
A highly polymorphic locus on chromosome 16q revealed by a probe from a chromosome-specific cosmid...
7
Hum Genet (1986) 74 : 425-431 © Springer-Verlag 1986 A highly polymorphic locus on chromosome 16q revealed by a probe from a chromosome-specific cosmid library L. Buflon 1 , T. K. Mohandas 3, R. E. Magenis I, R. Sheehy 1, R. K. Bestwick 2, and M. Litt 1'2 1Department of Medical Genetics, and 2Department of Biochemistry, Oregon Health Sciences University, 3181 S.W. Sam Jackson Park Road, Portland, OR 97201, USA SDivision of Medical Genetics, Harbor UCLA Medical Center, Torrance, CA 90509, USA Summary. A cosmid library was constructed from genomic DNA of a human-mouse somatic cell hybrid containing an 11q-16q translocation chromosome as the only human DNA. Cosmids with human inserts were prehybridized with total human DNA and were screened to find probes that revealed highly polymorphic loci. From one such cosmid, CF33-79, a single-copy subclone was isolated which revealed an insertion/ deletion polymorphism with at least 11 alleles and a PIC of 0.77. Using a somatic cell hybrid mapping panel, the subclone was mapped to chromosome 16. By in situ hybridization with the entire cosmid used as a probe, chromosomal localization was shown at 16q22---~24. Introduction Restriction fragment length polymorphisms (RFLP) are use- ful markers for mapping the human genome (Botstein et al. 1980). As the number of known RFLPs increases, they are be- coming ever more useful in the prenatal or early diagnosis of numerous hereditary diseases. Also of significance is the no- tion that mapping of a disease gene to a specific chromosomal location may serve as the first step in the cloning of the gene. Diseases that have been mapped by linkage studies with RFLPs include Huntington's disease (Gusella et al. 1983), Duchenne's muscular dystrophy (Murray et al. 1982), X- linked retinitis pigmentosa (Bhattacharya et al. 1984), adult polycystic kidney disease (Reeders et al. 1985) and cystic fibrosis (Tsui et al. 1985). RFLPs have also been crucial to the elucidation of mechanisms underlying certain hereditary can- cer syndromes frequently associated with chromosome dele- tions such as retinoblastoma (Cavenee et al. 1983) and Wilm's tumor (Koufos et al. 1984). In the future, RFLPs may be use- ful in characterizing the genetic contributions to suscep- tibilities to common diseases that tend to cluster in families, such as colon cancer and schizophrenia (White et al. 1985). For the construction of linkage maps of human chromo- somes, marker loci with multiple alleles and polymorphism in- formation contents (PIC) (Botstein et al. 1980) near 1.0 are very useful. Among the 333 RFLPs reported to date, only about 10% have PIC values greater than 0.5 (Willard et al. 1985). To address this problem, we have used a new method for rapidly screening cosmids and other repeat-containing DNA clones to identify those with inserts homologous to Offprint requests to: M. Litt, Department of Medical Genetics, Ore- gon Health Sciences University, 3181 S. W. Sam Jackson Park Road, Portland, OR 97201, USA genomic regions especially rich in RFLPs (Litt and White 1985; Sealey et al. 1985). Radioactively labelled probes are prehybridized with a vast excess of non-radioactive total human DNA under conditions which drive repetitive (but not single-copy) DNA into duplex form. These probes are used directly on Southern blots of restricted DNAs from panels of unrelated individuals to visualize low- and single-copy bands. Probes that display multiple polymorphisms by this method are further studied to obtain single-copy subclones which re- veal RFLPs. We have successfully used these methods to characterize highly polymorphic loci on chromosomes 2 (Litt and White 1985; Litt et al. 1986a), 12 (Buroker et al. 1986), 19 (Burton et al. 1986) and 20 (Litt et al. 1986b). The distribution of reported RFLPs is very uneven. Whereas chromosomes such as 4, 21 and the X are well en- dowed with RFLPs, others, such as chromosomes 8, 10, 15, 18 and 20 have 4 or fewer (Willard et al. 1985). Furthermore, certain chromosome arms are nearly devoid of RFLPs with PIC values > 0.5. In particular, only three polymorphic loci have been assigned to the long arm of chromosome 11, whereas only two have been assigned to the long arm of chro- mosome 16; of these five loci, only APOAI on llq has a PIC > 0.5 (Willard et al. 1985). In order to fill in these gaps in the human linkage map, we have made genomic libraries from a mouse-human somatic cell hybrid containing an 11q-16q translocation as the only human chromosome and we are using these libraries as sources of probes to reveal high PIC RFLPs on these chromosome arms. In this paper we describe a highly polymorphic locus on the distal half of the long arm of chromosome 16 revealed by a probe isolated from a cosmid library of the llq-16q translocation hybrid. Methods Somatic cell hybrids The cell hybrid used for library construction was isolated from a fusion of mouse A9 cells (GM0346, Institute for Medical Re- search, Camden, N.J.), deficient in adenine phosphoribosyl transferase (APRT) and human fibroblasts containing a trans- location between chromosomes 16 and 11 (46,XX,t(ll;16) (q13;p11); GM3005). Hybrids were isolated in medium that selects for APRT as described previously (Koeffler et al. 1981). Human APRT is located on the long arm of human chromosome 16 (Tischfield and Ruddle 1974; Kahan et al. 1974) and as a result the derived chromosome [der(16)] con- sisting of the long arm (pll-~qter) of 16 and most of the long arm of 11 (q13--~qter) is selectively retained in hybrid cells
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A highly polymorphic locus on chromosome 16q revealed by a probe from a chromosome-specific cosmid...

