded by MAGE-1 (van der Bruggen et al., 1994b). Others appeared to recognize another antigen, because they failed to recognize COS-7 cells transfected with HLA- Cw'1601 and MAGE-?. CTL 82•82 belonged to this group and its target antigen was named Ba. In addition to MZ2- MEL, CTL 82•82 also recognized two other melanoma cell lines carrying the HLA-Cw* 1601 allele. These two mela- noma cell lines stimulated tumor necrosis factor (TNF) release by CTL 82•82 (van der Bruggen et al., 1994b) and were lysed by it, albeit to a lesser extent than the MZ2-MEL cells (Figure 1B).
Transcript of BAGE: a new gene encoding an antigen recognized on human melanomas by cytolytic T lymphocytes

Immunity, Vol. 2, 167-175, February, 1995, Copyright © 1995 by Cell Press
BAGE: a New Gene Encoding an Antigen Recognized on Human Melanomas by Cytolytic T Lymphocytes
Pascale BoiH, * Claude Wildmann,* Maria Luisa Sensi,t Robert Brasseur,~ Jean-Christophe Renauld,* Pierre Coulie,* Thierry Boon,* and Pierre van der Bruggen* • Ludwig Institute for Cancer Research Brussels Branch 74 avenue Hippocrate Universite Catholique de Louvain 74.59
and Cellular Genetics Unit Universite Catholique de Louvain B1200 Brussels Belgium flstituto Nazionale per Io Studio e la Cura dei Tumori Division of Experimental Oncology D 20133 Milano Italy $Centre de Biophysique Moleculaire Numer[que Facult~ des Sciences Agronomiques de Gembloux B5030 Gembloux, Belgium
Summary
Several tumor antigens are recognized by autologous cytolytic T lymphocytes (CTL) on human melanoma MZ2-MEL. Some of them are encoded by genes MAGE.1 and MAGE.3, which are not expressed in normal tis- sues except in testis. Here, we report the identification of a new gene that codes for another of these antigens. This gene, named BAGE, codes for a putative protein of 43 aa and seems to belong to a family of several genes. The antigen recognized by the autologous CTL consists of BAGE-encoded peptide AARAVFLAL bound to an HLA-Cw*1601 molecule. Gene BAGE is expressed in 22% of melanomas, 30% of infiltrating bladder carcinomas, 1'0% of mammary carcinomas, 8% of head and neck squamous cell carcinomas, and 6% of non-small cell lung carcinomas. Like the MAGE genes, it is silent in normal tissues with the exception of testis. Because of its tumor-specific expression, the BAGE-encoded antigen may prove useful for cancer immunotherapy.
Introduction
Cytolytic T lymphocytes (CTL) directed against human tu- rnor cells can often be obtained by culture of irradiated tumor cells with autologous iymphocytes (reviewed by Boon et al., 1994). From such a responder T cell population obtained against melanoma cell line MZ2-MEL, it was pos- sible to derive a large number of stable anti-tumor CTL clones, which showed specificity for the tumor cells inso- far as they did not lyse natural killer target cells, such as K562, autologous fibroblasts, and autologous Epstein- Barr virus (EBV)-transformed lymphobtastoid cells (Herin et al., 1987). These CTL clones were used to select resis- tant variants of MZ2-MEL and the analysis of these anti-
gen-loss variants led to the conclusion that several distinct antigens were recognized on MZ2-MEL by autologous CTL (Van den Eynde et al., 1989).
A gene transfection approach led to the identification of the gene coding for one of these antigens, namely MZ2-E. This gene, named MAGE-1, is not expressed in normal tissues except testis and it is expressed in approximately
'37% of melanoma tumor samples (van der Bruggen et al., 1991; De Smet et al., 1994). Antigen MZ2-E consists of a MAGE-l-encoded nonapeptide presented by an HLA-A1 molecule (Traversari et al., 1992a). Another antigen recog- nized by CTL on melanoma MZ2-MEL is also encoded by MAGE-1 and it is presented by an HLA-Cw* 1601 molecule (van der Bruggen et al., 1994b). Gene MAGE-1 belongs to a family of at least 12 closely related genes, and another member of this gene family, namely MAGE-3, codes for a third antigen present on MZ2-M EL. Like MZ2-E, this anti- gen is presented by HLA-A1 (De Plaen et al., 1994; Gaugler et al., 1994).
We report here the identification of a new gene, which codes for yet another antigen recognized on MZ2-MEL by autologous CTL. This gene is not related to the MAGE family.
Results
CTL 82•82 Recognizes an Antigen Presented by HLA-Cw* 1601 Melanoma cell line MZ2-MEL was derived from an abdomi- nal metastasis of patient MZ2 and clonal line MZ2-MEL.3.0 was obtained by limiting dilution. After more than 150 gen- erations in culture, this subline was found to be resistant to a large fraction of autologo us CTL clones that had strong lytic activity on MZ2-MEL.3.0. This resistant subline was named MZ2-MEL.3.1 (Figure 1A; Van den Eynde et al., 1989). The target antigen of the CTL that failed to lyse MZ2-MEL.3.1 was named MZ2-B, under the erroneous assumption that a single antigen had been lost by this subline. Subsequent analysis indicated that subline MZ2- MEL.3.1 had lost the genes coding for HLA-A29, B44, and Cw'1601 (van der Bruggen et al., 1994b). Transfection experiments with HLA-Cw* 1601 indicated that several of the anti-B CTL recognized an antigen presented by the product of this HLA-C allele (Figure 1A). Some of these CTL recognized an antigen, named Bb, that turned out to be encoded by MAGE-1 (van der Bruggen et al., 1994b). Others appeared to recognize another antigen, because they failed to recognize COS-7 cells transfected with HLA- Cw'1601 and MAGE-?. CTL 82•82 belonged to this group and its target antigen was named Ba. In addition to MZ2- MEL, CTL 82•82 also recognized two other melanoma cell lines carrying the HLA-Cw* 1601 allele. These two mela- noma cell lines stimulated tumor necrosis factor (TNF) release by CTL 82•82 (van der Bruggen et al., 1994b) and were lysed by it, albeit to a lesser extent than the MZ2-MEL cells (Figure 1B).

Immunity 168
A
8o 03
~, 60 O
,+- ~ 40- o_ (,9 o~ " 20-
o
TARGET
MZ2-MEL3.0
/ .1 .3 i 3 1'o .t
B
MZ2-MEL.3.1 MZ2-MEL.B.TC.4
T . • , , , .3 1 3 10 .1 .3 1 3 10
Effector to target ratios
TARGET
MZ2-MEL.43 MI4024/1-MEL LB17-MEL 80
~>, 60
o 40- Q .
