
Gland formation of human colon cancer cells combined with foetal rat mesenchyme in organ culture: an...
8
Gland formation of human colon cancer cells combined with foetal rat mesenchyme in organ culture: an ultrastructural study HIROSHI FUKAMACHI*, TAKEO MIZUNO Zoological Institute, Faculty of Science, University of Tokyo, Hongo, 7-3-1, Bunkyo-ku, Tokyo 113, Japan and YOUNG S. KIM Gastrointestinal Research Laboratory!, Veterans Administration Medical Center, and Department of Medicine, University of California School of Medicine, CA 94143, USA * Author for correspondence Summary The morphogenetic potential of human colon cancer cells was examined by combining cells with foetal rat mesenchyme in organ culture. Out of four cell lines examined, LS174T cells formed glandular structures composed of a simple col- umnar or cuboidal epithelium with a lumen in the centre of the cell mass. Ultrastructurally, these gland-forming LS174T cells exhibited po- larity, with nuclei located basally, microvilli pro- jecting into the lumen, tight junctions at the cell- cell junction facing the lumen, desmosomes at the subluminal region, and basal laminae at the epithelial-mesenchymal interface. Other cell lines did not form such glandular structures, while some cells could form microvilli, tight junctions or basal laminae, which were arranged randomly. It is concluded that it is not the exist- ence but the ordered arrangement of these struc- tures that is essential for gland formation by epithelial cells. The mechanism of gland forma- tion is discussed with special reference to the arrangement of different structures. Key words: intestinal epithelium, morphogenesis, mesenchyme, organ culture. Introduction The interaction between the epithelium and surround- ing mesenchyme has been intensively studied by epi- thelial-mesenchymal recombination experiments. The results indicate that mesenchymes play major roles in determining epithelial morphogenesis in many organs, including the salivary gland and mammary gland (for reviews see Goldin, 1980; Sawyer & Fallon, 1983; Yamada, 1983; Cunha et al. 1985). In the developing digestive organs of mammals, in contrast, such an instructive tissue-specific effect of mesenchyme has not been found, and mesenchyme has been reported to support epithelial morphogenesis permissively in both stomach (Fukamachi et al. 1979) and intestine (Fuka- machi & Takayama, 1980). The nature of epithelial-mesenchymal interaction is not fully understood, but experiments with salivary glands indicate that extracellular matrix, including Journal of Cell Science 87, 615-621 (1987) Printed in Great Britain © The Company of Biologists Limited 1987 basal laminae (Thompson & Spooner, 1982) and col- lagen fibres (Nakanishi et al. 1986a,6), play an es- sential role in the interaction. On the other hand, investigations of the process of gland formation indi- cate the importance of the formation of interepithelial junctions for the establishment of epithelial polarity in the developing liver (Montesano et al. 1975), thyroid gland (Luciano et al. 1979) and intestine (Madaraef al. 1981). Formation of extracellular matrix, establish- ment of polarity of epithelial cells, and formation of interepithelial junctions could be closely related, and there may exist complicated interactions between them for controlling normal morphogenesis of epithelial cells. But the mechanism of the interaction has not been extensively studied, partly because model systems have not been fully developed. In a previous paper we reported that LS174T human colon cancer cells formed glandular structures in com- bination with foetal rat mesenchyme in organ culture, while HT29 cells did not (Fukamachi et al. 1986). 615
Transcript of Gland formation of human colon cancer cells combined with foetal rat mesenchyme in organ culture: an...

Gland formation of human colon cancer cells combined with foetal rat
mesenchyme in organ culture: an ultrastructural study
HIROSHI FUKAMACHI*, TAKEO MIZUNO
Zoological Institute, Faculty of Science, University of Tokyo, Hongo, 7-3-1, Bunkyo-ku, Tokyo 113, Japan
and YOUNG S. KIM
Gastrointestinal Research Laboratory!, Veterans Administration Medical Center, and Department of Medicine,University of California School of Medicine, CA 94143, USA
* Author for correspondence
Summary
The morphogenetic potential of human coloncancer cells was examined by combining cellswith foetal rat mesenchyme in organ culture. Outof four cell lines examined, LS174T cells formedglandular structures composed of a simple col-umnar or cuboidal epithelium with a lumen inthe centre of the cell mass. Ultrastructurally,these gland-forming LS174T cells exhibited po-larity, with nuclei located basally, microvilli pro-jecting into the lumen, tight junctions at the cell-cell junction facing the lumen, desmosomes atthe subluminal region, and basal laminae at the
epithelial-mesenchymal interface. Other celllines did not form such glandular structures,while some cells could form microvilli, tightjunctions or basal laminae, which were arrangedrandomly. It is concluded that it is not the exist-ence but the ordered arrangement of these struc-tures that is essential for gland formation byepithelial cells. The mechanism of gland forma-tion is discussed with special reference to thearrangement of different structures.
