
Homology between the HrpO protein of Pseudomonas solanacearum and bacterial proteins implicated in a...
16
Mol Gen Genêt (1993) 239:378-392 MGG © Springer-Verlag 1993 Homology between thé HrpO protein of Pseudomonas solanacearum and bacterial proteins implicated in a signal peptide-independent sécrétion mechanism Clare L. Gough, Stéphane Genin, Viviane Lopes, Christian A. Boucher Laboratoire de Biologie Moléculaire des Relations Plantes Microorganismes, INRA-CNRS, F-31326 Castanet-Tolosan, BP27, Cedex, France Receîved: 3 August 1992/Acceptée: 19January 1993 Âbstract. A'région of approximately 22 kb of DNA de- fines thé large hrp gène cluster of strain GMI1000 of Pseudomonas solanacearum, The majority of mutants that map to this région hâve lost thé ability to induce disease sympîoms on tomato plants and are no longer able to elicit a hypersensitive reaction (HR) on tobacco, a non- host plant. In this study we présent thé complémentation analysis and nucleotide séquence of a 4772 bp région of this hrp gène cluster. Three complète open reading frames (ORFs) are predicted within this région. The correspond- ing putative proteins, HrpN, HrpO and HpaP, hâve pre- dicted sizes of 357, 690 and 197 amino acids, respectively, and predicted molecular weights of 38607, 73990 and 21959 dalton, respectively. HrpN and HrpO are both predicted to be hydrophobic proteins with potential membrane-spanning domains and HpaP is rien in pro- line residues. A mutation in hpaP (for hrp associated) does not affect thé HR on tobacco or thé disease on tomato plants. None of thé proteins is predicted to hâve an N-terminal signal séquence, which would hâve indi- cated that thé proteins are exported. Considérable sé- quence similarities were found between HrpO and eight known or predicted prokaryotic proteins: LcrD of Yer*inia pestis and Y. enterocolitica, FlbF of Caulobacter crescentus, FlhA of Bacillus subtilis, MxiA and VirH of Shigeîla flexneri, InvA of Salmonella typhimurium and HrpC2 of Xanthomonas campestris pv. vesicatoria. Thèse homologies suggest that certain hrp gènes of phytopatho- genic bacteria code for components of a secretory System, which is related to thé Systems for sécrétion of flagellar proteins, Ipa proteins of Shigeîla flexneri and thé Yersinïa Yop proteins. Furthermore, thèse homologous proteins hâve thé common feature of being implicated in a distinct secretory mechanism, which does not require thé cleav- age of a signal peptide. The séquence similarity between HrpO and HrpC2 is particularly high (66% identity and . 81% similarity) and thé amino acid séquence comparison between thèse two proteins présentée! hère reveals thé Communicated by H. Hennecke Correspondence to: C.L. Gough first such séquence similarity to be shown between Hrp proteins of P. solanacearum and X. campestris. An efflux of plant electrolytes was found to be associated with thé interactions between P. solanacearum and both tomato and tobacco leaves. This phenomenon may be part of thé mechanism by which hrp gène products control and dé- termine plant-bacterial interactions, since hrpO mutants induced levels of leakage which were significantly lower than those induced by thé wild type on each plant. Key words: Pseudomonas solanacearum - hrp gènes - Sé- crétion - Electrolyte leakage Introduction Pseudomonas solanacearum is thé causal agent of bac- terial wilt of solanaceous crops and many other plants (Buddenhagen and Kelman 1964). Leaf infiltration of P. solanacearum into nonhost plants often induces a reac- tion known as thé hypersensitive reaction (HR), which is associated with résistance and is characterised by thé rapid, localised necrosis of plant tissue. The identification and characterisation of mutants, which hâve simul- taneously lost thé ability to provoke disease symptoms on a host plant (tomato) and thé ability to induce an HR on a nonhost plant (tobacco), has led to thé discovery in strain GMI1000 of P. solanacearum of hrp gènes (/zyper- sensitive réaction and^athogenicity; Boucher et al. 1985, 1987). There are actually two loci of P. solanacearum hrp gènes; thé one that has been described in strain GMI1000, and another in strain K60 (Huang et al. 1990). The first locus, which is thé larger of thé two, consists of hrp gènes clustered in a région of approximately 22 kb of thé megaplasmid of P. solanacearum (Boucher et al. 1986; Arlat et al. 1991). This large cluster of hrp gènes has recently been shown to be organised in a minimum of six transcription units, and factors involved in thé régulation of expression of thé gènes hâve been described (Arlat et al. 1992; Genin et al. 1992b).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Homology between the HrpO protein of Pseudomonas solanacearum and bacterial proteins implicated in a...

Mol Gen Genêt (1993) 239:378-392 MGG© Springer-Verlag 1993
Homology between thé HrpO protein of Pseudomonas solanacearumand bacterial proteins implicated in a signal peptide-independentsécrétion mechanismClare L. Gough, Stéphane Genin, Viviane Lopes, Christian A. Boucher
Laboratoire de Biologie Moléculaire des Relations Plantes Microorganismes, INRA-CNRS,F-31326 Castanet-Tolosan, BP27, Cedex, France
Receîved: 3 August 1992/Acceptée: 19January 1993
Âbstract. A'région of approximately 22 kb of DNA de-fines thé large hrp gène cluster of strain GMI1000 ofPseudomonas solanacearum, The majority of mutants thatmap to this région hâve lost thé ability to induce diseasesympîoms on tomato plants and are no longer able toelicit a hypersensitive reaction (HR) on tobacco, a non-host plant. In this study we présent thé complémentationanalysis and nucleotide séquence of a 4772 bp région ofthis hrp gène cluster. Three complète open reading frames(ORFs) are predicted within this région. The correspond-ing putative proteins, HrpN, HrpO and HpaP, hâve pre-dicted sizes of 357, 690 and 197 amino acids, respectively,and predicted molecular weights of 38607, 73990 and21959 dalton, respectively. HrpN and HrpO are bothpredicted to be hydrophobic proteins with potentialmembrane-spanning domains and HpaP is rien in pro-line residues. A mutation in hpaP (for hrp associated)does not affect thé HR on tobacco or thé disease ontomato plants. None of thé proteins is predicted to hâvean N-terminal signal séquence, which would hâve indi-cated that thé proteins are exported. Considérable sé-quence similarities were found between HrpO and eightknown or predicted prokaryotic proteins: LcrD ofYer*inia pestis and Y. enterocolitica, FlbF of Caulobactercrescentus, FlhA of Bacillus subtilis, MxiA and VirH ofShigeîla flexneri, InvA of Salmonella typhimurium andHrpC2 of Xanthomonas campestris pv. vesicatoria. Thèsehomologies suggest that certain hrp gènes of phytopatho-genic bacteria code for components of a secretory System,which is related to thé Systems for sécrétion of flagellarproteins, Ipa proteins of Shigeîla flexneri and thé YersinïaYop proteins. Furthermore, thèse homologous proteinshâve thé common feature of being implicated in a distinctsecretory mechanism, which does not require thé cleav-age of a signal peptide. The séquence similarity betweenHrpO and HrpC2 is particularly high (66% identity and .81% similarity) and thé amino acid séquence comparisonbetween thèse two proteins présentée! hère reveals thé
Communicated by H. Hennecke
Correspondence to: C.L. Gough
first such séquence similarity to be shown between Hrpproteins of P. solanacearum and X. campestris. An effluxof plant electrolytes was found to be associated with théinteractions between P. solanacearum and both tomatoand tobacco leaves. This phenomenon may be part of thémechanism by which hrp gène products control and dé-termine plant-bacterial interactions, since hrpO mutantsinduced levels of leakage which were significantly lowerthan those induced by thé wild type on each plant.
Key words: Pseudomonas solanacearum - hrp gènes - Sé-crétion - Electrolyte leakage
Introduction
Pseudomonas solanacearum is thé causal agent of bac-terial wilt of solanaceous crops and many other plants(Buddenhagen and Kelman 1964). Leaf infiltration ofP. solanacearum into nonhost plants often induces a reac-tion known as thé hypersensitive reaction (HR), whichis associated with résistance and is characterised by thérapid, localised necrosis of plant tissue. The identificationand characterisation of mutants, which hâve simul-taneously lost thé ability to provoke disease symptomson a host plant (tomato) and thé ability to induce an HRon a nonhost plant (tobacco), has led to thé discovery instrain GMI1000 of P. solanacearum of hrp gènes (/zyper-sensitive réaction and^athogenicity; Boucher et al. 1985,1987). There are actually two loci of P. solanacearum hrpgènes; thé one that has been described in strainGMI1000, and another in strain K60 (Huang et al. 1990).The first locus, which is thé larger of thé two, consists ofhrp gènes clustered in a région of approximately 22 kb ofthé megaplasmid of P. solanacearum (Boucher et al.1986; Arlat et al. 1991). This large cluster of hrp gèneshas recently been shown to be organised in a minimumof six transcription units, and factors involved in thérégulation of expression of thé gènes hâve been described(Arlat et al. 1992; Genin et al. 1992b).

379
hrp gènes hâve been identified in most of thé maingroups of Gram-negative phytopathogenic bacteria (Wil-lis et a!. 1991). Between related bacteria thèse gènes arefunctionally conserved: within pathovars of Pseudo-monas syringae (Lindgren et al. 1988), pathovars ofXanthomonas campestris (Bonas et al. 1991 ; Arlat et al.1991), strains of P. solanacearum (Boucher et al. 1988)and probably even between two différent bacteriaï spe-cies, P. solanacearum and X. campestris (Arlat et al.1991). Also, DNA homology has been detected betweenhrp gènes of phylogenetically distinct bacteria, betweenvarious Erwinia species and between Erwinia amyiovoraand P. syringae (Laby and Béer 1990). This widespreadoccurrence and conservation of hrp gènes among phy-topathogenic bacteria implies that many différent bac-terial plant diseases involve common mechanisms. How-ever, while thé transcriptional organisation of several hrpgène clusters has been described (Lindgren et al. 1989;Barnyetal. 1990; Bonas et al. 1991 ; Rahme et al. 1991;Arlat et al. 1991, 1992; Bauer and Béer 1991), and certainhrp loci hâve been sequenced (Mukhopadhyay et al.1988; Grimm and Panopoulos 1989; Sneath et al. 1990;Gough et al. 1992; Fenselau et al. 1992), it is not knownhow hrp gène products control and détermine thé out-come of plant-pathogen interactions. TnphoA analysis ofP. syringae pv. syringae hrp gènes has led to thé discoverythat two Hrp proteins are either exported or hâve peri-plasmic domains (Huang et al. 1991). Based on this, andséquence homologies with Yersinia virulence déter-minants, we hâve proposed that hrp gènes code for aspécifie export System which is responsible for thé exportof an elicitor (Gough et al. 1992). Recently it has beenshown that an hrp gène of E. amyiovora codes for a cellenvelope-associated elicitor of thé hypersensitive re-sponse (Wei et al. 1992).
In this paper we report thé complémentation andDNA séquence analyses of a région of thé large hrp gènecluster of P. solanacearum. We also présent séquencehomologies to one of thé putative Hrp proteins, HrpO,which is encoded by this région. The homology betweenHrpO of P. solanacearum and LcrD of Yersinia pestis hasalready been shown (Gough et al. 1992), but we nowshow additional homologies and a multiple séquencealignment between ail thèse proteins. The high level ofséquence similarity which we hâve found between HrpOand a predicted Hrp protein ofX. campestris pv. vesicaîo-ria is very exciting, because it implies that Hrp proteinsof taxonomically unrelated bacteria may be functionallyconserved. Following thé findings of Atkinson et al.(1985a) that P. solanacearum can induce electrolyte leak-age from plant cells, we présent data concerning thélevels of electrolyte leakage induced by P. solanacearumfrom both tobacco and tomato leaves. For thé iîrst timewe investigate whether hrp mutants of P. solanacearumcan induce electrolyte leakage and from thé results ofthèse experiments we speculate on thé rôle played by hrpgènes in this induction.