~ 2 0 - ~
O- ~ ~ .1 .3 i 3 1'0 '.1 .3 i 3 1'0 .t .3 i ~ 10
EIfector to t a rge t ra t ios
Figure 1. Lytic Activity of CTL Clone 82/82 on MZ2-MELSublinesand Other Melanoma Lines Chromium release was measured after 4 hr. (A) Subline MZ2-MEL.3.1 was obtained by extending the culture of sub- clone MZ2-MEL.3.0 for more than 150 generations. MZ2-MEL.B.TC.4 was obtained by transfecting the HLA-Cw*1601 gene into a subline derived from MZ2-MEL.3.1. (B) Clonal line MZ2-MEL.43 was derived from MZ2-MEL.3.0 and mela- noma lines MI4024/1-MEL and LB17-MEL were derived from HLA- Cw* 1601 patients.
Isolation of a cDNA Coding for Antigen MZ2-Ba We set out to identify the gene coding for the antigen recognized by CTL 82•82. Our approach involved the co- transfection of the HLA-Cw*1601 cDNA with a cDNA li- brary derived from clonal subline MZ2-MEL.43, which was also lysed by CTL 82182 (Figure 1B). The cDNAs were cloned in expression vector pcD-SR~, which contains the replication origin of SV40, and they were transfected into COS-7 cells. This vector provides considerable episomal multiplication in the transfected cells and, therefore, high expression of the antigen (Seed and Aruffo 1987; Brichard et al., 1993; Coulie et al., 1994).
The library, which contained approximately 66,000 in- serts, was divided into pools of either 400 or 200 bacteria. Each cDNA pool was cotransfected with the HLA-Cw* 1601 construct into duplicate microcultures of COS-7 cells. CTL 82•82 was added 2 days later to each transfected microcul- ture. After 24 hr, the concentration of TNF in the superna- tant was estimated bymeasuring its cytotoxic effect on cells of WEHI-164 clone 13. Among the 384 pools of cDNAs (297 of 200 bacteria and 87 of 400) that were transfected, two produced positive supernatants con-
Stimulator cells
601+ cDNA-AD5 HLA-Cw*I COS-7 cells transfected with cDNA-AD
H LA-Cw* 16
MZ2-MEL.43
MZ2-MEL.2.2.5
0 20 4'0 6'0 80 100 120 TNF released by CTL82/82 (pg/ml)
Figure 2. Stimulation of CTL 82•82 by COS-7 Cells Transfected with cDNA-AD5 and with an HLA-Cw*1601 cDNA, or with Each of These cDNAs Alone The cDNAs were inserted in expression vector pcD-SRm CTL 82•82 was added 1 day after the transfection and the TNF content of the supernatant was estimated 1 day later by testing its toxicity on cells of WEHI-164 clone 13. Positive and negative controls were made with MZ2-MEL.43 and MZ2-MEL.2.2.5 cells.
taining more than 40 pg/ml of TNF, whereas TNF concen- tration in all the microcultures transfected with the other pools was lower than 5 pg/ml. From one of these cDNA pools, which contained 400 independent bacteria, we sub- cloned 800 bacteria. Plasmid DNA was extracted from each of them and transfected into COS-7 cells together with the HLA-Cw*1601 construct. Twelve clones con- ferred recognition by CTL 82182. The result obtained with one of them, namely cDNA-AD5, is represented in Fig- ure 2.
To confirm the result obtained with cDNA-AD5 in tran- sient transfections, stable transfectants were also pre- pared. P1.HTR, a highlytransfectable variant derived from mouse tumor cell line P815 (Van Pel et al., 1985), was transfected with both HLA-Cw*1601 and cDNA-AD5. Transfected cells were lysed by CTL 82•82, indicating that the antigen can also be processed in these mouse cells (Figure 3A).
Sequence of Gene BAGE Even though the sequence of cDNA-AD5 is 1004 bp long, the largest open reading frame comprises only 132 bp and encodes a protein of 43 aa (Figure 4). The sequence bears no significant similarity to any other sequence presently recorded in data banks, except for an Alu repeat sequence (nucleotides 385-484) located outside the coding part. We propose to name BAGE the gene that corresponds to cDNA-AD5.
We prepared a Southern blot with DNA extracted from blood lymphocytes of patient MZ2 and from the melanoma line MZ2-MEL.3.0. This blot was hybridized with a 121 bp probe including almost all the BAGE coding sequence. We observed four bands in lanes containing DNA digested with EcoRI and six bands after Hindlll digestion (Figure 5). Considering thesmall size of the probe and considering the absence of EcoRI and Hindlll restriction sites in the coding sequence, these results indicate that BAGE be- longs to a family of several related genes.
Expression of Gene BAGE A Northern blot prepared with poly(A) + RNA of subline MZ2-MEL.43 was hybridized with a BAGE probe covering

Gene BAGE Encodes a Human Tumor Antigen 169
8O
_~ 6o ._o :~ 40.
0..9 " 20
A
TARGET: PI.HTR transfected with
HLA-Cw*1601 HLA-Cw*1601 + cDNA-AD5
.1 .3 i 3 1'0 .1 .3 1 3 10 .'1 .3 1" 3 1"0 Effector to target ratios
B
P1.HTR.Cw*160" + peptide
AARAVFLAL
/ .'1 .3 i ~ lo
Figure 3. Lysis of Mouse Cells Expressing HLA-Cw*1601 and Trans- fected with BAGE or Incubated with the BAGE-Encoded Peptide (A) Lysis by CTL clone 82/82 of PI.HTR mouse cells cotransfected with expression vectors carrying H LA-Cw*1601 and cDNA-AD5. Un- transfected P 1.HTR and P 1. HTR transfected with H LA-Cw * 1601 alone were also tested. (B) Lysis by CTL clone 82/82 of P 1. HTR transfected with H LA-Cw * 1601 and incubated with 1 I~M of the BAGE-encoded nonapeptide. Lysis of chromium-labeled cells was tested after 4 hr.
Kb
E c o R I H i n d l I I I II I
nucleotides 100-385. Two bands of approximately 1 and 2.4 kb were observed (Figure 6). The expression of BAGE was also studied by reverse transcription and polymerase chain reaction (PCR) amplification, with primers located on both sides of the coding part of the gene (see Figure 4). These primers are located in two different exons: the PCR amplified a fragment of 1.7 kb on DNA of MZ2- MEL.43 and a fragment of 286 bp on cDNA (Figure 7).
No expression of gene BAGE was found in normal adult t issues except in testis (Figure 7; Table 1). The gene was also silent in placenta and umbilical cord and in several tissue samples f ro~ fetuses older than 20 weeks. We found no expression of BAGE in the twelve EBV-trans- formed lymphoblastoid cell l ines and in blood lymphocytes
st imulated with phytohemagglutinin. Of tumors of various histological origins, 600 samples
were analyzed for expression of BAGE (Table 2; Figure 7). The gene is expressed mainly in melanomas (22%),
1 0 1 ~
8 •
6 • ~ . . . . . . . . . .