Hum Genet (1986) 74 : 425-431
© Springer-Verlag 1986
A highly polymorphic locus on chromosome 16q revealed by a probe from a chromosome-specific cosmid library
L. Buflon 1 , T. K. Mohandas 3, R. E. Magenis I , R. Sheehy 1, R. K. Bestwick 2, and M. Litt 1'2
1Department of Medical Genetics, and 2Department of Biochemistry, Oregon Health Sciences University, 3181 S.W. Sam Jackson Park Road, Portland, OR 97201, USA SDivision of Medical Genetics, Harbor UCLA Medical Center, Torrance, CA 90509, USA
Summary. A cosmid library was constructed from genomic D N A of a human-mouse somatic cell hybrid containing an 11q-16q translocation chromosome as the only human DNA. Cosmids with human inserts were prehybridized with total human D N A and were screened to find probes that revealed highly polymorphic loci. From one such cosmid, CF33-79, a single-copy subclone was isolated which revealed an insertion/ deletion polymorphism with at least 11 alleles and a PIC of 0.77. Using a somatic cell hybrid mapping panel, the subclone was mapped to chromosome 16. By in situ hybridization with the entire cosmid used as a probe, chromosomal localization was shown at 16q22---~24.
Introduction
Restriction fragment length polymorphisms (RFLP) are use- ful markers for mapping the human genome (Botstein et al. 1980). As the number of known RFLPs increases, they are be- coming ever more useful in the prenatal or early diagnosis of numerous hereditary diseases. Also of significance is the no- tion that mapping of a disease gene to a specific chromosomal location may serve as the first step in the cloning of the gene.
Diseases that have been mapped by linkage studies with RFLPs include Huntington's disease (Gusella et al. 1983), Duchenne's muscular dystrophy (Murray et al. 1982), X- linked retinitis pigmentosa (Bhattacharya et al. 1984), adult polycystic kidney disease (Reeders et al. 1985) and cystic fibrosis (Tsui et al. 1985). RFLPs have also been crucial to the elucidation of mechanisms underlying certain hereditary can- cer syndromes frequently associated with chromosome dele- tions such as retinoblastoma (Cavenee et al. 1983) and Wilm's tumor (Koufos et al. 1984). In the future, RFLPs may be use- ful in characterizing the genetic contributions to suscep- tibilities to common diseases that tend to cluster in families, such as colon cancer and schizophrenia (White et al. 1985).
For the construction of linkage maps of human chromo- somes, marker loci with multiple alleles and polymorphism in- formation contents (PIC) (Botstein et al. 1980) near 1.0 are very useful. Among the 333 RFLPs reported to date, only about 10% have PIC values greater than 0.5 (Willard et al. 1985). To address this problem, we have used a new method for rapidly screening cosmids and other repeat-containing D N A clones to identify those with inserts homologous to
Offprint requests to: M. Litt, Department of Medical Genetics, Ore- gon Health Sciences University, 3181 S. W. Sam Jackson Park Road, Portland, OR 97201, USA
genomic regions especially rich in RFLPs (Litt and White 1985; Sealey et al. 1985). Radioactively labelled probes are prehybridized with a vast excess of non-radioactive total human D N A under conditions which drive repetitive (but not single-copy) DNA into duplex form. These probes are used directly on Southern blots of restricted DNAs from panels of unrelated individuals to visualize low- and single-copy bands. Probes that display multiple polymorphisms by this method are further studied to obtain single-copy subclones which re- veal RFLPs. We have successfully used these methods to characterize highly polymorphic loci on chromosomes 2 (Litt and White 1985; Litt et al. 1986a), 12 (Buroker et al. 1986), 19 (Burton et al. 1986) and 20 (Litt et al. 1986b).
The distribution of reported RFLPs is very uneven. Whereas chromosomes such as 4, 21 and the X are well en- dowed with RFLPs, others, such as chromosomes 8, 10, 15, 18 and 20 have 4 or fewer (Willard et al. 1985). Furthermore, certain chromosome arms are nearly devoid of RFLPs with PIC values > 0.5. In particular, only three polymorphic loci have been assigned to the long arm of chromosome 11, whereas only two have been assigned to the long arm of chro- mosome 16; of these five loci, only A P O A I on l l q has a PIC > 0.5 (Willard et al. 1985). In order to fill in these gaps in the human linkage map, we have made genomic libraries from a mouse-human somatic cell hybrid containing an 11q-16q translocation as the only human chromosome and we are using these libraries as sources of probes to reveal high PIC RFLPs on these chromosome arms. In this paper we describe a highly polymorphic locus on the distal half of the long arm of chromosome 16 revealed by a probe isolated from a cosmid library of the l l q -16q translocation hybrid.
