
Targeted ex vivo reduction of CD64-positive monocytes in chronic myelomonocytic leukemia and acute...
12
Targeted ex vivo reduction of CD64-positive monocytes in chronic myelomonocytic leukemia and acute myelomonocytic leukemia using human granzyme B-based cytolytic fusion proteins Sonja Schiffer 1,2 , Reinhard Rosinke 2 , Edgar Jost 3 , Grit Hehmann-Titt 4 , Michael Huhn 1 , Georg Melmer 4 , Stefan Barth 1,2* and Theo Thepen 2* 1 Department of Experimental Medicine and Immunotherapy, Institute for Biomedical Engineering, University Hospital RWTH Aachen, Aachen, Germany 2 Department of Pharmaceutical Product Development, Fraunhofer IME, Aachen, Germany 3 Department of Hematology and Oncology (Internal Medicine IV), University Hospital RWTH Aachen, Germany 4 Pharmedartis GmbH, Aachen, Germany CMML (chronic myelomonocytic leukemia) belongs to the group of myeloid neoplasms known as myelodysplastic and myelo- proliferative diseases. In some patients with a history of CMML, the disease transforms to acute myelomonocytic leukemia (AMML). There are no specific treatment options for patients suffering from CMML except for supportive care and DNA methyl- transferase inhibitors in patients with advanced disease. New treatment strategies are urgently required, so we have investi- gated the use of immunotherapeutic directed cytolytic fusion proteins (CFPs), which are chimeric proteins comprising a selective domain and a toxic component (preferably of human origin to avoid immunogenicity). The human serine protease granzyme B is a prominent candidate for tumor immunotherapy because it is expressed in cytotoxic T lymphocytes and natural killer cells. Here, we report the use of CD64 as a novel target for specific CMML and AMML therapy, and correlate CD64 expression with typical surface markers representing these diseases. We demonstrate that CD64-specific human CFPs kill CMML and AMML cells ex vivo, and that the mutant granzyme B protein R201K is more cytotoxic than the wild-type enzyme in the presence of the granzyme B inhibitor PI9. Besides, the human CFP based on the granzyme B mutant was also able to kill AMML or CMML probes resistant to Pseudomonas exotoxin A. The clonal hematopoietic stem cell disorder CMML (chronic myelomonocytic leukemia), is a highly aggressive and resist- ant form of leukemia. It is characterized by persistent mono- cytosis in the peripheral blood, at least one dysplasia component in the bone marrow, and <20% promonocytes and blasts in peripheral blood and bone marrow. 1 CMML can be classified further according to blast num- bers, that is, CMML1 (<5% blasts in peripheral blood and <10% blasts in bone marrow) and CMML2 (<20% blasts in peripheral blood and 10–19% blasts in bone marrow). 2,3 The disease occurs mainly in elderly people with a median sur- vival of 15–20 months and leukemic transformation rates of 15–30%, both factors depending on the disease subtype. 4 Other diagnostic criteria include immunophenotyping based on the overlapping antigen expression patterns in CMML and acute myelomonocytic leukemia (AMML). The expres- sion of CD56 combined with reductions in the levels of mye- loid markers such as CD15 and CD13 is a unique signature of CMML monocytes. 5 Furthermore, CD14 is expressed at higher levels on bone marrow monocytes in CMML com- pared with reactive monocytosis and normal marrow sam- ples. 5 High levels of CD33 have also been reported. 6 Clonal cytogenetic abnormalities are found in 20–40% of CMML patients, but these do not appear to be specific. 7–9 The close relationship between CMML and other myelodysplastic (MDS)/MPNs makes correct diagnosis and treatment a sig- nificant challenge. Novel molecular markers described more recently include specific alleles of TET2 and CBL, but these Key words: serpin B9, immunotoxin, therapy-resistance, leukemia Additional Supporting Information may be found in the online version of this article. This is an open access article under the terms of the Creative Com- mons Attribution-Non-Commercial-NoDerivs Licence, which per- mits use and distribution in any medium, provided the original work is properly cited, the use is non-commercial and no modifica- tions or adaptations are made. *S.B. and T.T. contributed equally to this work Conflict of Interest disclosure: S.S., G.H.-T. and S.B. are co-inventors on a corresponding patent assigned to Pharmedartis GmbH. G. M. is the CEO of Pharmedartis GmbH. The authors otherwise disclose no conflicts of interest. DOI: 10.1002/ijc.28786 History: Received 1 Dec 2013; Accepted 30 Jan 2014; Online 13 Feb 2014 Correspondence to: Stefan Barth, Fraunhofer IME, Forckenbeckstr. 6, 52074 Aachen, Germany, Tel.: [49241608511060/1], Fax: 1[49- 241-6085-10000], E-mail: [email protected] Cancer Therapy Int. J. Cancer: 00, 00–00 (2013) V C 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC. International Journal of Cancer IJC
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Targeted ex vivo reduction of CD64-positive monocytes in chronic myelomonocytic leukemia and acute...

Targeted ex vivo reduction of CD64-positive monocytes inchronic myelomonocytic leukemia and acute myelomonocyticleukemia using human granzyme B-based cytolytic fusionproteins
Sonja Schiffer1,2, Reinhard Rosinke2, Edgar Jost3, Grit Hehmann-Titt4, Michael Huhn1, Georg Melmer4,
Stefan Barth1,2* and Theo Thepen2*
1 Department of Experimental Medicine and Immunotherapy, Institute for Biomedical Engineering, University Hospital RWTH Aachen, Aachen, Germany2 Department of Pharmaceutical Product Development, Fraunhofer IME, Aachen, Germany3 Department of Hematology and Oncology (Internal Medicine IV), University Hospital RWTH Aachen, Germany4 Pharmedartis GmbH, Aachen, Germany
CMML (chronic myelomonocytic leukemia) belongs to the group of myeloid neoplasms known as myelodysplastic and myelo-
proliferative diseases. In some patients with a history of CMML, the disease transforms to acute myelomonocytic leukemia
(AMML). There are no specific treatment options for patients suffering from CMML except for supportive care and DNA methyl-
transferase inhibitors in patients with advanced disease. New treatment strategies are urgently required, so we have investi-
gated the use of immunotherapeutic directed cytolytic fusion proteins (CFPs), which are chimeric proteins comprising a
selective domain and a toxic component (preferably of human origin to avoid immunogenicity). The human serine protease
granzyme B is a prominent candidate for tumor immunotherapy because it is expressed in cytotoxic T lymphocytes and natural
killer cells. Here, we report the use of CD64 as a novel target for specific CMML and AMML therapy, and correlate CD64
expression with typical surface markers representing these diseases. We demonstrate that CD64-specific human CFPs kill
CMML and AMML cells ex vivo, and that the mutant granzyme B protein R201K is more cytotoxic than the wild-type enzyme in
the presence of the granzyme B inhibitor PI9. Besides, the human CFP based on the granzyme B mutant was also able to kill
AMML or CMML probes resistant to Pseudomonas exotoxin A.