Key words: intestinal epithelium, morphogenesis,mesenchyme, organ culture.
Introduction
The interaction between the epithelium and surround-ing mesenchyme has been intensively studied by epi-thelial-mesenchymal recombination experiments. Theresults indicate that mesenchymes play major roles indetermining epithelial morphogenesis in many organs,including the salivary gland and mammary gland (forreviews see Goldin, 1980; Sawyer & Fallon, 1983;Yamada, 1983; Cunha et al. 1985). In the developingdigestive organs of mammals, in contrast, such aninstructive tissue-specific effect of mesenchyme has notbeen found, and mesenchyme has been reported tosupport epithelial morphogenesis permissively in bothstomach (Fukamachi et al. 1979) and intestine (Fuka-machi & Takayama, 1980).
The nature of epithelial-mesenchymal interaction isnot fully understood, but experiments with salivaryglands indicate that extracellular matrix, including
Journal of Cell Science 87, 615-621 (1987)Printed in Great Britain © The Company of Biologists Limited 1987
basal laminae (Thompson & Spooner, 1982) and col-lagen fibres (Nakanishi et al. 1986a,6), play an es-sential role in the interaction. On the other hand,investigations of the process of gland formation indi-cate the importance of the formation of interepithelialjunctions for the establishment of epithelial polarity inthe developing liver (Montesano et al. 1975), thyroidgland (Luciano et al. 1979) and intestine (Madaraef al.1981). Formation of extracellular matrix, establish-ment of polarity of epithelial cells, and formation ofinterepithelial junctions could be closely related, andthere may exist complicated interactions between themfor controlling normal morphogenesis of epithelialcells. But the mechanism of the interaction has notbeen extensively studied, partly because model systemshave not been fully developed.
In a previous paper we reported that LS174T humancolon cancer cells formed glandular structures in com-bination with foetal rat mesenchyme in organ culture,while HT29 cells did not (Fukamachi et al. 1986).
615

Using this organ culture system, we have here exam-ined the morphogenetic potential of four human coloncancer cell lines, and compared their ultrastructure toclarify the relationship between the epithelial morpho-genetic potency and the ultrastructural characteristics.
Materials and methods
CellsFour human colon cancer cell lines were used in this study.LS174T was kindly provided by Dr B. D. Kahan of theUniversity of Texas (Houston, TX, USA), HT29 andSKCO-1 by Dr J. Fogh of the Sloan-Kettering Institute forCancer Research (Rye, NY, USA), and HCT48 by Dr R. M.Schultz of the National Cancer Institute (Bethesda, MD,USA). Cells were maintained as monolayer cultures inDulbecco's modified Eagle's medium (GIBCO Lab., USA)supplemented with 10% foetal bovine serum (Flow Lab.,USA) and antibiotics (penicillin 100 units ml"1, strepto-mycin lOOftgmF1, and Fungizone 0-25 j jgmF 1 ; MA Bio-products, USA) in a humidified atmosphere of 10% CO2 inair. The cells were cultured in a 25 cm2 plastic flask (LuxScientific Corp., USA) at 37°C, and medium was changedtwice weekly. They were subcultured every 2 weeks at thesplit ratios of 1:25 (LS174T), 1:50 (HT29), l:50(HCT48)and 1:5 (SKCO-1), after being suspended by treatment with0-25% trypsin and 0 0 2 % EDTA (GIBCO Lab.).
Combination of human colon cancer cells with foetalrat mesenchymeThe method of combination of human colon cancer cells withfoetal rat mesenchyme has been reported (Fukamachi et al.1986). Cells were seeded on membrane filters (MilliporeCorp., USA; type HAVVP) at 104 cells cm"2 (day 0). On day5, tissues were obtained from 16-5-day-old foetuses of Wis-tar/Mishina rats (kindly provided by the late Dr T. Noguchiof the National Institute of Genetics, Mishima, Japan)maintained in our laboratory by sister—brother mating.Forestomach, glandular stomach and duodenal tissue frag-ments were dissected out, and the epithelium and mesen-chyme were separated with the aid of collagenase as described(Fukamachi et al. 1979, 1986). The foetal mesenchyme wascombined with cancer cells by putting the mesenchyme onthe membrane filter on which cancer cells had been pre-viously cultured for 5 days, thus allowing the cells to beoverlaid with the mesenchyme. Then the membrane filterwith cancer cells and mesenchyme was laid on a stainless steelgrid, and the combination was cultured for 4 more dayswithout medium change.