4 •
s • O g
2 •
CGCCAATTTA GGGTCTCCGG TATCTCCCGC TGAGCTGCTC TGTTCCCGGC TTAGAGGACC 60
AGGAGAAGGG GGAGCTGGAG GCTGGAGCCT GTAACACCGT GGCTCGTCTC ACTCTGGATG 120
GTGGTGGCAA CAGAGATGGC AGCGCAGCTG GAGTGTTAGG AGGGCGGCCT GAGCGGTAGG 180
M IA A R A V F L A L i S A Q 13 AGTGGGGCTG GAGCAGTAAG ATGGCGGCCA GAGCGGTTTT TCTGGCATTU TCTGCCCAGC 240
L L Q A R L M K E E S P V V S W R L E P 33
TGCTCCAAGC CAGGCTGATG AAGGAGGAGT CCCCTGTGGT GAGCTGGAGG TTGGAGCCTG 300
E D G T A L C F I F 43 AAGACGGCAC AGCTCTGTGC TTCATCTTCT GAGGTTGTGG CAGCCACGGT GATGGAGACG 360
GCAGCT~AAC AGGAGCAATA GGAGGAGATG GAGTTTCACT GTGTCAGCCA GGATGGTCTC 420
GATCTCCTGA CCTCGTGATC CGCCCGCCTT GGCCTTCCAA AGTGCCGAGA TTACAGCGAT 480
GTGCATTTTG TAAGCACTTT GGAGCCACTA TC~AATGCTG TGAAGAGAAA TGTACCCAGA 540
TGTATCATTA TCCTTGTGCT GCAGGAGCCG GCTCCTTTCA GGATTTCAGT CACATCTTCC 600
TGCTTTGTCC AGAACACATT GACCAAGCTC CTGAAAGATG TAAGTTTACT ACGCATAGAC 660
TTTT~AACTT CAACCAATGT ATTTACTGAA AATAACAAAT GTTGTA~TT CCCTGAGTGT 720
TATTCTACTT GTATTAAAAG GTAATAATAC ATAATCATTA AAATCTGAGG GATCATTGCC 780
AGAGATTGTT GGGGAGGGAA ATGTTATCAA CGGTTTCATT GAAATTAAAT CCA~GTT 840
ATTTCCTCAG AAAAATCAAA TAAAGTTTGC ATGTTTTTTA TTCTTAAAAC ATTTTAAAAA 900
CCACTGTAGA ATGATGTAAA TAGGGACTGT GCAGTATTTC TGACATATAC TATAAAATTA 960
TTAAAAAGTC AATCAGTATT CA~CATCTTT TACACTAA~A AGCC 1004
Figure 4. Sequence of BAGE cDNA-AD5 and of the Putative Protein of 43 aa Encoded by the Largest Open Reading Frame The sequence corresponding to the peptide recognized in association with HLA-Cw*1601 by MZ2-CTL 82/82 is indicated in a box. The se- quence of primers VDB85 (sense) and VDB86 (antisense) used for PCR amplification are underlined with arrows. The sequence of cDNA-A5 is available from EMBL/GenBank/DDBJ under accession number U19180.
Figure 5. Southern Blot of DNA Extracted from Melanoma Cell Line MZ2-MEL3.0, Blood Lymphocytes from Patient MZ2, and Mouse Cell Line P1 .HTR The DNA was digested with EcoRI or Hindlll. The membrane was hybridized with a probe including almost all the BAGE coding sequence (nucleotides 211-331) and it was washed under stringent conditions.
bladder carcinomas (15%), mammary carcinomas (10%), and head and neck squamous cell carcinomas (8%). A smaller proportion of posit ive samples was found in sarco- mas (6%) and in non-small cell lung carcinomas (6%). No expression of BAGE was found in renal, cotorectal, and prostatic carcinomas, leukemias, or lymphomas. With very few exceptions, tumor samples that expressed BAGE also expressed one of the MAGE genes (data not shown).
BAGE was more frequently expressed in metastatic le- sions of melanomas (26%) than in primary lesions (8%).

Immunity 170
P r o b e s
oo
. j .9_ ILl - ~
N =E E
B A G E 5.1 kb
,4 2 . 0 k b
I -actin
Figure 6. Northern Blot Analysis of the Expression of Gene BAGE in MZ2-MEL.43 Cells Lanes contained 5 l~g of poly(A) + RNA. The membrane was hybridized with a probe including the BAGE coding sequence (nucleotides 100- 385) and washed under stringent conditions. Control hybridization was performed on the same membrane with a [5-actin probe.
in transitional cell carcinomas of the urinary bladder, 30% of invasive tumors (stages T2-T4 (Mostofi et al., 1973)) expressed BAGE, while no expression was observed in superficial tumors (stages Ta-T1). BAGE was expressed in a higher proportion of tumor cell lines than of tumor samples: 32 of 60 melanoma (53%) and 3 of 15 colorectal carcinoma (20%) cell lines were positive. This has also been observed with MAGE genes. It may be owing to the fact that tumor cell lines are more readily derived from metastatic tumors.
Identification of the Antigenic Peptide To identify the region of the BAGE sequence that codes for the antigenic peptide presented by HLA-Cw* 1601, we produced a large number of truncated BAGEcDNA clones. By digesting BAGE cDNA-AD5 with exonuclease III for various incubation times, we generated progressive de- letions starting from the 3' end. The truncated cDNAs were transfected into COS-7 cells, together with the HLA-
Table 1. Expression of Gene BAGE by Normal Adult and Fetal Tissues
Adult tissues Expression*
Adrenal gland Bone marrow Brain Breast Cerebellum Colon Heart Kidney Liver Lung Melanocytes Muscle Ovary Prostate Skin Sperm Splenocytes Stomach Testis + Thymocytes Urinary bladder Uterus Placenta Umbilical cord Benign naevus
Fetal tissues1" Expression
Fibroblasts Brain Liver Spleen Thymus Testis
* Expression of gene BAGE was tested by RT-PCR amplification of total RNA, with the primers shown on Figure 4. Absence of PCR prod- uct is indicated by (minus) and presence by (plus). 1" Fetal tissues derived from fetuses older than 20 weeks.
Cw'1601 cDNA, to test for their ability to code for the antigen. Sequence comparison of the longest negative (nucleotides 1-206) and the shortest positive cDNA (nucle- otides 1-267) indicated that the antigenic peptide was en- coded by the first 67 nt of the open reading frame.
