
Comparison of Human Chromosome 6p25 with Mouse Chromosome 13 Reveals a Greatly Expanded Ov-Serpin...
14
GENOMICS Vol. 79, Number 3, March 2002 Copyright © 2002 Elsevier Science (USA). All rights reserved. 0888-7543/02 $35.00 349 Article doi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL INTRODUCTION Proteins of the serpin superfamily are found in multicellular organisms and some viruses. Many serpins are extracellular protease inhibitors involved in diverse processes such as the regulation of blood coagulation, complement activation, fib- rinolysis, and matrix remodeling. Serpin mutants are impli- cated in tumor progression, thrombosis, emphysema, angioedema, and familial dementia [reviewed in 1]. The most striking feature of a serpin is an exposed reactive center loop (RCL) located 30–40 amino acids from the carboxy terminus that resembles the natural substrate of the target protease, and contains a peptide bond cleaved during the protease–ser- pin interaction [2]. Cleavage causes conformational change in the serpin, accompanied by major structural distortion and inactivation of the protease. The protease and serpin remain Comparison of Human Chromosome 6p25 with Mouse Chromosome 13 Reveals a Greatly Expanded Ov-Serpin Gene Repertoire in the Mouse Dion Kaiserman, 1 Susan Knaggs, 2 Katrina L. Scarff, 1 Anneliese Gillard, 1 Ghazala Mirza, 2 Matthew Cadman, 3 Richard McKeone, 3 Paul Denny, 3 Jessica Cooley, 4 Charaf Benarafa, 4 Eileen Remold-O’Donnell, 4 Jiannis Ragoussis, 2 and Phillip I. Bird 1, * 1 Department of Biochemistry and Molecular Biology, Monash University 3800, Australia 2 The Genomics Laboratory, Division of Medical Molecular Genetics, GKT Medical School, King’s College, London SE1 9RT, UK 3 MRC UK Mouse Genome Centre and Mammalian Genetics Unit, Harwell, Oxfordshire OX11 ORD, UK 4 Center for Blood Research, Harvard Medical School, Boston, Massachusetts 02115, USA *To whom correspondence and reprint requests should be addressed. Fax: (+61) 3 9905 4699. E-mail: [email protected]. Ov-serpins are intracellular proteinase inhibitors implicated in the regulation of tumor pro- gression, inflammation, and cell death. The 13 human ov-serpin genes are clustered at 6p25 (3 genes) and 18q21 (10 genes), and share common structures. We show here that a 1-Mb region on mouse chromosome 13 contains at least 15 ov-serpin genes compared with the three ov-serpin genes within 0.35 Mb at human 6p25 (SERPINB1 (MNEI), SERPINB6 (PI-6), SER- PINB9 (PI-9)). The mouse serpins have characteristics of functional inhibitors and fall into three groups on the basis of similarity to MNEI, PI-6, or PI-9. The genes map between the mouse orthologs of the Werner helicase interacting protein and NAD(P)H menadioine oxi- doreductase 2 genes, in a region that contains the markers D13Mit136 and D13Mit116. They have the seven-exon structure typical of human 6p25 ov-serpin genes, with identical intron phasing. Most show restricted patterns of expression, with common sites of synthesis being the placenta and immune tissue. Compared with human, this larger mouse serpin repertoire probably reflects the need to regulate a larger proteinase repertoire arising from differing evo- lutionary pressures on the reproductive and immune systems. Key Words: serpin, ov-serpin, SPI3, SPI6, PI-6, PI-9, MNEI, phylogeny, gene structure covalently linked, and the complex is eventually removed from the system and degraded. Thus serpins function as sui- cide substrates regulating proteolysis [1]. Phylogenic studies clearly show that serpins fall into a number of distinct clades [3], and as a consequence human serpin gene nomenclature is now clade-based [1]. Comparison of serpin gene localization and structures shows that the genes for members of the same clade are often clustered, and have very similar numbers and positioning of introns [4,5]. In humans there are nine serpin clades: the largest, clade A, com- prises the 1-antitrypsin-like serpins; the next largest, clade B, comprises the ovalbumin (ov)-like serpins [3]. Ov-serpins are emerging as a large class of serpins involved in the regulation of tumor progression, cell differ- entiation, and cell survival [reviewed in 1]. They are pre- dominantly intracellular molecules, although release from cells can occur under certain circumstances, for example in
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Comparison of Human Chromosome 6p25 with Mouse Chromosome 13 Reveals a Greatly Expanded Ov-Serpin...

Articledoi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
Comparison of Human Chromosome 6p25 with MouseChromosome 13 Reveals a Greatly Expanded Ov-Serpin
Gene Repertoire in the MouseDion Kaiserman,1 Susan Knaggs,2 Katrina L. Scarff,1 Anneliese Gillard,1 Ghazala Mirza,2Matthew Cadman,3 Richard McKeone,3 Paul Denny,3 Jessica Cooley,4 Charaf Benarafa,4
Eileen Remold-O’Donnell,4 Jiannis Ragoussis,2 and Phillip I. Bird1,*
1Department of Biochemistry and Molecular Biology, Monash University 3800, Australia2The Genomics Laboratory, Division of Medical Molecular Genetics, GKT Medical School, King’s College, London SE1 9RT, UK
3MRC UK Mouse Genome Centre and Mammalian Genetics Unit, Harwell, Oxfordshire OX11 ORD, UK4Center for Blood Research, Harvard Medical School, Boston, Massachusetts 02115, USA
*To whom correspondence and reprint requests should be addressed. Fax: (+61) 3 9905 4699. E-mail: [email protected].
Ov-serpins are intracellular proteinase inhibitors implicated in the regulation of tumor pro-gression, inflammation, and cell death. The 13 human ov-serpin genes are clustered at 6p25(3 genes) and 18q21 (10 genes), and share common structures. We show here that a 1-Mbregion on mouse chromosome 13 contains at least 15 ov-serpin genes compared with the threeov-serpin genes within 0.35 Mb at human 6p25 (SERPINB1 (MNEI), SERPINB6 (PI-6), SER-PINB9 (PI-9)). The mouse serpins have characteristics of functional inhibitors and fall intothree groups on the basis of similarity to MNEI, PI-6, or PI-9. The genes map between themouse orthologs of the Werner helicase interacting protein and NAD(P)H menadioine oxi-doreductase 2 genes, in a region that contains the markers D13Mit136 and D13Mit116. Theyhave the seven-exon structure typical of human 6p25 ov-serpin genes, with identical intronphasing. Most show restricted patterns of expression, with common sites of synthesis beingthe placenta and immune tissue. Compared with human, this larger mouse serpin repertoireprobably reflects the need to regulate a larger proteinase repertoire arising from differing evo-lutionary pressures on the reproductive and immune systems.
Key Words: serpin, ov-serpin, SPI3, SPI6, PI-6, PI-9, MNEI, phylogeny, gene structure
covalently linked, and the complex is eventually removed
INTRODUCTIONProteins of the serpin superfamily are found in multicellularorganisms and some viruses. Many serpins are extracellularprotease inhibitors involved in diverse processes such as theregulation of blood coagulation, complement activation, fib-rinolysis, and matrix remodeling. Serpin mutants are impli-cated in tumor progression, thrombosis, emphysema,angioedema, and familial dementia [reviewed in 1]. The moststriking feature of a serpin is an exposed reactive center loop(RCL) located 30–40 amino acids from the carboxy terminusthat resembles the natural substrate of the target protease,and contains a peptide bond cleaved during the protease–ser-pin interaction [2]. Cleavage causes conformational change inthe serpin, accompanied by major structural distortion andinactivation of the protease. The protease and serpin remain
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.0888-7543/02 $35.00
from the system and degraded. Thus serpins function as sui-cide substrates regulating proteolysis [1].
Phylogenic studies clearly show that serpins fall into anumber of distinct clades [3], and as a consequence humanserpin gene nomenclature is now clade-based [1]. Comparisonof serpin gene localization and structures shows that thegenes for members of the same clade are often clustered, andhave very similar numbers and positioning of introns [4,5]. Inhumans there are nine serpin clades: the largest, clade A, com-prises the �1-antitrypsin-like serpins; the next largest, cladeB, comprises the ovalbumin (ov)-like serpins [3].
Ov-serpins are emerging as a large class of serpinsinvolved in the regulation of tumor progression, cell differ-entiation, and cell survival [reviewed in 1]. They are pre-dominantly intracellular molecules, although release fromcells can occur under certain circumstances, for example in
349

Article doi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
FIG. 1. Amino acid sequence alignment of the mouse chromosome 13ov-serpins. New serpin sequences were deduced from direct sequenc-ing of cDNA clones (SPI3C, R86, NK10, NK13, NK21B, NK26, EIA,EIB), or predicted from public nonredundant and htgs data (EIC, SPI3B,SPI3D, NK9, NK21) or the Celera database (AT2). Also shown are thesequences of the related human ov-serpins, PI-6, PI-8, PI-9, and MNEI(shaded). See Table 1 for accession numbers. Asterisks indicate pre-dicted sequences; intron/exon boundaries are indicated by arrowsabove the alignment.
response to an inflammatory stimulus or during cell necro-sis. Several ov-serpins are associated with disease: maspin isa tumor suppressor, the squamous-cell carcinoma antigens(SCCAs) appear in the circulation of patients with squamous-cell carcinoma, megsin is upregulated in IgA nephropathy,and increased plasminogen activator inhibitor-2 is correlatedwith a favorable prognosis in breast cancer patients.
The 13 human ov-serpin genes are split into two clusters,with 3 genes found on chromosome 6 and 10 on chromosome18 [4,6]. The region at chromosome 6p25 containing the ser-pin gene cluster is within a segment positioned between thegenetic markers D6S344/AFMB34ya5 and D6S1617 showingloss of heterozygosity in cervical carcinoma [7], implying thatone or more of these serpins are tumor suppressors. The 6p25
350
serpin gene cluster is also in a region deleted in patients withcraniofacial and anterior eye anomalies [8]. Similar abnor-malities are seen in Del(13)Svea36H mice hemizygous for a syn-tenic region on mouse chromosome 13 [9]. Most ov-serpingenes have eight exons identically arranged to those in thechicken ovalbumin gene in terms of both position andintron/exon boundary phasing [5]; this suggests that a pri-mordial ov-serpin gene was present in a common ancestor ofbirds and mammals. However, five of the human ov-serpingenes contain only seven exons [4], having lost the exonencoding a variable interhelical loop in the protein. Otherwisethey are identical in structure to the eight-exon genes. Takenwith the phylogenic analyses, the structure and localizationof the human ov-serpin genes suggest that the clusters have
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.
