
Enzyme-Amplified Array Sensing of Proteins in Solution and in Biofluids
Upload
independentCategory
view
1download
0
www.fems-microbiology.org
FEMS Microbiology Letters 240 (2004) 87–97
Evaluation of randomly amplified polymorphic DNA and pulsedfield gel electrophoresis techniques for molecular typing of
Dermatophilus congolensis
Jose Larrasa a, Alfredo Garcıa-Sanchez b, Nicholas C. Ambrose c,1,Alberto Parra a, Juan M. Alonso d, Joaquın M. Rey d, Miguel Hermoso-de-Mendoza d,
Javier Hermoso-de-Mendoza d,*
a Departamento de Microbiologıa, Laboratorios Larrasa S.L., Corredera Hernando de Soto 13-A, Jerez de los Caballeros, 06380 Badajoz, Spainb Bacteriology Division, Moredun Research Institute, Pentlands Science Park, Bush Loan, Midlothian, EH26 0PZ, Scotland, UKc Centre for Tropical Veterinary Medicine, Royal (Dick) School of Veterinary Studies, University of Edinburgh, Scotland, UK
d Catedra de Patologıa Infecciosa, Facultad de Veterinaria, Universidad de Extremadura,
Campus U. EX., Av. de la Universidad s/n, 10071Caceres, Spain
Received 31 May 2004; received in revised form 14 September 2004
First published online 28 September 2004
Edited by M.R. Soria
Abstract
This study aimed to evaluate molecular typing methods useful for standardization of strains in experimental work on dermato-
philosis. Fifty Dermatophilus congolensis isolates, collected from sheep, cattle, horse and a deer, were analyzed by randomly ampli-
fied polymorphic DNA (RAPD) method using twenty-one different primers, and the results were compared with those obtained by
typing with a pulsed field gel electrophoresis (PFGE) method using the restriction digest enzyme Sse8387I. The typeability, repro-
ducibility and discriminatory power of RAPD and Sse8387I-PFGE typing were calculated. Both typing methods were highly repro-
ducible. Of the two techniques, Sse8387I-PFGE was the least discriminating (Dice Index (DI), 0.663) and could not distinguish
between epidemiologically related isolates, whereas RAPD showed an excellent discriminatory power (DI, 0.7694–0.9722). Overall,
the degree of correlation between RAPD and PFGE typing was significantly high (r, 0.8822). We conclude that the DNA profiles
generated by either RAPD or PFGE can be used to differentiate epidemiologically unrelated isolates. The results of this study
strongly suggest that at least two independent primers are used for RAPD typing in order to improve its discriminatory power,
and that PFGE is used for confirmation of RAPD results.
� 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Dermatophilus; RAPD; PFGE; Molecular typing
0378-1097/$22.00 � 2004 Federation of European Microbiological Societies
doi:10.1016/j.femsle.2004.09.016
* Corresponding author. Tel.: +34 927 25 71 29; fax: +34 927 25 71
10.
E-mail address: [email protected] (J. Hermoso-de-Mendoza).1 Present address: Scottish Executive Environment and Rural
Affairs Department (SEERAD), Pentland House, 47 Robbs Loan,
Edinburgh EH14 1TY, Scotland, UK.
1. Introduction
Dermatophilosis is an exudative dermatitis that af-
fects a wide range of animal species, including humans.
The aetiological agent of dermatophilosis is the branch-
ing filamentous actinomycete bacterium Dermatophilus
congolensis. It has a significant economic impact on
. Published by Elsevier B.V. All rights reserved.

88 J. Larrasa et al. / FEMS Microbiology Letters 240 (2004) 87–97
cattle production in West and Central Africa and in the
Caribbean islands, and on sheep production in Aus-
tralia. Acute, subacute, chronic and also latent forms
have been described in many of the affected species
and between breeds in cattle and sheep [1], affecting in
a generalised form or located in diverse body areas: dor-sal region, feet areas, external genital areas, mammary
skin and head area [2–4]. In the tropics, the disease in
cattle is associated with tick-mediated immunosuppres-
sion with extent and nature of lesions varying consider-
ably between breeds. In Australia, dermatophilosis
manifests as lumpy wool in fine-wooled sheep breeds
as Merinos. In horses the disease takes several forms
the most typical being rain scald. In all species the com-monly observed features are proliferation of the epider-
mis to produce thickened crusts or scabs formed from
palisaded layers of keratinocytes, serous exudates and
infiltrates of neutrophils [5].
To date, strategies to control dermatophilosis have
included immunoprophylaxis and treatment with anti-
biotics. However, the available literature indicates that
these strategies have not been successful. In additionto the aforementioned species and breed variation in
clinical symptoms one reason for failure of vaccina-
tion and therapy may be the antigenic, enzymatic, vir-
ulence or pathogenic factor differences between strains
of D. congolensis described by several authors [5–13].
Evidence also points to genetic-based differences be-
tween strains, for example, in a recent study Larrasa
et al. [14], developed a simple RAPD method for gen-otyping field isolates of D. congolensis, where strain
differences were evident. Typing D. congolensis isolates
is a priority for dermatophilosis research. It is re-
quired in order to standardise the experimental use
of different strains for vaccine, diagnostic or antimi-
crobial susceptibility trials between different research
groups and for inter-laboratory exchange of tech-
niques. Standardization of strain characterisation im-prove the chances of success in the field application
of results [15].
According to recent guidelines, the typing efficiency
of RAPD can be improved by the parallel use of sev-
eral primers, and to be certain that two isolates with
the same fingerprint are genetically linked results
should be compared with ‘‘gold standard’’ molecular
typing methods such as pulsed field gel electrophoresis(PFGE) [16]. Comparisons are based in criteria like
typeability, or the ability of the typing method to pro-
duce unmistaken results [17]; reproducibility or ability
to produce the same results in different moments,
places, using different devices or performed by differ-
ent operators [16,18,19]; discriminatory power or the
probability that two randomly selected isolates from
a population of epidemiologically unrelated ones willbe recognized as actually different by the technique
[20].
In addition, special attention should be paid to the
selection of an appropriate set of isolates as studies to
evaluate the typeability and discriminatory power of a
typing system should be carried out with collections of
epidemiologically unrelated isolates [17].
In this study, we evaluated the typeability, reproduc-ibility and discriminatory power of RAPD using 21 dif-
ferent primers, and PFGE using the restriction enzyme
Sse8387I as methods to distinguish between 50 D. cong-
olensis isolates from different geographic origins and
host species. The efficacy of the two techniques to type
D. congolensis strains was then compared.
2. Materials and methods
2.1. Bacterial strains and culture conditions
The fifty D. congolensis isolates used in this study
came from collections held at the University of Edin-
burgh and the University of Extremadura, except two
horse isolates which came from the Institut fur Mikrobi-ologie und Tierseuchen, Tierarztlichen Hochschule
Hannover, Germany, and three type isolates from Brit-
ish and American type culture collections. The isolates,
their host species and geographic origin are listed in
Table 1.
The isolates were cultured on brain heart infusion
agar and in 30 ml of 2% (w/v) brain heart infusion/
1.7% neutralised soya peptone (Oxoid Ltd, Basingstoke,UK), using glass McCartney bottles. Cultures were incu-
bated at 37 �C in 5% CO2 for 24–48 h to produce a dense
growth of hyphae and filaments with a clear supernatant
in broth.
The broth cultures were centrifuged at 4.000g for 15
min at 4 �C using Corning tubes. The pellet was resus-
pended in 10 ml of TE to wash the cells again by
centrifugation.
2.2. RAPD
Template DNA was extracted using a fast non-phe-
nolic method as previously described [14]. A total of
21 arbitrary primers were selected from a group of
33 (see Table 2), on the basis of their ability to pro-
duce polymorphism, visible as a number of clearly de-fined electrophoretic bands in trial experiments using a
generic RAPD protocol [14]. Sequential optimization
of each component of the reaction and the amplifica-
tion conditions were carried out following a previously
described protocol [14]. The final reaction composition
and amplification conditions are shown in Tables 3
and 4.
Randomly amplified products (10 ll) were size sepa-rated on 2% agarose gels by electrophoresis and were
visualized with ethidium bromide.

