
Optimizing Soluble Cues for Salivary Gland Tissue Mimetics ...

Amphibia-Reptilia 30 (2009): 561-569
The mental gland of Bolitoglossa nicefori (Caudata: Plethodontidae)
Ezequiel González-León, Martha Patricia Ramírez-Pinilla*
Abstract. The mental gland of the plethodontid salamander Bolitoglossa nicefori was examined by macroscopic observationand light microscopy and compared with morphological descriptions of other species of Plethodontidae. The mental gland isan oval shaped pad which develops exclusively in reproductive males. It is composed of 255 to 710 simple glands of merocrinesecretion arranged in a dorso-ventral disposition. The outlets of the glands are spindle-shaped or fusiform. Negative stainreaction for AB and PAS is consistent with results of the glandular components of mental glands from other Plethodontidae.Males of this species produce sperm continuously throughout the year: all examined adult males showed conspicuous mentalglands. However, an evident and significant increase in the surface area of the mental gland pad, in the density of the glands,and in the number of tubular glands was observed in the males collected during the breeding season. At the histological level,a slight variation was observed among males in the height and diameter of the simple tubular glands, and significant variationwas found in the diameter of the secretory granules. This variation may reflect the fact that, although males potentially canmate during all months of the year, females oviposit seasonally. During this defined season, the hypertrophy of the mentalgland is the greatest.
Keywords: Bolitoglossa nicefori, courtship, mental gland, Plethodontidae, salamander.
Introduction
In salamanders, chemical signals have a keyimportance in the courtship process. In manyspecies, the male produces courtship phero-mones from a specialized submandibular glandthat hypertrophies only during the mating sea-son (Weichert, 1945; Sever, 1975; Rollmann,Houck and Heldhoff, 1999, 2000, 2003; Houckand Arnold, 2003). Courtship pheromones canincrease female receptivity (Rollmann, Houckand Heldhoff, 1999, 2003; Houck and Arnold,2003; Rajchard, 2005), facilitate the continuityof courtship (Sever, 1976b; Rollmann, Houckand Heldhoff, 2003), reduce the courtship du-ration which in turn decreases the probabil-ity of disruption of mating by a rival maleor predators (Rollmann, Houck and Feldhoff,2003), and is probably involved in mate choice(Rajchard, 2005). The cluster of individualglands in the submandibular region of maleswas named the mental gland by Noble (1927).These glands occur widely in Plethodontidae
Laboratorio de Biología Reproductiva de Vertebrados,Escuela de Biología, Universidad Industrial de San-tander, Bucaramanga, Colombia*Corresponding author; e-mail: [email protected]
(Sever, 1976b, 2003), and functionally simi-lar glands have been reported in males of onespecies in the Salamandridae (Sever, 2003). Ac-cording to Sever (1976b), many authors recog-nized that some intergeneric variation existedin the morphology of mental glands. The men-tal glands in Plethodontidae vary from round,oval, or broadened posteriorly, to elongate andbifurcate (Valentine, 1963; Sever, 1975; Doddand Brodie, 1976). Generally, the mental glandconsists of either (a) groups of short, dorso-ventrally oriented tubules that secrete over asmall-to-large area (Sever, 2003) via excre-tory ducts in the epidermis of the mental re-gion (Testa Riva et al., 1993); or (b) a fan-shaped cluster of short or long tubules, antero-posteriorly oriented that secrete at the apex ofthe lower jaw (Sever, 2003). The mental glandis hypertrophied during the breeding season andcan be used as a sexually dimorphic characterto distinguish between the sexes (Noble, 1929;Weichert, 1945).
Bolitoglossa nicefori (San Gil Mushroom-tongue Salamander) is a Neotropical and en-demic species from the Colombian CordilleraOriental. This species is found in disturbed hu-mid forests and in coffee plantations, among
© Koninklijke Brill NV, Leiden, 2009. Also available online - www.brill.nl/amre

562 E. González-León, M.P. Ramírez-Pinilla
fallen leaves, herbaceous vegetation and epi-phytes (Ramírez-Pinilla et al., 2004). The analy-sis of the annual reproductive activity of a popu-lation of this species showed a distinct breed-ing season, defined as the time when femalesoviposited. Histological analyses of testes andexcurrent ducts showed the presence of freespermatozoa through the year, suggesting con-tinuous reproductive activity for males. How-ever, there was a significant variation in themonthly behavior of testis volume. Maximaltesticular volume was observed by Decemberwhen the females are periovulatory, and testisvolume fell during the following months per-haps in response to the increased mating oppor-tunities because matings occur around the timeof egg-laying (Ortega, Monares and Ramírez-Pinilla, 2009). In a parallel study on femalecloacal histology of this same population of B.nicefori we have found that females only havesperm stored in their spermathecae during thebreeding season, suggesting that males and fe-males mate during this time. Non-reproductivefemales had no cloacal or stored sperm in theirspermathecae, which suggests that they becamenon-receptive to males following oviposition(González-León and Ramírez Pinilla, unpub-lished). The objective of this study was to de-scribe and compare the anatomy and histol-ogy of the mental glands in juveniles and adultmales of this species to allow intra- and inter-specific comparisons.