Materials and methods
Bacterial strains and plasmids. Thèse are listed in Table 1.
Bacterial growth, antibiotic sélection and bacterial con~jugation. Techniques described by Arlat et al, (1992) wereused.
DNA purification, restriction enzyme fragment lengthanalysis, DNA ligations and transformation. Techniquesdescribed by Maniatis et al. (1982) were used, except thatplasmid DNA was extracted from Escherichia coli by thérapid boiling technique of Holmes and Quigley (1981)and from P. solanacearum by thé alkaline lysis techniqueof Birnboim and Doly (1979),
Marker-exchange of Tn5-B20 insertions from thé chro-mosome onto thé plasmid pG M1737. By conjugation usingthé helper plasmid pRK2073 (Leong et al. 1982),pGMI737 was introduced into genomic TnJ-B20 mu-tants and transconjugants were selected for tetracyclineand kanamycin résistance (TcrKmr). pGMI737 was thentransferred from thèse strains into thé P. solanacearumstrain GMI1336, again by conjugation using thé helperplasmid pRK2073, and acquisition of TnJ-B20 on théplasmid was selected for by TcrKmr, with counterselec-tion against thé donor strains provided by thé strep-tomycin and rifampicin résistances (SmrRiP) ofGMI1336. GMI1336 is an acridine orange-résistant mu-tant (Boucher et al. 1987), which carries a deletion en-compassing thé pVir2 région and is a good récipientstrain for such conjugation experiments. DNA was ex-tracted from TcrKmrSmrRifr GMI1336 transconjugantsand transformed into E. coli. TcrKmf transformants wereselected and plasmid DNA was analysed by restrictionenzyme fragment analysis.
DNA sequencing. Deletion derivatives of thé plasmidspBS2.1 and pBS2.2 were generated exactly as describedby HenikofT(1984). Selected deletions were chosen afterrestriction enzyme fragment length analysis. Single-stranded DNA was purified from thé corresponding bac-terial TG2 strains (Gibson et al. 1987), after infectionwith thé phage M13K07 (Vieira and Messing 1987) usingtechniques described by Stratagene and specifically de-signed for Bluescript vectors (DNA Sequencing Instruc-tion Manual, Stratagene). DNA sequencing reactionswere performed by thé dideoxy-termination method(Sanger et al. 1977), with 5'([35S]thio)triphosphate (Dû-Pont, NEN Research Products) as thé radioactive label(Biggin et al. 1983), using thé same Stratagene protocols.Problems arising from compressions were overcome bysubstituting 7-deazaguanosine-5'-triphosphate for dGTP(Mizusawa et al. 1986). Buffer gradient polyacrylamidegels were mostly used (Biggin et al. 1983) in thé analysisof products of thé sequencing reaction. The complètenucleotide séquence was analysed using computer pro-grains in version 7.0 of thé University of WisconsinGenetics Computer Group Séquence Analysis SoftwarePackage (Devereux et al. 1984). In particular, théséquence alignments were produced using thé programs

380
Table 1. Bacterial strains and plasmîds
Strain/Plasmid Relevant characteristics Re fe rence/ Source
Pseudomonas solanacearum
GMIiOOOGMI1495GMI1494GMII400GMI1499GMI1502GMI1501GMI1415GMI1488GMI1336
Escherichia coli
TG2
PlasmidspGMI737
pGMI737::1495pGMI737::l494pGMI737::I499pGMÏ737::1502pGMI737::1488pBluescript KS ~pBS2.1, pBS2.2
PRK2073
Wîld typeTn5-B20 derivative of GMIIOOOTn5-B20 derivative of GMIIOOOTn5-B20 derivative of GMIIOOOTn5-B20 derivative of GMIIOOOTn5-B20 derivative of GMIIOOOTn5-B20 derivative of GMIIOOOTn5-B20 derivative of GMIIOOOTn5-B20 derivative of GMIIOOOSmr Rifr Acrr derivative of GMII305
F' (traD 36,proAB, lacfi, /acZAMIS)à(lac,pro), supE, thi, recA, sri::TnlOTcr
Subclone of pVir2: thé left-hand most 13.6 kb of insert DNArecloned in pLAFR3Tn5-B20 derivative of pGMI737Tn5-B20 derivative of pGMI737Tn5-B20 derivative of pGMI737Tn5-B20 derivative of pGMI737Tn5-B20 derivative of pGMI737Sequencing vector, Apr
2 orientations of thé 8 kb Eco RI fragment of pVir2 cloned inpBluescript KS —Spr::Tn7 Ira, mob ColEl replicon
Boucher etArlat et al.Arlat et al.Arlat et al.Arlat et al.Arlat et al.Arlat et al.Arlal et al.Arlat et al.Boucher et
al. (1985)(1992)(1992)(1992)(1992)(1992)(1992)(1992)(1992)al. (1987)
Gibson étal. (1987)
Boucher et
This workThis workThis workThis workThis workStratageneThis work,
Leong et al
al. (1987)
Arlat et al. (1992)
. (1982)
PILEUP and GAP from this package and thé programFASTA and TFASTA were used to search release 20.0of thé SWISSPROT (protein) database and releases 70.0and 29.0 of thé databases of Genbank and EMBL (res-pectively), which together constitute thé nucleotideséquence library GenEmbl. The protein séquences wereanalysed for potential N-terminal signal séquences andmembrane-spanning domains were predicted using re-lease 5.0 of thé PC/Gène Program, Department of Médi-cal Biochemistry, University of Geneva, Switzerland.
Plant tests. Bacteria were assayed for their ability toproduce an HR after infiltration into tobacco leaves asdescribed by Boucher et al. (1985).
Electrolyte leakage measurements. This protocol wasadapted from Brisset and Paulin (1991). Leaves weresurface sterilised in 1.25% sodium hypochlorite .for2 min, rinsed twice in stérile distilled water and leaf dises(5 mm diameter) were eut with a cork borer. Bacteriawere grown in minimal médium (Boucher et al. 1985)overnight at 30° C, washed and resuspended in stérileMES buffer (1 mM morpholinoethane sulphonic acidadjusted to pH 6,0 with NaOH) to a final concentrationof 2x 109cfu/ml. Leaf dises were transferred into tubes,containing either 1 ml of bacterial suspension or 1 ml ofstérile MES buffer, which were then connected to a vac-'uum pump. After several seconds with thé dises wellsubmerged thé vacuum was broken thereby forcing thébacteria to infiltrate thé leaves. The leaf dises were blot-ted dry and transferred into pièces of folded gauze (six
dises each), which were introduced into plastic tubescontaining 2.5ml MES buffer and a magnetic stirrer.Dises were incubated at 25° C with continuons stirringand continuons light. The electrolyte leakage was deter-mined by measuring changes in thé conductivity of théincubation médium.
Results
Définition of complémentation unit 3
The large hrp gène cluster of P. solanacearum has beenshown to be organisée in a minimum of six transcriptionunits on thé basis of thé orientation and p-galactosidaseactivity of TnJ-B20 gène fusions in inducing conditions(Arlat et al. 1992). In particular, transcription unit 3 wasdefined as a région within which three TnJ-B20 insertionsoriented in one direction expressed high levels of p-galac-tosidase activity and two oriented in thé reverse directionexpressed thé background level of activity. This régionwas distinguished from adjacent transcription units by awild-type insertion on each side and from unit 2 by areversai of thé direction of transcription.
In order to perforai complémentation analysis of thérégion corresponding to transcription unit 3 an appro-priate plasmid was needed. Due to thé instability inP. solanacearum of thé cosmid pVir2, which carries thémajority of thé large hrp gène cluster (Boucher et al.1987), a subclone was chosen with which stable merodi-ploid strains could be produced. This plasmid, pGMI737

PGM1737
381
R R
Compiernentingplasmids
pGMI737pGMI737::1495pGM!737::1494pGMl737;:1499pGM!737::1502pGMI737::1488
Fig. 1. Complémentation analysis of transcription unit 3 of thé largehrp gène cluster of Pseudomonas solanacearum. The EcoRl (R)restriction enzyme map of thé whole cluster is represented and ailsix putative transcription units as defined by Arlat et al. (1992) areindicated by arrows. The DNA insert carried by thé plasmidpGMI737 is represented by thé shaded bar. The positions of théTn5-B20 insertions used for complémentation analysis are indicatedand thé insertions are identified by numbers according to Arlat etal. (1992). The two insertion numbers which are boxed (1491 and
5 kb
1415), correspond to insertion mutants with a wild-typephenotype;thèse were not used in thé complémentation analysis. The combina-tions of mutants and complementing plasmids used in thé analysisare shown below thé map. Plus ( + ) and minus ( — ) symbols indicatecomplémentation and lack of complémentation, respectively. Thecombination that sometimes gave complémentation and sometimesdid not is indicated by both a plus and a minus sign ( + / — ). Theresponses of thé combinations of mutants and plasmids whichenabled complémentation unit 3 to be defined are boxed
(Boucher et al. 1987), which has as ils insert DNA théleft-hand most 13.7kb of thé P. solanacearum DNAinsert of pVir2, should contain transcription unit 3 in itsentirety (Fig. 1). Relevant Tn5-B20 insertion derivativesof pGMI737 were constructed by marker exchange fromthé corresponding genomic mutants, (only those mutantshaving an Hrp" phenotype were used). ThèsepGMI737 : :Tn5-B20 plasmids were introduced into vari-ous genomic mutants of P. solanacearum by conjugationand thé abilities of thé resulting merodiploid strains toproduce an HR were determined after infiltration intotobacco leaves.
The results shown in Fig. 1. demonstrate that a régionof approximately 4500 bp between insertions 1494 and1488 defined a single complémentation unit, with 1494and 1495 representing thé start of a separate complé-mentation unit to thé left and 1488 being thé first inser-tion in a complémentation unit on thé right-hand side.The mutant strain GMI1488 (thé genomic mutant ofinsertion 1488) was not complemented by any of théplasmids, even by thé plasmid pGMI737 itself withoutinsertion. When thé insertion 1488 was carried on aplasmid, however, this was able to complément ail thégenomic mutants tested except GMI1488. This findingindicates that this insertion defines a new complémenta-tion unit extending into thé région which is not represen-ted on pGMI737, beyond thé £coRI site just to thé rightof insertion 1488. The limits of thé complémentation unitthat we hâve described hère can be more precisely definedby thé wild-type insertions 1491 and 1415. It thereforecorresponds exactly to transcription unit 3 (Arlat et al.1992) and has been designated complémentation unit 3.
The three insertions (1400, 1499 and 1502) that mapwithin complémentation unit 3 were generally unable tocomplément each other. However, inoculation ofGMI1502/pGMI737: :1499 did sometimes resuit in thé
appearance of an HR and therefore in complémentation.In thèse cases thé HR was not always apparent after 24 h,but it was always clear within 48 h. This complémenta-tion was only seen when insertion 1499 was carried ona plasmid; thé alternative combination of insertions 1499and 1502, (GMI1499/pGMI737: :1502), did not resuit incomplémentation. The effect of copy number maytherefore be important.