Histology
Cultures were fixed with Bourn's fluid on day 9, and wereprocessed for paraffin sectioning and staining with PAS-haematoxylin. Some combinations were also fixed with 2-5 %glutaraldehyde in 0 1 M-cacodylate buffer (pH7-4) for 2h at4°C, washed with the buffer, and post-fixed with 1 % osmiumtetroxide in the buffer for 2h at 4°C. They were dehydratedthrough a graded series of ethyl alcohol and w-butyl glycidylether, and embedded in Embedding Resin (Taab Lab., UK).
Ultrathin sections were stained with uranyl acetate and leadcitrate, and observed with a JEOL 100CX electron micro-scope. For light microscopy, semi-thin sections were cut, andstained with Toluidine Blue.
Results
Characteristics of human colon cancer cells in cellculture
In cell culture in plastic flasks LS174T cells prolifer-ated relatively slowly with an approximate doublingtime of 32h. They did not migrate on the plastic, butpiled up to form cell masses. LS174T cells attached soclosely to each other on the surface of the cell massesthat cell boundaries could hardly be distinguished byphase-contrast microscopy (Fig. 1A). But transmissionelectron microscopy showed that LS174T cells at-tached loosely to each other in the cell mass. There wasa broad intercellular space between them, and noepithelial polarity was observed (see Fig. 3A).
In contrast, HT29 and HCT48 cells proliferatedrapidly, with an approximate doubling time of 20 and24 h, respectively. HT29 cells formed a typical polyg-onal cell sheet characteristic of epithelial cells(Fig. IB). HCT48 cells did not form a cell sheet, butindividual cells were oriented randomly, often exhibit-ing a criss-cross pattern or piling up (Fig. 1C). SKCO-1 cells proliferated very slowly, with an approximatedoubling time of 56h. They were round, with arelatively large cell body, and they never formed a cellsheet (Fig. ID).
Ultrastructural analysis of morphogenesis of humancolon cancer cells combined with foetal ratmesenchyme
In organ culture with foetal rat mesenchyme, humancolon cancer cells usually migrated into the mesen-chyme to proliferate and differentiate in it.
LS174T cells always formed cell masses in themesenchyme, and in about 25-40% of cases lumenswere found in the centre of the cell mass. These lumen-forming cells were columnar, with nuclei locatedbasally, and PAS-positive mucus was present in thelumen, indicating that cellular polarity had beeninduced by the surrounding mesenchyme (Figs 2A,3B). Ultrastructural analysis showed that the glandularstructure formed by LS174T cells was accompanied bytight junctions, microvilli, desmosomes and basal lam-inae arranged in the same order as in normal intestinalepithelial cells. Cells attached closely to each other withlittle intercellular space between them (Fig. 3B,C). Atmost (85-90%) cell-cell junctions facing a lumen,cells were connected by tight junctions (Fig. 3C).Microvilli with core filaments projected into the lumen,and desmosomes were observed at the subluminalregion (Fig. 3C). About 5-20% of the basal surfaces
