
Ultrastructural study of the mouse antimesometrial decidua
12
Anat Embryol (1983) 166:263 274 Anatomy and Embryology Springer-Verlag I983 Ultrastructural Study of the Mouse Antimesometrial Decidua Paulo A. Abrahamsohn Department of Histology, Institute of Biomedical Sciences, University of S~o Paulo, Caixa Postal 4365, 01000 Sao Paulo, Brazil Summary. The ultrastructure of mouse antimesometrial decidual cells was analyzed during the development of the decidua between days 5 and 8 of pregnancy. The first decidual cells, appearing on the 5th day, are polygonal with rounded nuclei and prominent nucleoli; free ribo- somes predominate in the cytoplasm. On the 6th to the 8th days the cytoplasm of these cells is typically that of cells actively engaged in macromolecular synthesis. Large numbers of granular and agranular endoplasmic reticulum cisternae are present in addition to well-devel- oped Golgi complexes, mitochondria and lysosomes. Many bundles of microfilaments and lipid droplets occur during this period. An intense accumulation of autophagosomes and lysosomes with very heteroge- neous content was noted on the 7th and especially the 8th days. The presence of these organelles is an indication that involution of this part of the decidua has begun. Key words: Decidua --Ultrastructure - Mouse Introduction The establishment of the decidua is one of the most obvious modifications seen in the uterus of rodents during embryo implantation. In the mouse, typical decidual cells may be identified from the 5th day of pregnancy (d.p.) on, while the blastocyst reaches the attachment stage of implantation (Reinius 1967; Potts 1968; Nilsson 1970). The first decidual cells surround the antimesometrial extremity of the uterine epithelial crypts containing each embryo (Krehbiel 1937). Decidualization proceeds as further decidual cells accumulate by means of transformation of spindle-shaped endometrial stromal cells. The population of decidual cells in the antimesometrial wall Offprint requests to: Dr. P.A. Abrahamsohn, Department of Histology, Institute of Biomedical Sciences, University of S. Paulo, Caixa Postal 4365, 01000 S. Paulo, SP, Brasil
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Ultrastructural study of the mouse antimesometrial decidua

Anat Embryol (1983) 166:263 274 Anatomy and Embryology �9 Springer-Verlag I983
Ultrastructural Study of the Mouse Antimesometrial Decidua
Paulo A. Abrahamsohn Department of Histology, Institute of Biomedical Sciences, University of S~o Paulo, Caixa Postal 4365, 01000 Sao Paulo, Brazil
Summary. The ultrastructure of mouse antimesometrial decidual cells was analyzed during the development of the decidua between days 5 and 8 of pregnancy. The first decidual cells, appearing on the 5th day, are polygonal with rounded nuclei and prominent nucleoli; free ribo- somes predominate in the cytoplasm. On the 6th to the 8th days the cytoplasm of these cells is typically that of cells actively engaged in macromolecular synthesis. Large numbers of granular and agranular endoplasmic reticulum cisternae are present in addition to well-devel- oped Golgi complexes, mitochondria and lysosomes. Many bundles of microfilaments and lipid droplets occur during this period. An intense accumulation of autophagosomes and lysosomes with very heteroge- neous content was noted on the 7th and especially the 8th days. The presence of these organelles is an indication that involution of this part of the decidua has begun.
Key words: Decidua --Ult ras t ructure - Mouse
Introduction
The establishment of the decidua is one of the most obvious modifications seen in the uterus of rodents during embryo implantation. In the mouse, typical decidual cells may be identified from the 5th day of pregnancy (d.p.) on, while the blastocyst reaches the attachment stage of implantation (Reinius 1967; Potts 1968; Nilsson 1970). The first decidual cells surround the antimesometrial extremity of the uterine epithelial crypts containing each embryo (Krehbiel 1937). Decidualization proceeds as further decidual cells accumulate by means of transformation of spindle-shaped endometrial stromal cells. The population of decidual cells in the antimesometrial wall
Offprint requests to: Dr. P.A. Abrahamsohn, Department of Histology, Institute of Biomedical Sciences, University of S. Paulo, Caixa Postal 4365, 01000 S. Paulo, SP, Brasil