Nucleotide séquence of complémentation unit 3
The nucleotide séquence of thé 4772 bp DNA régionbetween insertions 1494 and 1488 was determined and isshown in Fig. 2. The predicted amino acid séquences ofthree complète open reading frames (ORFs) are in-dicated. The corresponding gènes hâve been designatedhrpN, hrpO and hpaP. The high G+C content for thisrégion (69.3%) is reflected in thé codon usage of thé threeORFs, in thé same way that bacterial gènes usually hâvea codon usage characteristic of their overall base com-position (Bibb et al. 1984). In fact, as a resuit of a highpréférence for G or C in thé third codon position, andto a lesser degree in thé first position, there is a nonran-dom use of codons, with seven not being used at ail andeight only being used once among ail three ORFs. Sincethis is completely normal for bacteria with a high G + Ccontent (Bibb et al. 1984), thé same bias was not foundin other ORFs, and since we used a codon usage tableprepared from published séquence data of P. solanacea-rum (Akiyoshi et al. 1989), thé prédictions we hâve madefor coding régions are probably reliable. Each gène isappropriately preceded by a potential ribosome-bindingsite (Shine and Dalgarno 1974), usually at a distance thatis optimal for translation initiation (Curry and Tomich

382
101 GTCGAGCTCCTTGAAGTTCGGACGCAGCCCGCGCAGCGCTACCAGCATCGCCTGGGCGTTTTCGAACTGGCCGCTTCGCGTGGCATACATCAGCACTTCG 200
i rzrn^irT"^ "n nrrr zzirirtit^501 CGCGGCCGATAGCAGGGGCCAGATCATGTCGGACGAAAAGACCGAACAGCCCACAGACAAGAAACTCGAGGATCGGCACCGGGACGGCGAGACGGGCAAG 600
hrpN M S D E K T E Q P T D K K L E D A H R D G E T A K
601 AGCGCAGACCTGACCGCGGCGGCGGTGCTGCTCTCCGGCTGCCTGCTGCTGGCACTGACCGCCTCCGTCTTCGGCGAGCGATGGCGGGCCCTGCTCGACC 700
S A D L T A A A V L L S G C L L L A L T A S V F G E R W R A L L D L
701 TCGCGCTGGATGTCGACAGCAGCCGGCACCCGTTGA'PGACGCTCAAGCAGACGATCAGCCATTTCGCCCTGCAGCTCGTGCTGATGACGCTGCCGGTGGG 800
A L D V D S S R H P L M T L K Q T I S H F A L Q L V L M T L P V G
900901 CTTCGTCTTCG
F V F A L V A W I A T W A Q T G V V L S F K P V E L K M S A I N F
TGGCGGCGGCGGTCTGGAAGCTCA 100:
A S G L K R I P S V R S M I D L V K M I I K G V A V A A A V W K
1001 TCCTGATCCTGATGCCGTCCATCGTCGGCGCCGCCTATCAGTCCGTCATGGACATCGCCGAGATCGGCATGACGCTGCTGGTCCGGCTGCTGGCGGCGGG 1100
L I L M P S I V G A A Y Q S V M D I A E I G M T L L V R L L A A G
GAAGTCAAGCGCGAG 1200
G G L F L I L G A A D F G I Q R W L F I R D H R M S K D E V K R E
H K N S E G D P H I K G E R K K L A R E L A D E A K P K Q S V A G A
Q A V V V N P T H Y A V A I R Y A P E E Y G L P R I I A K G V D D
E A L A L R E E A A A L G I P I V G N P P L A R S L Y R V D L V G
P V P E P L F E T V A E V L A W V G E M G A S G T P G A E P Q H
1601 CCTGACCGGACCCTGACATGGCCAAAAAGAATGCAATCCAGGATTTCAGCGGCGAAATCGGTATCGCCGCGCTAGTGGTGGCGGTGGTGGCGCTGATGGT 17QO
hrpO M A K K N A I Q D F S G E I G I A A L V V A V V A L M V
rACATTCCGTCGGCGACC 1800
L P L P T M L I D A L L G L N : T L S V V L L H V T M Y I P S A T
S L S A F P S L L L F T T L L R L S L N I A S T K S I L L H A O ^
H I I E S F G K L V V G G N L V V G L V V F L J I T T V Q F I V I
2001 CGCCAAGGGTT
A K G S E R V A E V G A R F T L D A M P G K Q M S I D A D L R A G
H L S P E E A R K R R A L L A M E S Q L H G G M D G A M K F V K G D
A I A G L V I T L V N I L A G I V I G I T Y H N M T A G E A A N R
2301 CTTCGCGGTGCTGTCGATCGGCGACGC
F A V L S I G D A M V S Q I P S L L I S V A A G V M I T R V S D E
2500
E Q A H K Q S S L G M E I V R Q L S T S A R A M F T A S A L I , H G F
2501 TCGCGCTGGTGCCGGGCTTTCCGTCGTTCCTCTTCGTCGCGCTGGCGACGCTGATCT*I'CGTGTTCGGCTACACGCTGCGCAACCGCGCCAAGGAGGGCGA 2600
A L V P G F P S F L F V A L A T L I F V F G Y T L R N R A K E G DFig. 2. (for continuation see page 383)

383
2601 CGGCGACGAAGGCGACGCCCTGCCCGCGCTGCTGCGCGAGGGCTCGAAGGGCAAGGCGCCGACCATCGCCGAACAGGCCCCGTCGTTTACCGTGCCGGTG 2 ? f - 0
G D S G D A L P A L L R E G S K G K A P T I A E Q A P S F T V P V
27Q1 GGCGTGCGGCTCGGCGCGGAGCTGGCCAAGGGCCTGGACGTGCCCGCCCTCGATACCGCCTTCCAGCAGGGCCGCCACGCGC1X5GCCGAAGCGCTCGGCC 2800
G V R L G A E L A K G L D V P A L D T A F Q Q G R H A L A E A L G L
2801 TGCCCTTCCCCGGCATCGCGATCTGGA/^GGCCGACGCCCTGCAGCCCGACAGCTACGAGGTGCGCGTGCACGATATCCCGGGCC^AGrrn'-TCGCGGTGCC 2 9 0 0
L A I W K A D A L C D S Y E V R V H D I P G E P V A v P
XÎCACTGGATCGCG 3000
D G H L L . I P D L P E A L A Q A V E A A G L P N H P A P H W I A
P A H V A Q D A A L S A T G Q R V E R V I A D H V V H V L R R S A H
L F V G L Q E T Q W M L E R V T T D Y P G L V A E A Q K A V P A Q
3201
R î A D V L R R L E E Q V P I R N M R A I E S : , V V W G P K E
K D T L M L V E Y V R G D L G R Q I A H Q A T G G T R Q M P A I L I
3401 TGGACCTGTCGGTCGAGCAGACCGTGCGCCAGGCCATCAAGCCGACGCCGGCGGGCAACTTCCTGACGCTCGATCCGCAGCAGGTGGAGGCCATCATCAT 3500
D L S V E Q T V R Q A I K P T P A G N F L T L D P Q Q V E A I I M
R L R G I M Q G N P V E T P S A L A I V T S M D I R R Y V R R M I
GCGGCGGCC 3700
hpâPE P H L Q A L N V Y S F Q E L G G Y V D L R P V G K L V L
GGCCCGCATGACGCGC 3800
M T Q A V R L W R P Q P V D P A P P ^ R R R T P L R T P G P H D A L
3801 TGCCCTATGTGCCGCCGCCCCGCCCCGAGCCCCCGGAGCCCGTCGAGGACGACTTCCCGGAGCCCCGGCAGGACACCCCGGCCAGCGATCCGCCCCCCGA 3900
P Y V P P P R P E P P E P V E D D F ' P E P R Q D T P A S D P P P E
T3901 GGCGCGCACCGACGCCAGCCCACCGCAGCCCGCCGCCGACCCCGATGCCTCGCTCCACACCGCGGCCATCGCCAGCCGCCTGGTGCGGACCTGCTCGGAG 4000
À R T D A S P P Ç P A A D P D A S L H T A A I A S R L V R T C S E
4001 ATCGGCGCGGGCGAGACCATGACCGAGCACCTTGCCCGGCTGCTCTCCCAGTTCTGTGCCTCGCAGGCGATCCGCTCCGGCGGCGAATGCTGGGAAATCT 4100
I G A G E T M T E H L A R L L S Q F C A S Q A I R S G G E C W E I S
4101 CACTCGATCTTGATCCCAÀGATCCTGCCCGAAACCCGCCTCACGCTGCGGCTTTCGCCGCACACGCTATCGCTTCGCTTCGAGGCCGGACACCCTCGqTC 4 2 0 0
L D L D P K I L P E T R L T L R L S P H T L S L R F E A G H P R S
\ 4300
4301 CACCCTCCGACAT^AACGCCGGGCCTTCTTTCTC'TCCGCTGGAACCCCACCTGCGCGCCTTCACGCCGGCACACGCCGCGCTTACGCGCCTGCTGGACGG 4 4 0 0
4701 CCCCGTCCTGCCGCTGCAGATCGCCAGCGCCGCCCACGCATGCCGCTGCGACATCCACGCGCTGGACTGGCA ^ 7 7 2
Fig. 2. Nucleotide séquence and deduced amino acid séquence of thé ribosome-binding séquences are underlined. The séquence withrégion designated complémentation unit 3. The séquence is given homologyto thé motif found upstream ofseveral avirulence and hrpbetween Tn5-B20 insertions 1494 and 1488; thé position of insertion gènes is underlined and given in bold lettering. The positions of1494 is immediately before nucleotide no. 1 and that of insertion Tn5-B20 insertions are indicated by arowheads; 1491 between nu-1488 is immediately after nucleotide no. 4772. The three open cleotides 513 and 514; 1492 between nucleotides 845 and 846; 1499reading frames (ORFs) corresponding to hrpN, hrpO and hpaP are between nucleotides 1499 and 1500; 1502 between nucleotides 2341shown with their deduced amino acid séquences specified in one- and 2342; 1501 between nucleotides 3610 and 3611; and 1415letter code below thé first nucleotide of each codon. Potential between nucleotides 3992 and 3993

384
1988), providing strong évidence that each gène beginswith its assigned start codon.
The direction of transcription of thèse gènes is thésame as that anticipated on thé basis of Tn5-B20 orienta-tion and p-galactosidase activity (Arlat et al. 1992) andno ORFs with high coding probabilities were predictedfor thé opposite direction. A partial ORF was found toprécède hrpN, but oriented in thé opposite direction andanother that starts after hpaP in thé same direction ashpaP (data not shown). Thèse presumably correspond tothé first ORFs in transcription units 2 and 4, respectively.No séquences located 5' of hrpN and showing homologyto thé consensus E. coli or Pseudomonas promoterséquences were found (Hawley and McClure 1983; De-retic et al, 1989). However, an 11 bp région (TGGAAC-CATAG) shows good homology to a motif found 5' ofseveral avirulence gènes (Jenner et al. 1991) and hrpgènes, thé so-called "harp box" (Fellay et al. 1991;Fig. 2). There is no région of dyad symmetry 3' of hpaP,but there is a région of five thymine residues (TTCTTTat positions 4326-4331 ; Fig. 2), indicating that this maybe thé site ôf transcription termination (Platt 1986). Theconsensus séquences CGGGC/G and TCTG found up-stream and downstream, respectively, of many prokary-otic RNA endpoints are also présent, located on eitherside of thé group of thymine residues.
Positioning of Tn5-B20 insertions
The insertion sites of relevant Tn5-B20 insertions weredetermined using appropriate insertion derivatives of thérégion corresponding to complémentation unit 3 sub-cloned in Bluescript vectors and oligonucleotide primersmade complementary to thé nucleotide séquence at bothextrême ends of thé transposon Tn5-B20. The positionsare shown on thé nucleotide séquence in Fig. 2. Theorientations of thèse insertions were confirmed bysequencing: insertions 1491, 1492, 1499 and 1501 (whichresuit in significant p-galactosidase activity) are ailoriented so that transcription of thé lacZ codingséquence is in thé same direction as that of hrpN, hrpOand hpaP; insertions 1502 and 1415 (which resuit inbackground levels of p-galactosidase activity) are orient-ed in thé reverse direction. Insertion 1494 was found tobe oriented in thé opposite direction to thé transcriptionof hrpN, hrpO and hpaP and insertion 1488 in thé samedirection as that of hrpN, hrpO and hpaP. Both of thèseinsertions resuit in significant p-galactosidase activity.