Articledoi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
Tab
le 1
con
tinu
ed o
n ne
xt p
age
TA
BL
E 1
: Acc
essi
on n
umbe
rs a
nd d
etai
ls o
f mou
se s
erpi
n cD
NA
s, E
STs,
BA
C c
lone
s, a
nd p
rim
ersa
Mou
seA
cc. n
o.G
ene
Rep
rese
ntat
ive
Htg
s cl
ones
hit
RC
L (s
) and
3’-U
TR (a
/s) p
rim
ers
used
H
uman
se
rpin
sym
bol
dbES
T cl
one
for
RT-
PCR
and
gen
omic
map
ping
orth
olog
SPI3
U25
844
(F)
Serp
inb6
RP2
3-26
2J21
(c)
No.
259
(s) 5
’-aca
gctg
gcat
gatg
acg-
3’PI
-6(A
cc. n
o. A
L450
406)
No.
370
(a/s
) 5’-g
gcaa
ttgt
gctc
aggg
agag
gaga
acc-
3’(Z
2265
8)SP
I6U
9670
0 (F
)Se
rpin
b9R
P23-
391I
11 (c
)N
o. 6
45 (s
) 5’-g
ccat
cata
gaat
tttg
ctgt
gcct
c-3
PI-9
(Acc
. no.
AL5
8987
1)N
o. 5
20 (a
/s) 5
’-ggg
atac
tgaa
gaga
gaac
tctc
cctg
tg-3
’(U
7136
4)R
P23-
312D
21(A
cc. n
o. A
L606
533)
AT2
U96
704
(S)
Serp
inb9
dN
one
foun
dR
P23-
414F
19 (p
)N
o. 6
90 (s
) 5’-g
ctgt
ttag
gttc
ttat
cccc
-3’
No.
691
(a/s
) 5’-g
gtgg
tgga
gttg
ccaa
gaga
g-3’
NK
9U
9670
6 (S
)Se
rpin
b9c
AW
5383
90 (S
)R
P23-
414F
19 (p
)N
o. 6
77 (s
) 5’-g
ccac
agct
gatg
atac
tgta
tgtt
c-3’
AK
0144
48 (F
)N
o. 6
74 (a
/s) 5
’-ggg
caca
gatg
agtg
tcag
gg-3
’
NK
10U
9670
3 (F
)Se
rpin
b8A
W90
7882
(S)
RP2
3-13
0F18
(c)
No.
636
(s) 5
’-gtg
atta
ggaa
cgcc
cggt
gctg
tag-
3’PI
-8BG
2457
99N
o. 6
37 (a
/s) 5
’-gat
cggc
aggt
tggc
acca
tcat
g-3’
(L40
377)
NK
13U
9670
7 (F
)Se
rpin
b6b
AW
2604
89 (F
)R
P23-
39I1
1 (c
)N
o. 6
04 (s
) 5’-g
ccaa
tata
ggtt
ttag
gtgt
atgg
tcc-
3’A
W25
9940
(Acc
. no.
AL5
8987
1)N
o. 6
05(a
/s) 5
’-gct
atgc
agtt
gagg
ctag
ccct
gcat
g-3’
RP2
3-31
2D21
(c)
(Acc
. no.
AL6
0653
3)N
K21
U96
708
(S)
Serp
inb9
fBG
O77
981
RP2
3-41
L21
(c)
No.
646
(s) 5
’-gca
gcct
ctgc
tgta
gaat
ttat
attt
ttat
gttc
a-3’
(Acc
. no.
AL4
5033
1)N
o. 6
35 (a
/s) 5
’-tcc
tcac
agag
cagt
aagg
c-3’
RP2
3-41
4F19
(p)
NK
21B
AF4
2508
3 (F
)Se
rpin
b9g
AA
4082
35 (F
)R
P23-
41L2
1 (c
)N
o. 6
46 (s
) 5’-g
cagc
ctct
gctg
taga
attt
atat
tttt
atgt
tca-
3’A
A40
8234
(Acc
. no.
AL4
5033
1)N
o. 6
35 (a
/s) 5
’-tcc
tcac
agag
cagt
aagg
c-3’
NK
21C
Serp
inb9
-ps1
NK
26U
9670
9 (F
)Se
rpin
b9e
AA
4094
18 (F
)R
P23-
414F
19 (p
)N
o. 6
23 (s
) 5’-t
tgca
gcct
ctgc
tggc
aaaa
ttat
actt
ttct
gtga
t-3’
AA
4094
17N
o. 6
24 (a
/s) 5
’-gtc
catt
ctca
caga
gcag
taga
ag-3
’R
86U
9670
5 (F
)Se
rpin
b9b
AA
3891
24 (F
)R
P23-
391I
11 (c
)N
o. 2
60 (s
) 5’-t
ccgc
tgag
ggca
tcat
t-3’
AK
0054
91 (F
)(A
cc. n
o. A
L589
871)
No.
496
(a/s
) 5’-a
cccc
acat
agta
atgt
gcc-
3’R
P23-
312D
21(A
cc. n
o. A
L606
533)
R86
BN
ot a
ssig
ned
Non
e fo
und
RP2
3-39
1I11
(c)
(Acc
. no.
AL5
8987
1)R
P23-
312D
21
(Acc
. no.
AL6
0653
3)
SPI3
BSe
rpin
b6e
AI5
0652
7 (S
)R
P23-
262J
21 (c
)N
o. 6
18 (s
) 5’-g
aaat
agtc
acag
tggg
gtca
cc-3
’(A
cc. n
o. A
L450
406)
No.
621
(a/s
) 5’-g
cact
tatg
gata
aggt
cact
tggg
-3’
SPI3
CA
F425
084
(F)
Serp
inb6
cBG
0788
82R
P23-
262J
21 (c
)N
o. 6
19 (s
) 5’-c
aatt
gttc
tgaa
gggg
tcat
cacg
-3’
BG06
5606
(S)
(Acc
. no.
AL4
5040
6)N
o. 6
21 (a
/s) 5
’-gca
ctta
tgga
taag
gtca
cttg
gg-3
’
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.
351
Article doi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
TAB
LE
1: c
ontin
ued
Mou
seA
cc. n
o.G
ene
Rep
rese
ntat
ive
Htg
s cl
ones
hit
RC
L (s
) and
3’-U
TR (a
/s) p
rim
ers
used
H
uman
se
rpin
sym
bol
dbES
T cl
one
for
RT-
PCR
and
gen
omic
map
ping
orth
olog
SPI3
DSe
rpin
b6d
AA
1683
65 (S
)R
P23-
3N21
No.
620
(s) 5
’-gtg
atga
tggg
tgca
tcac
caac
t-3’
(Acc
. no.
AL5
1302
2)N
o. 6
22 (a
/s) 5
’-gca
cata
tgga
taag
ataa
agtg
gg-3
’SP
I3E
Serp
inb6
-ps1
Non
e fo
und
RP2
3-26
2J21
No.
625
(s) 5
’-aat
tgtg
atga
tggg
tgta
ccac
c-3’
(Acc
. no.
AL4
5040
6)N
o. 6
26 (a
/s) 5
’-ctc
agtt
ctga
agat
ggga
tgcc
-3’
RP2
3-3N
21(A
cc. n
o. A
L513
022)
EIA
AF4
2602
4 (F
)Se
rpin
b1A
A20
1023
(F)
RP2
3-21
7N23
(c)
No.
420
(s) 5
’-gag
gcat
tgct
acat
tctg
t-3’
MN
EIR
P23-
427P
14 (c
)N
o. 4
21 (a
/s) 5
’-gag
tcct
gctt
tcat
tgta
c-3’
CT7
-236
L20
(Acc
. no.
AC
0736
66)
(M93
056)
EIB
AF4
2602
5 (F
)Se
rpin
b1b
BE65
0533
(S)
RP2
3-39
I11
(c)
No.
608
(s) 5
’-ggc
atta
ttca
ggtg
ctct
gcga
gaag
-3’
AK
0182
26 (F
)(A
cc. n
o. A
L589
871)
No.
609
(a/s
) 5’-g
ggaa
tgaa
tagc
taag
ctct
gcct
tc-3
’EI
CSe
rpin
b1c
Non
e fo
und
RP2
3-21
7N23
(c)
No.
692
(s) 5
’-gtt
ggat
gctg
cctg
atgc
cc-3
’R
P23-
312D
21N
o. 6
93 (a
/s) 5
’-tta
tggt
caag
ggca
agtg
-3’
(Acc
. no.
AL6
0653
3)C
T7-2
36L2
0(A
cc. n
o. A
C07
3666
)EI
DSe
rpin
b1-p
s1N
one
foun
dC
T7-2
36L2
0(A
cc. n
o. A
C07
3666
)M
ark
erP
rim
ers
use
d f
or m
app
ing
Nm
or2
AF2
5226
0 (F
)N
o. 5
36 (s
) 5’-t
gctc
atcg
tcta
tgca
cacc
aaga
-3’
NM
OR
2N
o. 5
37 (a
/s) 5
’-ttt
cttg
tggc
cctt
ggct
caa-
3’(N
M_0
0090
4)P
her1
RP2
3-26
2J21
(c)
No.
534
(s) 5
’-agc
atgc
agtt
attc
ttct
gtgt
ctgg
a-3’
(Acc
. no.
AL4
5040
6)N
o. 5
35 (a
/s) 5
’-gta
gcac
ttgg
gggc
aaag
atca
a-3’
Whi
pF2
0804
6 (F
)R
P23-
217N
23 (c
)R
UV
BL (s
) 5’-t
agtt
ttat
tggc
tcat
aaga
cctt
tgc-
3’W
HIP
(RU
VBL
1)C
T7-2
36L2
0R
UV
BL (a
/s) 5
’- ac
ctgg
cagc
tttg
tgca
atta
agta
at-3
’(N
M_0
2013
5)(A
cc. n
o. A
C07
3666
)D
13M
it11
6.1
RP2
3-41
l21
(c)
No.
702
5’-g
ctct
gatc
caag
agag
gttg
ata-
3’(A
cc. n
o. A
L450
331)
No.