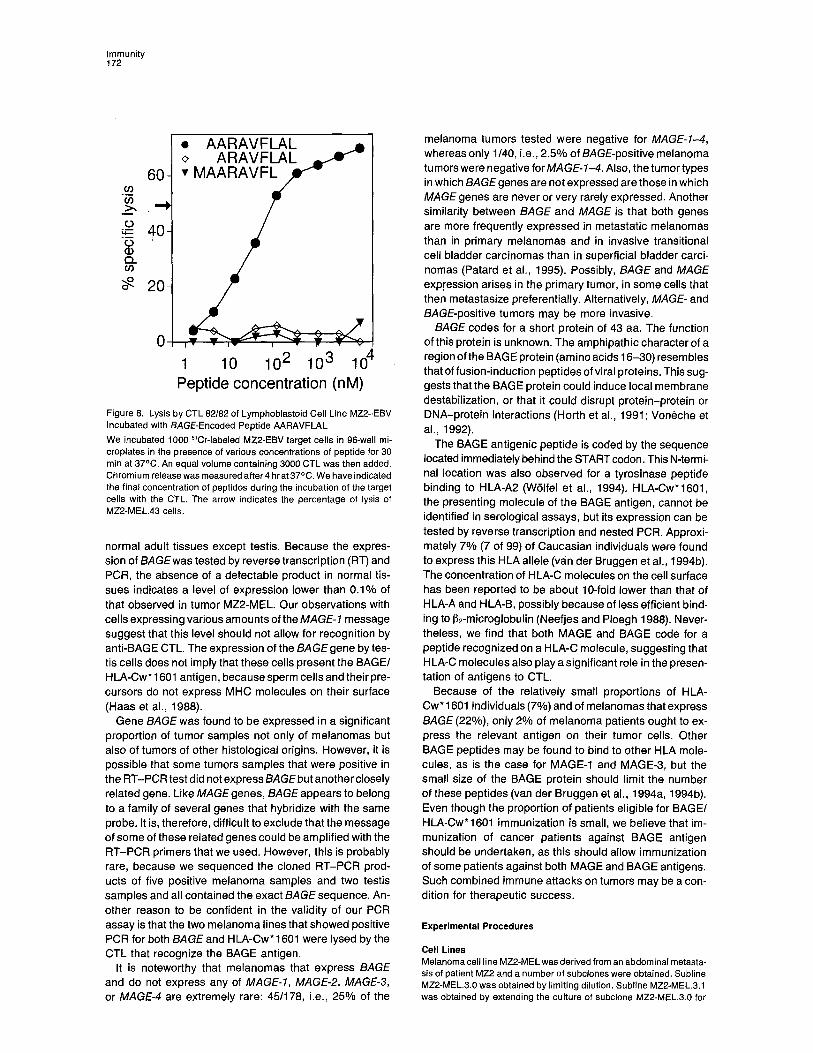
Several synthetic peptides were prepared on this basis. Sensitization of a lymphoblastoid cell line from patient MZ2 to lysis by CTL 82/82 was observed with nonapeptide AAR- AVFLAL (amino acids 2-10; see Figure 4). Half-maximal lysis was obtained at a peptide concentration of 30 nM (Figure 8). Nonapeptides that did not include the N-terminal Ala or the C-terminal Leu were not able to sensitize target cells to lysis. When P1.HTR cells transfected with HLA- Cw* 1601 were incubated with nonapeptide AARAVFLAL, they were also lysed by CTL 82/82 (see Figure 3B).
From two MLTC set up with different blood samples of patient MZ2, we derived six CTL c/ones that recognized the BAGE/HLA-Cw* 1601 antigen on MZ2-MEL cells. They also produced TNF in the presence of COS-7 cells that had been either cotransfected with HLA-Cw*1601 and BAGEcDNA-AD5, or that had been transfected with HLA- Cw* 1601 and incubated with nonapeptide AARAVFLAL

Gene BAGE Encodes a Human Tumor Antigen 171
cDNA DNA
O t h e r Norma l Melanomas tumors tissues
(D
~=, "¢ ¢.O '~" ¢~ E .J ,,, = , m
-
-= =E ~ E E
E. E
Figure 7. PCR Amplification of cDNAs from Melanoma Lines, Tumor and Normal Tissue Samples, and of Genomic DNA from Subline MZ2-MEL.43 The cDNA was obtained with an oligo(dT) primer. PCR amplification was performed on 100 ng of cDNA with BAGE-specific primers VDB85 (sense) and VDB8S (antisense). Hu- man ~-actin amplification was also performed on the cDNA samples to check the quality of RNA preparations. PCR products were visual- ized on a 1.5% agarose gel stained with ethid- ium bromide.
BAGE
1.7 kb
286 bp
I~-actin
Table 2. Expression of Gene BAGE by Tumor Samples
Number of BAGE Histological type positive tumors*
Melanomas 40/178 primary lesions 3/38 metastatic lesions 37/140
Bladder carcinomas 9/62 superficial tumor~ 0/32 infiltrating tumors 9/30
Mammary carcinomas 8•79 Head and neck squamous cell carcinomas 4/53 Lung carcinomas NSCLCt 4/64 Sarcomas 1/18 Renal carcinomas 0/50 Colorectal carcinomas 0/42 Prostatic carcinomas 0/22 Leukemias and lymphomas 0/22
* Expression of gene BAGE was tested by RT-PCR amplification of total RNA with the primers shown on Figure 4. t NSCLC, non-small cell lung carcinomas.
(data not shown). CTL clone 82/1 expressed V~2 and V~3 and V[~13. CTL clone 25/244 expressed Va8 and VI38, and CTL clone 82/82 expressed V~3 and Ve4 and V~13. It appears, therefore, that at least three different CTL precur- sors from the same patient can recognize the same BAGE antigenic peptide. This was previously observed for the MAGE antigens presented by HLA-Cw* 1601 (van der Brug- gen et al., 1994b) and HLA-A1 (P. Romero and P. C., un- published data). Several reports have described the oc-
curence of mature ~ T cells that have undergone two productive Ve rearrangements and have, therefore, the potential to express two V~ genes and two T cell receptors (reviewed by Padovan et al., 1993).
Discussion
So far, two main classes of antigens recognized by autolo- gous CTL have been found on human melanoma. The antigens of the first class are encoded by genes that are expressed very specifically in tumors. An antigen encoded by gene MAGE-1 was the first example (van der Bruggen et al., 1991), followed by other antigens encoded by genes MAGE-1 and MAGE-3 (Gaugler et al., 1994; van der Brug- gen et al., 1994a, 1994b). A tumor rejection antigen ob- served on mouse mastocytoma P815 also resulted from the activation of a gene that is silent in all normal adult tissues with the exception of testis (Van den Eynde et al., 1991). The second class of antigens represents differenti- ation antigens encoded by genes that are expressed only in melanocytes and melanomas. Antigens encoded by ty- rosinase were examples of this class (Brichard et al., 1993; Robbins et al., 1994; W61fel et al., 1994), which also com- prises antigens encoded by Melan-A/MART-1 (Coulie et al., 1994; Kawakami et al., 1994) and gp100/pmell 7 (Bak- ker et al., 1994; Cox et al., 1994).
The antigen encoded by gene BAGE is a new member of the first class. We decided to use the suffix "AGE" in the names of the antigen-coding genes that are silent in
Immunity 172
6 0 - oo
o ~= 40 - o (b o.. t J)
o "9- 20-
0
• A A R A V F L A L <> A R A V F L A L • M A A R A V F L J
1 10 102 10 3 104 Peptide concentration (nM)
Figure 8. Lysis by CTL 82•82 of Lymphoblastoid Cell Line MZ2-EBV Incubated with BAGE-Encoded Peptide AARAVFLAL We incubated 1000 51Cr-labeled MZ2-EBV target cells in 96-well mi- croplates in the presence of various concentrations of peptide for 30 min at 37°C. An equal volume containing 3000 CTL was then added. Chromium release was measured after 4 hr at 37°C. We have indicated the-final concentration of peptides during the incubation of the target cells with the CTL. The arrow indicates the percentage of lysis of MZ2-MEL.43 cells.
normal adult tissues except testis. Because the expres- sion of BAGE was tested by reverse transcription (RT) and PCR, the absence of a detectable product in normal tis- sues indicates a level of expression lower than 0.1% of that observed in tumor MZ2-MEL. Our observations with cells expressing various amounts of the MAGE- 1 message suggest that this level should not allow for recognition by anti-BAGE CTL. The expression of the BAGE gene by tes- tis cells does not imply that these cells present the BAGEl HLA-Cw* 1601 antigen, because sperm cells and their pre- cursors do not express MHC molecules on their surface (Haas et al., 1988).