264 P.A. Abrahamsohn
of the endometrium constitutes the antimesometrial decidua (Snell 1941). The fully-developed decidual cells are large, polygonal, densely packed and have an epithelioid character (Krehbiel 1937). The initial steps of transfor- mation of rat stromal cells following decidual induction have been analyzed by electron microscopy (Lundkvist and Ljunkvist 1977; Lundkvist 1978). The decidual cells of the rat were described by Enders and Schlafke (1967) and Tachi et al. (1970) in the course of studies on embryo implantation. However, few reports have dealt specifically with the ultrastructure of the fully-developed decidua (rat: Jollie and Bencosme 1965; hamster: Orsini et al. 1970; Parkening 1976).
This paper is part of a systematic study on the development and involu- tion of the mouse antimesometrial decidua. The present work was under- taken to study the ultrastructural features of decidual cells between the 5th and 8th gestational days, a period comprising formation and maximal development of the antimesometrial decidua.
Material and Methods
Three month old, random-bred Swiss mice were used. Virgin females were mated overnight and were examined the following morning for the presence of a vaginal plug. The day on which a plug was found was designated day 1 of pregnancy (d.p.).
Animals from 5 to 8 days pregnant were used for ultrastructural analysis (three to four animals for each day). Three virgin animals in oestrus were also used. Fifteen minutes before sacrificing the animals, 0.15 ml of a solution of 0.5% Evans blue in 0.9% NaC1 was injected into each animal's tail vein in order to detect implantation sites (Psychoyos 1960). Decidua- bearing uteri of mice are usually difficult to fix adequately. Cytoplasmic extraction and vacuoli- zation of mitochondria are very frequent artifacts. Fixation was made in 2% glutaraldehyde in 0.1 M phosphate or cacodylate buffer at pH 7.2, either by immersion of tissue portions or by perfusion of the uterus through the thoracic aorta. The latter method generally gave better results and was preferred because it also maintained the topographical relations between embryo and endometrium. The implantation sites were sliced transversely to the long axis of the uterus and post-fixed in buffered osmium tetroxide. Following dehydration in a graded ethanol series the tissues were embedded in Araldite. Semi-serial sections, 0.25 ~tm thick, were made in order to locate the embryos. Different areas of the antimesometrial decidua were trimmed for further sectioning. Ultrathin sections, cut on a Sorvall MT-II ultramicrotome, were stained with uranyl acetate and lead citrate and examined in a Zeiss EM 9A electron microscope.
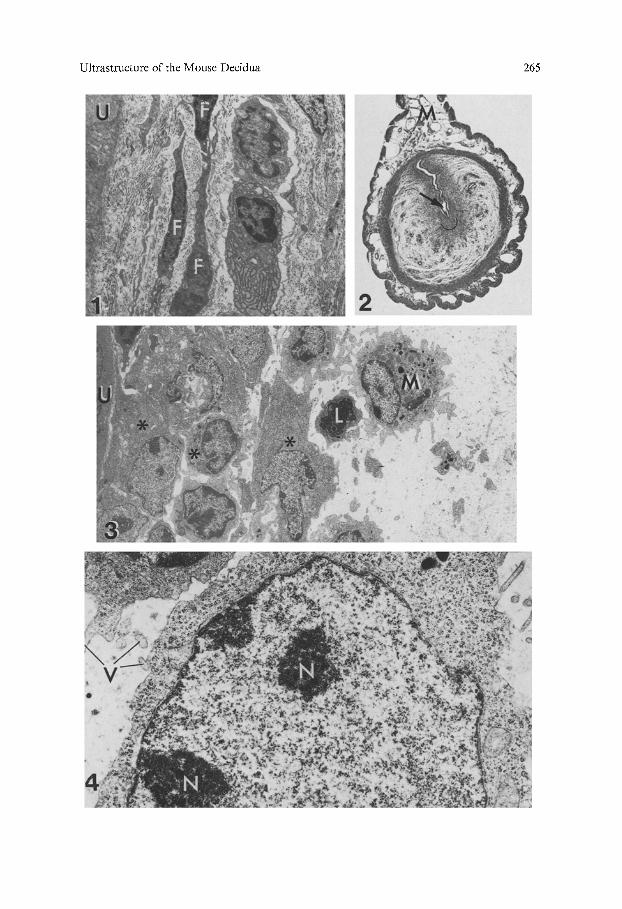
Fig. 1. Endometrium of a virgin mouse. The stromal cells (F) are spindle-shaped and sur- rounded by abundant collagen fibers. U uterine epithelium, x 4,400
Fig, 2. Transverse section of paraffin-embedded implantation site on the 5th d.p. The embryo (arrow) situated at the antimesometrial end of the uterine lumen is enveloped by a group of decidual cells whose approximate limits are indicated by the semicircle. M mesometrium. Hematoxylin-eosin stained, x 25
Fig. 3, On the 5th d.p. a few layers of polygonal decidual cells (*) surround the uterine epitheli- um (U). A macrophage (M) and a lymphocyte (L) are present at the border of the decidua. A large, intercellular oedema is present around the decidua, x 3,000
Fig. 4. Decidual cell on the 5th d.p. The nucleus contains dispersed chromatin and nucleoli (N). Free ribosomes predominate in the cytoplasm. V microvilli, x 14,700
Ultrastructure of the Mouse Decidua 265