The clonal hematopoietic stem cell disorder CMML (chronicmyelomonocytic leukemia), is a highly aggressive and resist-ant form of leukemia. It is characterized by persistent mono-cytosis in the peripheral blood, at least one dysplasia
component in the bone marrow, and <20% promonocytesand blasts in peripheral blood and bone marrow.1
CMML can be classified further according to blast num-bers, that is, CMML1 (<5% blasts in peripheral blood and<10% blasts in bone marrow) and CMML2 (<20% blasts inperipheral blood and 10–19% blasts in bone marrow).2,3 Thedisease occurs mainly in elderly people with a median sur-vival of 15–20 months and leukemic transformation rates of15–30%, both factors depending on the disease subtype.4
Other diagnostic criteria include immunophenotyping basedon the overlapping antigen expression patterns in CMMLand acute myelomonocytic leukemia (AMML). The expres-sion of CD56 combined with reductions in the levels of mye-loid markers such as CD15 and CD13 is a unique signatureof CMML monocytes.5 Furthermore, CD14 is expressed athigher levels on bone marrow monocytes in CMML com-pared with reactive monocytosis and normal marrow sam-ples.5 High levels of CD33 have also been reported.6 Clonalcytogenetic abnormalities are found in 20–40% of CMMLpatients, but these do not appear to be specific.7–9 The closerelationship between CMML and other myelodysplastic(MDS)/MPNs makes correct diagnosis and treatment a sig-nificant challenge. Novel molecular markers described morerecently include specific alleles of TET2 and CBL, but these
Key words: serpin B9, immunotoxin, therapy-resistance, leukemia
Additional Supporting Information may be found in the online
version of this article.
This is an open access article under the terms of the Creative Com-
mons Attribution-Non-Commercial-NoDerivs Licence, which per-
mits use and distribution in any medium, provided the original
work is properly cited, the use is non-commercial and no modifica-
tions or adaptations are made.
*S.B. and T.T. contributed equally to this work
Conflict of Interest disclosure: S.S., G.H.-T. and S.B. are
co-inventors on a corresponding patent assigned to Pharmedartis
GmbH. G. M. is the CEO of Pharmedartis GmbH. The authors
otherwise disclose no conflicts of interest.
DOI: 10.1002/ijc.28786
History: Received 1 Dec 2013; Accepted 30 Jan 2014; Online 13 Feb
2014
Correspondence to: Stefan Barth, Fraunhofer IME, Forckenbeckstr.
6, 52074 Aachen, Germany, Tel.: [49241608511060/1], Fax: 1[49-
241-6085-10000], E-mail: [email protected]
Can
cerTherapy
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.
International Journal of Cancer
IJC

are not specific and there are no therapeutic strategies avail-able to target the corresponding proteins.10
Currently, the only curative therapeutic approach forCMML is allogeneic hematopoietic stem cell transplantation,but this treatment option is not suitable for elderly patients,which represent most of the affected population. Palliativetreatments for advanced CMML include transfusions andhydroxyurea to reduce the high cell count in peripheralblood,11 although DNA methyltransferase inhibitors may alsobe administered to CMML2 patients to delay transformationto AMML and reduce the frequency of transfusions.12 AMMLrepresents 15–20% of all AML cases. It occurs in all agegroups but is more common in older adults, and is initiallytreated with chemotherapy in some cases followed by alloge-neic hematopoietic stem cell transplantation. Myeloblasts typ-ically express CD13, CD33, CD34 and CD117, whereas themoncytic lineage including monoblasts and promonocytesexpress CD11b, CD11c, CD13, CD14, CD33 and CD64.2,13
Standard cancer therapies generally lack specificity result-ing in significant off-target effects on normal cells. Morespecific and targeted approaches include the use of immuno-toxins, which are chimeric proteins comprising a specifictumor-binding component and a toxic component that indu-ces apoptosis. The tumor-binding component is usually a full-length antibody or single chain fragment derivative, whichbinds to the target cells triggering internalization so that thetoxic component can enter the cell. Most immunotoxins havebeen designed to target hematological disorders because theyare accessible, and many different combinations of target andtoxin components have therefore been studied for the treat-ment of leukemias and lymphomas.14 The only antibodybased targeted therapy used in human acute myeloid leukemiais gemtuzumab ozogamycin. However, clinical studies withthis conjugate were not convincing and approval for the drughas been withdrawn.15
Initially, the toxins originated from plants or bacteria andwere therefore immunogenic,16 but human cytolytic fusionproteins (hCFPs) have been investigated as a more recentalternative to circumvent this undesirable effect whenrepeated administration is necessary.
Human granzyme B is one of the most promising toxincandidates for immunotherapy17 and many studies have dem-onstrated its potential for cancer therapy, even for the treat-ment of tumor cell lines that show resistance against manycytotoxic drugs.18–20 Granzyme B is a serine protease primarilyexpressed in cytotoxic T lymphocytes (CTLs) and natural
killer (NK) cells.21,22 It activates apoptosis via both thecaspase-dependent and caspase-independent pathways.23–26
However, one of the major drawbacks of granzyme B is thepresence of its specific inhibitor serpin B9 (PI9). This proteinis a member of the clade B serpin family and like granzyme Bit is expressed in CTLs and NK cells. Recent studies haveshown that it is also expressed in cancer cells allowing them tocircumvent the natural immune response.27–30 We haveaddressed this challenge by developing a PI9-independent var-iant of granzyme B.31
We previously showed that human granzyme B fused tothe humanized single chain antibody H22 (which binds spe-cifically and with high affinity to CD64) induces apoptosis inAML cells.32 CD64 (FcgRI) is a 72 kDa glycoprotein thatmediates endocytosis, phagocytosis, antibody-dependent cel-lular cytotoxicity and the production of cytokines and super-oxide.33 We here show that CD64 is overexpressed onprimary CMML and AMML cells and is coexpressed withother receptors such as CD33, CD14 and CD56. Viabilityand apoptosis assays on cell lines and primary CMML andAMML cells demonstrated the effectiveness of Gb-H22(scFv)and mutant GbR201K-H22(scFv) compared with H22(scFv)-ETA0 which has been evaluated extensively in previous stud-ies.34 We also discuss the ability of primary CMML andAMML cells to upregulate the granzyme B inhibitor PI9 dur-ing cultivation and correlate these effects with the greaterperformance of the mutant hCFP GbR201K-H22(scFv) com-pared to its wild-type counterpart Gb-H22(scFv).
Material and MethodsProtein expression, purification and activity
The pMS plasmid carrying the EGb-H22(scFv) constructincluding a N-terminal enterokinase cleavage site, and theexpression and purification strategy, has been described pre-viously.32 The mutations R201K and S184A were introducedinto the human granzyme B sequence by site-directed muta-genesis via the restriction sites NheI and BlpI. The immuno-toxin was activated by adding enterokinase to the protein(0.02 U/lg) in the presence of 2 mM CaCl2 and incubatingfor 16 hr at 23�C. The protein concentration was calculatedfollowing analysis by SDS-PAGE and staining with Coomas-sie Brilliant Blue using AIDA Image Analyzer Software. Thecontrol protein H22(scFv)-ETA0 was expressed by inducingperiplasmic stress in Escherichia coli and was purified byusing affinity chromatography to capture the His6-tag.
34
What’s new?
New treatment strategies for chronic (CMML) and acute myelomonocytic leukemia (AMML) are urgently needed. This study sug-
gests that CD64 is not only a promising diagnostic marker but also a novel target for specific CMML and AMML therapy. The
data show that CD64-specific human cytolytic fusion proteins kill CMML and AMML cells ex vivo, with the mutant granzyme B
protein R201K being more cytotoxic than its wild type in the presence of the granzyme B inhibitor PI-9. GbR201K-H22(scFv)
also shows superior activity to a state-of-the-art anti-CD64 immunotoxin based on Pseudomonas exotoxin A on PI-9-positive
and negative primary CMML and AMML cells.
Can
cerTherapy
2 Granzyme B based immunotherapy against CD641
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.

The activities of the wild-type and mutant proteins weredetermined by measuring Km values using the Caspase-8Assay kit (Calbiochem) according to the manufacturer’sinstructions, with Ac-IETD-pNA as a colorimetric granzymeB substrate. The OD405 was recorded for 12 min on anELISA Plate Reader (BioTek Instruments, Epoch) and Km
values were derived by non-linear curve fitting to theMichaelis–Menten equation using GraphPad Prism v4.0.
