
Mouse chromosome 3
11
Mammalian Genome 4, $47-$57 (1993). 9 Springer-Vedag NewYork Inc. 1993 Mouse Chromosome 3 Michael F. Seldin, 1 Jan-Bas Prins, z Nanda R. Rodrigues, z John A. Todd, z Miriam H. Meisler 3'* 1Duke University Medical Center, Departments of Medicine and Microbiology, Research Drive, Room 247, CARL Building, Box 3380, Durham, North Carolina 27710-3380, USA 2Nuffield Department of Surgery, John Radcliffe Hospital, Headington, Oxford OX3 9 DU, UK 3Department of Human Genetics, University of Michigan, Ann Arbor, Michigan 48109-0618, USA Received: 10 May 1993 Introduction During the past year, more than 80 new loci have been assigned to Chromosome (Chr) 3, including 15 genes and a large number of anonymous DNA markers. This wealth of markers makes it increasingly difficult to represent the genetic maps of mouse chromosomes in traditional formats. In this report, map positions for Chr 3 loci are presented in tabular form. A traditional graphic representation of Chr 3 is available in the first and second committee reports (103, 103a). This report provides an updated locus list and map, updated strain distribution patterns for recombinant inbred lines, recombination data from seven large mul- tilocus crosses, a new map entirely based on PCR- based microsatellite loci that span Chr 3, and primer sequences for a large number of markers that can be detected by the PCR. Locus list and chromosome map The main features of Chr 3 loci are presented in Table 1. Map positions were calculated by Seldin using the methods previously described (103,103a). Entries that have been added or changed since the previous report are marked with an asterisk. The map positions are composites based on a large number of measurements. The 95% confidence intervals for the primary data are in most cases greater than 2 cM, and those for the composite data are greater than 5 cM. As discussed in the previous reports, there may be errors in the indi- cated gene orders for closely linked loci that have not been mapped in the same cross. * Chair of Committee for Mouse Chromosome 3 The map positions that are listed alphabetically in Table 1 are also listed in order of increasing distance from the centromere in Table 2, providing a tabular representation of the Chr 3 map. Data on gene order obtained from seven multilocus crosses are summarized in Fig. 1A. Unambiguous gene order can be determined only for loci that were mapped within the same cross. The data from the seven crosses generate a consistent gene order. The quantitative data from these crosses are presented in Fig. lB. PCR primers for Chr 3 loci The development of PCR-based assays that detect ge- netic variation has greatly reduced the time and effort required for genotyping, as well as the amount of ge- nomic DNA required per assay. PCR primers amplify products of different lengths as a result of variation in simple sequence repeat length. Published gene se- quences have been used to derive primers that detect variation at known loci. In addition, a large number of primers that detect (CA)n repeat length variation have been developed from anonymous genomic clones by the Mouse Genome Center at the Massachusetts Insti- tute of Technology (MIT) (37). Primer sequences for 72 Chr 3 loci of both types are presented in Table 3. Microsateilite map of Chr 3 During the past year, microsatellite markers have been widely adopted by many investigators as convenient and reliable genetic markers. The presence of varia- tion among inbred strains, the reproducibility of as-
-
Upload
khangminh22 -
Category
Documents
-
view
7 -
download
0
Transcript of Mouse chromosome 3

Mammalian Genome 4, $47-$57 (1993).
�9 Springer-Vedag New York Inc. 1993
Mouse Chromosome 3
Michael F. Seldin, 1 Jan-Bas Prins, z Nanda R. Rodrigues, z John A. Todd, z Miriam H. Meisler 3'*
1Duke University Medical Center, Departments of Medicine and Microbiology, Research Drive, Room 247, CARL Building, Box 3380, Durham, North Carolina 27710-3380, USA 2Nuffield Department of Surgery, John Radcliffe Hospital, Headington, Oxford OX3 9 DU, UK 3Department of Human Genetics, University of Michigan, Ann Arbor, Michigan 48109-0618, USA
Received: 10 May 1993
Introduction
During the past year, more than 80 new loci have been assigned to Chromosome (Chr) 3, including 15 genes and a large number of anonymous DNA markers. This wealth of markers makes it increasingly difficult to represent the genetic maps of mouse chromosomes in traditional formats. In this report, map positions for Chr 3 loci are presented in tabular form. A traditional graphic representation of Chr 3 is available in the first and second committee reports (103, 103a).
This report provides an updated locus list and map, updated strain distribution patterns for recombinant inbred lines, recombination data from seven large mul- tilocus crosses, a new map entirely based on PCR- based microsatellite loci that span Chr 3, and primer sequences for a large number of markers that can be detected by the PCR.
Locus list and chromosome map
The main features of Chr 3 loci are presented in Table 1. Map positions were calculated by Seldin using the methods previously described (103,103a). Entries that have been added or changed since the previous report are marked with an asterisk. The map positions are composites based on a large number of measurements. The 95% confidence intervals for the primary data are in most cases greater than 2 cM, and those for the composite data are greater than 5 cM. As discussed in the previous reports, there may be errors in the indi- cated gene orders for closely linked loci that have not been mapped in the same cross.
* Chair of Committee for Mouse Chromosome 3
The map positions that are listed alphabetically in Table 1 are also listed in order of increasing distance from the centromere in Table 2, providing a tabular representation of the Chr 3 map.
Data on gene order obtained from seven multilocus crosses are summarized in Fig. 1A. Unambiguous gene order can be determined only for loci that were mapped within the same cross. The data from the seven crosses generate a consistent gene order. The quantitative data from these crosses are presented in Fig. lB.
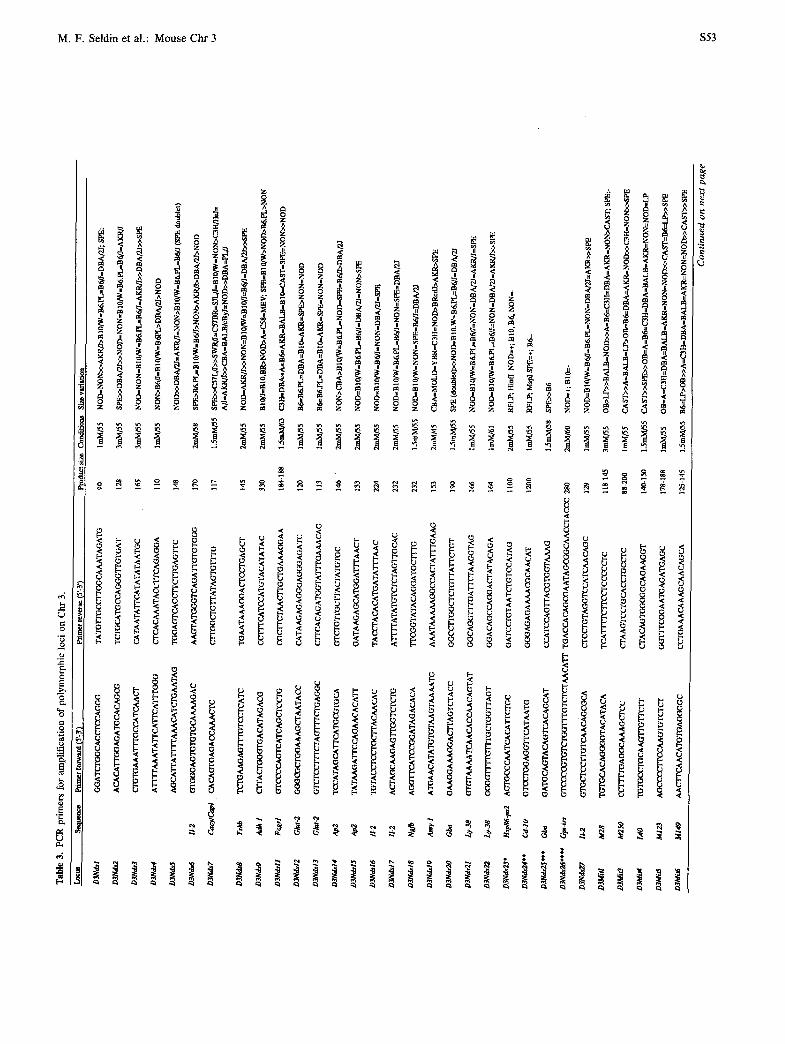
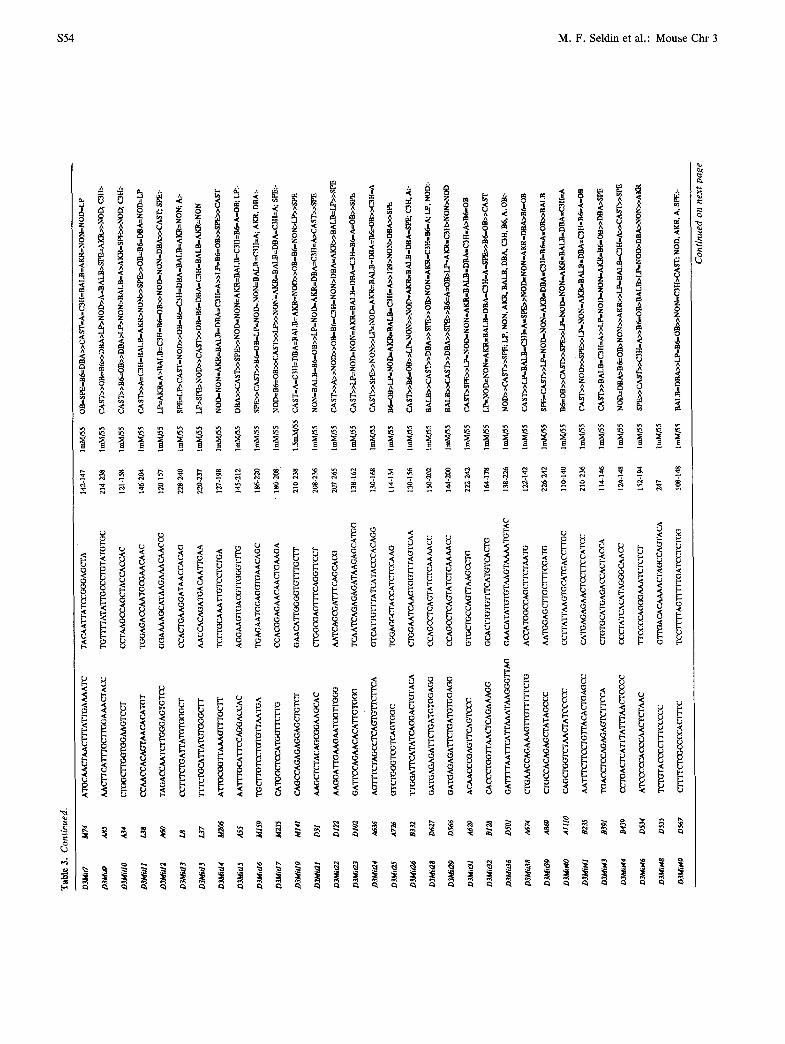
PCR primers for Chr 3 loci
The development of PCR-based assays that detect ge- netic variation has greatly reduced the time and effort required for genotyping, as well as the amount of ge- nomic DNA required per assay. PCR primers amplify products of different lengths as a result of variation in simple sequence repeat length. Published gene se- quences have been used to derive primers that detect variation at known loci. In addition, a large number of primers that detect (CA)n repeat length variation have been developed from anonymous genomic clones by the Mouse Genome Center at the Massachusetts Insti- tute of Technology (MIT) (37). Primer sequences for 72 Chr 3 loci of both types are presented in Table 3.