266 P.A. Abrahamsohn
Results
Although the decidua is composed of a very homogeneous cell population, some morphological differences are always observable among cells on each day of pregnancy. Therefore, only the features common to the majority of cells will be described.
Virgin Animals: The stromal cells are mostly spindle-shaped and have long cytoplasmic processes (Fig. I). Only few granular endoplasmic reticu- lum cisternae and Golgi complexes are found in the cytoplasm. The nuclei are flat and contain some clumps of condensed chromatin.
5th d.p. : On the morning of the 5th d.p. the trophoblast and the uterine epithelium establish a very intimate contact. At this time, three to six poly- gonal decidual cell layers are present around the uterine epithelium sur- rounding the blastocysts (Fig. 3).
The nuclei of the decidual cells are round and usually indented. The chromatin is mostly dispersed and prominent nucleoli are observed. The cytoplasm is full of free ribosomes joined into polyribosomes, and exhibits a few strands of granular endoplasmic reticulum. Small Golgi complexes, a few lipid droplets and some lysosomes are occasionally found (Fig. 4).
The stromal cells situated near the myometrium are spindle-shaped. Be- tween these cells and the decidual cell layer lies a population of cells with intermediate features. The most striking characteristic of these intermediate cells is the cell body, which is round or polygonal but has cytoplasmic processes. The nuclei are round or elliptic (Fig. 5). These cells are in process of transformation into decidual cells and will be denominated predecidual cells.
The disposition of the decidual cells is compact, while an extensive inter- cellular space occurs between the predecidual cells. The space has few con- nective tissue fibres indicating the endometrial oedema typical of this im- plantation stage (Finn and McLaren 1967). Portions of the decidual cell surfaces are apposed and intercellular junctions are established between small areas of the cell membranes. Other cell types like neutrophils, lympho- cytes and macrophages may be found interspersed among transforming or spindle-shaped cells but not between decidual cells (Fig. 5).
6th d.p. : A large population of decidual cells surrounds the embryo (Fig. 6). The epithelium of the antimesometrial crypt is no longer present. The basement membrane of the epithelium is maintained for a certain period of time and separates the trophoblast from the decidual cells (Enders and Schlafke 1967). These cells are large, polygonal and have very narrow inter- cellular spaces. Their nuclei are larger but otherwise similar to those of cells of the preceding day. The prominent nucleoli frequently touch the inner nuclear membrane. Some multinucleate cells are found (Fig. 8). Most decidual cells contain large amounts of polyribosomes and granular endo- plasmic reticulum in the cytoplasm; they also exhibit several Golgi com- plexes, usually surrounded by vacuoles and coated vesicles (Figs. 7 and 8). The number of mitochondria, lysosomes and lipid droplets is greater than that of the 5th d.p. cells. Large quantities of microfilaments, frequently

Ultrastructure of the Mouse Decidua 267
Fig. 5. Predecidual cells enveloping decidual cells (not shown) on the 5th d.p. Note the poly- gonal cell body and rounded nucleus in association with long cell extensions, x 3,500
Fig. 6. Transverse section of paraffin-embedded uterus on the 6th d.p. The area enclosed within the semicircle indicates the antimesometriat decidua. Arrow embryo; M insertion of the mesometrium. Hematoxylin-eosin stained, x 25
Fig. 7. Binucleate decidual cell on the 6th d.p. The cytoplasm contains free ribosomes, cisternae of granular endoplasmic reticulum (E), several Golgi complexes (G), lipid droplets (L) and bundles of microfilaments (3//). x 13,300