616 H. Fukamachi et al.
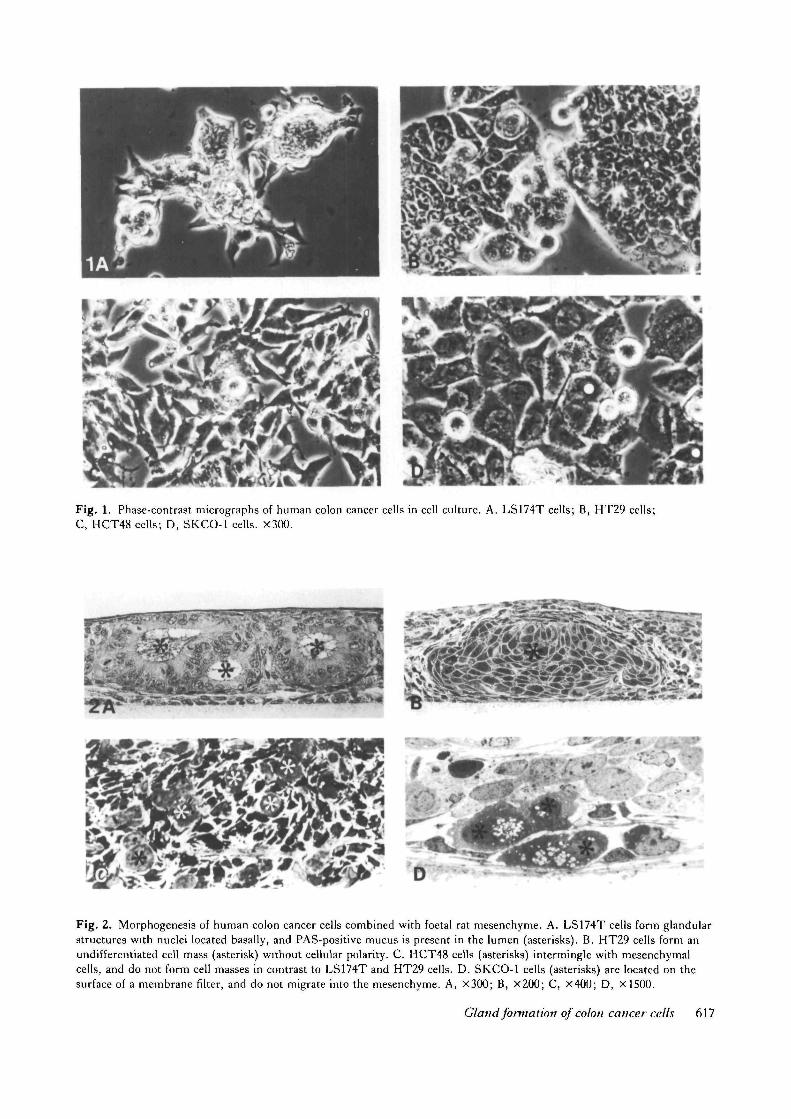
Fig. 1. Phase-contrast micrographs of human colon cancer cells in cell culture. A. LS174T cells; B, HT29 cells;C, HCT48 cells; D, SKCO-1 cells. X300.
7*f c* "y
Fig. 2. Morphogenesis of human colon cancer cells combined with foetal rat mesenchyme. A. LS174T cells form glandularstructures with nuclei located basally, and PAS-positive mucus is present in the lumen (asterisks). B. HT29 cells form anundifferentiated cell mass (asterisk) without cellular polarity. C. HCT48 cells (asterisks) intermingle with mesenchymalcells, and do not form cell masses in contrast to LS174T and HT29 cells. D. SKCO-1 cells (asterisks) are located on thesurface of a membrane filter, and do not migrate into the mesenchyme. A, X300; B, X200; C, X400; D, X1500.
Gland formation of colon cancer cells 617

Fig. 3. Transmission electron micrographs of LS174T cells. A. In cell culture on a coverglass, cells attach closely to eachother to form a smooth surface, but there is a broad intercellular space in the cell mass (asterisks). B. When combined witha foetal rat mesenchyme (stars), LS174T cells acquire cellular polarity, and are closely attached to each other to form alumen (asterisk). C. Higher magnification of a cell-cell junctional site facing a lumen (asterisk) induced by combinationwith a mesenchyme. Cells are connected by a tight junction (tj), and a desmosome (d). Microvilli (mv) with core filaments(arrowheads) are projecting into the lumen. D. Higher magnification of an interface of LS174T cells (asterisk) and amesenchyme. A basal lamina (arrowheads) is formed between them. A, X6300; B, X5800; C, X85OOO; D, XS0 0O0.
(at the epithelial-mesenchymal interface) was coveredwith basal lamina (Fig. 3D).
HT29 cells also formed cell masses with occasionalintraepithelial vacuoles, but never formed glandularstructures. Nuclei were located in the centre of a cell,and no polarity was observed in the cell mass (Fig. 2B).