Table 1
Dermatophilus congolensis isolates
Isolate Host Origin
OA1 (NCTC 7915) Sheep UK
OPOA Sheep Spain
OPOB Sheep Spain
OPOE Sheep Spain
OPOF Sheep Spain
OMR Sheep Spain
ORL Sheep Spain
OT8 Sheep Spain
OT10 Sheep Spain
OTS1 Sheep Spain
OG1 Sheep Spain
OA3 (16793) Sheep UK
OA4 (S3) Sheep UK
OA5 (A1) Sheep UK
OA6 (S598) Sheep UK
OA7 (A5N) Sheep UK
OA8 (NM) Sheep Australia
OA9 (S164) Sheep UK
BE3 (ATCC 14637) Cattle Zimbabwe
B556 Cattle Spain
BTB1 Cattle Spain
BG45 (IMTV M6445) Cattle Guadeloupe
BF13 (5.11A) Cattle Nigeria
BE5 (16.3) Cattle Nigeria
BE7 (Gh�88) Cattle Ghana
BE11 (Gh�89) Cattle Ghana
BE6 (Z8) Cattle Zimbabwe
BE2 (ANU931) Cattle Antigua
BE12 (ANU932) Cattle Antigua
BE4 (SB�86) Cattle Antigua
BE10 (SC�86) Cattle Antigua
BE1 Cattle Santa Lucıa
BE8 (Gh�93B) Cattle Ghana
BE9 (Gh�93C) Cattle Ghana
BC87 (IMVT) Cattle France
BH6 (IMVT) Cattle France
EA2 (NCTC 5175) Horse UK
EDC (43037) Horse UK
EEQ Horse Spain
EWAN Horse Spain
EATI Horse Spain
EISL Horse Spain
EH1 (17174) Horse UK
EH2 (1161) Horse UK
EH3 (1274) Horse UK
EA11 (3.86) Horse UK
ED201 Horse Germany
EK628 Horse Germany
EA10 (Dec84) Horse UK
CH8 Deer UK
Table 2
RAPD: Oligonucleotide primers used
Primer Target DNA Sequence 50 ! 3 0
IS 1* IS 6110 [78] GGCTGAGGTCTCAGATCAG (19 b)
IS 2* IS 6110 [78] ACCCCATCCTTTCCAAGAAC (20 b)
MPTR 1 MPRT [78] GCCGGTGTTGGTGTG (15 b)
Pntb 1* PGRS [79] CCGTTGCCGTACAGCTG (17 b)
Pntb 2* PGRS [79] CCTAGCCGAACCCTTTG (17 b)
DAF 7 Arbitrary [78] GCGCACGGTC (10 b)
RAPD 5 Arbitrary [78] AATGCAGCTGGCTCG (15 b)
OPA 2 Arbitrary [80] TGCCGAGCTG (10 b)
OPC 20 Arbitrary [80] ACTTCGCCAC (10 b)
OPM2 Arbitrary TGCCGAGCTG (10 b)
AP1 Arbitrary GCCACGCCAGTCCAGCTGTG (20 b)
D1 Arbitrary CTGGTCGACT (10 b)
D2* Arbitrary AAGTCCTGCC (10 b)
D3* Arbitrary GTCGGTGTCA (10 b)
D4* Arbitrary CTGGTCACGT (10 b)
D5* Arbitrary AGGTCACCGA (10 b)
D6 Arbitrary TCCAGGACAG (10 b)
D7 Arbitrary GCACCTGGAA (10 b)
D8 Arbitrary GGTAGCCACT (10 b)
D9 Arbitrary CCAGCGTTGA (10 b)
D10 Arbitrary GTTACCCGGA (10 b)
DP1* Arbitrary CCTACCTCAC (10 b)
DP2 Arbitrary TTCCGATGCC (10 b)
DP3 Arbitrary GAAGCAGGGA (10 b)
DP4* Arbitrary CTCCACCTTC (10 b)
DP5 Arbitrary AGAAGCAGCC (10 b)
DP6 Arbitrary TGGTGCGAGT (10 b)
DP7* Arbitrary GCCAGAGTGA (10 b)
DP8* Arbitrary ATTGGCTCGG (10 b)
DP9 Arbitrary ACCTTGCCAG (10 b)
DP10 Arbitrary GCCAAGCAGT (10 b)
DL1 Arbitrary AGAGGTGGCAACAGGTGCGA(20 b)
DL2 Arbitrary CTTCACCTCGTTGTCCACCC (20 b)
Discarded ones are marked with an asterisk.
J. Larrasa et al. / FEMS Microbiology Letters 240 (2004) 87–97 89
2.3. PFGE-sample preparation
Initially, in order to standardize a procedure for
PFGE analysis we assessed four different methods to
prepare intact chromosomal DNA embedded in low-
melting-point agarose. The methods were those de-
scribed by Levy-Frebault et al. [21], Udo and Grubb
[22], Matushek et al. [23], and Cornillot et al. [24]. Agar-ose plugs containing a range of DNA concentrations
were tested in order to determine the optimum.
The efficacy of the DNA preparation methods was
evaluated using the following quality criteria [21]: (a)
lack of ‘‘sandy’’ appearance in the blocks, indicative of
enough enzymatic treatment; (b) lack of a DNA ‘‘track’’
after electrophoresis, a sign of efficacy of the enzymes
used to break cells.
Although not wholly successful in its unmodified
form, only Levy-Frebault�s protocol had some apparentefficacy on D. congolensis cells. Therefore, it was modi-
fied by additional homogenization and washing steps,
adjustments of cell concentration, increasing the lyso-
zyme concentration, increasing treatment time with liso-
zyme and proteinase K and increasing the number of
wash steps after enzymatic treatment with simultaneous
reduction of the washing solution concentration. The
modified protocol proved successful for sample prepara-tion, and is described in the results below.
2.4. PFGE-restriction enzyme digestion
Twenty-two restriction enzymes (Amersham Bio-
sciences) (see Table 5) and a variety of electrophoresis
parameters were examined in order to determine the
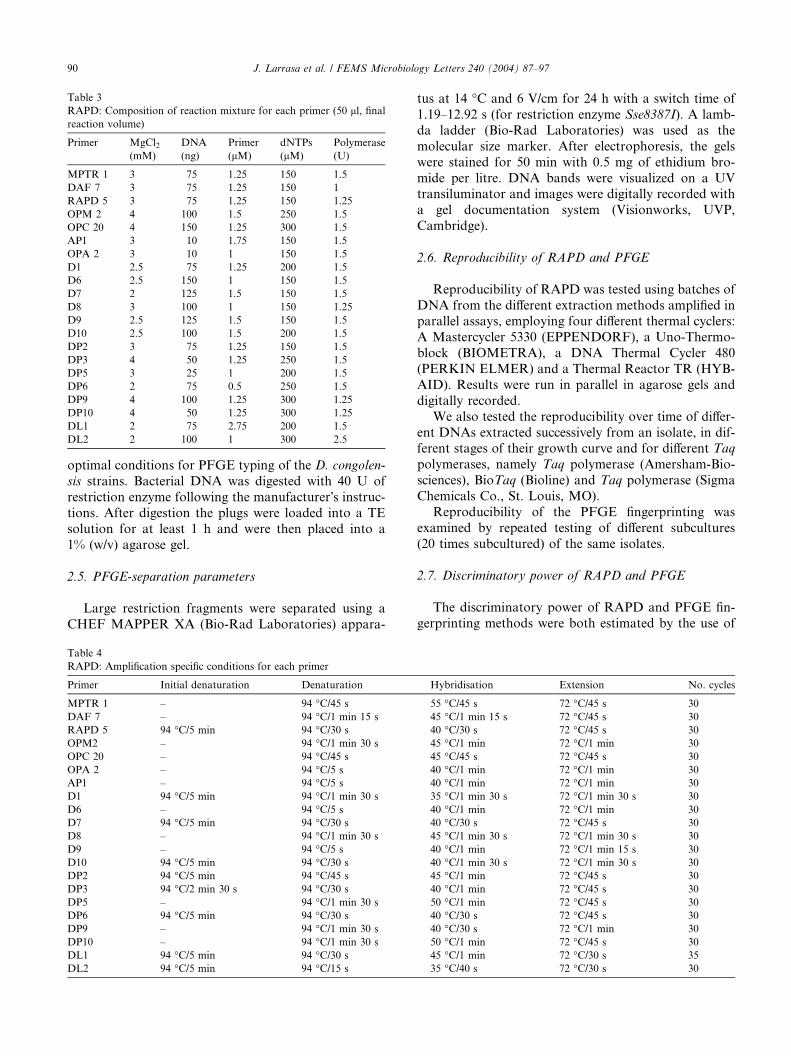
Table 3
RAPD: Composition of reaction mixture for each primer (50 ll, finalreaction volume)
Primer MgCl2(mM)
DNA
(ng)
Primer
(lM)
dNTPs
(lM)
Polymerase
(U)
MPTR 1 3 75 1.25 150 1.5
DAF 7 3 75 1.25 150 1
RAPD 5 3 75 1.25 150 1.25
OPM 2 4 100 1.5 250 1.5
OPC 20 4 150 1.25 300 1.5
AP1 3 10 1.75 150 1.5
OPA 2 3 10 1 150 1.5
D1 2.5 75 1.25 200 1.5
D6 2.5 150 1 150 1.5
D7 2 125 1.5 150 1.5
D8 3 100 1 150 1.25
D9 2.5 125 1.5 150 1.5
D10 2.5 100 1.5 200 1.5
DP2 3 75 1.25 150 1.5
DP3 4 50 1.25 250 1.5
DP5 3 25 1 200 1.5
DP6 2 75 0.5 250 1.5
DP9 4 100 1.25 300 1.25
DP10 4 50 1.25 300 1.25
DL1 2 75 2.75 200 1.5
DL2 2 100 1 300 2.5
90 J. Larrasa et al. / FEMS Microbiology Letters 240 (2004) 87–97
optimal conditions for PFGE typing of the D. congolen-
sis strains. Bacterial DNA was digested with 40 U of
restriction enzyme following the manufacturer�s instruc-tions. After digestion the plugs were loaded into a TE
solution for at least 1 h and were then placed into a
1% (w/v) agarose gel.