Microsateilite map of Chr 3
During the past year, microsatellite markers have been widely adopted by many investigators as convenient and reliable genetic markers. The presence of varia- tion among inbred strains, the reproducibility of as-

$48 M . F . S e l d i n e t a l . : M o u s e C h r 3
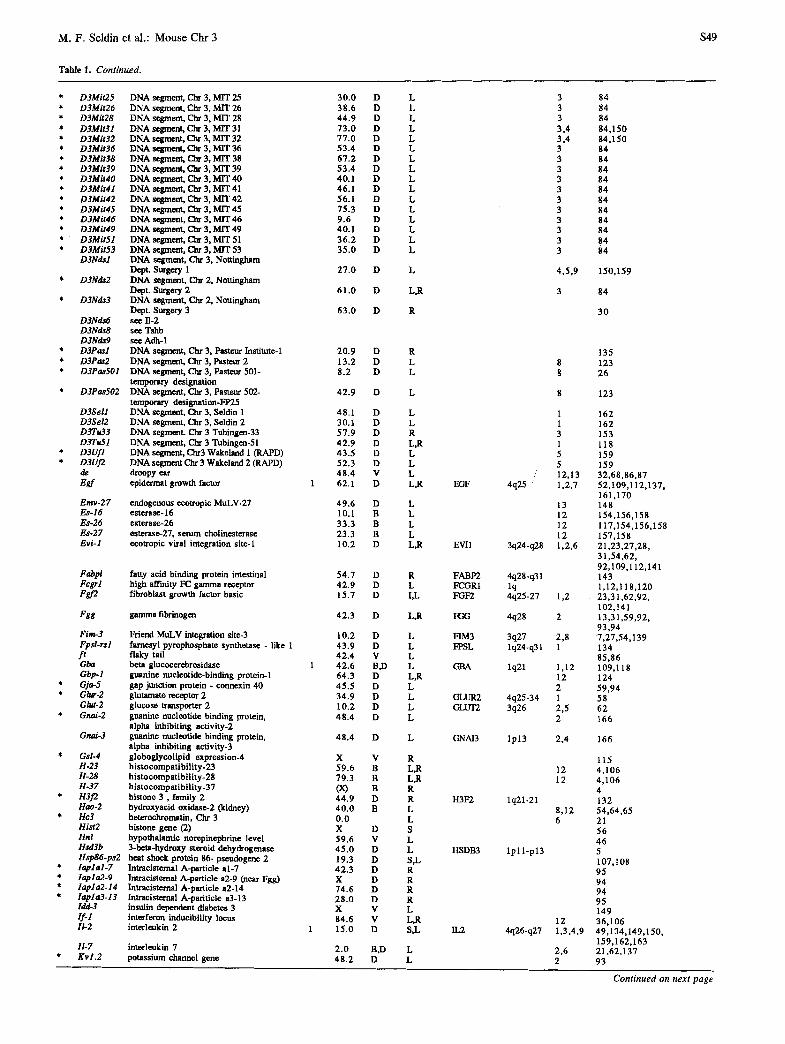
Table 1. Locus fist for mouse Chr 3.
New S~'mbol Name A Map T Method
Acrb-2 acetylcholine receptor beta 2 neural 43.0 D L Acts skeletal alpha actin X D S,R Adh-I alcohol dehydrngenase-1 1 68.1 B,D L,R
Adh-lps alcohol dehydrogenase-1 l~eudogene 52.0 D R Adh-lt alcohol dehydrogenase-tempural 68.1 B L Adh-3 alcohol dehydrogenase-3 68. i B R,L
Adh-3t alcohol dehydrngenaae-3-tempural 68.1 B L Adh-5 alcohol dehydrogenase-5 X B S Ahr-I aldehyde reduetase-1 68.1 B R,L Ampd-1 AMP dearninase-I (muscle form) 47.4 D L Ampd-2 AMP deaminase-2 (nonmnscle form) 50.2 D L Amy-1 amylase, salivary 1 49.6 B,D L,R
Amy-2 amylase, pancreatic 1 49.6 B,D L,R Ank-2 brain specific ankyrin-2 58.4 D L Ap2 adipocyte protein aP2 9.7 B,D R,L Arnt aryl hydocarbon receptor nuclear translator X B S Atplal Na, K ATPase alpha-I 47.3 D L Atpa-I see Atplal Bglap bone matrix Gla protein D S B r a n Beta-marmosidese activity (liver, kidney) 68.1 B R BRS-3 see Odc-rs3 Cacy calcyclin 42.9 D L Calll calpactin I light chain 42.9 D R Calla see Mine Capl calcium binding protein, placental 42.9 D R Car-1 carbonic anhydrase-1 6.3 B L Car-2 carbonic anhydrase-2 1 6.3 B,D L,R
$
Car-3 carbonic anhydrase-3 7.5 D L Ccna Cyclin a 15.0 D L Cdl cluster designation I 43.9 D L Cd2 cluster designation 2 46.4 D L Cdl 0 see Mrae Cd53 cluster designation 53 47.4 D L cdm cadmium resistance 61.6 V R Cf-3 coagulatioon factor 3 49.6 D R Cap-2 cyclic nuclentide phosphodiesterase-2 38.8 D R Cnx40 connexin (see Gja-5) coa cocoa 8.3 V L Csfm colony stimulating factor, macrophage 48.4 B,D,V L
(alternative for op) D3Byul DNA segment, Brig Young Univ. 1 (RAPD) 6.9 D R D3Byu2 DNA segment, Brig Young Univ. 2 (RAPD 6.9 D R D3Byu3 DNA segment, Brig Young Univ. 3 (RAPD) 8.1 D R D3Byu4 DNA segment, Brig Young Univ. 4 (RAPD) 45.3 D R D3Byu5 DNA segment, Brig Young Univ. 5 (RAID) 34.4 D R D3Byu6 DNA segment, Brig Young Univ. 6 (RAPD) 42.3 D R D3Byul 7 DNA segment, Brig Young Univ. 17 (RAPD) 30.1 D R D3Dol P40-4 done ? high affinity laminin 76.4 D R D3Hunl DNA segment, Chr 3, Hunter 1 56.1 D L D3J1 DNA segment, Chr 3, Jackson Lab 1 33.1 D R D3J2 DNA segment, Chr 3, Jackson Lab 2 65.6 D R D3J3 DNA segment, Chr 3, Jackson Lab 3 54.7 D R D3Jfrl DNA segment, Oar 3, MJeffers-1 47.4 D P D31knl DNA segment, Chr 3, Ian Jackson 1 69.0 D R D3Lehl DNA segment, Cbr 3, Lehrach 1 17.6 D L D3Leh2 DNA segment, Chr 3, Lehrach 2 10.2 D L D3Lerl DNA segment, Chr 3, Leroy 1 41.5 D L D3Ler2 DNA segment, Chr 3, Leroy 2 41.5 D L D3Mitl DNA segment, Chr 3, M1T 1 7 .0 D L D3Mit3 DNA segment, oar 3, MIT 3 20.0 D L D3Mit4 DNA segment, Chr 3, M_IT 4 20.0 D L D3Mit5 DNA segment, Chr 3, MIT 5 1 24.0 D L D3Mit6 DNA segment, oar 3, MIT 6 22.0 D L D3Mit7 DNA segment, Oar 3, MIT 7 30.0 D L D3Mit9 DNA segment, Oar 3, M1T 9 38.6 D L D3MitlO DNA segmctat, Chr 3, MIT 10 48.6 D L D3Mitll DNA segment, Chr 3, MIT 11 48.6 D L D3Mitl2 DNA segmen4 Chf 3, MIT 12 50.3 D L D3Mitl3 DNA segment, Chr 3, MIT 1 57.7 D L D3Mitl4 DNA segment, Chr 3, MIT 14 61.0 D L D3Mitl5 DNA segment, Chr 3, M1T 15 63.1 D L D3Mit16 DNA segment, Chr 3, MIT 16 63.1 D L D3Mit17 DNA segment, Chr 3, MIT 17 68.7 D L D3Mit18 DNA segment, Chr 3, MIT 18 73.0 D L D3Mit19 DNA segment, Chr 3, MIT 19 1 81.0 D L D3Mit21 see 11-2 D3Mit22 DNA segment, Cbr 3, MIT 22 1 35.0 D L D3Mit23 DNA segment, Oar 3, MIT 23 0 .0 D L D3Mit24 DNA segment, Cb.r 3, M r r 24 20.0 D L
H. s]tmbol
ACTAI ADHI
ADH3
ADH5
AMPD1
AMY1
AMY2
ARNT ATP1A1
BGLAP
CACY
C A ~ CA1 CA2
CA3
CD1 CD2
F3
CSF1.
H. location Notes References
8 11 lp21-qter 8,12 33 4q21-q23 4,5 13,15,20,22,54,
68,150,159 20
13 6,71 4q21 -q23 12 43,67,68,69,72,
112,124 12 66,69
4q21-q25 53 12 43
lp13 1 79 1 79,111,118
lp21 8.10,12 13,15,45,52. 59,86,121. 137,138,156
lp21 1,2 13,14,109,121 12 122 4 61,150
lpter-q12 17 lp13 1.13 77,79,81,109
76 97
1q21-q25 1,12 39,79,109,118 131,151
lq12-q22 39,151 8q13-q22 13 44 8q13-q22 1,7,8,13 15,23,28,44,
47,109, i17,121,156,161
8qI3-q22 13 9 1,2 92,129,134
lq22-23 1 23,110,111,118 Ip13 1 78,79,109,118
1 168 145,146
lp22-p21 98 10
13 117,142 lp13-21 2,8 18,54,86,93,169
167 167 167 167 167 167 167 40
1 73 114 114 114 75 99
2 31 2 31 8 91 8 91 3,4,5 37,150,159 3 37 3 37 3,4 37,150 3 37,150 3 37 3 37 4,5 37,159 3,4 37,150 3 37 3 84 3 37 3 37 3 37 3 37 1,3 37,134,161 1,3,4 134,150,159
3,4 84,150 3 84 3 84
Continued on next page
M. F. Seldin et al.: Mouse Chr 3
Table 1. Continued.
$49
* D3Mit25 DNA segment, Chr 3, MIT 25 30.0 D L * D3Mit26 DNA segment, Chr 3, MIT 26 38.6 D L * D3Mit28 DNA segment, Chr 3, MIT 28 44.9 D L * D3Mit31 DNA segment, Chr 3, MIT 31 73.0 D L * D3Mit32 DNA segment, Chr 3, MIT 32 77.0 D L * DJMit36 DNA segment, Chr 3, MIT 36 53.4 D L * D3Mit38 DNA segment, Cl3r 3, MIT 38 67.2 D L * D3Mit39 DNA segment, Chr 3, MIT 39 53.4 D L * D3MiI40 DNA segment, Chr 3, MIT 40 40.1 D L * DJMit41 DNA segment, Chr 3, M1T 41 46.1 D L * DJMit42 DNA segment, Chr 3, MIT 42 56.1 D L * D3Mit45 DNA segment, Chr 3, M1T 45 75.3 D L * D3Mit46 DNA segraent, Chr 3, MIT 46 9.6 D L * D3Mit49 DNA segment, Chr 3, M1T 49 40.1 D L * D3Mit51 DNA segment, Chr 3, MIT 51 36.2 D L * D3Mit53 DNA segment, Chr 3, M1T 53 35.0 D L
D3Ndsl DNA segment, Chr 3, Nottingham Dept. Surgery 1 27.0 D L
* D3Nds2 DNA segment, C~" 2, Nottingham Dept. Surgery 2 61.0 D L,R
* D3Nds3 DNA segment, Chr 2, Nottingham DepL Surgery 3 63.0 D R
D3Nds8 see II-2 D3Nds8 see Tshb D3Nds9 see Adh-I
* D3Pasl DNA segment, Chr 3, Pasteur Institute-1 20.9 D R * D3Pas2 DNA segment, Chr 3, Pastetw 2 13.2 D L * D3PasS01 DNA segment, Chr 3, Pasteur 501- 8.2 D L
temporary designation * D3Pas502 DNA segment, Chr 3, Pasteur 502- 42.9 D L
temporary designation-FP25 D3$ell DNA segment, Chr 3, Seldin 1 48.1 D L D3Sel2 DNA segment, Chr 3, Seldin 2 30.1 D L D3Tu33 DNA segment. Chr 3 Tubingen-33 57.9 D R D3Tu51 DNA segment, Chr 3 Tubingen-51 42.9 D L,R
* D3Ufl DNA segment, Chr3 Wakeland 1 (RAPD) 43.5 D L * D3Uf2 DNA segment Chr 3 Wakeland 2 (RAPE)) 52.3 D L
de droopy ear 48,4 V L Egf epidermal growth factor I 62.1 D L,R
Emv-27 endogenous r MuLV-27 49.6 D L Es-16 esterase- 16 10.1 B L Es-26 esterase-26 33.3 B L Es-27 esterase-27, serum eholinesterase 23.3 B L Evi-I ecotropic viral integration sited 10.2 D L,R
Fabpi fatty acid binding protein intestinal 54.7 D R Fcgrl high affinity FC gamma receptor 42.9 D L Fgr fibroblast growth factor basic 15.7 D I,L
Fgg gamma fibrinogen 42.3 D L,R
Fire-3 Friend MuLV integration site-3 10.2 D L Fpsl.rsl famesyl pyrophosphate synthetase - like 1 43.9 D L f t flaky tail 42.4 V L Gba beta glueocerebrosidase 1 42.6 B,D L Gbp-1 guanine nuelentide-binding protein-1 64.3 D L,R
* Gjw5 gap junction protein - connexin 40 45.5 D L * Glut.2 glutamate receptor 2 34.9 D L
Glut-2 glucose transporter 2 10.2 D L * Gnai-2 guanine nucleotide binding protein, 48.4 D L
alpha inhibiting activity-2 Gnal-3 guanine nueleotide binding protein, 48.4 D L
alpha inhibiting activity-3 * Gsl.4 globoglycolipid expression-4 X V R
H-23 histocompatibility-23 59.6 B L,R H-28 histoeompatibility-28 79.3 B L,R H-37 histoeompatibility-37 CA") :8 R
* H3f2 histone 3 , family 2 44.9 D R Hao-2 hydroxyacid oxidase-2 (kidney) 40.0 B L
* He3 heterochromatin, Chr 3 0.0 L Hist2 histone gene (2) X D S Hnl hypothalamie norepinephrine level 59.6 V L Hsd3b 3-beta-hydroxy steroid dehydrogeaaase 45.0 D L Hsp86-ps2 beat shock protein 86- pseudogene 2 19.3 D S,L
* lap la l -7 Intraeistemal A-particle al-7 42.3 D R * lapin2-9 Intracisternal A-particle a2-9 (near Fgg) X D R * lapla2-14 Intracisternal A-particle a2-14 74.6 D R * lapla3-13 Intraeisternal A-pariticle a3-13 28.0 D R
ldd.3 insulin dependent diabetes 3 X V L If-1 interferon indneibility locus 84.6 V L,R II-2 interleukin 2 1 15.0 D S,L
II-7 interleukin 7 2.0 B,D L * K v l . 2 potassium channel gene 48.2 D L
BGF
EVIl
FABP2 FCGR1 FGF2
FIM3 F I l L
GIlA
GLUR2 GLIYI'2
GNAI3
H3F2
HSDB3
11.2
4q25
3q24-q28
4q28-q31 lq 4q25-27
4q28
3q27 lq24-q31
lq21
4q25-34 3q26
lp13
lq21-21
1 p l l - p l 3
4q26-q27
3 3 3 3,4 3,4 3 3 3 3 3 3 3 3 3 3 3
4,5,9
3
8 8
8
1 1 3 1 5 5 12,13 1,2,7
13 12 12 12 1,2,6
1,2
2
2,8 1
1,12 12 2 1 2,5 2
2,4
12 12
8,12 6
12 1,3,4,9
2,6 2
84 84 84 84,150 84,150 84 84 84 84 84 84 84 84 84 84 84
150,159
84
3 0
135 123 26
123
162 162 153 118 159 159 32,68,86,87 52,109,112,137, 161,170 148 154,156,158 117,154,156,158 157,158 21,23,27,28, 31,54,62, 92,109,112,141 143 1,12,118,120 23,31,62,92, 102,141 13,31,59,92, 93,94 7,27,54,139 134 85,86 109,118 124 59,94 58 62 166
166
115 4,106 4,106 4 132 54,64,65 21 56 46 5 107,108 95 94 94 95 149 36,106 49,134,149,150, 159,162,163 21,62.137 93
Continued on next page

$50
Table 1. Continued.