Ultrastructurally, HT29 cells did not attach closely toeach other, and a broad intercellular space was alwaysobserved in the cell mass (Fig. 4A,B). As in the case ofLS174T cells, tight junctions, desmosomes, microvilliand basal laminae were also observed in the cell massformed by HT29 cells (Fig. 4A,B). In contrast to
618 H. Fukamachi et al.
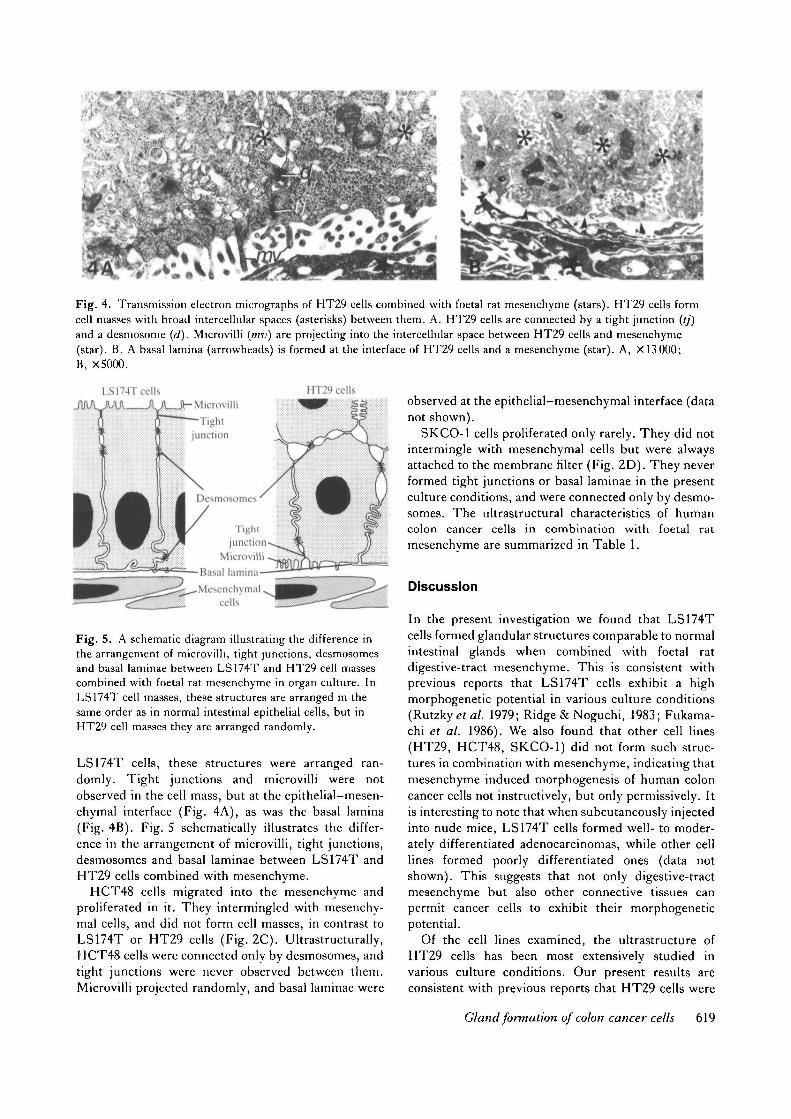
Fig. 4. Transmission electron micrographs of HT29 cells combined with foetal rat mesenchyme (stars). HT29 cells formcell masses with broad intercellular spaces (asterisks) between them. A. HT29 cells are connected by a tight junction (//)and a desmosome (d). Microvilli (mv) are projecting into the intercellular space between HT29 cells and mesenchyme(star). B. A basal lamina (arrowheads) is formed at the interface of HT29 cells and a mesenchyme (star). A, X 13000;B, X5000.
HT29 cellsLS174T cellsL _ _ J U L _ J - Microvilli
J T i g h tjunction
Desmosomes
Tightjunction
MicrovilliBasal kuninaMesenchymal
cells
Fig. 5. A schematic diagram illustrating the difference inthe arrangement of microvilli, tight junctions, desmosomesand basal laminae between LS174T and HT29 cell massescombined with foetal rat mesenchyme in organ culture. InLS174T cell masses, these structures are arranged in thesame order as in normal intestinal epithelial cells, but inHT29 cell masses they are arranged randomly.
LS174T cells, these structures were arranged ran-domly. Tight junctions and microvilli were notobserved in the cell mass, but at the epithelial-mesen-chymal interface (Fig. 4A), as was the basal lamina(Fig. 4B). Fig. 5 schematically illustrates the differ-ence in the arrangement of microvilli, tight junctions,desmosomes and basal laminae between LS174T andHT29 cells combined with mesenchyme.
HCT48 cells migrated into the mesenchyme andproliferated in it. They intermingled with mesenchy-mal cells, and did not form cell masses, in contrast toLS174T or HT29 cells (Fig. 2C). Ultrastructurally,HCT48 cells were connected only by desmosomes, andtight junctions were never observed between them.Microvilli projected randomly, and basal laminae were
observed at the epithelial-mesenchymal interface (datanot shown).