2.5. PFGE-separation parameters
Large restriction fragments were separated using a
CHEF MAPPER XA (Bio-Rad Laboratories) appara-
Table 4
RAPD: Amplification specific conditions for each primer
Primer Initial denaturation Denaturation
MPTR 1 – 94 �C/45 s
DAF 7 – 94 �C/1 min 15 s
RAPD 5 94 �C/5 min 94 �C/30 s
OPM2 – 94 �C/1 min 30 s
OPC 20 – 94 �C/45 s
OPA 2 – 94 �C/5 s
AP1 – 94 �C/5 s
D1 94 �C/5 min 94 �C/1 min 30 s
D6 – 94 �C/5 s
D7 94 �C/5 min 94 �C/30 s
D8 – 94 �C/1 min 30 s
D9 – 94 �C/5 s
D10 94 �C/5 min 94 �C/30 s
DP2 94 �C/5 min 94 �C/45 s
DP3 94 �C/2 min 30 s 94 �C/30 s
DP5 – 94 �C/1 min 30 s
DP6 94 �C/5 min 94 �C/30 s
DP9 – 94 �C/1 min 30 s
DP10 – 94 �C/1 min 30 s
DL1 94 �C/5 min 94 �C/30 s
DL2 94 �C/5 min 94 �C/15 s
tus at 14 �C and 6 V/cm for 24 h with a switch time of
1.19–12.92 s (for restriction enzyme Sse8387I). A lamb-
da ladder (Bio-Rad Laboratories) was used as the
molecular size marker. After electrophoresis, the gels
were stained for 50 min with 0.5 mg of ethidium bro-
mide per litre. DNA bands were visualized on a UVtransiluminator and images were digitally recorded with
a gel documentation system (Visionworks, UVP,
Cambridge).
2.6. Reproducibility of RAPD and PFGE
Reproducibility of RAPD was tested using batches of
DNA from the different extraction methods amplified inparallel assays, employing four different thermal cyclers:
A Mastercycler 5330 (EPPENDORF), a Uno-Thermo-
block (BIOMETRA), a DNA Thermal Cycler 480
(PERKIN ELMER) and a Thermal Reactor TR (HYB-
AID). Results were run in parallel in agarose gels and
digitally recorded.
We also tested the reproducibility over time of differ-
ent DNAs extracted successively from an isolate, in dif-ferent stages of their growth curve and for different Taq
polymerases, namely Taq polymerase (Amersham-Bio-
sciences), BioTaq (Bioline) and Taq polymerase (Sigma
Chemicals Co., St. Louis, MO).
Reproducibility of the PFGE fingerprinting was
examined by repeated testing of different subcultures
(20 times subcultured) of the same isolates.
2.7. Discriminatory power of RAPD and PFGE
The discriminatory power of RAPD and PFGE fin-
gerprinting methods were both estimated by the use of
Hybridisation Extension No. cycles
55 �C/45 s 72 �C/45 s 30
45 �C/1 min 15 s 72 �C/45 s 30
40 �C/30 s 72 �C/45 s 30
45 �C/1 min 72 �C/1 min 30
45 �C/45 s 72 �C/45 s 30
40 �C/1 min 72 �C/1 min 30
40 �C/1 min 72 �C/1 min 30
35 �C/1 min 30 s 72 �C/1 min 30 s 30
40 �C/1 min 72 �C/1 min 30
40 �C/30 s 72 �C/45 s 30
45 �C/1 min 30 s 72 �C/1 min 30 s 30
40 �C/1 min 72 �C/1 min 15 s 30
40 �C/1 min 30 s 72 �C/1 min 30 s 30
45 �C/1 min 72 �C/45 s 30
40 �C/1 min 72 �C/45 s 30
50 �C/1 min 72 �C/45 s 30
40 �C/30 s 72 �C/45 s 30
40 �C/30 s 72 �C/1 min 30
50 �C/1 min 72 �C/45 s 30
45 �C/1 min 72 �C/30 s 35
35 �C/40 s 72 �C/30 s 30
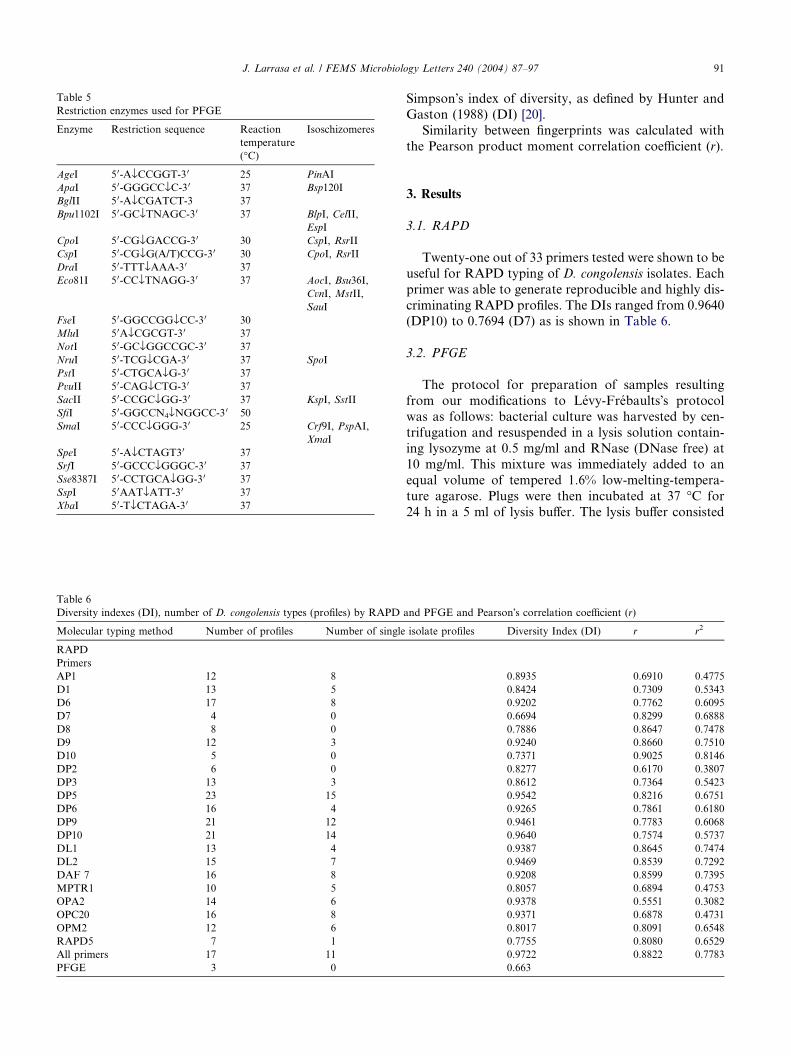
Table 6
Diversity indexes (DI), number of D. congolensis types (profiles) by RAPD
Molecular typing method Number of profiles Number of single
RAPD
Primers
AP1 12 8
D1 13 5
D6 17 8
D7 4 0
D8 8 0
D9 12 3
D10 5 0
DP2 6 0
DP3 13 3
DP5 23 15
DP6 16 4
DP9 21 12
DP10 21 14
DL1 13 4
DL2 15 7
DAF 7 16 8
MPTR1 10 5
OPA2 14 6
OPC20 16 8
OPM2 12 6
RAPD5 7 1
All primers 17 11
PFGE 3 0
Table 5
Restriction enzymes used for PFGE
Enzyme Restriction sequence Reaction
temperature
(�C)
Isoschizomeres
AgeI 5 0-AflCCGGT-3 0 25 PinAI
ApaI 5 0-GGGCCflC-30 37 Bsp120I
BglII 5 0-AflCGATCT-3 37
Bpu1102I 5 0-GCflTNAGC-30 37 BlpI, CelII,
EspI
CpoI 5 0-CGflGACCG-30 30 CspI, RsrII
CspI 5 0-CGflG(A/T)CCG-3 0 30 CpoI, RsrII
DraI 5 0-TTTflAAA-30 37
Eco81I 5 0-CCflTNAGG-30 37 AocI, Bsu36I,
CvnI, MstII,
SauI
FseI 5 0-GGCCGGflCC-30 30
MluI 5 0AflCGCGT-3 0 37
NotI 5 0-GCflGGCCGC-30 37
NruI 5 0-TCGflCGA-3 0 37 SpoI
PstI 5 0-CTGCAflG-3 0 37
PvuII 5 0-CAGflCTG-30 37
SacII 5 0-CCGCflGG-30 37 KspI, SstII
SfiI 5 0-GGCCN4flNGGCC-3 0 50
SmaI 5 0-CCCflGGG-30 25 Crf9I, PspAI,
XmaI
SpeI 5 0-AflCTAGT30 37
SrfI 5 0-GCCCflGGGC-30 37
Sse8387I 5 0-CCTGCAflGG-30 37
SspI 5 0AATflATT-30 37
XbaI 5 0-TflCTAGA-30 37
J. Larrasa et al. / FEMS Microbiology Letters 240 (2004) 87–97 91
Simpson�s index of diversity, as defined by Hunter and
Gaston (1988) (DI) [20].