M. F. Se ld in et al . : M o u s e C h r 3
* Kvl .3 potassium channel gene 48.2 D L * Kvl.3rs3 potassium channel gene 1.3 related sequence 48.4 D L
Lef-1 lymphoid enhancer-binding factor 1 X D R,S * M6pr-ps cation-dependent mannose 6- 45.5 D L
phosphate receptor pseudo gene ma matted 40.4 V L Mine membrane metallo-endo peptidase 30.1 D L
(neutral endopeptidase) Mrav-2 MCF endogenous viros-2 X D S Mmv-12 MCF endogenous vires-12 X D S Mov-lO Moloney leukemia virus-10 X D S Mpmv-9 modified polytropic murine leukemia virus-9 88.2 D L,R Mpmv-20 modified polytropic murine leukemia virus-20 9.6 D R my blebs 30.4 V L Ngfb nerve growth factor beta 47.4 D L
Nras Nras oncogene 47.4 D L * Nsel-2 see Tau-1
Oat-rs2 orolthine aminotransferase related sequence 2 52.4 D L Odc-r$3 omithine decarboxylase-3 X D R op osteop~rosis (Csfm mutation see Csfm) Otf-3rs3 octamer transcription factor -3 related sequence 3 0,0 D L Otf-3rs4 octamer transcription factor-3 related sequence 4 62.1 D
* Otf-3rs9 octarner transcription factor-3 related sequence 9 16.4 D Oua-I ousbain resistance- 1 X V S Pgk-lps3 phosphoglycerate kinase-1 pseudogene 3 8.7 D S,R Pk-1 pyruvate kinase (may be the same as Pklr) 33.6 B L Pk./r pyruvate kinase liver, red blood cells (see Pk-1) 42.6 D
* Prop-1 peroxisomal membrane protein C/0k) 54.6 D L Pray-26 polytropic routine virus-26 71.8 D R Pray-28 polytropic marine virus-28 42.9 D R Pray-38 polytropic marine virus-38 43.1 D R Pray-39 polytropic routine virus-39 53.8 D R Rapla member of RAS oncogene family 47.4 D L rcm rostral cerebellar malformation 65.4 V L
* Rn7s-3 7s RNA related sequence -3 63.0 D R Rnulb-1 Ulbl small nuclear RNA 42.7 D R Rnulb-3 Ulb3 small nuclear RNA 42.9 D R soc soft coat 43.4 V L spa spastic 37.4 V L suc-1 see Suc- lr Sac-lr sacrase-isomaltase, regulatory 33.3 B L Sac.Is sucrase-isomaltase, structural 33.3 D R sut subtle gray 12.2 V L Tau-1 basic domain helix-loop-helix CoHLH') 50.6 D L
* Thbs3 thrombospondin 3 43.9 D L * Tkr tyrosine kinase receptor (Ngf is ligand) 43.9 D L
Tmevd-2 TMEV induced demyelinating disease susceptibility 8.4 V R
* Tpi-2 triosephosphate isomerase related sequence-2 36.4 D L
Tshb thyrotropin stimulating hormone beta subunit 1 47.4 D L
Va varitint-waddler 71.6 V L
Xmmv-22 xenotropic-MCF leukemia virus - 22 42.3 D R Xmmv-47 xenotropic-MCF leukemia virus - 47 31.0 D R Xmmv-65 xenotropic-MCF leukemia virus - 65 42.3 D L,R
* Ybld YB-I DNA binding protein related sequence d 14.6 D L
2 93 2 93
LEF1 4q23-q25 105 2 94
10,12 85,86,87,106 MME 3q21-27 1,7 23,161
63 63 74,113
11 51 51
10 19,35,45 NGFB lp13 1,2 18,42,52,59,79,94,
109,118,166,171 NRAS lp13 1,2 18,93,118,128
125 126
2,7 137,161 L 1,2,7,137,161 L 7,161
83 1
8 54,139 L PKLR lq21 1,118 PMP1 lp21-22 2 52
5O 50 5O 50
RAPIA lp12-p13 1 41 12 89,90
147 RNU1 96
13,96 10 45,140 10 85,86
12 SI 3q25-26 12
12 88 12 55 1 38 1 80
104
2 138
TSI-IB lp13 1 13,42,79,82,109, 116,118,150,159
10,12 32,43,45,67,68, 86,87,106 13 164
12 164 2 141
The Chr 3 map positions are an estimate of distances, in cM, from the cen- tromere. The position of the centromere is determined by heterochromatin mapping data (21,101). Since recombination frequencies may vary depending on the specific cross, composite map positions may distort gene order when loci have not been mapped in an individual backcross. In deriving the com- posite map, RI strain data was used to determine gene position only as a supplement to backcross data. For a fuller discussion of the generation of map positions, see text and (133). In addition to the references cited for each locus, data used to derive map positions are described in the following "Notes : " (1) Duke University cross: complete haplotypes in 114 and incomplete haplo- types in 338 interspecific [(C3H/HeJ-g/d • M. spretus)F 1 • C3H/HeJ-gld] backcross mice (134); ( la) same cross as above, but fewer than 40 meiotic events examined; (2) Frederick cross: complete haplotypes in 83-198 inter- specific [(C57BL/6J • M. spretus)F1 • C57BL/6J] backcross mice; (2a) same cross as 2, but fewer than 50 meiotic events examined; (3) Rockefeller Uni- versity cross and MIT microsatellite mapping panel: complete haplotype data
in 40-48 (C57BL/6J-ob • CAST/Ei)F 2 intersubspecific intercross mice; (4) Eu- ropean Collaborative Interspecific Backcross [(C57BL/6 x SPR)F~ x SPR] produced by the UK's Human Genome Mapping Project, with the support of the Medical Research Council (15); (5) data from the cross [(NOD/Uf x C57BL/6J)F 1 • NOD/Uf] from the laboratory of E.K. Wakeland. The RAPD polymorphism D3Ufl was detected with the primer GTGCCTAACC, and D3 Uf2 with the primer TGCTCACTGA; (6) RPMI cross: complete haplotypes in 130--140 [(C57BL/6 • M. spretus)Fl • M. spretus]; (7) Duke University cross #2: complete haplotype data in 100-182 [(MRL/MpJ-Ipr • CAST/Ei)FI x MRL/MpJ-lpr] intersubspecific backcross mice (161); (8) Pasteur cross: incomplete haplotypes in 38-74 interspecific backcross mice (J.-L. Gurnet, unpublished data); (9) same as 4 with complete haplotype data in 92-299 mice; (10) included in nine overlapping three- or four-point crosses that derive from analysis of 125-500 meiotic events in each of multiple individual crosses; (11) haplotype data in 75 interspecific backcross mice (W.N. Frankel, unpublished data); (12) three-point mapping data; (13) two-point mapping data.
M. F. Seldin et al.: Mouse Chr 3 $51
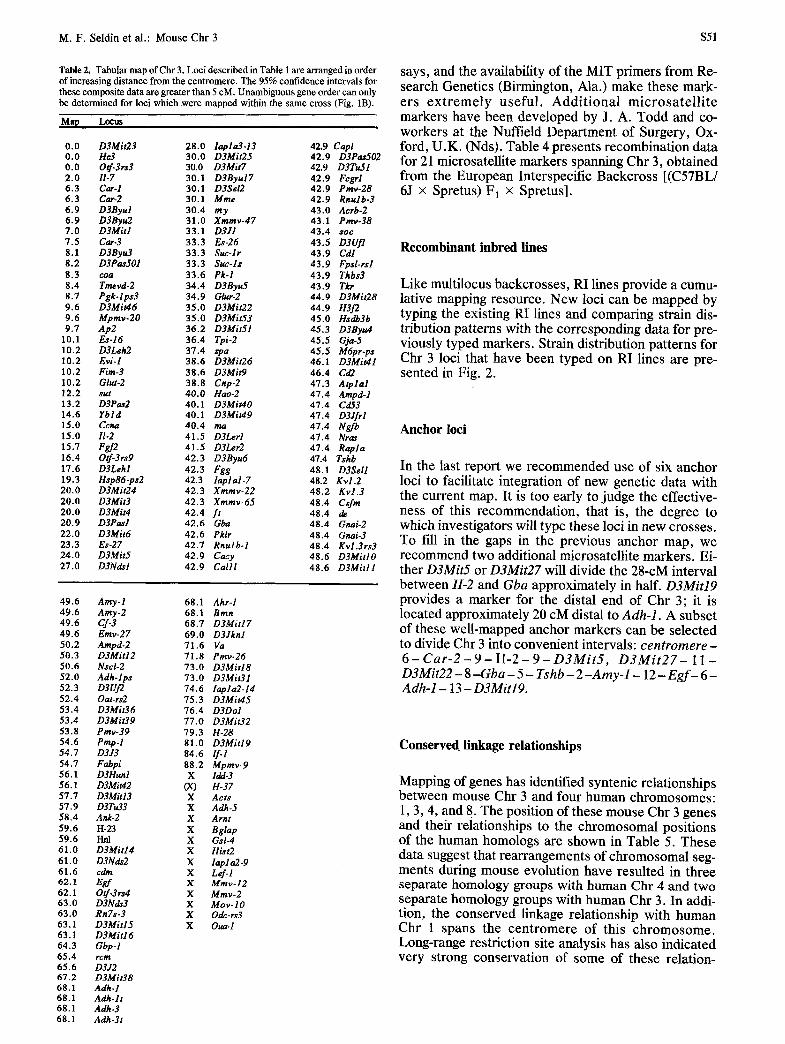
Table 2. Tabular map of Chr 3. Loci described in Table 1 are arranged in order of increasing distance from the centromere. The 95% confidence intervals for these composite data are greater than 5 cM. Unambiguous gene order can only be determined for loci which were mapped within the same cross (Fig. 1B).