Cell lines and primary cells
Cell lines HL60 (kindly provided by Department of Immu-nology, University Medical Center Utrecht), K562 (DSMZ,ACC 10) and HEK293T (ATCC, CRL-11268) were grown inRPMI 1640 plus Gluta-MAX-I supplemented with 10% (v/v)FCS and 100 lg/ml penicillin and streptomycin (R10) at37�C and 5% CO2. HEK293T cells were selected by adding100 lg/ml zeocin after transfection.
Blood samples were taken from CMML and AMMLpatients (diagnosis based on the WHO classification as shownin Table 1) after written consent was obtained according tothe biobank rules of the University of Aachen Medical Faculty(permit number EK206/09) and approval was granted by thelocal ethics committee. Primary mononuclear cells were iso-lated from peripheral blood by density gradient centrifugationusing Biocoll with a density 1.077 g/ml (Biochrom AG). Cellswere cultured in RPMI complex medium as above.
Apoptosis assays
Cells were stimulated 24 hr (HL60) or 1–2 hr (primary cells)prior to the addition of each immunotoxin by adding 50(HL60) or 200 (primary cells) U/ml interferon c. Apoptosis-
inducing activity was measured in 12-well plates containing 1ml R10 and 2–4 3 105 cells. The cells were incubated at37�C and 5% CO2 for 24 or 48 hr following the addition ofthe immunotoxin and apoptosis was detected by their abilityto bind annexin V conjugated to FITC (Invitrogen) and totake up propidium iodide (PI). The cells were washed oncewith PBS and incubated for 20 min at room temperaturewith annexin V-FITC according to the manufacturer’sinstructions, then washed with incubation buffer (10 mMHEPES pH 7.5, 150 mM NaCl, 5 mM KCl and 2 mM CaCl2)and resuspended in the same buffer containing 5 lg/ml PI.Cell staining was analyzed by flow cytometry. The untreatedbuffer control was set to 100% viability and treated sampleswere correlated accordingly. Caspase 3/7 activity was meas-ured using the Caspase-GloTM 3/7 Assay kit (Promega)according to the manufacturer’s instructions.
Viability assays
Cytotoxicity was determined using an XTT assay. Cells wereincubated in 96-well-plates with 5000 HL60 cells or 20,000 pri-mary cells per 100 ll of medium. The immunotoxins were addedto different total concentrations and the cells were incubated for72 hr at 37�C. Independently, XTT was used to determine viablecells after incubation with single doses of CFPs in 12-well-plates.Therefore 100 ll cells were transferred to 96-well-plates andincubated with 50 ll XTT for at least 4 hr. Viability curves andIC50 values were evaluated with GraphPad Prism v4.0.
Detection of PI9 in cell lysates
Cells were lysed with PBS containing 1% Triton X-100 fol-lowed by incubation on ice for 30 min. After centrifugation,the cell lysates were boiled with Laemmli buffer and 40 lg oftotal protein was separated by SDS-PAGE. The proteins weretransferred to a membrane and probed with mouse anti-human PI9 (7D8, Santa Cruz) and goat anti-mouse IgG con-jugated to horseradish peroxidase. The resulting signal wasdetected using the enhanced chemiluminescence system (BDBioScience). The total protein concentration in the cell lysateswas determined by Bradford analysis.
Flow cytometry
The binding of immunotoxins to target cells was analyzed byincubating 2–4 3 105 cells with 500–1000 ng of each proteinfor 30 min on ice, washing with PBS and detecting boundproteins with anti-granzyme B conjugated to phycoerythrin(PE; Invitrogen) or anti-His6 conjugated to Alexa 488 (Qia-gen). The binding of antibodies to the cell surface was carriedout in the same manner to investigate the distribution of sur-face proteins. We used anti-CD64 PE, anti-CD14 allophyco-cyanin (APC), anti-CD56 APC, anti-CD56 PE, anti-CD33APC and anti-CD71 PE reagents from eBioscience, anti-CD64 Alexa 488 from BioLegend, anti-CD14 FITC fromDako and anti-CD64 Alexa647 and anti-CD64 FITC fromABD Serotec. The cells were incubated for 15 min at roomtemperature with antibody concentrations recommended by
Table 1. Characteristics of the 7 patients
Parameter Number
Number of patients 7
Age, years (median and range) 62.3 (45 – 83)
Gender
Male 6
Female 1
WHO subtype
CMML-I 4
AMML 3
Laboratory parameters (median and range)
WBC (G/l) 86,74 (31.5 – 268)
% Monocytes 29,8 (7 – 70)
Hemoglobin (g/l) 95.4 (86 – 111)
Platelet count (G/l) 311 (20 – 1212)
Previous treatment
No previous treatment 5
Hydroxyurea 2
(CMML II and III)
Can
cerTherapy
Schiffer et al. 3
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.

the manufacturers. Kd values were evaluated for differentconcentrations of Gb-H22(scFv) and GbR201K-H22(scFv) bynonlinear regression in GraphPad Prism v4.0.
Confocal microscopy
Antibody binding and internalization were confirmed byincubating 4 3 105 cells with the antibodies described aboveat 4�C and monitoring the cells by confocal microscopy. Forinternalization studies, purified H22(scFv)-SNAP was coupledto 50 nM BG-Vista-Green (NEB) as previously described.35
After washing, binding and internalization was observedusing a Leica confocal microscope (TCS SP5).
Serum stability
The stability of each immunotoxin was determined by incu-bating 70 ng/ll purified protein with 50% human serum at37�C for different time periods, then taking 7.5 ll samplesand incubating 100 ll of cells with a 400-fold dilution of theprotein-serum mixture and measuring the amount of func-tional immunotoxin remaining after incubation by FACS asdescribed.36
Statistics
A two-tailed t-test was used to determine the statistical sig-nificance of the data using GraphPad Prism v4.0 and a signif-icance threshold of p < 0.05.
ResultsEnzymatic and specific cytotoxic activity of Gb-H22(scFv)
and GbR201K-H22(scFv)
The yield of each purified protein was 1–2 mg/l of superna-tant, as shown in Figure 1a. Cleavage of the pro-peptide withenterokinase exposed the free N-terminus required for activ-ity, and the KM values were comparable for both constructswith 104 lM for Gb-H22(scFv) and 105 lM for GbR201K-H22(scFv) (data not shown). Flow cytometry using CD641
HL60 cells and CD642 control cells confirmed specific bind-ing to CD64 (Fig. 1b). The affinity constant of both proteinswas 2.1 nM indicating specific, high-affinity binding to thereceptor (data not shown).
Efficient, CD64-specific internalization of the proteins intoHL60 cells was demonstrated using H22(scFv)-SNAP-BG-647, revealing the intracellular accumulation of the red-labeled protein after incubation for 30 min 37�C, whereas nointernalization was detected at 4�C (Fig. 1c). Cell viabilitywas measured after incubating HL60 cells with different con-centrations of Gb-H22(scFv), GbR201K-H22(scFv) or thebuffer control for 72 hr (Fig. 1d). The IC50 values for bothconstructs fell within the range 4–7 nM revealing no signifi-cant differences in toxicity.
The induction of apoptosis was confirmed by exposingcells to 21 nM of each immunotoxin followed by stainingwith annexin V and PI. This revealed that >50% of theCD641 HL60 cells underwent apoptosis within 48 hr whereasthere was no impact on the CD64– CML cell line K562 (Fig.
