
Characterization of polymorphic markers in the von Willebrand factor gene and pseudogene
Upload
independentCategory
view
0download
0
1 23
Conservation Genetics Resources ISSN 1877-7252 Conservation Genet ResourDOI 10.1007/s12686-012-9846-8
Development of polymorphic microsatellitemarkers of the common toad,Duttaphrynus melanostictus useful forgenetic studies
S. Jegath Janani, Richa Sharma,Karthikeyan Vasudevan, Sushil K. Dutta& Ramesh K. Aggarwal

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media Dordrecht. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

TECHNICAL NOTE
Development of polymorphic microsatellite markersof the common toad, Duttaphrynus melanostictus usefulfor genetic studies
S. Jegath Janani • Richa Sharma •
Karthikeyan Vasudevan • Sushil K. Dutta •
Ramesh K. Aggarwal
Received: 2 October 2012 / Accepted: 15 December 2012
� Springer Science+Business Media Dordrecht 2012
Abstract Asian common toad, Duttaphrynus melanos-
tictus, is a widespread habitat generalist species of Indian-
subcontinent and South-East Asia. Here we report, 14 new
microsatellite markers (SSRs) for this toad, developed from
a SSR-enriched genomic library, and validated using 44
unrelated samples of Asian toad and *20 samples belong-
ing to four congeners. Nine SSRs were highly polymor-
phic; amplified 6–22 alleles/marker with Ho and PIC
ranging from 0.477 to 0.955 and 0.518 to 0.918, respec-
tively, while one was found to be monomorphic. Of the
remaining four SSRs, two (DMccmb02, DMccmb17)
amplified only two apparently fixed alleles in all samples,
while the other two (DMccmb22, DMccmb29) amplified
1–4alleles/sample; these four SSRs probably represent
duplicated loci. Interestingly, all the 14 SSRs showed
cross-species transferability to varying extent, with four
SSRs showing 100 % transferability. It is hoped that these
SSRs will greatly facilitate population genetic studies on
the Asian toad species across their geographic distribution.
Keywords Duttaphrynus melanostictus � Asian common
toad � Microsatellite markers � Conservation/population
genetics � Cross-species transferability
The Asian common toad Duttaphrynus melanostictus
belongs to the family ‘‘Bufonidae’’, called true toads, com-
prising of more than 500 species that are wide spread around
the world except Australia, Madagascar and The Antarctic.
The Asian common toad is native to Indian subcontinent and
South-East Asia, and is identified as species complex con-
sisting of more than one species. Traits such as large body
size, use of terrestrial adult niche, presence of parotid glands
make the toad more adaptable to broad range of habitats
(Van Bocxlaer et al. 2010). The species is commonly seen in
human dominated and disturbed areas and breeds through-
out the year using all types of water bodies. The dispersal
and life history traits greatly influence the geographical scale
of the genetic structure of the continuously distributed
population within the species’ distribution range. Though
most of the amphibian populations are considered as meta-
populations, toads like Bufo americanus show complete
absence of isolation by distance within its expanding range
(Leblois et al. 2000). The Asian toad offers to be a model for
understanding the population biology and biogeography of
anurans adapted to diverse landscapes. Moreover, it appears
to be an ideal candidate bio-indicator species for environ-
mental health and ecosystem function (pending validation),
by virtue of being habitat generalist. However, any such
studies will require understanding the genetic structure and
dynamics of its extant populations. To achieve this, genomic
DNA variation based molecular/genetic markers hold great
promise, especially the microsatellite markers that target
hypervariable repetitive simple sequence repeats (SSRs) in
the genome. These are most informative and efficient for the
population level studies, but no such markers have been
developed to-date for the Asian toad. Here, we describe the
first set of 14 SSR markers for the common toad D. mela-
nostictus that would facilitate population genetic studies on
this important bufonid species.
