
Ultrastructural analysis of vascular features in cerebral cavernous malformations
7
Clinical Neurology and Neurosurgery 115 (2013) 438–444 Contents lists available at SciVerse ScienceDirect Clinical Neurology and Neurosurgery j o ur nal homep age: www.elsevier.com/locate/clineuro Ultrastructural analysis of vascular features in cerebral cavernous malformations Gamze Tanriover a , Berna Sozen a , Askin Seker b , Turker Kilic b , Murat Gunel c , Necdet Demir a,∗ a Department of Histology and Embryology, Akdeniz University, School of Medicine, Antalya 07070, Turkey b Institute of Neurological Sciences, Marmara University, 34840 Istanbul, Turkey c Department of Neurosurgery, Yale University, School of Medicine, 333 Cedar Street, 06510 New Haven, CT, USA a r t i c l e i n f o Article history: Received 21 March 2011 Received in revised form 6 June 2012 Accepted 16 June 2012 Available online 7 July 2012 Keywords: Cerebral cavernous malformation Ultrastructure TEM SEM Immunohistochemistry CCM2 CCM3 a b s t r a c t Objective: Investigation of the structure of vascular malformations highlights the pathogenic mechanisms underlying their clinical behavior. One of the vascular malformations is called cerebral cavernous mal- formation (CCM). However, the ultrastructural features of the vascular malformations are not defined in detail. Methods: We aimed to investigate the ultrastructural features of CCMs using transmission (TEM), scanning (SEM) electron microscopy, and also immunohistochemistry methods with antibodies against CCM pro- teins such as CCM2 and CCM3. CCM tissues (n = 6) microsurgically excised from patients for conventional indications. Results: CCM2 and CCM3 were strongly detected in the vascular endothelium. However, there was a very weak immunostaining in stroma. SEM observations revealed that there were ruptures and damages in the luminal endothelium, possibly due to the damage of intercellular junctions. TEM observations also showed a few ruptures and detachments between the endothelium and basal lamina as observed with partially damages and disconnections. The architecture of pericytes showed protrusions and shrinkages. Our results suggest that the thin vessel walls of CCMs were lacking of subendothelial support and intact basal lamina underlying the endothelial cells. Conclusion: This study is so far the first study attempting to show human CCM lesions with SEM. We believe that an understanding of the ultrastructural features of these lesions by light and electron microscopy techniques would help to understand the pathology of these diseases. © 2012 Elsevier B.V. All rights reserved. 1. Introduction The vascular network of the brain is dependent on the blood flow and resistance of the large arteries and the smaller, terminal arteri- oles. Alterations in the vessel ultrastructure potentially contribute to alterations in both blood flow and the transport of materials [1,2]. The supportive system of the capillary such as endothelial cells, basal lamina, pericytes are also important for the vascu- lar architecture, blood flow and transport of materials. Moreover, there is another important supportive system around the capillar- ies namely membrana limitans glia which is composed of astrocytic processes. Brain vascular endothelial cells and their intercellular tight junc- tions form a cellular interface between the circulating blood and neural environment. The endothelial cells are surrounded by a ∗ Corresponding author at: Department of Histology and Embryology, Faculty of Medicine, Akdeniz University, 07070 Campus, Antalya, Turkey. Tel.: +90 242 249 6884; fax: +90 242 227 4486. E-mail address: [email protected] (N. Demir). 30–40 nm thick basal lamina which is a target of investigation under pathophysiological conditions [3–7]. Also, pericytes, which are relatively undifferentiated cells communicate with the cells of the neurovascular unit [8]. The neurovascular unit that is a specific interplay between the neuron and the endothelium is crucial to the development of the brain and its vasculature [9]. Cerebral cavernous malformation (CCM) is one of the most com- mon types of vascular malformations. CCMs can occur anywhere in the body, but usually produce serious signs and symptoms only when they occur in the central nervous system [10]. CCM lesions can also lead, albeit rarely, to gross hemorrhages causing clinical signs and symptoms reflecting the size and location of the bleed- ing. CCMs are collections of “irregular and enlarged” capillaries in the brain [11]. When viewed under the light microscope, these vas- cular abnormalities are made up of fairly large blood-filled channels or “caverns”. Endothelial cells play a role in progression of various pathological processes. CCMs consist of blood or thrombus-filled caverns lined by a single layer of endothelial cells, while its inter cavernous spaces lack mature vessel wall elements [12]. Therefore, it is possible that endothelial cells of CCMs, as the major component of the vessel wall, may play a central role in the pathogenesis of the 0303-8467/$ – see front matter © 2012 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.clineuro.2012.06.023
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Ultrastructural analysis of vascular features in cerebral cavernous malformations