1e). The granzyme B mutant was shown to induce apoptosisvia caspase 3/7 in HL60 cells exposed to 33 nM GbR201K-H22(scFv) for 48 hr (Fig. 1f).
Analysis of cell surface markers on primary cells
The auditability of CD64 as a target for the treatment ofAMML and CMML is unclear, so we investigated its expres-sion profile compared with the known AMML and CMMLmarkers CD56, CD33 and CD14. Normal monocytes showalso expression of CD64, CD14 and CD33, but CD56 is con-sidered to be an aberrant cell surface marker in CMML andAML. The marker profile was determined by flow cytometryand confocal microscopy using specific detection reagents(Fig. 2a). CD64 was expressed on 56–98% of the cells in allpatient samples, and was coexpressed with all three of themarkers tested. Cells from patient CMML-I clearly showeddouble staining for CD64 and CD56 (Fig. 2b) whereas cellsfrom patient CMML-III showed double staining with CD64and CD56, CD33 and CD14 (Fig. 2c). In contrast to that, thecoexpression of CD64 with the established monocyte markerCD14 was between 6 and 9% within blood samples from 6healthy volunteers (data not shown).
After the detection of CD64 expression, specific bindingof the H22(scFv)-derived fusion proteins was confirmed asdescribed above for HL60 cells (data not shown). CD64-dependent internalization was also demonstrated after theincubation of primary cells with H22(scFv)-BG-Vista-Greenas shown for patient CMML-IV in Figure 2d. The presenceof some unstained cells confirmed the absence of nonspecificbinding to CD64– cells.
Endogenous PI9 expression in target cells
Cell lysates from each of the primary cell cultures were pre-pared and tested for the expression of PI9, to determinewhether a correlation existed between the PI9-dependent inhi-bition of granzyme B and the observed differences in the proa-poptotic efficacy of the constructs (Fig. 5a). The primary cellswere harvested and lysed immediately after isolation fromperipheral blood and again after different time points either inthe presence or absence of IFNg to evaluate whether endoge-nous PI9 was upregulated during cultivation or in response toexternal stimuli. Figure 5b shows the representative results forthe cells derived from patient CMML-IV. These data showthat PI9 expression depends on the cultivation time but notthe presence or absence of IFNg. All CMML samples with theexception of CMML-II expressed PI9 after at least 14 hr incu-bation in R10 medium, but among the AMML samples onlyAMML-I was positive for PI9. The PI9-negative cells did notexpress PI9 after 5 days in supplemented RPMI medium.
Cytotoxic activity of Gb-H22(scFv) and GbR201K-H22(scFv)
against primary CMML and AMML cells
Primary cells were incubated with Gb-H22(scFv) and its PI9resistant derivative GbR201K-H22(scFv) with H22(scFv)-ETA0 used as a control. The cells were stimulated with IFNg
Can
cerTherapy
4 Granzyme B based immunotherapy against CD641
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.

to ensure continued activation, as confirmed by flow cytome-try using CD64-specific antibodies. In the absence of IFNg,the CFPs did not induce apoptosis in the targeted cells (datanot shown). Apoptosis and viability were measured asdescribed above using the XTT assay (Fig. 3a) and combinedstaining with annexin V and PI (Fig. 3b).
A statistically significant reduction in cell viabilitywas observed for GbR201K-H22(scFv) compared with thewild-type protein in cells taken from patients CMML-I,CMML-III, CMML-IV and AMML-I (comparison with agranzyme M-based CFP partially published in Ref. 37). Thisdifference was most evident in cells from patients CMML-IV
Figure 1. (a) SDS-PAGE of the purified fusion protein before (EGb-H22) and after (Gb-H22, 60 kDa) Enterokinase cleavage. M, prestained
protein marker (New England Biolabs); (b) Specific binding to CD64 positive HL60 cells (left) detected via anti-His Alexa 488 during flow
cytometry, no binding to CD64 negative cell line Ramos (right). Anti-His Alexa 488 control (dotted curve), GbR201K-H22(scFv) (gray curve)
and Gb-H22(scFv) (black curve); (c) Internalization of H22(scFv)-SNAP 647 into HL60 detected via confocal microscopy. (d,f): Specific cyto-
toxicity on CD641 cancer cell line HL60. (d) Cytotoxic effect of Gb-H22(scFv) and GbR201KH22 (scFv) measured with XTT: Cells were incu-
bated for 72 hr with different protein concentrations in 96 well plates. Data represent mean values with standard deviations of triplicates.
(e) Specific toxic effect of Gb-H22(scFv) and GbR201K-H22(scFv) on HL60 cells whereas no toxic effect seen on CD64- CML cell line K562,
measured via AnnexinV-FITC 1 PI. (f) Result of caspase 3/7 assay from Promega after incubation of HL60 with 33 nM immunotoxin or Zeo-
cin for 48 hr. Error bars represent standard deviations of at least three independent experiments.
Can
cerTherapy
Schiffer et al. 5
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.

and AMML-I. In contrast, there was no significant differencebetween the constructs in cytotoxicity towards cells frompatients CMML-II, AMML-II and AMML-III (Fig. 3a). Thecytotoxicity of H22(scFv)-ETA0 was equivalent to that of themutant granzyme B construct against cells from patientCMML-III but significantly lower than both Gb-H22(scFv)and GbR201K-H22(scFv) in the serpin B9-negative AMML
III, and CMML II, as well as the serpin B9-positive CMMLIV patient probes. Annexin V and PI staining confirmed thatcytotoxicity was in all cases mediated by inducing apoptosis.The relative cytotoxicity of each construct followed the sametrend demonstrated in the XTT assays, although the overallviability was much higher (Fig. 3b). The reason for the largerdifference seen for AMML-I might be due to a different well
Figure 2. Phenotyping of primary leukemic cells. PBMCs were isolated from peripheral blood of leukemic patients (CMML or AMML) and
incubated with specific labeled antibodies. Binding was detected by flow cytometry or confocal microscopy. (a) Overview of expression pro-
file of patient-derived primary leukemic cells determined via flow cytometry; %: portion of receptor positive cells within the whole isolated
cell population; n.d.: not determined; numbers in brackets: strong CD64 expression detected. (b,c) Confocal microscopy of cells from
patient CMML I (b) and CMML III (c) stained with antibodies as indicated. (d) 50 nM H22(scFv)-SNAP-BG-Vista-Green was incubated with
4*105 CD64-positive primary cells (exemplary shown for patient CMML IV) for 10 min at 37�C and internalization was detected with confo-
cal microscopy.
Can
cerTherapy
6 Granzyme B based immunotherapy against CD641
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.