Of thé two Tn5-B20 insertions in this région that giverise to wild-type phenotypes in plant tests, insertion 1491is located exactly within thé proposed ribosome-bindingsite of hrpN. In order to explain how HrpN is expressedin GMI1491 despite thé fact that thé predicted ribosome-binding site is interrupted, we deduced thé DNAséquence created at thé site of insertion of 1491. Thisrevealed that a potential ribosome-binding site (AG-.AGGG) is reconstituted exactly at thé site of insertion,which together with thé présence in thé transposon ofséquences recognised as a promoter by P. solanacearumcould explain thé wild-type phenotype of GMI1491. This
mutant produces a level of p-galactosidase activity whichis similar to that of GMI 1492 in inducing conditions, andthé expression of which is regulated in thé same way asthat of most other insertion mutants with respect to invitro culture conditions (Arlat et al. 1992). Therefore,while thé wild-type phenotype of GMI1491 may be dueto a nonpolar effect of thé transposon, thé expression ofp-galactosidase activity of this mutant seems to be underthé control of an hrp promoter. The second wild-typeinsertion 1415, is positioned approximately in thé middleof hpaP. The désignation of hpaP as an hrp gène istherefore strictly speaking incorrect. It is possible that théproduct of this gène is involved in interactions betweenP. solanaceraum and plants other than tomato and to-bacco, in which case thé nomenclature hrp could be used.However, we hâve décidée to avoid confusion by usingthé new three-letter prefix hpa (for hrp associated), hencehpaP (and HpaP for thé protein), to indicate that thisgène may not be a genuine hrp gène.
Characterislics of thé predicted proteins
By computer analysis, prédictions were made for théthree complète predicted protein séquences, HrpN,HrpO and HpaP, with respect to their molecular weights,hydrophobicity, présence of N-terminal signal séquences,other particular consensus séquences and homologieswith other known proteins. None of thé proteins ispredicted to hâve an N-terminal signal séquence and nohomologies were found between HrpN or HpaP andknown proteins in thé databases.
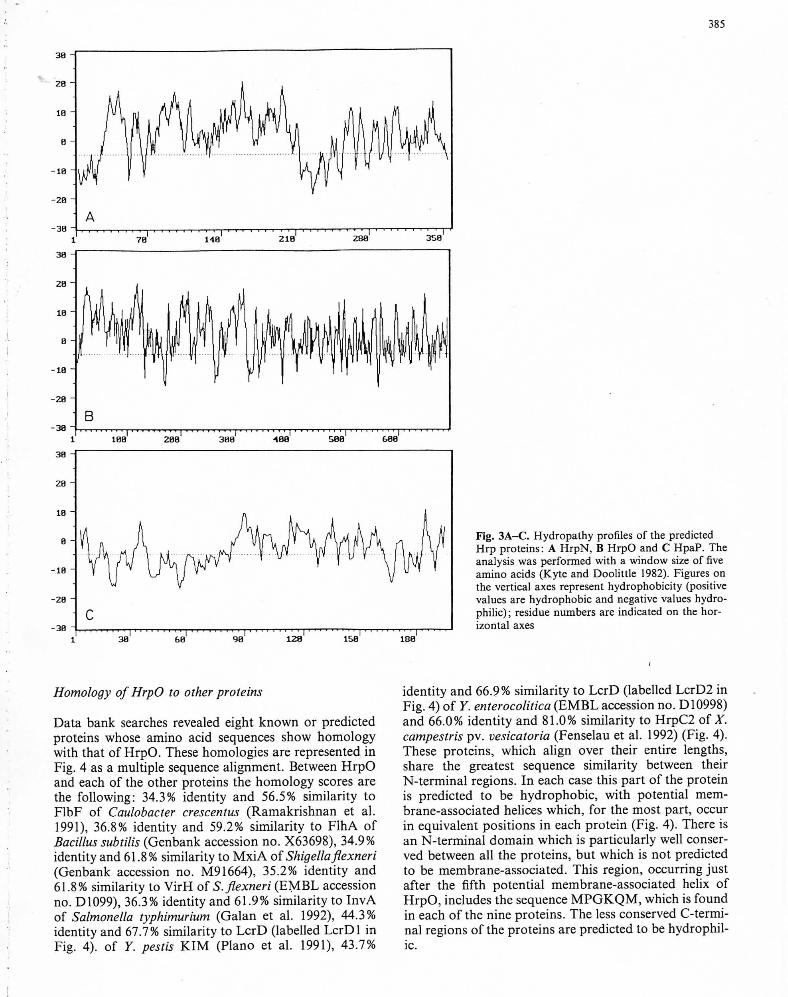
(1) HrpN is predicted to be a protein of 357 aminoacids and molecular weight (Mr) 38 607. HrpN is predict-ed to be a hydrophobic protein (Fig. 3A); there are fivepotential membrane-spanning domains, four in théN-terminal and central parts of thé protein and one atthé C-terminus.
(2) The nucleotide séquence ofhrpO predicts a proteinof 690 amino acids and Mr 73 990. The N-terminal andcentral parts of HrpO are very hydrophobic (Fig. 3B)with nine potential membrane-spanning domains. Thereare positively charged amino acids between thèse do-mains, which probably contribute to thé déterminationof thé orientation of thé membrane-spanning domainswithin thé membrane (von Heijne 1986). The C-terminalpart of thé protein is hydrophilic.
(3) The predicted protein HpaP bas 197 amino acids,Mr 21 959 and no hydrophobic domains (Fig. 3C). It isunusually rich in proline residues, with a total of 30representing 15% of thé total number of amino acids.This high frequency of proline may hâve important ef-fects on thé tertiary structure of thé protein. For exam-ple, proline is often used to produce a bend in a proteinchain and pro!ine-rich proteins often hâve structuralfunctions (Zubay 1983), since thé cyclic structure of thisamino acid is very rigid and proline residues are unableto form intrachain hydrogen bonds.
385
28 -
18 -
8 -1
-18 -
-29 -
-38 H
28 -
-18 -
-28 -
-38 -1
38 -
28 -
18 -
8 -
-18 -
-28 -
-38 -1
A
B
268 388 580
Fig. 3A-C. Hydropathy profiles of thé predictedHrp proteins: A HrpN, B HrpO and C HpaP. Theanalysis was perforraed with a window size of fiveamino acids (Kyte and Doolittle 1982). Figures onthé vertical axes represent hydrophobicity (positivevalues are hydrophobic and négative values hydro-philic); resîdue numbers are indicated on thé hor-izontal axes
128 ise
Homology of HrpO to other proteins
Data bank searches revealed eight known or predictedproteins whose amino acid séquences show homologywith that of HrpO. Thèse homologies are represented inFig. 4 as a multiple séquence alignment. Between HrpOand each of thé other proteins thé homology scores arethé following: 34.3% identity and 56.5% similarity toFlbF of Caulobacîer crescenîus (Ramakrishnan et al.1991), 36.8% identity and 59.2% similarity to FlhA ofBacillus subtilis (Genbank accession no. X63698), 34.9%identity and 61.8% similarity to MxiA oîShigellaflexneri(Genbank accession no. M91664), 35.2% identity and61.8% similarity to VirH of S.flexneri (EMBL accessionno. D1099), 36.3% identity and 61.9% similarity to InvAof Salmonella lyphimuriwn (Galan et al. 1992), 44.3%identity and 67.7% similarity to LcrD (labelled LcrDl inFig. 4). of Y. pestis KIM (Piano et al. 1991), 43.7%
identity and 66.9% similarity to LcrD (labelled LcrD2 inFig. 4) of Y. enterocolitica (EMBL accession no. D10998)and 66.0% identity and 81.0% similarity to HrpC2 of A'.campestris pv. vesicaloria (Fenselau et al. 1992) (Fig. 4).Thèse proteins, which align over their entire lengths,share thé greatest séquence similarity between theirN-terminal régions. In each case this part of thé proteinis predicted to be hydrophobic, with potential mem-brane-associated hélices which, for thé most part, occurin équivalent positions in each protein (Fig. 4). There isan N-terminal domain which is particularly weîl conser-ved between ail thé proteins, but which is not predictedto be membrane-associated. This région, occurring justafter thé fifth potential membrane-associated hélix ofHrpO, includes thé séquence MPGKQM, which is foundin each of thé nine proteins. The less conserved C-termi-nal régions of thé proteins are predicted to be hydrophil-ic.

386
• l 2 *i . . . . -. 1Q5
FlbF madaaaDnassmDsakslIdQlmraFjnQ'a ' :avVaII_vLLIiPvPapL iDvLja isl_tasVl iLMt a i l IkkpLeFt sFPtvLLvTTLFRLqLji la s c R l l L s h gFlhA .•nstrDlsvlisvV nVaivViP^PpwLlsiLj- i iN ' is la l ivLLt tmnmaeaLaFS JFPs lLLUTLFRLgLnVst tRsILshaMxiA mVmlIaHLliPLPtyLvDELiglNivlallvFMasfYIer JLsFStFPsvLLiTTLFRLaLsIstsRllLvdAVirH miqs f IkqvsckpE l l i l y lmVml IaMLI iPLP tvLvD f LialNivlaÛyFMgsEYIerÀI^gZStFPsvL^iTT^FRLaLsIstsRl ILvdAInvA mlls l lnsar l rpEl l ilxJrriVml IsMJ^/iPLJ'tvLvDf LialNj,vlaUvFMasf Yîdr iLsFSLFPayLLiTTLj^RLa^sIscsRlILieALcrDl mnphdlewlnriqe.rkDimJ^ylIlaVVfMMVlPLPplvl^^LcrD2 mnphdlewlnrige.rkPim lavjl laWfHMVlPLPplylDi^iavNRit isVvlLMiaiYJnspLQFSaFPavLLvTTLFRL^LgYs t C R m I L l a AHrpC2 mladrvratryfaysqEva iaaJvVaVIaLMIlPLPcpMiDtLlqiNit IsVvlI^yCmYVpdsisL^aF^slL^fTTj^RLgLDla stksjLlhAHrpO makkna iad f sgEia ia_alvVaVVaLM'/IPLPcniLiD3Llg.INi^_;5Vyj.LMvtfnYlpsaCs^SaF_PslLLfTT[ JLRLpLnIa stkslLlhAcons _' P _ P L F P - - L L - - T L - R L - L IL---
106FlbFFlhAMxiAVirHInvALcrDlLcrD2HrpC2HrpOcons
_ 210
E ..... qGkVVetFGnFVVGqjwJ.YGlVWilAilIOFUA/ITKGaERVsEVaARFtLDaMPGKQMSIDADLnAGmlteqeAkhRRekvarEadFYGamDGAsKFVD ..... rGkII t tFGQFVIGdsLaVGfVIFsIVTVVQ_F_iVXTKGSËRVAEVaARFsLDqHPGKQM5IDADLkAGiIdaagAkeRRsilerESQLYGsfDGAMKFID. ,.. . .AGklI^tFGQFVIGQsLaV^fVIFsIVTV'/OFj.VITKGSERVAEVaARFsLDqHPGKQMSIDADLkAGildaaqAkeRRsilerESQLYGsËDGAHKPID. ^.. . .&GeIlAt:FGQFVIGdsL3YSfWFsIVTWOFiVITKGSERVAEVaARFsLDqMPGKQMSIDADLkAGiIdadaAreRRsvlerESQLYG5fCGAMKFID_. ^^^.^^V^^FGnfVVGqnLi^GjVJJlIITIVOF^VITKGSERVAEVsARFsLDaHPGKOMSIDqDMrAGvIdvneAreRRatiekESQMFGsmDGAMKFVD. . „ . AGalVvtFGngWGq^I^iVgiVIFlI ITIVOFlVITKGSERVAEVsARFsLDaMPGKQMSIDgDMrAGvIdvneAreRRat iekESQMFGsmDGAMKFVE. . „ .AGhIIegFGeLWGqr)LvVGl\/FIIITrVOFi lVIdKG5ERVAEVQARFtLDaMPGKQMSIDADLrqGnltadeArrkRarlamE5QLhGqmDGAMKFVD. . L. . AGhlIejFGkLWGCLrjLvVGlVVFlIITtVOFiVIaKGSERVAEVaARFCLDaHPGKQHSIDADLrAGhlspeeArkRRallamESQLhGqmDGAMKFV------- G ---- -FG ---------- G-- -F- I ----- F- V--KG- -R- -EV-ARF-LD-MPGKQM- ID-D- -- G ------ A - - - R ----- E ---- G- -DGA-KF-