SKCO-1 cells proliferated only rarely. They did notintermingle with mesenchymal cells but were alwaysattached to the membrane filter (Fig. 2D). They neverformed tight junctions or basal laminae in the presentculture conditions, and were connected only by desmo-somes. The ultrastructural characteristics of humancolon cancer cells in combination with foetal ratmesenchyme are summarized in Table 1.
Discussion
In the present investigation we found that LS174Tcells formed glandular structures comparable to normalintestinal glands when combined with foetal ratdigestive-tract mesenchyme. This is consistent withprevious reports that LS174T cells exhibit a highmorphogenetic potential in various culture conditions(Rutzky et al. 1979; Ridge & Noguchi, 1983; Fukama-chi et al. 1986). We also found that other cell lines(HT29, HCT48, SKCO-1) did not form such struc-tures in combination with mesenchyme, indicating thatmesenchyme induced morphogenesis of human coloncancer cells not instructively, but only permissively. Itis interesting to note that when subcutaneously injectedinto nude mice, LS174T cells formed well- to moder-ately differentiated adenocarcinomas, while other celllines formed poorly differentiated ones (data notshown). This suggests that not only digestive-tractmesenchyme but also other connective tissues canpermit cancer cells to exhibit their morphogeneticpotential.
Of the cell lines examined, the ultrastructure ofHT29 cells has been most extensively studied invarious culture conditions. Our present results areconsistent with previous reports that HT29 cells were
Gland formation of colon cancer cells 619

Table 1. Ultrastructural characteristics of human colon cancer cells combined with foetal rat mesenchyme inorgan culture
Cell lineMorphogenesisin organ culture Microv Desmosomes
Basallaminae
Tightjunctions
LS174T Glandular structureHT29 Cell massHCT48 Intermingle with mesenchymal cellsSKCO-1 Do not migrate into mesenchyme
able to form tight junctions (Augeron & Laboisse,1984; Cohtnet al. 1985), microvilli (Remyet al. 1984)and desmosomes (Augeron & Laboisse, 1984). Whileultrastructurally functional differentiation of HT29cells has been reported (Augeron & Laboisse, 1984),we (Fukamachi et al. 1986; present study) and others(Haffen et al. 1981) have not found any gland forma-tion of HT29 cells in combination with mesenchyme.Thus, it may be concluded that ultrastructural differ-entiation does not necessarily result in gland formation.
We found in the present investigation that bothLS174T and HT29 cells were able to form tightjunctions, microvilli, desmosomes and basal laminae incombination with mesenchyme, while LS174T cellsformed glandular structures, and HT29 cells did not.The difference between HT29 and LS174T cells wasthat these structures were arranged in correct order inthe glandular structures formed by LS174T cells, butwere arranged randomly in the cell mass formed byHT29 cells (Fig. 5). This indicates that an essentialprocess of the formation of glandular structures is notthe formation but the arrangement of structures, andthat formation of these structures does not necessarilyresult in gland formation, though it may be a prerequi-site for gland formation.
Tight junctions were not observed in cultures ofHCT48 or SKCO-1 cells, irrespective of the combi-nation with mesenchyme, and basal laminae were notobserved in cultures of SKCO-1 cells, while all celllines examined could form desmosomes and microvilli(Table 1). This suggests that the formation of desmo-somes and microvilli might be a fundamental feature ofcolon epithelial cells, rarely lost by transformation,unlike the formation of tight junctions or basal laminae.This is consistent with the report that desmosomes arethe principal and fundamental adhesive intercellularjunctions of epithelia, and are widely shared and highlyconserved between different vertebrate classes (Cowin& Garrod, 1983).