Methods
Somatic cell hybrids
The cell hybrid used for library construction was isolated from a fusion of mouse A9 cells (GM0346, Institute for Medical Re- search, Camden, N.J.), deficient in adenine phosphoribosyl transferase (APRT) and human fibroblasts containing a trans- location between chromosomes 16 and 11 (46,XX,t(l l ;16) (q13;p11); GM3005). Hybrids were isolated in medium that selects for APRT as described previously (Koeffler et al. 1981). Human APRT is located on the long arm of human chromosome 16 (Tischfield and Ruddle 1974; Kahan et al. 1974) and as a result the derived chromosome [der(16)] con- sisting of the long arm (p l l -~q t e r ) of 16 and most of the long arm of 11 (q13--~qter) is selectively retained in hybrid cells
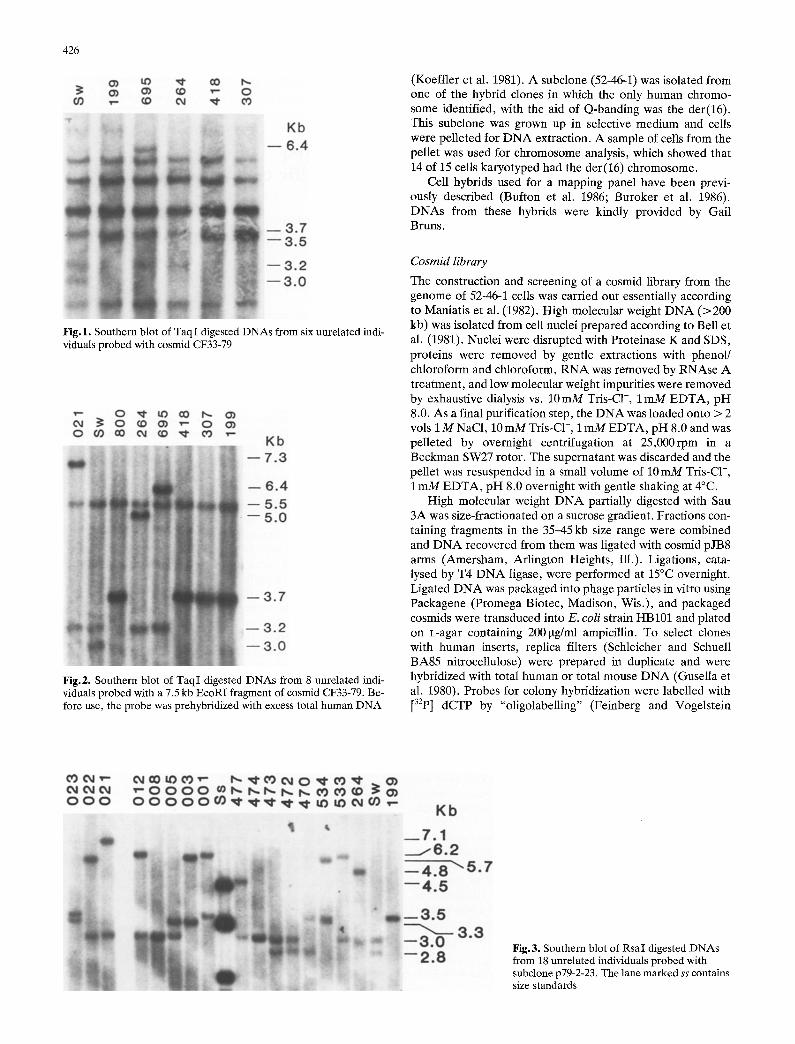
426
Fig. 1. Southern blot of TaqI digested DNAs from six unrelated indi- viduals probed with cosmid CF33-79
Fig.2. Southern blot of TaqI digested DNAs from 8 unrelated indi- viduals probed with a 7.5 kb EcoRI fragment of cosmid CF33-79. Be- fore use, the probe was prehybridized with excess total human DNA
(Koeffler et al. 1981). A subclone (52-46-1) was isolated from one of the hybrid clones in which the only human chromo- some identified, with the aid of Q-banding was the der(16). This subclone was grown up in selective medium and cells were pelleted for D N A extraction. A sample of cells from the pellet was used for chromosome analysis, which showed that 14 of 15 cells karyotyped had the der(16) chromosome.
Cell hybrids used for a mapping panel have been previ- ously described (Burton et al. 1986; Buroker et al. 1986). DNAs from these hybrids were kindly provided by Gaff Bruns.
Cosmid library
The construction and screening of a cosmid library from the genome of 52-46-1 cells was carried out essentially according to Maniatis et al. (1982). High molecular weight DNA (> 200 kb) was isolated from cell nuclei prepared according to Bell et al. (1981). Nuclei were disrupted with Proteinase K and SDS, proteins were removed by gentle extractions with phenol/ chloroform and chloroform, RNA was removed by RNAse A treatment, and low molecular weight impurities were removed by exhaustive dialysis vs. 10 mM Tris-Cl-, i mM EDTA, pH 8.0. As a final purification step, the D N A was loaded onto > 2 vols 1 M NaC1, 10 mM Tris-Cl-, 1 mM EDTA, pH 8.0 and was pelleted by overnight centrifugation at 25,000rpm in a Beckman SW27 rotor. The supernatant was discarded and the pellet was resuspended in a small volume of 10raM Tris-Cl-, i mM EDTA, pH 8.0 overnight with gentle shaking at 4°C.