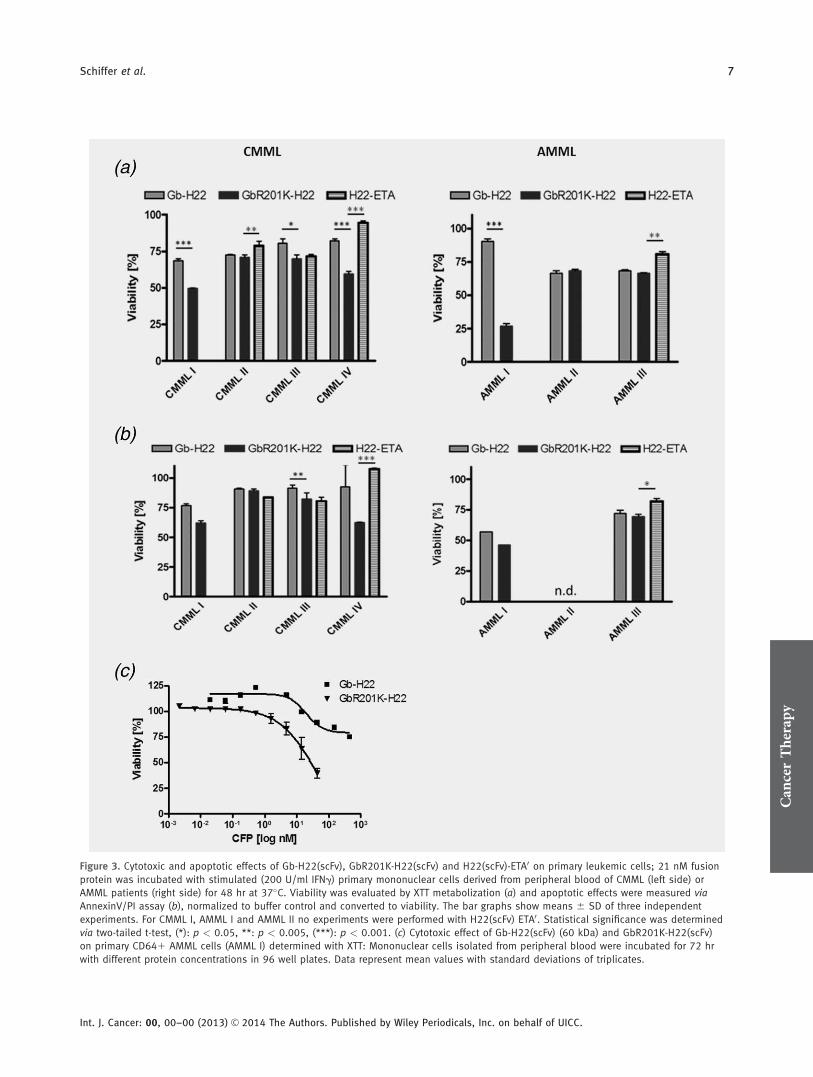
Figure 3. Cytotoxic and apoptotic effects of Gb-H22(scFv), GbR201K-H22(scFv) and H22(scFv)-ETA0 on primary leukemic cells; 21 nM fusion
protein was incubated with stimulated (200 U/ml IFNg) primary mononuclear cells derived from peripheral blood of CMML (left side) or
AMML patients (right side) for 48 hr at 37�C. Viability was evaluated by XTT metabolization (a) and apoptotic effects were measured via
AnnexinV/PI assay (b), normalized to buffer control and converted to viability. The bar graphs show means 6 SD of three independent
experiments. For CMML I, AMML I and AMML II no experiments were performed with H22(scFv) ETA0. Statistical significance was determined
via two-tailed t-test, (*): p < 0.05, **: p < 0.005, (***): p < 0.001. (c) Cytotoxic effect of Gb-H22(scFv) (60 kDa) and GbR201K-H22(scFv)
on primary CD641 AMML cells (AMML I) determined with XTT: Mononuclear cells isolated from peripheral blood were incubated for 72 hr
with different protein concentrations in 96 well plates. Data represent mean values with standard deviations of triplicates.
Can
cerTherapy
Schiffer et al. 7
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.

plate format used in the assays. Incubation of the primarycells with the inactive variant GbS184A-H22(scFv)38 and thenonbinding control GbR201K-Ki4(scFv)39 had no impact oncell viability (data shown in Schiffer et al.37 and SupportingInformation Fig. S1, respectively).
The concentration-dependent cytotoxicity of the granzymeB constructs was also determined using the XTT assay, whichconfirmed that the difference between constructs was mostevident in cells from patient AMML-I (Fig. 3c).
Specificity of toxicity toward primary cells
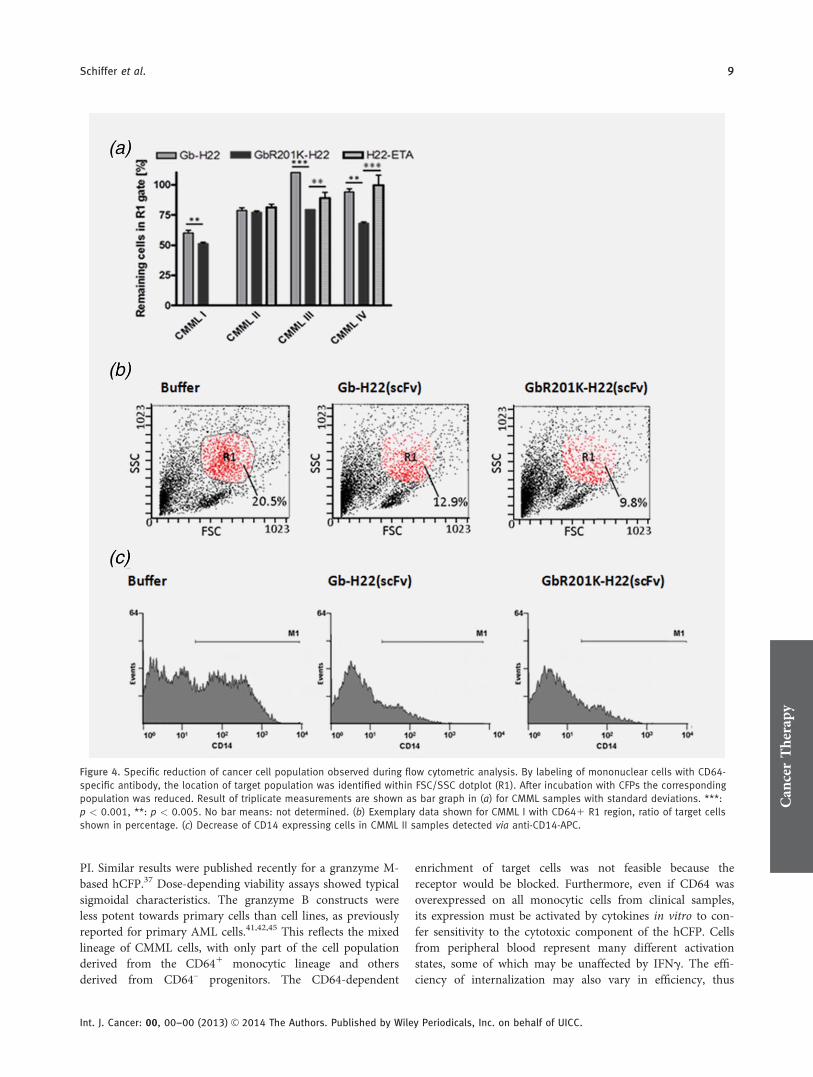
The targeted cytotoxicity of the granzyme B constructstowards malignant cells was confirmed using independentapproaches. First, the location of the target cell population(R1) within the FCS/SSC dotplot was established using aCD64-specific antibody. After treatment with Gb-H22(scFv)or GbR201K-H22(scFv), this particular population declinedby 50%, as shown for the cells from CMML patients in Fig-ure 4a. The data representing cells from patient CMML-I areshown in Figure 4b, where the R1 population declined to agreater extent when treated with GbR201K-H22(scFv) com-pared with the wild-type construct. These data support theresults from our XTT and annexin V/PI assays. Second, thetreated cells were stained with antibodies against cell surfacereceptors that are coexpressed with CD64, allowing us tomonitor the population dynamics of cells displaying particu-lar markers. The results for cells from patient CMML II areshown in Figure 4c, revealing a clear reduction in the treatedsamples compared to the buffer control and thus indicatingthat cell death is targeted and specific.