Gene BAGE was found to be expressed in a significant proportion of tumor samples not only of melanomas but also of tumors of other histological origins. However, it is possible that some tumors samples that were positive in the RT-PCR test did not express BAGEbut another closely related gene. Like MAGE genes, BAGE appears to belong to a family of several genes that hybridize with the same probe. It is, therefore, difficult to exclude that the message of some of these related genes could be amplified with the RT-PCR primers that we used. However, this is probably rare, because we sequenced the cloned RT-PCR prod- ucts of five positive melanoma samples and two testis samples and all contained the exact BAGE sequence. An- other reason to be confident in the validity of our PCR assay is that the two melanoma lines that showed positive PCR for both BAGE and H LA-Cw* 1601 were lysed by the CTL that recognize the BAGE antigen.
It is noteworthy that melanomas that express BAGE and do not express any of MAGE-1, MAGE-2, MAGE-3, or MAGE-4 are extremely rare: 45/178, i.e., 25% of the
melanoma tumors tested were negative for MAGE-1-4, whereas only 1/40, i.e., 2.5% of BAGE-positive melanoma tumors were negative for MAGE-1-4. Also, the tumor types in which BAGE genes are not expressed are those in which MAGE genes are never or very rarely expressed. Another similarity between BAGE and MAGE is that both genes are more frequently expressed in metastatic melanomas than in primary melanomas and in invasive transitional cell bladder carcinomas than in superficial bladder carci- nomas (Patard et al., 1995). Possibly, BAGE and MAGE expression arises in the primary tumor, in some cells that then metastasize preferentially. Alternatively, MAGE- and BAGE-positive tumors may be more invasive.
BAGE codes for a short protein of 43 aa. The function of this protein is unknown. The amphipathic character of a region of the BAGE protein (amino acids 16-30) resembles that of fusion-induction peptides of viral proteins. This sug- gests that the BAGE protein could induce local membrane destabilization, or that it could disrupt protein-protein or DNA-protein interactions (Horth et al., 1991; Von~che et al., 1992).
The BAGE antigenic peptide is coded by the sequence located immediately behind the START codon. This N-termi- nal location was also observed for a tyrosinase peptide binding to HLA-A2 (WSIfel et al., 1994), HLA-Cw*1601, the presenting molecule of the BAGE antigen, cannot be identified in serological assays, but its expression can be tested by reverse transcription and nested PCR. Approxi- mately 7% (7 of 99) of Caucasian individuals were found to express this HLA allele (van der Bruggen et al., 1994b). The concentration of HLA-C molecules on the cell surface has been reported to be about 10-fold lower than that of HLA-A and HLA-B, possibly because of less efficient bind- ing to 132-microglobulin (Neefjes and PIoegh 1988). Never- theless, we find that both MAGE and BAGE code for a peptide recognized on a HLA-C molecule, suggesting that HLA-C molecules also play a significant role in the presen- tation of antigens to CTL.
Because of the relatively small proportions of HLA- Cw* 1601 individuals (7%) and of melanomas that express BAGE (22%), only 2% of melanoma patients ought to ex- press the relevant antigen on their tumor cells. Other BAGE peptides may be found to bind to other HLA mole- cules, as is the case for MAGE-1 and MAGE-3, but the small size of the BAGE protein should limit the number of these peptides (van der Bruggen et al., 1994a, 1994b). Even though the proportion of patients eligible for BAGEl HLA-Cw*1601 immunization is small, we believe that im- munization of cancer patients against BAGE antigen should be undertaken, as this should allow immunization of some patients against both MAGE and BAGE antigens. Such combined immune attacks on tumors may be a con- dition for therapeutic success.
Experimental Procedures
Cell Lines Melanoma cell line MZ2-MEL was derived from an abdominal metasta- sis of patient MZ2 and a number of subclones were obtained, Subline MZ2-ME/.3,0 was obtained by limiting dilution, Subline MZ2-MEL.3,1 was obtained by extending the culture of subclone MZ2-MEL.3,0 for

Gene BAGE Encodes a Human Tumor Antigen 173
more than 150 generations. Subline MZ2-MEL.43 was derived by lim- iting dilution from MZ2-MEL.3.0 cells that had survived to a mutagen treatment (H~rin et al., 1987; Van den Eynde et al., 1989). Clonal subline MZ2-MEL.2.2, which does not express antigen MZ2-E, was selected from subclone MZ2-MEL.3.1 with an autologous anti-MZ2-E CTL clone (Van den Eynde et al., 1989). Subline MZ2-MEL.2.2.5 was selected from subline MZ2-MEL.2.2 with an anti-MZ2-F CTL clone. MZ2-MEL.B.TC.4 was obtained by transfecting the HLA-Cw*1601 gene into subline MZ2-MEL.2.2.5 (van der Bruggen et al., 1994b). Melanoma cell lines were grown as previously described (Van den Eynde et al., 1989; Traversari et al., 1992b). Autologous CTL clone 82182 was derived from peripheral blood lymphocytes (PBL) of patient MZ2 and grown in conditions similar to those previously described (Traversari et al., 1992b). Lymphoblastoid cell line MZ2-EBV was de- rived from patient MZ2 by standard techniques. Mouse cell line PI.HTR was derived from mastocytoma P815 (Van Pel et al., 1985). PI.HTR cells were cotransfected by the phosphate calcium precipitate method with plasmid pSVtkneol~ conferring resistance to geneticin (Nicolas and Berg, 1983) and HLA-Cw*1601 alone, or HLA-Cw*1601 and BAGE cDNA-AD5, cDNAs were inserted in expression vector pcD- SR~ (received from K. Moore, DNAx, Palo Alto, California; Takebe et al., 1988). Cional sublines were isolated from a geneticin-resistant transfected population.
Construction of the cDNA Library Poly(A) ÷ RNA was extracted from MZ2-MEL.43 cells using mRNA ex- traction kit Fastrack (Invitrogen Corporation, Oxon, United Kingdom). mRNA was converted to cDNA using random primers, Iigated to adap- tors as described in the SuperScript plasmid system kit (GIBCO BRL, Gaithersburg, Maryland), and inserted into the EcoRI site of expression vector pcD-SR*z. Recombinant plasmids were electroporated into Escherichia coil JM101 and selected with ampicillin (50 i~g/ml). The library contained 66,000 inserts and was divided into 87 pools of 400 bacteria and 297 pools of 200 bacteria. Each of these pools comprised approximately 280 or 140 different cDNAs, respectively, as about 70% of the plasmids carried an insert. Each pool of bacteria was amplified to saturation and plasmid DNA was extracted by the alkaline lysis method.