703
5’-a
ccca
gaac
actg
gtga
gtg-
3’R
P23-
414F
19 (c
)D
13M
it13
6R
P23-
262J
21 (c
)N
o. 5
44 5
’-ttt
tatc
tatt
gagt
agat
tcat
ggtg
-3’
(Acc
. no.
AL4
5040
6)N
o. 5
45 5
’-tat
gcct
ggag
gaaa
acag
g-3’
D13
Mit
241
No.
546
5’-c
cacc
cttt
gctg
tttt
gtt-
3’N
o. 5
47 5
’-aca
gttc
caga
catc
cacc
tg-3
’M
it26
5N
o. 5
48 5
’-ggc
gtcc
aagg
tcct
tcc-
3’N
o. 5
49 5
’-aac
gtgt
gcct
acaa
tgca
c-3’
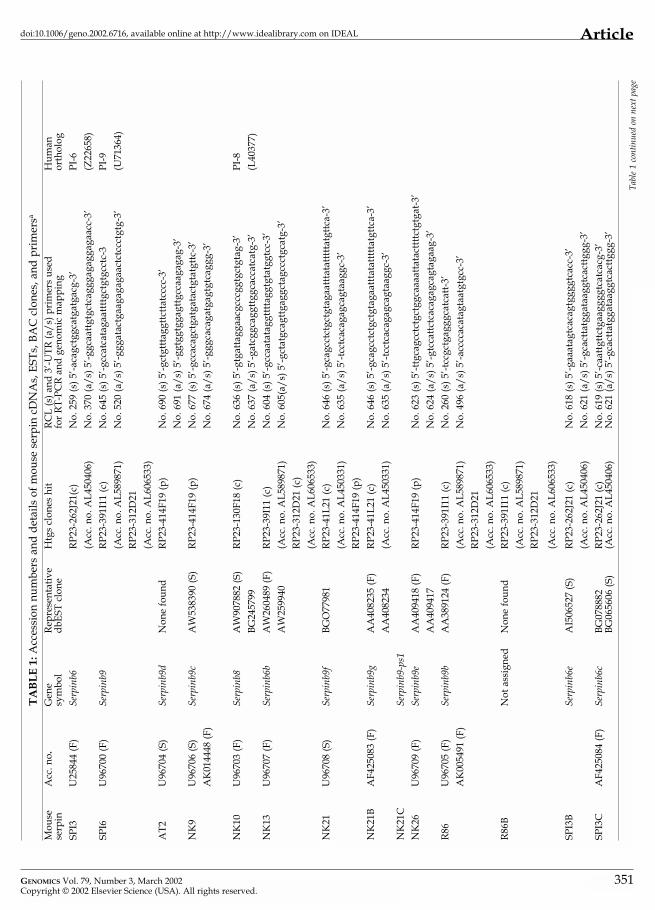
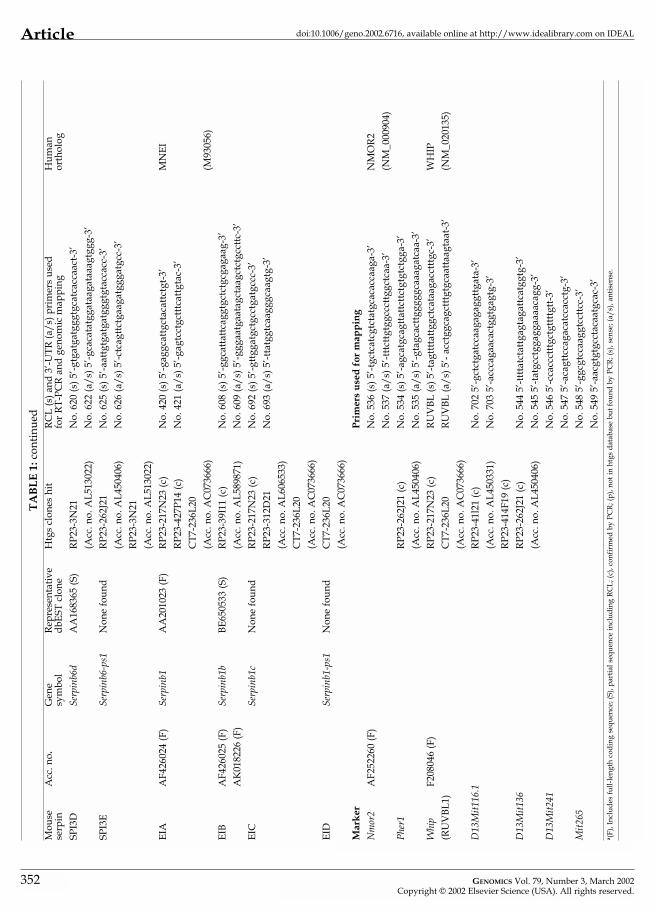
a (F)
, Inc
lude
s fu
ll-le
ngth
cod
ing
sequ
ence
; (S)
, par
tial s
eque
nce
incl
udin
g R
CL;
(c),
conf
irm
ed b
y PC
R; (
p), n
ot in
htg
s da
taba
se b
ut fo
und
by P
CR
; (s)
, sen
se; (
a/s)
, ant
isen
se.
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.
352

Articledoi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
evolved through at least two interchromosomal duplicationsand several intrachromosomal duplications [4].
To understand the biology of ov-serpins and ov-serpingene evolution in more detail, we have begun to identify andexamine mouse ov-serpin genes, and compare their localiza-tion and structures with human genes. Studies so far haveshown that at least three mouse ov-serpin genes are found inregions syntenic with human chromosomes 6 and 18 [10–12].Analysis of two of these genes has demonstrated that both theeight-exon and the seven-exon structures are conserved inthe mouse [10,12]. However, RT-PCR-based screening ofmouse immune tissue using degenerate primers that amplifythe variable RCL has indicated that there are a number ofmouse serpins that have no human counterparts [11]. Here weshow that the mouse ov-serpin repertoire on chromosome 13(syntenic with human chromosome 6) comprises at least 16genes and several pseudogenes, with all those analyzed hav-ing the seven-exon gene structure. RT-PCR analysis indicatesthat most of these mouse genes have restricted expression,with many found in reproductive tissue. These results indi-cate that the mouse ov-serpin gene repertoire is significantlylarger than human, and that it has arisen through intrachro-mosomal gene duplication.
RESULTS
Identification and Characterization of New Mouse Ov-SerpinsWe previously described the mouse ortholog of human SERPINB6 (PI-6 gene), which is designated Spi3 [10]. Spi3has the seven-exon ov-serpin structure and maps to mouse chromosome 13 between Pl1 and Ctla2a in a region syntenicwith human chromosome 6 [10]. We have also described aparalog of human SERPINB9 (PI-9 gene), termed Spi6, whichmaps to the same region as Spi3 [11]. As part of the latterstudy we carried out a survey for other ov-serpins expressedin mouse immune cells and tissue. Using RT-PCR withdegenerate primers designed to amplify the variable RCLsequences from ov-serpins, we cloned and sequenced par-tial cDNA sequences encoding seven new proteins, termedAT2, NK9, NK10, NK13, NK21, NK26, and R86 (GenBankaccession numbers are given in Table 1). On the basis ofsequence similarity between RCLs, it is likely that the NK10gene represents the mouse ortholog of SERPINB8(PI-8 gene), but none of the others have obvious humancounterparts. (Although SERPINB8 is in the human chro-mosome 18 ov-serpin cluster, it is closely related to SERPINB6 and SERPINB9 and shares the same seven-exonstructure.) Two other partial serpin cDNA sequences werecloned from a bone marrow library in the same study(mBM2A and mBM17 [11]). Subsequent investigations haverevealed that the library was contaminated with ratsequences and that mBM2A and mBM17 probably representthe rat counterparts of SPI6 and NK10, respectively (P.I.B.,unpublished data).
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.
To obtain full-length sequence data for the new mouseserpins, we iteratively scanned the nonredundant, mouseEST, and high-throughput genomic sequence (htgs) GenBankdatabases by BLASTN analysis for sequences similar but notidentical to SPI3 and SPI6, and for sequences that matched theRCLs of the new serpins. This process yielded hits on theexpressed sequence tag (EST) or nonredundant databases forNK9, NK10, NK13, NK21, NK26, and R86 (Table 1). No hitswere obtained for AT2 in these databases. Apart from NK9and NK26, these sequences also appeared in the htgs data-base, with all except NK10 falling on mouse chromosome 13genomic clones (the clone containing NK10 has not beenassigned to a specific chromosome, but is likely to belong tochromosome 1, which is syntenic with human 18q21). Weidentified four new sequences in the htgs database thatstrongly resemble SPI3 but have clearly distinct RCLs (Fig. 1).These are present in two overlapping chromosome 13genomic clones (262J21 and 3N21), one of which also containsSPI3 (Table 1). We termed these new SPI3-related serpinsSPI3B, SPI3C, SPI3D, and SPI3E. Finally, inspection of theCelera mouse database revealed sequences on chromosome13 matching the previously published RCL of AT2 [11], andthe first two exons of a gene closely resembling R86, whichwe termed R86B.
Detailed analysis of genomic sequence data from BACRP23-41L21 revealed two copies of NK21 (designated NK21and NK21B) that are 98% identical through both exons andintrons, with only slight differences in the lengths of intronsE and F. Overall, 5 of 377 codons in the putative NK21 andNK21B open reading frames are different, with a key dis-tinction at P5 in the RCL (glutamic acid and lysine, respec-tively). NK21 and NK21B are represented in the EST database,indicating that both are expressed (Table 1). In addition weidentified sequences from a third serpin gene on RP23-41L21that closely resembles NK21 but probably represents apseudogene because of multiple frameshifts and terminationcodons in the putative exons. We termed this gene NK21C.