Similarity between fingerprints was calculated with
the Pearson product moment correlation coefficient (r).
3. Results
3.1. RAPD
Twenty-one out of 33 primers tested were shown to be
useful for RAPD typing of D. congolensis isolates. Each
primer was able to generate reproducible and highly dis-
criminating RAPD profiles. The DIs ranged from 0.9640(DP10) to 0.7694 (D7) as is shown in Table 6.
3.2. PFGE
The protocol for preparation of samples resulting
from our modifications to Levy-Frebaults�s protocol
was as follows: bacterial culture was harvested by cen-
trifugation and resuspended in a lysis solution contain-ing lysozyme at 0.5 mg/ml and RNase (DNase free) at
10 mg/ml. This mixture was immediately added to an
equal volume of tempered 1.6% low-melting-tempera-
ture agarose. Plugs were then incubated at 37 �C for
24 h in a 5 ml of lysis buffer. The lysis buffer consisted
and PFGE and Pearson�s correlation coefficient (r)
isolate profiles Diversity Index (DI) r r2
0.8935 0.6910 0.4775
0.8424 0.7309 0.5343
0.9202 0.7762 0.6095
0.6694 0.8299 0.6888
0.7886 0.8647 0.7478
0.9240 0.8660 0.7510
0.7371 0.9025 0.8146
0.8277 0.6170 0.3807
0.8612 0.7364 0.5423
0.9542 0.8216 0.6751
0.9265 0.7861 0.6180
0.9461 0.7783 0.6068
0.9640 0.7574 0.5737
0.9387 0.8645 0.7474
0.9469 0.8539 0.7292
0.9208 0.8599 0.7395
0.8057 0.6894 0.4753
0.9378 0.5551 0.3082
0.9371 0.6878 0.4731
0.8017 0.8091 0.6548
0.7755 0.8080 0.6529
0.9722 0.8822 0.7783
0.663

Fig. 1. Electrophoresis patterns obtained by PFGE on several ovine origin strains of Dermatophilus congolensis. M (k ladder); m (k DNA-HindIII);
m 0 (kilobase DNA marker).
92 J. Larrasa et al. / FEMS Microbiology Letters 240 (2004) 87–97
of 5 ml of proteolysis buffer (1% sodium lauryl sarco-
syne, 0.5 M EDTA, pH 9.5) and 60 mg of proteinase
K (Sigma Chemicals Co., St. Louis, MO). This solution
was incubated with gentle shaking at 55 �C for 2 days.
Finally, the blocks were washed 25 times for 20 min each
at 4 �C in 50 ml of TE buffer.
Only Sse8387I restriction enzyme was effective for
PFGE typing. All D. congolensis isolates were typeableby Sse8387I-PFGE. The DNA typing profiles of each
isolate generated by PFGE were found to be stable
and reproducible. PFGE of Sse8387I digests yielded
13–23 bands in the 10–200 kb size range for D. congolen-
sis isolates. Three banding profile types were evident,
corresponding in general with the animal source of the
strain, thus the ovine type exhibited 13 bands (see Fig.
1), the bovine type 18 bands, and equine type 23 bands.The discriminatory power of PFGE as well as the poly-
morphism were lower than in RAPD, as indicated by the
comparison of the indexes of diversity and number of
types (see Table 6).
4. Discussion
4.1. Typeability of RAPD
Subtyping or strain classification can be accom-
plished by a number of different techniques. All of these
methods must meet several criteria in order to be
broadly useful: typeability, reproducibility, discrimina-
tory power and simplicity to perform.
In our study, RAPD has shown an excellent typeabil-ity (100%), following the exclusion of primers that did
not generate reproducible and discriminative profiles.
This typeability accords with other authors� findings
that RAPD is superior to ribotyping, southern blotting
or even to PFGE. For example, Van Belkum et al. [25]
consider that PCR-mediated typing is preferable, in
principle, over phage typing, plasmid typing, some of
the RFLP approaches, and IS mapping, as all of these
leave an appreciable percentage of strains un-typed.
4.2. Reproducibility of RAPD
Reproducibility refers to the ability of a technique toyield the same result when a particular strain is repeat-
edly tested. Reproducibility is especially important for
the construction of reliable databases containing all
known strains within a species to which unknown
organisms can be compared for classification.
In order to obtain good reproducibility it is essential
to define the appropriate PCR reagents, conditions,
primers, and analysis methods that will produce reliable,discriminatory, and reproducible results in different lab-
oratories [18].
In the current study, the reproducibility of RAPD
analysis was evaluated, as this is an essential character-
istic to confirm performance as acceptable.
Thus, under our experimental conditions, of the 33
primers assayed we retained 21 primers, which produced
discriminatory and reproducible profiles.RAPDs for all isolates examined in this study were
reproducible over time. Furthermore profiles were stable
for DNAs extracted from successive cultures of an iso-
late and in different stages of their life cycle.
The reproducibility of the RAPD patterns was also
consistent by using different DNA preparations, various
thermal cyclers and diverse Taq polymerases.
For D. congolensis this reproducibility was in agree-ment with other studies [26–38], showing that RAPD
can be reliable at the practical level if the following con-
ditions are addressed: (i) standardized reagents (quality
controlled), (ii) standardized extraction of DNA, and
(iii) standardized amplification conditions. In this study,

J. Larrasa et al. / FEMS Microbiology Letters 240 (2004) 87–97 93
12 out of 33 primers were rejected as they were found to
produce non-reproducible RAPD results probably due
to the critical or suboptimal reagent concentrations or
reaction conditions [39,40], or perhaps because the tar-
get amplification is extremely rare in the D. congolensis
genome [41,42].It is important that a RAPD typing method should
be reliable enough to allow between-laboratory compar-
isons. Several reports have found difficulties in obtaining
reproducible RAPD banding patterns in different labo-
ratories [19]. Our results emphasize again that each pri-
mer to be used in RAPD should be carefully tested for
reproducibility of the banding pattern, because, for
RAPD typing, the selection of primers that producereproducible and easily interpretable DNA fingerprints
is essential. Only primers that provide consistent band-
ing patterns in different environments should be chosen
for inter-laboratory use. Primers showing variability in
the banding patterns can only be used within one labo-
ratory, if they produce consistent results in the thermo-
cycler used.