U
Ga
b
c
a
ARRAA
KCUTSICC
1
aot[cltip
tn
MT
0h
Clinical Neurology and Neurosurgery 115 (2013) 438– 444
Contents lists available at SciVerse ScienceDirect
Clinical Neurology and Neurosurgery
j o ur nal homep age: www.elsev ier .com/ locate /c l ineuro
ltrastructural analysis of vascular features in cerebral cavernous malformations
amze Tanriovera, Berna Sozena, Askin Sekerb, Turker Kilicb, Murat Gunelc, Necdet Demira,∗
Department of Histology and Embryology, Akdeniz University, School of Medicine, Antalya 07070, TurkeyInstitute of Neurological Sciences, Marmara University, 34840 Istanbul, TurkeyDepartment of Neurosurgery, Yale University, School of Medicine, 333 Cedar Street, 06510 New Haven, CT, USA
r t i c l e i n f o
rticle history:eceived 21 March 2011eceived in revised form 6 June 2012ccepted 16 June 2012vailable online 7 July 2012
eywords:erebral cavernous malformationltrastructureEMEMmmunohistochemistryCM2CM3
a b s t r a c t
Objective: Investigation of the structure of vascular malformations highlights the pathogenic mechanismsunderlying their clinical behavior. One of the vascular malformations is called cerebral cavernous mal-formation (CCM). However, the ultrastructural features of the vascular malformations are not defined indetail.Methods: We aimed to investigate the ultrastructural features of CCMs using transmission (TEM), scanning(SEM) electron microscopy, and also immunohistochemistry methods with antibodies against CCM pro-teins such as CCM2 and CCM3. CCM tissues (n = 6) microsurgically excised from patients for conventionalindications.Results: CCM2 and CCM3 were strongly detected in the vascular endothelium. However, there was a veryweak immunostaining in stroma. SEM observations revealed that there were ruptures and damages inthe luminal endothelium, possibly due to the damage of intercellular junctions. TEM observations alsoshowed a few ruptures and detachments between the endothelium and basal lamina as observed with
partially damages and disconnections. The architecture of pericytes showed protrusions and shrinkages.Our results suggest that the thin vessel walls of CCMs were lacking of subendothelial support and intactbasal lamina underlying the endothelial cells.Conclusion: This study is so far the first study attempting to show human CCM lesions with SEM. Webelieve that an understanding of the ultrastructural features of these lesions by light and electronmicroscopy techniques would help to understand the pathology of these diseases.© 2012 Elsevier B.V. All rights reserved.
. Introduction
The vascular network of the brain is dependent on the blood flownd resistance of the large arteries and the smaller, terminal arteri-les. Alterations in the vessel ultrastructure potentially contributeo alterations in both blood flow and the transport of materials1,2]. The supportive system of the capillary such as endothelialells, basal lamina, pericytes are also important for the vascu-ar architecture, blood flow and transport of materials. Moreover,here is another important supportive system around the capillar-es namely membrana limitans glia which is composed of astrocyticrocesses.
Brain vascular endothelial cells and their intercellular tight junc-ions form a cellular interface between the circulating blood andeural environment. The endothelial cells are surrounded by a
∗ Corresponding author at: Department of Histology and Embryology, Faculty ofedicine, Akdeniz University, 07070 Campus, Antalya, Turkey.
el.: +90 242 249 6884; fax: +90 242 227 4486.E-mail address: [email protected] (N. Demir).
303-8467/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.clineuro.2012.06.023
30–40 nm thick basal lamina which is a target of investigationunder pathophysiological conditions [3–7]. Also, pericytes, whichare relatively undifferentiated cells communicate with the cells ofthe neurovascular unit [8]. The neurovascular unit that is a specificinterplay between the neuron and the endothelium is crucial to thedevelopment of the brain and its vasculature [9].
Cerebral cavernous malformation (CCM) is one of the most com-mon types of vascular malformations. CCMs can occur anywhere inthe body, but usually produce serious signs and symptoms onlywhen they occur in the central nervous system [10]. CCM lesionscan also lead, albeit rarely, to gross hemorrhages causing clinicalsigns and symptoms reflecting the size and location of the bleed-ing. CCMs are collections of “irregular and enlarged” capillaries inthe brain [11]. When viewed under the light microscope, these vas-cular abnormalities are made up of fairly large blood-filled channelsor “caverns”. Endothelial cells play a role in progression of variouspathological processes. CCMs consist of blood or thrombus-filled
caverns lined by a single layer of endothelial cells, while its intercavernous spaces lack mature vessel wall elements [12]. Therefore,it is possible that endothelial cells of CCMs, as the major componentof the vessel wall, may play a central role in the pathogenesis of the
y and
lrf
ftmTtsda
ttmgwc
t
2
2
MctAtMcm
ecaphowa
2
tp
2
wtpt
2
soAV
G. Tanriover et al. / Clinical Neurolog
esion. Previous descriptive studies on CCM lesions indicates matu-ation phase of angiogenesis and its involved structural moleculesor CCM pathogenesis [13].