Using the human SERPINB1 (encoding monocyte/neu-trophil elastase inhibitor (MNEI)) sequence in the BLASTNprogram, we identified, obtained, and sequenced severalrelated mouse EST clones. One clone (dbEST 638727; acc. no.AA201023) was full-length and was sequenced in both direc-tions. On the basis of sequence similarity, particularly theRCL, we identified this serpin as the mouse counterpart ofhuman MNEI, and it was designated EIA (Table 1). SeveralBACs containing portions of mouse chromosome 13 and alsocontaining EIA sequences were found in the htgs database(Table 1), supporting the idea that the counterparts of humanPI-6, PI-9, and MNEI also form a gene cluster in the mouse.Further analysis identified three other genes related to EIA onchromosome 13, termed EIB, EIC, and EID. Almost completecDNA structures containing full-length open reading frameshave been deduced for EIB and EIC (Fig. 1; E.R.-O., unpub-lished data). Information on EID is incomplete, but nucleotidesequence alignments indicate it most closely resembles EIC (> 90% homology; data not shown); however, the absence of
353

Article doi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
an identifiable exon 7 and the presence of multiple stopcodons and frameshifts in other exons suggest it is a pseudo-gene. EIB is also represented in the nonredundant databaseby a RIKEN clone (Table 1), and preliminary results indicateboth EIB and EIC are expressed (E.R.-O., unpublished data).
We obtained and entirely sequenced full-length IMAGE ESTclones encoding NK13, NK21B, NK26, R86, and SPI3C. Usinginformation from EST and genomic databases we deduced thecoding sequences for six of the remaining serpins (NK9, NK10,NK21, AT2, SPI3B, and SPI3D). We were unable to deduce full-length coding sequences for EID, NK21C, or SPI3E because ofa lack of EST data, incomplete htgs data, or assembly errors inthe htgs data. However, it is likely that EID, NK21C, and SPI3Eare pseudogenes because stop codons, deletions, andframeshifts were evident in almost all exons analyzed.
Amino acid sequence alignments and comparisons (Fig. 1)showed that the new proteins are very similar to earlierdescribed mouse ov-serpins such as SPI3 and SPI6. Each hasfeatures typical of serpins in general, and ov-serpins in par-ticular. For example, they have the clearly identifiable, con-served proximal and distal hinge (serpin signature) motifsthat flank the RCL in almost all serpins, indicating that eachis potentially a functional protease inhibitor. By contrast, nonehas a discernible N-terminal signal peptide, and each isshorter than the serpin prototype �1-antitrypsin, both ofwhich are characteristic of ov-serpins. In addition, many carrythe FCAD (or very similar) motif in the distal hinge regionthat is typical of human PI-6, PI-8, and PI-9, and most termi-nate with FSSP (typical of many ov-serpins). Taken together,these observations indicate that all are new members of cladeB of the serpin superfamily.
PhylogenyTo establish the evolutionary relationships between themouse serpins, and their similarity or otherwise to human
354
clade B ov-serpins, we constructed a phylogenic tree usingmouse antithrombin as a common ancestor (Fig. 2). Thisclearly shows that the mouse serpins fall into four distinctgroups comprising SPI3-like proteins (SPI3, NK13, SPI3B–D),SPI6-like proteins (SPI6, R86, NK9, NK21, NK21B, NK26,AT2), EI-like proteins (EIA, EIB, EIC), and NK10. Addinghuman serpins to the tree illustrates that each mouse groupcorresponds to a single human serpin, suggesting that a com-mon ancestor of humans and mice possessed single counter-parts of PI-6, PI-8, PI-9, and MNEI, and that the PI-6, PI-9, andMNEI genes were duplicated in mice after the divergencefrom humans. This is supported by the high homologybetween EIA, EIB, and EIC (88%), NK21 and NK21B (98%),NK21 and NK26 (93%), SPI3 and NK13 (80%), and SPI6 andR86 (80%).
Gene NomenclatureHuman serpin gene nomenclature has recently been revisedto accommodate phylogenic information [1]. In the new sys-tem, a serpin gene is given the descriptor SERPIN, followedby a capital letter indicating which clade [3] the encoded pro-tein belongs to, and finally by a number to act as an identi-fier within the clade. Thus the gene encoding MNEI is SER-PINB1, the gene encoding PI-6 is SERPINB6, and the geneencoding PI-9 is SERPINB9. The letter P is reserved forpseudogenes.
The Mouse Gene Nomenclature Committee (http://www.informatics.jax.org/mgihome/nomen/) recommendsthat for orthologous genes the same root symbol is used in themouse as in human. Major clade designators are representedby a lowercase letter following the root symbol, the genenumber is an arabic numeral, and paralogs not present inhumans are indicated by a lowercase letter following the genenumber. A hyphen followed by “ps” and an arabic numeralis used to indicate pseudogenes. Accordingly, we have usedour phylogenic data to assign symbols for the mouse chro-mosome 13 ov-serpin genes (Table 1). These have beenapproved by the Mouse Gene Nomenclature Committee. Forexample, the previously identified gene Spi3, which encodesthe ortholog to human PI-6, becomes Serpinb6, and the genefor the related serpin NK13 is assigned the symbol Serpinb6b.(Note that for maximum consistency with human nomencla-ture the designator Serpinb6a is not used for the SERPINB6ortholog.) The pseudogene SPI3E becomes Serpinb6-ps1. Thenew gene encoding EIA becomes Serpinb1 because it is orthol-ogous to SERPINB1.
FIG. 2. Relationships between mouse and human ov-serpins. The amino acidsequences of the mouse chromosome 13 serpins and related human serpinswere aligned using ClustalW. The alignment was used to calculate a phylo-genic tree based on the neighbor-joining method using 1000 bootstraps. Mouseantithrombin (AT-III) was defined as the outgroup. Human serpins are boxed.NK10 is not part of the mouse chromosome 13 serpin gene cluster but is closelyrelated and shown for comparison. The scale bar represents the number of sub-stitutions per 100 amino acids. Bootstrap numbers are indicated at the nodes.
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.

GENO
Copy
Articledoi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
TABLE 2: Expression of mouse ov-serpin genes
Tissue Ov-serpin
SPI3 SPI3C SPI3B SPI3E SPI3D NK21 NK26 AT2 NK9 R86 SPI6 NK13 EIA
Skin ++ + + +
Brain ++ +
Eye ++ +
Heart ++ + ++
Lung + ++
Kidney ++ ++ +
Liver ++ + +
Pancreas + +
Stomach ++ +
Lower intestine ++ + ++ +++
Small intestine ++ + +
Muscle + + +
Lymph node ++ + +++
Thymus ++ ++ +
Spleen ++ + + +
Testis ++ + +
Uterus ++ + +
Placenta ++ + + + + + ++ +
Embryo (d13) +
ES cell ++ + +
Tissue Distribution of Mouse Ov-SerpinsWith the exception of AT2, EIC, and R86B, all the new genesare represented in the nonredundant cDNA and/or EST data-bases (Table 1), indicating that they are expressed in mouse tis-sue and are unlikely to represent pseudogenes. To establishthe distribution of the various serpins, we prepared RNA froma range of mouse tissues for use in RT-PCR analysis. Theprimers used in the analysis are shown in Table 1, and in eachcase to ensure specificity they were designed to anneal to theRCL (which is the most variable sequence between serpins)and to the 3�-untranslated region (3�-UTR). Because both theRCL and 3�-UTR are in the last exon, the presence of contam-inating genomic DNA could not be assessed by amplificationacross an intron/exon boundary, so genomic DNA traces wereremoved by DNase I treatment before cDNA synthesis. In eachcase a control reaction was run without addition of RT. Sampleswere also checked and normalized by amplification of mRNAencoding the housekeeping enzyme, GAPDH.
Results of the analysis are shown in Table 2. SPI3 and EIAshowed a broad tissue distribution, which is consistent withpreviously published results [10] and the expression patternsof their human counterparts PI-6 and MNEI [13–15]. SPI6 wasdetected in heart, large intestine, spleen, lymph node, andplacenta. Contrary to previous results [11], SPI6 was notdetected in lung. This discrepancy may be due to lack of
MICS Vol. 79, Number 3, March 2002right © 2002 Elsevier Science (USA). All rights reserved.
hybridization stringency in the original northern blot analy-sis, such that the SPI6 probe cross-reacted with other mouseserpin sequences. Nevertheless, expression of SPI6 in immunetissue and placenta is consistent with the observation that itsclosest human counterpart (PI-9) is present in immune tissueand placenta [16]. Consistent with previous results [11], wedid not detect SPI6 in testis, which contrasts with the presenceof PI-9 in human testis [16]. This result implies that SPI6expression is either induced in testis or not required to fulfilla similar role to PI-9 in this tissue.
Of the other serpins, NK13 showed a wide and overlappingdistribution with SPI3 and EIA, indicating its presence in sim-ilar cells to SPI3 and EIA. By contrast the remaining serpinsshowed very restricted expression, being present only in skin(SPI3B, SPI3C), muscle (SPI3D), embryonic stem (ES) cells(SPI3C), or placenta (R86, NK9, NK21, and/or NKL21, NK26).Given the very high similarity between NK21 and NK21B, wecannot be sure whether one or both genes are expressed in pla-centa. Expression of AT2 and SPI3E was not detected.
Gene StructuresAs outlined above, examination of human ov-serpin geneshas shown two very similar gene organizations (8-exon or 7-exon structures). The only distinction is lack of an exonencoding the interhelical loop in the 6p25 genes encoding
355
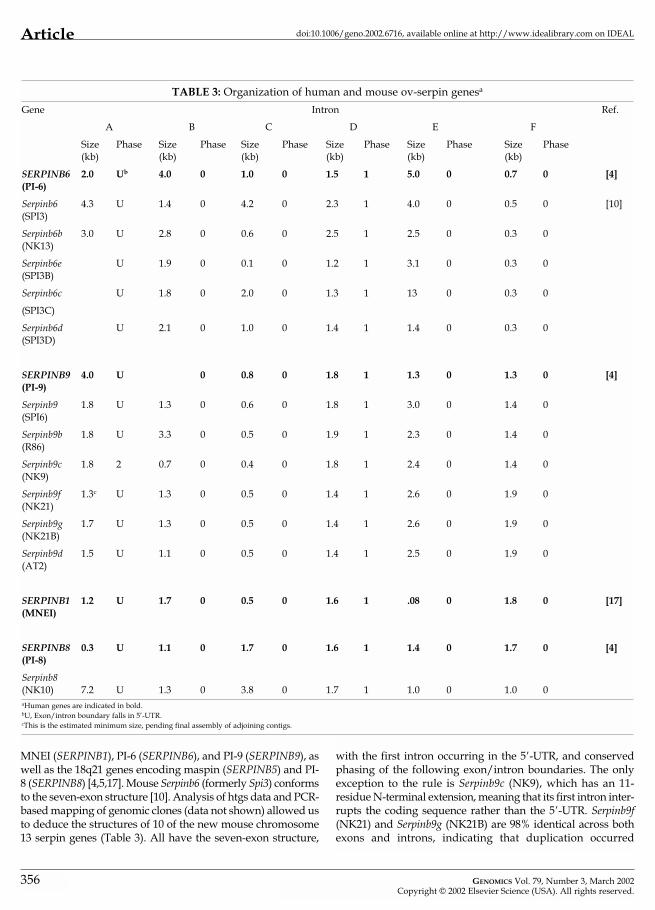
Article doi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
TABLE 3: Organization of human and mouse ov-serpin genesa
Gene Intron Ref.