Map Locus
0.0 D3Mit23 28.0 lapla3.13 42.9 Capl 0.0 He3 30.0 D3Mit25 42.9 D3Pas502 0.0 Otf-3rs3 30.0 D3Mit7 42.9 D3Tugl 2.0 II-7 30.1 D3Byu17 42.9 Fcgrl 6.3 Card 30.1 D3Sel2 42.9 Pray-28 6.3 Car-2 30.1 Mine 42.9 Rnulb-3 6.9 D3Byul 30.4 my 43.0 Acrb-2 6.9 D3Byu2 31.0 Xmmv-47 43.1 Pmv-38 7.0 D3Mitl 33.1 D3JI 43.4 s o c
7.5 Car-3 33.3 Es-26 43.5 D3Ufl 8.1 D3Bynd 33.3 Suc-lr 43.9 Cd/ 8.2 D3PasSOl 33.3 Suc-ls 43.9 Fpsl-rsl 8.3 coa 33.6 Pk-I 43.9 Thbs3 8.4 Tmevd-2 34.4 D3Byug 43.9 Tkr 8.7 Pgk-lps3 34.9 Glut-2 44.9 D3Mit28 9.6 D3MiuI6 35.0 D3Mit22 44.9 H3f2 9.6 Mpmv-20 35.0 D3Mit53 45.0 Hsdb3b 9.7 Ap2 36.2 D3MitSl 45.3 D3Byu4
10.1 Es-16 36.4 Tpi-2 45.5 Gja-5 10.2 D3Leh2 37.4 spa 45.5 M@r-ps 10.2 Evi-I 38.6 D3Mit26 46.1 D3Mit41 10.2 Fire-3 38.6 D3Mit9 46.4 Cd2 10.2 Glut-2 38.8 Cnp-2 47.3 Atplal 12.2 sut 40.0 Hao-2 47.4 Ampd-I 13.2 D3Pas2 40.1 D3Mit40 47.4 Cd53 14.6 Ybld 40.1 D3Mit49 47.4 D3Jfrl 15.0 Ccna 40.4 ma 47.4 Ngfb 15.0 II-2 41.5 D3Lerl 47.4 Nras 15.7 Fgf2 41.5 D3Ler2 47.4 Rapla 16.4 Otf-3rs9 42.3 D3Byu6 47.4 Tshb 17.6 D3Lehl 42.3 Fgg 48.1 D3Sell 19.3 Hsp86-ps2 42.3 iaplal-7 48.2 Kvl.2 20.0 D3Mit24 42.3 Xmmv-22 48.2 Kvl .3 20.0 D3Mit3 42.3 Xmmv-65 48.4 Csfm 20.0 D3Mit4 42.4 f t 48.4 de 20.9 D3Pasl 42.6 Gba 48.4 Gnai-2 22.0 D3Mit6 42.6 Pklr 48.4 Gnai-3 23.3 Es-27 42.7 Rnulb-1 48.4 Kvl.3rs3 24.0 D3Mit5 42.9 Cacy 48.6 D3MiHO 27.0 D3Ndsl 42.9 Calll 48.6 D3Mitll
Amy. l 68.1 Ahr-1 Aray-2 68.1 Bran Cf-3 68.7 D3Mitl7 Emv-27 69.0 D3Jknl Ampd-2 71.6 Va D3Mitl2 71.8 Pray-26 Nscl-2 73.0 D3Mitl8 Adh-lps 73.0 D3Mit31 D3Uf2 74.6 lapla2-14 Oat-rs2 75.3 D3Mit45 D3Mit36 76.4 D3Dol D3Mit39 77.0 D3Mit32 Pray-39 79.3 H-28 Pmp-I 81.0 D3Mitl9 D3J3 84.6 If.1 Fabpi 88.2 Mpmv-9 D3Hunl X ldd-3 D3Mit42 (X) H-37 D3Mit13 X Acts D3Tu33 X Adh-5 Ank-2 X Arnt H-23 X Bglap Hill X G s l-4 D3Mit14 X H&t2 D3Nds2 X lapl a2.9 cdm X Lef-I Egf X Mmv-12 00r X Mmv-2 D3Nds3 X Mov- l 0 RnTs-3 X Odc-rs3 D3Mit15 X Oua-I D3Mitl 6 Gbp-I rcrtz D3J2 D3Mit38 Adh-I Adh-lt Adh.3 Adh-3t
says, and the availability of the MIT primers from Re- search Genetics (Birmington, Ala.) make these mark- ers ex t remely useful. Addit ional microsate l l i te markers have been developed by J. A. Todd and co- workers at the Nuffield Department of Surgery, Ox- ford, U.K. (Nds). Table 4 presents recombination data for 21 microsatellite markers spanning Chr 3, obtained from the European Interspecific Backcross [(C57BL/ 6J • Spretus)F1 • Spretus].
Recombinant inbred lines
Like multilocus backcrosses, RI lines provide a cumu- lative mapping resource. New loci can be mapped by typing the existing RI lines and comparing strain dis- tribution patterns with the corresponding data for pre- viously typed markers. Strain distribution patterns for Chr 3 loci that have been typed on RI lines are pre- sented in Fig. 2.
Anchor loci
In the last report we recommended use of six anchor loci to facilitate integration of new genetic data with the current map. It is too early to judge the effective- ness of this recommendation, that is, the degree to which investigators will type these loci in new crosses. To fill in the gaps in the previous anchor map, we recommend two additional microsatellite markers. Ei- ther D 3 M i t 5 or D 3 M i t 2 7 will divide the 28-cM interval between 11-2 and G b a approximately in half. D 3 M i t 1 9
provides a marker for the distal end of Chr 3; it is located approximately 20 cM distal to A d h - 1 . A subset of these well-mapped anchor markers can be selected to divide Chr 3 into convenient intervals: c e n t r o m e r e -
6 - C a r - 2 - 9 - 11-2 - 9 - D 3 M i t 5 , D 3 M i t 2 7 - I 1 -
D 3 M i t 2 2 - 8 - G b a - 5 - T s h b - 2 - A m y - 1 - 12 - E g f - 6 - A d h - 1 - 13 - D 3 M i t l 9 .
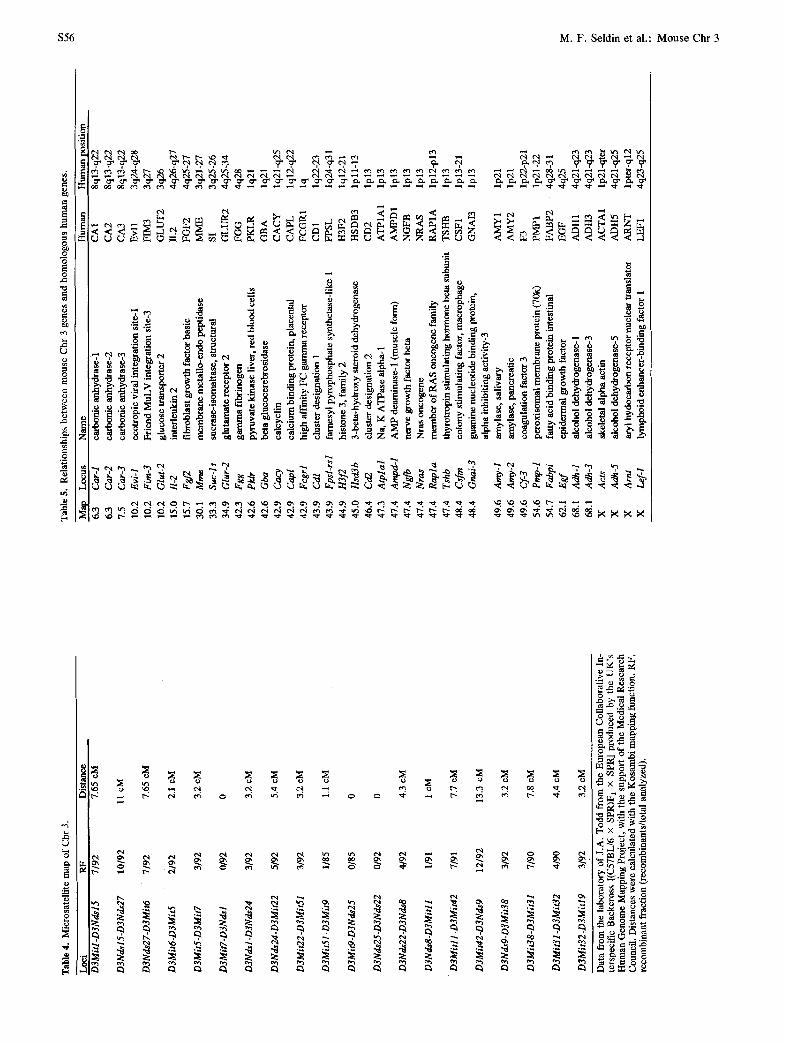
Conserved linkage relationships
Mapping of genes has identified syntenic relationships between mouse Chr 3 and four human chromosomes: 1, 3, 4, and 8. The position of these mouse Chr 3 genes and their relationships to the chromosomal positions of the human homologs are shown in Table 5. These data suggest that rearrangements of chromosomal seg- ments during mouse evolution have resulted in three separate homology groups with human Chr 4 and two separate homology groups with human Chr 3. In addi- tion, the conserved linkage relationship with human Chr 1 spans the centromere of this chromosome. Long-range restriction site analysis has also indicated very strong conservation of some of these relation-

$52 M . F . Seldin et al.: Mouse Chr 3
A I c. " ~ - ~ ~ I I I I
o r - a r ~ [ ] I I I I I I I ~ - ~ i 1 4 1 ) I r I I J I I
.Lt lt
I c-~-1 m m I I I i i-I I Nras I - - ~ I I I I I ~ ~ I I I I I I napla I � 9 I I I I I ] Gt~-2 I �9 I I I I �9 I rst~ I [ ] r I I I �9
�9 I l l I I l l l
IO3Mn111 I I I I ~ �9
Ampd-21 �9 [ I I I I I ~nY-2 i � 9 1 4 9
D~urz I I 1 [ I
~ ' ~ l I1~ I I ~ 1 1 1 'l I D 3 M ~ I ' �9 I I IO~n'~l I �9 I (
~l~r-2 �9 I I I I p,-np-1 I �9 I I I I D3Mt122 I [ ] [ ] D~/1i142 �9 I
o ~ , f ~ , ~ I �9 I D ~ I I ~ I I
,.,,, �9 . i ,.,,,,m". I 1 � 9
U.dl l D3Mtt18 D3Mit31
Cacy D3Mit45 | ' , D3ruS ~ �9 II D3M~32
D3Ufl
o-- II , IIII
B
5 6
I
I_j_ JI
I I m
1 I
CROSS 1: (C3HIHeJ-gldx M. spretuS)F1 x C3H/HeJ-gld (n = 117 to 338)
C a r - 2 - 3 . 5 - Evi-1 - 2 . 6 - 11-2 - 0 - Cyca - 0 . 9 - F g f 2 - 13 .5 - M m e - 4 .4 - G l u r - 2 - 7 . 1 0 -
G b a - 0 . 9 - Thbs3 - 0 - Pk i r - O - F s p h r s l - O - C d l - 0 - T rk - 0 . 6 - D 3 T u 5 1 - 0 - C a c y
- 0 . 9 - F c g r l - 2 . 6 - C d 2 - 0 . 9 - A t p l a l - O.3 - N ras - 0 - C d 5 3 - 0 - Ng fb - 0 - Tshb - 0
- A m p d - 1 - 1 .8 - A m y - 2 - 0 . 6 - A m p d - 2 - 13 .4 - E g f - 11 ,7 - D 3 M i t 1 8 - 9 . 3 - D 3 M i t 1 9
CROSS 9: (C57BL/SJ x M. spretus)F1 x C57BL/6J (n = 83 to 198)
Ot l -3 rs3 - 2 . 0 - 11-7 - 7 . 3 - Evi-1 - 0 - F im-3 - O - G lu t2 - 7 . 6 - Ybld - O - C c n a - 0 - Fg f2
- 2 1 . 8 - T p i - 2 - 6 . 0 - F g g - 6 . 8 - G ja -5 - 0 - M 6 p r - p s - 5 . 2 - N g f b - O - N r a s - 2 . 2 -
K v l . 2 - 0 - K v l , 3 - 0.6 - Csfm - 0 - K v l . 3 - r s 3 - 0 - G n a i - 2 - 0 - Gna i -3 - 1 . 5 . A m y - 2 -
4 , 9 - Pmp-1 - 5 . 5 - E g f - O - Ot f -3rs4
C R O S S 3: ( C 5 7 B L / 6 J - o b x CAST/Ei)F2 (n = 40 1o 48)
D3Mit23 - 6.8 - D3Mi t l - 1.4 - D3Mit46 - 2.4 - 11-2 - 2 .2 - D3Mi t4 - 0 - D3Mit3 - 0 -
D3Mi t24 - 1.1 - D3Mit5 - 0 - D3Mit6 - 4.7 - D3Mi t7 - O - D3Mit25 - 3.4 - D3Mit22 - 0 -
D3Mi t53 : 1.1 - D3Mit51 - 2.3 - D3Mi t26 - 0 - D3Mit9 - 1.4 - D3Mit40 - 0 - D3Mit49 -
4 6 - D3Mi t28 - 1.1 - D3Mit41 - 2 . 3 - D 3 M i t l 0 - 0 - D3Mit11 - 1.1 - D3Mi t12 - 1 .8 -
D3Mi t39 - 0.5 - D3Mi t36 - 1.8 - D3Mit42 - 1.1 - D3Mit13 - 2 .2 - D3Nds2 - 0 - D2Mi t14
- 1 . 1 - D3Mit16 - 0 - D3Mft15 - 3 . 5 - D3Mit38 - 1 . 2 - D 3 M i t 1 7 - 4 . 3 - D3Mit81 - 0 -
D3Mi t18 - 3.1 - D3Mi t45 - 2 .4 - D3Mi t32 - 7.3 -D3Mi t19
CROSS 4: C57BL/6J x SPR)F1 x S P R : D 3 M i t l - 7 ,6 - Ap2 - 10.8 - 11-2 - 7.6 -
DSMit6 - 2 . 0 - D3Mi t5 - 3 . 0 - D 3 N d s l - 8 . 6 - D3Mit22 - 8 . 6 - T s h b - 1 . 0 - D3Mit11 - 1 . 9
- Adh-1 - 1 0 , 8 - D 3 M i t 3 1 - 4 . 3 - D3Mit32 - 3 , 0 - D3Mit19
C R O S S 5: (NOD/Uf x 057BI - /6J)F1 x NOD/Uf
D3Mi t l - 1 9 . 0 - Glut -2 - 4 . 0 - 11-2 - 6 . 1 - D 3 N d s l - 1 8 . 8 - D3Uf l - 7 . 3 - T ~ h b - 4 . 3 -
D 3 M i t l 0 - 3 .2 - D3Uf2 - 14.1 - Adh-1 - 20.5 - D3Mit19
CROSS 6: (C57BL/6J x M. spretus)F1 x M. spretus (n = 130 to 140)
Hc3 - 0 .7 - 11-7 - 6.9 - Evi-1
CROSS 7: (MRLIMpJ-Ipr x C A S T / E i ) F t x M R L I M p J - I p r : Otf-3rs3 - 6.3 - Car-2 -
10 .6 - Otf-3rs9 - 1 4 . 4 - M m e - 2 4 . 3 - Otf-3rs4 - 0 - Egf - 2 6 . 8 - D3Mit18
Fig. 1. (A) Gene order in seven multilocus crosses. Loci are listed from proximal to distal on the chromosome. The filled boxes represent the loci that were typed in each cross. The shaded boxes represent loci that did not recombine with the locus listed directly above. The observed recombination frequencies from each of these crosses are presented in Fig. lB. See Table 1 and Fig. IB for additional information about the crosses.