21l1
FlbF KGDAJAGUltalNiiqGIilgYyqhkMpfqdAasCYtlMTIGDGLVSOIPALilsJaAGfnvVskaavEgFlhA KGDAIAGilIvHINii^GIvIGmlqaGHsiaeAashFtmLTVGDGiVSOIPALLlBtatGivVTRaasEaMxiA KGDAIAGilIiFVNligGIsVGrasqhGHslsaAlstYCILTIGDGLVSOIPALLISlsAGfmlTRVnqDsd. . . .Vi rH KGDAIAGi l I iFVNl iaGIsVGmsQhGMslsaAls tYCILTIGQGLVSOIPALLIs I sAGf iVTRVnaDsd . . . .InvA K G D A I A G i l I i F V N f i a G I s V G m t r h G H d l s s A l s t Y t m L T I G S G L V a O I P A L L I a l s A G f i V T R V n a D t d . . . .LcrDl KGDAIAGlI I iFVNi laGVtrGvtQkGLaaaeAla lYsILTVGDGMVSOVPALLIaTtAQi iVTRVssEds . . . .LcrD2 KGDAIAGlII iFVNilnGVIi IGvCQkGLaaaeAla lYsILTVGDGMVSOVPALLIalCAGiiVTRVssEds . . . .HrpC2 KGDAIAGlVIcKVNllaGIvVGvCvhGHtaadAanrFalLsVGDaMVSOIasLLIsVaAGvïïiITRVanEnet.riHrpO KGDAlAG]j/]:t:Ly^j,l3GIvIGitvhnMtaqeAanrFaVLsIGOaMVSOIPsLLIsVaAGvmITRVsdEeqahkqcons KGDAIAG-- I N G G A GD--V-Q L - I G
315. sadkalCtOlamnPvQLqmvsass oilal^.nLGhdltqOlEavPklLwaaat: imllq^i.nMGrnlmsOifanPfvLivCsa LalaioM.nHGrn lmsOi fanPfvLiv t saLa la igM. nMGrn lmtOI lnnPfvLwta iL t ismacsdLGsdlakCvvaaPkaMliQavLlllfqLsdLGsdlqkOvvaaPkaMliqavLll l fqLssLGldlgrOltsnaraLmaasvLlacfaFssI^GmelvrQlstsaraMf tasaLlmglâli
316 420FlbF iPGmPifpFaaMAlaaaalaYkrvqdakkpkaldpadleaaapse.DeeeDisaslaiddvkielqyqllt lindlDgrkLtdqira iRktLFlhA f tpigilltgpLAqllafgaYtilsksakekeevdeileeeaevdelksDesyyglIhidoj^gf ef aval ip ladanqggdLldrivmiRrqLMxiA IPGFPf JLvFLUAvtltalfYvkkwekekslsesd ssgycgt fdidnthdsslamienldrissecvPlill faenkinanDmegL , . . ieriRsqFVirH IPGFPfivFfllAvtltalf Yvkkwekekslsesd ssgvtqt fdidnshdsslamienldaisser vPl^1 If aenkinanDmeqL. . . ieriRsqFInvA IPGFPlDVFviLsvvlsvlfYf . kEreakrsaakpk tskgegpls ieekeass^ql iqdldkvstetvPljJ IvpksrredlEkagL . . .aerlRsqFLcrDl iPGFPEvtFlLLAllvacqgYmlsrkasrndeanqd.IqsiltsgsgapaartkaktsgankgrlgeqeafamtvPllidvdssqqealEanaLndelvrvRraLLcrD2 jPGFPt_v:tFliLAllvqcgqYmlsrkgsrndeanQd . Iqsilcsgsgapaartkaktsgankgrlgeqea famtvPllidvdssqqealEaiaLndelvrvRraLHrpC2 vPG^FPav^Fll^AaavgaQqYtiwrkardisgcdqrklpsasrkaakgeaDhir knapdfasPlsmrlspqlaallDparLdqaieseRrq LHrpO vEGJPsf^jVa^jAt 1 i fvf gYc Irnrakegdgdegda ) pal Iregskgkapt ia eqapsftvpvgvrlgaelakglCrvpaLdtafqqgRha^cons Y L R
421 525FlbF asEyGÈvMPpVrildnmrLanqgyairlkErneagagevrIgclmcmDprggqvelpgehvrepaf..glpatWIaddlrEeat frgytvvdpacvltthlteiLkFlhA alElGlviPvVriRdniaLqpneyrlklkgnevakgell Idhy lamsptpeddl iegietvepsf . .glpakWIseavkDeadmlgyCwdpasvvsthitekikMxiA t iDyGvrLPtllyRCsneLkvddivllInEVradsEniyfdkvcitDengdidalglpwstsy . . nervisWVdvsytEnltnidakiksaqdefyhqlsqaLlVirH f iDyGvrLPCIlyRtsneLkvddivllInEVradsÊniyfdkvcitDengdidalglpwstsy . . nervisWVdvsyt EnltnidakiksaqdefyhqlsqaLlInvA f iDyGvrLPeVllRdgegLddnsivllInEIrveqf tvy Edlmrvvnysdewsfglnptihqq. .gssqy EWVtheegEklrelgyvlrnaldelyhclavtvaLcrDl yIDlGvpFPglhlRfnegMgegeyiislqEVpvargelkagy1IvrEsvsqlelIgipyekgehlIpdqeafWVsveyeErleksqleffshsqvltwhlshvLrLcrD2 yIDlGvpFPglhlRfnegMgegey lis IqEVpvargelkagy1IvrEsvsqlel1gIpyekgehlIpdqetfWVsveyeErleksqleffshsqvltwhlshvLrHrpC2 Ye_llGl_DFPglaiWflj;esLqgmcjvevlIhDVpet:raelentd. . . . Dmqaalarqal. . . sp LhHrpO aea 1G Ipp'i'q la iWkadaLqpdsvevrVhDIpqepvavpdghl 1 ipDlpealraqaV . . .eaaglpnhpaphWIapahvaqdaalsatgqrverviadhvuhvLrcons G P
« » * *526 630
PlbF enmaDLlsyaEvqkLLkeLpetqkkLvdDlipgcvtattVqrVLqsLLrErVSIRdLpqlLEqvqeaaphtasvtqLvEqVRarLaRqlcwanrgddq a_LpiI_tL_PlhA qhahELiGrQETkqLiDhLkesYpvLveEVtpnplsvgdlqkVLakLLkEkVSIRNLvtlFEtLadYgkltcDsdlLCEytRqaLakqltaqfakenevLkvVtcMxiA nninEifGiQETknMLDqFenrYpdLlkEVfr.hvtiQRIsEVLqRLLgEnlSVRNLkllMEsLalWaprEKDvitLvEhVRasLsRylcskiavsge.ikvVMLVirH nninEifGiQETknHLDqFenrYpdLlkEVEr.hvtiQRTsEVLqRLLgEnlSVRNLkllMEsLalWaprEKDvitLvEhVRasLsRylcskiavsge,ikvVMLInvA rnvnEYfGiQETkhHLDqLeakFpdLlkEVlr.haCvQRIsEVLqRLLsErVSVRNMkllHEaLalWaprEKDvinLvEhIRgaMaRyIchk£angge,LraVMvLcrDl eyaeDFiGiQETryLLEqMeggYgeLikEVqr.ivplQRmtEILqRLvgEdISIRNMrsILEaMveWgqkEKDvvqLtEylRssLkRylcykyangnniLpayLFLcrD2 eyaeDFiGiQETryLLEqMeggYgeLikEVqr.ivplQRmtEILqRLvgEdlSIRNHrsILEaMveWgqkEKDvvqLtEylRssLkRyIcykyangnniLpayLFHrpC2 arahlFvGiQETqwMLEqvavdYpgLvaEVnk.ampaQRIaDVLrRLLeErlpVRNiksILEsLvvWgpkEKDllmLtEyVRcdLgRylahtatagtgqLpaVMLHrpO rsahlFvGlQETqwMLErvttdYpgLvaEaqk.avpaQRIaDVLrRLLeEqVpIRNMralLEstv-vWgpkEKDt ImLvEyVRgdLgRqlahqatggtrqHpaILLCons E L L--L--E R I-E L-E--R
631 725FlbF sadwEaafaesil iqpgddkqLaLpPsrlqdf irgvrdsferaalageapv . 1 ItSpgVRpYVrsilErErgqt WmSqnEihprarlkCvgmV. .FlhA sgrVEkaladgVqqTehGnYLsLEPdisesivrsvakeaeqlslrqetai . HcSppVRmYVkqLlEryFpdlpVlSYnEleanvevqs igvVdiMxiA sgylEdalRkglrqTsgGsFLnMDievsdevmetlahalrel.rnakknfvllvSvDIRRFVkrLIDnrFksiIViSYaEideaycinvlktI..VirH sgylEdalRkglrqTsgGsFLnMDievsdevmetlahalrel.rnakknfvllvSvDIRRFVkrLIDnrFksiIViSYaEideaytinvlktI..InvA saeVEdvlRkglrqTsgsCFLsLDPeasanlmdlitlklddl.UahkdlvlltSvDVRRFIkkMIEgrFpdleVlSFgEiadsksvnviktI..LcrDl dqeVEeklRsgVrqTsaGsYLaLEPavteslleqvrktigdlsqiqskpvliv.SmDIRRYVrkLIEseYyglpVlSYqEltqqiniqplgrlclLcrD2 dqeVEeklRsgVrqTsaGsYLaLDPavteslleqvrktigdlsqiqskpvliv.SmDIRRYVrkLIEseYyglpVlSYqEltqqiniqplgrVclHrpC2 dhaVEqlIRqsIraTaaGnFLaLpPeqanqlveqverivgdhaqh. . .plawaSmDVRRYVrrMIEarLtwlqVySFqElgsevqlqpigrVwHrpO aisVEqtVRqalkpTpaGnFLtLDPqqveaiirnrlrgimqgnpvetpsalaivtSmDIRRYVrrMIEphLqalnVySFqElggyvdlrpvgklvl- = -s -----E L S___ R V-S--E
Fig. 4. (for legend see page 387)

387
AU of thé proteins homologous to HrpO are im-plicated in sécrétion, except for LcrD and FlbF, whichhâve been reportée to hâve regulatory rôles, and HrpC2,which is thé predicted protein of unknown function of anhrp gène, hrpC2, of X. c. pv. vesicatoria (Fenselau et al.1992). The MxiA and VirH proteins, encoded by thé spalocus of S.flexneri, are involved in thé sécrétion of plas-mid-encoded invasion antigens, tcrmed [pa proteins,which are essential invasion déterminants (Venkatesan etal. 1992; Galan et al. 1992). FlhA is thought to be in-volved in thé export of flagellar proteins, which occursthrough thé nascent structure of thé flagellum by aflagellum-specific pathway (Vogler et al. 1991). InvA isessential for thé invasion phenotype of S. typhimuriumand may be involved in thé translocation of InvE (Galanet al. 1992). LcrD is thought to be a positive regulatorof thé Yersinia Iow-Ca2+ response (Lcr), which is man-ifested in vitro by thé requirement for millimolar quan-tities of Ca2 + for maximal growth yield at températuresabove 34° C and by thé température- and Ca24-regulatedexpression of virulence-associated proteins, Yops, andthé V antigen (Cornelis et al. 1989). An IcrD mutant doesnot undergo growth restriction after a température shiftfrom 26 to 37° C in thé présence or absence of Ca2+ andis unable to induce strong expression of thé V antigen orYops (Goguen et al. 1984; Piano et al. 1991). FlbF isencoded by a cell cycle-regulated flagellar gène, flbF ofC. crescentus, and is required for transcription fromsigma-54 promoters oîfla gènes expressed late in thé cellcycle (Newton et al. 1989). FlbF is hypothesised to func-tion as a signal transducer to regulate flagellum biosyn-thesis and other steps required for polar morphogenesis,by mediating thé response to changes in thé periplasmicspace or membrane (Ramakrishnan et al. 1991).