Serum stability of Gb-H22(scFv) and GbR201K-H22(scFv)
Serum stability is necessary for the successful systemic applica-tion of immunotoxins, so Gb-H22(scFv) and GbR201K-H22(scFv) were each incubated in either 50% human serumderived from patient CMML-III or in PBS as a control. Theproteins were incubated at 37�C for 1, 6 or 24 hr. Stability wasdetermined by measuring residual binding activity by flowcytometry (Fig. 5c). Both hCFP variants showed similar stabil-ity profiles, remaining stable for at least 6 hr and declining after24 hr. Residual binding activity remained after 24 hr but theloss was significant compared with incubation in PBS, whereonly 8% loss of binding activity was observed after 24 hr.
DiscussionThe value of CD64 as a target for specific CMML andAMML therapy was difficult to establish, so we investigatedthe expression of CD64 on CMML and AMML cells, its rela-tionship with established tumor markers, and the ability ofhCFP targeting CD64 to induce apoptosis specifically inCMML and AMML cells. We used hCFPs comprising thehumanized anti-CD64 scFv H22 and the human enzymegranzyme B, which induces apoptosis. We tested native gran-zyme B against a mutant derivative that lacks sensitivity tothe specific inhibitor PI9.
We found that both Gb-H22(scFv)32 and its PI9-resistantderivative GbR201K-H22(scFv)31 can induce apoptosis to thesame extent in the CD641 HL60 cell line which does notexpress PI9. We also found that CD64 is a potential targetfor the treatment of CMML and AMML and that CD64 isexpressed on the majority of CMML and AMML cells. Weshowed that CD64-specific immunotoxins specifically killCD641 CMML and AMML cells and that although the wild-type and PI9-resistant mutant are equally toxic towards PI9–
cells, the mutant version is advantageous if PI9 is upregu-lated, for example, when CMML and AMML cells are culti-vated. Both granzyme B hCFP were more cytotoxic than acontrol construct containing H22 and the bacterial toxinETA0. These hCFPs can also be used at higher doses and forrepetitive treatments because they might be less immunogenicthan toxins from bacteria or plants. We have previouslyshown that, although they do express CD64, normal mono-cytes are not affected by CD64-specific immunotoxins usingH22 as targeting moiety in the presence of IFNg.40
The CD641 AML cell line HL60 was exposed to each ofthe granzyme B constructs and we found that both the wild-type and mutant versions were cytotoxic with similar IC50
values. Annexin V staining and caspase 3/7 assays confirmedthe induction of apoptosis. CD642 cells were unaffected byeither construct, confirming the specificity of the approachand suggesting it could be used to target tumor cells even inmixed populations of PBMCs.
Several previous studies have reported the targeting of pri-mary AML cells using reagents that bind CD3341,42 orCD64,32,43 but these have not focused on the treatment ofCMML or AMML. The development of targeted therapeuticsfor these diseases is restricted by the lack of specific geneticaberrations that can be influenced by small molecules or anti-bodies and the lack of useful in vitro and in vivo diseasemodels. We therefore isolated monocytes from the freshperipheral blood of CMML and AMML patients, identifiedthe malignant cells using specific antibodies and determinedthe percentage of cells positive for CD64, CD56, CD14 andCD33 within the mixed cell population.44 CD64 is often dis-cussed in context of CMML but its abundance was reportedto decline in this disease based on the immunohistochemicalstaining of bone marrow.4,44 However, our flow cytometryand confocal microscopy experiments clearly demonstratedthat CD64 was overexpressed in all the CMML samples thatwere analyzed, and showed a tendency to be coexpressedwith the CMML-associated markers CD56, as well as CD33and CD14. These findings indicated that CD64 is not only avalid target for the treatment of AML, but also CMML, asCD64 coexpressing, CD56 positive malignant cells are alsoeliminated. Overexpression of CD64 on AMML cells hasbeen reported previously.2
The CD64-specific hCFPs based on granzyme B were ableto kill CD641 primary CMML and AMML cells, as con-firmed both by testing residual mitochondrial activity usingXTT assays, and by combined staining with annexin V and
Can
cerTherapy
8 Granzyme B based immunotherapy against CD641
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.
PI. Similar results were published recently for a granzyme M-based hCFP.37 Dose-depending viability assays showed typicalsigmoidal characteristics. The granzyme B constructs wereless potent towards primary cells than cell lines, as previouslyreported for primary AML cells.41,42,45 This reflects the mixedlineage of CMML cells, with only part of the cell populationderived from the CD641 monocytic lineage and othersderived from CD64– progenitors. The CD64-dependent
enrichment of target cells was not feasible because thereceptor would be blocked. Furthermore, even if CD64 wasoverexpressed on all monocytic cells from clinical samples,its expression must be activated by cytokines in vitro to con-fer sensitivity to the cytotoxic component of the hCFP. Cellsfrom peripheral blood represent many different activationstates, some of which may be unaffected by IFNg. The effi-ciency of internalization may also vary in efficiency, thus
Figure 4. Specific reduction of cancer cell population observed during flow cytometric analysis. By labeling of mononuclear cells with CD64-
specific antibody, the location of target population was identified within FSC/SSC dotplot (R1). After incubation with CFPs the corresponding
population was reduced. Result of triplicate measurements are shown as bar graph in (a) for CMML samples with standard deviations. ***:
p < 0.001, **: p < 0.005. No bar means: not determined. (b) Exemplary data shown for CMML I with CD641 R1 region, ratio of target cells
shown in percentage. (c) Decrease of CD14 expressing cells in CMML II samples detected via anti-CD14-APC.
Can
cerTherapy
Schiffer et al. 9
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.

affecting the likely response to treatment. At this stage, it isunclear whether CD64 is present only on mature monocytesor also on leukemic stem cells in CMML or AMML, so it isimpossible to tell whether CD64-targeting immunotoxins wouldreach all cells or only the monocytic component. The granzymeB hCFP may also be suitable for AML, which also has limitedoptions for targeted therapy and a poor prognosis.46
In contrast to the results obtained in the cell lines, thecontrol construct H22(scFv)-ETA0 demonstrated uncharacter-istically low cytotoxicity against the primary CMML cellsdespite its potency against other cancers.47 In fact, we wereable to demonstrate, that hCFPs based on our serpin B9-resistant granzyme B mutant were active also in primaryprobes showing insensitivity to Pseudomonas exotoxin A.This insensitivity may reflect innate resistance based on thelack of CMML cell proliferation ex vivo, as described previ-ously for constructs based on diphtheria toxin48 or for con-structs based on Pseudomonas exotoxin A.49
We also investigated the relationship between the cytotox-icity of the PI9-sensitive and PI9-resistant constructs and theabundance of PI9 immediately after the isolation of cellsfrom peripheral blood and again after cultivation for differenttime intervals. Western blots of all samples showed minimalsignals just after isolation but strong bands during cultiva-tion, suggesting that PI9 is upregulated in culture. PI9 wasalso expressed in lymphocytes and monocytes isolated fromPBMCs (data not shown) as previously described.50 It is
unclear whether PI9 expression is a hallmark of themalignant CMML and AMML cells or a natural backgroundsignal, but we did not observe any upregulation of PI9 byIFNg in contrast to previous reports,50 although this mayreflect differences in the detection method. Nevertheless, thepresence of PI9 in CMML and AMML target cells diminishesthe potential of granzyme B immunotherapy, and the PI9-resistant mutant would therefore be necessary to increasetherapeutic efficacy. Accordingly, we found a correlationbetween PI9 expression and efficacy of the GbR201K-H22(scFv) mutant, which was much more potent than thewild-type construct in the presence of PI9, whereas there wasno difference in cytotoxicity between the constructs when PI9was not expressed. Further investigation is required to deter-mine whether PI9 expression is a hallmark of CMML orAMML in vivo, and could be used as a diagnostic marker forthe selection of appropriate treatment approaches.