S. J. Janani � R. Sharma � R. K. Aggarwal (&)
Centre for Cellular and Molecular Biology (CSIR-CCMB),
Uppal Road, Tarnaka, Hyderabad 500007,
Andhra Pradesh, India
e-mail: [email protected]
K. Vasudevan
Wildlife Institute of India, P.O. Box 18, Dehradun 248 001, India
S. K. Dutta
P.G. Department of Zoology, North Orissa University,
Baripada 757003, India
123
Conservation Genet Resour
DOI 10.1007/s12686-012-9846-8
Author's personal copy
Ta
ble
1C
har
acte
rist
ics
of
the
new
mic
rosa
tell
ite
mar
ker
sd
evel
op
edfo
rD
utt
ap
hry
nu
sm
ela
no
stic
tus
Mar
ker
Rep
eat
mo
tif
Pri
mer
seq
uen
ceM
ost
freq
uen
t
alle
le(s
)
Ta
(�C
)
nS
ize
ran
ge
#
Na
Ho
He
PIC
HW
E
(Pv
alu
e)
Gen
Ban
k
acce
ssio
n
no
.
DM
ccm
b0
4(T
G) 2
6F
*C
AG
GT
TA
GG
GA
GC
GA
AT
TT
CA
GT
TT
16
7,
18
4,
19
76
24
31
60
–2
01
21
0.9
32
0.9
05
0.8
98
0.6
95
KC
28
71
73
RC
CC
GC
AC
TA
TC
TG
TC
CC
TT
CC
T
DM
ccm
b0
8(A
C) 1
2F
*C
TC
AG
GA
CC
AG
GC
GA
TT
TT
CA
GA
T2
00
,2
09
52
38
19
2–
23
01
70
.86
80
.90
80
.89
80
.06
4K
C2
87
17
4
RC
AT
CA
TA
AA
AA
AC
CC
CC
CA
TA
AA
AC
T
DM
ccm
b0
9(G
T) 2
0F
*G
CA
AA
AA
AT
GT
CT
AA
GT
CC
TA
AG
GG
TG
A1
67
,1
94
62
44
15
6–
22
01
90
.95
50
.92
40
.91
80
.84
4K
C2
87
17
5
RT
CA
CC
CT
GC
AC
TA
TC
TG
TT
CC
TT
CT
G
DM
ccm
b1
2(T
CT
A) 1
7F
*T
GC
TG
CT
TC
AT
TC
AT
TA
AA
CT
TC
AC
AT
A2
93
,3
31
,3
41
58
42
27
4–
42
92
20
.90
90
.91
40
.90
80
.32
2K
C2
87
17
6
RC
TT
CC
TT
CT
GG
GC
TC
CT
AA
CA
TC
TG
DM
ccm
b1
3(T
C) 1
6F
*G
CC
TG
TC
AG
TG
TC
CC
TT
TA
AT
TC
T1
60
,1
68
58
44
14
3–
18
21
30
.68
20
.87
70
.86
50
.00
2*
KC
28
71
77
RA
TT
GC
AT
GT
TT
TT
CC
CT
AT
AT
CT
GA
C
DM
ccm
b2
4(T
C) 3
3F
*A
GG
GT
GC
AG
CA
AC
AA
AG
TC
AT
AA
TG
13
15
54
31
26
–1
39
60
.47
70
.54
70
.51
80
.08
2K
C2
87
17
8
RT
TC
CA
AA
GA
CT
AA
TA
GA
TC
CA
TG
TA
CC
A
DM
ccm
b2
7(G
T) 4
,(G
T) 1
7F
*G
CT
GC
TG
CT
CA
CA
AA
TA
AA
AG
GG
A2
65
,2
69
62
44
25
2–
27
91