Materials and methods
Male Bolitoglossa nicefori collected in different breedingseasons were obtained from the Herpetological Collectionof the Museo de Historia Natural, Universidad Industrialde Santander. These salamanders were collected from anorganic coffee plantation located in Hacienda El Roble(06◦52′N, 73◦03′W, 1640 m altitude) or from a disturbedforest at Vereda Las Amarillas, Microcuenca La Venta(06◦58′N, 73◦01′W, 1400-2000 m altitude), Municipalityof Piedecuesta, Santander, Colombia. Individuals were col-lected for past studies of reproduction, diet and micro-habitat use (Ortega, Monares and Ramírez-Pinilla, 2009),cranial development (Dulcey Cala, Tarazona and Ramírez-Pinilla, 2009), and appendicular development (Cabrera etal., unpublished data), under collecting and research permits
granted by the local environmental authority (CorporaciónAmbiental para la Defensa de la Meseta de Bucaramanga).
We defined breeding season as the time when the vitel-logenic process has reached the maximum follicular size(Ortega, Monares and Ramírez-Pinilla, 2009) and femalespermathecae have evident stored sperm (González-Leònand Ramírez-Pinilla, unpublished data). According to Or-tega, Monares and Ramírez-Pinilla (2009), adult males ofthis species have sperm present in their testes and ductsthroughout the year; therefore for this study maturity wasascertained by the above characteristics. Males were sepa-rated into two groups: males collected during the breedingseason (November-February) and males collected during thenon-breeding season (March-September). We examined themental glands of six males collected in the breeding season,five males collected in the non-breeding season, and two ju-venile males.
Snout-vent length (SVL) was measured from the tip ofthe snout to the posterior edge of the cloacal orifice. Themental gland of each male was dissected and fixed in 10%buffered formalin for 8 hours, dehydrated in a series ofethanol and xylene, embedded in paraffin, serially sectionedat 7 to 10 μm with a rotary microtome. Some sectionsfrom each individual were stained with hematoxylin-eosin(general histology) and other sections were submitted tothe combined histochemical methods of Alcian blue (AB)pH 2.5 and periodic acid Schiff (PAS) for detection of acidand neutral glycoproteins respectively, according to the pro-tocol of Luna (1968) as follows: (1) deparaffinization andhydration of sections through a series of graded alcohol todistilled water; (2) staining with Alcian Blue Solution (Al-cian Blue, pH 2.5) for 30 minutes; (3) washing in waterfor 5 minutes; (4) oxidization in Periodic Acid, 1%, for10 minutes; (5) washing in running water for 5 minutes;(6) staining with Schiff’s Reagent, for 10 minutes; (7) rins-ing in Sodium Metabisulfite, 0.5% (aqueous, three changes,two minutes each); (8) washing in running water for tenminutes; (9) dehydratation in 95% Alcohol, Absolute Al-cohol and clear in Xylene, two changes each; and finally(10) Mounting with Entellan® media. By using this method,all polysaccharides and mucosubstances containing hexosesor deoxyhexoses with glycol groups and neutral mucosub-stances stain magenta to red. Hyaluronic acid, sialomucinsand all but the most strongly acidic sulfated mucosubstancesstain blue.
For each mental gland, we measured the length and widthmacroscopically; we calculated the area of each mentalgland pad using the formula of the surface area of a general“flat” ellipsoid. To establish the number of simple glandsconstituting the mental gland pad of each male we countedthe number of openings of the simple tubular glands ina specific area under a stereo-microscope and then werelated this result to the total pad area. The relationshipbetween the size of the mental gland pad and the malebody size (SVL) was examined with a linear regression.We used a microscope to measure the height and diameterof the simple glands (middle minor axis), diameter of thesecretory granules, and length and width of the ducts ofthe simple glands in their medial region. For each of theseparameters we measured at least 20 simple glands (n � 20)

The mental gland of Bolitoglossa nicefori 563
of different sections or profiles (n � 12). We used at-test to compare the difference between means with dataobtained for diameter and height of the simple tubularglands and the secretory granules from males collected indifferent breeding seasons. Additionally, testes were alsodissected, fixed in 10% buffered formalin, embedded inparaffin, serially sectioned at 10 μm, and stained withhematoxylin-eosin to determine the reproductive conditionof each male.