We carried out serum stability assays to determinewhether the constructs are suitable for in vivo applicationsand found that both constructs retained the majority of theircell-binding and cytotoxic activity after incubation in humanserum for 6 h. Since H22(scFv) internalizes rapidly and has ahigh binding affinity based on the observed Kd values, stabil-ity for 6 hr should be more than sufficient.51 The stability forH22(scFv)-ETA0 has been evaluated previously and retained77% activity after incubation for 24 hr.47 This is 10% morestable than the granzyme B constructs although it should be
Figure 5. (a,b) Expression of endogenous PI-9 in primary leukemic cells derived from peripheral blood of CMML or AMML patients. (a) Over-
view of PI-9 expression for all determined primary patient cells after at least 14 hr cultivation duration. (b) Exemplary data for patient
CMML IV. Cells were incubated in R10 in presence or absence of 200 U/ml IFNg for different time intervals, before harvesting. Total soluble
protein from cell lysates (40.5 g/lane) was separated by SDS-PAGE and transferred to a nitrocellulose membrane. Endogenous PI-9 was
detected in a western blot using anti-human PI-9 and GAM-PO. M: Prestained broad range marker in kDa; Pos.: recombinant PI 9 as positive
control. (c) Serum stability of Gb-H22(scFv) and GbR201K-H22(scFv) in human patient serum (CMML III) or PBS determined by flow cytome-
try according to residual binding activity to target cells.
Can
cerTherapy
10 Granzyme B based immunotherapy against CD641
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.

noted that human serum was used in this study and mouseserum was used previously.51
In conclusion, we have demonstrated the efficacy ofhuman wild-type and mutant granzyme B based hCFPs thattarget CD641 cells including cell line HL60 and primaryCMML and AMML cells. The constructs induced apoptosisspecifically in the target cells and therefore have a strongclinical potential because of their specificity, potency and theabsence of immunogenicity. Furthermore, we correlated theexpression of CD64 with known markers for CMML andAMML, suggesting CD64 could be a novel diagnostic markerand therapeutic target for the treatment of CMML andAMML patients. We were able to specifically kill CMML andAMML cells ex vivo and the GbR201K mutant showed highercytotoxicity than the wild-type construct, against cellsexpressing the inhibitor PI9 allowing these cells to escapefrom cytotoxic immune effector cell attack. Interestingly,GbR201K-H22(scFv) also demonstrated superior activity tosome AMML and CMML probes less sensitive to Pseudomo-nas exotoxin A. This might have major consequences for
future personalized treatment approaches allowing selectingfor the population of patients responding best to a cell sur-face receptor specific immunotherapeutic agent and mightconsequently pave the way for the identification of the bestagent from a portfolio of different immunotherapeutic agentsfor efficient treatment of a specific indication. Therefore, thePI9-resistant version of granzyme B is a better candidatecytotoxic component than its wild-type counterpart for tar-geted immunotherapy against CMML, AMML and probablyother cancers expressing this inhibitor.
Authors’ ContributionsS.S., E.J. and T.T. wrote the article; S.S., E.J., M.H. and R.R.conceived, executed and analyzed the experiments; G.H.-T.,G.M. and S.B. provided critical intellectual input and contrib-uted to writing the manuscript.
AcknowledgementsThe authors thank Richard M. Twyman for critical reading of themanuscript.
References
1. Vardiman JW, Imbert M, Pierre R, et al. Chronicmyelomonocytic leukaemia. In: Jaffe ES, HarrisNL, Sten H, Vardiman JW, eds. World HealthOrganization Classification of Tumors: Pathologyand Genetics of Tumours of Haematopoietic andLymphoid Tissues. Lyon: IARC Press, 2001. 49–52.
2. Vardiman JW, Harris NL, Brunning RD. TheWorld Health Organization (WHO) classificationof the myeloid neoplasms. Blood 2002;100:2292–302.
3. Hall J, Foucar K. Diagnosing myelodysplastic/myeloproliferative neoplasms: laboratory testingstrategies to exclude other disorders. Int J LabHematol 2010;32:559–71.
4. Orazi A, Germing U. The myelodysplastic/myelo-proliferative neoplasms: myeloproliferative dis-eases with dysplastic features. Leukemia 2008;22:1308–19.
5. Xu Y, McKenna RW, Karandikar NJ, et al. Flowcytometric analysis of monocytes as a tool fordistinguishing chronic myelomonocytic leukemiafrom reactive monocytosis. Am J Clin Pathol2005;124:799–806.
6. Vuckovic S, Fearnley DB, Gunningham S, et al.Dendritic cells in chronic myelomonocytic leu-kaemia. Brit J Haematol 1999;105:974–85.
7. Orazi A, Chiu R, O’Malley DP, et al. Chronicmyelomonocytic leukemia: the role of bone mar-row biopsy immunohistology. Mod Pathol 2006;19:1536–45.
8. Fenaux P, Beuscart R, Lai JL, et al. Prognosticfactors in adult chronic myelomonocytic leuke-mia: an analysis of 107 cases. J Clin Oncol 1988;6:1417–24.
9. Onida F, Ball G, Kantarjian HM, et al. Character-istics and outcome of patients with Philadelphiachromosome negative, bcr/abl negative chronicmyelogenous leukemia. Cancer 2002;95:1673–84.
10. Itzykson R, Kosmider O, Renneville A, et al. Clo-nal architecture of chronic myelomonocytic leu-kemias. Blood 2013;121:2186–98.
11. Germing U, Neukirchen J. How to treat patientswith CMML? Leuk Res 2013;37:605–606.
12. Fenaux P, Mufti GJ, Hellstrom-Lindberg E, et al.Efficacy of azacitidine compared with that of con-ventional care regimens in the treatment ofhigher-risk myelodysplastic syndromes: a rando-mised, open-label, phase III study. Lancet Oncol2009;10:223–232.
13. Rollins-Raval MA, Roth CG. The value of immu-nohistochemistry for CD14, CD123, CD33,myeloperoxidase and CD68R in the diagnosis ofacute and chronic myelomonocytic leukaemias.Histopathology 2012;60:933–42.
14. Kreitman RJ. Immunotoxins for targeted cancertherapy. AAPS J 2006;8:E532–51.
15. Walter RB, Appelbaum FR, Estey EH, et al. Acutemyeloid leukemia stem cells and CD33-targetedimmunotherapy. Blood 2012;119:6198–208.
16. Mathew M, Verma RS. Humanized immunotoxins:a new generation of immunotoxins for targetedcancer therapy. Cancer Sci 2009;100:1359–65.
17. Kurschus FC, Jenne DE. Delivery and therapeuticpotential of human granzyme B. Immunol Rev2010;235:159–71.
18. Liu Y, Cheung LH, Hittelman WN, et al. Tar-geted delivery of human pro-apoptotic enzymesto tumor cells: in vitro studies describing a novelclass of recombinant highly cytotoxic agents. MolCancer Ther 2003;2:1341–50.
19. Kurschus FC, Fellows E, Stegmann E, et al. Gran-zyme B delivery via perforin is restricted by size,but not by heparan sulfate-dependent endocytosis.Proc Natl Acad Sci U S A 2008;105:13799–804.
20. Zhang L, Zhao J, Wang T, et al. HER2-targetingrecombinant protein with truncated pseudomonasexotoxin A translocation domain efficiently killsbreast cancer cells. Cancer Biol Therapy 2008;7:1226–31.
21. Clement MV, Haddad P, Soulie A, et al. Involve-ment of granzyme B and perforin gene expressionin the lytic potential of human natural killer cells.Nouv Rev Fr Hematol 1990;32:349–52.