10
.75
00
.76
20
.73
70
.35
2K
C2
87
17
9
RG
GT
TA
GA
GA
AG
TC
GC
TG
GT
GA
CA
GA
DM
ccm
b2
8(C
A) 1
2F
*G
CC
CA
GA
AA
CA
GT
CA
CT
AA
GC
AC
CC
13
86
24
31
37
–1
57
11
0.6
05
0.8
40
0.8
25
0.0
00
*K
C2
87
18
0
RT
TT
GG
CT
GA
CC
CT
CT
AA
TG
TG
TA
TG
G
DM
ccm
b3
2(A
G) 7
…(A
G) 7
….(
AG
) 14
F*
TT
GC
CT
GG
AA
CC
TA
AT
AT
AA
TC
AT
GT
G2
07
,2
22
58
44
17
7–
26
32
00
.84
10
.88
50
.87
50
.09
1K
C2
87
18
1
RC
AA
AT
GC
TT
AG
GA
TG
TT
GT
GA
AC
TT
TT
A
DM
ccm
b1
6(G
T) 1
5F
*G
TT
TT
CT
GC
AA
TC
GA
GC
CC
CA
AT
AT
24
55
54
32
45
1M
on
om
orp
hic
KC
28
71
82
RT
GC
AA
TT
GC
TC
CA
CG
GT
TT
TA
TG
DM
ccm
b1
7#
(AG
) 25
F*
CA
CC
GT
TA
TA
AA
GG
CA
CT
GG
CG
TC
A1
55
,2
10
62
44
15
5,
21
02
ND
;p
uta
tiv
ed
up
lica
ted
loci
KC
28
71
83
RC
TG
AG
AC
CC
CT
GT
GC
AC
TG
AA
TG
AG
DM
ccm
b0
2#
(AC
) 24
F*
AG
TC
CC
AA
AA
CA
TC
CC
CC
CA
TA
12
6,
13
26
24
01
26
,1
32
2N
D;
pu
tati
ve
du
pli
cate
dlo
ciK
C2
87
18
4
RC
CA
CA
CC
TC
CC
TT
CC
TC
CA
CT
C
DM
ccm
b2
2}
(CA
) 27
F*
CC
TG
GT
CG
TG
AA
GG
GG
TT
AA
CA
CA
C1
29
,1
31
,1
35
60
44
11
7–
14
82
1N
D;
pu
tati
ve
du
pli
cate
dlo
ciK
C2
87
18
5
RG
CT
GA
AA
CG
TT
GC
CA
TC
GG
AA
TA
DM
ccm
b2
9}
(GA
) 9…
(AG
) 9F
*A
CC
GC
CA
GT
TG
CT
AG
AA
TG
AA
GG
17
4,
18
0,
23
16
24
41
74
–2
33
15
ND
;p
uta
tiv
ed
up
lica
ted
loci
KC
28
71
86
RC
CA
CT
GA
CA
GC
TA
GA
AC
GA
GG
AG
A
F*
forw
ard
pri
mer
wit
hF
AM
tag
atth
e50
end
,R
rev
erse
pri
mer
,T
aan
nea
lin
gte
mp
erat
ure
in�C
,n
nu
mb
ero
fsa
mp
les,
Na
nu
mb
ero
fal
lele
so
bse
rved
,H
o,
ob
serv
edH
eter
ozy
go
sity
,H
e
exp
ecte
dH
eter
ozy
go
sity
,P
ICp
oly
mo
rph
ism
info
rmat
ion
con
ten
t,#
,F
ixed
du
pli
cate
dlo
cus;
},p
oly
mo
rph
icd
up
lica
ted
locu
s;N
DN
ot
Det
erm
ined
.A
llel
eat
trib
ute
sw
ere
calc
ula
ted
usi
ng
Gen
AlE
x6
.4(h
ttp
://w
ww
.an
u.e
du
.au
/Bo
Zo
/Gen
AlE
x/i
nd
ex.p
hp
)an
dC
ER
VU
Sv
er3
.1(h
ttp
://w
ww
.fiel
dg
enet
ics.