268 P.A. Abrahamsohn
joined into thick bundles, are another feature consistently found in decidual cells from this stage onwards (Figs. 7 and 8). Organelles such as lysosomes and mitochondria were found associated with the microfilaments. A few spindle-shaped cells persisted between the decidual cells and the epithelial basement membrane. Their nuclei and cytoplasm are dark and show signs of degeneration. The predecidual cells present very similar characteristics to those seen on the 5th d.p. (Fig. 9).
The area of plasmalemma involved in the intercellular junctions is con- siderably larger than that of the preceding day. Many slender processes project from the cells. Other cell types besides endothelial cells are only rarely found in the decidua.
7th d.p.. Decidual cells occupy almost the whole width of the antimeso- metrial endometrium around each embryo.
The outline of the nuclei is more irregular than in the preceding days and clumps of chromatin are frequently found. The cytoplasm contains abundant granular and agranular endoplasmic reticulum cisternae. Golgi complexes and many bundles of microfilaments occupy significant areas of the cytoplasm (Figs. 10 and 11). A large number of primary and second- ary lysosomes is present in the cells (Fig. 12). The content of the latter is very heterogeneous; many lysosomes show lamellar or myelin figures, while other are autophagosomes that contain endoplasmic reticulum strands, mitochondria and ribosomes. Extensive areas of the surface mem- brane of contiguous cells are apposed and establish intercellular junctions.
8th d.p. : The antimesometrial decidua attains the maximal width on this day (Fig. 13). As in other days of pregnancy, the decidual cells display some degree of morphological diversity. Their nuclei have irregular surfaces and many clumps of chromatin. The number of multinucleated cells in- creases at this stage. The cytoplasm of most cells displays profuse quantities of granular and agranular endoplasmic reticulum cisternae and well-devel- oped Golgi complexes, always surrounded by coated and uncoated vesicles. The amount of lysosomes and autophagosomes is increased in many cells (Figs. 14 and 15). The lysosomes are found in a variety of forms, some with a rather homogeneous content but most display granules, long lamellae or circular "holes". Some membrane-bounded bodies, probably of lysoso- mal origin, contain a dense structure surrounded along its axis by a lucid material. Some cells have few organelles, the cytoplasm being filled with microfilaments (Figs. 16 and 17).
Fig. 8. Multinucleate decidual cell on the 6th d.p. Note bundles of microfilaments (*) and a Golgi complex (G). N nucleolus, x 13,800
Fig. 9. Predecidual cells on the 6th d.p. The cell body is thicker than that of spindle-shaped cells and the nuclei are elliptic. The lower cell is binucleate and is joined to the others by intercellular junctions (arrows). L lipid droplets, x 3,800
Fig. 10. Golgi complex in decidual cell on the 7th d.p. The organelle is surrounded by abundant vesicles and by cisternae of endoplasmic reticulum, x 19,000