A B C D E F
Size Phase Size Phase Size Phase Size Phase Size Phase Size Phase(kb) (kb) (kb) (kb) (kb) (kb)
SERPINB6 2.0 Ub 4.0 0 1.0 0 1.5 1 5.0 0 0.7 0 [4](PI-6)
Serpinb6 4.3 U 1.4 0 4.2 0 2.3 1 4.0 0 0.5 0 [10](SPI3)
Serpinb6b 3.0 U 2.8 0 0.6 0 2.5 1 2.5 0 0.3 0(NK13)
Serpinb6e U 1.9 0 0.1 0 1.2 1 3.1 0 0.3 0(SPI3B)
Serpinb6c U 1.8 0 2.0 0 1.3 1 13 0 0.3 0
(SPI3C)
Serpinb6d U 2.1 0 1.0 0 1.4 1 1.4 0 0.3 0(SPI3D)
SERPINB9 4.0 U 0 0.8 0 1.8 1 1.3 0 1.3 0 [4](PI-9)
Serpinb9 1.8 U 1.3 0 0.6 0 1.8 1 3.0 0 1.4 0(SPI6)
Serpinb9b 1.8 U 3.3 0 0.5 0 1.9 1 2.3 0 1.4 0(R86)
Serpinb9c 1.8 2 0.7 0 0.4 0 1.8 1 2.4 0 1.4 0(NK9)
Serpinb9f 1.3c U 1.3 0 0.5 0 1.4 1 2.6 0 1.9 0(NK21)
Serpinb9g 1.7 U 1.3 0 0.5 0 1.4 1 2.6 0 1.9 0(NK21B)
Serpinb9d 1.5 U 1.1 0 0.5 0 1.4 1 2.5 0 1.9 0(AT2)
SERPINB1 1.2 U 1.7 0 0.5 0 1.6 1 .08 0 1.8 0 [17](MNEI)
SERPINB8 0.3 U 1.1 0 1.7 0 1.6 1 1.4 0 1.7 0 [4](PI-8)
Serpinb8(NK10) 7.2 U 1.3 0 3.8 0 1.7 1 1.0 0 1.0 0aHuman genes are indicated in bold.bU, Exon/intron boundary falls in 5’-UTR.cThis is the estimated minimum size, pending final assembly of adjoining contigs.
MNEI (SERPINB1), PI-6 (SERPINB6), and PI-9 (SERPINB9), aswell as the 18q21 genes encoding maspin (SERPINB5) and PI-8 (SERPINB8) [4,5,17]. Mouse Serpinb6 (formerly Spi3) conformsto the seven-exon structure [10]. Analysis of htgs data and PCR-based mapping of genomic clones (data not shown) allowed usto deduce the structures of 10 of the new mouse chromosome13 serpin genes (Table 3). All have the seven-exon structure,
356
with the first intron occurring in the 5�-UTR, and conservedphasing of the following exon/intron boundaries. The onlyexception to the rule is Serpinb9c (NK9), which has an 11-residue N-terminal extension, meaning that its first intron inter-rupts the coding sequence rather than the 5�-UTR. Serpinb9f(NK21) and Serpinb9g (NK21B) are 98% identical across bothexons and introns, indicating that duplication occurred
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.

Articledoi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
recently. Introns B, C, and D are identical in size, while intronE is 32 bp larger in Serpinb9f (NK21B) and intron F in Serpinb9g(NK21) contains a 64-bp repeat. Sequence assembly for intronA of Serpinb9g (NK21) is incomplete, but the data show that itsminimum size is 1.3 kb and it is 98% identical to intron A ofSerpinb9f (NK21B).
Our ability to identify only two exons of R86B (exons 1and 2) implies that the gene either encodes a truncatedproduct or is a nonfunctional remnant. Given its uncertainstatus, a gene symbol was not assigned to R86B.
Chromosome 13 Ov-Serpin Gene Localization and OrderTo localize and order the chromosome 13 serpin genes, weused SPI3 and SPI6 cDNA probes to isolate 30 clones from a129sv mouse genomic PAC library [18]. We also obtained thoseBACs listed in the htgs database containing mouse chromo-some 13 ov-serpin genes (RP23-391I11, 262J21, 41L21, 3N21).Using fingerprint, sequence-tagged site (STS), and genomicsurvey sequence data, we identified overlapping BACs andconstructed a contig comprising 217N23-391I11-414F19-41L21-3N21-262J21. Two BACS that potentially overlapped 391I11and 217N23 were also identified (312D21, 427P14), as was aBAC from a different library (CT7-236L20). The localization ofthe new serpin genes to mouse chromosome 13 was confirmedby FISH using the PACs 600A11, 532I21, 507G8, 578H9, 672N8,and 358P7, as well as the BACs 262J21, 3N21, 414F19, 312D21,and 427P14 (see Table 4 for the genes on these clones). FISHwas carried out using metaphase chromosomes isolated frommice heterozygous for the Del(13)Svea36H deletion, which havelost a 25-Mb region of chromosome 13 syntenic with human6p21–6p25 [9]. All of the clones hybridized to the wild-typechromosome 13, within the region lost in the Del(13)Svea36Hdeletion (data not shown). Thus the serpin genes are clusteredin a region syntenic with human 6p25.
In humans, genes encoding the human receptor(TNFRSF)-interacting serine-threonine kinase 1 (RIPK1;NM_003804) and biphenyl hydrolase-like protein (BPHL;NM_004332) are close to the human 6p25 serpin genes, andtheir mouse counterparts (Bphl and Ripk1) have been mappedto mouse chromosome 13 (J.R., unpublished data). Thereforethe PACs were placed into contigs on the basis of hybridiza-tion with serpin probes and probes for Ripk1 and Bphl (datanot shown). These contig data indicate that in the mouse, asin human, the RIPK1, BPHL, and serpin genes are clustered.
Mapping and recent sequence analysis of the region ofhuman chromosome 6 containing the genes for PI-6, PI-9, andMNEI has identified two other neighboring genes, besidesRIPK1 and BPHL, which encode the Werner helicase inter-acting protein (WHIP; NM_020135) and NAD(P)H mena-dioine oxidoreductase 2 or quinone oxidoreductase (NMOR2;NM_000904; Fig. 3). In addition, our analysis of the sequenceof the mouse clone 262J21, which contains the gene for SPI3,also identified two intronless pseudogenes similar to the calcium-sensing receptor (CaSR) gene, which is a member ofthe pheromone receptor family. One pseudogene appears tobe derived from CaSR (AF110178), while the other appears to
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.
be derived from a differentially spliced variant (AF110179).We termed these pseudogenes Pher1 and Pher2 and designedPCR primers to Pher1 (Table 1).
To confirm synteny between this region of mouse chro-mosome 13 and human chromosome 6p24–p25, we testedwhether Nmor2, Pher1, and Whip are also present in thegenomic clones. As shown in Table 4, Whip, Nmor2, and Pher1are indeed associated with the mouse chromosome 13 ov-serpin genes. We also tested for the markers D13Mit241,D13Mit136, D13Mit116.1, and D13Mit265 thought to be in thisregion of mouse chromosome 13 [19]. D13Mit136 andD13Mit116.1 were present on a number of clones but nonewere positive for D13Mit241 or D13Mit265. Subsequentsequence analysis showed that D13Mit116.1 is within intronA of Serpinb6c (SPI3C).
The PAC and BAC genomic clones were screened for thepresence of ov-serpin and marker genes by PCR using theprimer pairs listed in Table 1. Nine clones were negative forall the probes tested. With the exception of Serpinb8 (NK10),each of the ov-serpin genes was found on one or more of theremaining clones (Table 4). Although it is not yet repre-sented in the htgs database, Serpinb9c (NK9) was found onthe same clone as Serpinb9f (NK21) and Serpinb9e (NK26),demonstrating that these genes are clustered. This PCRanalysis unequivocally placed the serpin and marker genesinto three clusters encoding Whip-Serpinb1-Serpinb1-ps1-Serpinb1c-Serpinb6b-Serpinb9-Serpinb9b-Serpinb1b, Serpinb6d-Serpinb6-ps1-Pher1-Serpinb6e-Serpinb6c-D13Mit136-Serpinb6-Nmor2, and Serpinb9c-Serpinb9d-Serpinb9e-Serpinb9f-D13Mit116.1. Subsequent analysis of htgs sequence datafrom 312D21 placed Serpinb1-ps1 (EID) between Serpinb1(EIA) and Serpinb1c (EIC), and R86B between Serpinb1c (EIC)and Serpinb6b (NK13), while data from 41L21 placedSerpinb9g (NK21B) and Serpinb9-ps1 (NK21C) distal toSerpinb9f (NK21). Celera sequence information suggests thatSerpinb9c (NK9) and Serpinb9d (AT2) are distal to Serpinb1b(EIB), but there is a 0.2-Mb gap between Serpinb9d (AT2)and the Serpinb6 cluster.
From the BAC fingerprint contig and STS content data wededuced linkage and the relative arrangement of the threeclusters using the following overlaps: 391I11 (containsSerpinb1b (EIB)) and 414F19 (contains Serpinb9c (NK9)); 414F19and 41L21 (contains Serpinb9-ps1 (NK21C)); 41L21 and 3N21(contains Serpinb6-ps1 (SPI3E)); and 3N21 and 262J21 (containsSerpinb6 (SPI3)). Thus the organization of the serpin gene clus-ter is cen-Whip-Serpinb1-Serpinb1-ps1-Serpinb1c-R86B-Serpinb6b-Serpinb9-Serpinb9b-Serpinb1b-Serpinb9c-Serpinb9d-Serpinb9e-Ser-pinb9f-D13Mit116.1-Serpinb9g-Serpinb9-ps1-Serpinb6d-Serpinb6-ps1-Pher1-Serpinb6e-Serpinb6c-D13Mit136-Serpinb6-Nmor2-tel(Fig. 3). Analysis of sequence and PCR data (not shown)allowed us to orient most of the genes and pseudogenes withrespect to the flanking markers (Fig. 3). Several other poten-tial pseudogenes or gene remnants are also evident (Fig. 3,diamonds). From sequence information we estimate that theminimum size of the mouse serpin cluster from Whip to Ripk1is 1-Mb, compared to 0.35-Mb in humans.