Results
Adult males collected during the breeding sea-son were similar in body size to adult males ofthe non-breeding season (t = −2.25, df = 9,P = 0.051). All the adult males had a con-spicuous and hypertrophied mental gland pad;however, the area of this pad was not signifi-cantly related to SVL (r2 = 0.032, F1,9 = 1.34,P = 0.28). The mental pad had a significantlyincreased area in males of the breeding season(mean ± SD = 8.8 ± 1.76 mm2) comparedto males collected during the non-breeding sea-son (5.1 ± 0.22 mm2, t = 4.6, df = 9, P =0.0015). The juvenile males (SVL > 40 mm)did not have any evident indication of a mentalgland in the submandibular region.
Anatomy
The mental gland exhibits an oval form incross-sections (fig. 1) that changes in shapefrom 1.82 mm long and 3.58 mm wide (malesfrom non-breeding season) to 2.82 mm longand 3.94 mm wide (males from breeding sea-
Figure 1. Pad-shaped mental gland of adult male Boli-toglossa nicefori. A-B – Note the oval pad-shape from (A)an outer view and (B) an inner view. In B, the presence ofmelanocytes is indicated by arrowheads. Ep, epidermis; Mg,mental gland. Scale bar = 2 mm for A; 1 mm for B.
son). The mental gland consists of many sim-ple tubular glands with dorso-ventral orienta-tion. The entire surface of the mental glandpad is secretory because each simple tubu-lar gland open in a simple tubule. The den-sity of the glands (number of gland open-ings per unit area) was significantly differ-ent in males from the breeding season and inmales from non-breeding season (60 ± 6.25ducts/mm2 and 51.51 ± 4.12 ducts/mm2, re-spectively; t = 17.55, df = 9, P = 0.015). Thenumber of tubules per male ranged from ap-proximately 255-386 in males from the non-breeding season to 318-710 tubules in malesfrom breeding season (fig. 2A, B).
Histology
The submandibular region of the juvenile malesonly has common epidermal granular glandsand no indication of any development of themental gland (fig. 3). Among reproductivemales (from both the breeding and non-breedingseasons), the simple tubular glands had a vari-able diameter and height (table 1). These sim-ple tubular glands were composed of columnarglandular cells with basophilic basal nuclei anda granular eosinophilic apex directed towardsthe lumen of the gland (figs 2, 4A). Surround-ing the simple tubular glands was a small layerof highly eosinophilic connective tissue. Severalmelanocytes surrounded the mental gland at dif-ferent levels (figs 1B, 2B, C).
Simple tubular glands opened individuallyat the bottom of the lower jaw, through smallspindle-shaped or fusiform ducts (fig. 2B, C). Inthe medial region, these ducts were 40 ± 7 μmlong and 8.7±3.7 μm wide (fig. 4B). Each ductconsisted of two cells. In turn, four epithelialcells surround these duct cells in a particulardisposition: two cells located longitudinally tothe major axis of the duct, and other two cellslocated perpendicular to this axis (fig. 4B).
Each simple tubular gland had a merocrinesecretion, and the secretory apical granuleswere liberated into the lumen of the gland asyn-chronously (fig. 4A). The apical granules were

564 E. González-León, M.P. Ramírez-Pinilla
Figure 2. Sagittal histological sections of the mental gland of a male B. nicefori collected during the breeding season. Sectionswere stained with hematoxylin-eosin (H-E). A – Note the transition between the simple tubular glands of the mental glandand the typical skin glands surrounding the mental gland. B – The simple tubular glands open in ducts at the bottom of thelower jaw (arrowhead). C – Detail of B showing a duct that opens in the submandibular region (arrowhead) through theepidermis; connective tissue surrounds each gland (arrow), and melanocytes are located between the simple tubular glandsand the epidermis. Cts, connective tissue; Ep, epidermis; Tg, simple tubular gland. Scale bar = 100 μm for A and B; 50 μmfor C.