com
).H
WE
,H
ard
y–
Wei
nb
erg
exac
tte
stfo
rh
eter
ozy
go
ted
efici
tca
lcu
late
d
usi
ng
Gen
epo
p(h
ttp
://g
enep
op
.cu
rtin
.ed
u.a
u/g
enep
op
_o
p1
.htm
l),
and
*p
rese
nce
of
nu
llal
lele
sas
ind
icat
edb
yM
ICR
OC
HE
CK
ER
(htt
p:/
/ww
w.m
icro
chec
ker
.hu
ll.a
c.u
k/)
Conservation Genet Resour
123
Author's personal copy

In this study, to identify and develop Asian toad specific
microsatellite markers, total genomic DNA was isolated
from the thigh muscle tissue using the standard phenol:
chloroform extraction method (Sambrook et al. 1989),
which was then used to construct a small-insert SSR-enri-
ched genomic library following the method described earlier
by Aggarwal et al. (2004). Briefly, the process involved:
digestion of genomic DNA with frequent 4-base cutters
(HaeIII and RsaI restriction enzymes), elution of agarose-
gel fractionated fragments of 0.3–1.0 kb in size, ligation of
the eluted fragments to Mlu-adaptors (Edwards et al. 1996),
SSR-enrichment of the adaptor-ligated genomic fragments
by Solution hybridization with biotinylated oligonucleotide
probes [(GA)15 (CA)15 (AGA)10 (CAA)10] and solid-phase
separation using strepdavidin coated magnetic beads (Dyn-
abeads, DYNAL), followed by cloning of the selected DNA
fragments into pTZ57R/T vector (Fermentas, USA) and
E. coli DH5a host cells to get the SSR-enriched genomic
library. Plasmids were isolated and cloned inserts were
amplified/sequenced using M13 universal primers on an
automated ABI 3730xl DNA Analyzer following manufac-
turer’s instructions (Applied Biosystems Inc.). Sequencing
of *150 clones, revealed 96 SSR-positive sequences, of
which [30 were found to be part of large tandem mini
satellite repeats and thus not considered for marker devel-
opment. Thirty-six of the remaining SSR-positive sequences
that had a minimum repeat core (simple or compound) of
18 bp were identified manually and used for primer design
using the Primer3 tool (http://biotools.umassmed.edu/bio
apps/primer3_www.cgi). Each forward primer was 50-tag-
ged with FAM fluorophore. The standardization of PCR
amplification conditions (annealing temperature and Mg??
ion concentration) were attempted for all the 36 primer pairs,
and the 14 working primer-pairs (that gave good, clean and
robust amplicons) were further tested for their potential
utility as genetic markers using a panel of 44 unrelated
samples of D. melanostictus collected from south India. All
PCR amplifications were performed in PTC-200 thermal-
cycler (MJ Research Inc.) in 15 ll reaction volumes that
comprised 5 ng template DNA, 2 pM of each primer, 1U
AmpliTaqGold DNA polymerase (Applied Biosystems,
USA), 1 9 PCR buffer, 100 lM of each dNTP and 1.5 mM
MgCl2. PCR conditions comprised: initial denaturation at
94 �C for 10 min, followed by 35 cycles of 3-step amplifi-
cation—(denaturation at 94 �C for 30 s, annealing at primer
specific temperature for 30 s and extension at 72 �C for
45 s), and a final extension at 72 �C for 15 min. The
SSR amplicons were resolved by fragment analysis on
ABI-3730xl DNA Analyzer with LIZ500 as internal size
standard, and were precisely sized using the software
GeneMapperTM ver3.7 (Applied Biosystems, USA). The 14
primer pairs/validated SSRs were also tested for their
cross-species transferability using four of the congeners
Duttaphrynus scaber, Duttaphrynus stomaticus, Duttaphry-
nus himalayanus and a new D. melanostictus cryptic species
(our unpublished data).