We found in the present study that tight junctionswere formed at most (85-90 %) cell-cell adhesion sitesfacing the lumen, while basal laminae were found onlyin some regions (5—20 %) of the epithelial-mesenchy-mal interface when LS174T cells formed glandularstructures. This indicates that basal lamina formationis not responsible for gland formation, but that forma-
tion of tight junctions may be involved in it. Theimportance of basal lamina/basement membrane ininducing epithelial polarity has been reported both innormal epithelial cells (Bernfield et al. 1972; Vracko,1974; Banerjee et al. 1977), and in transformed cells(Ingber et al. 1981; Reddy et al. 1986). However,experiments with epithelial cells on/in collagen gels(Bennett, 1980; Chambard et al. 1981; Hal loa / . 1982;Haeuptle et al. 1983; Garbi et al. 1984) or on agarose-coated dishes (Nitsch & Wollman, 1980; Nitsch et al.1984) have shown that epithelial polarity can beinduced without basal lamina formation on someappropriate substrata. Zimmermann et al. (1985) con-cluded that a basement membrane is not necessary forlung cell differentiation in vitro. Our results areconsistent with theirs, but we have examined humancolon cancer cells morphologically only, and it ispossible that some biochemical defect in the compo-sition of the basement membrane results in the loss ofpolarity in HT29 cells in contact with mesenchyme.Further quantitative analysis is needed for full under-standing of the relationship of gland formation andchanges in the ultrastructure of intestinal epithelialcells.
This work was supported by grants-in-aid for CancerResearch from the Ministry of Education, Science andCulture, Japan, and by the Veterans Administration MedicalResearch Service, USA.
References
AUGERON, C. & LABOISSE, C. L. (1984). Emergence of
permanently differentiated cell clones in a human coloniccancer cell line in culture after treatment with sodiumbutyrate. Cancer Res. 44, 3961-3969.
BANERJEE, S. D., COHN, R. H. & BERNFIELD, M. R.
(1977). Basal lamina of embryonic salivary epithelia:Production by the epithelium and role in maintaininglobular morphology. J. Cell Biol. 73, 445-463.
BENNETT, D. C. (1980). Morphogenesis of branchingtubules in cultures of cloned mammary epithelial cells.Nature, Land. 285, 657-659.
BERNFIELD, M. R., BANERJEE, S. D. & COHN, R. H.
(1972). Dependence of salivary epithelial morphologyand branching morphogenesis upon acidmucopolysaccharide-protein (proteoglycan) at theepithelial surface. J . Cell Biol. 52, 674-689.
620 H. Fukamachi et al.

CHAMBARD, M., GABRION, J. & MAUCHAMP, J. (1981).
Influence of collagen gel on the orientation of epithelialcell polarity: Follicle formation from isolated thyroidcells and from preformed monolayers. J. Cell Biol. 91,157-166.
COHEN, E., TALMON, A., FAFF, 0 . , BACHER, A. &
BEN-SHAUL, Y. (1985). Formation of tight junctions inepithelial cells. I. Induction by proteases in a humancolon carcinoma cell line. Expl Cell Res. 156, 103-116.
COWIN, P. & GARROD, D. R. (1983). Antibodies to
epithelial desmosomes show wide tissue and speciescross-reactivity. Nature, Land. 302, 148-150.
CUNHA, G. R., BlGSBY, R. M., COOKE, P. S. & SUGIMURA,Y. (1985). Stromal-epithelial interactions in adultorgans. Cell Differ. 17, 137-148.
FUKAMACHI, H., MIZUNO, T. & KIM, Y. S. (1986).
Morphogenesis of human colon cancer cells with fetal ratmesenchymes in organ culture. Experientia 42, 312-315.
FUKAMACHI, H., MIZUNO, T. & TAKAYAMA, S. (1979).
Epithelial-mesenchymal interactions in differentiation ofstomach epithelium in fetal mice. Anat. Embryol. 157,151-160.
FUKAMACHI, H. & TAKAYAMA, S. (1980).
Epithelial-mesenchymal interaction in differentiation ofduodenal epithelium of fetal rats in organ culture.Experientia 36, 335-336.
GARBI, C , NITSCH, L. & WOLLMAN, S. H. (1984).
Embedding in a collagen gel stabilizes the polarity ofepithelial cells in thyroid follicles in suspension culture.Expl Cell Res. 151, 458-465.
GOLDIN, G. V. (1980). Towards a mechanism formorphogenesis in epithelio-mesenchymal organs. Q. Rev.Biol. 55, 251-265.
HAEUPTLE, M.-T., SUARD, Y. L. M., BOGENMANN, E.,
REGGIO, H., RACINE, L. & KRAEHENBUHL, J.-P. (1983).
Effect of cell shape change on the function anddifferentiation of rabbit mammary cells in culture, jf. CellBiol. 96, 1425-1434.
HAFFEN, K., SIMON-ASSMANN, P., KEDINGER, M.,
GRENIER, J. F. & ZWEIBAUM, A. (1981). Effet permissifdes cellules canceVeuses coliques humaines (HT-29 etCaco-2) sur la diffeVenciation du muscle lisse intestinal.C. r. hebd. Seanc Acad. Set., Paris 293, 807-812.