4.3. Discriminatory power of RAPD
In 1988, Hunter and Gaston [20] used a single numer-
ical index of discrimination, the discriminatory index
(DI), based on the probability that two unrelated iso-
lates would be classified as the same type. Since then,
the DI has been widely used to compare the discrimina-
tory power of different typing methods, and the determi-nation of this index is now recommended in guidelines
for the evaluation of typing methods. This study is the
first to use DI to compare D. congolensis typing
methods.
In order to evaluate the discriminatory power of
RAPD analysis for typing of D. congolensis isolates,
we studied 50 unrelated D. congolensis strains isolated
over a significant period of time and from different geo-graphic origins. We also included and three type strains
from American Type Culture Collection and British
Type Culture Collection.
According to recent guidelines [17,20,43], a typing
system should achieve a DI of >0.95 for reliable assess-
ment of the clonal relatedness of isolates or, if typing re-
sults are to be interpreted with confidence, a DI of
greater than 0.90 is desirable [20,44,45].Our study has shown that RAPD offers significant
discrimination among D. congolensis isolates (DI,
0.9722). However, the level of discrimination depends
on the primer being used (see Table 6). Primer DP10
exhibited the greatest discriminatory power (DI,
0.9640) while primer D7 was the less discriminating
(DI, 0.7694).
These results are in agreement with those of otherauthors who found RAPD a satisfactory method for
typing microorganisms [36,43,44,46–61].
For comparison, we have applied Hunter and Gas-
ton formula on results of previously published com-
parisons of D. congolensis isolates, many of them
included in the present study, based on phenotype fea-
tures and carried out by other methods. Thus, the dis-
criminatory power of SDS–PAGE for characterizationof D. congolensis exoproteins [6] ranged from 0.6004
to 0.8464 while the discrimination power of API
ZYM system [9] was 0.9341. Our results showed that
RAPD is more discriminating than these, although
caution is required as these included a lower number
of isolates.
Considering its combination of discriminatory power
and ease of application, RAPD emerges as a particularlyattractive molecular technique especially if the simplified
RAPD technique using heated bacterial DNA templates
[14] is employed to reduce the time and labour required.
This study shows that when an appropriately chosen set
of primers is employed, RAPD analysis provides a ra-
pid, reproducible, and powerful genomic typing method
for D. congolensis.
4.4. PFGE sample preparation
The preparation and manipulation of very large
DNA molecules for pulsed field gel electrophoresis
(PFGE) is a laborious procedure which requires more
care than for smaller molecules. Generally DNA is
prepared in solid agarose blocks (plugs). However,
this is often difficult due to many factors, 4 of whichare discussed below: (a) Difficulty emulsifying cells.
The cells should be suspended homogeneously because
aggregation, evident as a lumpy appearance reduces
the efficiency of cell digestion. For example, the bands
generated by PFGE during typing of Flavimonas ory-
zihabitans were sometimes distorted, because it was
difficult to emulsify the colonies of this species into
homogeneous suspensions. As a consequence repeatedtesting was necessary before the patterns could be
compared with confidence [62]. In the present study,
we found it was easier to emulsify the colonies of
D. congolensis taken from solid media cultures where
the filaments tend to fragment into cocci more easily
than those from liquid media. Liquid media cultures
required more washes with detergent and gentle
shaking.(b) The samples are too concentrated. Pulsed field
gels are especially sensitive to sample overload. As the
amount of DNA increases, its rate of migration is re-
tarded correspondingly, resulting in larger than true
sizes. In general, it is recommended that an agarose plug
should contain 106–1010 cells; equivalent to between 100
ng and 1 lg [51,63]. In our study, we have optimised the
number of cells per plug, at 107 D. congolensis. This ena-bled us to assess the appropriate conditions to optimise
PFGE resolution.

94 J. Larrasa et al. / FEMS Microbiology Letters 240 (2004) 87–97
(c) Inadequate enzymatic treatment. Following inad-
equate enzymatic treatment plugs often have a sandy or
gritty aspect. In the present study, we firstly used DNA
prepared by Levy-Frebault et al. protocol [21]. How-
ever, the enzymatic treatment did not degrade the cell
wall of D. congolensis. On the other hand, when thegenomic DNA of the test strains was prepared in agar-
ose plugs by the rapid DNA extraction procedure de-
scribed by Matushek et al. [23] a small percentage of
the organisms did not lyse during the first 2 h incuba-
tion. However, adequate lysis did occur (the plugs had
a transparent aspect) following a second incubation in
lysis buffer for 24 h. Thus, mixing the bacterial suspen-
sion with lysis solution prior to adding low meltingpoint agarose was essential to complete the lysis of the
cell wall.
(d) Degradation of the DNA during extraction. A
number of studies have reported difficulty PFGE typing
bacterial species due to difficulties obtaining non-
degraded DNA [26,59,64]. The reason for this may be
the low yields associated of DNA isolation in these spe-
cies, further hampered by DNA degradation problemscaused by extracellular DNases. Proposed solutions to
prevent DNAase-related problems include formalde-
hyde fixation of cells, heat treatment of cells [65–67],
the use of lysis solution for the resuspension of cells
prior to mixing with the insert gel [23], inclusion of
hypertonic sucrose in the lysis solution [68], and shorten-
ing of the lysis and DNA plug wash steps [23,69,70].
We evaluated the effect of different in situ preparationmethods on the quality of DNA in order to optimize the
PFGE typing of D. congolensis strains. The protocol re-
ported here has been developed and tested to address the
above problematic factors. It provides an efficient and
reproducible method for preparation of D. congolensis
genomic DNA for PFGE.
4.5. PFGE-Restriction enzyme digestion
The selection of restriction enzymes for PFGE is spe-
cific for each microorganism, and must take account of
the G+C contents of the target DNAs [71]. For efficient
initial mapping of bacterial chromosomes it is advisable
to use enzymes, which have relatively few restriction
sites and give larger fragments from the target DNA.
It must be taken into account that pulsed field gel elect-rophoresis allows clear separation of very-large-molecu-
lar-length DNA fragments ranging from 10 to 800 kb
[51].
By contrast, enzymes producing a large number of
small fragments (10 kb) are unlikely to be useful for
PFGE and mapping. In general, only restriction enzymes
with six base-pair recognition sites, or greater, are useful.
In thios study we tested 24 restriction enzymes with 6–8base-pair recognition sites. Only Sse8387I yielded well-
resolved patterns composed of a limited number (1–20)
of fragments, as recommended [72]. Other enzymes
tested yielded excessive numbers of small fragments, that
were impossible to resolve by PFGE.
It is important to recognize the problems of partial
DNA digestion when using restriction enzymes in PFGE
analysis. Partial digestion can be overcome by an ex-tended overnight (20 h) incubation or by replacing the
reaction buffer after 6 h, adding fresh enzyme, and incu-
bating the mixture for another 6 h [73]. Many sub-
stances, such as the lysed bacterial material, proteinase
K, PMSF, and TE buffer, can decrease the activity of
the restriction enzymes. Hence, these substances must
be eliminated as efficiently as possible. Furthermore,
for optimal enzyme activity, it is important to washthe DNA blocks in the restriction enzyme reaction buf-
fer at 4 �C for 30 min before incubating the DNA blocks
in fresh restriction enzyme reaction buffer containing
endonuclease.
Results from using our modified protocol, which in-
cludes washing steps at the end of the DNA sample
preparation procedure and a distilled water washing
step prior to restriction enzyme digestion, suggest thatthe lack of proteinase K, PMSF, and TE buffer im-
proves efficient digestion of D. congolensis DNA.
4.6. Typeability of PFGE
Numerous factors influence the preparation of intact
chromosomal DNA for PFGE analysis and contribute
to the low typeability of PFGE reported in several stud-ies [43]. By contrast, the results of the present study, like
those of Louie et al [74] show that PFGE can have excel-
lent typeability: all strains of D. congolensis were amena-
ble to PFGE analysis.
4.7. Reproducibility of PFGE
Guidelines for PFGE typing indicate that variationsin some bands occur when strains of some species are cul-
tured repeatedly over time [75]. By contrast in this study
genome patterns (PFGE types) D. congolensis strains
were unchanged after 20 passages showing the reproduc-
ibility of the PFGE technique with D. congolensis.