High molecular weight D N A partially digested with Sau 3A was size-fractionated on a sucrose gradient. Fractions con- taining fragments in the 35-45 kb size range were combined and DNA recovered from them was ligated with cosmid pJB8 arms (Amersham, Arlington Heights, Ill.). Ligations, cata- lysed by T4 D N A ligase, were performed at 15°C overnight. Ligated DNA was packaged into phage particles in vitro using Packagene (Promega Biotec, Madison, Wis.), and packaged cosmids were transduced into E. coli strain HB101 and plated on L-agar containing 200~tg/ml ampicillin. To select clones with human inserts, replica filters (Schleicher and Schuell BA85 nitrocellulose) were prepared in duplicate and were hybridized with total human or total mouse D N A (Gusella et al. 1980). Probes for colony hybridization were labelled with [32p] dCTP by "oligolabelling" (Feinberg and Vogelstein
Fig. 3. Southern blot of Rsa I digested DNAs from 18 unrelated individuals probed with subclone p79-2-23. The lane marked ss contains size standards
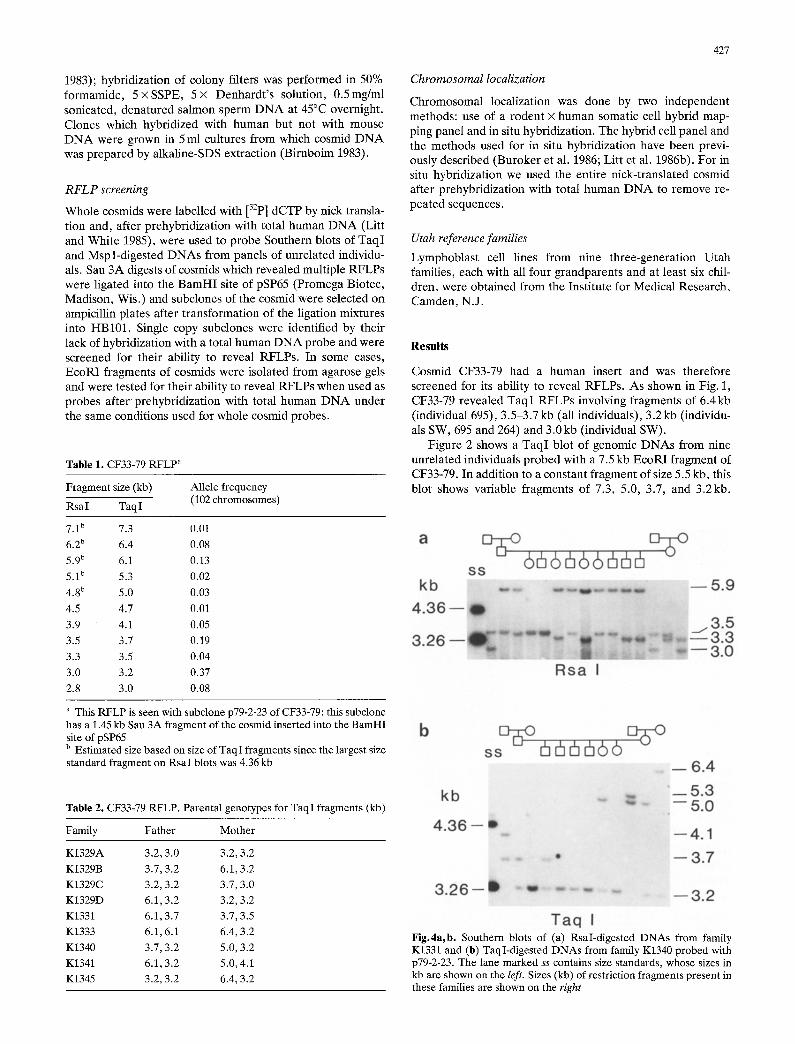
1983); hybridization of colony filters was performed in 50% formamide, 5×SSPE, 5× Denhardt's solution, 0.5mg/ml sonicated, denatured salmon sperm DNA at 45°C overnight. Clones which hybridized with human but not with mouse DNA were grown in 5 ml cultures from which cosmid DNA was prepared by alkaline-SDS extraction (Birnboim 1983).
RFLP screening
Whole cosmids were labelled with [32p] dCTP by nick transla- tion and, after prehybridization with total human DNA (Litt and White 1985), were used to probe Southern blots of TaqI and MspI-digested DNAs from panels of unrelated individu- als. Sau 3A digests of cosmids which revealed multiple RFLPs were ligated into the BamHI site of pSP65 (Promega Biotec, Madison, Wis.) and subclones of the cosmid were selected on ampicillin plates after transformation of the ligation mixtures into HB101. Single copy subclones were identified by their lack of hybridization with a total human DNA probe and were screened for their ability to reveal RFLPs. In some cases, EcoRI fragments of cosmids were isolated from agarose gels and were tested for their ability to reveal RFLPs when used as probes after prehybridization with total human DNA under the same conditions used for whole cosmid probes.
Table 1. CF33-79 RFLP a
Fragment size (kb)
RsaI TaqI
Allele frequency (102 chromosomes)
7.1 b 7.3 0.01 6.2 b 6.4 0.08 5.9 b 6.1 0.13 5.1 b 5.3 0.02 4.8 b 5.0 0.03
4.5 4.7 0.01
3.9 4.1 0.05 3.5 3.7 0.19 3.3 3.5 0.04 3.0 3.2 0.37 2.8 3.0 0.08
This RFLP is seen with subclone p79-2-23 of CF33-79: this subclone has a 1.45 kb Sau 3A fragment of the cosmid inserted into the BamHI site of pSP65 b Estimated size based on size of TaqI fragments since the largest size standard fragment on RsaI blots was 4.36 kb
427
Chromosomal localization
Chromosomal localization was done by two independent methods: use of a rodent x human somatic cell hybrid map- ping panel and in situ hybridization. The hybrid cell panel and the methods used for in situ hybridization have been previ- ously described (Buroker et al. 1986; Litt et al. 1986b). For in situ hybridization we used the entire nick-translated cosmid after prehybridization with total human DNA to remove re- peated sequences.
Utah reference families
Lymphoblast cell lines from nine three-generation Utah families, each with all four grandparents and at least six chil- dren, were obtained from the Institute for Medical Research, Camden, N.J.
Results
Cosmid CF33-79 had a human insert and was therefore screened for its ability to reveal RFLPs. As shown in Fig. 1, CF33-79 revealed TaqI RFLPs involving fragments of 6.4kb (individual 695), 3.5-3.7 kb (all individuals), 3.2 kb (individu- als SW, 695 and 264) and 3.0 kb (individual SW).
Figure 2 shows a TaqI blot of genomic DNAs from nine unrelated individuals probed with a 7.5 kb EcoRI fragment of CF33-79. In addition to a constant fragment of size 5.5 kb, this blot shows variable fragments of 7.3, 5.0, 3.7, and 3.2kb.