Ultrastructure of the Mouse Decidua 269

270 P.A. Abrahamsohn
Discussion
The transformation of endometrial stromal cells into decidual cells is accom- plished by profound morphological modifications of the nucleus, cytoplasm and cell surface. The magnitude of the modifications may be judged by comparing the spindle-shaped cells of virgin animals with the ribosome-rich cytoplasm of cells from the 5th d.p. and the highly developed cytoplasm of the 6th d.p. cells. The structure of the 6th d.p. cells is similar to that of the rat and hamster (Jollie and Bencosme 1965; Enders and Schlafke 1967; Tachi etal. 1970; Parkening 1976), except that glycogen was not detected in our material and the agranular endoplasmic reticulum seems to be present in smaller amounts in the mouse. A noteworthy indication of stromal cell transformation was the presence of binucleate cells already on the 6th d.p. On subsequent days many cells presented four or five nuclei as revealed in thick or thin sections. These cells arise through mitosis without subsequent cytokinesis and not by cell fusion (Ansell et al. 1974). Decidual cells from the 6th d.p. can be considered mature and fully developed, since their morphology does not undergo remarkable changes during the follow- ing days. The transformation of individual cells is very rapid, considering that maturity can be achieved in less than 24 h. The speed of cell transforma- tion in terms of cells added between the 5th and 6th d.p. is another remark- able feature of the decidua. This can easily be evaluated by comparing the width of the antimesometrial decidua on both days.
The greatest rate of development of the mouse antimesometrial decidua seems to occur during the period from the early embryo-uterine attachment until the beginning of contact between the embryo and decidua that follows the epithelial breakdown. The rapid initial growth is probably important in establishing a barrier around the embryo which is assumed to control the trophoblastic invasion (Kirby 1965).
Decidualization proceeds from the subepithelial stroma towards the myometrium. The growth of the decidua results from mitosis, increase in cellular volume and addition of newly transformed cells around the already existant population of mature cells. The new cells undergo gradual modifica- tions during which intermediate forms (predecidual cells) between mature and untransformed cells can be recognized. These intermediate cells are morphologically similar to the stromal ceils present during induction of
Fig, 11. Cytoplasm of two 7th d.p. decidual cells. E endoplasmic reticulum; G Golgi complex; M bundle of microfilaments; L lipid droplets, x 14,500
Fig. 12. Possible fusion of lysosomes in a 7th d.p. cell. x 28,500
Fig. 13. Transverse section of paraffin-embedded uterus on the 8th d.p. Most of the antimeso- metrial wall is occupied by the decidua (D). Arrow embryo ; M insertion of the mesometrium. Hematoxylin - eosin stained, x 25
Fig. 14. Accumulation of lysosomes in the cytoplasm o f a decidual cell on the 7th d.p. x 11,000
Fig. 15. Autophagosomes (A) in deciduat cell on the 8th d.p. x 30,000

Ultrastructure of the Mouse Decidua 271

272 P.A. Abrahamsohn
Fig. 16, Cells on the 8th d.p. The cytoplasm of many cells at this stage is filled with secondary lysosomes and microfilaments (M). x 4,600. Inset: detail of lysosomes, x 16,000
Fig. 17. Cytoplasm of decidual cell on the 8th d.p. filled with microfilaments, x 19,000
decidualization in the rat (Lundkvist 1978). Human predecidual cells are present from the mid-luteal part of the cycle extending until late pregnancy (Lawn et al. 1971; Stark and Kaufmann 1973; Liebig and Stegner 1977). Such cells are probably homologous to those of rodents as they are generally enlarged due to cytoplasmic growth and present increased amounts of gran- ular endoplasmic reticulum (Lawn et al. 1971; Liebig and Stegner 1977). They do, however, show some features typical of human decidual cells such as small club-shaped processes and gap junctions between areas of the same cell (Liebig and Stegner 1977). Another interesting feature of human predeci- dual cells is the presence of large amounts of 17-fi-hydroxysteroid dehy- drogenase indicating that a certain degree of differentiation has been reached during the process of decidual cell transformation (Stark and Kaufmann 1973).
Towards the 7th d.p., the decidual cells accumulate large numbers of primary and secondary lysosomes. Autophagosomes and residual bodies are also present. All these structures are the first manifestation of the exten- sive involution which most of the antimesometrial decidua will undergo during the next few days of pregnancy (Katz and Abrahamsohn 1981). The increasing presence of lysosomes on the 7th d.p. signifies that the disor- ganization of the decidual cells begins as early as 48 h after their first appear- ance.