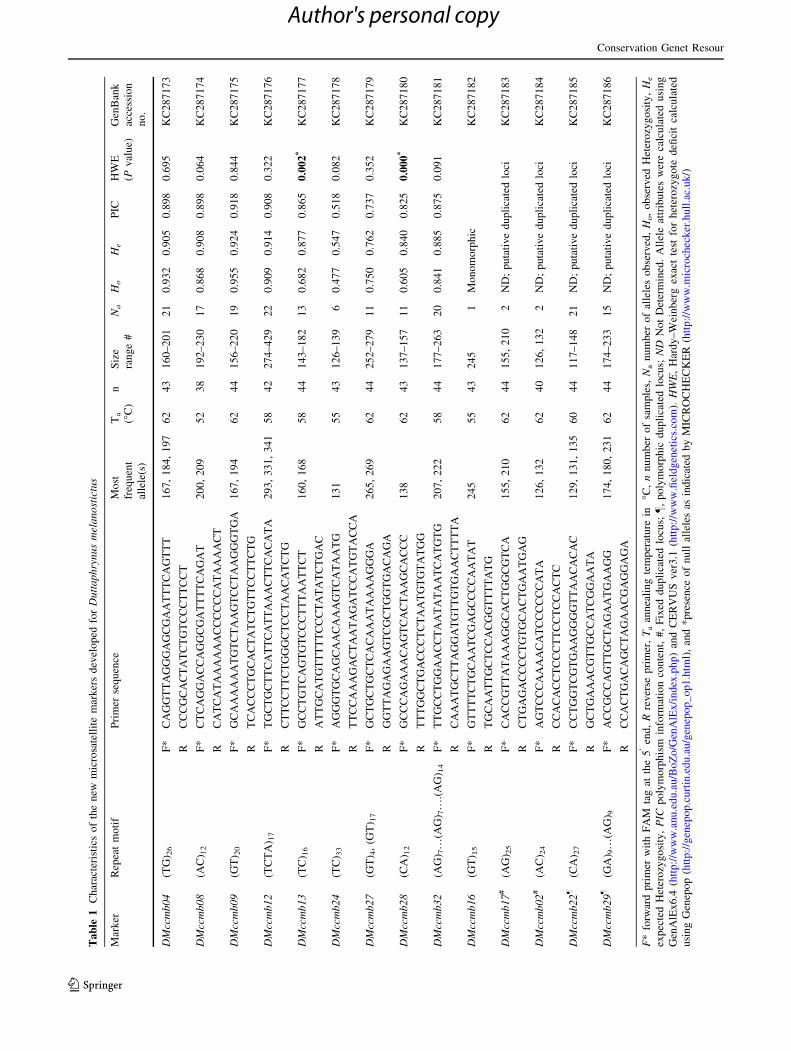
Characteristics of the new SSR markers, such as, marker
designation, repeat motifs, primer sequences, allele attri-
butes (number of alleles, size range of alleles, observed and
expected heterozygosity, polymorphism information con-
tent) and GenBank accessions are summarized in Table 1,
while their cross-species transferability status is shown in
Table 2. Nine of the 14 SSRs were found to be highly
polymorphic in the 44 Asian toad individuals, while one
Table 2 Cross-species transferability of the D. melanostictus microsatellites
Locus D. scaber D. stomaticus D. himalayanus D. melanostictus sp2
n/status/Na Allele range N/status/Na Allele range N/status/Na Allele range N/status/Na Allele range
DMccmb04 NA 4/P/6 150–193 NA 12/P/12 158–209
DMccmb08 NA NA NA 10/P/10 191–248
DMccmb09 NA NA NA 10/P/7 152–169
DMccmb12 NA NA 3/P/6 259–295 10/P/7 200–252
DMccmb13 3/P/3 128–158 2/P/2 178, 211 NA 11/P/13 136–171
DMccmb24 4/P/5 120–136 4/P/5 124–134 3/M/1 124, 130 12/P/9 124–175
DMccmb27 4/P/6 252–279 4/P/3 266–274 3/M/1 249 12/P/9 259–289
DMccmb28 4/P/5 121–155 4/P/4 120–132 3/P/3 121–128 11/P/8 120–141
DMccmb32 4/P/6 185–201 NA 3/P/2 187, 191 9/P/7 176–239
DMccmb16 3/P/4 174–183 3/P/3 162–218 3/M/2 171, 250 9/P/8 181–245
DMccmb17 NA 4/M/1 188 NA 10/P/13 150–200
DMccmb02 4/P/3 100–136 4/P/4 130–146 3/M/2 126, 133 9/M/2 126, 133
DMccmb22 NA NA NA 10/P/10 122–151
DMccmb29 4/P/4 180–211 4/P/11 172–233 3/P/3 182–202 12/P/17 152–231
n number of samples, Na number of alleles observed, NA no amplification, p polymorphic
Conservation Genet Resour
123
Author's personal copy

(DMccmb16) was monomorphic in the tested samples. For
the nine polymorphic markers, the PIC values ranged from
0.518 to 0.918 amplifying 6–22 alleles/markers with
observed heterozygosity ranging from 0.477 to 0.955. None
of these markers showed linkage disequilibrium, but two
(DMccmb13, DMccmb28) deviated from Hardy–Weinberg
equilibrium showing heterozygote deficit, which is indicated
to be due to null alleles (Table 1). Of the remaining four
markers, two markers, DMccmb22 and DMccmb29,
amplified 1–4 alleles per individual and were highly poly-
morphic in the test samples, whereas the other two markers
DMccmb02 and DMccmb17 amplified only two alleles that