Table 2. CF33,79 RFLP. Parental genotypes for TaqI fragments (kb)
Family Father Mother
K1329A KI329B K1329C K1329D K1331 K1333 K1340 K1341 K1345
3.2, 3.0 3.7.3.2 3.2.3.2 6.1.3.2 6.1.3.7 6.1.6.1 3.7.3.2 6.1.3.2 3.2.3.2
3.2,3.2 6.1, 3.2 3.7, 3.0 3.2, 3.2 3.7,3.5 6.4, 3.2 5.0, 3.2 5.0,4.1 6.4, 3.2
Fig.4a, b. Southern blots of (a) RsaI-digested DNAs from family K1331 and (b) TaqI-digested DNAs from family K1340 probed with p79-2-23. The lane marked ss contains size standards, whose sizes in kb are shown on the left. Sizes (kb) of restriction fragments present in these families are shown on the right

428
Fig. 5. S o u t h e r n blot of H i n d I I I digests of D N A s f r o m p a r e n t a l and hybr id cell lines p r o b e d wi th p79-2-23. E36 and R A G are the h a m s t e r and
m o u s e pa ren t s . T h e l ane m a r k e d ss conta ins size s tandards . Sizes of human-spec i f i c H i n d I I I f r a g m e n t s which hybr id ize to the p r o b e a re shown
on the left
T a b l e 3. C h r o m o s o m e con ten t s of hyb r id cell lines. T h e des igna t ions are: + , p r e s e n c e o r - , absence of a h u m a n c h r o m o s o m e ; R, r e a r r a n g e d
c h r o m o s o m e as d e t e r m i n e d by d is rupt ion of a Syntenic g r o u p o r by a cy togene t i c abno rma l i t y ; + , c h r o m o s o m e p re sen t in less t han 15% of cells and /o r the i sozyme o r D N A p r o b e charac te r i s t ic of the c h r o m o s o m e w e a k l y posi t ive; p, p r e s e n c e of a m a r k e r for the shor t a r m and absence of a
m a r k e r for the long a r m ; q, p r e s e n c e of a m a r k e r for the long a r m and absence of a m a r k e r for the shor t a rm; a, Xq24---~qter; N D , no t done . Fo r ca lcula t ion o f the d i sco rdancy f ract ions , hybr ids wi th a r e a r r a n g e d c h r o m o s o m e or those w h e r e the c h r o m o s o m e was p r e sen t in less t han 15% of
the cells w e r e exc luded f r o m analysis. T h e c o l u m n des igna ted M indicates the p r e s e n c e or absence of human-spec i f i c T a q I f r a g m e n t s in Sou the rn
t r ans fe r p r o b e d wi th p79-2-23
Cell line C h r o m o s o m e c o m p l e m e n t Scor-
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 X Y ing M
G 3 5 D 5
G35F3
G 3 5 D 2
G 3 5 E 3
G 3 5 A 2
G35C2
G35C4
G35B5
G35F5
G 3 5 D 3
G35C5
G 3 5 A 4
G 3 5 B 4
G 3 5 E 4
G175
G 2 4 A 9
G 2 4 A 4
G24B5
G 8 9 E 5 a
G 9 5 A 4
G 3 5 A 5 A z A b
G 3 5 A 5 A z F b
R R P 5 - 7
R R P 3 - 6 B 1
G35F1
4- 4- 4- 4- 4- 4-
-- 4- 4- -- 4- -- 4- 4- -- 4- + 4-
___ 4- 4- 4- -- 4- 4-
4- + -- 4-
4- _+ -- 4-
4- 4-
+ R
R +
4- 4- 4- + 4- 4- 4- 4- - 4- a -
4- - 4- 4- 4- a -
4- - - + R + - + a R -
+ + + + - + - + + + + + a -
4- 4- - _ 4- 4- 4- 4- 4- -
R + - + R R -
4- - 4- - - - _+ 4- 4- 4- - a -
+ p + - + - + + a -
___ - 4- 4- -- 4-
-- p - _+ + + + +
p - + - + + + + - p
4- -- 4- 4- -- 4- -- 4- 4-
4- 4- 4- 4- 4-
R R
_+ 4- - 4- - 4- - 4-
- 4- - + - 4- - 4- 4- 4-
-+ p + + + - + - + - + + + +
4- - 4- 4- 4- 4- 4- 4- 4- 4- - 4-
+ + + -- +
+ 4- 4- 4- 4- 4- 4-
4- 4-
+ - + - + + - + + + + _+ + + + -
+ - + + - + + + + a -
+ + + - + + + + a -
+ - + + + + - a -
_ _ + + + + -
+ - + a -
+ + + - + + a -
+ + + - + + + - + - q -
+ - + - q -
+ - + + + -
+ -
_ - } - 4 - - -
+
+ - + N D + - - + + - + + R
+ + + + + + + R - + - + +
+ - + - p - + - - - + + + + - a -
D i s c o r d a n c y 8 7 5 8 8 10 9 8 8 11 6 9 9 9 11 0 10 12 10 7 10 11 5 8
F rac t ion 19 21 22 20 23 22 23 23 20 22 21 20 21 22 23 24 22 22 20 21 23 23 8 22
+
+
+
+
+
+
+
+
+
+
+
+