Transfection of COS-7 Cells Transfection experiments were performed by the DEAE-dextran-chlo- roquine method (Seed and Aruffo 1987; Brichard et al., 1993; Coulie et ai., 1994). In brief, 1.5 x 104 COS-7 cells were transfected with 100 ng of plasmid pcD-SR(~ containing cDNA of HLA-Cw*1601, and 100 ng of a poolOf the cDNA library or 100 ng of a cDNA clone. The HLA-Cw*1601 cDNA was isolated from a cDNA library prepared with RNA extracted from subline MZ2-MEL.43 (van der Bruggen et al., 1994b). Transfected COS-7 cells were tested in a CTL stimulation assay after 24 or 48 hr.
CTL Stimulation Assay Transfectants were tested for their ability to stimulate the production of TNF by CTL (Traversari et al., 1992b). In brief, in microwells containing target cells, 1500 CTL were added in 100 i~1 of Iscove medium (GIBCO BRL) containing 10% human serum and 20 U/ml r-hulL-2. After 24 hr, the supernatant was collected and its TNF content was determined by testing its cytotoxic effect on cells of WEHI-164 clone 13 (Espevik and Nissen-Meyer, 1986) in a M'FT colorimetric assay (Hansen et al., 1989; Traversad et al., 1992b).
DNA Sequencing and Homology Search DNA sequencing analysis was performed by specific priming with syn- thetic oligonucleotides. The sequencing reactions were performed by the dideoxy-chain termination method (T7 Sequencing Kit, Pharmacia, Uppsala, Sweden, ATaq Cycle-Sequencing Kit, United States Bio- chemical, Cleveland, Ohio). The computer search for the sequence homology was done with programs FASTA@EMBL-Heidelberg and [email protected]. The sequence of BAGE cDNA-AD5 is avail- able from EMBL/GenBank/DDBJ under accession number U19180.
Southern Blot Analysis DNA from melanoma cell line MZ2-MEL.3.0, PBL of patient MZ2, and mouse cell line P1.HTR were digested with EcoRI or Hindlll. DNA
capillary transfer was done by alkaline blotting on a Zeta-Probe mem- brane (Bio-Rad). Following transfer, the membrane was rinsed in 2 x SSC, baked for 1 hr at 80°C and pretreated for 30 min at 60°C in 6 x SSC, 10x Denhardt's solution. The membrane was then hybridized for 18 hr at 65°C in 3.5x SSC, l x Denhardt's solution, 25 mM NaH2PO4 (pH 7.0), 0.5% SDS, 2 mM EDTA, 100 p.g/ml of herring sperm DNA, and 2 x 106 cpm/ml of a 121 bp 32P-labeled probe (BAGE cDNA-AD5, nucleotides 211-331) produced by PCR. The membrane was then washed at 65°C two times for 15 min in 2x SSC, 0.5% SDS, then for 15 min in 0.2 x SSC, 0.1% SDS, and autoradiographed for 10 days.
Northern Blot Analysis Poly(A) + RNA from MZ2-MEL.43 was prepared using mRNA extraction kit Fastrack (Invitrogen Corporation, Oxon, United Kingdom). Total RNA from mouse kidney tissue was extracted by the guanidine-iso- thiocyanate procedure as described (Davis et al., 1986). Poly(A)* RNA was purified from total RNA on an oligo(dT) column. For the Northern blot analysis, 5 p.g of poly(A) ÷ RNA from subline MZ2-MEL.43 and 5 p.g of poly(A) + RNA from mouse kidney cells were fractionated on 1% agarose gel containing 0.66 M formaldehyde and transfered on a mem- brane (Hybond C-extra, Amersham) in 10 x SSC. The membrane was prehybridized for 15 rain at 60°C in 10% dextran sulfate, 1% SDS, and 1 M NaCI and hybridized overnight at 60°C in the same solution with 2 x 106 cpm/ml of a 286 bp 3~P-labeled probe (BAGE cDNA-AD5, nucleotides 100-385) produced by PCR and 100 i~g/ml of herring sperm DNA. The membrane was washed at room temperature in 0.2 x SSC for 10 min and then washed two times for 20 rain at 60°C in 0.2 x SSC supplemented with 0.1% SDS, and autoradiographed for 15 hr. Control hybridization was performed on the same membrane with a mouse I~-actin probe.
PCR Assay for BAGE Expression Total RNA was extracted by the guanidine-isothiocyanate procedure as described (Davis et al., 1986). Reverse transcription was performed on 2 p.g of total RNA in a reaction volume of 20 p.I with 4 p,I of 5x reverse transcriptase buffer (GIBCO BRL), 2 pl of a 20 mM solution of oligo(dT)~s primer, 20 U of RNasin (Promega Biotec, Madison, Wis- consin), 2 I~1 of 0.1 M dithiotreitol, and 200 U of MoMLV reverse tran- scriptase (GIBCO BRL). The reaction was incubated at 42°C for 60 rain. Of the cDNA product, one-twentieth was then supplemented with 5 I~1 of 10 x thermostable DNA polymerase buffer (Finnzymes OY, Espoo, Finland), 1 I11 each of 10 mM solution of dNTP, 1 Id each of 25 mM solution of primers, 1 U of DynaZyme (Finnzymes OY, Espoo, Finland), and water to a final volume of 50 p,l. The PCR primers were 5'-TGGCTCGTCTCACTCTGG-3' (VDB85, sense, nucleotides 100- 117) and 5'-CCTCCTATTGCTCCTGTTG-3' (VDB86, antisense, nucle- otides 367-385). The PCR was performed for 30 cycles (1 rain at 94°C, 2 min at 62°C, and 2 min at 73°C). Of the PCR product, 10 pl was size fractionated on a 1,5% agarose gel. The quality of RNA prepara- tions was tested by PCR amplification of human ~-actin cDNA with primers 5'-GGCATCGTGATGGACTCCG-3' (exon 3, sense) and 5'- GTCGGAAGGTGGACAGCGA-3' (exon 6, antisense) for 21 cycles of 1 min at 94°C, 2 rain at 68°C, and 2 min at 72°C by AmpliTaq DNA polymerase (Perkin-Elmer Cetus, Norwalk, Connecticut).
Production of Progressive Deletions in BAGE cDNA-AD5 BAGE cDNA-AD5 was inserted in the EcoRI site of pcDNAI/Amp (In- vitrogen Corporation, San Diego, California). The plasmid was opened with Notl and Sphl before being digested with exonuclease III. This treatment was performed with the Erase-a-base System (Promega, Madison, Wisconsin). After ligation, the plasmids were electroporated in E. coil DH5~F'IQ and selected with ampicillin (50 i~g/ml). Clones were isolated, plasmid DNA was extracted from each clone and transfected into COS-7 cells together with an HLA-Cw*1601 cDNA.