Moreover, using our protocol we found the banding
pattern was also consistent when using DNA samples
prepared 5 months, in common with McEllistrem et al.(2000) [76] when PFGE typing Streptococcus
pneumoniae.
4.8. Discriminatory power of PFGE
Applying the guidelines [75] to our results, the dis-
criminatory power of Sse8387I-PFGE in the present
study was low (DI, 0.663). Although it is desirablethat any subtyping method has a high differentiation
power, epidemiological data should be taken in ac-

J. Larrasa et al. / FEMS Microbiology Letters 240 (2004) 87–97 95
count when deciding whether genetically related
strains are also epidemiologically related. Thus, the
generation of excessive numbers of types introduces
the possibility that the relationship between the typing
data and epidemiological characteristics will start to
deteriorate [25].Results of a PFGE typing study of Staphylococcus
aureus [55] showed that the resolution of this method ex-
ceeded that of restriction fragment length polymorphism
analysis with ribosomal or toxin A DNA probes and
that it did not correlated with epidemiological data.
Gori et al. [77] showed that RAPD and PFGE were
useful for typing Klebsiella pneumoniae strains in epide-
miological investigations, and although RAPD analysiswas more efficient, more subclonal variants were deter-
mined among epidemic clones by PFGE analysis than
by RAPD analysis.
In our opinion, the PFGE typing data obtained in the
present study interpreted in the context of epidemiolog-
ical information on the host and geographic origin dem-
onstrates the relationship of D. congolensis isolates,
while correlating with RAPD results. Indeed the degreeof correlation between RAPD and PFGE typing was
significantly high (r, 0.8822).
The low discriminatory power of PFGE can be ex-
plained because Sse8387I-PFGE could not distinguish
between D. congolensis strains from the same host.
Interestingly this association with host species accords
with results of an examination of D. congolensis pro-
teases [6]. In that study distinct patterns of enzymemigration in PAGE gels were observed to be consistent
within isolates from the 3 main host species (r, 0.9416).
In conclusion, we have found RAPD and PFGE to
be satisfactory methods for typing D. congolensis that
can distinguish between D. congolensis strains with a
high degree of correlation between two techniques. We
recommend that at least two independent primers
should be used for RAPD typing in order to improveits discriminatory power and that PFGE be used for
confirmation of RAPD results.
Acknowledgements
This research was supported by the Spanish Govern-
ment Ministerio de Educacion y Cultura, grant PB96-0478 and by the European Commission INCO-DC Pro-
gram Project Contract CT980334.
The authors would like to thank Alex Morrow and
Ian Heron, University of Edinburgh, for providing part
of the D. congolensis isolates used in this study. Thanks
also to Dr. K.H. Bohm, Institut fur Mikrobiologie und
Tierseuchen, Tierarztlichen Hochschule Hannover, Ger-
many, for providing German horse isolates of D.
congolensis.
References
[1] Roberts, D.S. (1965) The histopathology of epidermal infection
with the actinomycete Dermatophilus congolensis. J. Pathol.
Bacteriol. 90, 213–216.
[2] Thierry, G. and Memery, G. (1961) La streptothricose cutanee IV.
Etiologie-traitement-prophylaxie. Rev. Elev. Med. Vet. Pays
Trop. 14, 413–427.
[3] Perreau, P. (1981) La dermatophilose infection a Dermatophilus
congolensis. Polycopie d�enseignement. I.E.M.V.T.
[4] Barouh, B.J.E. (1984) La dermatophilose animale. Situation en
France. These Doct. Vet., Ecole Nationale Veterinaire d�Alfort.,
Alfort.
[5] Ambrose, N., Lloyd, D. and Maillard, J.C. (1999) Immune
responses to Dermatophilus congolensis infections. Parasitol.
Today 15, 295–300.
[6] Ambrose, N.C., Mijinyawa, M.S. and Hermoso de Mendoza, J.
(1998) Preliminary characterisation of extracellular serine pro-
teases of Dermatophilus congolensis isolates from cattle, sheep and
horses. Vet. Microbiol. 62, 321–335.
[7] Ellis, T.M., Masters, A.M., Sutherland, S.S., Carson, J.M. and
Gregory, A.R. (1993) Variation in cultural, morphological,
biochemical properties and infectivity of Australian isolates of
Dermatophilus congolensis. Vet. Microbiol. 38, 81–102.
[8] Hanel, H., Kalisch, J., Keil, M., Marsch, W.C. and Buslau, M.
(1991) Quantification of keratinolytic activity from Dermatophilus
congolensis. Med. Microbiol. Immunol. 180, 45–51.
[9] Hermoso de Mendoza, J., Arenas, A., Alonso, J.M., Rey, J.M.,
Gil, M.C., Anton, J.M. and Hermoso de Mendoza, M. (1993)
Enzymatic activities of Dermatophilus congolensis measured by
API ZYM. Vet. Microbiol. 37, 175–179.
[10] How, S.J. and Lloyd, D.H. (1990) The effect of recent vaccination
on the dose-response to experimental Dermatophilus congolensis
infection in rabbits. J. Comp. Pathol. 102, 157–163.
[11] Makinde, A.A. and Gyles, C.L. (1999) A comparison of extracted
proteins of isolates of Dermatophilus congolensis by sodium
dodecyl sulphate-polyacrylamide gel electrophoresis and Western
blotting. Vet. Microbiol. 67, 251–262.
[12] Skalka, B. and Pospisil, L. (1992) Hemolytic interactions of
Dermatophilus congolensis. Zentralbl Veterinarmed [B] 39, 139–
143.
[13] Skalka, B. and Votava, M. (1997) Hemolysis and hemolytic
interactions of bacteria. Epidemiol. Mikrobiol. Imunol. 46, 87–98.
[14] Larrasa, J., Garcıa, A., Ambrose, N.C., Alonso, J.M., Parra, A.,
Hermoso de Mendoza, M., Salazar, J., Rey, J. and Hermoso de
Mendoza, J. (2002) A simple random amplified polymorphic
DNA genotyping method for field isolates of Dermatophilus
congolensis. J. Vet. Med. B, 135–141.
[15] Makinde, A.A. (1991) Immunoserology of Dermatophilus congo-
lensis infection. In: Presented at the 2nd International Symposium
on Dermatophilosis, Vom, Nigeria.
[16] Olive, D.M. and Bean, P. (1999) Principles and applications of
methods for DNA-based typing of microbial organisms. J. Clin.
Microbiol. 37, 1661–1669.
[17] Struelens, M.J. (1996) Consensus guidelines for appropriate use
and evaluation of microbial epidemiologic typing systems. In:
Presented at the European Estudy Group of epidemiological
Markers (ESGEM), of European Society for Clinical Microbiol-
ogy and Infectious Diseases (ESCMID).
[18] Arbeit, R.D. (1995) Laboratory procedures for the epidemiologic
analysis of microorganisms, sixth ed. ASM Press, Washington,
DC.
[19] Tyler, K.D., Wang, G., Tyler, S.D. and Johnson, W.M. (1997)
Factors affecting reliability and reproducibility of amplification-
based DNA fingerprinting of representative bacterial pathogens.
J. Clin. Microbiol. 35, 339–346.

96 J. Larrasa et al. / FEMS Microbiology Letters 240 (2004) 87–97
[20] Hunter, P.R. and Gaston, M.A. (1988) Numerical index of
the discriminatory ability of typing systems: an application of
Simpson�s index of diversity. J. Clin. Microbiol. 26, 2465–
2466.
[21] Levy-Frebault, V.V., Thorel, M.F., Varnerot, A. and Gicquel, B.
(1989) DNA polymorphism in Mycobacterium paratuberculosis,
‘‘wood pigeon mycobacteria, and related mycobacteria analyzed
by field inversion gel electrophoresis. J. Clin. Microbiol. 27, 2823–
2826.
[22] Udo, E.E. and Grubb, W.B. (1993) Genetic analysis of methicil-
lin-resistant Staphylococcus aureus from a Nigerian hospital. J.
Med. Microbiol. 38, 203–208.
[23] Matushek, M.G., Bonten, M.J. and Hayden, M.K. (1996) Rapid
preparation of bacterial DNA for pulsed-field gel electrophoresis.