A model of LcrD transmembrane topology has beenpresented, in which LcrD is predicted to be an innermembrane-bound protein and in which thé positions ofLcrD-phoA fusions, predicted membrane-spanning seg-ments and charged amino acid residues indicate periplas-mic and cytoplasmic domains of thé protein (Piano et al.
Fig. 4. Multiple séquence alignaient of HrpO (P. solanacearum),with FlbF (Caulobacfer crescentus), FlhA (Bacillus subtilis), MxiA(Shigella flexnerï), VirH (S. flexneri). InvA (Salmonella ty-phimurium), LcrD (Yersiniapestis KIM; labelled LcrDl), LcrD (Y.enterocolilica ; labelled LcrD2) and HrpC2 (X. campestris pv. vesi-catoria). Amino acids that are shared identically or conservativelyare written in capital leiters and thé consensus Une of thé alignment(cons) contaîns amino acids that are identical in ail nine proteins.The numbering represents thé alignment length including gaps; théactua! predicted number of amino acid of each protein are: 700(FlbF), 677 (FlhA), 666 (MxiA), 686 (VirH), 686 (InvA), 704(LcrD), 705 (LcrD2), 645 (HrpC2) and 690 (HrpO). Underlinedamino acids represent potential membrane-associated hélices, aspredicted by thé PC/Gène program (see thc Materials and meth-ods), using thé method of Eisenberg et al. (1984). Bars labelled 1 to8 above thé séquences represent thé régions predicted by Piano etal. (1991) to be membrane-spanning segments in LcrD. Asterisksabove thé séquences correspond to lysine and arginine residues.which are conserved between LcrD and thé majority of thé otherproteins. Percentage identity and similariîy values are given in thétext. This alignment was produced using thé PILEUP program
1991). By comparison, thé similar positions of predictedmembrane-associated hélices and of charged amino acidresidues (lysine and arginine) in HrpO and in thé otherproteins shown in Fig. 4 indicate that thèse proteins ailshare a similar secondary structure and transmembranetopology.
According to this topology thé conserved lysine andarginine residues are predicted to be in thé cytoplasmicdomains of thé proteins, in agreement with thé "positive -inside rule" of von Heijne (1986). TnphoA analysis ofInvA of S. typhimurium has also indicated that InvA is aninner membrane-bound protein (Galan et al. 1992). Ac-cording to thé prédiction of membrane-associated hélicesîn Fig. 4 produced by thé method of Eisenberg et al.(1984), LcrD has a hélix covering amino acids 66 to 86which is not predicted by Piano et al. (1991) to be mem-brane-spanning. This région is also predicted to be mem-brane-assocïttted in ail thé other proteins presented inFig. 4 ; however, it is predicted to be located at thé surfaceof thé membrane rather than in a transmembrane posi-tion, which does not contradict thé model of LcrD topol-ogy. Within thé C-terminal parts of most of thèse pro-teins there is a final membrane-associated région, but itsposition is not well conserved among thé nine proteins.This région is only in équivalent positions in VirH andMxiA, which are highly homologous to each other and,interestingly, for thé two Hrp proteins, HrpO andHrpC2, thé last predicted membrane-associated région isin thé same position in each protein. The extremely highlevel of séquence identity between HrpO and HrpC2(66%), in fact reflects thé very high level of homology inthé 200 or so C-terminal amino acids of each protein, aswell as in thé N-terrninal régions. There is only a centralpart of HrpO which is not conserved with HrpC2.
Since LcrD and FlbF are both thought to be involvedin thé régulation of thé expression of other gènes, weperformed computer searches on HrpO for motifs typi-cally found in regulatory proteins. No such motifs wererevealed and no homologies were found to other regula-tory proteins; in particular there is no significant homol-ogy to HrpB of P. solanacearum, which appears to be amember of thé AraC family of regulatory proteins(Genin et al. 1992b), or to HrpS of P. syringae. pv.phaseolicola, which appears to be a member of thé NtrCfamily of regulatory proteins (Grimm and Panopoulos1989). Nevertheless, we investigated whether HrpO isinvolved in thé control of thé expression of other hrpgènes. Two potential targets, consisting of thé hrp gènepromoters of hrpB (Genin et al. 1992b) and of transcrip-tion unit 2, cloned in front of thé coding séquence of thégène lacZ, were transferred into both an hrpO mutantand thé wild-type strain GMI1000. No différences werefound, however, between thé levels of p-galactosidaseactivity produced in each case (in inducing conditions) bythé mutant and wild-type strains, indicating that théexpression of hrpB and of transcription unit 2 is notdépendent on HrpO (our unpublished data).

388
Tobacco Tomato
T -f
Strain Strain
Fig. 5. Conductivity changes induced over 20 h in thé incubationmédium contaîning (a) tobacco or (b) tomato leaf dises infiltratedwith thé P. solanacearum strains GMI1000 (wild type), GMI1415(Hrp* Tn5-B20 mutant), GMI1501 and GMI1502 (hrpO mutants)or MES (stérile médium). Values are means of 15 îndependentmeasurements and thé vertical Unes represent SEM
Induction of elecîrolyte îeakage in P.solanacearum-/7te; interactions
One of thé first détectable events associated with thé HRat thé host plant plasmalemma is thé activation of a K +
efflux/H+ influx exchange mechanism, termed thé XR(Yucel et al. 1989). The resulting net efflux of electrolytesacross thé plant cell membrane has been proposed to leadto cell death and thé characteristic necrotic symptoms ofthé HR (Atkinson et al. 1985b). Given that hrp mutantsof P. syringae pv. syringae (Huang et al. 1991) andE. amylovora (Brisset and Paulin 1991) are unable toelicit either an HR or thé XR, hrp gènes may be involvedin thé production of a bacterial factor that is responsiblefor both host responses.
We investigated thé rôle of HrpO in thé elicitation ofthé XR by infiltrating thé P. solanacearum strainsGMI1000 (wild type), GMI1415 (Hrp+ Tn5-B20 mu-tant), GMI1501 and GMI1502 (hrpO mutants) into to-bacco and tomato leaves and measuring conductivitychanges produced in thé incubation médium. For bothplants conductivity changes were complète within 20 h(thé rate for tomato was always slower than for tobacco),and thé values presented are thé net increases in conduc-tivity over this time.
Figure 5 shows that GMI1501 and GMI1502 produceonly low levels of conductivity change on both plantscompared to thé wild type and GMI1415. For a par-ticular plant thé levels associated with thé mutants aresignificantly différent from those of thé wild type andGMI1415, according to Tukey's method of multiplecompaiïson at thé 0.05 level (Scheffé 1959); conductivitychanges produced by GMI1000 and GMI1415 are notsignificantly différent from each other according to thistest. For tobacco thé results for control infiltrations withstérile médium (labelled MES in Fig. 5) are not signifi-
cantly différent from those of thé hrp mutants GMI 1501and GMI 1502. This is not true for tomato, although théchanges produced by GMI1501 and GMI 1502 on toma-to are not significantly différent from other hrp mutantson tomato (data not shown).
Discussion
By complémentation and nucîeotide séquence analysesand by thé mapping of Tn5-B20 insertions, we hâvecharacterised in détail a région of thé large hrp gènecluster of P. solanacearum. The complémentation datademonstrate that thé previously defined transcriptionunit 3 (Arlat et al. 1992) corresponds to a single complé-mentation unit, designated complémentation unit 3. Thepredicted positions and direction of transcription of thégènes hrpN, hrpO and hpaP correspond completely tothose that were implied by thé positions, orientations,p-galactosidase activities and Hrp phenotypes of Tn5-B20 insertion mutations. From this we assume that thécoding régions are probably correctly predicted and inaddition, thé amino acid séquence of HrpO aligns wellwith those of several known or predicted proteins. At-tempts to confirm thèse prédictions by detecting thé pro-teins HrpN, HrpO and HpaP by expression under con-trol of thé T7 or lac promoters, hâve so far been unsuc-cessful, as hâve RNA studies. Similar problems hâvebeen encountered with other hrp gène loci of P. solana-cearum.
hrpN, hrpO and hpaP appear to constitute an operon.The régulation of expression of thèse gènes is presumablycontrolled, at least in part, by séquences located 5' ofhrpN, although no consensus promoter séquences couldbe found. In order to explain thé fact thaï thé plasmidpGMI737: ;1499 was sometimes able to complémentGMI 1502, we assume that in thèse cases either insertion1499 is nonpolar or there is a weak promoter présentwithin thé complémentation unit, in addition to thé mainpromoter of thé operon. Since thé subséquent position-ing of thé two relevant insertions showed that insertion1499 is inserted in hrpN and insertion 1502 in hrpO, thésecond explanation implies that hrpO may hâve its ownpromoter. If this promoter does exist then it is presum-ably either only weakly active relative to thé main one oris only active under certain conditions since complé-mentation was not always found. As a conséquence ofthé séquence similarity between HrpO of -P. solanacear-um and FlbF of C. crescentus we looked for nucîeotideséquence éléments similar to those found in promoterstranscribed by thé sigma-28 form of RNA polymerase,which has recently been shown to be essential for théfunctioning of theflbF promoter of C. crescentus (San-ders et al. 1992). Such séquence éléments are not obviousin thé région 5' of hrpO or in thé putative promoterrégion of thé operon 5' of hrpN. Since hrp gène ex-pression in P. syringae pv. phaseolicola is dépendent onsigma-54 (Fellay et al. 1991), and since there is a séquencemotif 5' of hrpS of P. syringae pv. phaseolicola resem-bling thé — 24/— 12 régions of promoters that are recog-nised by thé form of RNA polymerase containing sigma-

389
54 (Grimm and Panopoulos 1989), we also looked forsuch a séquence motif, but were unable to find any.
The considérable séquence similarities found betweenHrpO of P. solanacearum and what appears to be a newfamily of bacterial proteins, indicate that thèse proteinsprobably hâve structural and functional features in com-mon, particularly among thé highly conserved N-termi-nal domains. Despite thé fact that LcrD and FlbF hâveregulatory rôles, no homologies were found to otherknown regulatory proteins. One of thé cloned promotersused to investigate a potential regulatory rôle for HrpOwas that of hrpB thé product of which, HrpB, has beenshown to positively regulate thé expression of severalother hrp gènes in thé cluster (Genin et al. 1992b).Therefore, by analogy to thé position of FlbF at théhighest level of thé regulatory hierarchy of fia gène ex-pression (Newton et al. 1989), we might hâve expectedHrpO to control thé expression of HrpB. This does notseem to be thé case. However, in Yersinia there is asécrétion system for thé Yop proteins, encoded by gènesin thé virA and virC loci (Michiels et al. 1991), and it hasbeen suggested that LcrD of Y. pestis could be involvedin thé sécrétion or export of Yops to thé outer membrane(Piano et al. 1991). The regulatory phenotype ascribed toan LcrD mutant could therefore be explained if, forexample, this translocation function included thé move-ment of a sensor/regulator protein or of components ofthé secretory system, whose accumulation in an LcrDmutant rnight hâve regulatory feedback effects. Prelimi-nary data in fact suggest a secretory rôle for HrpO(C. Boucher, unpublished results), which, together withthé fact that MxiA, VirH and FlhA hâve well charac-terised secretory functions, implies that this family ofproteins consists principally of proteins with rôles insécrétion.