Fig. 1. (B) Recombination frequencies in multilocus crosses. Crosses 1-7 are described in Table 1 and Fig. 1A. The approximate lengths, in cM, of the intervals observed in each cross are presented here. We included only crosses with at least six markers analyzed in at least 80 meiotic events , with the exception of cross 6, which provides data for anchoring the composite maps with respect to the centromere. (Note: Cross 4, n = 92)
Tab
le 3
. P
CR
pri
mer
s fo
r am
plif
icat
ion
of p
olym
orph
ic lo
ci o
n C
hr 3
.
Lect
m
Sequ
en~
Prim
er fo
rwar
d (5'-Y)
DJN
dzl
GG
AT
CrG
GC
AC
CT
CC
AG
GG
D3
Nd
s2
AC
AC
A']W
GG
AG
ATG
CA
CA
G @
G
D3N
da3
Cr'G
TG
AA
A'IT
f'GC
CA
TCAACT
D3N
dg4
AI -
ITI'
AA
AT
AT
rCA
TT
CA
~
D3Nds5
AGCATrAI-I- I - I'AAACATCTGAATAG
D3N
d.v6
1
1-2
G
TG
GG
AG
TG
TG
TG
CA
AA
AG
A C
D3
Nd
J7
Ca
cylC
~p
l CA
CA
GTG
AG
AC
CA
AA
CTC
D3N
ds8
Tsh
b
TC
TG
AA
GA
GT
ITG
TC
CT
CA
TC
D3N
d.4)
A
dh-I
C
ITA
CrC
,.-C
JGT~
A C
AT
AG
AC
G
D3N
dsll
F
cgrl
G
TO
L~C
AG
'I~A
TC
AG
CI'C
CI'G
D3N
dM2
Glu
t-2
G
GG
CG
CT
GG
AA
AG
CI'A
AT
A C
C
D3N
dM3
Gh
a-2
O
TCTC
L- i T
I C
TAG
'IT]'C
T'G
AC
-C-C
D3N
dsl4
A
p2
TC
CA
TA
GC
AT
FCA
TG
CG
TG
CA
D3N
dsl5
A
p2
TA
TA
AG
AT
rCC
AG
AA
CA
CA
Tr
D3N
dM6
11
-2
TG
TA
C~
AC
AA
CA
C
D3N
dsl7
1
1-2
A
CrA
GC
AA
GA
G~
TC
rCT
G
D3N
dsl8
N
sfo
AG
GT
rCA
TC
CG
GA
TA
GA
CA
CA
D3N
dM9
Am
y-1
A
TGA
AC
ATA
TGTG
TAA
GTA
AA
.ATG
D3N
ds~7
G
ba
GA
AG
GA
AA
GG
AC
rTA
GT
CT
AC
C
D3N
da21
L
y-3
8
GT
GT
A A
AA
TC
A A
CA
CC
A A
CA
G T
AT
DJN
ds22
Ly
-38
CK3G~ I-I-I-I~3T]~G ~'I'FAGT
D3N
da23
* H
~86-
ps2
AG
TGC
CC
AA
TCA
CA
'V['C
TGC
D~N
ds24
**
Cd-
IO
D3N
ds25
***
C, ba
Prim
er re
vers
e {5
'-3')
Prod
tmt s
ize
Con
ditio
ns
TA
TG
TF
GC
~C
AA
AT
AG
AT
G
90
1 laM
/55
TCTG CATG C
CAGGGTI'GTGAT
128
3mM
155
CA
TA
AT
AT
FC
AT
AT
AT
AA
TG
C
165
3mM
f55
CTC
AC
AA
ATA
CC
TTC
AG
AG
GA
I I
0 I r
aM/5
5
TG
GA
GT
CA
CC
TT
CT
TG
AG
Tr C
14
8
AA
GT
AT
GG
GT
CA
GA
TT
GT
GT
GG
G
] 70
2mM
/58
CI'I
'GG
CI'G
'VI'A
TA G
TC.3
"I'V
I'G
117
1.5m
M/'5
5
TG
AA
TA
AA
GG
A C
TC
CT
GA
G C
l"
145
2mM
/55
CC
FIT
CA
TC
CA
TG
TA
CA
TA
TA
C
330
2mM
/55
OG
CT'
FCTA
AC
TTG
CTG
A A
AG
G A
A
184-
188
1,5m
IVl~
3
CA
TA
AG
AG
AG
GG
AG
GG
AG
AT
C
120
ImM
~5
C1T
CA
CA
GA
'~.G
TA
T'IT
GA
AA
CA
G
113
lmM
/55
GT
CT
G~
AC
rAT
GT
GC
14
6 ~
2mM
/55
GA
TA
AG
AG
CA
TG
GA
TT
TA
AC
T
133
2mM
/55
TA
CC
TA C
A C
AT
GA
TA
T]~
A A
C
224
2mM
/55
AT
ITI'A
TA
TG
TC
TC
TA
GT
'I'G
CA
C
232
2mM
/55
TI'
CC
~T
AT
AC
AG
GA
T~
G
232
1.5m
M/5
5
A A
AT
AA
A A
AG
GC
CA
CT
A'IT
rGA
AG
15
3 2m
M/4
5
GG
C cr
TG
GC
TC
TG
'I~i'A
'ITC
TG
T
190
1.5m
M/5
5
GG
CA
GG
TI'T
GA
TFC
FA A
GG
TA
G
166
1 raM
/55
GG
A C
AG
CC
AG
G A
CT
AT
AC
AG
A
164
I raM
/61
GA
TC
CI'G
TA
AT
CT
GT
CC
AT
AG
11
00
2mM
/55
hnM
/55
1.5m
M/5
8
2raM
/60
CFC
CT
GT
AG
GT
CC
AT
CA
A C
AG
C
129
lmM
/55
TCA
TI'/W
CTI
"CC
I'CC
CC
CTC
11
8-14
5 3m
M/5
5
CTA
AG
TC
CT
G C
.A C
C'I~
CC
TC
88
-200
lm
M/5
5
CT
AC
AG
TG
GG
GG
CA
GA
AG
GT
14
0-15
0 1.
5mM
/55
GG
'IT
rCG
GA
AT
G A
GA
TG
AG
C
178-
188
lmM
/55
CC
TGA
AA
CA
AA
GC
AA
CA
GC
A
125-
145
1.5m
M/5
5
D3Nds26****
Gpi-irs
D3N
d~27
1
1-2
G
~T
CA
AC
AG
CG
CA
DJM
itl
M28
T
GT
GC
AC
AG
GG
GT
AC
AT
AC
A
D3M
iB
M25
0 C
L-I !
I-I
C-A
GG
CA
AA
GC
TCC
D3M
iJ4
L40
TG
TG
CC
rGC
AA
GT
FG
TF
CrT
D3MiB
M123
A~
AG
TG
TC
TC
T
D3Mi~
M149
AACTrCAACATGTGAGGGGC
GT
CC
rGG
AG
GT
FCA
TA
AT
G
GG
GA
GA
GA
AA
AC
GC
AA
CA
T 1200
GA
TG
CA
GT
AC
AG
TC
A C
AG
CA
T
CC
AT
CC
AG
TrA
CG
TG
GT
AA
AG
GT
CC
CO
OT
GT
CT
C~T
FI'G
TC
rCT
AA
CA
'IT
TG
AC
CA
CA
GC
GA
ATA
GC
GG
CA
AC
CTA
CC
C
280
Size
var
iati
on
NO
Dff
iNO
N>>
AK
R/J
>B ]
0/W
=B6 J
JL~B
6/$
= DB
A f2
J; S
PE:-
S PE
>> D
B A
/73>
> N
OD
~ N
O N
ffiB
10/W
=B6.
FLfB
6/Jf
fiA K
I~
NO
D~
NO
N-B
10/W
=B6.
PL~
B 6/
J.A
IGT
~>> D
BA
/2J>
>S P
E
N O
N>B
6/J-
B 10
/Wff
iB ~J
L>
DB
A/2
J>N
OD
NO
D>>
DB
A/2
JffiA
KR
/J.N
ON
>B
10/W
ffiB
&PL
=B6/
j" ($
PE d
oubl
et)
SPE
>B6.
PL-B
I 0/W
ffiB
6/$>
NO
N>A
KIT
JJ>D
BA
/2J>
NO
D
SPF-
>>C
57L/
J>>S
WR
/Jff
iC57
BR
fSJL
/Jff
iB 1
0/W
.NO
N>C
3 H
/HeJ
ffi
A/J
~A K
R/J
>>C
BA
ffiB
A LB
/cB
yJff
iNO
D>>
DB
AfP
Lf/
N O
D~A
KR
/J>>
NO
Nffi
B 10
/Wff
iB 10
/Jff
iB6/
J= D
BA
/2I>
>SPE
B I 0
/Jff
iB 10
.BR
>NO
D>A
=C58
~MEV
; SP
EffiB
10/W
>NO
D>B
6.PL
>NO
N
C3H
-DB
A=A
fB6f
fiAK
Rff
iBA
LBff
iB 1
0~C
AST
ffiS
PEff
iNO
N>>
NO
D
B6f
fiB6J
~=D
BA
ffiB
10f
fiAK
R=$
PE>N
ON
fNO
D
B6f
fiBfi,
PLzD
BA
=B 1
0ffiA
KR
ffiS
PF~N
ON
fNO
D
NO
N>C
BA
>B 10
/W=B
6.P
L= N
OD
~SPE
-B 6/
J> D
BA
~J
N O
Dff
iB 10
/Wff
iB 6.