Electron microscopy technique is a highly valuable techniqueor deep understanding of the structural problems of the lesion,herefore, the analysis of the microscopic anatomy of the CCMs
ay be very useful to clarify the histopathology of this disease.he present study was designed to compare the ultrastructural fea-ures between CCMs and normal human brain vessels. In the CCMpecimens, we examined the components of the vessel and suben-othelial wall structures such as basal lamina, pericytes, stromand neuroglial components.
As a second aim, it is known that the disease is most likelyo have a genetic cause. Screening for the CCM genes may iden-ify a mutation in around 50% of cases. Three CCM loci have been
apped as Ccm1, Ccm2, Ccm3 genes in CCMs. In order to investi-ate whether CCM proteins may participate in CCM pathogenesis,e report here also the expression patterns of CCM2 and CCM3 in
avernoma lesions by means of immunohistochemistry.To our knowledge, this study is so far the first demonstration of
he SEM observations on human CCM lesions.
. Materials and methods
.1. Tissue collection
Tissues were supplied from the Department of Neurosurgery,armara University School of Medicine, Istanbul, Turkey. Informed
onsent forms and protocols were approved by the Ethical Commit-ee of School of Medicine, Marmara University, Istanbul, Turkey.lso, two normal cerebral tissues as control of the vascular struc-
ure from Department of Pathology, Akdeniz University School ofedicine were used in the study. None of the control specimens had
erebral malformation or cerebral ischemic/hemorrhagic abnor-alities and were in normal neurological conditions.For this study we selected surgically easily resectable non-
loquently located cavernoma samples from the patients. So theavernoma tissues were removed entirely without any partition orny piece. After the excision the tissues were washed with phos-hate buffer saline (pH: 7.4) and the blood was removed. This couldelp us to protect the endothelial cells and the inner environmentf cavernoma tissues. During this period total 20 cavernoma casesere operated and only six of them were selected for the study
ccording to this criteria.
.2. Histological evaluation
Tissue sections from CCM samples were stained with hema-oxylin and eosin for basic histological evaluation using standardrotocols.
.3. CCM2 and CCM3 antibody production
A CCM3/PDCD10 (CCM3)-specific rabbit polyclonal antibodyas synthesized and affinity-purified against the antigenic pep-
ide KIPDEINDRVRFLQTIKD (Open Biosystems, Huntsville, AL) asreviously described [14]. The CCM2 antibody synthesis, charac-erization, and validation have been previously described [15].
.4. Immunohistochemistry
For CCM2, CCM3 and CD31/PECAM-1 immunohistochemistry
ections were deparaffinized and blocked for endogenous per-xidase activity with methanol containing 3% H2O2 for 30 min.fterwards, sections were incubated in a blocking serum (UltraBlock, TP-060-HL; Neo-Marker, Fremont, CA, USA) for 7 min in
Neurosurgery 115 (2013) 438– 444 439
order to block non-specific binding without the washing step. Sec-tions were subsequently incubated over night at 4 ◦C with eitheranti-rabbit CCM2 or CCM3. They are diluted in dilution buffer1/250 (LabVision, Fremont, CA, USA) for the identification of CCMproteins or mouse monoclonal anti-human CD31/PECAM-1 (MS-1873-R7; Clone 1A10; prediluted, NeoMarkers, Fremont, CA, USA)to identify the vascular endothelium of the caverns. As a negativecontrol, the primary antibodies were replaced by normal rabbitand mouse IgG serum (Vector Lab., Burlingame, CA, USA) at thesame concentration. Sections were incubated with biotinylatedgoat anti-rabbit IgG or biotinylated anti-mouse secondary anti-body (1/400 dilution, Vector Lab. Burlingame, CA, USA) for 30 minfollowed by LSAB streptavidin-peroxidase complex (Dako, Carpin-teria, CA, USA) incubation for 30 min. The resulting signal wasdeveloped with diaminobenzidine (DAB) (K3466; Dako). Each stepwas followed by three washes in phosphate buffered saline (PBS).Sections were counterstained with Mayer’s hematoxylin (Dako),dehydrated, mounted and examined under the light microscope(Zeiss-Axioplan, Oberkochen, Germany).
2.5. Scanning electron microscopy (SEM)
Tissue samples from the CCMs were prepared for SEM analy-sis. Following the same fixation procedure used for TEM, sampleswere dehydrated through a gradually increasing acetone series. Thetissues were kept in n-amyl acetate for critical point drying (E300,Polaron, ThermoVG Micro Tech, England). Tissues were coated withgold-palladium particles and examined using a Zeiss 1430 scanningelectron microscope (Leo 1430, Zeiss, Germany).