Figure 3. Sagittal histological section of the submandibularregion of a juvenile male of Bolitoglossa nicefori showingthat only granular glands are observed. Sections stained withhematoxylin-eosin (H-E). Melanocytes are indicated by thearrows. Cts, connective tissue; Ep, epidermis; Gg, granulargland. Scale bar = 40 μm.
round or oval with variable diameters in malesfrom different reproductive seasons (table 1).One male (SVL 44.52 mm) collected duringthe last month of the breeding season (mid-February), had few secretory granules. More-over, these granules had the lowest mean size(diameter 2.3 mm) among all males collectedduring the breeding season. Staining reactionsto AB and PAS were negative in all cases for theglandular content of tubules. However, the nu-clei of secretory cells and connective tissue sur-rounding the mental gland showed, respectively,a moderate and weak staining for PAS (fig. 5).Only the diameter of the granules showed astatistically significant difference (t = −1.86,df = 9, P < 0.05) between males from the

The mental gland of Bolitoglossa nicefori 565
Table 1. Measurements of mental gland parameters in adult male Bolitoglossa nicefori. These males are reproductivethroughout the year and were separated in two groups: males collected during the breeding season (November-February),and males collected in the non-breeding season (March-September).
N Simple tubular gland diameter Simple tubular gland height Granule diameter
Range Mean SD Range Mean SD Range Mean SD
Males frombreeding season 6 121.2-152.4 137.5 12.48 225.6-497.5 353.7 96.12 2.3-5.3 4.1 0.94
Males fromnon-breeding season 5 123.3-147 135.6 9.43 213.8-375.0 295.3 56.98 2.2-3.9 3.0 0.72
Figure 4. Transverse sections of the mental gland of an adult male of Bolitoglossa nicefori from the non-breeding season.Sections stained with hematoxylin-eosin (H-E). A – Simple tubular glands (Tg) compose the mental gland. A comparison oftubules reveals the asynchronous merocrine secretion of eosinophilic secretory granules. Some simple tubular glands presenta lumen devoid of secretion as these glandular cells already have discharged their secretory granules (arrowhead). In othersimple tubular glands, secretory granules obscure the lumen (Asterisk). B – Detail of the region showing the ducts (arrowhead)of the simple simple tubular glands. Note the special form of the ducts (arrowhead) and the particular disposition of the cellsthat surround each duct. Tg, simple tubular gland. Scale bar = 100 μm for A; 20 μm for B.
breeding and non-breading seasons. The otherparameters (diameter and height of the sim-ple tubular glands) were not statistically differ-ent among males from the breeding and non-breading seasons (t = −0.24, df = 9, P =0.81, and t = −0.97, df = 9, P = 0.35, respec-tively).
The testes of the juvenile males had severalcysts in different stages of spermatogenesis.However, cysts of early spermatids were thelatest stage found and the excurrent ducts were
empty. In the testes of adult males collectedduring breeding and non-breeding seasons, weobserved cysts of all the different stages ofspermatogenesis and spermiogenesis, includingmature sperm at the lumen of the seminiferoustubules and in the collecting ducts.
Discussion
The anatomy and histology of the mental glandof Bolitoglossa nicefori is similar to that de-

566 E. González-León, M.P. Ramírez-Pinilla
Figure 5. Sagittal histological section of the mental glandof a male B. nicefori collected during the breeding season.Sections were stained by AB/PAS method. In the tubularglands (Tg), the secretory granules are negative to ABand PAS reactions. The nuclei of secretory cells (arrows),epithelium (Ep) and connective tissue (Cts) are moderatlyto weakly positive to PAS. Scale bar = 100 μm.
scribed for the plethodontid genus Bolitoglossaby Truffelli (1954). Sever (2003) expandedupon Trufelli’s description by proposing fivetypes of glands within the Plethodontidae. Severbased his descriptions on the different glandshapes: (1) pad-shaped, relatively large andoval, in Plethodon and Bolitoglossa; (2) pad-shaped, relatively small, in Hemidactylium andEurycea; (3) anterior protrusion, in Desmog-nathus; (4) elongate and bifurcated, in Desmog-nathus wrighti; (5) fan-shaped and elongated, inEurycea and various Bolitoglossine genera. Ac-cording to this classification, the mental glandof B. nicefori is of the first type (a relativelylarge, oval pad), which fits with Sever’s char-acterization of glands in Bolitoglossa species.
In plethodontid salamanders with type onemental glands, the average size of these glandshave been measured for 12 species (table 2).Among these species, B. nicefori males col-
lected during the breeding season have a mentalgland of intermediate size. In the genus Pletho-don, P. glutinosus and P. jordani have cylindri-cal glands, whereas in Eurycea, these glands aretubular or elliptical (Sever, 1976b). The diffe-rence in size, number, and shape of the simpletubular glands in males from the breeding andnon-breeding seasons of B. nicefori probably isrelated to the gland diameters (fig. 4A). Similarconsequences of differences in gland diametersalso were observed by Truffelli (1954) in Tho-rius, Pseudoeurycea, and some species of Boli-toglossa that have glands with a markedly non-uniform diameter.