were almost fixed in the test population. In our opinion,
these markers probably represent duplicated loci as have
been observed in other animal/plant systems (Hendre et al.
2008). Notably, all the new markers also showed cross
species transferability to varying degree (overall 60 % for
three congeners and 100 % to closely related cryptic
species).
Thus, in this study, we have developed and validated 14
new SSR markers of Asian toad, which are expected to be
greatly helpful in undertaking population genetics studies
on this important generalist species, as well as, other
related bufonids.
Acknowledgments The authors thank the Department of Biotech-
nology, India for the financial support under the CCMB-WII-NOU
collaborative project (DBT, Govt of India, BT/PR8354/NDB/51/141/
2006) on anuran DNA barcoding; and the Council for Scientific and
Industrial Research for the lab facilities; Phanindranath and Justin for
the technical support extended for the study.
References
Aggarwal RK, Velavan T, Udaykumar D, Hendre P, Kartik S,
Choudhury B, Singh L (2004) Development and characterization
of novel microsatellite markers from the olive ridley sea turtle
(Lepidochelys olivacea). Mol Ecol Notes 4:77–79
Edwards KJ, Barker JHA, Daly A, Jones C, Karp A (1996)
Microsatellite libraries enriched for several microsatellite
sequences in plants. Biotechniques 20:758–760
Hendre PS, Phanindranath R, Annapurna V, Lalremruata A, Aggarwal
RK (2008) Development of new genomic microsatellite markers
from robusta coffee (Coffea canephora Pierre ex A. Froehner)showing broad cross-species transferability and utility in genetic
studies. BMC Plant Biol 8:51
Leblois R, Rousset F, Tikel D, Moritz C, Estoup A (2000) Absence of
evidence for isolation by distance in an expanding cane toad
(Bufo marinus) population: an individual-based analysis of
microsatellite genotypes. Mol Ecol 9:1905–1909
Sambrook J, Fritsch E, Maniatis T (1989) Molecular cloning: a
laboratory manual, 2nd edn. Cold Spring Harbor Press, New York
Van Bocxlaer I, Loader SP, Roelants K, Biju SD, Menegon M,
Bossuyt F (2010) Gradual adaptation toward a range-expansion
phenotype initiated the global radiation of toads. Science 327:
679–682
Conservation Genet Resour
123
Author's personal copy
Copyright © 2022 FDOKUMEN




