PL~B
6/J-
DB
Af2
JffiN
ON
>SPE
NO
I)=B
10/W
-B 6/
J- N
ON
ffi D
BA
f2Jf
fiSPE
NO
Dff
iB 10
/Wff
iB&
PLff
iB6/
JffiN
ON
ffiS
PEff
iDB
A/2
J
NO
D~B
10/
Wff
iNO
N=$
PEff
iB6/
JffiD
BA
/2J
CB
Aff
iMO
LDff
iYB
Rff
iC3H
=NO
D>B
R~d
J>A
KR
>SPE
SPE
(dou
ble0
>> N
O D
-B 1
0.W
ffiB
6.PL
ffiB
6/Jf
fi DB
A/2
)"
NO
Dff
iB 10
/W=B
KPL
~B6/
Y-N
ON
-DB
A/2
Yff
iAK
R/J
ffiS
pE
N O
D=B
10/W
ffiB
6.PL
ffiB
6/~'
ffiN
ON
ffi DB
A/2
I=A
KR
/3">
>S pE
RFL
I~ H
infl
NO
D-+
; B
I0,B
6. N
ON
ffi-
RFL
P; M
spI S
PE=+
; B
6=-
SPE>
mB
6
NO
D=-
+; B
10=-
N O
DfB
10/
W=B
6/J
=B6.
IK,=
NO
N=D
B A
/2J=
A K
R>>
SPE
OB
>LF>
>BA
LB=N
OD
>>A
=B6=
C3H
ffiD
BA
=AK
Rff
iNO
N>C
AST
; SP
E:-
CA
ffF>
>A=B
ALB
=LI~
OB
ffiB
6= D
BA
fA K
R= N
O D
>> C
3 H= N
ON
>>Sp
E
CA
ST>>
SPE>
>OB
=A=B
6ffiC
3Hff
iDB
A=B
ALB
=AK
R=N
ON
=NO
D~L
P
OB
ffiA
=C3H
=DB
A=B
ALB
=AK
R=N
ON
= N
OD
>>C
AST
=B6=
LP>>
S PE
B6'
ffiL
I~O
B>>
A=C
3 H
=DB
A=B
ALB
=A K
R=N
ON
=NO
D>>
CA
ST>>
SPE
Co
nti
nu
ed
o
n n
ex
t p
ag
e
O C3
L~
(./3
Tab
le 3
. C
ontin
ued.
D3M
it'7
M74
D3M
it9
A85
D3M
itlO
A
34
D3M
itll
L
38
D3M
itl2
A
60
D 3
Mid
3 1,
8
DM
ditl
3 L
37
AT
GC
AA
CrA
AC
r1T
AT
I*G
A A
AA
TC
AA
CT
rCA
~A
AA
C]'
AC
C
CrG
GC
TrG
GT
GG
AA
GT
CC
T
CC
AA
CC
AC
AG
TA
AC
AC
AT
GT
TA
GA
CC
AA
T~
AG
TG
TC
C
CC
TrT
CT
GA
TT
AT
GT
GC
K~ C
T
TFI'CrC, CATFAT~TC, C
~
D3M
itl 4
M
206
A~
GG
TrA
AA
G'I
TrG
CT
T
D3M
itl5
A
55
AA
Trr
G C
A'['
FCC
AG
GA
CC
AC
D3M
itl6
M
159
TG
CT
TG
TC
CT
GT
GT
TA
AT
GA
D 3M
it17
M
235
CA
~ T
GG
2WC
I'I'
G
D 3
Mid
9 M
141
CA
GC
CA
GA
GA
GG
AG
CT
GT
CT
D3M
it21
D
31
AA
GC
rCT
AC
AG
OG
GA
AG
CA
C
D3M
it22
D
122
AAGGATIX3 AAGAATOM3Ti~
D3M
iz23
D
I02
G A
'IT
CC
AG
A A
C A
CA
T'~
TG
GG
D3M
it24
A
636
AG
TI'
FC
TA
G C
CF
CA
GT
G'[
'rcI
TC
A
DfM
it25
A
726
GT
CI'
GG
GT
CG
TC
AG
TG
G C
D3Mi
t26
B332
T
rGG
AT
rCA
TA
TC
AG
GA
CF
GT
AC
A
D3M
i,O8
D62
7 O
AT
G A
GA
GA
TY
C]'G
AT
GT
GG
AG
G
D3M
it29
D
566
GA
TG
AG
AG
A'I
WC
rG A
TG
TG
GA
GG
D 3
MiB
I A
629
AC
AA
CC
CG
AG
TT
CA
GT
CC
C
D 3
Mit3
2 B
128
CA
CC
CT
C~
TrA
AC
TC
AG
A A
AG
G
TA
CA
AT
rAT
CC
GG
GA
GC
rA
TG
TIT
r AT
AT
FG
CC
CT
GT
AT
GT
G C
CC
TA
AG
CC
AG
CT
AC
CA
CC
AC
TG
GA
GA
CC
AA
TG
CG
AA
CA
AC
GG
AA
A A
GC
AT
AA
G A
AA
CA
AC
CG
CC
AC
TG
AA
GG
AT
AA
CC
AC
AG
AA
CC
AC
AG
AT
GA
CA
ATI
X3A
A
TC
CT
GC
AA
AT
I'G
TC
CT
CrG
A
AG
G A
AG
TG
A C
GT
['G
GG
TF
rG
TG
AG
AA
TG
GA
GG
TG
AA
CA
GC
CC
AC
GG
AG
AA
CA
AC
rGA
AG
A
GA
AC
AT
rGG
GG
TG
TIT
GC
IT
CT
GG
GG
AG
'IT
I'C
AG
GT
I'C
CT
AA
TC
AG
CG
A~
'TC
AG
CA
CG
TC
A A
TC
AG
AG
AG
AT
A A
GA
GC
AT
GG
GT
CA
T]'
GT
rTA
TC
AT
A C
CC
AC
AG
G
TG
GA
GG
C~A
CC
AT
CT
CC
A A
G
~A
AT
CA
AG
TG
GT
FF
AG
TC
AA
CC
AG
CC
TC
AG
TA
TC
TC
AA
AA
CC
CC
AG
CC
TC
AG
TA
TC
TC
AA
AA
CC
GT
GC
]W.~
CA
G~r
A A
G ~
'TG
GC
A C
FD
GT
GT
ITC
AT
GT
CA
CFG
D3M
it36
D
SOI
GA
TI'
I'FA
A"f
fI'C
AT
rAA
AT
AA
GG
GT
TA
G
GA
A C
AT
AT
GT
GT
AA
GT
AA
AA
TG
TA
C
D3M
it38
A
674
C'I
~A
AC
CA
GA
AA
GT
rGI -
F 1 -
|-rC
r'G
D3M
it3O
A
86P
C
TG
CC
AC
AG
AG
CI'
AT
AG
CC
C
D3M
i~IO
A
IIlO
C
AG
~T
CT
AA
CT
AT
CC
CC
C
D3M
it41
B23
5 A
AT
I'F
CT
FC
CT
GT
rAC
A C
TG
AG
CC
D3M
iN3
B391
T
G A
CC
I"C
CA
G A
GA
GT
CT
FC
CA
D3M
iN4
B43
9 C
CT
GA
CT
CA
T'f
TA
2~i'A
AC
TC
CC
C
D3M
iN6
D53
4 A
TC
CC
CC
AC
CC
AA
CT
CrA
AC
D3M
iPI8
D
535
TC
r~T
AC
CC
CT
Trc
cccc
D3M
iN9
D56
7 C
ITL
TC
TC
GC
CC
CA
L-t
l- t C
AC
CA
TG
GC
CA
GC
TT
CT
AA
TG
A A
TG
GA
G C
Tl~
CF'
FFC
GA
TG
CC
qTA
TI'
A A
GT
GC
AT
G A
CC
I'F
G C
CA
TG
AG
AG
A A
CT
CC
TT
C C
AT
CC
CT
GT
GC
AT
GA
GA
CC
AC
TA
CC
A
CC
CT
AT
CA
CA
TA
GG
GC
AA
CC
"Frc
ccC
AG
GG
AA
AT
CT
CF
cr
GT
TG
AC
AC
AA
A A
CT
AG
CC
AG
TA
CA
TC
t-I~
I 1
l AG
TIT
ITG
AT
CC
TC
TG
G
142-
147
lmM
/55
OB
~S P
E=B
6= D
BA
>>C
A S
T~A
= C
3H=
BA
L]~
-A K
R=N
ON
=NO
Dff
iLP
214-
238
1 raM
/55
CA
ST
>>
OB
=B6~
>DB
A>I
..P>N
OD
>A=B
ALB
>SPE
-~A
KR
.>>N
OD
; C
3H:-
121-
158
1 mM
/55
CA
$'I'>
>B6=
OB
>> D
BA
>LP>
NO
N>B
AL
B=A
>A K
R=S
PE>>
NO
D;
C3H
:-
146-
204
lmM
/55
CA
ST
>>
A=
C3H
fBA
LB
=A
KR
>N
ON
>>
SP
E>
>O
B=
B6=
DB
AfN
OI~
LP
120-
157
lmM
/55
LP=
AK
R=A
>BA
LB
=C3H
=B6=
OB
>>N
OD
~NO
N=D
BA
>>C
AST
; SP
E:-
228-
240
1 raM
/55
SP
E=
LP
>C
AS
T, N
OD
>>O
B=B
6=C
3 H
=D
BA
-BA
LB
=A
KR
=N
ON
; A
:-
220-
237
lmM
/55
LP>S
t~>N
OD
>>C
AST
>>O
Bff
iB6f
fiDB
A=C
3H=B
ALB
ffiA
KR
=NO
N
12%
198
lmM
/55
N O
Dff
i NO
N~A
KR
fBA
LB
- DB
A=
C3
HfA
>>
LP~
B6f
OB
>>
SPE
>>
CA
ST
145-
212
1 raM
/55
DB
A>
>C
AS
T>
>S
PE
>>
NO
D-N
ON
=A
KR
ffiB
ALB
=C3H
ffiB
6=A
ffiO
B; L
P:-
186-
220
1mM
/55
SPE>
>CA
ST~>
B6=
OB
~LPf
fiNO
D=N
ON
fBA
LBff
iC3H
ffiA
; AKR, D
BA
:-
�9 1
80-2
081
lmM
/55
NO
Dff
iB6f
fiOB
>>C
AST
>>LP
>>N
ON
fAK
RfB
ALB
ffiD
BA
fC3H
ffiA
; SP
E:-
210-
238
1.5m
M/5
5 C
A S
TfA
fC3H
ffiD
BA
ffiB
AL
B=A
KR
fNO
D>>
OB
fB6'
,,,N
ON
>LP>
>SPE
208-
236
1 raM
/55
NO
NfB
ALB
=B6=
OB
>>LP
ffiN
OD
ffiA
K
R=
DB
A=
C3H
fA>
CA
ST>
>SP
E
207-
265
lmM
/55
CA
ST>>
A>>
NO
D>>
OB
=B6=
C3
H=N
ON
>DB
Aff
iAK
R>>
BA
LBff
iLP>
>SPE
138-
162
lmM
/55
CA
ST~.