The characteristic spindle-shaped or fusiformnature of the simple tubular gland ducts in themental gland of B. nicefori is similar to thatdescribed by Testa Riva et al. (1993) for Hy-dromantes genei, a European plethodontid (Sar-dinia, Italy). The ducts present an appearanceof elliptical stomata lined by one or two cells.Other studies on the morphology of the mentalgland only show longitudinal sections, so littleis known regarding the form and cell position ofthese ducts.
The mental gland has been described as aprominent feature in reproductive males (Sever,1976b). Rollmann, Houck and Heldhoff (2003)report for P. shermani that glands hypertro-phy only during breeding season. In contrast,all adult B. nicefori have conspicuous mentalglands throughout the year, without significantdifferences in the diameter and height of thesimple tubular glands in males from breedingand non-breading seasons. Mental gland devel-opment may be a response to the secretion oftestosterone evidenced in the continuously ac-tive testes of B. nicefori. This response wassuggested for Eurycea quadridigitata, in whichtestosterone aids in the maintenance of spermin vas deferens; in turn, males show little sea-sonal variation in the presence and hypertro-phy of the mental gland (Sever, 1975). In adifferent study, Sever (1976a) injected femaleE. quadridigitata with testosterone solution andthe chin area eventually was stimulated to hy-
The mental gland of Bolitoglossa nicefori 567
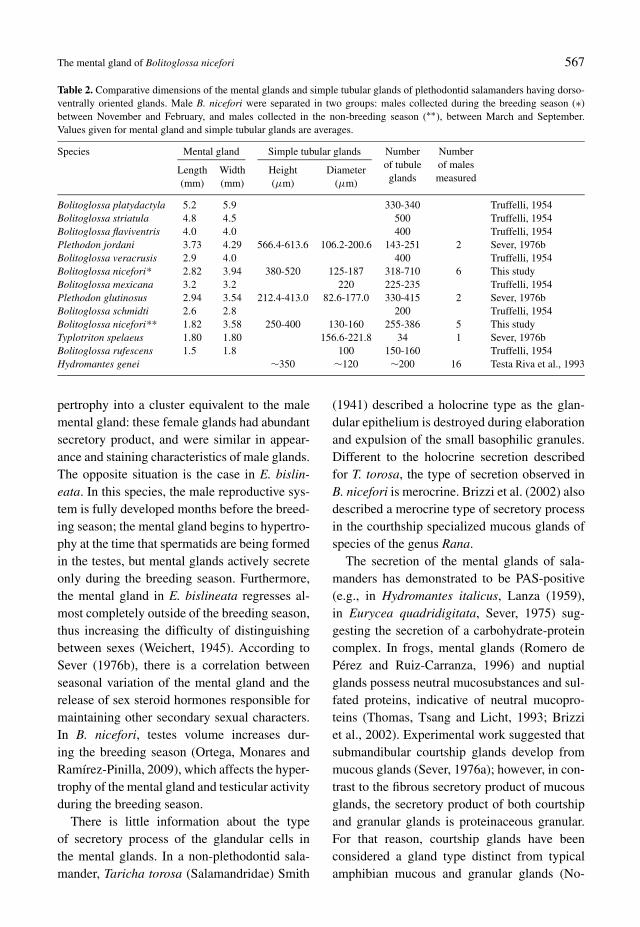
Table 2. Comparative dimensions of the mental glands and simple tubular glands of plethodontid salamanders having dorso-ventrally oriented glands. Male B. nicefori were separated in two groups: males collected during the breeding season (∗)between November and February, and males collected in the non-breeding season (∗∗), between March and September.Values given for mental gland and simple tubular glands are averages.