Antigenic Peptides and CTL Assay Peptides were synthesized on solid phase using Fmoc for transient NH2-terminal protection as described by Atherton et al. (1981) and characterized by mass spectrometry. All peptides were >90% pure as indicated by analytical high pressure liquid chromatography. Lyoph- ilized peptides were dissolved at 20 mg/ml in DMSO, diluted at 2 rag/ ml in 10 mM acetic acid, and stored at -80°C. Peptides were tested in

Immunity 174
a CTL stimulation assay with COS-7 cells transfected by H LA-Cw * 1601 and incubated with the peptides. They were also tested by chromium release assay as previously described (Boon et al., 1980). In this pep- tide sensitization assay, target cells were ~lCr-labeled for 1 hr at 37°C and washed extensively. Then 1000 target cells were incubated in 96-well microplates in the presence of various concentrations of pep- tide for 30 min at 37°C before CTL 82•82 cells were added. Chromium release was measured after 4 hr at 37°C.
TCR Va and VI} PCR Amplification Total RNA from the different CTL clones was prepared by using RNA- zol* B (Cinna/Biotecx, Friendswood, Texas) (Chomczynski and Sacchi, 1987). Single-stranded cDNA synthesis was carried with oligo(dr) and Moloney murine leukemia virus-derived reverse transcriptase without RNAse H activity (MMLV RT RNAse H, Superscript, GIBCO BRL, Rockville, Maryland). PCR was carried out by amplification of TCR~ and TCRI~ cDNA with the oligonucleotide primers complementary to TCR variable (V(~1-W29, VI~l-W24) and constant (Co{, CI~) region se- quences described by Genev~e et al. (1992). Specificity of TCR V(1 and VI3 PCR amplification was assessed by Southern blotting and hybridization with ~2P-labeled C(x or Cl~ oligonucleotides internal to the ones used for amplification.
Acknowledgments
The secretarial assistance of S. Khaoulali is gratefully acknowledged. This work is partially supported by the Belgian program on Interuniver- sity Poles of Attraction initiated bythe Belgian state, Office of the Prime Minister, Office for Science, Technology, and Culture. The scientific responsibility is assumed by the authors. This research is partially subsidized by the Dr. Steiner prize (Switzerland), and by the Associa- tion centre le Cancer (Belgium). P. B. is Research Assistant of the Fends National de la Recherche Scientifique, Brussels, Belgium. C. W. is supported by a grant, TELEVIE, from the Fonds National de la Recherche Scientifique, Brussels, Belgium. M. L. S. is partially supported by a grant from the Italian Association for Cancer Research (Milan, Italy). R. B. is Research Director of the Fonds National de la Recherche Scientifique, Brussels, Belgium.
Received November 7, 1994; revised January 9, 1995.
References
Atherton, E., Logan, C. J., and Sheppard, R. C. (1981). Peptide synthe- sis. II. Procedures for solid phase synthesis using N(x-fluorenylmeth- oxycarbamylamino-acid on polymide supports: synthesis of substance P and of acyl carrier protein 65-74 decapeptide. J. Chem. Soc. Lond. Perkin Trans. 1,538-546.
Bakker, A. B. H., Schreurs, M. W. J., de Boer, A. J., Kawakami, Y., Rosenberg, S. A., Adema, G. J., and Figdor, C. G. (1994). Melanocyte lineage-specific antigen gpl00 is recognized by melanoma-derived tumor-infiltrating lymphocytes. J. Exp. Med. 179, 1005-1009.
Boon, T., Cerottini, J.-C., Van den Eynde, B., van der Bruggen, P., and Van Pel, A. (1994). Tumor antigens recognized by T lymphocytes. Annu. Rev. Immunol. 12, 337-365.
Boon, T., Van Snick, J., Van Pel, A., Uyttenhove, C., and Marchand, M. (1980). Immunogenic variants obtained by mutagenesis of mouse mastocytoma P815. I1. T lymphocyte-mediated cytolysis. J. Exp. Med. 152, 1184-1193.
Brichard, V., Van Pel, A., WtHfel, T., W61fel, C., De Plaen, E., Leth6, B., Coulie, P., and Boon, T. (1993). The tyrosinase gene codes for an antigen recognized by autologous cytolytic T lymphocytes on HLA-A2 melanomas. J. Exp. Med. 178, 489-495.
Chomczynski, P., and Sacchi, N. (1987). Single-step method of RNA isolation by acid guanidium thiocyanate-phenol-chloroform extrac- tion. Annal Biochem 162, 156-159.
Coulie, P. G., Brichard, V., Van Pel, A., WSIfel, T., Schneider, J., Traversari, C., De Plaen, E., Lurquin, C., Szikora, J.-P., Renauld, J.-C., and Boon, T. (1994). A new gene coding for a differentiation antigen recognized by autologous cytolytic T lymphocytes on HLA-A2 melano- mas. J. Exp. Med. 180, 35-42.
Cox, A. L., Skipper, J., Chen, Y., Henderson, R. A., Darrow, T. L., Shabanowitz, J., Engelhard, V. H., Hunt, D. F., and Slingluff, C. L., Jr. (1994). Identification of a peptide recognized by five melanoma- specific human cytotoxic T cell lines. Science 264, 716-719.
Davis, L. G., Dibner, M. D., and Battey, J. F. (1986). Basic Methods in Molecular Biology (New York: Elsevier), pp. 130-135.
De Plaen, E., Arden, K., Traversari, C., Gaforio, J. J., Szikora, J.-P., De Smet, C., Brasseur, F., van der Bruggen, P., Lethe, B., Lurquin, C., Brasseur, R., Chomez, P., De Backer, O., Cavenee, W., and Boon, T. (1994). Structure, chromosomal localization and expression of twelve genes of the MAGE family. Immunogenetics 40, 360-369.
De Smet, C., Lurquin, C., van der Bruggen, P., De Plaen, E., Brasseur, F., and Boon, T. (!994). Sequence and expression pattern of the hu- man MAGE2 gene. Immunogenetics 39, 121-129.
Espevik, T., and Nissen-Meyer, J. (1986). A highly sensitive cell line, WEHI 164 clone 13, for measuring cytotoxic factor/tumor necrosis factor frem human monocytes. J. Immunol. Meth. 95, 99-105.
Gaugler, B., Van den Eynde, B., van der Bruggen, P., Romero, P., Gaforio, J. J., De Plaen, E., Lethe, B., Brasseur, F., and Boon, T. (1994). Human gene MAGE-3 codes for an antigen recognized on a melanoma by autologous cytolytic T lymphocytes. J. Exp. Med. 179, 921-930.
Genev~e, C., Diu, A., Nierat, J., Caignard, A., Dietrich, P. Y., Ferradini, L., Roman-Roman, S., Triebel, F., and Hercend, T. (1992). An experi- mentally validated panel of subfamily-specific oligonucleotide primers (V~l-w29NI31-w24) for the study o f human T cell receptor variable V gene segment usage by polymerase chain reaction. Eur. J. Immunol. 22, 1261-1269.