>LP=
NO
D,=
NO
NfA
KR
ffiB
ALB
=DB
A=C
3H=B
6=A
=OB
>>SP
E
130-
168
1mM
/55
CA
ST>>
SPE
>> N
ON
>>
Lt~
NO
D=
AK
R=
BA
LB
=D
BA
=B
6=O
B>
>C
3H=
A
114-
134
1 mM
t55
B6~
OB
>L
P~N
OD
,=A
KI~
BA
LI~
C3H
fA>
>12
9>N
ON
>D
BA
>>
SP
E
130-
156
1 raM
/55
CA
ST>>
B6,
* O
B>
>L
I~N
ON
>>
NO
D,,
,AK
I~B
AL
B-D
B A
= SP
E; C
3H, A
:-
1.50
-202
I r
aM/5
5 B
A.L
B>
>C
AST
>>
DB
A>
>SP
E>
>O
B>
NO
NfA
KR
=C
3H-B
6-A
; L
P, N
OD
:-
14,4
-2~0
lm
M/5
5 B
A L
B>>
CA
ST>>
DB
A>>
SPE
>>B
6-A
ffiO
B>L
P=A
KR
=C3H
>NO
N>
NO
D
222-
242
lroM
/55
CA
ST
>SPE
>>L
P=N
OD
iN O
Nff
iAK
R=B
ALB
=DB
A- C
3 H=A
>B6=
OB
164-
178
lmM
/55
LP=
NO
D,=
NO
Nff
iA K
Rff
iBA
LB=
DB
A=C
3 HfA
-SP
E>
>B
6=O
B>
> C
A S
T
138-
226
l raM
/55
NO
D>>
CA
ST>>
SPE
; L
P, N
ON
, AK
I~ B
AL
B,
DB
A, C
3H, B
6, A
, OB
:-
122-
142
lmM
/55
CA
ST
>>
Lt~
BA
L B
=C3 H
fAfS
F~K
>>N
OD
= NO
NfA
KR
=DB
A>B
6=O
B
226-
242
lmM
/55
S PE
=CA
ST
>>
Lt~
N O
D,,,
N O
NfA
KR
=D
BA
-C3
Hff
iB6=
A-O
B>>
B A
LB
110-
140
lmM
/55
B6=
OB
>>C
A S
T>>
SPE
.>>L
I~ N
O I~
NO
N=A
KR
=BA
LB
= D
BA
ffiC
3 H=A
210-
236
lmM
/55
CA
ST
~> N
O D
>>SP
E>>L
IX~
NO
N=
A K
/~B
AL
B=
DB
A=C
3 H=B
6~A
fOB
114-
146
lmM
/55
CA
ST>>
BA
L]g=
C3
Hff
iA>>
LI~
NO
D=
NO
NfA
KR
=B
6-O
B>
> D
B A
>S PE
124-
148
l raM
/55
N O
1~ D
B A
>B 6=
OB
>N O
N>>
A K
R>>
LP=
BA
LB
~C3
H=A
>>C
AST
~>S P
E
152-
194
lmM
/55
SPE>
>C.A
ST
>>C
3H,A
>>
B6,
,OB
> B
AL
B>
Lf~
NO
D>
DB
A>N
O N
>>A
KR
247
lmM
/55
108-
148
1 raM
/55
BA
LB
=DB
A>>
LP=
B6=
OB
>>N
ON
-C3H
>CA
ST;
NO
D, A
KR
. A, S
PE:-
Co
nti
nu
ed
o
n n
ex
t p
ag
e
o O
t= g ('3

Tab
le 3
. C
onti
nued
.
Loc
us
Sequ
ence
P
rim
er fo
nvtr
d (5
'-3')
Prim
er re
vers
e (5'
.Y~
Prod
uct s
ize
Con
ditio
~, S
ize
vaH
ltinv
l
D3M
itSO
17
107
168-
198
lmM
/55
D3M
itSl
.I8
GG
CA
CT
GA
T
AG
CA
GG
CC
I'A G
T
CT
CT
YC
I'G
GT
AT
YI'
CC
I'I'
CC
G
230-
258
I raM
/55
DJM
it52
AI0
81
A
GC
CA
GG
AT
AT
GG
AA
TA
TG
CC
T
GA
CC
AG
AT
TG
CA
TG
CA
TIT
19
6-20
4 lm
M/5
5
D3M
iI54
BS
T2
T
I'G
G'I
'FC
CA
CA
GC
AA
C'I
'AC
G
CA
GG
GA
AT
GT
AT
GT
CA
A
TG
A G
G
122-
148
lmM
/55
D3M
it~5
B5
36
C
I'G
GG
A
CC
A C
CA
GT
AG
TA
CC
A
TC
AG
GA
CT
GC
AA
CI~
AG
GC
11
6-14
4 lm
M/5
5
D3M
i66
B71
3 T
CT
AG
CI'A
TG
TG
AT
GA
GT
GT
GT
CG
C
AG
GA
'IT
I'C
CA
A
AA
A C
AT
CC
A
138-
148
lmM
/55
D3M
it57
B4
93
T
CC
AG
TF
A~
TG
AA
CF
CC
.A
AT
AT
GT
GT
AC
AT
GT
YC
AT
GG
TG
TG
14
8-17
6 Im
M/5
5
D3M
it$8
B52
7 A
CA
TC
AG
AA
GA
GT
CA
TI'
CA
'IT
rCA
G
CT
CT
rCA
GT
CA
CA
G
CI'C
I'GC
1
40
-15
2
lmM
/55
D3M
it59
B5
43
G
TrG
AT
GC
CC
AA
GG
AA
TG
AT
L
'I'A
CI'G
CA
TC
CT
GG
CA
CA
GA
20
4-21
2 lm
M/5
5
CA
ST~>
LPffi
AK
Rffi
BA
LB-ff
i DB
A>C
3 H=A
> N O
Dffi
NO
Nffi
B 6=
OB
; SPE
:-
AK
R=
BA
LB~D
BA
>N
OD
ffiN
ON
>>
LI~>
C3H
=B
6ffiA
=O
B>
>C
AST
; SP
E:-
Affi
SPE>
OB
>NO
Dffi
NO
Nffi
AK
Rffi
DB
Affi
C3 H
>L
P~B
AL
I~B
6; C
AST
:-
N O
Dff
iBA
LB
fC3 H
=B6f
fiAff
iOB
>CA
S'I~
NO
N>>
LP.
A KR
= DB
A>>
SPE
C3 H
ffiA
>>L
t~-N
O D
=B6>
A 1C
Rffi
BA
LBffi
DB
A.C
A S
T>>
NO
N>>
S pE
; OB
:-
A>L
PffiN
OD
ffiN
ON
=AK
Rff
iDB
A=C
3n=B
6~O
B>B
AL
B>>
CA
ST; SP
E:-
CA
ST~>
SPE
>>
Lt~
B6>
NO
N=
AK
R.B
AL
B=
C3H
.A.O
B>
>N
OD
; DB
A:-
CA
ST>
>N
OD
=N
ON
ffiB
ALB
=D
BA
ffiC
3H=
B6,
~A=
OB
>>
LPffi
AK
P4
SPE
:-
OB
>CA
ST>L
P=A
KR
ffiB
6ffiN
O N=B
A LB
ffiD
BA
.C3H
ffiA
ffiS
PE> N
OD
Rec
omm
ende
d m
agne
sium
con
cent
rati
ons
and
anne
alin
g te
mpe
ratu
res
are
indi
cate
d fo
r m
ost
loci
. *D
3Nds
23: H
sp86
-ps2
, unp
ubli
shed
seq
uenc
e w
as k
indl
y pr
ovid
ed b
y S.
M
oore
, Foo
d an
d D
rug
Adm
inis
trat
ion,
Div
isio
n of
Met
abol
ism
and
End
ocri
ne D
rug
Pro
duct
s, R
ockv
i!le
, Md.
; the
ST
S an
d po
lym
orph
ism
are
from
N.
Rod
rigu
es. *
*D3N
ds24
: C
DIO
, clo
ne o
btai
ned
from
M.
Ship
p, D
ana-
Far
ber
Can
cer
Inst
itut
e, B
osto
n, M
ass.
; th
e ST
S an
d po
lym
orph
ism
are
fro
m N
. R
odri
gues
. **
*D3N
ds25
: G
ba,
(ref
, 11
9).
****
D3N
ds26
: G
pi-i
rs, u
npub
lish
ed S
TS
from
J.
Jone
s, M
RC
Rad
iobi
olog
y U
nit,
UK
.
t~
Tab
le 4
. M
icro
sate
llit
e m
ap o
f C
hr 3
.
Loc
i R
F D
ista
nce
D3M
itl-
D3N
dsl5
7/
92
7.65
cM
D3N
ds15
-D3N
ds27
10
/92
11 c
M
D3N
ds27
-D3M
it6
7/92
7.
65 e
M
D3M
it6-D
3Mit5
2/
92
2.1
eM
D3M
it5-
D3M
it7
3/92
3.
2 eM
D3M
it7-
D3N
dsl
0/92
0
D3N
da1.
D3N
ds24
3t
92
3.2
cM
D3N
ds24
-D3M
it22
5/92
5.
4 cM
D3M
it22
-D3M
itSl
3/
92
3.2
cM
D3M
it51
-D3M
it9
1/85
1.
1 cM
D3M
it9-D
3Nds
25
0/85
0
D3N
ds25
-D3N
ds22
0/
92
0
D3N
ds22
-D3N
ds8
4/92
4.
3 eM
D3N
dsS-
D3M
itl I
1/
91
1 cM
�9 D3M
itll
-D3M
it42
7191
7.
7 cM
D3M
it42-
D3N
ds9
1219
2 13
.3 c
M
D3N
ds9.
D3M
it38
3/
92
3.2
cM
D3M
it38-
D3M
it31
7/90
7.
8 eM
D3M
it31
-D3M
it32
4/
90
4.4
cM
D3M
it32
-D3M
it19
3/
92
3.2
cM
Dat
a fr
om th
e la
bora
tory
of
J.A
. T
odd
from
the
Eur
opea
n C
olla
bora
tive
In-
te
rspe
cifi
c B
ackc
ross
[(C
57B
L/6
x
SPR
)FI
• SP
R]
prod
uced
by
the
UK
's
Hum
an G
enom
e M
appi
ng P
roje
ct,
wit
h th
e su
ppor
t of
the
Med
ical
Res
earc
h C
ounc
il. D
ista
nces
wer
e ca
lcul
ated
wit
h th
e K
osam
bi m
appi
ng fu
ncti
on. R
F,
reco
mbi
nant
frac
tion
(re
com
bina
nts/
tota
l ana
lyze
d).
Tab
le 5
. R
elat
ions
hips
bet
wee
n m
ouse
Chr
3 g
enes
and
hom
olog
ous
hum
an g
enes
.
Map
L
ocus
N
ame
Hum
an
Hum
an p
osit
ion
6.3
Car
-I
carb
onic
anh
ydra
se-i
C
A 1
8q
13-q
22
6.3
Car
-2
carb
onic
anh
ydra
se-2
C
A2
8q13
-q22
7.