Species Mental gland Simple tubular glands Numberof tubuleglands
Numberof malesmeasured
Length Width Height Diameter(mm) (mm) (μm) (μm)
Bolitoglossa platydactyla 5.2 5.9 330-340 Truffelli, 1954Bolitoglossa striatula 4.8 4.5 500 Truffelli, 1954Bolitoglossa flaviventris 4.0 4.0 400 Truffelli, 1954Plethodon jordani 3.73 4.29 566.4-613.6 106.2-200.6 143-251 2 Sever, 1976bBolitoglossa veracrusis 2.9 4.0 400 Truffelli, 1954Bolitoglossa nicefori* 2.82 3.94 380-520 125-187 318-710 6 This studyBolitoglossa mexicana 3.2 3.2 220 225-235 Truffelli, 1954Plethodon glutinosus 2.94 3.54 212.4-413.0 82.6-177.0 330-415 2 Sever, 1976bBolitoglossa schmidti 2.6 2.8 200 Truffelli, 1954Bolitoglossa nicefori** 1.82 3.58 250-400 130-160 255-386 5 This studyTyplotriton spelaeus 1.80 1.80 156.6-221.8 34 1 Sever, 1976bBolitoglossa rufescens 1.5 1.8 100 150-160 Truffelli, 1954Hydromantes genei ∼350 ∼120 ∼200 16 Testa Riva et al., 1993
pertrophy into a cluster equivalent to the malemental gland: these female glands had abundantsecretory product, and were similar in appear-ance and staining characteristics of male glands.The opposite situation is the case in E. bislin-eata. In this species, the male reproductive sys-tem is fully developed months before the breed-ing season; the mental gland begins to hypertro-phy at the time that spermatids are being formedin the testes, but mental glands actively secreteonly during the breeding season. Furthermore,the mental gland in E. bislineata regresses al-most completely outside of the breeding season,thus increasing the difficulty of distinguishingbetween sexes (Weichert, 1945). According toSever (1976b), there is a correlation betweenseasonal variation of the mental gland and therelease of sex steroid hormones responsible formaintaining other secondary sexual characters.In B. nicefori, testes volume increases dur-ing the breeding season (Ortega, Monares andRamírez-Pinilla, 2009), which affects the hyper-trophy of the mental gland and testicular activityduring the breeding season.
There is little information about the typeof secretory process of the glandular cells inthe mental glands. In a non-plethodontid sala-mander, Taricha torosa (Salamandridae) Smith
(1941) described a holocrine type as the glan-dular epithelium is destroyed during elaborationand expulsion of the small basophilic granules.Different to the holocrine secretion describedfor T. torosa, the type of secretion observed inB. nicefori is merocrine. Brizzi et al. (2002) alsodescribed a merocrine type of secretory processin the courthship specialized mucous glands ofspecies of the genus Rana.
The secretion of the mental glands of sala-manders has demonstrated to be PAS-positive(e.g., in Hydromantes italicus, Lanza (1959),in Eurycea quadridigitata, Sever, 1975) sug-gesting the secretion of a carbohydrate-proteincomplex. In frogs, mental glands (Romero dePérez and Ruiz-Carranza, 1996) and nuptialglands possess neutral mucosubstances and sul-fated proteins, indicative of neutral mucopro-teins (Thomas, Tsang and Licht, 1993; Brizziet al., 2002). Experimental work suggested thatsubmandibular courtship glands develop frommucous glands (Sever, 1976a); however, in con-trast to the fibrous secretory product of mucousglands, the secretory product of both courtshipand granular glands is proteinaceous granular.For that reason, courtship glands have beenconsidered a gland type distinct from typicalamphibian mucous and granular glands (No-

568 E. González-León, M.P. Ramírez-Pinilla
ble, 1929; Thomas, Tsang and Licth, 1993;Brizzi, Delfino and Pellegrini, 2002). Althoughthe method AB-PAS did not clearly detect gly-coproteinaceous substances in the mental glandof B. nicefori, this not means that these glandsnot secrete some neutral carbohydrates. In fact,Brizzi, Delfino and Pellegrini (2002) also foundspecialized mucous courtship glands in Ranaiberica that fails to react with this method. Itis possible that the histochemical tests reveala mainly proteinaceous content bounded witha very scarce amount of neutral carbohydratesthat are interpreted as negative or slightly PASpositive by this method.
Strongly eosinophilic secretory granuleswere observed filling the apex of most of glan-dular cells in adult males from both breed-ing and non-breeding seasons. This result in B.nicefori is consistent with results from Roll-mann, Houck and Heldhoff (1999, 2000) forPlethodon salamanders: glandular secretions ofthe mental gland include protein componentssuch as PRF (plethodontid receptivity factor),a pheromone that has multiple isoforms. Inmales, variation among isoforms and in the rel-ative proportions of these isoforms may varytheir ability to stimulate females (Rollmann,Houck and Heldhoff, 2000). It is possible thata seasonal variation exists in the chemical com-position of the secreting products of the men-tal gland of B. nicefori allowing courtship of re-productive females during the breeding season.This possibility assumes that females are morelikely to mate during the period when ovipo-sition occurs as has been suggested for otherspecies of neotropical plethodontids (Houck,1977; Chan, 2003).