2.6. Transmission electron microscopy (TEM)
The CCMs and normal brain tissues were fixed with 2.5% glu-taraldehyde in 0.1 M phosphate buffer, pH 7.4. The tissue sampleswere post-fixed in 1% osmium tetroxide solution using an isotonic0.1 M phosphate buffer, pH 7.4 solution at 4 ◦C for 1 h. Samples weredehydrated through a gradually increasing series of ethanol andembedded in Araldite resin (Araldite CY 212, 91 ml + dodecenyl suc-cinic anhydride (DDSA) 84 ml + N,N-dimethylbenzylamine (BDMA)3.5 ml, TAAB Company, England). Semi-thin (1 mm) and ultra-thin (40–60 nm) sections were cut using diamond knives on aLeica ultramicrotome (Leica ultracut, UCT, Leica MZ6, England).Ultra-thin sections were collected on copper grids (Mesh 100,TAAB, England; coated with Formvar 15/95 Resin polyvinyl for-mol, EMS-Electron Microscopy Sciences, Fort Washington, PA,USA). In semi-thin sections, routine toluidine blue staining methodwas applied for orientation. In ultra-thin sections, double-contraststaining was applied with uranyl acetate (100 ml methanol and 5 guranyl acetate) and Reynold’s lead nitrate solution (1.76 g sodiumcitrate, 1.33 g lead nitrate, 50 ml distilled water and 8 ml 1 N NaOH)[16]. Ultra-thin sections were examined using a LEO 906 transmis-sion electron microscope (Leo 906E, Zeiss, Germany).
3. Results
3.1. Histological evaluation of the CCM samples
Histological evaluation of the CCM lesion composed of endothe-lial cell-lined caverns filled with blood cells and thrombus wereobserved under a light microscope with Hematoxylin and eosinstaining (Fig. 1A and B). The caverns were separated with inter-
cavernous matrix consisting of amorphous matrix without anyneural components such as neuropil or neurons. Also, denser inter-cavernous matrix containing some leukocyte infiltration areas waswidely distributed. Moreover, we have categorized the caverns
440 G. Tanriover et al. / Clinical Neurology and Neurosurgery 115 (2013) 438– 444
F lin anA L, leu
ao
3
eTcs
Fe
ig. 1. Representative image of the histological sections that stained with hematoxyrrowhead: blood cells and thrombus. B, M, and S: big, medium and small caverns;
s big, medium and small. The small caverns were commonlybserved in the inter-cavernous area.
.2. Immunohistochemistry
CCM2 and CCM3 proteins were strongly detected in the vascular
ndothelium with very weak staining pattern in stroma (Fig. 2A–D).o clarify the cell types, CD31/PECAM-1 staining was used toonfirm the endothelial staining of CCM2 and CCM3 on serialections of the CCM samples. CCM2 and CCM3 expressions wereig. 2. Representative pictures of CCM2 (A), CCM3 (C) and CD31/PECAM-1 (E and F) protndothelium (arrows) and capillaries (F, arrows) in cavernoma tissues. Scale bars represe
d eosin. They are consisted of vascular sinusoids lined by endothelial cells (arrows).kocyte infiltration area. Scale bars represent 50 �m.
detected in the vascular endothelium, which was also labeled withCD31/PECAM-1 in CCMs. In addition, CD31/PECAM immunore-activity was also detected in some of the endothelium of smallcapillaries in inter-cavernous areas (Fig. 2E and F).
3.3. Scanning electron microscopy (SEM)
The SEM analysis of the cavernoma revealed that the three-dimensional appearance of the cavern shapes appeared in a normalstructure (Fig. 3). The caverns were observed with different sizes
eins expression in cavernoma. An intense immunostainings were seen in vascularnt 50 �m.

G. Tanriover et al. / Clinical Neurology and Neurosurgery 115 (2013) 438– 444 441
Fig. 3. SEM micrographs of CCMs. (A and B) The caverns were connected with each other constituting a labyrinth like structure (arrows). The higher magnification of thea caveri like lue
awmcr(otv
3
ltrAwecobci
rea from B is presented as C. This image was shown that the cavern (asterix). Then regular endothelial structures (B, arrowheads) however, some of them were gridndothelium were observed in a different size and diameters (K, arrows).
nd luminal morphology. We observed that they were connectedith each other constituting a labyrinth like structure. Further-ore, the big caverns were observed to be connected to the small
averns in the stroma of CCM. The endothelium of CCM revealeduptured luminal lining and partial detachments in the cavernsFig. 3C–K). In addition to normal endothelial structures in somef the caverns, a grid like luminal surfaces with endothelial rup-ures in different sizes and diameters were also observed (Fig. 3Bs Fig. 3H).