5 C
ar-3
ca
rbon
ic a
nhyd
rase
-3
CA
3 8q
13-q
22
10.2
E
vi-
1
eeot
ropi
c vi
ral i
nteg
rati
on s
ite-
I E
vil
3q24
-q28
10
.2
Fir
e-3
Fr
iend
MuL
V i
nteg
rati
on s
ite-
3 FI
M3
3q27
10
.2
Glu
t-2
gluc
ose
tran
spor
ter
2 G
LU
T2
3q26
15
.0
11
-2
inte
rleu
kin
2 ]1
.2
4q26
-q27
15
.7
Fgf2
fi
brob
last
gro
wth
fact
or b
asic
F
GF
2 4q
25-2
7 30
.1
Min
e m
embr
ane
met
alio
-end
o pe
ptid
ase
MM
E
3q21
-27
33.3
Sa
c-Is
su
cras
e-is
orna
ltas
e, st
ruct
ural
SI
3q
25-2
6 34
.9
Glu
t-2
glut
amat
e re
cept
or 2
G
LU
R2
4q25
-34
42.3
F
gg
ganm
aa fi
brin
ogan
F
GG
4q
28
42.6
P
klr
pyru
vate
kin
ase
live
r, r
ed b
lood
cel
ls
PK
LR
lq
21
42.6
G
ba
beta
glu
cece
rebr
osid
ase
GB
A
lq21
42
.9
Cac
y ca
lcyc
lin
CA
CY
lq
21-q
25
42.9
C
apl
calc
ium
bin
ding
pro
tein
, pla
cent
al
CA
PL
lq
12-q
22
42.9
F
cgrl
hi
gh a
ffin
ity
FC
gam
ma
rece
ptor
FC
GR
1 lq
43
.9
Cd/
cl
uste
r des
igna
tion
1
CD
1
lq22
-23
43.9
F
psl-
rsl
farn
esyl
pyr
opho
spha
te s
ynth
etas
e-li
ke 1
FP
SL
lq24
-q31
44
.9
H3f
2 hi
ston
e 3,
fam
ily
2 H
3F2
lq12
-21
45.0
H
sd.3
b
3-be
ta-h
ydro
xy s
tero
id d
ehyd
roge
nase
H
SDB
3 lp
ll-1
3 46
.4
Cd2
cl
uste
r de
sign
atio
n 2
CD
2 lp
13
47.3
A
tpla
l N
a, K
AT
Pas
e al
pha-
1 A
TP1
A1
lp13
47
.4
Am
pd
-1
AM
P d
eam
inas
e-1
(mus
cle
form
) A
MPD
1 Ip
13
47.4
N
g/b
nerv
e gr
owth
fact
or b
eta
NG
FB
lp13
47
.4
Nra
s N
ras
onco
gone
N
RA
S lp
13
47.4
R
apla
m
embe
r ofR
AS
onc
ogen
e fa
mil
y R
AP
1A
lp12
-p13
47
.4
Tsh
b th
yrot
ropi
n st
imul
atin
g ho
rmtm
e be
ta s
ubun
it
TS
HB
lp
13
48.4
C
sfm
co
lony
sti
mul
atin
g fa
ctor
, mac
roph
age
CSF
1 lp
13-2
1 48
.4
Gna
i-3
guan
ine
nucl
eoti
de b
indi
ng p
rote
in,
GN
AI3
lp
13
alph
a in
hibi
ting
act
ivit
y-3
49.6
A
my
-1
amyl
ase,
sal
ivar
y A
MY
1 lp
21
49.6
A
my-
2 am
ylas
e, p
ancr
eati
c A
MY
2 lp
21
49.6
C
f-3
coag
ulat
ion
fact
or 3
F3
lp
22-p
21
54.6
P
rop
-1
pero
xiso
mal
mem
bran
e pr
otei
n (7
0k)
PMP1
1p
21-2
2 54
.7
Fab
pi
fatt
y ac
id b
indi
ng p
rote
in i
ntes
tina
l F
AB
P2
4q28
-31
62.1
E
g]"
ep
ider
mal
gro
wth
fac
tor
EG
F 4q
25
68.1
A
dh
-1
alco
hol d
ehyd
roge
nase
-I
AD
H1
4q21
-q23
68
.1
Adh
-3
alco
hol d
ehy&
ogen
ase-
3 A
DH
3 4q
21-q
23
X
Act
s sk
elet
al a
lpha
act
in
AC
TA
1 lp
21-q
ter
X
Adh
-5
alco
hol d
ehyd
toge
nase
-5
AD
H5
4q21
-q25
X
A
rm
aryl
hyd
ecar
bon
rece
ptor
nuc
lear
tran
slat
or
AR
NT
lp
ter-
ql2
X
Le
f-1
ly
mph
oid
enha
ncer
-bin
ding
fact
or 1
L
EF1
4q
23-q
25
O~
t~
(3
L~

M. F. Seldin et al.: Mouse Chr 3 $57
Locus AXL Lines Ref. Locus AXB Lines Ref.
1111112222233 11111111/222222 567892346791458978 123456789~/2345789~12345
Cur-2 LALL-ALALAAALALLAA /21 Cor-2 BBkM~BABABABAMAB BB - B BBA 73,1/2 ApZ LAL LLAL~LAN~LALL LA 61 Evi-1 BA-BABABABAA-A-A~BABAA 1/2 Mpmv-2~ LLAALALALLLLLALLLL 51 O3Mit6 BA-ABB-A- BA- BAA-AAA- - BS- 73 Evi-1 L LAALAAALALL LALLAL 1/2 D3Mit7 BB-ABA- B- BA-AAA-A,~,- - BS- 73 Xr~v-47 LLLLLAAALLALAALLLL 164 Fgg BBAABABBBSAB-AAABESB- BSA 13 X~v- 65 ALALLAAALLLLALLLAA 164 Thbs3 BB-ABA- B- BA-AAA- BBS- -BB- 73 I~v-28 ALALLAAALLLLALLLAA 50 Rnulb-1 ABAABABBBBAB-AABBBSB- BBA 13 Colll ALALLAAA- LLLALLLAA ]31 gnulb-3 AAAABABBBBABAAABBBSB- BS- 13 O3Tu51 LLALLAAALLLLA-LLAA 153 D3Mit14 AA-A~-A- BA-AAB-AAA- -AB- 73 Amy ALALLAAAL L LLA~AL LA /21 ~any-l,2 AJ~ue~k~BABBAB -ABSBA-A-A B - 13 O3Tu33 AAL-AN~AL LLLA-AL LA 153 EgF AA~.~BABSAAAA-AA-AA~BB 11Z Pmv-26 AAa_~LA L L LAAALAL LA 50 ~dh-3 AMABA BAABAAAA BBAAABAAB - 13
M;mv-9 BAABAABAABABAAABAAABAABA 51,1@0u O~Mit18 AA-BAA-A-BA-AAB-A-A- -AB- 73 O3M~t/9 AA-AAA-A-AA-AAB-AAA- -AB- 73
LOCUS BXD Lines Ref.
11111111222222222333 /25689/2345689e12345789~/2 Locus BXA Lines Ref.
Car-2 DDDBDBDDDOBBDOBBBBBBDBDDBB 121 Ap2 DDOBDBDDDOBBD- BSB BSBDBDDDD 61 1111111111222222 D3Byu2 DDOBO BDDOOBBDDBBBESBOBDDDB 167 1234567890/234567890/2345 O3ByuJ DOOBOBODDOBSDOBBBBBBDBOD08 167 Con-2 AABB- ABAA- BAABBBAA- - -AABA 73,112 O3Byu3 DDBBOBBDDDBSDOBBBBSBDBDDD6 167 Evi-1 AA- B- B-AABBAAB--- B8- -AABB 112 Evi-i DDBBDBBDBBBSBDBDBBSBDDODD8 112 O3Mit6 AB-B--BBB-BAAB-BB . . . . . . BA 73 I1-2 DDBBDBDDDDDBDDBDBBSBDOBDDB 37 D3Mit7 AB-B--BBB-BAAB-BB ...... BA ?3 D3Mit5 -DBBDDDOBDBBDDBDBBBBDDB--- 37 Fgg A-ABBB-BBABA-B---AB ...... 13 DOByul7 BBDBDDOOBBDBBBBDDBBDOBODB8 167 Tsp3 AB-A--ABB-BAAB-BB . . . . . . BA 73 Cnp-2 BBDBBDODBBDBBBBDO B8 [X)OBDB8 10 Rnulb-1 A-AABB- BB-BA-B-- -AAA-B-- - 13 03_71 BBDBBBOBBBDBBBB- DBSOBDBDB8 114 Rnulb-3 ABAABBABB- BAB8- - BAAA- B- BB 13 O3Byu5 BBDBSDODBSDSBBBODBBDBD6OBB 167 O3Mit14 AA-A- -ABB-AABS-BB . . . . . . BB 73 D3Mit22 - BDBBIBOOBBDBBBBODBBDBDB- - - 37 Amy-l,2 A-ABBB-BBABA-B---AAB . . . . . 13 Xnmv-~ BBDBBBOBBBC6BBBDOBBDBDBOBB 164 Egf AA-A- BBBBAAABA- --AA- -A- - B 112 D3MZt9 -BDBBBOBBBDBBBBDDBSDBOB--- 37 Adh-3 AAAAABABBAABBA- --AAA- B- - B 13 Fgg BBDBBBOBBBDBBBBDDBBDBDBDBB 13 MCTnv-9 AB-A-AA - BAABBA-AAAAA- ABAB 51,100o O3Byu6 BBO~DBB8BO()BOOBDBO~ 167 D3MItI8 AA-A- -AAB-ABBA-AA . . . . . . AB 73 Flnv-38 BBSBSBOBBBDBBSBODBBDBDBDBB 50 D3MIt19 AB-B--AAB-ABBS-AA . . . . . . AA 73 O3MitlO -BBI~BBDSBBBDDBBDBOB--- 37 Copl BBB88B~- BDBBS- - D-BDBDBDBB 39 Colll BBSBSBDBBSDBBS- - DBSDBDBDB8 131
D3Tu51 BBSBSB- BD6DBBS- DDBS- BDBD- - 153 Locus AXD Lines Ref. D3Byu4 BBO6BBOB88D6BSBO BBOOB DBDB8 167 Amy BDBBOBDOOB~BSBDDBBDBDBDBB 121 11111111/2.222222;'2 Amy CB B~BBI)B)OI~OSBSBODB8 DIBDBDBB 13 /23456789012345678e/2345678 Fol~i BDOBDBDODBDSB- DOOBS-BOSBBO 143 Nlpmv-20 DDA- -AAADAAADA- A-AADODADADA 51 D333 BDBBDIg)O(ODBBSO- - BSDSDBBBO 114 I I -2 ADA- -AAADAAADADA-AAADADAADA 60 Adh-lps BDSBDBI)ODSDSBSOODBDDSDSBBD 20 Cnp-2 ADA--AADAAADAADA-A6,ADAt~AAA 10 c~ ~DBDSDBB{)BDBB88 145 O3Ndsl A DA - - AAD/~AI)AADA-AAADADAAAA 30 Peev-39 BDBIg)BODOBDBDBODDDBDBDBBBO SO Rnv-28 DDA - - AA DDAADAADADAAADADDCJu~ 50 Oat-rs2 B~6E~([~I~O(~6DBD6BBO 125 Pmv-38 DDA- -AADDAADAADADAAADDODDAA 50 Egf ~ D B O B D B B B D 112 Tshb DDA- -AADAAADADDA-Ak~ADOODAA 2 O3t4it15 - ~ ~ ~ - - - 37 Pmv-39 DDO- - DDD/~DOADDADADOADODDAD 50 Adh-3 BOSIBDSOD-B88DDDDBDBDSDSB~ 112 Oat - rs2 DDD- -AADAADOADDADADOADO- DAD 125 Aah-3 PJt 8DS~-B~-DBDSDSDSBBD 72 O3Nds2 DDD--DADAADOADDA-ADDAAOOflAD Be Mh-1 BDSBDBDOOBBSDDDDBDSOBDBBBO Ze Egf D DOADDADADDOADDADDOOAADOOAD 112 O3Nds3 BOBBDBDIX)B880000BDBDB~B BBD 30 D311ds2 -088DBDD~88DDDDSDSDBDB--- 37
BDBBDBDO-BBBDO-DB-BDBDB- BO 97 D3Jknl BDBBOBDOOBBSDOODBCBOBDBBBB 99 Pmv-2~ BDBBDO[X)DB88DODOB68DBDBBBD 50 I>40-4 DDBBSDODDBBSDODBDBBDODBB BO 40 D3Nit~ - ~DOBD(O)O~DOBOB~- -- 37 D3Nitl7 - BSCODDBDBB8DODOBBBDBDB- - - 37
Locus BXH Lines Ref.
11111 2345678901249
Cor-2 HBH-BHSBSBSBB 121 Pgkps3 HBH-BHSB8BHBB 1 EvI-I HBHBBHBBSHHBB 112 I~uJb3 BBS-BHBBHHHBB 13 Colll BBB-BHBBHHHBB 131 D3Tu51 BBS-BHBBHHHHB 153
BBS-BHBHHHBBB 121 Ode -rs3 HBH-BHBHBHBBB 126 0at-rs2 BBS-BHBHHHBBB 125 Egf BBSHBHBHHEBHB 112 Adh-3 BB8-BHBBHHBBB 124 6bp-/ BBS-BHBBHHBBB 124 H3f'2 BBB-BHBBHHHBB 132 Mpmv-9 BBB-BBBBBHBHB 51
Locus CXB Lines Ref.
/234S67 Cor-2 CCBBs 47 Cor-2 RE CCBBBCB 121 Xalm, V-65 BCBBEBC 164 H-37 BCCBCBC 4
CCCBCCB 121 /4 -23 CCCCCCB 4 Adh-3 CCCBCCB 43 Ahr-1 CCCBCCB 43 Gbp-J CCCBCCB 124 H-28 CCB~CB 4 I f -1 CCBB(C8 36 /r BCBBCcB 51
Locus NXO Lines Ref.
11 /257~
Xmmv-65 NONOQO 164 Gbp-1 NNOONO $24
Locus SXL Lines Ref.
11111 4724S67
Calll SLLLSSL ]31 LLLLSSL 121
Fig. 2. Strain distribution patterns for recombinant inbred strains. Data from Benjamin Taylor and Beverly Paigen (The Jackson Laboratory).
ships in both gene order and approximate physical length (79).
Acknowledgments. Jane Santoro provided invaluable assistance in preparation of the tables and figures for this report. We thank the
individuals who contributed unpublished data so that this report could be as complete as possible. We would appreciate receiving information from readers regarding corrections and additions. Prep- aration of this report was supported by U.S. Public Health Service Grants HG00101 and DK42923 (M.F. Seldin) and GM24872 (M.H. Meisler).




