For plethodontid salamanders Houck et al.(1998) found that courtship pheromones func-tion to increase female receptivity, thus in-creasing the probability that the female willbe inseminated. In fact, the male delivers thepheromone by bringing his mental gland indirect contact with the female (Houck et al.,2007). Pheromones are absorbed by the fe-male skin circulatory system and, then, stimu-
late female receptivity. For temperate plethod-ontids, Rollmann, Houck and Heldfhoff (1999)suggested that maximal receptivity, insemina-tion, and ovulation are not closely linked be-cause oviposition typically occurs weeks oreven months after pheromone delivery. Thus,the pheromone-induced change in receptivity isthe only known function of PRF. For neotrop-ical plethodontids it is not possible to general-ize a prolonged period between the pheromonerelease/mating and the effective storage ofsperm/oviposition. For example for B. rostrataand B. occidentalis Chan (2003) suggestedthat a greater proportion of matings take placearound the time of egg-laying. Houck (1977)did not find sperm stored in the spermathecaof most of the females of B. rostrata withvitellogenic follicles with diameters higher than2.0 mm, as we observed in B. nicefori fe-males: sperm was observed in the spermath-eca of those females with maximum follicu-lar diameters close to ovulation. These obser-vations suggest that in these species there isnot a decoupling of mating and ovulation/egg-laying as observed in temperate plethodontidsalamanders. Houck et al. (1998) observed thatcourtship pheromones are delivered only after amate has been located and after initial courtshipinteractions have commenced. Thus, it is possi-ble to suggest that pheromone liberation occursduring the breeding season.
Mental gland development seems to be amale secondary sexual feature related to hor-monal control. Variations in the size of thegland (pad area, density and number of simpletubular glands, and diameter of secretory gran-ules) differed between males from the breed-ing season (i.e., when mating and ovipositionoccurred) and the non-breeding seasons in B.nicefori. This seasonal difference indicates that,although the males are producing spermatozoaevery month of the year and their mental glandsare active continuously, the gland hypertrophyis maximal during the season when females areovipositing. Subsequent studies may determinethe content of pheromones in the mental gland

The mental gland of Bolitoglossa nicefori 569
of B. nicefori, and their interactions related tothe timing of ovipositing in the females.
Acknowledgements. We thank the Colección Herpetológ-ica of the Museo de Historia Natural, Universidad Indus-trial de Santander for the loan of the Bolitoglossa niceforispecimens and the Laboratorio de Biología Reproductiva deVertebrados, Universidad Industrial de Santander for the in-frastructure and materials provided for the realization of thiswork. We thank J.C. Afanador, C. Dulcey and L. Cabrera fortheir help in the field; and F. Leal for their help with the his-tological techniques and critical reading of the manuscript.We thank to L. Houck and two other anonymous reviewerswho offered constructive and insightful comments on themanuscript. We especially thank to L. Houck who also pro-vided a detailed language review.
References
Brizzi, R., Delfino, G., Pellegrini, R. (2002): Specializedmucous glands and their possible adaptive role in themales of some species of Rana (Amphibia, Anura). J.Morphol. 254: 328-341.
Chan, L.M. (2003): Seasonality, microhabitat and crypticvariation in tropical salamander reproductive cycles.Biol. J. Linn. Soc. 78: 489-496.
Dodd, C.K., Brodie, E.D. (1976): Observations on the men-tal hedonic gland-cluster of eastern salamanders of thegenus Plethodon. Chesap. Sci. 17: 129-131.
Dulcey, C.J., Tarazona, O.A., Ramírez-Pinilla, M.P. (2009):Morphology and postnatal development of the skull ofBolitoglossa nicefori (Caudata: Plethodontidae). Zool-ogy 112: 227-239.
Houck, L.D. (1977): Reproductive biology of a neotropicalsalamander, Bolitoglossa rostrata. Copeia 1977: 70-82.
Houck, L.D., Arnold, S.J. (2003): Courtship and MatingBehavior. In: Reproductive Biology and Phylogeny ofUrodela, p. 383-424. Sever, D.M., Ed., Enfield, NH,Science Publisher.
Houck, L.D., Bell, A.M., Reagan-Wallin, N.L., Feldhoff,R.C. (1998): Effects of experimental delivery of malecourtship pheromones on the timing of courtship ina terrestrial salamander, Plethodon jordani (Caudata:Plethodontidae). Copeia 1998: 214-219.