The gènes coding for each of thé proteins of thisfamily belong to clusters of gènes, which code for sé-crétion Systems. Diverse proteins are secreted by thèsevarious Systems, including flagellar proteins, which areassembled to form an extracellular structure, Ipa pro-teins of S. flexneri, which are surface-located antigens,and Yops, which are proteins of Yersinia exported intothé extracellular médium. Furthermore, thèse secretedproteins perform différent rôles in their respective bac-terial hosts ; they are virulence déterminants in thé caseof thé Ipa proteins of S. flexneri and of thé Yops ofYersinia, but thé flagellum of S. typhimurium is involvedin motility not virulence. Despite this apparent diversityin thèse sécrétion Systems, both in terms of thé nature ofthé secreted proteins and of their rôles, other proteinhomologies hâve already been found between thé dif-férent Systems. For example, we hâve found homologiesbetween other P. solanacearum Hrp proteins and Yer-sinia proteins involved in Yop sécrétion (Gough et al.1992) and thèse same homologies were found betweenthé corresponding Hrp proteins of X. c. pv. vesicatoriaand Yop sécrétion proteins of Yersinia (Fenselau et al.1992). At least two other predicted proteins from thé spalocus of S. flexneri show séquence similarity to predictedHrp proteins of P. solanacearum (unpublished data) andthere is good homology between one of thèse Spa pro-
teins of S. flexneri (Spa.41) and Flil of S. typhimurium andORF4 of thé B. subtilis flaA locus (Venkatesan et al.1992). In addition, other Salmonella proteins from thésame locus as InvA hâve been found to share séquencesimilarity with Yersinia virulence plasmid-encoded pro-teins implicated in thé sécrétion of Yops (Galan et al.1992).
There is in fact, a commun feature among those pro-teins having characterised secretory functions, namelythat thé secretory function itself involves thé sécrétion ofgroups of proteins which do not hâve thé N-terminalsignal séquences characteristic of many secreted or inté-gral membrane proteins. This is true for thé Ipas of S.flexneri (Venkatesan et al. 1992), thé Yops of Yersinia(Michiels and Cornelis 1991) and thé flagellar com-ponents of S. typhimurium, B. subtilis and E. coli (Vogleret al. 1991). It appears therefore that thèse sécrétionSystems probably involve a common mechanism for se-creting proteins without signal séquences, and this mech-anism seems to be différent from other signal sequence-independent sécrétion pathways (Wandersman 1992). Inthé case of Yops, it has been shown that thé récognitionsignal of différent exported proteins is located in théN-terminal part of thé protein, but no consensus motifhas been found, indicating that this récognition signal isconformational rather than séquence spécifie (Michielsand Cornelis 1991). The discovery of thèse homologiesnot only supports thé hypothesis already made, thatsome hrp gene-encoded proteins are involved in thé ex-tracellular production of one or more macromolecules(Gough et al. 1992), but also indicates that thé sécrétionof thèse molécules involves a distinct, widely occurringmechanism that is signal sequence-independent.
Following thé discovery by Boucher et al. (1987) thatail tested X. campeslris (X. c) pathovars contain DNAwhich hybridises to thé large hrp gène cluster of P. sola-nacearum, it has recently been shown by hybridisationexperiments that X. c. pv. campeslris contains an hrp gènecluster which is homoîogous and colinear over a con-tinuous 19 kb DNA région with thé P. solanacearum hrpgène cluster (Arlat et al, 1991). hrp gènes homoîogous toP. solanacearum hrp gènes were also isolated from X. c.pv. vitians and were found to be functionally inter-changeable with those from X. c. pv. campesîris (Arlat etal. 1991). Comparison of thé complémentation groupsand transcriptional organisation of thé P. solanacearumand X. c. pv. vesicatoria hrp gène clusters indicated thatcomplémentation group 3 of thé P. solanacearum hrpgène cluster described in this paper could correspond tothé hrpC locus of thé X. c. pv. vesicatoria hrp gène cluster(Bonas et al. 1991). Thèse two régions are approximatelythé same size, are located in équivalent positions in thétwo clusters and are transcribed in thé same direction.The discovery of séquence similarity between HrpO anda predicted Hrp protein from thé équivalent position inthé X. c, pv. vesicatoria hrp gène cluster is thereforecompletely consistent with ail thèse data. In addition,HrpC2 has recently been reported to show séquencesimilarit> to LcrD of Y. pestis KIM (Fenselau et al.1992). The highly conserved proteins HrpO and HrpC2are probably functionally équivalent, which would con-

390
firm thé preliminary functional complémentation de-tected between some of thé hrp séquences of X. c. pv.campestris and P. solanacearum (Arlat et al. 1991). Werecently demonstrated DNA homology between hrpO of/*. solanacearum and thé hrp gène clusters of X. c. pv.campestris (Arlat et al. 1991), E. amylovora (Barny et al.1990) and P, syringae pv. phaseolicola (Rahme et al.1991), indicating that HrpO is conserved among thèsevarious bacteria (Gough et al. 1992) and may be a com-mon feature of alJ hrp gène clusters.
Two other predicted Hrp proteins of P. solanacearumalso hâve high levels of séquence similarity with predictedX. c. pv. vesicatoria Hrp proteins (Gough et al. 1992).Based on thèse results we compared our séquence datafor thé hrp gène cluster of P, solanacearum with thatrecently published for pathogenicity gènes from X. c. pv.glycines (Hwang et al. 1992). This comparison revealedthree ORFs with amino acid séquence similarity (unpub-lished data), including a potential HpaP homologue(31% identity over 134 aa) located 5' of ORF1 of thé X.c. pv. glycines séquence. The three régions are in équiv-alent relative positions in thé two DNA séquences, whichis again consistent with thé finding of Arlat et al. (1991),that thé X. campestris and P. solanacearum hrp gèneclusters are homologous and colinear. This homologysupports thé hypothesis that hpaP is a coding séquence,although we were unable to detect thé protein HpaP inexpression Systems.
The electrolyte leakage data demonstrate that P. sola-nacearum is like other phytopathogenic bacteria in thatthé HR produced by P. solanacearum on a nonhost plantis correlated with thé induction of an XR (Atkinson etal. 1985a, b; Baker et al. 1987; Brisset and Paulin 1991).Also demonstrated in this paper is thé finding that hrpmutants of P, solanacearum are unable to induce eitherplant response, and thus resemble typical hrp mutants ofphytopathogenic bacteria (Huang et al. 1991 ; Brisset andPaulin 1991). The compatible reaction between P. solana-cearum and thé tomato is also associated with an XR, asis thé case for compatible plant-bacterial interactionsinvolving P. syringae pv. syringae (Atkinson and Baker1987) and E. amylovora (Brisset and Paulin 1991). Theelectrolyte leakage induced by P, syringae pv. syringae inbean leaf tissue (host plant) is associated with an increasein thé pH of plant intercellular fiuids and with bacteria]multiplication. In facl increased intercellular pH hasbeen proposed to lead to increased nutrient levels, whichpromote bacterial multiplication (Atkinson and Baker1987). As is thé case for most hrp mutants, P. solanacea-rum hrp mutants that are able to infect and colonisetomato plants hâve been demonstrated to reach in plantapopulation levels much lower than those of thé wild type(Trigalet and Demery 1986). Considering this and ourresults demonstrating that thé induction of electrolyteleakage by P. solanacearum in host tissue is hrp gcne-dependent, hrp gene-encoded functions may be involvedin thé compatible interaction in diverting plant meta-bolites for bacterial growth, as has been suggested byBoucher et al. (1992). In accordance with thé model wehâve proposed, in which it is hypothesised that hrp gènescode for a spécifie export System (Gough et al. 1992), thé
protein(s) exported could be responsible for triggeringelectrolyte leakage in thé plant and thé recently dis-covered extracellular HR-inducing factor of P. solana-cearum is a likely candidate for such an elicitor (Geninet al. 1992a). Harpin, thé proteinaceous elicitor of théHR recently isolated from E. amylovora has already beenshown to induce an XR in tobacco cell suspension cul-tures (Wei et al. 1992).
Acknowledgements. We would like to thank M.N. Brisset (Stationde Pathologie Végétale, INRA, Beaucouzé) for help with thé proto-col for measurements of electroJyte leakage, A. Moisan (Station deBiométrie, INRA, Toulouse) for thé statistical analysis of conduc-tivity changes, C. Rosenberg for invaluable help with Computingand F. Van Gijsegem for protein expression studies. We are alsogratefu] to U. Bonas (IGF, Berlin) for giving us séquence data prîorto publication. This work was supported by grants from thé Euro-pean Economie Community (BIOT-CT90-0168) and thé NorthAtlantic Treaty Organisation [(21)880310/88].