.4. Ultrastructural analysis of CCMs
According to TEM analysis, the vascular endothelium, basalamina and pericytes of the vascular unit showed normal struc-ural organization in the normal brain vessels. Perivascular areaevealed regular and normal structural view in vascular unit.
few pinocytotic vesicles and irregular protoplasmic processesere seen through the lumen in the vascular endothelium. How-
ver, endothelial cells did not reveal any inter-digitations in theytoplasm. The subcellular components showed normal morphol-
gy throughout the cytoplasm of endothelium. Also, basal laminaetween endothelium and pericytes was in regular thickness andontinuity. Astrocyte feet around the vascular structures were alson normal condition.ns with different size and luminal morphology (D–G, asterix). Some caverns wereminal surface (H, arrows). Ruptured luminal lining and partial detachments of the
In the CCM lesions, the ultrastructural feature of theendothelium was observed with distinctive and prominent hete-rochromatic nuclei. Moreover, the endothelium was consisted ofenlarged pinocytotic vesicles (Fig. 4A–C), chaotic protoplasmic pro-trusions (Fig. 4B) and also showed intra-cytoplasmic canaliculi. Thestructure of the pericytes revealed protrusions and shrinkages asit was observed in the vascular endothelium. Also, the consider-able accumulations of amorphous substance with different sizesand irregular shapes were situated in the caverns. The cavernwalls in CCMs were composed of an amorphous matrix with-out significant support from organized connective tissue matrix(Fig. 4D).
The basal lamina showed a very thin and partial membranousstructure underlying the endothelium (Fig. 4E–G). The endothe-lium was exposed directly to the amorphous matrix with edemaand collagen fibers. Therefore, the basal lamina was not clearlyobserved in these areas (Fig. 4G and L). In the peripheral regionsof the CCMs, capillaries were often surrounded by erythrocytes(Fig. 4H and K).
The macrophages were clearly observed with phagocytic nuclei
and electron dense bodies including phagosome and lysosomesin the cytoplasm (Fig. 4M). The labyrinth-like cells were detectedwith irregular shape and heterochromatic nuclei and dispersedorganelles around the nucleus (Fig. 4N and P).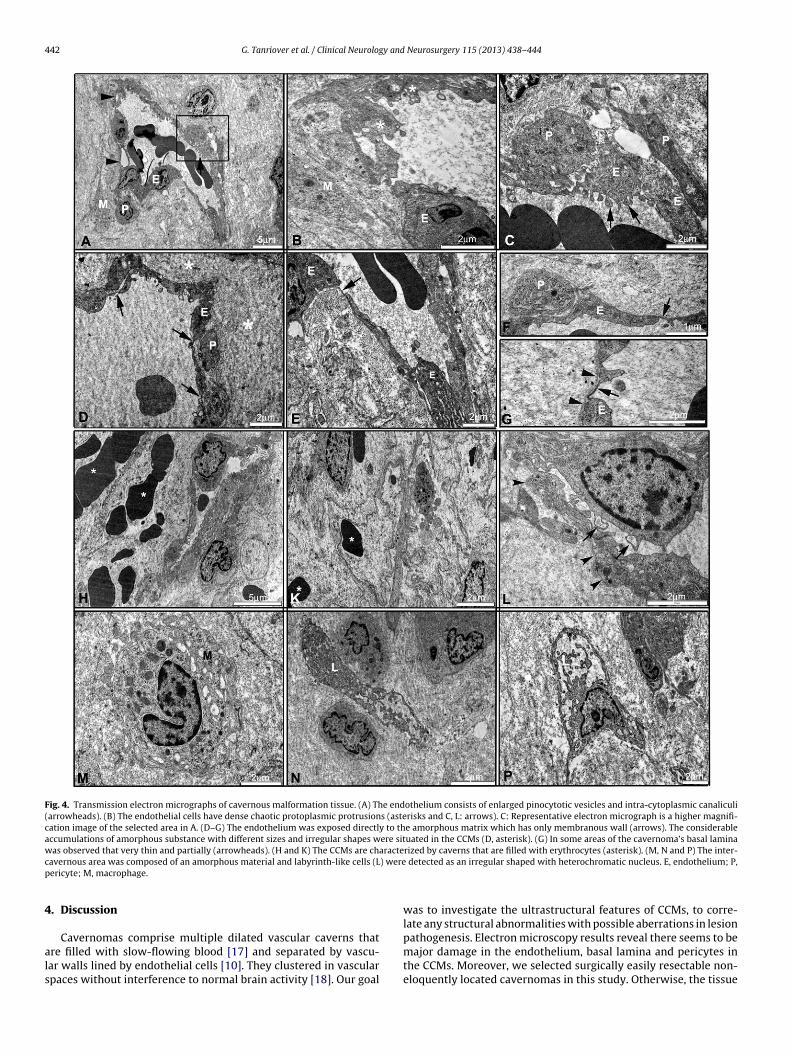
442 G. Tanriover et al. / Clinical Neurology and Neurosurgery 115 (2013) 438– 444
Fig. 4. Transmission electron micrographs of cavernous malformation tissue. (A) The endothelium consists of enlarged pinocytotic vesicles and intra-cytoplasmic canaliculi(arrowheads). (B) The endothelial cells have dense chaotic protoplasmic protrusions (asterisks and C, L: arrows). C: Representative electron micrograph is a higher magnifi-cation image of the selected area in A. (D–G) The endothelium was exposed directly to the amorphous matrix which has only membranous wall (arrows). The considerableaccumulations of amorphous substance with different sizes and irregular shapes were situated in the CCMs (D, asterisk). (G) In some areas of the cavernoma’s basal laminawas observed that very thin and partially (arrowheads). (H and K) The CCMs are characterized by caverns that are filled with erythrocytes (asterisk). (M, N and P) The inter-c ) werep
4
als
avernous area was composed of an amorphous material and labyrinth-like cells (Lericyte; M, macrophage.