22. Raja SM, Wang B, Dantuluri M, et al.Cytotoxic cell granule-mediated apoptosis. Char-acterization of the macromolecular complex ofgranzyme B with serglycin. J Biol Chem 2002;277:49523–30.
23. Barry M, Heibein JA, Pinkoski MJ, et al. Gran-zyme B short-circuits the need for caspase 8activity during granule-mediated cytotoxic T-lymphocyte killing by directly cleaving Bid. MolCell Biol 2000;20:3781–94.
24. Metkar SS, Wang B, Ebbs ML, et al. Granzyme Bactivates procaspase-3 which signals a mitochon-drial amplification loop for maximal apoptosis. JCell Biol 2003;160:875–85.
25. Trapani JA, Jans DA, Jans PJ, et al. Efficientnuclear targeting of granzyme B and the nuclearconsequences of apoptosis induced by granzymeB and perforin are caspase-dependent, but celldeath is caspase-independent. J Biol Chem 1998;273:27934–8.
26. Andrade F, Roy S, Nicholson D, et al. GranzymeB directly and efficiently cleaves severaldownstream caspase substrates: implicationsfor CTL-induced apoptosis. Immunity 1998;8:451–60.
27. Ray M, Hostetter DR, Loeb CR, et al. Inhibitionof Granzyme B by PI-9 protects prostate cancercells from apoptosis. Prostate, in press.
28. Rousalova I, Krepela E, Prochazka J, et al.Expression of proteinase inhibitor-9/serpinB9 innon-small cell lung carcinoma cells and tissues.Int J Oncol 2010;36:275–83.
29. ten Berge RL, Meijer CJ, Dukers DF, et al.Expression levels of apoptosis-related proteinspredict clinical outcome in anaplastic large celllymphoma. Blood 2002;99:4540–6.
30. Bladergroen BA, Meijer CJ, ten Berge RL, et al.Expression of the granzyme B inhibitor, proteaseinhibitor 9, by tumor cells in patients with non-Hodgkin and Hodgkin lymphoma: a novel pro-tective mechanism for tumor cells to circumventthe immune system? Blood 2002;99:232–7.
Can
cerTherapy
Schiffer et al. 11
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.

31. Losasso V, Schiffer S, Barth S, et al. Design ofhuman granzyme B variants resistant to serpinB9. Proteins 2012;80:2514–22.
32. Stahnke B, Thepen T, Stocker M, et al. GranzymeB-H22(scFv), a human immunotoxin targetingCD64 in acute myeloid leukemia of monocyticsubtypes. Mol Cancer Ther 2008;7:2924–32.
33. van de Winkel JG, Anderson CL. Biology ofhuman immunoglobulin G Fc receptors. J LeukocBiol 1991;49:511–24.
34. Tur MK, Huhn M, Thepen T, et al. RecombinantCD64-specific single chain immunotoxin exhibitsspecific cytotoxicity against acute myeloid leuke-mia cells. Cancer Res 2003;63:8414–9.
35. Kampmeier F, Niesen J, Koers A, et al. Rapidoptical imaging of EGF receptor expressionwith a single-chain antibody SNAP-tag fusionprotein. Eur J Nucl Med Mol Imaging 2010;37:1926–34.
36. Hetzel C, Bachran C, Fischer R, et al. Smallcleavable adapters enhance the specific cytotoxic-ity of a humanized immunotoxin directedagainst CD64-positive cells. J Immunother 2008;31:370–6.
37. Schiffer S, Letzian S, Jost E, et al. Granzyme M asa novel effector molecule for human cytolyticfusion proteins: CD64-specific cytotxicity of Gm-H22(scFv) against leukemic cells. Cancer Lett.2013;341:178–85.
38. Silverman GA, Bird PI, Carrell RW, et al. Theserpins are an expanding superfamily of structur-ally similar but functionally diverse proteins. Evo-lution, mechanism of inhibition, novel functions,and a revised nomenclature. J Biol Chem 2001;276:33293–6.
39. Schiffer S, Hansen H, Hehmann-Titt G, et al.Efficacy of an adapted Granzyme B-basedanti-CD30 cytolytic fusion protein against PI-9-positive classical Hodgkin lymphoma cells in amurine model. Blood Cancer J 2013;3:e106.
40. Thepen T, Huhn M, Melmer G, et al. Fcgammareceptor 1 (CD64), a target beyond cancer. CurrPharm Des 2009;15:2712–8.
41. ten Cate B, Bremer E, de Bruyn M, et al. A novelAML-selective TRAIL fusion protein that is supe-rior to Gemtuzumab Ozogamicin in terms of invitro selectivity, activity and stability. Leukemia2009;23:1389–97.
42. Schwemmlein M, Peipp M, Barbin K, et al. ACD33-specific single-chain immunotoxinmediates potent apoptosis of cultured humanmyeloid leukaemia cells. Br J Haematol 2006;133:141–51.
43. Zhong RK, van de Winkel JG, Thepen T, et al.Cytotoxicity of anti-CD64-ricin a chain immuno-toxin against human acute myeloid leukemia cellsin vitro and in SCID mice. J Hematother StemCell Res 2001;10:95–105.
44. Santos IM, Franzon CM, Koga AH. Laboratorydiagnosis of chronic myelomonocytic leukemiaand progression to acute leukemia inassociation with chronic lymphocytic leukemia:morphological features and immunophenotypicprofile. Rev Bras Hematol Hemoter 2012;34:242–4.
45. Nordigarden A, Zetterblad J, Trinks C, et al. Irre-versible pan-ERBB inhibitor canertinib elicitsanti-leukaemic effects and induces the regressionof FLT3-ITD transformed cells in mice. Br J Hae-matol 2011;155:198–208.
46. Krauter J, Wagner K, Schafer I, et al. Prognosticfactors in adult patients up to 60 years old withacute myeloid leukemia and translocations ofchromosome band 11q23: individual patient data-based meta-analysis of the German Acute Mye-loid Leukemia Intergroup. J Clin Oncol 2009;27:3000–6.
47. Tur MK, Huhn M, Jost E, et al. In vivo efficacyof the recombinant anti-CD64 immunotoxinH22(scFv)-ETA0 in a human acute myeloid leuke-mia xenograft tumor model. Int J Cancer 2011;129:1277–82.
48. Jedema I, Barge RM, Frankel AE, et al. Acutemyeloid leukemia cells in G0 phase of the cellcycle that are unresponsive to conventionalchemotherapy are sensitive to treatment withgranulocyte-macrophage colony-stimulating fac-tor/diphtheria toxin fusion proteins. Exp Hematol2004;32:188–94.
49. Wei H, Bera TK, Wayne AS, et al. A modifiedform of diphthamide causes immunotoxin resist-ance in a lymphoma cell line with a deletion ofthe WDR85 gene. J Biol Chem 2013;288:12305–12.
50. Classen CF, Bird PI, Debatin KM. Modulation ofthe granzyme B inhibitor proteinase inhibitor 9(PI-9) by activation of lymphocytes and mono-cytes in vitro and by Epstein-Barr virus and bac-terial infection. Clin Exp Immunol 2006;143:534–42.
51. Graziano RF, Tempest PR, White P, et al. Con-struction and characterization of a humanizedanti-gamma-Ig receptor type I (Fc gamma RI)monoclonal antibody. J Immunol 1995;155:4996–5002.
Can
cerTherapy
12 Granzyme B based immunotherapy against CD641
Int. J. Cancer: 00, 00–00 (2013) VC 2014 The Authors. Published by Wiley Periodicals, Inc. on behalf of UICC.




