357

358
Article doi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
TABLE 4: Identification of serpin genes and markers on PACs and BACsa
600 A11 + + N + N N
584 B16 N N N + +
532 I21 N N N + + + + +
661 D24 N N N + + + + + +
598 A11 N N N + + +
507 G8 + + N + N + + N
578 H9 N N + + + + N
672 N8 N N + + + + N + + + +
548 D1 N N N + + + + +
577 C18 N N N
455 G5 N N + + + N
358 P7 N N N + + +
628 P21 N N N + +
339 H23 N N + + + + N
346 O17 N N N + + + +
470 B12 N N N + + + +
442 N20 N N N + + +
339 I24 N N + + + + N
473 C19 N N N + + + +
530 C20 N N N + + + +
642 K6 N N N + +
414 F19 N N + + + + + N
391 I11 N S + + + + N
262 J21 N N N S + + + + +
41 L21 N N + + S S
3 N21 N N N S S
217 N23 + + N + N N N N N N N N N N N N N N N
312 D21 S S S + S S N N N N N N N N N N N N
427 P14 + + N N N N N N N N N N N N N N N N
236L20 S S S S N N N N N N N N N N N N N N N N N N N NaClones 600A11–642K6 are from the RPCI21 library. Clones 414F19–427P14 are from the RP23 BAC library. 236L20 is from the CT7 library. +, Positive by PCR; blank, negative by PCR; S, positive by database sequence information; N, PCR not done. Serpin genes are indicated withoutthe Serpin root symbol.
Whi
p
b1(E
IA)
b1-p
s1 (E
ID)
b1c
(EIC
)
b9-p
s2 (R
86B)
b6b
(NK
13)
b9(S
PI6)
b9b
(R86
)
b1b
(EIB
)
b9c
(NK
9)
b9d
(AT2
)
b9e
(NK
26)
b9f(
NK
21)
Mit1
16
b9g
(NK
21B)
b9-p
s1(N
K21
C)
b6d
(SPI
3D)
b6-p
s1 (S
PI3E
)
Phe
r1
b6e
(SPI
3B)
b6c
(SPI
3C)
Mit1
36
b6(S
PI3)
PAC/BAC
DISCUSSION
There are 13 ov-serpins in humans, and they are implicated inprocesses such as tumor progression, inflammation, and theregulation of apoptosis. Three human ov-serpin genes are clus-tered at 6p25, with the remainder at 18q21, and all have
essentially the same structures. In this study we have identi-fied and characterized 15 serpin genes on mouse chromosome13, in a region syntenic to human 6p25. Synteny is illustratedby the presence of genes encoding RIPK1, BPHL, WHIP, andNMOR2 near the ov-serpin clusters in both humans and mice.Although the gene order is conserved in mouse and human, itis likely that the orientation of each locus on the chromosome
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.

Articledoi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
FIG. 3. Comparison of the human and mouse ov-serpin gene clusters. The relationship of the htgs RP23 BACs to the mouse cluster is indicated, as deduced fromgenomic survey sequence data and PCR analysis. The Celera scaffold designations for this part of mouse chromosome 13 are shown; information on clones usedto sequence the region can be found at http://www.celera.com. Arrows indicate gene orientation; closed circles indicate named pseudogenes; closed diamondsindicate unnamed pseudogenes or remnants; closed square indicates truncated gene. Accession numbers for the serpin sequences, markers, and htgs RP23 BACsare listed in Table 1. The clone 236L20 is from the CT7 library. Note that the map is not to scale. Sequence information indicates that the mouse cluster spans ~ 1-Mb, whereas the human cluster spans ~ 0.35-Mb. The structure and predicted orientation of the human 6p25 locus serpin is as shown in the NCBI mapviewer (http://www.ncbi.nlm.nih.gov/cgi-bin/Entrez/maps.cgi?org=hum&chr=6).
may differ between species. Thus the gene for BPHL is telom-eric in the mouse, but is centromeric in humans according tothe draft genomic sequence. At present the orientation of themouse locus is supported by analysis of the Del(13)Svea36H dele-tion mice [9], but the orientation of the human locus has yet tobe confirmed by joining of the adjacent sequence contigs.Hence the formal possibility remains that the human locus isin the same orientation as the mouse.
Although there has been a significant expansion in thenumber of serpin genes in the mouse cluster compared tohuman, gene structures have been conserved in terms of exonnumber and intron phasing. At least seven potential serpinpseudogenes or gene remnants have been observed at mousechromosome 13 (EID, NK21C, SPI3E, and four unnamedsequences) and one at human 6p25 (PI8L1 [4]), but untilsequencing of the region is completed we cannot rule out theexistence of additional genes or pseudogenes. The possibilityof additional genes is illustrated by one PAC (577C18) thatwas identified as carrying serpin sequences by probing forSPI3/SPI6 and is positive for Bphl, yet is negative for Nmor2and all of the serpin genes described here. The several otherPACs isolated in the same way that are negative for all mark-ers may contain chromosome 1 serpin sequences (from aregion syntenic to human chromosome 18q21).
Phylogenic analysis indicates that the mouse serpinsencoded by genes on chromosome 13 fall into three groups,
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.
each with similarity to one of the three 6p25 human serpins.Hence mice have at least three MNEI-like serpins, seven PI-9-like serpins, and five PI-6-like serpins. The simplest explana-tion for the evolution of the mouse chromosome 13 cluster isthat a common ancestor of human and mice possessed a cor-responding primordial cluster of three ov-serpin genes thatwere independently duplicated after the human and mouselineages diverged. This would explain the conservation of genestructure. An alternative view that the common ancestor had16 or more serpin genes in the primordial cluster, and thatmost were lost in humans after divergence, is not supportedby human genomic sequence data, which show no evidence ofgene remnants. One or more duplications of the entire pri-mordial cluster is also unlikely to have occurred in micebecause there are different numbers of serpins in each of thethree subgroups. Our map of the chromosome 13 genes is alsoinconsistent with such a “duplication en-masse” model, whichwould predict conserved gene order within each subcluster(for example, PI-6-like, then PI-9-like, then MNEI-like).
What are the likely functions of the mouse chromosome13 serpins? On the basis of sequence similarity, functionalstudies and expression patterns, we have earlier suggestedthat SPI3 and SPI6 are counterparts of the human 6p25 serpins PI-6 and PI-9, respectively [10,11]. The work reportedhere supports this view by extending the expression patterndata, and showing that SPI6 has an identical gene structure
359

Article doi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
to PI-9. It is also clear from sequence comparisons that EIA isthe counterpart of MNEI (Fig. 1). The 6p25 human serpins arewidely expressed proteinase inhibitors thought to have intracellular cytoprotective roles [reviewed in 20]. Known tar-gets of PI-6, PI-9, and MNEI are the leukocyte granule pro-teinases cathepsin G, granzyme B, and neutrophil elastase,respectively [14,21,22]. Although they may have additionaltargets and sites of action [15,23], the main physiological roleof the 6p25 serpins is probably the protection of proteinase-producing leukocytes, bystander cells, and lining cells frommisdirected proteolysis. For example, PI-9 is produced bycytotoxic, dendritic, endothelial, and epithelial cells [16,24,25],and has been shown to protect cells from apoptosis inducedby misdirected granzyme B [21]. It is likely that in the mouseSPI6 has a similar role to PI-9, and that SPI3 and EIA mirrorPI-6 and MNEI, respectively. On the basis of our earlier stud-ies identifying AT2, R86, NK9, NK13, NK21 and NK26 inmouse immune tissue [11], it is possible that these serpinsalso protect cells and tissues against mouse granule or secre-tory proteinases. As there are more granule proteinases in themouse than in human [26], it is reasonable to suggest that thelarger number of mouse ov-serpins reflects the need to regu-late a larger number of proteinases.
Inspection of the sequences of the chromosome 13 mouseserpins shows conservation of the proximal and distal hingemotifs flanking the RCL, suggesting that all are functionalproteinase inhibitors. From RCL comparisons alone it is notpossible to predict what proteinases are potential targets ofthese serpins. Although the overall homology between theserpins is high, their RCLs show substantial sequence varia-tion, suggesting that they inhibit different proteinases andare unlikely to be functionally redundant. The exceptions areNK21 and NK21B, which are essentially identical in theirRCLs and probably inhibit the same proteinase(s). At presentwe do not know whether NK21 and NK21B are differentiallyexpressed, but it is possible they inhibit the same proteinasein different contexts, for example at different sites or inresponse to different stimuli. Our RT-PCR analysis also showsthat apart from NK13, the new serpins have a very restrictedexpression pattern in normal adult tissue, suggesting thattheir targets are also restricted. However it remains to be seenwhether expression of one or more of these genes also occursduring development or is induced by specific signals, forexample in response to immune challenge or other stress.
An interesting question raised by our analysis of mousechromosome 13 is whether a similar expansion has occurredin the region of mouse chromosome 1 syntenic with humanchromosome 18q21 (which contains the larger number ofhuman ov-serpin genes). Expansion of mouse serpin genenumber compared to human is not limited to the chromo-some 13 ov-serpin subgroup, as genes corresponding to thesingle-copy human plasma serpins �1-antitrypsin and �1-antichymotrypsin have undergone duplication on mousechromosome 12 to form clusters of 5 and 10 genes, respec-tively [27,28]. Members of each of these plasma serpin geneclusters show considerable divergence in their RCLs but
360
otherwise are very closely related, suggesting that they havealso evolved to regulate different proteinases [29]. It is clearfrom database searches that there are indeed other mouse ov-serpins that have no obvious human counterparts, and areprobably not encoded in the cluster we have analyzed. Atpresent we do not know whether they represent genes onchromosome 13, chromosome 1 or elsewhere. Our prelimi-nary analysis of the region on chromosome 1 encoding NK10(the mouse homolog of PI-8) suggests that it has not beenduplicated, and there is no evidence of additional NK10-likegenes in the EST or htgs databases. The possibility thereforeremains open that the chromosome 1 ov-serpin gene clusterhas not undergone significant expansion.