. Discussion
Cavernomas comprise multiple dilated vascular caverns thatre filled with slow-flowing blood [17] and separated by vascu-ar walls lined by endothelial cells [10]. They clustered in vascularpaces without interference to normal brain activity [18]. Our goal
detected as an irregular shaped with heterochromatic nucleus. E, endothelium; P,
was to investigate the ultrastructural features of CCMs, to corre-late any structural abnormalities with possible aberrations in lesion
pathogenesis. Electron microscopy results reveal there seems to bemajor damage in the endothelium, basal lamina and pericytes inthe CCMs. Moreover, we selected surgically easily resectable non-eloquently located cavernomas in this study. Otherwise, the tissue
y and
ibt[ra
oimfplspi
rbsvsdp
weolthbMebtfvdbthpwogo
gpmaiClt
slogMsVfd
G. Tanriover et al. / Clinical Neurolog
s often thrombosed, reorganized, and could be artificially alteredy the surgical manipulation. Therefore, the endothelial cells lininghe unaltered caverns could only represent a small subset of cells19]. Thus, we dissected the gliotic tissue around the lesion andemoved the cavernoma without dividing the tissue into the partsnd without using electrosurgical coagulation.
Our TEM and SEM findings confirm and extend the previ-us structural findings suggesting that vascular alterations aremportant in the pathophysiology of cavernous malformations. The
arked differences in brain cavernous malformation architecturerom normal brain microvasculature may play a part in the patho-hysiology of these lesions. Apparently, the membranous basal
amina, cytoplasmic protrusions, local damage of the endothelialurface and structurally weakened vessels were prominent. So, theresent findings are in agreement with results from previous stud-
es using the electron microscopy techniques [11,20].Moreover, one of the other cerebrovascular disorders is an arte-
iovenous malformation (AVM). AVM is an abnormal connectionetween the arteries and veins in the brain. Tu et al. examined AVMamples that observed pericytes were rich in pinocytotic vesicles,acuoles, and filaments [21]. But, in CCMs, pericytes were rarelyeen and showed protrusions and shrinkages [20] which is in accor-ance with our results. Although the morphological changes to theericytes might have evolved in the structure of the lesions.
Our study showed that the basal lamina underlying the ECshether is partially damaged or not encountered in CCMs. Appar-
nt abnormality in the basal lamina has been described in somef the other studies [11,20]. Therefore, it is possible to specu-ate that the thin and partially membranous structure underlyinghe endothelial cells or without basal lamina might contribute toemorrhages. Also, the erythrocytes have easily passed throughetween endothelial cells and the poorly organized cavern walls.oreover, our results showed that the endothelial cells were
xposed directly to the amorphous matrix which have a thin mem-ranous interactions. It is already known that CCM specimens havehin vessel walls that are lined by endothelial cells. So, their dif-erent bleeding rate may be due to the different structure of theessel walls, the blood flow and the intravascular pressure in theifferent lesions [22]. Also, the endothelial differences were seeny using scanning electron microscopy. It is known that the distinc-ive structure of the lesions may be important in their high risk ofemorrhage [11]. This would indicate that the capillary lumen anderivascular components were directly interact with membranousall. However, endothelial cells were disrupted, with preservation
f the underlying subendothelial vessel wall in AVMs [23]. It is sug-ested that a weakness in the vessel wall might be an important rolef hemorrhage in AVM structure.
CCM is inherited in an autosomal dominant manner, and theenes mutated in three forms have been identified [24]. We hadreviously hypothesized that these genes all participate in a com-on pathway [14] and recent evidence suggests that these genes
re expressed in an over-lapping fashion in the brain [14]. Find-ngs of this study supports our previous hypothesis that CCM2 andCM3 proteins have similar expression patterns in the cavernoma
esions, which indicates common pathway in CCM development byhree different genes.