a Cell l ine G 8 9 E 5 conta ins an X c h r o m o s o m e in all cells plus an un iden t i f i ed h u m a n - h a m s t e r t r ans loca t ion in abou t 20% of the cells b Cell lines G 3 5 A 5 A z A and G 3 5 A 5 A z F are subc lones of G 3 5 A 5 de l ec t ed in 8 -azaguan ine for loss of the de r (19 ) t rans loca t ion c h r o m o s o m e t ( X ; 1 9 ) q 2 3 - 2 5 : : q13 (La t t et al. 1976). A l t h o u g h these cell lines h a v e no t b e e n ex tens ive ly cha rac t e r i zed , they bo th hybr id ized to a p r o b e for
hap tog lob in , k n o w n to res ide on c h r o m o s o m e [email protected] (Wil lard et al. 1985)
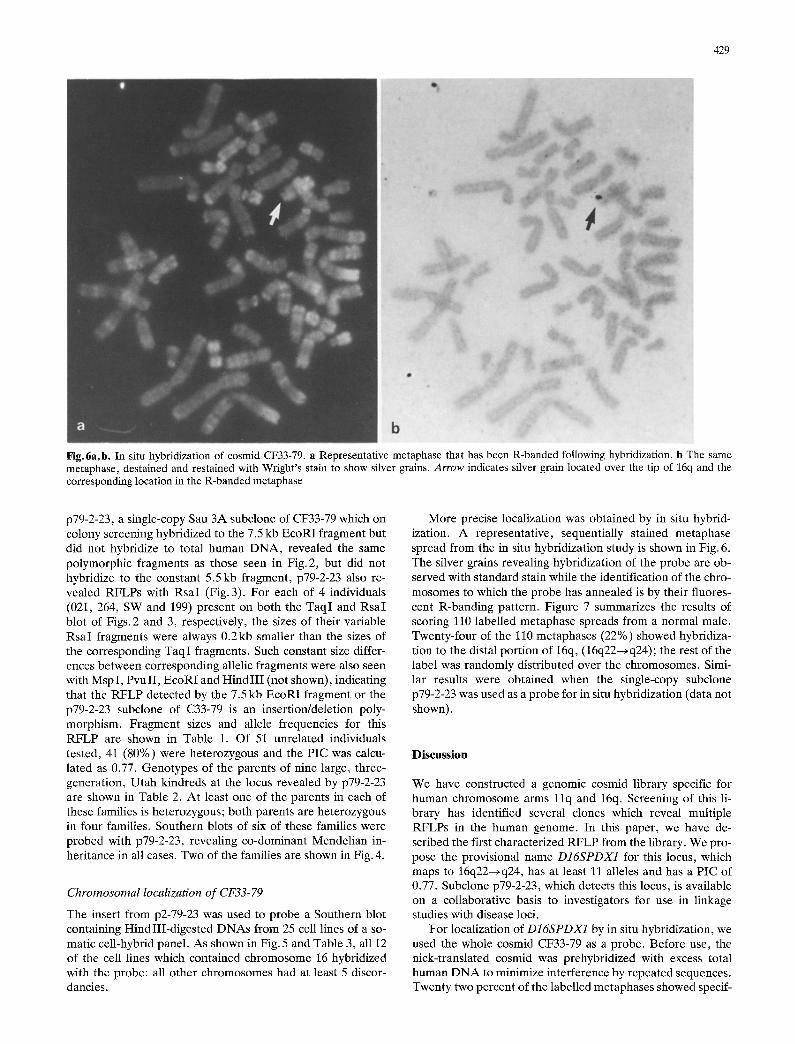
429
Fig.6a, b. In situ hybridization of cosmid CF33-79. a Representative metaphase that has been R-banded following hybridization, b The same metaphase, destained and restained with Wright's stain to show silver grains. Arrow indicates silver grain located over the tip of 16q and the corresponding location in the R-banded metaphase
p79-2-23, a single-copy Sau 3A subclone of CF33-79 which on colony screening hybridized to the 7.5 kb EcoRI fragment but did not hybridize to total human DNA, revealed the same polymorphic fragments as those seen in Fig. 2, but did not hybridize to the constant 5.5 kb fragment, p79-2-23 also re- vealed RFLPs with RsaI (Fig. 3). For each of 4 individuals (021, 264, SW and 199) present on both the TaqI and RsaI blot of Figs. 2 and 3, respectively, the sizes of their variable RsaI fragments were always 0.2kb smaller than the sizes of the corresponding TaqI fragments. Such constant size differ- ences between corresponding allelic fragments were also seen with Msp I, Pvu II, EcoRI and Hind III (not shown), indicating that the RFLP detected by the 7.5 kb EcoRI fragment or the p79-2-23 subclone of C33-79 is an insertion/deletion poly- morphism. Fragment sizes and allele frequencies for this RFLP are shown in Table 1. Of 51 unrelated individuals tested, 41 (80%) were heterozygous and the PIC was calcu- lated as 0.77. Genotypes of the parents of nine large, three- generation, Utah kindreds at the locus revealed by p79-2-23 are shown in Table 2. At least one of the parents in each of these families is heterozygous; both parents are heterozygous in four families. Southern blots of six of these families were probed with p79-2-23, revealing co-dominant Mendelian in- heritance in all cases. Two of the families are shown in Fig. 4.
Chromosomal localization o f CF33-79
The insert from p2-79-23 was used to probe a Southern blot containing HindIII-digested DNAs from 25 cell lines of a so- matic cell-hybrid panel. As shown in Fig. 5 and Table 3, all 12 of the cell lines which contained chromosome 16 hybridized with the probe: all other chromosomes had at least 5 discor- dancies.
More precise localization was obtained by in situ hybrid- ization. A representative, sequentially stained metaphase spread from the in situ hybridization study is shown in Fig. 6. The silver grains revealing hybridization of the probe are ob- served with standard stain while the identification of the chro- mosomes to which the probe has annealed is by their fluores- cent R-banding pattern. Figure 7 summarizes the results of scoring 110 labelled metaphase spreads from a normal male. Twenty-four of the 110 metaphases (22%) showed hybridiza- tion to the distal portion of 16q, (16q22--*q24); the rest of the label was randomly distributed over the chromosomes. Simi- lar results were obtained when the single-copy subclone p79-2-23 was used as a probe for in situ hybridization (data not shown).