Ultrastructure of the Mouse Decidua 273
Very few "secretory" products have been associated with the decidual tissue. The longer lifetime of corpora lutea concomitant with the presence of deciduomata in the rat suggests an influence of the decidua on the ovary (Olsen et al. 1951). More recently, Basuray and Gibori (1980) demonstrated the production of a luteotropic "hormone" by rat decidual cells. The human decidua has been held responsible for the production of prolactin (Rosen- berg et al. 1980; Meuris et al. 1980), and rat decidual cells have been shown to produce prostaglandins in vitro (Vladimirsky et al. 1977). The richness of cytoplasmic organelles involved in synthetic activities revealed by ultra- structural studies in the mouse and other rodents, together with the dispersed chromatin and prominent nucleoli, indicate that decidual cells must be meta- bolically very active; this suggests that these cells might be involved in the production of several substances other than the few already ascribed to them.
Acknowledgements. The technical help of Mr. G.F. Lima and Mr. H. Correa as well as the photographical assistance of Mr. S.C. Silva and Mr. O.C. Silva are gratefully acknowledged. This work was made with help of grants from FAPESP, CNPq and PLAMIRH.
References
Ansell JD, Barlow PW, McLaren A (1974) Binucleate and polyploid cells in the decidua of the mouse. J Embryol Exp Morphol 31:223-227
Basuray R, Gibori G (1980) Luteotropic action of decidual tissue in the pregnant rat. Biol Reprod 23 : 507-512
Enders AC, Schlafke S (1967) A morphological analysis of the early implantation stages in the rat. Am J Anat 120:185-226
Finn CA, McLaren A (1967) A study of the early stages of implantation in mice. J Reprod Fertil 13:259-267
Jollie WP, Bencosme SA (1965) Electron microscopic observations on primary decidua forma- tion in the rat. Am J Anat 116:271-236
Katz S, Abrahamsohn PA (1981) Ultrastructural observations on the involution of the mouse antimesometrial decidua. Acta Anat 111 : 72-73
Kirby DRS (1965) The invasiveness of the trophoblast. In: Park WW (ed) The early conceptus, normal and abnormal. Livingstone, Edinburgh-London
Krehbiel RH (1937) Cytological studies of the decidual reaction in the rat during early pregnan- cy and in the production of deciduomata. Physiol Zool 10:212233
Lawn AM, Wilson EW, Finn CA (1971) The ultrastructure of human decidual and predecidual cells. J Reprod Fert 26:85-90
Liebig W, Stegner H-E (1977) Die Dezidualisation der endometriale Stromazelle. Elektronen- mikroskopische Untersuchungen. Arch Gyn/ik 223:19-31
Lundkvist 0 (1978) Morphological studies of the stromal changes after artificial induction of the decidual reaction in rats. Cell Tissue Res 194:287-296
Lundkvist O, Ljungkvist I (1977) Morphology of the rat endometrial stroma at the appearance of the pontamine blue reaction during implantation after an experimental delay. Cell Tissue Res 184:453-466
Meuris S, Soumenkoff G, Malengreau A, Bobyn C (1980) Immunoenzymatic localization of prolactin-like immunoreactivity in decidual cells of the endometrium from pregnant and nonpregnant women. J Histochem Cytochem 28:1347-1350
Nilsson O (1970) Some ultrastructural aspects of implantation. In: Hubinont PO, Leroy F, Robyn C, Leleux P (eds) Ovoimplantation, human gonadotrophins and prolactin. S Karger, Basel Mfinchen New York
Olsen AG, Velardo JT, Hisaw FL, Dawson AB, Braverman LE 0951) Prolongation of pseudo- pregnancy associated with the presence of deciduomata. Anat Rec 111:460

274 P.A. Abrahamsohn
Orsini MW, Wynn RM, Harris JA, Bulmash JM (1970) Comparative ultrastructure of the decidua in pregnancy and pseudo-pregnancy. Am J Obstet Gynec 106:14-25
Parkening T (1976) An ultrastructural study of implantation in the golden hamster. III. Initial formation and differentiation of decidual cells. J Anat 122:485-498
Potts DM (1968) The ultrastructure of implantation in the mouse. J Anat 103:77-90 Psychoyos A (1960) La r6action d6ciduale est pr~c~d6e de modifications pr~coces de la permea-
bilit6 capillaire de l'ut6rus. CR Soc Biol (Paris) 154:1384-1387 Reinius S (1967) Ultrastructure of blastocyst attachment in the mouse. Z Zellforsch 77 : 257-266 Rosenberg SM, Maslar IA, Riddick DH (1980) Decidual production of prolactin in late gesta-
tion: further evidence for a decidual source of amniotic fluid prolactin. Am J Obstet Gynecol 138:681-685
Snell GD (1941) The early embryology of the mouse. In: Snell GD Biology of the laboratory mouse. Dover, New York, p 1-54
Stark J, Kaufmann P (1973) Die Basalplatte der reifen menschlichen Placenta. III. Bindege- webs- und Deciduazellen. Arch Gyn/ik 213:399 417
Tachi S, Tachi C, Lindner HR (1970) Ultrastructural features of blastocyst attachment and trophoblastic invasion in the rat. J Reprod Fertil 21 : 37-56
Vladimirsky F, Chen L, Amsterdam A, Zor U, Lindner HR (1977) Differentiation of decidual cells in cultures of rat endometrium. J Reprod Fertil 49:61-68
Accepted November 22, 1982




