HALL, H. G., FARSON, D. A. & BISSELL, M. J. (1982).
Lumen formation by epithelial cell lines in response tocollagen overlay: A morphogenetic model in culture.Proc. naln. Acad. Sci. U.SA. 79, 4672-4676.
INGBER, D. E., MADRI, J. A. & JAMIESON, J. D. (1981).
Role of basal lamina in neoplastic disorganization oftissue architecture. Proc. natn. Acad. Sci. U.SA. 78,3901-3905.
LUCIANO, L., THIELE, J. & REALE, E. (1979).
Development of follicles and of occluding junctionsbetween the follicular cells of the thyroid gland. A thin-section and freeze-fracture study in the fetal rat.J. Ultrastruct. Res. 66, 164-181.
MADARA, J. L., NEUTRA, M. R. & TRIER, J. S. (1981).
Junctional complexes in fetal rat small intestine duringmorphogenesis. Devi Biol. 86, 170-178.
MONTESANO, R., FRIEND, D. S., PERRELET, A. & ORCI, L.
(1975). In vivo assembly of tight junctions in fetal ratliver. J. Cell Biol. 67, 310-319.
NAKANISHI, Y., SUGIURA, F., KISHI, J. & HAYAKAWA, T.
(1986a). Collagenase inhibitor stimulates cleft formationduring early morphogenesis of mouse salivary gland.Devi Biol. 113,201-206.
NAKANISHI, Y., SUGIURA, F., KISHI, J. & HAYAKAWA, T.
(19866). Local effects of implanted elvax chipscontaining collagenase inhibitor and bacterial collagenaseon branching morphogenesis of mouse embryonicsubmandibular glands in vitro. Zool. Sci. 3, 479-486.
NITSCH, L., TACCHETTI, C , TRAMONTANO, D. & AMBESI-
IMPIOMBATO, F. S. (1984). Suspension culture reveals amorphogenetic property of a thyroid epithelial cell line.Expl Cell Res. 152, 22-30.
NITSCH, L. & WOLLMAN, S. H. (1980). infrastructure of
intermediate stages in polarity reversal of thyroidepithelium in follicles in suspension culture. J. Cell Biol.86, 875-880.
REDDY, J. K., KANWAR, Y. S., RAO, M. S., WATANABE,
T. K., REDDY, M. K., PARSA, I., LONGNECKER, D. S.
& TAFURI, S. (1986). Duct-like morphogenesis ofLongnecker pancreatic acinar carcinoma cells maintainedin vitro on seminiferous tubular basement membranes.Cancer Res. 46, 347-354.
REMY, L., MARVALDI, J., RUA, S., SECCHI, J. & LECHENE
DE LA PORTE, P. (1984). The role of intracellular luminain the repolarization process of a colonic adenocarcinomacell line. Virchows Arch. Cell Path. 46, 297-305.
RIDGE, J. & NOGUCHI, P. D. (1983). Histotypic expressionof tumor architecture by human cell lines in chickembryonic tissue organ culture. In Vitro 19, 274.
RUTZKY, L. P., TOMITA, J. T., CALENOFF, M. A. &
KAHAN, B. D. (1979). Human colon adenocarcinomacells. III. In vitro organoid expression andcarcinoembryonic antigen kinetics in hollow fiberculture. J. natn. Cancer Inst. 63, 893-902.
SAWYER, R. H. & FALLON, J. F. (1983). Epithelial-
Mesenchymal Interactions in Development. New York:Praeger.
THOMPSON, H. A. & SPOONER, B. S. (1982). Inhibition of
branching morphogenesis and alteration ofglycosaminoglycan biosynthesis in salivary glands treatedwith |3-D-xyloside. Devi Biol. 89, 417-424.
VRACKO, R. (1974). Basal lamina scaffold. Anatomy andsignificance for maintenance of orderly tissue structure.Am. J. Path. 77, 314-346.
YAMADA, K. M. (1983). Cell Interactions andDevelopment. New York: John Wiley and Sons Inc.
ZlMMERMANN, B., BARRACH, H.-J., MERKER, H.-J. &HlNZ, N. (1985). Basement membrane formation andlung cell differentiation in vitro. Eur. J. Cell Biol. 36,66-73.
{Received 12 February 19S7 - Accepted I April I9S7)
Gland formation of colon cancer cells 621





