Houck, L.D., Palmer, C.A., Watts, R.A., Arnold, S.J., Feld-hoff, P.W., Feldhoff, R.C. (2007): A new vertebratecourtship pheromone, PMF, affects female receptivity ina terrestrial salamander. Anim. Behav. 73: 315-320.
Lanza, B. (1959): I1 corpo ghiandolare mentoniero dei“Plethodontidae” (Amphibia, Caudata). Monit. Zool.Ital. 67: 15-53.
Luna, L.G. (1968): Manual of Histologic Staining Methodsof the Armed Forces Intitute of the Pathology. 3rd Edi-tion. New York, McGraw-Hill.
Noble, G.K. (1927): The plethodontid salamanders; someaspects of their evolution. Am. Mus. Nov. 249: 1-26.
Noble, G.K. (1929): The relation of courtship to the sec-ondary sexual characters of the two-lined salamander,Eurycea bislineata (Green). Am. Mus. Nov. 362: 1-5.
Ortega, J.E., Monares, J.M., Ramírez-Pinilla, M.P. (2009):Reproductive activity, diet and microhabitat use in Boli-toglossa nicefori (Caudata: Plethodontidae). J. Herpetol.43: 1-10.
Rajchard, J. (2005): Sex pheromones in amphibians: a re-view. Vet. Med-Czech. 50: 385-389.
Ramírez-Pinilla, M.P., Osorno-Muñoz, M., Rueda, J.V.,Amézquita, A., Ardila-Robayo, M.C. (2004): Boli-toglossa nicefori. IUCN, Conservation International,and NatureServe 2006. Global Amphibian Assessment.www.globalamphibians.org. Downloaded on 30 May2008.
Rollmann, S.M., Houck, L.D., Feldhoff, R.C. (1999): Pro-teinaceous pheromone affecting female receptivity in aterrestrial salamander. Science 285: 1907-1909.
Rollmann, S.M., Houck, L.D., Feldhoff, R.C. (2000): Popu-lation variation in salamander courtship pheromones. J.Chem. Ecol. 26: 2713-2724.
Rollmann, S.M., Houck, L.D., Feldhoff, R.C. (2003): Cone-specific and heterospecific pheromone effects on femalereceptivity. Anim. Behav. 66: 857-861.
Romero de Pérez, G., Ruíz-Carranza, P.M. (1996): His-tología, histoquímica y estructura fina de la glándulamentoniana de dos especies de Hyla (grupo bogoten-sis) y del antebrazo de Phrynopus adenobatrachius. Rev.Acad. Col. Cien. Exact. Fís. Nat. 20: 575-584.
Sever, D.M. (1975): Morphology and seasonal variation ofthe mental hedonic glands of the Dwarf Salamander,Eurycea quadridigitata (Holbrook). Herpetologica 31:241-251.
Sever, D.M. (1976a): Induction of secondary sexual charac-ters in Eurycea quadridigitata. Copeia 1976: 830-833.
Sever, D.M. (1976b): Morphology of the mental hedonicgland clusters of plethodontid salamanders (Amphibia,Urodela, Plethodontidae). J. Herpetol. 10: 227-239.
Sever, D.M. (2003): Courtship and Mating Glands. In: Re-productive Biology and Phylogeny of Urodela, p. 383-424. Sever, D.M., Ed., Enfield, NH, Science Publisher.
Smith, R.E. (1941): Mating behavior in Triturus torosus andrelated Newts. Copeia 1941: 255-262.
Testa Riva, R.F., Serra, G.P., Loffredo, F., Riva, A. (1993):Ultrastructural study of the mental body of Hydro-mantes genei (Amphibia: Plethodontidae). J. Morphol.217: 75-86.
Thomas, E.O., Tsang, L., Licht, P. (1993): Comparativehistochemistry of the sexually dimorphic skin glands ofanuran amphibians. Copeia 1993: 133-143.
Truffelli, G.T. (1954): A macroscopic and microscopicstudy of the mental hedonic gland-clusters of someplethodontid salamanders. U. Kansas Sci. Bull. 36: 1-39.
Valentine, B.D. (1963): The mental gland of the salamanderDesmognathus wrighti King. Ohio J. Sci. 63: 25-26.
Weichert, C.K. (1945): Seasonal variation in the mentalgland and reproductive organs of the male Euryceabislineata. Copeia 1945: 78-84.
Received: October 5, 2008. Accepted: May 22, 2009.
Copyright © 2022 FDOKUMEN




