Discussion
We have constructed a genomic cosmid library specific for human chromosome arms 11q and 16q. Screening of this li- brary has identified several clones which reveal multiple RFLPs in the human genome. In this paper, we have de- scribed the first characterized RFLP from the library. We pro- pose the provisional name D16SPDX1 for this locus, which maps to 16q22---~q24, has at least 11 alleles and has a PIC of 0.77. Subclone p79-2-23, which detects this locus, is available on a collaborative basis to investigators for use in linkage studies with disease loci.
For localization of D16SPDX1 by in situ hybridization, we used the whole cosmid CF33-79 as a probe. Before use, the nick-translated cosmid was prehybridized with excess total human DNA to minimize interference by repeated sequences. Twenty two percent of the labelled metaphases showed specif-

430
cc ¢g
U. 0
o l ~ • •
' ' : ' r ' " : " 1 2 3 4 5
6 7 8 9 10 11 12 Z
13
t o o l l e Q l a Q
• • • • • g • • •
14 15 16 17 18 19 2 0 21 2 2 X Y
Fig.7. Histogram of chromosomal distribution of silver grains from in situ hybridization of cosmid CF33-79 to chromosomes of a normal male. Each dot represents a grain observed over a specific chromosomal region. Twenty-four of 110 cells examined had a grain localized to the region 16q22--+ q24
ic hybridization, a value similar to that commonly found with single-copy probes (Bufton et al. 1986; Litt et al. 1986a,b). This approach also succeeded for cosmid 1-11 (Buroker et al. 1986) as well as for several bacteriophage clones (Donlon et al. 1986; R.Sheehy, personal communication); we propose that it may be generally useful for in situ hybridization of probes containing repeated sequences.
Two other polymorphic loci have been mapped to 16q. These are adenine phosphoribosyl transferase (APRT) at 16q22 and haptoglobin, at 16q22.1 (Willard et al. 1985). Al- though each of these loci contains several RFLPs, the publish- ed data do not allow evaluation of their heterozygosities or PICs. Other genes assigned to 16q include tyrosine amino- transferase (Barton et al. 1986), diaphorase-4 (D1A4) (Lavinha et al. 1984), glutamic-oxaloacetic transaminase-2 (GOT2) (Jeremiah et al. 1982), lecithin-cholesterol acyl- transferase (LCAT) (Azoulay et al. 1985) and the metallo- thionein gene family (MT1 and MT2) (Le Beau et al. 1985). Two fragile sites (FRA16B, Magenis et ah 1970; and FRA16D, Sutherland 1983) have been mapped to 16q.
Several other clones from our l lq -16q specific cosmid library and also from a phage library of the 11q-16q transloca- tion hybrid (C.Maslen-McClure, personal communication) reveal multiple RFLPs which have been localized to chromo- some 11 or 16. Probes which reveal such RFLPs will be used to type the CEPH families (Marx 1985), facilitating construc- tion of linkage maps of the long arms of these chromosomes.
Acknowledgements. Excellent technical assistance was provided by Ruth Litt. We thank Gail Bruns for providing the somatic cell hybrid mapping panel. This work was supported by NIH Grant RO1- GM32500 and by Maternal and Child Health grant MCT 920. Lind• Burton was supported by a Medical Scientist Scholarship from the Life and Health Insurance Medical Research Fund.
References
Azoulay M, Henry I, Tata F, Weil D, Grzeschik KH, Chavez E, McIntyre N, Williamson R, Humphries SE, Junien C (1985) The structural gene for human lecithin: cholesterol acyl transferase (LCAT) maps to 16q22. (8th International Workshop on Human Gene Mapping) Cytogenet Cell Genet 40 : 573
Barton DE, Yang-Feng TL, Francke U (1986) The human tyrosine aminotransferase gene mapped to the long arm of chromosome 16 (region 16q22--*24) by somatic cell hybrid analysis and in situ hy- bridization. Hum Genet 72 : 221-224
Bell GI, Karam JH, Rutter WJ (1981) Polymorphic DNA region adja- cent to the 5' end of the human insulin gene. Proc Natl Acad Sci USA 78: 5759-5763
Bhattacharya SS, Wright AF, Clayton JF, Price WH, Phillips CI, McKeown CME, Jay M, Bird AC, Pearson PL, Southern EM, Evans HJ (1984) Close genetic linkage between X-linked retinitis pigmentosa and a restriction fragment length polymorphism iden- tified by recombinant DNA probe L1.28. Nature 309 : 253-255
Birnboim HC (1983) A rapid alkaline extraction method for the isola- tion of plasmid DNA. Methods Enzymol 100 : 243-249

431
Botstein D, White RL, Skolnick M, Davis RW (1980) Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am J Hum Genet 32:314-331
Bufton L, Bruns GAP, Magenis RE, Tomar D, Shaw D, Brook D, Litt M (1986) Four restriction fragment length polymorphisms re- vealed by probes from a single cosmid map to chromosome 19. Am J Hum Genet 38 : 447-460
Buroker NE, Magenis RE, Weliky K, Brnns GAP, Litt M (1986) Four restriction fragment length polymorphisms revealed by probes from a single cosmid map to human chromosome 12q. Hum Genet 72: 86-94