In closing, it should be noted that our work illustrates theimportance of using complementary techniques to character-ize clusters of closely-related genes, and the potential diffi-culties in relying on genomic nucleotide sequence informationalone. At the time of writing neither the public nor privatemouse genome sequence databases contain the entire, cor-rectly constructed and ordered chromosome 13 serpin clus-ter. Although the assembly phase is complete, the Celera data-base has a 0.2-Mb gap in the middle of the serpin cluster, andis missing or failed to predict EID, NK21, NK21B, NK21C,NK26, SPI3B, SPI3D and SPI3E. By contrast, the public data-base sequences (htgs) remain mainly unassembled, and lackAT2, NK9 and NK26. In addition, the presence of introns inthe 5’-untranslated regions of these serpin genes complicatesidentification of the first splice donor and the upstream pro-moter region using gene prediction programs. Comparison ofEST and genomic sequences is required to reliably delineateand validate these and other intron/exon boundaries, there-fore it is important that large-scale or genome-wide projectscharacterizing full-length ESTs are continued, and that therange of source tissues used in library generation is expanded.
MATERIALS AND METHODS
Mouse EST and genomic clones. The mouse PAC library RPCI21 was con-structed using female 129/SvevTACfBr mouse spleen genomic DNA [18]. Itwas obtained in the form of DNA on membranes from the HGMP ResourceCentre, Hinxton, UK. ESTs were obtained from the IMAGE collection (ATCC).Clones from the RPCI23 C57BL6/J BAC library were obtained from the HGMPResource Centre (Hinxton, UK) or from PAC/BAC Resources at the MurdochInstitute (Melbourne, Australia). Clone DNA was prepared by standard procedures [30].
Database mining and sequence analysis. Genbank accession numbers for theclones identified or used in this study are shown in Table 1. Mouse counter-parts of human PI-6 (SPI3) and PI-9 (SPI6) have been reported [10,11]. Usingthe human MNEI amino acid sequence in the tBLASTn program(http://www.ncbi.nlm.nih.gov/BLAST/ [31]), we found a clone in the GEN-BANK mouse EST database that we identified as the mouse counterpart ofhuman MNEI, on the basis of very high homology in the RCL. We designatedthis serpin EIA. A full-length RIKEN clone was released and identified later inthe GENBANK non-redundant database.
In all human and mouse ov-serpin genes characterized so far, the RCLand 3’-UTR are encoded by the last exon [4,5]. Our previous study identified100 bp fragments of new, expressed mouse serpin genes on the basis of RCLsequence comparisons [11]. Each of these 100 bp fragments comprised part of
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.

Articledoi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
the coding sequence and 3’-UTRs were not identified. As a first step towardsobtaining full-length sequences of the new ov-serpins, the BLASTn program(http://www.ncbi.nlm.nih.gov/BLAST/ [31]) was used to search the GEN-BANK mouse EST database with the previously identified RCL fragments [11]for EST clones containing both RCL and 3’-UTR sequences. Of the many clonesidentified, one was apparently full length with essentially complete sequencein the database (NK9), and three (EIB, NK21 and NK26) had inserts of suffi-cient length (> 1.2 kb) to encode a full-length serpin. ESTs encoding EIB, NK21and NK26 were obtained from the ATCC IMAGE collection and sequenced onboth strands. The full-length cDNA sequences of NK21 and NK26 were alignedto SPI6 and SPI3 using ClustalW (http://www.clustalw.genome.ad.jp/), anda consensus nucleotide sequence for the mouse ov-serpin coding region wasderived. (Subsequent studies showed that the NK21 EST actually represents ahighly homologous gene, designated NK21B.)
The consensus sequence was used with BLASTn to identify clones in theGENBANK high throughput genomic sequence (htgs) database that containmouse ov-serpin sequences. These htgs sequences were then compared byBLASTn to the known RCL sequences [11]. Several BACs carrying EIA, NK10,NK13, NK21, R86, SPI3 and SPI6 were identified. In addition, six previouslyunknown serpin genes (SPI3B-E, NK21B, NK21C) were identified by BLASTnanalysis using the consensus sequence. No htgs sequences corresponding toAT2, NK9 or NK26 were identified. Inspection of data in the Celera mouse data-base (http://www.celera.com) revealed genomic sequences containing NK9and AT2. The gene prediction program GENSCAN (http://genes.mit.edu/GENSCAN.html [32]) was then used to analyze the htgs data to predict the full-length open reading frames of NK10, NK13, R86, AT2, and the five novelserpins.
Intron size and phasing for the ov-serpin genes was predicted by compar-ing the experimentally determined (SPI3, SPI6, NK21B) and predicted (NK10,NK13, R86, SPI3B-E, AT2) cDNA sequences to the corresponding genomic (htgs)sequences using the program EST_GENOME (http://zeno.well.ox.ac.uk/git-bin/est_genome [33]). Because ov-serpin genes have an intron in the 5�-UTR, it was difficult to locate exon I in htgs data de novo using BLASTn andGENSCAN, and hence to identify the beginning of the relevant genes.Accordingly, exon II of each predicted gene was used with BLASTn to identifycorresponding clones in the mouse EST database that contain sequencesupstream of exon II. This approach identified EST clones for NK10, NK13, R86and SPI3C, which were obtained from the relevant EST collection and completelysequenced. In each case the sequencing data yielded the exon I sequence, andconfirmed the predicted open reading frames. The full-length cDNA sequencewas then used to identify the corresponding exon I in the htgs data, and com-plete the determination of intron size and phasing using EST_GENOME. Becauseno ESTs containing exon II of SPI3B or SPI3D were identified, the beginning ofthese genes and size of intron A could not be identified. Due to incomplete orincorrect assembly of htgs data, the full nucleotide sequence and structure of theEIA, EID and SPI3E genes could not be determined.
Phylogenic trees based on the amino acid sequences of the new serpinswere constructed as described [3].
DNA sequencing. Plasmids containing EIA, EIB, NK10, NK13, NK21B, NK26,R86, and SPI3C ESTs were sequenced on both strands using an “oligonucleotidewalk” strategy. Primers were purchased from Sigma and used with purifiedplasmid template DNA in the Big-Dye system (Amersham Pharmacia Biotech).Samples were run on an Applied Biosciences 373 DNA sequencer.
RT-PCR analysis. Extraction of RNA from mouse (C57BL6) tissue sampleswas performed using the RNAzol B RNA isolation system (Tel-Test). Toremove any contaminating DNA, 5 �g of RNA was incubated with 2 units ofRQ1 RNase-free DNAse (Promega) in Taq DNA polymerase buffer and 3.75mM MgCl2 at room temperature for 30 minutes. To inactivate the DNase, EDTAwas added to a final concentration of 1 mM, and the samples were phenol-extracted. Approximately 2.5 �g of DNA-free RNA was reverse transcribedwith 1 �g of oligo dT (Amersham Pharmacia Biotech) in the presence of 20 unitsRNase inhibitor and 200 units M-MLV reverse transcriptase (Promega) in atotal volume of 25 �l. Parallel reactions lacking reverse transcriptase were per-formed to ensure the absence of genomic DNA. The reactions were incubatedat 42�C for 2 hours. To assess the efficiency of cDNA synthesis and absence ofcontaminating genomic DNA, 1 �l from each reaction was used in a 30 cyclePCR (10 mM Tris, pH 9, 2.5 mM MgCl2 , 50 mM KCl, 0.1% Triton X-100, 200 �MdNTP) with 10 pmol primers specific for GAPDH (5�-gaccccttcattgacctcaac-3�
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.
and 5�-gatgaccttgcccacagcctt-3�). Approximately 1 �l of cDNA was used ineach PCR used to amplify a mouse serpin sequence with 20 pmol of the appro-priate primer combination listed in Table 1. 30 cycle PCR reactions were per-formed in 50 �l containing 1.5 or 2.5 mM MgCl2 and the highest possible anneal-ing temperature (usually 60–65�C) empirically determined for each oligonu-cleotide combination on a genomic DNA template. Amplified samples wereassessed by electrophoresis of approximately 20 �l on 2% ethidium bromideagarose gels.
PAC mapping. Genomic PAC clones containing ov-serpin genes were identi-fied in the RPCI21 mouse genomic PAC library by screening with cDNA probescorresponding to SPI3, SPI6 or human PI8. Plasmids containing these cDNAswere cleaved with the appropriate restriction endonuclease(s) to release theinsert, which was gel purified using a BIO 101 kit and the concentration deter-mined by gel electrophoresis. DNA (50 ng) was labeled using a Rediprime IIkit (Amersham Pharmacea Biotech) with 0.05 mCi/1.85 MBq of [�-32P]dCTPand incubated at 37�C for 30 minutes. PAC library membranes were pre-hybridized for 1 hour at 65�C in hybridization solution (10% Dextran SO4, 10�Denharts, 50 mM Tris HCl, 6� SSC, 1% Sarkosyl). The probe was denaturedat 95�C for 5 minutes, added to pre-warmed hybridization mix and hybridizedfor 16 hours at 65�C. The filters were then washed in 2� SSC, 0.1% SDS at 65�Cfor 20 minutes, followed by two 20-minute washes in 1� SSC, 0.1% SDS.Positive clones were identified and ordered from the HGMP Resource Centre,Hinxton, UK.
Contig building. DNA from the positive clones was prepared by standardprocedures [30]. Approximately 200 ng of clone DNA was incubated with 10U EcoRI at 37�C for 2 hours, and the sample was split and resolved on two0.8% ethidium bromide agarose gels. DNA was transferred to membranes byalkali blotting techniques as recommended by the manufacturer (AmershamHybond N+) and hybridized to SPI3, SPI6 or human PI8 probes. From theseresults it was possible to identify the serpin genes contained in individualclones and to build contigs. Two IMAGE clones, IMAGE 739159 and IMAGE592125, corresponding to the Ripk1 and Bphl genes were obtained from theHGMP Resource Centre, Hinxton, UK, digested and used as probes asdescribed above on the clones to aid contig construction and further refinethe map of the region.