The pathogenesis of CCM is still unknown, but it has beenuggested that their development may be linked to aberrant vascu-ogenesis [25]. In our study these proteins are strongly expressedn the vascular endothelium that CCMs might exhibit more angio-enic activity than normal vascular structures of the adult brain.oreover, Kilic et al., have previously shown the aberrant expres-
ion of some angiogenic proteins on CCMs [13]. Additionally, bothEGF and growth factor receptors, that are thought to be important
or vasculogenesis and angiogenesis, also have been shown to beeregulated in cerebrovascular malformations [12]. Dependent on
Neurosurgery 115 (2013) 438– 444 443
the recent findings regarding the role of CCM2 and CCM3 [14,26],we may speculate that either co-expression of these proteins ortheir individual expressions may contribute to the development ofaberrant architecture of the lesions.
In addition to vascular endothelial expression, these proteins arelocalized very weak staining in stroma that might be a fibrocyte ofcaverns.
CCM2 and CCM3 gets role in cell proliferation and differentiation[27]. CCM2 protein is required for endothelial cell-cell junctionallocalization, and impaired protein causes destabilization of bar-rier structures [28]. Stockton et al. suggested that KRIT1-CCM2interaction regulates vascular barrier function and this interac-tion is dysregulated in human CCM endothelium [28]. Moreover,CCM3 promotes VEGFR2 signaling during vascular development[29]. Thus, we suggest that these proteins could ameliorate bothCCM disease and vascular leak.
Formation of competent cerebral microvasculature relies onappropriate signaling between adjacent endothelial cells, pericytes,astrocytes, and the stroma. Genetic abnormalities affecting theseendothelial receptors such as Tie1, Tie2 have caused some prob-lems in the blood vessel formation [30]. The strong expression ofthe CCM2 and CCM3 proteins might be important in the normalformation and function of the vasculature.
In conclusion, our results showed that thin vessel walls of CCMsare composed of amorphous material lacking endothelial supportand also, this may contribute to the known propensity of CCMsfor recurrent hemorrhages. Additionally, further ultrastructuralexaminations of CCMs are needed to understand the accuracy andrelevance of these findings. Further studies should aim to find othermolecules such as angiogenic and vasculogenic markers interact-ing with CCM2 and CCM3, and to identify which signaling pathwaysare affected by these proteins. It is clear that further studies relatedto these proteins or genes expression, their possible contributionsto pathogenesis of CCMs in central nervous system.
Acknowledgments
This study was partially supported by the Akdeniz UniversityResearch Foundation, Antalya, Turkey. The authors would like tothank E. Inanc GURER for her excellent scientific collaborationfrom the Department of Pathology, Akdeniz University School ofMedicine and to thank Arife Demirtop and Hakan Er for their excel-lent technical assistance.
References
[1] Farkas E, Luiten PG. Cerebral microvascular pathology in aging and Alzheimer’sdisease. Progress in Neurobiology 2001;64:575–611.
[2] Kalaria RN. Cerebral vessels in ageing and Alzheimer’s disease. Pharmacology& Therapeutics 1996;72:193–214.
[3] Perlmutter LS, Chui HC, Saperia D, Athanikar J. Microangiopathy and the colo-calization of heparan sulfate proteoglycan with amyloid in senile plaques ofAlzheimer’s disease. Brain Research 1990;508:13–9.
[4] Claudio L. Ultrastructural features of the blood–brain barrier in biopsy tissuefrom Alzheimer’s disease patients. Acta Neuropathologica 1996;91:6–14.
[5] Kalaria RN, Premkumar DR, Pax AB, Cohen DL, Lieberburg I. Production andincreased detection of amyloid beta protein and amyloidogenic fragments inbrain microvessels, meningeal vessels and choroid plexus in Alzheimer’s dis-ease. Brain Research: Molecular Brain Research 1996;35:58–68.
[6] Elhusseiny A, Cohen Z, Olivier A, Stanimirovic DB, Hamel E. Functionalacetylcholine muscarinic receptor subtypes in human brain microcirculation:identification and cellular localization. Journal of Cerebral Blood Flow andMetabolism 1999;19:794–802.
[7] Luiten PG, de Jong GI, Van der Zee EA, van Dijken H. Ultrastructural localiza-tion of cholinergic muscarinic receptors in rat brain cortical capillaries. BrainResearch 1996;720:225–9.
[8] Kelley C, D’Amore P, Hechtman HB, Shepro D. Microvascular pericyte contrac-tility in vitro: comparison with other cells of the vascular wall. Journal of CellBiology 1987;104:483–90.
[9] del Zoppo GJ. Relationship of neurovascular elements to neuron injury duringischemia. Cerebrovascular Diseases 2009;27(Suppl. 1):65–76.

4 y and
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
44 G. Tanriover et al. / Clinical Neurolog
10] Dashti SR, Hoffer A, Hu YC, Selman WR. Molecular genetics of familial cerebralcavernous malformations. Neurosurgical Focus 2006;21:e2.