Références
Akiyoshi DE, Regier DA, Gordon MP (1989) Nucleotide séquenceof thé tzs gène from Pseudomonas solanacearum strain K60.Nucleic Acids Res 17:8886
Arlat M, Gough CL, Barber CE, Boucher C, Daniels MJ (1991)Xanthomonas campesîris contains a cluster of hrp gènes relatedto thé larger hrp cluster of Pseudomonas solanacearum. MolPlant-Microbe Internet 4:593-601
Arlat M, Gough CL, Zischek C, Barberis PA, Trigalet A, BoucherÇA (1992) Transcriptional organisation and expression of thélarge hrp gène cluster of Pseudomonas solanacearum. Mol Plant-Microbe Interact 5:187-193
Atkinson MM, Baker CJ (1987) Association of host plasma mem-brane K+/H+ exchange with multiplication of Pseudomonassyringae pv. syringae in Phaseolus vulgaris. Phytopathology77:1273-1279
Atkinson MM, Huang JS, Knopp JA (I985a) Hypersensitivity ofsuspension-cultured tobacco cells to pathogenic bacteria. Phy-topathology 75:1270-1274
Atkinson MM, Huang JS, Knopp JA (I985b) The hypersensitivereaction of tobacco to Pseudomonas syringae pv. pisi. PlantPhysiol 79:843-847
Baker CJ, Atkinson MM, Collmer A (1987) Concurrent loss in Tn5mutants of Pseudomonas syringae pv. syringae of thé ability toinduce thé hypersensitive response and host plasma membraneK+/H+ exchange in tobacco. Phytopathology 77:1268-1272
Barny MA, Guinebretière MH, Marçais B, Coissac E, Paulin JP,Laurent J (1990) Cloning of a large gène cluster involved inErwînia amylovora CFBP1430 virulence. Mol Microbîol4:777-786
Bauer DW, Béer SV (1991) Further characterization of an hrp gènecluster of Envinia amylovora. Mol Plant-Microbe Interact4:493-499
Bibb MJ, Findlay PR, Johnson MW (1984) The relationship be-tween base composition and codon usage in bacterial gènes andils use for thé simple and reliable identification of protein-codingséquences. Gène 30:157-166
Biggin MD, Gibson TJ, Hong GF (1983) Buffer gradient gels and35S label as an aid to rapid DNA séquence détermination. ProcNatI Acad Sci USA 80:3963-3965
Birnboim HC, Doly J (1979) A rapid alkaline extraction procédurefor screening recombinant plasmid DNA. Nucleic Acids Res7:1513-1523
Bonas U, Schûlte R, Fenselau S, MJnsavage GV, Staskawicz BJ,Stall RE (1991) Isolation of a gène cluster from Xanthomonascampestris pv. vesicatoria that détermines paîhogenicity and théhypersensitive response on pepper and tomato. Mol Plant-Microbe Interact 4:81-88

391
Boucher ÇA, Barberis P, Trigalet A, Demery D (1985) Transposonmutagenesis of Pseudomonas solanacearum : isolation of Tn5induced avirulent mutants. J Gen Microbiol 131:2449-2457
Boucher C, Martinel A, Barberis P, Alloing G, Zischek C (1986)Virulence gènes are carried by a megaplasmid of thé plantpathogen Pseudomonas solanacearum. Mol Gcn Genêt205:270-275
Boucher ÇA, Van Gijsegem F, Barberis PA, Arlat M, Zischek C(1987) Pseudomonas solanacearum gènes controlling bolhpathogenicity and hypersensitivity on tobacco are clustered.J Bacteriol 169:5626-5632
Boucher ÇA, Barberis PA, Arlat M (1988) Acridine orange sélectsfor deletion of hrp gènes in ail races of Pseudomonas solanacea-rum. Mol Plant-Microbe Interact 1:282-288
Boucher ÇA, Gough CL, Arlat M (1992) Molecular genetics ofpathogenicity déterminants of Pseudomonas solanacearum withspécial emphasis on hrp gènes. Annu Rev Phytopathol30:443^161
Brisset MN, Paulin JP (1991) Relationships between electrolyteleakage from Pyrus communis and virulence of Erwinia amylovo-ra. Physiol Mol Plant Pathol 39:443-453
Buddenhagen IW, Kelman A (1964) Biological and physiologicalaspects of bacterial with caused by Pseudomonas solanacearum.Annu Rev Phytopathol 2:203-230
Cornelis GR, Biot T, Lambert de Rouvroit C, Mîchiels T, MulderB, Sluiters C, Sory MP, Van bouchaute M, Vanooteghem JC(1989) The Yersinia yop regulon. Mol Microbiol 3:1455-1459
Curry KA, Tomich C-SC (1988) Effect of ribosome binding site ongène expression in Escherichia coll. DNA 7:173-179
Deretic V, Konyecsni WM, Mohr CD, Martin DW, Hibler NS(1989) Common determinators of promoter control in Pseudo-monas and other bacteria. Bio/Technology 7:1249-1254
Devereux J, Haeberli P, Smithîes O (1984) A comprehensive set ofséquence analysis programs for thé VAX. Nucleic Acids Res12:387-395
Eisenberg D, Schwarz E, Komaromy M, Wall R (1984) Analysis ofmembrane and surface protein séquences with thé hydrophobicmoment plot. J Mol Biol 179:125-142
Fellay R, Rahme LG, Mindrinos MN, Frederick RD, Pisi A,Panopoulos NJ (1991) Gènes and signais controlling théPseudomonas syringae pv. phaseolicola-planl interaction. In:Heynecke H, Verma DPS (eds) Advances in molecular geneticsof plant-microbe interactions. Kluwer Académie Publishers,Dordrecht, Netherlands, pp 45-52
Fenselau S, Balbo I, Bonas U (1992) Déterminants of pathogenicityin Xanthomonas campesîris pv. vesicaîoria are related to proteinsinvolved in sécrétion in bacterial pathogens of animais. MolPlant-Microbe Interact 5:390-396
Galan JE, Ginocchio C, Costeas P (1992) Molecular and functionalcharacterization of thé Salmonella invasion gène invA : homol-ogy of InvA to members of a new protein family. J Bacteriol174:4338-4349
Genin S, Gough CL, Arlat M, Zischek C, Van Gijsegem F, BarberisP, Boucher ÇA (1992a) Involvement of Pseudomonas solana-cearum hrp gènes in thé sécrétion of a bacterial compound whichinduces a hypersensitive-like response on tobacco. In: NesterEW, Verma DPS (eds.) Sixth International Symposium onMolecular Plant-Microbe Interactions, July 11-16 1992, Seattle;Kluwer Académie Publishers, Dordrecht, Netherlands
Genin S, Gough CL, Zischek C, Boucher ÇA (1992b) Evidence thatthé hrpB gène encodes a positive regulator of pathogenicitygènes from Pseudomonas solanacearum. Mol Microbiol6:3065-3076
Gibson TJ, Coulson AR, Sulston JE, Little PFR (1987) Lorist2, acosmid with transcriptional terminators insulating vector gènesfrom interférence by promoters within thé insert : efTect on DNAyield and cloned insert frequency. Gène 53:275-281
Goguen JD, Yother J, Straley SC ( 1984) Genetic analysis of thé lowcalcium response in Yersinia pestis Mu dl(Ap lac) insertionmutants. J Bacteriol 160:842-848
Gough CL, Genin S, Zischek C, Boucher ÇA (1992) hrp gènes ofPseudomonas solanacearum are homologous to pathogenicily
déterminants of animal pathogenic bacteria and are conservedamong plant pathogenic bacteria. Mol Plant-Microbe Interact5:384-389
Grimm C, Panopoulos NJ (1989). The prcdicted protein product ofa pathogenicity locus from Pseudomonas syringae pv. phaseo-licola is homologous to a highly conserved domain of severalprocaryotic regulatory proteins. J Bacteriol 171:5031-5038
Hawley DK, McClure WR (1983) Compilation and analysis ofEscherichia coli promoter DNA séquences. Nucleînc Acids Res11:2237-2255
Henikoff S (1984) Unidirectional digestion with exonuclease IIIcréâtes targeted breakpoints for DNA sequencing. Gène28:351-359
Holmes DS, Quigley M (1981) A rapid boiling method for thépréparation of bacterial plasmids. Anal Biochem 114:193-197
Huang H-C, Hutcheson SW, Collmer A (1991) Characterizalion ofthé hrp cluster from Pseudomonas syringae pv. syringae 61 andThphoA tagging of gènes encoding exportcd or membrane-span-ning Hrp proteins. Mol Plant-Microbe Interact 4:469-476
Huang Y, Xu P, Sequeira L (1990) A second cluster of gènes thatspecify pathogenicity and host response in Pseudomonas solana-cearum. Mol Plant-Microbe Interact 3:48-53
Hwang I, Lim SM, Shaw PD (1992) Cloning and characterisationof pathogenicity gènes from Xanthomonas campesîris pv. glyci-nes. J Bacteriol 174:1923-1931
Jenner C, Hitchin E, Mansfield J, Walters K, Betteridge P, TeversonD, Taylor J (1991) Gene-for-gene interactions between Pseudo-monas syringae pv. phaseolicola and Phaseolus. Mol Plant-Microbe Interact 4:553-562
Kyte J, Doolittle RF (1982) A simple method for displaying théhydrophathic character of a protein. J Mol Biol 157:105-132
Laby RJ, Béer SV (1990) The hrp gène cluster of Erwinia amylovorashares DNA homology with other bacteria. Phytopathology80:1038-1039
Leong SA, Ditta GS, Helinski DR (1982) Heme biosynthesis inRhizobium. Identification of a cloned gène coding for a-ami-nolevulinic acid synthetase from Rhizobium meliloti. J BiolChem 257:8724-8730
Lindgren PB, Panopoulos NJ, Staskawicz BJ, Dahlbeck D (1988)Gènes required for pathogenicity and hypersensitivity are con-served and interchangeable among pathovars of Pseudomonassyringae. Mol Gen Genêt 211:499-506
Lindgren PB, Frederick R, Govindarajan AG, Panopoulos NJ,Staskawicz BJ, Lindow SE (1989) An ice nucleation reportergène System: identification of inducible pathogenicity gènes inPseudomonas syringae pv. phaseolicola. EMBO J 8:1291-1301
Maniatis T, Fritsch E, Sambrook J (1982) Molecular cloning: alaboratory manual. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, New York
Michiels T, Cornelis G (1991) Sécrétion of hybrid proteins by théYersinia export System. J Bacteriol 173:1677-1685
Michiels T, Vanooteghem J-C, Lambert de Rouvroit C, China B,Gustin A, Boudry P, Cornelis GR (1991) Analysis of virC, anoperon involved in thé sécrétion of Yop proteins by Yersiniaenlerolilica. J Bacteriol 173:4994-5009
Mizusawa S, Nishimura S, Seela F (1986) Improvement of thédideoxy chain termination method of DNA sequencing by useof deoxy-7-deazaguanosine triphosphate in place of dGTP. Nu-cleic Acids Res 14:1319-1324
Mukhopadhyay P, Williams J, Mills D (1988) Molecular analysisof a pathogenicity locus in Pseudomonas syringae pv. syringae.J Bacîeriol 170:5479-5488
Newton A, Ohta N, Ramakrishnan G, Mullin D, Raymond G( 1989) Genetic switching in thé flagellar gène hierarchy of Caul-obacîer requires négative as well as positive régulation of tran-scription. Proc Natl Acad Sci USA 86:6651-6655
Piano GV, Barve SS, Straley SC (1991) LcrD, a membrane-boundregulator of thé Yersinia pesîis low-calcium response. J Bacteriol173:7293-7303
Platt T (1986) Transcription termination and thé régulation of gèneexpression. Annu Rev Biochem 55:339-372

392
Rahme LG, Mindrinos MN, Panopoulos NJ (1991) Genetic andIranscriptional organization of thé hrp cluster of Pseudomonassyringae pv. phaseolicola. J Bacteriol 173:575-586
Ramakrishnan G, Zhao J-L, Newton A (1991) The cell cycle-regulated flagella genej^W7 of Cauîobacter crescentus is homolo-gous îo a virulence locus (IcrD) of Yersinia pestis. J Bacteriol173:7283-7292
Sanders LA, Van Way S, Mullin DA (1992) Characterization ofthé Cauîobacter crescentus flbF promoter and identification ofthé inferred FlbF product as a homolog of thé LcrD proteinfrom a Yersinia enterocoîitica virulence plasmid. J Bacteriol174:857-866
Sanger F, Nicklen S, Coulson AR (1977) DNA sequencîng withchain-terminating inhibîtors. Proc Natl Acad Sci USA74:5463-5467
Scheffé H (1959) Tukey's method of multiple comparison. In: Theanalysis of varîance. Wiley, pp 73-75
Shine J, Dalgarno L (1974) The 3'-terminaI séquence of Escherichiacoli 16S ribosomal RNA: complementarity to nonsense tripletsand ribosome binding sites. Proc Natl Acad Sci USA71:1342-1346
Sneath BJ, Howson JM, Béer SV (1990) A pathogenicity gène fromErwinia amylovora encodes a predicted protein product homolo-gous to a family of prokaryotic response regulators. Phytopath-ology 80:'l038
Trigalet A, Demery D (1986) Invasiveness in tomato plant of Tn5-induced Pseudomonas solanacearum, Physiol Mol Plant Pathol28:423-430
Venkatesan MM, Buysse JM, Oaks EV (1992) Surface présentationof Shigeîla flexneri invasion plasmid antigens requires théproducts of thé spa locus. J Bacteriol 174:1990-2001
Vieira J, Messing J (1987) Production of single-stranded plasmidDNA. Methods Enzymol 153:3-11
Vogler AP, Homma M, Irikura VM, Macnab RM (1991) Sal-monella typhimurium mutants defective in flagellar filament re-growth and séquence similarity of Flil to FOF1, vacuolar, andarchaebacterial ATPase subunits. J Bacteriol 173:3564-3572
von Heijne G (1986) The distribution of positively charged residuesin bacterial inner membrane proteins correlates with thé trans-membrane topology. EMBO J 5:3021-3027
Wandersman C (1992) Sécrétion across thé bacterial outer mem-brane. Trends Genêt 8:317-322
Wei Z-M, Laby RJ, ZumorT CH, Bauer DW, He SY, Collmer A,Béer SV (1992) Harpin, elicitor of thé hypersensuive responseproduced by thé plant pathogen Erwinia amylovora. Science257:85-88
Willis DK, Rich JJ, Hrabak EM (1991) hrp gènes of phytopath-ogenic bacteria. Mol Plant-Microbe Interact 4:132-138
Yucel I, Xiao Y, Hutcheson SW (1989) Influence of Pseudomonassyringae culture conditions on initiation of thé hypersensitiveresponse of cultured tobacco cells. Appl Environ Microbiol55:1724-1729
Zubay G (1983) Biochemistry. Addison-Wesley, Reading, Mass





