The RP23 BAC contig was built using FPC [34] with a combination of STScontent (as described below, and M.C., R.M., Chris Sellick, and P.D., unpub-lished data) and restriction endonuclease digestion data (Genome SequenceCentre; BC Cancer Research Centre; http://www.bcgsc.bc.ca/projects/mouse_mapping/). Confirmation of BAC order was performed by BLASTnanalysis using the following genomic survey sequences, GenBank acc. nos.AQ983039, AQ929164, AZ110066, AZ110072, AZ250279, AZ250271, AZ060024,AZ561201, AZ561203, AAZ027750, AQ970854, AAZ088339, and AZ060148.Overlap of BACs 41L21 and 414F19 was established using the STS marker M-05614 (http://www-genome.wi.mit.edu [35]).FISH. Ear explants from Del(13)Svea36H mice were washed in 100% ethanol andthen twice in phosphate-buffered saline (PBS). Each ear was chopped into 4 partsand transferred into a 25 cm3 flask containing 5 ml Dulbecco’s medium contain-ing 10% fetal calf serum, 4 mM L-glutamine, and 50 �g/ml gentamycin. Theexplants were grown at 37�C in 5% CO2 until fibroblasts had reached about 80%confluence, then Colcemid (Gibco) was added to a final concentration of 75ng/ml. After 2 hours the cells were washed with PBS and incubated for 5 min-utes with 0.5 ml of 0.25% trypsin, 0.02% EDTA (Sigma). We added 3 ml of 75mM KCl to the detached cells, which were transferred to a centrifuge tube. After20 minutes at 37�C, 10 drops of pre-fixative (3:1 methanol: acetic acid) were mixedin well. The cells were collected by centrifugation at 1000 rpm in a Jouan C312centrifuge, resuspended in fixative then collected again by centrifugation. Thisstep was repeated before finally resuspending the pellet in 1 ml of fixative. TheFISH procedure was as described [36] except mouse cot1 DNA was used.
ACKNOWLEDGMENTSWe thank Pannos Ioannou (Murdoch Institute, Melbourne) for providing BACs, RuthArkell (MRC UK Mouse Genome Centre & Mammalian Genetics Unit, Harwell) forDel(13)Svea36H mice, Chris Sellick and Anne Southwall (MRC, Harwell) for techni-cal assistance, Owen McCann and Simon Gregory (Sanger Centre) for assistance withFPC, and Nicos Tripodis (Division of Medical Molecular Genetics, GKT MedicalSchool) for fibroblast preparation, and James Irving (Monash University) for discus-
361

Article doi:10.1006/geno.2002.6716, available online at http://www.idealibrary.com on IDEAL
sions. Parts of these data were generated through use of the Celera Discovery Systemand Celera’s associated databases. This work was supported by the National Health andMedical Research Council of Australia, the Wellcome Trust (grant 060759/Z), theMedical Research Council (UK), the National Institutes of Health (grant HL66548),and Monash University.
RECEIVED FOR PUBLICATION OCTOBER 23; ACCEPTED DECEMBER 27, 2001.
REFERENCES1. Silverman, G. A., et al. (2001). The serpins are an expanding superfamily of structurally
similar but functionally diverse proteins. J. Biol. Chem. 276: 33293–33296.2. Huntington, J. A., Read, R. A., and Carrell, R. W. (2000). Structure of a serpin–protease
complex shows inhibition by deformation. Nature 407: 923–926.3. Irving, J. A., Pike, R. N., Lesk, A. M., and Whisstock, J. C. (2000). Phylogeny of the ser-
pin superfamily: implications of patterns of amino acid conservation for structure andfunction. Genome Res. 10: 1845–1864.
4. Scott, F. L., et al. (1999). Human ovalbumin serpin evolution: phylogenic analysis, geneorganization, and identification of new PI-8-related genes suggest that two interchro-mosomal and several intrachromosomal duplications generated the gene clusters at18q21–q23 and 6p25. Genomics 62: 490–499.
5. Ragg, H., Lokot, T., Kamp, P.-B., Atchley, W. R., and Dress, A. (2001). Vertebrate ser-pins:construction of a conflict-free phylogeny by combining exon–intron and diagnos-tic site analyses. Mol. Biol. Evol. 18: 577–584.
6. Bartuski, A. J., Kamachi, Y., Schick, C., Overhauser, J., and Silverman, G. A. (1997).Cytoplasmic antiproteinase 2 (PI8) and bomapin (PI10) map to the serpin cluster at18q21.3. Genomics 43: 321–328.
7. Chatterjee, A., et al. (2001). Mapping the sites of putative tumor suppressor genes at 6p25and 6p21.3 in cervical carcinoma: occurrence of allelic deletions in precancerous lesions.Cancer Res. 61: 2119–2123.
8. Davies, A. F., et al. (1999). Delineation of two distinct 6p deletion syndromes. Hum.Genet. 104: 64–72.
9. Arkell, R. M., et al. (2001). Genetic, physical and phenotypic characterization of theDel(13)Svea36H mouse. Mamm. Genome 12: 687–694.
10. Sun, J., Rose, J. B., and Bird, P. (1995). Gene structure, chromosomal localization andexpression of the murine homologue of human proteinase inhibitor 6 (PI6) suggestsdivergence of PI6 from the ovalbumin serpins. J. Biol. Chem. 270: 16089–16096.
11. Sun, J., et al. (1997). A new family of 10 murine ovalbumin serpins includes twohomologs of proteinase inhibitor 8 and two homologs of the granzyme B inhibitor (pro-teinase inhibitor 9). J. Biol. Chem. 272: 15434–15441.
12. Dougherty, K. M., et al. (1999). The plasminogen activator inhibitor-2 gene is notrequired for normal murine development or survival. Proc. Natl. Acad. Sci. USA 96:686–691.
13. Coughlin, P., Sun, J., Cerruti, L., Salem, H. H., and Bird, P. (1993). Cloning and molec-ular characterization of a human intracellular proteinase inhibitor. Proc. Natl. Acad..Sci.USA 90: 9417–9421.
14. Remold-O’Donnell, E., Chin, J., and Alberts, M. (1992). Sequence and molecular char-acterisation of human monocyte/neutrophil elastase inhibitor. Proc. Natl. Acad. Sci.USA 89: 5635–5639.
362
15. Scott, F., et al. (1998). Proteinase inhibitor 6 (PI-6) expression in human skin: inductionof PI-6 and a PI-6/proteinase complex during keratinocyte differentiation. Exp. CellRes. 245: 263–271.
16. Sun, J., et al. (1996). A cytosolic granzyme B inhibitor related to the viral apoptotic reg-ulator cytokine response modifier A is present in cytotoxic lymphocytes. J. Biol. Chem.271: 27802–27809.
17. Zeng, W., Silverman, G. A., and Remold-O’Donnell, E. (1998). Structure and sequenceof human M/NEI (monocyte/neutrophil elastase inhibitor), an ov-serpin family gene.Gene 213: 179–187.
18. Osoegawa, K., et al. (2000). Bacterial artificial chromosome libraries for mouse sequenc-ing and functional analysis. Genome Res. 10: 116–128.
19. Hong, H. -K., Lass, J. H., and Chakravarti, A. (1999). Pleiotropic skeletal and ocular phe-notypes of the mouse mutation congenital hydrocephalus (ch/Mf1) arise from a wingedhelix/forkhead transcription factor gene. Hum. Mol. Genet. 8: 625–637.
20. Bird, P. I. (1999). Regulation of pro-apoptotic leucocyte granule serine proteinases byintracellular serpins. Immunol.Cell Biol. 77: 47–57.
21. Bird, C. H., et al. (1998). Selective regulation of apoptosis: the cytotoxic lymphocyte ser-pin proteinase inhibitor 9 protects against granzyme B-mediated apoptosis withoutperturbing the Fas cell death pathway. Mol. Cell. Biol. 18: 6387–6398.
22. Scott, F. L., et al. (1999). The intracellular serpin proteinase inhibitor 6 (PI-6) is expressedin monocytes and granulocytes and is a potent inhibitor of the azurophilic granule pro-teinase, cathepsin G. Blood 93: 2089–2097.
23. Kato, K., et al. (2001). Serine proteinase inhibitor 3 and murinoglobulin I are potentinhibitors of neuropsin in adult mouse brain. J. Biol. Chem. 276: 14562–14571.
24. Buzza, M. S., et al. (2001). The granzyme B inhibitor, PI-9, is present in endothelial andmesothelial cells, suggesting it protects bystander cells during immune responses. Cell.Immunol. 210: 21–29.
25. Bladergroen, B. A., et al. (2001). The granzyme B inhibitor, protease inhibitor 9, is mainlyexpressed by dendritic cells and at immune-privileged sites. J. Immunol. 166: 3218–3225.
26. Smyth, M. J., and Trapani, J. A. (1995). Granzymes: exogenous proteinases that inducetarget cell apoptosis. Immunol. Today 16: 202–206.
27. Borriello, F., and Krauter, K. S. (1991). Multiple murine alpha 1-protease inhibitor genesshow unusual evolutionary divergence. Proc. Natl. Acad. Sci. USA 88: 9417–9421.
28. Inglis, J. D., and Hill, R. E. (1991). The murine Spi-2 proteinase inhibitor locus: a multi-gene family with a hypervariable reactive site domain. EMBO J. 10: 255–261.
29. Paterson, T., and Moore, S. (1996). The expression and characterization of five recom-binant murine � 1-protease inhibitor proteins. Biochem. Biophys. Res. Commun. 219: 64–69.
30. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular Cloning. A LaboratoryManual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
31. Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic localalignment search tool. J. Mol. Biol. 215: 403–410.
32. Burge, C., and Karlin, S. (1997). Prediction of complete gene structures in humangenomic DNA. J. Mol. Biol. 268: 78–94.
33. Mott, R. (1997). EST_GENOME: a program to align spliced DNA sequences to unsplicedDNA. Comput. Appl. Biosci. 13: 477–478.
34. Soderland, C., Humphrey, S., Dunham, A., and French, L. (2000). Contigs built with fin-gerprints, markers and FPC V4.7. Genome Res. 10: 1772–1787.
35. Nusbaum, C., et al. (1999). A YAC-based physical map of the mouse genome. Nat. Genet.22: 388–393.
36. Davies, A. F., et al. (1995). Evidence of a locus for orofacial clefting on human chromo-some 6p24 and STS map of the region. Hum. Mol. Genet. 4: 121–128.
GENOMICS Vol. 79, Number 3, March 2002Copyright © 2002 Elsevier Science (USA). All rights reserved.




