11] Clatterbuck RE, Eberhart CG, Crain BJ, Rigamonti D. Ultrastructural andimmunocytochemical evidence that an incompetent blood–brain barrier isrelated to the pathophysiology of cavernous malformations. Journal of Neu-rology, Neurosurgery and Psychiatry 2001;71:188–92.
12] Uranishi R, Baev NI, Ng PY, Kim JH, Awad IA. Expression of endothelial cell angio-genesis receptors in human cerebrovascular malformations. Neurosurgery2001;48:359–67 [discussion 367–8].
13] Kilic T, Pamir MN, Kullu S, Eren F, Ozek MM, Black PM. Expression of structuralproteins and angiogenic factors in cerebrovascular anomalies. Neurosurgery2000;46:1179–91 [discussion 1191–2].
14] Tanriover G, Boylan AJ, Diluna ML, Pricola KL, Louvi A, Gunel M. PDCD10, thegene mutated in cerebral cavernous malformation 3, is expressed in the neu-rovascular unit. Neurosurgery 2008;62:930–8 [discussion 938].
15] Seker A, Pricola KL, Guclu B, Ozturk AK, Louvi A, Gunel M. CCM2 expressionparallels that of CCM1. Stroke 2006;37:518–23.
16] Berkman Z, Tanriover G, Acar G, Sati L, Altug T, Demir R. Changes in thebrain cortex of rabbits on a cholesterol-rich diet following supplementa-tion with a herbal extract of Tribulus terrestris. Histology and Histopathology2009;24:683–92.
17] Voigt K, Yasargil MG. Cerebral cavernous haemangiomas or cavernomas.Incidence, pathology, localization, diagnosis, clinical features and treatment.Review of the literature and report of an unusual case. Neurochirurgia1976;19:59–68.
18] Russell DS RW. Pathology of Tumours of the Nervous System. Baltimore;
1989.19] Pagenstecher A, Stahl S, Sure U, Felbor U. A two-hit mechanism causescerebral cavernous malformations: complete inactivation of CCM1, CCM2or CCM3 in affected endothelial cells. Human Molecular Genetics 2009;18:911–8.
[
Neurosurgery 115 (2013) 438– 444
20] Tu J, Stoodley MA, Morgan MK, Storer KP. Ultrastructural characteristics of hem-orrhagic, nonhemorrhagic, and recurrent cavernous malformations. Journal ofNeurosurgery 2005;103:903–9.
21] Tu J, Stoodley MA, Morgan MK, Storer KP. Ultrastructure of perinidal capillar-ies in cerebral arteriovenous malformations. Neurosurgery 2006;58:961–70[discussion 961–70].
22] Tu J, Stoodley MA, Morgan MK, Storer KP, Smee R. Different responses ofcavernous malformations and arteriovenous malformations to radiosurgery.Journal of Clinical Neuroscience 2009;16:945–9.
23] Wong JH, Awad IA, Kim JH. Ultrastructural pathological features of cerebrovas-cular malformations: a preliminary report. Neurosurgery 2000;46:1454–9.
24] Bergametti F, Denier C, Labauge P, Arnoult M, Boetto S, Clanet M, et al. Muta-tions within the programmed cell death 10 gene cause cerebral cavernousmalformations. American Journal of Human Genetics 2005;76:42–51.
25] Shalaby F, Rossant J, Yamaguchi TP, Gertsenstein M, Wu XF, Breitman ML, et al.Failure of blood-island formation and vasculogenesis in Flk-1-deficient mice.Nature 1995;376:62–6.
26] Chen L, Tanriover G, Yano H, Friedlander R, Louvi A, Gunel M. Apoptotic func-tions of PDCD10/CCM3, the gene mutated in cerebral cavernous malformation3. Stroke 2009;40:1474–81.
27] Nebreda AR, Porras A. p38 MAP kinases: beyond the stress response. Trends inBiochemical Sciences 2000;25:257–60.
28] Stockton RA, Shenkar R, Awad IA, Ginsberg MH. Cerebral cavernous malfor-mations proteins inhibit Rho kinase to stabilize vascular integrity. Journal ofExperimental Medicine 2010;207:881–96.
29] He Y, Zhang H, Yu L, Gunel M, Boggon TJ, Chen H, et al. Stabilization of VEGFR2
signaling by cerebral cavernous malformation 3 is critical for vascular devel-opment. Science Signaling 2010;3:ra26.30] Sato TN, Tozawa Y, Deutsch U, Wolburg-Buchholz K, Fujiwara Y, Gendron-Maguire M, et al. Distinct roles of the receptor tyrosine kinases Tie-1 and Tie-2in blood vessel formation. Nature 1995;376:70–4.




