Cavenee WK, Dryja TP, Phillips RA, Benedict WF, Godbout R, Gal- lic BR, Murphree AL, Strong LC, White RL (1983) Expression of recessive alleles by chromosomal mechanisms in retinoblastoma. Nature 305 : 779-784
Donlon TA, Lalande M, Wyman A, Bruns G, Latt SA (1986) Isola- tion of molecular probes associated with the chromosome 15 insta- bility in the Prader-Willi syndrome. Proc Natl Acad Sci USA 83 : 4408-4412
Feinberg AP, Vogelstein B (1983) A technique for radiolabelling DNA restriction endonuclease fragments to high specific activity. Anal Biochem 132: 6-13
Gusella JF, Keys C, Varsanyi-Breiner A, Kao F-T, Jones C, Puck TT, Housman D (1980) Isolation and localization of DNA segments from specific human chromosomes. Proc Natl Acad Sci USA 77 : 2829-2833
Gusella JF, Wexler NS, Coneally PM, Naylor SL, Anderson MA, Tanzi RE, Watkins PC, Ottina K, Wallace MR, Sakaguchi AY, Young AB, Shoulson I, Bonilla E, Martin JB (1983) A poly- morphic DNA marker genetically linked to Huntington's disease. Nature 306 : 234238
Jeremiah SJ, Povey S, Burley MW, Kielty C, Lee M, Spowart G, Cor- ney C, Cook PJL (1982) Mapping studies on human mitochondrial glutamate oxaloacetate transaminase. Ann Hum Genet 46:145- 152
Kahan B, Held HR, Demars R (1974) The locus for human adenine phosphoribosyl transferase is on chromosome 16. Genetics 78: 1143-1156
Koeffler HP, Sparkes RS, Stang H, Mohandas T (1981) Regional as- signment of genes for human u-globin and phosphoglycollate phosphatase to the short arm of chromosome 16. Proc Natl Acad Sci USA 78 : 7015-7018
Kuofos A, Hansen MF, Lampkin BC, Workman ML, Copeland NG, Jenkins NA, Cavenee WK (1984) Loss of alleles at loci on human chromosome l i during genesis of Wilm's tumour. Nature 309: 170-172
Latt SA, Willard HF, Gerald PS (1976) BrDU-33258 Hoeehst analysis of DNA replication in human lymphocytes with supernumerary or structurally abnormal chromosomes. Chromosoma 57:135-153
Lavinha J, Morrison N, Glasgow L, Ferguson-Smith MA (1984) Fur- ther evidence for the regional localization of human APRT and DIA4 on chromosome 16. (7th International Workshop on Human Gene Mapping) Cytogenet Cell Genet 37:517
Le Beau MM, Diaz MO, Karin M, Rowley JD (1985) Metallothionein gene cluster is split by chromosome 16 rearrangements in myelo- monocytic leukemia. Nature 313 : 709-711
Litt M, White R (1985) A highly polymorphic locus in human DNA revealed by cosmid-derived probes. Proc Natl Acad Sci USA 82 : 6206-6210
Litt M, Bruns GA, Sheehy R, Magenis RE (1986a) A highly poly- morphic locus in human DNA revealed by probes from cosmid 1- 5 maps to chromosome 2q35~37. Am J Hum Genet 38:288-296
Litt M, Sheehy R, Brnns GA, Magenis RE (1986b) A polymorphic locus on the long arm of chromosome 20 defined by two probes from a single cosmid. Hum Genet 73 : 340-345
Magenis RE, Hecht F, Lovrien EW (1970) Heritable fragile site on chromosome 16: probable localization of haptoglobin locus in man. Science 170 : 85-87
Maniatis T, Fritsch EF, Sambrook J (1982) Molecular cloning: a lab- oratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
Marx JL (1985) Putting the human genome on the map. Science 29 : 150-151
Murray JM, Davies KE, Harper PS, Meredith L, Mueller CR, Wil- liamson R (1982) Linkage relationship of a cloned DNA sequence on the short arm of the X chromosome to Duchenne muscular dystrophy. Nature 300 : 542-544
Reeders ST, Breuning MH, Davies KE, Nicholls RD, Jarman AP, Higgs DR, Pearson PL, Weatherall DJ (1985) A highly poly- morphic DNA marker linked to adult polycystic kidney disease on chromosome 16. Nature 317:542-544
Sealey PG, Whittaker PA, Southern EM (1985) Removal of repeated sequences from hybridization probes. Nucleic Acids Res 13 : 1905- 1922
Sutherland GR (1983) The fragile X chromosome. Int Rev Cytol 81 : 107-143
Tischfield JA, Ruddle FH (1974) Assignment of the gene for adenine phosphoribosyltransferase to human chromosome 16 by mouse- human somatic cell hybridization. Proc Natl Acad Sci USA 71 : 45-49
Tsui LC, Buchwald M, Barker D, Braman JC, Knowlton R, Schumm JW, Eiberg H, Mohr J, Kennedy D, Plasvic N, Zsiga M, Mar- kiewicz D, Akots G, Brown V, Helms C, Gravius T, Parker C, Rediker K, Donis-Keller H (1985) Cystic fibrosis locus defined by a genetically linked polymorphic DNA marker. Science 230: 1054--1056
White R, Leppert M, Bishop DT, Barker D, Berkowitz J, Brown C, Callahan P, Holm T, Jerominski L (1985) Construction of linkage maps with DNA markers for human chromosomes. Nature 313 : 101-105
Willard HF, Skolnick MH, Pearson PL, Mandel J-L (1985) Report of the Committee on Human Gene Mapping by Recombinant DNA Techniques. (8th International Workshop on Human Gene Map- ping) Cytogenet Cell Genet 40 : 360M90
Received May 16, 1986 / Revised July 23, 1986




