Haas, G. G., Jr., D'Cruz, O. J., and De Bault, L. E. (1988). Distribution of human leukocyte antigen-ABC and -D/DR antigens in the unfixed human testis. Am. J. Reprod. Immunol. Microbiol. 18, 47-51.
Hansen, M. B., Nielsen, S. E., and Berg, K. (1989). Re-examination and further development of a precise and rapid dye method for measur- ing cell growth/cell kill. J. Immunol. Meth. 119, 203-210.
Herin, M., Lemoine, C., Weynants, P., Vessi6re, F., Van Pel, A., Knuth, A., Devos, R., and Boon, T. (1987). Production of stable cytolytic T-cell clones directed against autologous human melanoma. Int. J. Cancer 39, 390-396.
Horth, M., Lambrecht, B., Chuah Lay Khim, M., Bex, F., Thiriart, C., Ruysschaert, J.-M., Burny, A., and Brasseur, R. (1991). Theoretical and functional analysis of the SIV fusion peptide. EMBO J. 10, 2747- 2755.
Kawakami, Y., Eliyahu, S., Delgado, C. H., Robbins, P. F., Rivoltini, L., Topalian, S. L., Miki, T., and Rosenberg, S. A.'(1994). Cloning of the gene coding for a shared human melanoma antigen recognized by autologous T cells infiltrating into tumor. Proc. Natl. Acad. Sci. USA 91, 3515-3519.
Mostofi, F. K., Sobin, L. H., and Torlini, H. (1973). Histological typing of urinary bladder tumors. WHO International Histological Classifica- tion of Tumors (Geneva: WHO).
Neefjes, J. J., and Ploegh, H. L. (1988). Allele and locus-specific differ- ences in cell surface expression and the association of HLA class I heavy chain with 132-microglobulin: differential effects of inhibition of glycosylation on class I subunit association. Eur. J. Immunol. 18, 801- 810.
Nicolas, J. F., and Berg, P. (1983). Regulation of expression of genes transduced into embryonal carcinoma cells. Cold Spring Harbor Con- ferences Cell Prolif. 10, 469-485.
Padovan, E., Casorati, G., Dellabona, P., Meyer, S., Brockhaus, M., and Lanzavecchia, A. (1993). Expression of two T cell receptor alpha chains: dual receptor T cells. Science 262, 422-424.
Patard, J.-J., Brasseur, F., GiI-Diez, S., Radvanyi, F., Marchand, M., Frangois, P., Abi Aad, A., Van Cangh, P., Abbou, C. C., Chopin, D., and Boon, T. (1995). Expression of MAGE genes in transitional-cell carcinomas of the urinary bladder. Int. J. Cancer, in press.
Robbins, P. F., EI-Gamil, M., Kawakami, Y., and Rosenberg, S. A. (1994). Recognition of tyrosinase by tumor-infiltrating lymphocytes

Gene BAGE Encodes a Human Tumor Antigen 175
from a patient responding to immunotherapy. Cancer Res. 54, 3124- 3126.
Seed, B., and Aruffo, A. (1987). Molecular cloning of the CD2 antigen, the T-cell erythrocyte receptor, by a rapid immunoselection procedure. Proc. Natl. Acad. Sci. USA 84, 3365-3369.
Takebe, Y., Seiki, M, Fujisawa, J.-I., Hoy, P., Yokota, K., Arai, K.-I., Yoshida, M., and Arai, N. (1988). SR(~ promoter: an efficient and versa- tile mammalian cDNA expression system composed of the simian virus 40 early promoter and the R-U5 segment of human T-cell leukemia virus type 1 long terminal repeat. MoI. Cell. Biol. 8, 466-472.
Traversari, C., van der Bruggen, P., Luescher, I. F., Lurquin, C., Cho- mez, P., Van Pel, A., De Plaen, E., Amar-Costesec, A., and Boon, T. (1992a). A nonapeptide encoded by human gene MAGE-1 is recog- nized on HLA-A1 by cytolytic T lymphocytes directed against tumor antigen MZ2-E. J. Exp. Med. 176, 1453-1457.
Traversari, C., van der Bruggen, P., Van den Eynde, B., Hainaut, P., Lemoine, C., Ohta, N., Old, L., and Boon, T. (1992b). Transfection and expression of a gene coding for a human melanoma antigen recog- nized by autologous cytolytic T lymphocytes. Immunogenetics 35, 145- 152.
Van den Eynde, B., Hainaut, P., Herin, M., Knuth, A., Lemoine, C., Weynants, P., van der Bruggen, P., Fauchet, R., and Boon, T. (1989). Presence on a human melanoma of multiple antigens recognized by autologous CTL. Int. J. Cancer 44,634-640.
Van den Eynde, B., Lethe, B., Van Pel, A., De Plaen, E., and Boon, T. (1991). The gene coding for a major tumor rejection antigen of tumor P815 is identical to the normal gene of syngeneic DBA/2 mice. J. Exp. Med. 173, 1373-1384.
van der Bruggen, P., Traversari, C., Chomez, P., Lurquin, C., De Plaen, E., Van den Eynde, B., Knuth, A., and Boon, T. (1991). A gene encoding an antigen recognized by cytolytic T lymphocytes on a hu- man melanoma. Science 254, 1643-1647.
van der Bruggen, P., Bastin, J., Gajewski, T., Coulie, P. G., Bo~l, P., De Smet, C., Traversari, C., Townsend, A., and Boon, T. (1994a). A peptide encoded by human gene MAGE-3 and presented by HLA-A2 induces cytolytic T lymphocytes that recognize tumor cells expressing MAGE-3. Eur. J. Immunol. 24, 3036-3043~
van der Bruggen, P., Szikora, J.-P., Bo~l, P., Wildmann, C., Somville, M., Sensi, M., and Boon, T. (1994b). Autologous cytolytic T lympho- cytes recognize a MAGE-1 nonapeptide on melanomas expressing HLA-Cw*1601. Eur. J. Immunol. 24, 2134-2140.
Van Pel, A., De Plaen, E., and Boon, T. (1985). Selection of a highly transfectable variant from" mouse mastocytoma P815. Som. Cell Genet. 11,467-475.
Voneche, V., Protetelle~ D., Kettmann, R., Willems, L., Limbach, K., Paoletti, E., Ruysschaert, J. M., Burny, A., and Brasseur, R. (1992). Fusogenic segments of bovine leukemia virus and simian immunodefi- ciency virus are interchangeable and mediate fusion by means of oblique insertion in the lipid bilayer of their target cells. Proc. Natl. Acad. Sci. USA 89, 3810-3814.
WSIfel, T., Van Pel, A., Brichard, V., Schneider, J., Seliger, B., Meyer zum B0schenfelde, K.-H., and Boon, T. (1994). Two tyrosinase nona- peptides recognized on HLA-A2 melanomas by autologous cytolytic T lymphocytes. Eur. J. Immunol. 24, 759-764.




