J. Clin. Microbiol. 34, 2598–2600.
[24] Cornillot, E., Croux, C. and Soucaille, P. (1997) Physical and
genetic map of the Clostridium acetobutylicum ATCC824 chro-
mosome. J. Bacteriol. 179, 7426–7434.
[25] van Belkum, A., Kluytmans, J., van Leeuwen, W., Bax, R., Quint,
W., Peters, E., Fluit, A., Vandenbroucke-Grauls, C., van den
Brule, A. and Koeleman, H., et al. (1995) Multicenter evaluation
of arbitrarily primed PCR for typing of Staphylococcus aureus
strains. J. Clin. Microbiol. 33, 1537–1547.
[26] Zhang, Y., Rajagopalan, M., Brown, B.A. and Wallace Jr., R.J.
(1997) Randomly amplified polymorphic DNA PCR for compar-
ison of Mycobacterium abscessus strains from nosocomial out-
breaks. J. Clin. Microbiol. 35, 3132–3139.
[27] Yao, J.D., Conly, J.M. and Krajden, M. (1995) Molecular typing
of Stenotrophomonas (Xanthomonas) maltophilia by DNA macro-
restriction analysis and random amplified polymorphic DNA
analysis. J. Clin. Microbiol. 33, 2195–2198.
[28] Van Couwenberghe, C.J., Cohen, S.H., Tang, Y.J., Gumerlock,
P.H. and Silva Jr., J. (1995) Genomic fingerprinting of epidemic
and endemic strains of Stenotrophomonas maltophilia formerly
Xanthomonas maltophilia by arbitrarily primed PCR. J. Clin.
Microbiol. 33, 1289–1291.
[29] Sloos, J.H., Dijkshoorn, L., Vogel, L. and van Boven, C.P. (2000)
Performance of phenotypic and genotypic methods to determine
the clinical relevance of serial blood isolates of Staphylococcus
epidermidis in patients with septicemia. J. Clin. Microbiol. 38,
2488–2493.
[30] Salama, S.M., Jiang, Q., Chang, N., Sherbaniuk, R.W. and
Taylor, D.E. (1995) Characterization of chromosomal DNA
profiles from Helicobacter pylori strains isolated from sequential
gastric biopsy specimens. J. Clin. Microbiol. 33, 2496–2497.
[31] Saito, M., Umeda, A. and Yoshida, S. (1999) Subtyping of
Haemophilus influenzae strains by pulsed-field gel electrophoresis.
J. Clin. Microbiol. 37, 2142–2147.
[32] Okazaki, M., Watanabe, T., Morita, K., Higurashi, Y., Araki, K.,
Shukuya, N., Baba, S., Watanabe, N., Egami, T., Furuya, N.,
Kanamori, M., Shimazaki, S. and Uchimura, H. (1999) Molecular
epidemiological investigation using a randomly amplified poly-
morphic DNA assay of Burkholderia cepacia isolates from
nosocomial outbreaks. J. Clin. Microbiol. 37, 3809–3814.
[33] Mahenthiralingam, E., Campbell, M.E., Foster, J., Lam, J.S. and
Speert, D.P. (1996) Random amplified polymorphic DNA typing
of Pseudomonas aeruginosa isolates recovered from patients with
cystic fibrosis. J. Clin. Microbiol. 34, 1129–1135.
[34] Giovannacci, I., Ragimbeau, C., Queguiner, S., Salvat, G.,
Vendeuvre, J.L., Carlier, V. and Ermel, G. (1999) Listeria
monocytogenes in pork slaughtering and cutting plants. Use of
RAPD, PFGE and PCR-REA for tracing and molecular epide-
miology. Int. J. Food Microbiol. 53, 127–140.
[35] Connor, K.M., Quirie, M.M., Baird, G. and Donachie, W. (2000)
Characterization of United Kingdom isolates of Corynebacterium
pseudotuberculosis using pulsed-field gel electrophoresis. J. Clin.
Microbiol. 38, 2633–2637.
[36] Clemons, K.V., Feroze, F., Holmberg, K. and Stevens, D.A.
(1997) Comparative analysis of genetic variability among Candida
albicans isolates from different geographic locales by three
genotypic methods. J. Clin. Microbiol. 35, 1332–1336.
[37] Bansal, N.S. and McDonell, F. (1997) Identification and DNA
fingerprinting of Legionella strains by randomly amplified poly-
morphic DNA analysis. J. Clin. Microbiol. 35, 2310–2314.
[38] Atia, M. (1981) Preliminary investigation on the ecology of
Dermatophilus congolensis. Mykosen. 24, 153–155.
[39] Bradley, A.F. (1992). Tecnologıa PCR: Fundamentos. Perkin
Elmer Hispania.
[40] Innis, M.S., Myambo, K.B., Gelfand, D.H. and Brown, M.A.
(1992) DNA sequencing with Thermus acquaticus DNA polym-
erase and direct sequencing of polymerase chain reaction-ampli-
fied DNA. 1988. Biotechnology 24, 6–10.
[41] Karlin, S. and Brendel, V. (1992) Chance and statistical signif-
icance in protein and DNA sequence analysis. Science 257, 39–49.
[42] Burge, C., Campbell, A.M. and Karlin, S. (1992) Over- and
under-representation of short oligonucleotides in DNA sequences.
Proc. Natl. Acad. Sci. USA 89, 1358–1362.
[43] Burucoa, C., Lhomme, V. and Fauchere, J.L. (1999) Performance
criteria of DNA fingerprinting methods for typing of Helicobacter
pylori isolates: experimental results and meta-analysis. J. Clin.
Microbiol. 37, 4071–4080.
[44] Van Looveren, M., Ison, C.A., Ieven, M., Vandamme, P., Martin,
I.M., Vermeulen, K., Renton, A. and Goossens, H. (1999)
Evaluation of the discriminatory power of typing methods for
Neisseria gonorrhoeae. J. Clin. Microbiol. 37, 2183–2188.
[45] Martinez, G., Harel, J., Higgins, R., Lacouture, S., Daignault, D.
and Gottschalk, M. (2000) Characterization of Streptococcus
agalactiae isolates of bovine and human origin by randomly
amplified polymorphic DNA analysis. J. Clin. Microbiol. 38, 71–
78.
[46] Weel, J.F., van der Hulst, R.W., Gerrits, Y., Tytgat, G.N., van
der Ende, A. and Dankert, J. (1996) Heterogeneity in suscepti-
bility to metronidazole among Helicobacter pylori isolates from
patients with gastritis or peptic ulcer disease. J. Clin. Microbiol.
34, 2158–2162.
[47] Wada, S., Matsuda, M., Kikuchi, M., Kodama, T., Takei, I.,
Ogawa, S., Takahashi, S., Shingaki, M. and Itoh, T. (1994)
Genome DNA analysis and genotyping of clinical isolates of
Helicobacter pylori. Cytobios 80, 109–116.
[48] Speijer, H., Savelkoul, P.H., Bonten, M.J., Stobberingh, E.E. and
Tjhie, J.H. (1999) Application of different genotyping methods for
Pseudomonas aeruginosa in a setting of endemicity in an intensive
care unit. J. Clin. Microbiol. 37, 3654–3661.
[49] Soto, S.M., Guerra, B., Gonzalez-Hevia, M.A. and Mendoza,
M.C. (1999) Potential of three-way randomly amplified polymor-
phic DNA analysis as a typing method for twelve Salmonella
serotypes. Appl. Environ. Microbiol. 65, 4830–4836.
[50] Schutze, K., Hentschel, E., Dragosics, B. and Hirschl, A.M.
(1995) Helicobacter pylori reinfection with identical organisms:
transmission by the patients� spouses. Gut 36, 831–833.
[51] Monaco, A.P. (1995) Pulse Field Electrophoresis: A Practical
Approach, first ed. Oxcford University Press, New York.
[52] Marois, C., Dufour-Gesbert, F. and Kempf, I. (2001) Comparison
of pulsed-field gel electrophoresis with random amplified poly-
morphic DNA for typing of Mycoplasma synoviae. Vet. Micro-
biol. 79, 1–9.
[53] Malorny, B., Schroeter, A., Bunge, C., Hoog, B., Steinbeck, A.
and Helmuth, R. (2001) Evaluation of molecular typing methods
for Salmonella enterica serovar typhimurium DT104 isolated in
Germany from healthy pigs. Vet. Res. 32, 119–129.
[54] Limansky, A.S., Sutich, E.G., Guardati, M.C., Toresani, I.E. and
Viale, A.M. (1998) Genomic diversity among Streptococcus
agalactiae isolates detected by a degenerate oligonucleotide-
primed amplification assay. J. Infect. Dis. 177, 1308–1313.

J. Larrasa et al. / FEMS Microbiology Letters 240 (2004) 87–97 97
[55] Jorgensen, M., Givney, R., Pegler, M., Vickery, A. and Funnell,
G. (1996) Typing multidrug-resistant Staphylococcus aureus:
conflicting epidemiological data produced by genotypic and
phenotypic methods clarified by phylogenetic analysis. J. Clin.
Microbiol. 34, 398–403.
[56] Dzierzanowska, D., Gzyl, A., Rozynek, E., Augustynowicz, E.,
Wojda, U., Celinska-Cedro, D., Sankowska, M. and Wad-
strom, T. (1996) PCR for identification and typing of
Helicobacter pylori isolated from children. J. Physiol. Pharma-
col. 47, 101–114.
[57] Chatellier, S., Ramanantsoa, C., Harriau, P., Rolland, K.,
Rosenau, A. and Quentin, R. (1997) Characterization of Strep-
tococcus agalactiae strains by randomly amplified polymorphic
DNA analysis. J. Clin. Microbiol. 35, 2573–2579.
[58] Calcagno, A.M., Nino-Vega, G., San-Blas, F. and San-Blas, G.
(1998) Geographic discrimination of Paracoccidioides brasiliensis
strains by randomly amplified polymorphic DNA analysis. J.
Clin. Microbiol. 36, 1733–1736.
[59] Bidet, P., Lalande, V., Salauze, B., Burghoffer, B., Avesani, V.,
Delmee, M., Rossier, A., Barbut, F. and Petit, J.C. (2000)
Comparison of PCR-ribotyping, arbitrarily primed PCR, and
pulsed-field gel electrophoresis for typing Clostridium difficile. J.
Clin. Microbiol. 38, 2484–2487.
[60] Bert, F., Branger, C. and Lambert-Zechovsky, N. (1997) Pulsed-
field gel electrophoresis is more discriminating than multilocus
enzyme electrophoresis and random amplified polymorphic DNA
analysis for typing pyogenic streptococci. Curr. Microbiol. 34,
226–229.
[61] Akopyanz, N., N.O, Bukanov, T.U., Westblom, S., Kresovich, A.
and Berg, D.E. (1992) DNA diversity among clinical isolates of
Helicobacter pylori detected by PCR-based RAPD fingerprinting.
Nucl. Acids Res. 20, 5137–5142.
[62] Liu, P.Y., Shi, Z.Y., Lau, Y.J., Hu, B.S., Shyr, J.M., Tsai, W.S.,
Lin, Y.H. and Tseng, C.Y. (1996) Epidemiological typing of
Flavimonas oryzihabitans by PCR and pulsed-field gel electropho-
resis. J. Clin. Microbiol. 34, 68–70.
[63] Birren, B.W., Lai, E., Clark, S.M., Hood, L. and Simon, M.I.
(1988) Optimized conditions for pulsed field gel electrophoretic
separations of DNA. Nucl. Acids Res. 16, 7563–7582.
[64] Owen, R.J., Sutherland, K., Fitzgerald, C., Gibson, J., Borman,
P. and Stanley, J. (1995) Molecular subtyping scheme for
serotypes HS1 and HS4 of Campylobacter jejuni. J. Clin.
Microbiol. 33, 872–877.
[65] Lin, W.J. and Johnson, E.A. (1995) Genome analysis of Clostri-
dium botulinum type A by pulsed-field gel electrophoresis. Appl.
Environ. Microbiol. 61, 4441–4447.
[66] Lawson, P.A., Gharbia, S.E., Shah, H.N. and Clark, D.R. (1989)
Recognition of Fusobacterium nucleatum subgroups Fn-1, Fn-2
and Fn-3 by ribosomal RNA gene restriction patterns. FEMS
Microbiol. Lett. 53, 41–45.
[67] Kristjansson, M., Samore, M.H., Gerding, D.N., DeGirolami,
P.C., Bettin, K.M., Karchmer, A.W. and Arbeit, R.D. (1994)
Comparison of restriction endonuclease analysis, ribotyping,
and pulsed-field gel electrophoresis for molecular differentiation
of Clostridium difficile strains. J. Clin. Microbiol. 32, 1963–
1969.
[68] Wilkinson, S.R. and Young, M. (1993) Wide diversity of genome
size among different strains of Clostridium acetobutylicum. J. Gen.
Microbiol, 1069–1076.
[69] Gibson, J.R., Sutherland, K. and Owen, R.J. (1994) Inhibition of
DNAse activity in PFGE analysis of DNA from Campylobacter
jejuni. Lett. Appl. Microbiol. 19, 357–358.
[70] Chachaty, E., Saulnier, P., Martin, A., Mario, N. and Andre-
mont, A. (1994) Comparison of ribotyping, pulsed-field gel
electrophoresis and random amplified polymorphic DNA for
typing Clostridium difficile strains. FEMS Microbiol. Lett. 122,
61–68.
[71] Hood, M.T. and Stachow, C.S. (1995) Electroporation of
Schizosaccharomyces pombe. Methods Mol. Biol. 47, 273–278.
[72] Tenover, F.C., Arbeit, R.D., Goering, R.V., Mickelsen, P.A.,
Murray, B.E., Persing, D.H. and Swaminathan, B. (1995)
Interpreting chromosomal DNA restriction patterns produced
by pulsed-field gel electrophoresis: criteria for bacterial strain
typing. J. Clin. Microbiol. 33, 2233–2239.
[73] Haertl, R. and Bandlow, G. (1993) Molecular typing of Enterob-
acter cloacae by pulsed-field gel electrophoresis of genomic
restriction fragments. J. Hosp. Infect. 25, 109–116.
[74] Louie, M., Jayaratne, P., Luchsinger, I., Devenish, J., Yao, J.,
Schlech, W. and Simor, A. (1996) Comparison of ribotyping,
arbitrarily primed PCR, and pulsed-field gel electrophoresis for
molecular typing of Listeria monocytogenes. J. Clin. Microbiol.
34, 15–19.
[75] Tenover, F.C., Arbeit, R.D. and Goering, R.V. (1997) How to
select and interpret molecular strain typing methods for epidemi-
ological studies of bacterial infections: a review for healthcare
epidemiologists. Molecular Typing Working Group of the Society
for Healthcare Epidemiology of America. Infect. Control Hosp.
Epidemiol. 18, 426–439.
[76] McEllistrem, M.C., Pass, M., Elliott, J.A., Whitney, C.G. and
Harrison, L.H. (2000) Clonal Groups of Penicillin-Nonsusceptible
Streptococcus pneumoniae in Baltimore, Maryland: a Population-
Based, Molecular Epidemiologic Study. J. Clin. Microbiol. 38,
4367–4372.
[77] Gori, A., Espinasse, F., Deplano, A., Nonhoff, C., Nicolas, M.H.
and Struelens, M.J. (1996) Comparison of pulsed-field gel
electrophoresis and randomly amplified DNA polymorphism
analysis for typing extended-spectrum-beta-lactamase- producing
Klebsiella pneumoniae. J. Clin. Microbiol. 34, 2448–2453.
[78] Glennon, M., Jager, B., Dowdall, D., Maher, M., Dawson, M.,
Quigley, F., Costello, E. and Smith, T. (1997) PCR-based
fingerprinting of Mycobacterium bovis isolates. Vet Microbiol.
54, 235–245.
[79] Friedman, C.R., Stoeckle, M.Y., Johnson Jr., W.D. and Riley,
L.W. (1995) Double-repetitive-element PCR method for subtyp-
ing Mycobacterium tuberculosis clinical isolates. J Clin Micro-
biol. 33, 1383–1384.
[80] Aranaz, A., Liebana, E., Mateos, A., Dominguez, L., Vidal, D.,
Domingo, M., Gonzolez, O., Rodriguez-Ferri, E.F., Bunschoten,
A.E., Van Embden, J.D. and Cousins, D. (1996) Spacer oligonu-
cleotide typing of Mycobacterium bovis strains from cattle and
other animals: a tool for studying epidemiology of tuberculosis. J
Clin Microbiol. 34, 2734–2740.
Copyright © 2022 FDOKUMEN




















